
 1500-2000 видов")






")


")

































")

































")
 Семейство: ПОРФИРИДИЕВЫЕ (PORPHYRIDIACEAE)Вид: ПОРФИРИДИУМ БАГРЯНЫЙ (ПАРФIРЫДЫУМ")




")






 карпогона Batrachospermum:")

")








 biology
biologySimilar presentations:



")
")




. Лекция - 5")
Отделы Бурые и Красные водоросли (лекция 5)
1. ЛЕКЦИЯ 5 Отделы Бурые, Красные водоросли
2. Отдел Бурые водоросли (Phaeophyta) 1500-2000 видов
3.
4. Бурые водоросли – ближайшие родственники диатомей
Спермий Laminaria angustataКлетка с хлоропластом
А – вид с
вентральной
стороны
Б – базальные
тела жгутиков и
основания
жгутиковых
корешков
А – хлоропласт, Б – пиреноид, В – обкладка пиреноида, Г – ядро, Д – ядрышко, Е
– ядерная оболочка, сосоящая из двух мембран и окружающая хлоропласт, Ж –
диктиосома, З – вакуоль, М – митохондрия, К – фрагмент канала ЭПС, Л –
оболочка хлоропласта (две внутренние мембраны).
5. Другие особенности строения клеток:
• Клеточная стенка всегда двухслойная:внутренний слой целлюлозный,
• наружный слой пектиновый с включением
альгиновой кислоты и её солей,
фукоидана, способных сильно набухать и
ослизняться.
6.
Часто встречаются физоды – очень
мелкие (до 4 мкм в диаметре) вакуоли,
заполненные фукозаном (особого типа
дубильное вещество, подобное танину). В
молодых клетках физоды бесцветные, в
старых – окрашены в желтый или бурый
цвет.
к пигментному составу хризофитов
добавляется -каротин;
запасные углеводы - -1,3 полигликан
ламинарин и сахароспирт маннитол,
танины и жиры.
7.
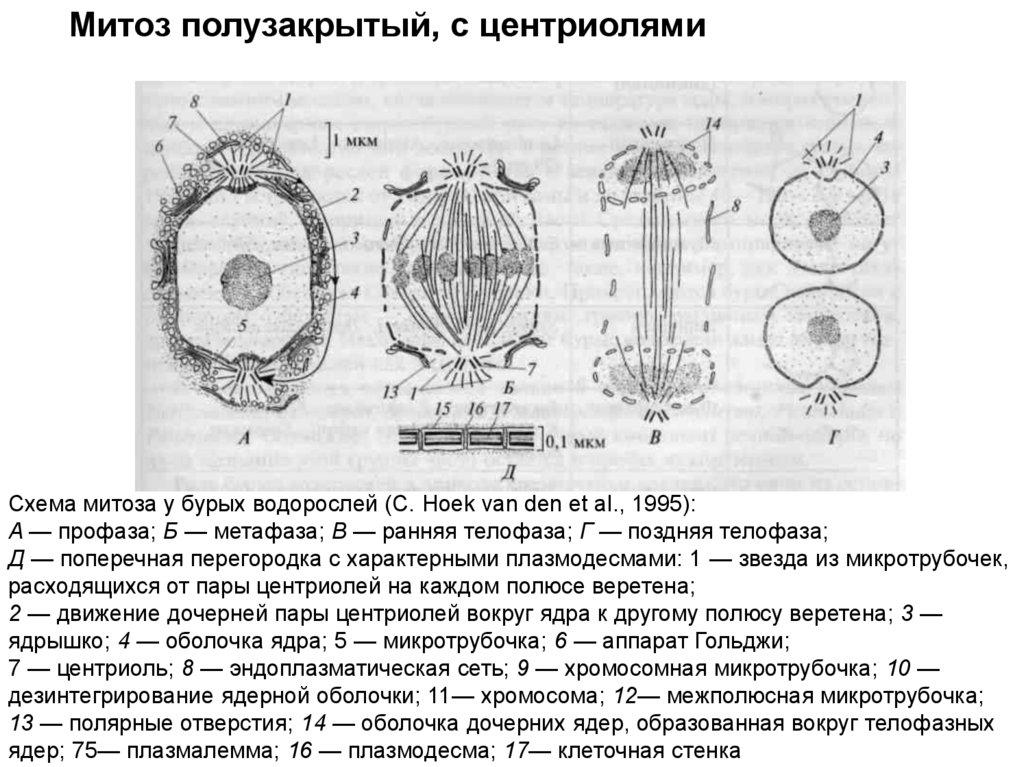
Митоз полузакрытый, с центриолямиСхема митоза у бурых водорослей (С. Hoek van den et al., 1995):
A — профаза; Б — метафаза; В — ранняя телофаза; Г — поздняя телофаза;
Д — поперечная перегородка с характерными плазмодесмами: 1 — звезда из микротрубочек,
расходящихся от пары центриолей на каждом полюсе веретена;
2 — движение дочерней пары центриолей вокруг ядра к другому полюсу веретена; 3 —
ядрышко; 4 — оболочка ядра; 5 — микротрубочка; 6 — аппарат Гольджи;
7 — центриоль; 8 — эндоплазматическая сеть; 9 — хромосомная микротрубочка; 10 —
дезинтегрирование ядерной оболочки; 11— хромосома; 12— межполюсная микротрубочка;
13 — полярные отверстия; 14 — оболочка дочерних ядер, образованная вокруг телофазных
ядер; 75— плазмалемма; 16 — плазмодесма; 17— клеточная стенка
8.
Цитокинез у большинства бурых водорослей
происходит путем впячивания мембраны. У Fucus
и Ascophyllum в центре клетки при делении
формируется пластинка, которая растет от
центра к периферии, как у зеленых растений.
Плазмодесмы (рис. 279, Д) встречаются,
вероятно, в поперечных перегородках всех бурых
водорослей, даже у тех, у которых перегородка
образуется за счет впячивания мембраны.
Через плазмодесмы осуществляется связь
цитоплазмы соседних клеток.
9. Отдел Бурые водоросли, Феофиты – ближайшие родственники диатомей
многоклеточная организация бурых водорослей варьирует от нитчатой до
талломной с хорошо развитой дифференцировкой клеток, у высших
представителей превращается истинную тканевую
при размножении образуются подвижные клетки (зооспоры или гаметы),
грушевидные или почковидные, несущие на боковой стороне два жгутика
переходная спираль в основании жгутика отсутствует
• к пигментному составу хризофитов добавляется -каротин;
•запасные
углеводы - -1,3 полигликан ламинарин и сахароспирт маннитол.
клеточная стенка всегда двухслойная, внутри целлюлозная, а снаружи
пектиновая с включением сильно ослизняющих ее веществ – фукоидана,
альгиновой кислоты и её солей
Жизненный цикл включает гаплоидную и диплоидную фазы,
чередующиеся правильно или неправильно, одинаковые или
морфологически различные
10. Подавляющее число бурых водоросдей – прикреплённые макрофиты которые обитают в основном в верхней сублиторали (до глубины 15 м)
холодныхморей.
Средообразующая роль бурых водорослей на литорали (побережье
Калифорнии). За исключением Corallina основные макрофиты – бурые
водоросли. По Woods Lytgoe из Casper, 1974.
11. Талломы
многоклеточная организация бурых водорослей варьирует от нитчатой до
талломной с хорошо развитой дифференцировкой клеток, у высших
представителей превращается истинную тканевую
• форма самая разнообразная: стелющиеся или
вертикально стоящие нити, корочки, пластинки
(простые или рассеченные), мешки, ветвящиеся
кустики.
• размеры талломы могут быть микроскопическими
или достигать нескольких десятков метров
(например, у Macrocystis, Nereocystis).
• Прикрепление талломов осуществляется с
помощью ризоидов или подошвы.
• Для удержания в вертикальном положении у ряда
бурых водорослей образуются воздушные пузыри,
заполненные газом.
12.
Рисунок 5. Бурые водоросли. Ламинария.13. Продольный разрез слоевища ламинарии (Laminaria)
14.
Laminaria. Продольный разрез молодого слоевища:1 — при малом увеличении; 2 — при большом увеличении:
а — трубчатые нити.
Меристодерма (делящаяся покровная ткань); Кора (ассимиляционная ткань)
Промежуточный слой (запасающая ткань);
Сердцевина (проводящая и механическая ткань) (трубчатые нити и ситовидные
пластинки и тонкие нити с толстыми продольными оболочками, соответственно)
15. У ламинариевых и фукусовых
• кора имеет значительную толщину и состоит из двух слоев:меристодермы и коры;
• а сердцевина делится на
промежуточный слой и собственно сердцевину.
В сердцевине образуются ситовидные трубки и трубчатые
нити. Сердцевина выполняет не только транспортную
функцию, но и механическую, так как в ней находятся нити
с толстыми продольными стенками. Между корой и
сердцевиной у многих бурых водорослей может находиться
промежуточный слой из крупных бесцветных клеток.
16. Окрашенная часть таллома - кора
• Меристодерма – делящаяся покровнаяткань. Состоит из 4-х слоев мелких клеток,
вытянутых по направлению к поверхности.
Они способны активно делиться и
формировать волоски и органы
размножения.
• Собственно кора – состоит из более крупных
окрашенных клеток, выполняет функцию
фотосинтеза.
17. Бесцветная часть таллома
• Промежуточный слой – крупные бесцветныеклетки, выполняющие запасающую
функцию.
• Сердцевина – рыхло или плотно
расположенные нити с сильно вытянутыми
клетками: ситовидные трубки и трубчатые
нити с расширенными воронковидными
концами клеток – для транспорта продуктов
фотосинтеза; тонкие нити с толстыми
продольными оболочками – механическая
функция.
18.
Laminaria (Л.Л. Великанови др., 1991):
Меристодерма (делящаяся покровная
ткань)
Кора (ассимиляционная ткань)
Промежуточный слой (запасающая ткань)
Сердцевина
(проводящая и механическая ткань)
трубчатые нити и ситовидные пластинки
А — продольный разрез черешка; Б — поперечный разрез черешка; В
— разрез листовой пластинки с сорусами зооспорангиев
19.
Laminaria.A — продольный
разрез черешка;
Б — поперечный
разрез черешка;
В- Г—разрез
листовой пластинки
с сорусом
спорангиев
20. Размножение
вегетативное, бесполое и половое.• Вегетативное размножение осуществляется
специализированными участками таллома и
выводковыми почками.
• Бесполое размножение происходит при помощи
двужгутиковых зооспор или неподвижных апланоспор
(тетраспор).
• Половой процесс – изо-, гетеро-, и оогамия.
• У преобладающего большинства видов цикл
воспроизведения со сменой поколений.
• Жизненный цикл включает гаплоидную и диплоидную
фазы,
чередующиеся правильно или неправильно,
одинаковые или морфологически различные. У
фукусовых – преобладает диплоидная фаза.
21.
• Схемажизненного цикла
представителей
рода
• ламинарии
(Laminaria)
22. Класс Фэозооспоровые PHAEOZOOSPOROPHYCEAE
• Водоросли разнообразной величины, формы истроения: от микроскопических слабо
разветвленных однорядных нитей до крупных
паренхиматозных талломов.
• В цикле развития обычно наблюдается
чередование двух самостоятельно живущих форм
или поколений – спорофита и гаметофита. У
примитивных форм строгая смена поколений отсутствует,
т.к. и спорофит и гаметофит способны к
самовоспроизведению.
• Преимущественно морские водоросли. В пресных
водах умеренных широт обнаружено 5 видов.
23.
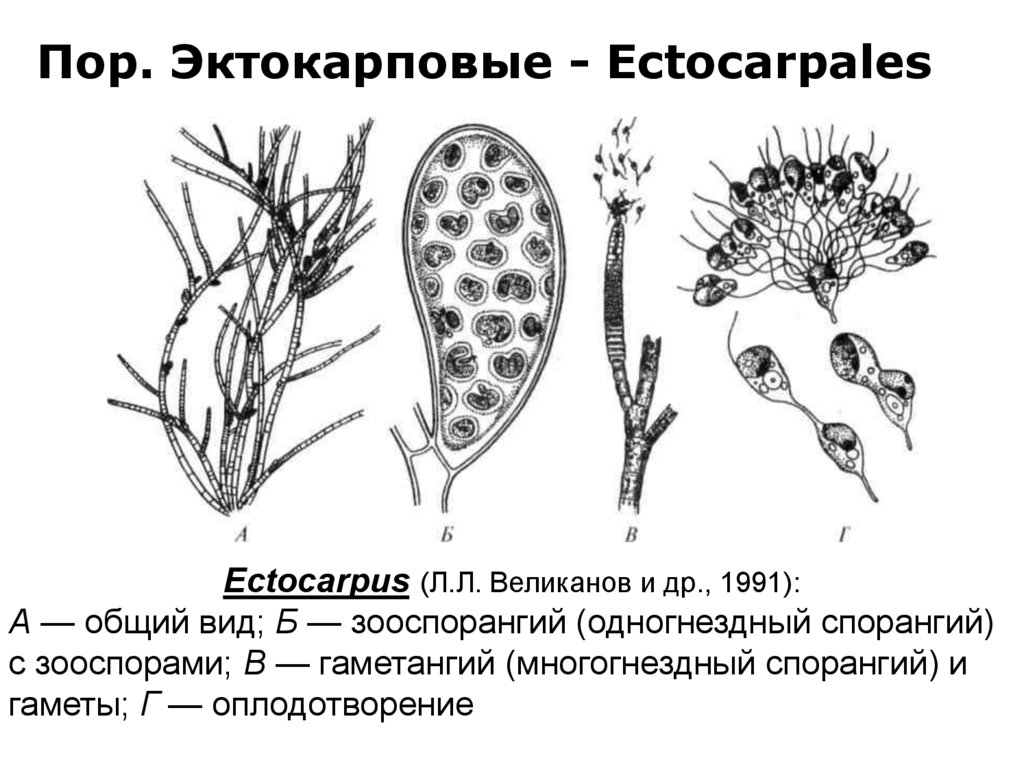
Пор. Эктокарповые - EctocarpalesEctocarpus (Л.Л. Великанов и др., 1991):
А — общий вид; Б — зооспорангий (одногнездный спорангий)
с зооспорами; В — гаметангий (многогнездный спорангий) и
гаметы; Г — оплодотворение
24.
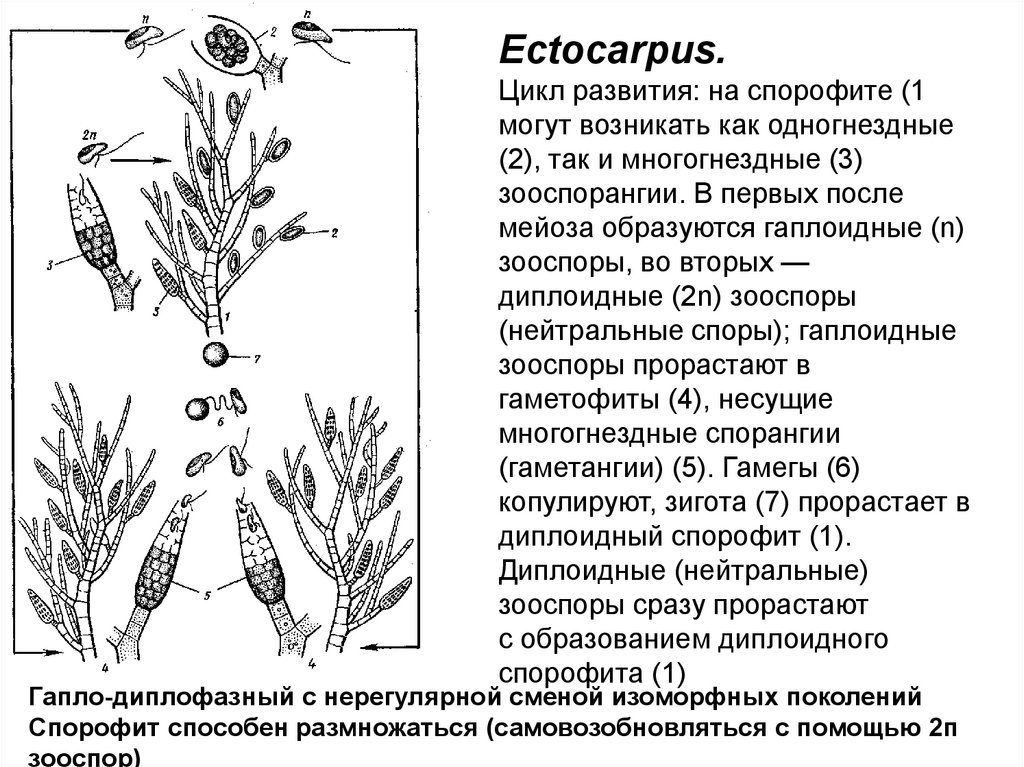
Ectocarpus.Цикл развития: на спорофите (1
могут возникать как одногнездные
(2), так и многогнездные (3)
зооспорангии. В первых после
мейоза образуются гаплоидные (n)
зооспоры, во вторых —
диплоидные (2n) зооспоры
(нейтральные споры); гаплоидные
зооспоры прорастают в
гаметофиты (4), несущие
многогнездные спорангии
(гаметангии) (5). Гамегы (6)
копулируют, зигота (7) прорастает в
диплоидный спорофит (1).
Диплоидные (нейтральные)
зооспоры сразу прорастают
с образованием диплоидного
спорофита (1)
Гапло-диплофазный с нерегулярной сменой изоморфных поколений
Спорофит способен размножаться (самовозобновляться с помощью 2п
зооспор)
25.
Возможныежизненные циклы у
Ectocarpus
(Н.П.Горбунова, 1991):
о — одногнездные
спорангии;
м — многогнездные
спорангии;
р! — редукционное
деление
26.
Пор. Ламинаривые - Laminariales27.
Рисунок 5. Бурые водоросли. Ламинария.28.
Laminaria.A — продольный
разрез черешка;
Б — поперечный
разрез черешка;
В- Г—разрез
листовой пластинки
с сорусом
спорангиев
29.
Жизненный цикл ламинарии (Laminaria sp.)1 – слоевище со спорами: а – пятно спорангиев, 2 – плавающие зооспоры,
3 – зооспора прикрепилась передним жгутиком к грунту, 4 – эмбриоспоры с
проростоковой трубкой, 6 – образование первых клеток гаметофитов (слева –
женский, справа – мужской), 7 – мужской гаметофит: б – антеридий,
в – антерозоиды, 8 – многоклеточный женский гаметофит, 9 – одноклеточный
женский гаметофит после выхода яйцеклетки из оогония: г – оболочка
эмбриоспоры, 10–11 – начало развития слоевища спорофита: д – первичный
30.
Laminaria. Цикл развития(Л.Л.Великанов и др., 1991).
На спорофите (1) развиваются
сорусы зооспорангиев (2), в
которых после редукционного
деления образуются зооспоры (3).
Они вырастают в
микроскопические гаметофиты (4)
— мужские с антеридиями (5) и
женские с оогониями (6).
Оплодотворенная сперматозоидом
(7) яйцеклетка (8) немедленно
прорастает с образованием
спорофита
31.
Циклразвития
ламинариевых
Гаплодиплофазный с
гетероморфной
сменой поколений
и преобладанием в
цикле развития
спорофита
32. Класс Циклоспорвые Cyclosporophyceae
• Талломкрупный,
дифференцирован
на
меристодерму, кору, промежуточный слой и
сердцевину.
• В цикле развития отсутствуют две самостоятельно
растущие
формы
развития.
Гаметангии
развиваются в концептакулах на слое клеток,
возникающем из одной инициальной клетки,
называемой проспорой.
• Мейоз происходит при образовании гамет. Цикл
развития диплофазный.
• Половой процесс у большинства – оогамия.
• Морские водоросли.
33.
Пор. Фукусовые - Fucalesслева — фукус пузырчатый (Fucus vesiculosus);
справа — фукус двусторонний (F. distichus)
Ремневидный дихотомически ветвящийся таллом (пластинчатый)
34. Пор. Фукусовые - Fucales
На снимке показаны бурые водоросли.IGDA/P. Donnini
35.
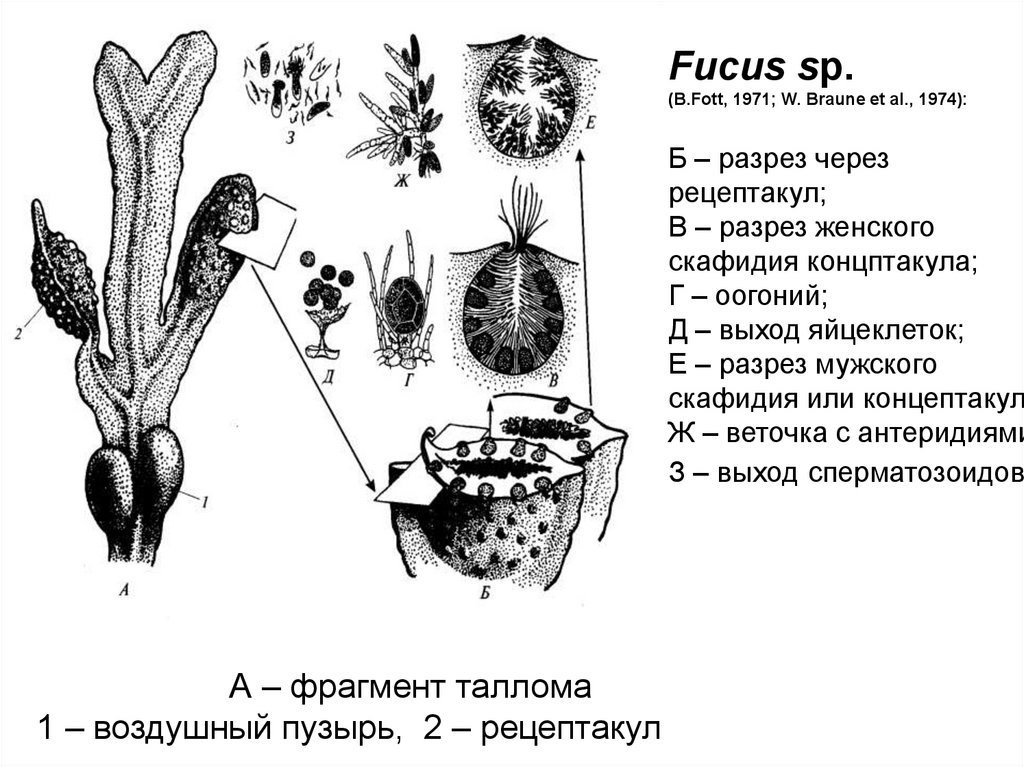
Fucus sp.(B.Fott, 1971; W. Braune et al., 1974):
Б – разрез через
рецептакул;
В – разрез женского
скафидия концптакула;
Г – оогоний;
Д – выход яйцеклеток;
Е – разрез мужского
скафидия или концептакул
Ж – веточка с антеридиями
З – выход сперматозоидов
А – фрагмент таллома
1 – воздушный пузырь, 2 – рецептакул
36.
Цикл развития Fucus sp.А – взрослый таллом, Б – рецептакул, В – скафидий (концептакул) на срезе,
Г – оогоний, Д – образование яйцеклеток, Е – выход яйцеклеток,
Ж – антеридий и образование сперматозоидов,
З – копуляция яйцеклетки и сперматозоида, И – зигота,
К, Л – образование диплоидного таллома.
37.
Циклразвития
фукусовых
Диплофазный
Мейоз
гаметический
38.
Виды Cystoseira (1-2), Sargassum (3)39.
Саргасум достигает длины 2-8 м, обладает коротким стволом, длиннымиветвями, листовые пластинки с ребром. В саргассовом море обитают
неприкрепленные виды с плавающим и погруженным талломом.
Рисунок 8. Саргассум
40. Филогения
• Бурые водоросли – очень древние организмы.• Их окаменелости известны в силурских и
девонских пластах палеозойской эры.
• Достоверные остатки ископаемых бурых
водорослей обнаружены в триасовых отложениях
мезозойской эры.
• Молекулярные методы определяют возраст бурых
водорослей как 155 — 200 млн лет.
41. Происхождение
• Бурые водоросли, вероятно, произошли откаких-то первичных жгутиковых, у которых
преобладали бурые пигменты.
• Однако связь их со жгутиковыми более
отдаленная, чем у сходных с ними
золотистых и желтозеленых, так как прямые
переходные формы (монадные, коккоидные,
трихальные) у них отсутствуют.
42. Эволюция
• Бурые — монофилетичная группа, но связи внутринее до конца не понятны.
• В эволюции бурых водорослей, вероятно,
произошел переход от интеркалярного роста
(эктокарповые, кутлериевые, ламинариевые) к
верхушечному (сфацеляриевые, диктиотовые,
фукусовые).
• Согласно второй схеме филогенеза, общие предки
бурых водорослей дали три эволюционные ветви,
которые отличаются циклами развития и
морфологическим строением талломов (классы
изогенератные, гетерогенератные и
циклоспоровые).
43.
Бурые водоросли (Phaeophyta)Бурые водоросли имеют крупные многоклеточные талломы.
Обитают в теплых и холодных морях. Наиболее распространены в
морях умеренных и полярных широт.
Талломы бурых водорослей имеют форму нитей, пластин,
корочек, крупные виды напоминают листостебельные растения и
имеют примитивные ткани.
Рост таллома осуществляется
интеркалярно или апикально
Клеточная оболочка содержит целлюлозу, пектиновые вещества,
альгины и камедиеподобные вещества.
Протопласт клетки содержит одно ядро, и хлоропласты. Окраска
бурых водорослей обусловлена преобладанием ксантинов: фуко-,
виоло и антероксантина и каротина над хлорофиллом. Запасные
вещества: ламинарин, маннит, танины и жиры.
44.
Размножение вегетативное, бесполое и половое.Вегетативное размножение осуществляется специализированными
участками таллома и выводковыми почками.
Бесполое размножение происходит при помощи двужгутиковых зооспор
или неподвижных апланоспор (тетраспор).
Половой процесс – изо-, гетеро-, и оогамия. У преобладающего
большинства видов цикл воспроизведения со сменой поколений.
Бурые водоросли служат местообитанием, источником пищи и укрытием
для морской фауны. Ч.Дарвин назвал их тропическими лесами в морях.
В хозяйственной деятельности бурые водоросли служат единственным
источником альгинатов, которые используют как стабилизаторы при
консервировании. Альгинаты также применяют для улучшения свойств
смазывающих, красящих и клеящих материалов, в книгопечатании,
производстве тканей, пластмасс. Альгинат натрия придает стойкость
лакокрасящим покрытиям и строительным материалам. Из него готовят
фармацевтические и парфюмерные пасты. Бурые водоросли используют в
медицине: они содержат йод, бромистые соли, витамины; а также в качестве
заменителей крови. Бурые водоросли выводят из организма радиоактивные
вещества.
45.
Ламинария (Laminaria) обитает в холодных водах южного и северногополушария, где образует заросли в полосе сублиторали (до глубины 20 м).
Талломы ламинарии имеют вид листовой пластинки размером до 2 м и более,
прикрепленной к плотному черешку с когтевидными мощными ризоидами,
которыми водоросль прикрепляется к подводным камням.
Таллом ламинарии многолетний: зимуют ризоиды и черешок, а листовая
пластинка ежегодно сбрасывается осенью и отрастает весной за счет
интеркалярной меристемы, расположенной на верхушке черешка.
Зооспорангии образуются у основания листовидной пластины на ее
поверхности большими группами – сорусами. Спорообразование происходит
летом. Зооспоры с перистым и бичевидным жгутиками сначала плавают, затем
теряют активность, прикрепляются к подводным предметам и дают начало
микроскопическим нитевидным гаметофитам – заросткам. На женском
гаметофите образуются оогонии с одной яйцеклеткой, которая при созревании
выходит, но не отделяется от оогония. Оплодотворенная яйцеклетка – зигота или
ооспора, без периода покоя прорастает в новое растение ламинарии (спорофит).
В анатомическом строении выделяют: меристодерму, кору, промежуточный
слой и сердцевину. Меристодерма образует покровную ткань, в коре
располагается ассимиляционная ткань, в промежуточном неокрашенном слое –
запасающая ткань, в сердцевине – проводящая и механическая ткань.
Ламинария японская или морская капуста самый промысловый продукт
среди бурых водорослей.
46.
Фукусы (Fucus). Виды фукуса распространены у берегов холодных иумеренных морей северного полушария.
Таллом Фукуса имеет вид дихотомически разветвленного бурого кустика,
длиной от 30-40 см до 2 м. Ветви таллома плоские, кожистые, со средней
жилкой. У большинства видов на них есть воздушные пузыри.
Основание таллома имеет вид округлого стволика с конической подошвой,
которой он прикрепляется к субстрату.
Под покровами таллома располагаются полости с волосками, служащие
для поглощения питательных веществ. По бокам от «средней жилки»
формируются воздушные пузыри, удерживающие таллом на поверхности воды.
Слоевище дифференцировано на меристодерму, кору с многочисленными
хлоропластами, глубже – промежуточный слой, а в центре сердцевина с
нитевидными клетками, выполняющими функцию проводящей ткани.
Бесполого размножения нет. Половой процесс – оогамный. Половые
органы находятся в особых вместилищах – скафидиях, или концептакулах,
которые располагаются под корой и открываются наружу отверстиями. От
стенок скафидиев в полость отходят парафизы, между которыми формируются
половые органы. В оогониях образуются яйцеклетки, а в антеридиях –
сперматозоиды. Оплодотворение происходит в воде. Зигота сразу прорастает,
образуя диплоидный таллом фукуса. Следовательно, у фукуса смена
поколений отсутствует.
47. Красные водоросли Отдел Rodophyta Царство Rodobionta
В отделе около 4000 видов.Преимущественно морские в тропических морях .
В пресных и солоноватых континентальных водоемах – 50
Большинство многоклеточные, есть 1 клеточные коккоидные.
В оболочке агар
Имеют красные и синие пигменты (фикобилины)
Запасают багрянковый крахмал
Нет подвижных стадий
В цикле развития 3 поколения
Название происходит от греческого слова rhodon – розовый.
Окраска таллома от светло-розовой, красной, фиолетовой, желтой до
голубовато-зеленой.
класс Bangiophyceae
класс Florideophyceae
48.
Их многоклеточные слоевища имеют вид корки, пленки,шнура,
кустика,
пластинки,
кораллов;
известны
и
одноклеточные формы. Очень крупных форм нет, лишь у
отдельных видов талломы достигают 1,5 – 2 м. Есть
микроскопические виды 1 клеточные коккоидные
49.
Красные водоросли (багрянки)Бягрянки – преимущественно морские водоросли, произрастающие в зоне
тропиков, самые глубоководные, достигают глубины 100 – 200 м, реже
встречаются в пресных водоемах.
Их многоклеточные слоевища имеют вид корки, пленки, шнура, кустика,
пластинки, кораллов; известны и одноклеточные формы. Очень крупных форм нет,
лишь у отдельных видов талломы достигают 1,5 – 2 м.
Окраска таллома от светло-розовой, красной, фиолетовой, желтой до
голубовато-зеленой.
Окраску обуславливают хлорофиллы a и d каротин, ксантофилл, фикоэритрин
и фикоцианин. Теневыносливы, улавливают даже незначительный свет.
Оболочки клеток состоят из целлюлозы и пектиновых веществ, способны
сильно ослизняться, у некоторых видов пропитаны карбонатами кальция и магния
и тогда таллом становится твердым, напоминающим кораллы. Цитоплазма вязкая,
чувствительная к солености, центральная вакуоль. Ядро в клетке одно, реже –
несколько. Хлоропласты дисковидной, пластинчатой, реже звездчатой формы.
Запасные вещества – багрянковый крахмал и масло.
50. Подавляющее число бурых водоросдей – прикреплённые макрофиты которые обитают в основном в верхней сублиторали (до глубины 15 м)
холодных морей.Средообразующая роль бурых водорослей на литорали (побережье
Калифорнии). За исключением Corallina основные макрофиты – бурые
водоросли. По Woods Lytgoe из Casper, 1974.
51.
https://cf2.ppt-online.org/files2/slide/x/XdRTp5DuV6AKk7vWMQsxU8haCiY0fgeyItrqNEnLG/slide-2.jpg52.
ПигментыХлорофиллы
Хл. а, Хл. б,
Хл. с, Хл. д
Каротиноиды
Каротины:
α-каротин, β-, γ-, ε-
Ксантофиллы:
фукоксантин,
зеаксантин,
диатоксантин и др.
Фикобилины
фикоэритрин,
фикоцианин,
аллофикоцианин
53.
54.
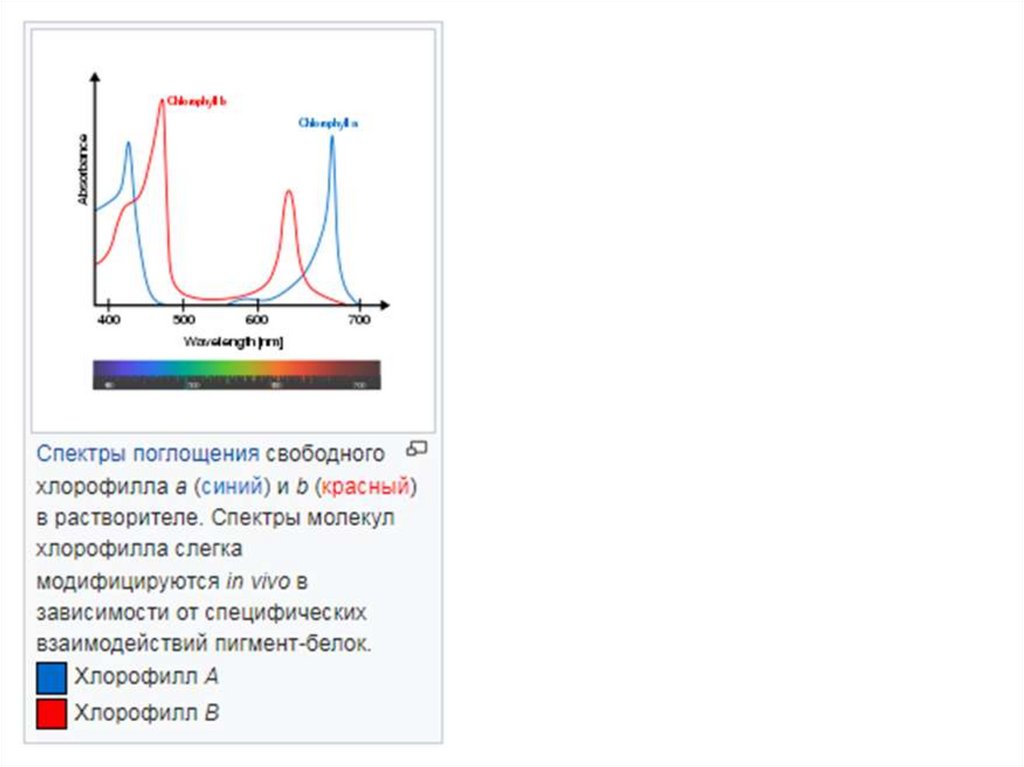
Хлорофиллы наиболее сильно поглощают свет в синей части электромагнитного спектра, а также вкрасной части.[4] И наоборот, он плохо поглощает зеленую и почти зеленую части
спектра. Следовательно, ткани, содержащие хлорофилл, выглядят зелеными, потому что зеленый
свет, рассеянно отраженный структурами, такими как клеточные стенки, поглощается меньше. [1] В
фотосистемах зеленых растений существуют два типа хлорофилла: хлорофилл а и в[5].
https://translated.turbopages.org/proxy_u/en-ru.ru.ef38695d-6362dec2-c0cfa6dc-74722d776562/https/en.wikipedia.org/wiki/Leafgreen
Известно более 1100 каротиноидов[8], которые можно разделить на два класса: ксантофиллы (которые
содержат кислород) и каротины (которые являются чистыми углеводородами и не содержат
кислорода).[1] Все они являются производными тетратерпенов, что означает, что они образуются из 8
молекул изопрена и содержат 40 атомов углерода. Как правило, каротиноиды поглощают волны
длиной от 400 до 550 нанометров (от фиолетового до зеленого света). Это приводит к тому, что
соединения приобретают насыщенный желтый, оранжевый или красный цвет.
https://translated.turbopages.org/proxy_u/en-ru.ru.80a11574-6362dc4b-a51157ae-74722d776562/https/en.wikipedia.org/wiki/Carotenoid
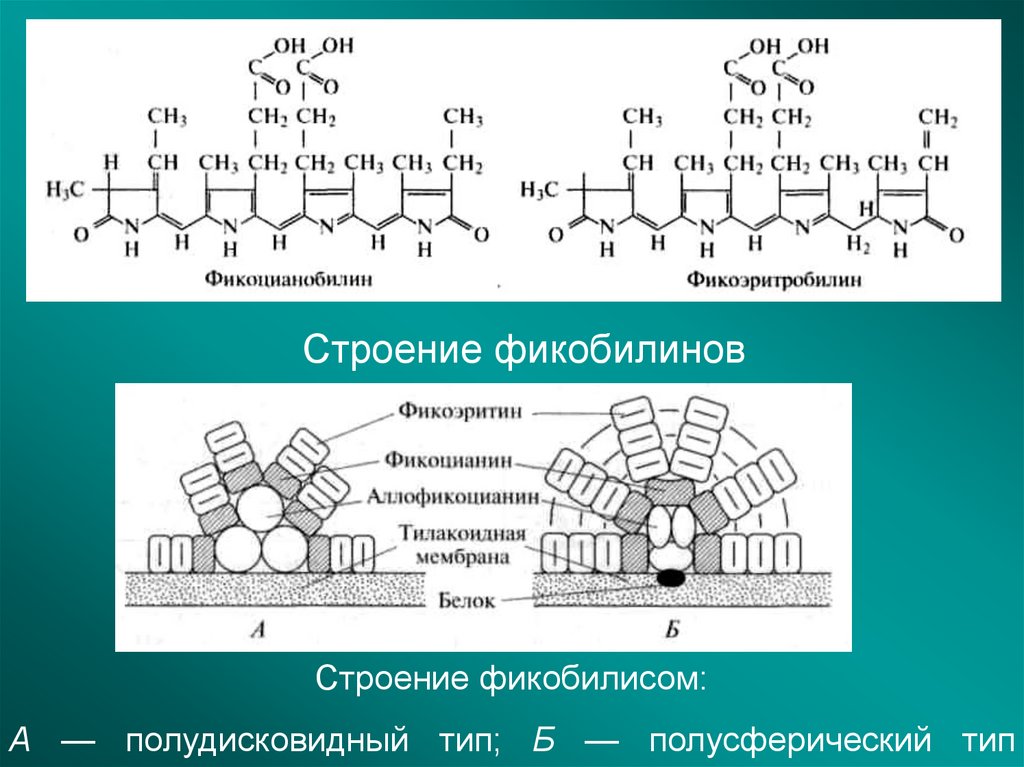
Фикобилины (от греческого: φύκος (phykos), что означает "водоросль", и от латинского: bilis, что
означает "желчь") - это улавливающие свет билины, обнаруженные в цианобактериях и
в хлоропластах красных водорослей, глаукофитов и некоторых криптомонад (хотя и не в зеленых
водорослях и растениях). Большая часть их молекул состоит из хромофора, который придает им
цвет. Они уникальны среди фотосинтетических пигментов тем, что они связаны с определенными
водорастворимыми белками, известными как фикобилипротеины. Фикобилипротеины затем передают
световую энергию хлорофиллам для фотосинтеза.
Фикобилины особенно эффективны при поглощении красного, оранжевого, желтого и зеленого света,
длины волн, которые плохо поглощаются хлорофиллом А. Организмы, растущие на мелководье, как
правило, содержат фикобилины, которые могут улавливать желтый / красный свет, в то время как
организмы на большей глубине часто содержат больше фикобилинов, которые могут улавливать
зеленый свет, который там относительно более распространен.
Фикобилины флуоресцируют на определенной длине волны и поэтому часто используются в
исследованиях в качестве химических меток, например, путем связывания фикобилипротеинов
с антителами методом, известным как иммунофлуоресценция.
https://translated.turbopages.org/proxy_u/en-ru.ru.81726cba-6362e091-11a3fdd7-74722d776562/https/en.wikipedia.org/wiki/Phycobili
55.
ПигментыХлорофиллы
Хл. а, Хл. б,
Хл. с, Хл. д
Каротиноиды
Каротины:
α-каротин, β-, γ-, ε-
Ксантофиллы:
фукоксантин,
зеаксантин,
диатоксантин и др.
Фикобилины
фикоэритрин,
фикоцианин,
аллофикоцианин
Окраска клеток от светло-желтой до темно-бурой за счет преобладания
фукоксантина.
(Фукоксантина нет у желтозеленых водорослей)
56.
ПигментыХлорофиллы
Хл. а, Хл. б,
Хл. с, Хл. д
Каротиноиды
Каротины:
α-каротин, β-, γ-, ε-
Ксантофиллы:
фукоксантин,
зеаксантин,
диатоксантин и др.
Фикобилины
фикоэритрин,
фикоцианин,
аллофикоцианин
57.
ПигментыХлорофиллы
Хл. а, Хл. б,
Хл. с, Хл. д
Каротиноиды
Каротины:
α-каротин, β-, γ-, ε-
Ксантофиллы:
фукоксантин,
зеаксантин,
диатоксантин и др.
лютеин,
антероксантин,
виолаксантин и др.
Фикобилины
фикоэритрин,
фикоцианин,
аллофикоцианин
58.
Строение фикобилиновСтроение фикобилисом:
А — полудисковидный тип; Б — полусферический тип
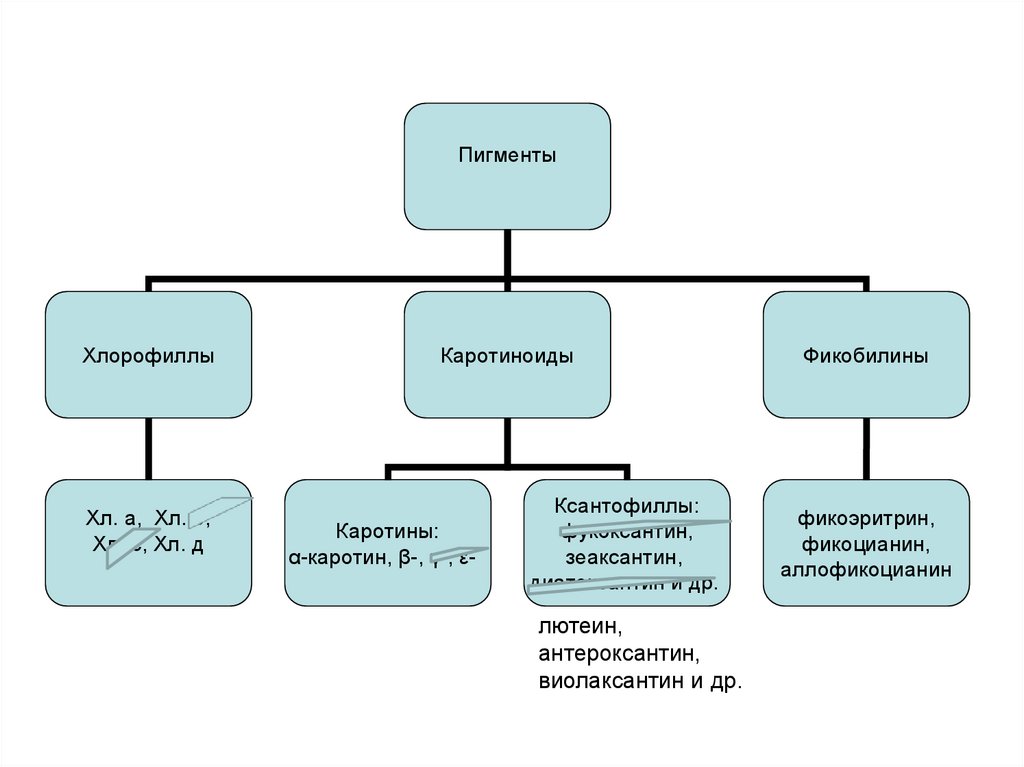
59. Пигменты
• хлорофиллы a и d• каротиноиды:
каротины α, β;
ксантофиллы: лютеин, зеаксантин, антероксантин,
виолаксантин и др.
• фикобилины
Фикоэритрин, фикоцианин, аллофикоцианин.
Теневыносливы, улавливают даже незначительный
коротковолновой свет (синий, зеленый, фиолетовый).
«Хроматическая адаптация»
(явление, представляющее собой приспособительное изменение окраски
водорослей под влиянием изменения спектрального состава света за счет
увеличения количества пигментов, имеющих окраску, дополнительную к цвету
падающих лучей. Характерно для цианопрокариот и красных водорослей
(Rhodophyta).
60.
61.
62.
63.
64.
1 — порфира (Porphyra variegata);2 — дюмонтия (Dumontia incrassata);
3 — эутора (Euthora cristata);
4 — хондрус (Chondrus yendoi).
1 — ломентария (Lomentaria articulata);
2 — хондрус (Chondrus pinnulatus);
3 — хризимения (Chryzymenia wrightii);
4 – родимения (Rhodimenia pertusa
Пластинки, шнуры, кустики, сильно рассеченные
65.
1 — одонталия (Odonthalia);2 — токидодендрон (Tokidodendron);
3 — родомела (Rhodomela);
4 — дазия (Dasia)
66.
членистыекораллиновые
водоросли
Corallina
https://ce63106.tmweb.ru/upload/resize_cache/iblock/af6/640_480_025be301
23f8603dbb1b476c4fe38b5a8/af621759f4561f0a120082400c8571ec.jpg
корковые
кораллиновые
водоросли
Lithothamnion
https://live.staticflickr.com/2562/5769153550_ee
906b489d_b.jpg
67. Типы морфологической дифференциации таллома
Коккоидный ипальмеллоидный
Нитчатый
68. Типы морфологической дифференциации таллома
РазнонитчатыйПластинчатый
69. Типы морфологической дифференциации таллома
Эволюция структуры флоридеевых:1—Kulinia;
2— часть слоевища Batrachospermum;
з — продольный срез через слоевище Lemanea: а — моноспоры, б — ризоиды,
в — гонимобласт, г — центральные клетки, д — перицентральная клетка.
70. Типы морфологической дифференциации таллома Псевдопаренхиматозный с одноосевым типом строения
Таллом красных водорослей содноосевым типом строения
Слева Batrachospermum:
a — инициальная клетка;
б — клетки центральной нити;
в — кольца веточек ограниченного
роста;
г — образование побегов
междоузлий;
д — нити коры на междоузлиях
Справа Таллом Lemanea:
1—в продольном разрезе;
2 — в поперечном разрезе;
а — клетки осевой нити;
б — крестообразно расположенные
клетки;
в — боковые нити;
г — карпогон;
д — трихогина;
е — гонимобласт;
ж — подкоровые клетки;
з — коровые клетки
71. Типы морфологической дифференциации таллома Псевдопаренхиматозный с многоосевым типом строения
многоосевой (фонтанный) типстроения в продольном
разрезе (Furcellaria fastigiata)
Псевдопаренхиматозный тип
строения в продольном разрезе
(Thorea ramosissima)
72. Клетки красных водорослей
целлюлоза ипектиновые вещества
агар,
каррагинан,
агароза).
(
Таллом напоминает хрящ.
Оболочки клеток
красных водорослей
(видна слоистость оболочки)
У некоторых видов в
оболочках откладывается
карбонат кальция и магния.
Таллом становится твердым,
напоминает кораллы или
окаменелости.
73.
Клетки красных водорослей:митохондрии с плоскими кристами
Рис. 22. Различные типы крист (к) митохондрий
(С.А.Карпов, 2001; О. Г.Кусакин, А.Л.Дроздов, 1994):
А — пластинчатые; Б — трубчатые; В —
дисковидные
74. Клетки красных водорослей: хлоропласты
тилакоиды расположены поодному, отсутствует
хлоропластная ЭПС
1 — рибосомы;
2 — оболочка хлоропласта;
3 — опоясывающий тилакоид;
4 — ДНК;
5 — фикобилисомы;
6 — крахмал;
Хлоропласты красных водорослей:
a — хлоропласты; б —- пиреноиды;
в — ядра
Хлоропласты дисковидной, пластинчатой,
реже звездчатой формы..
75.
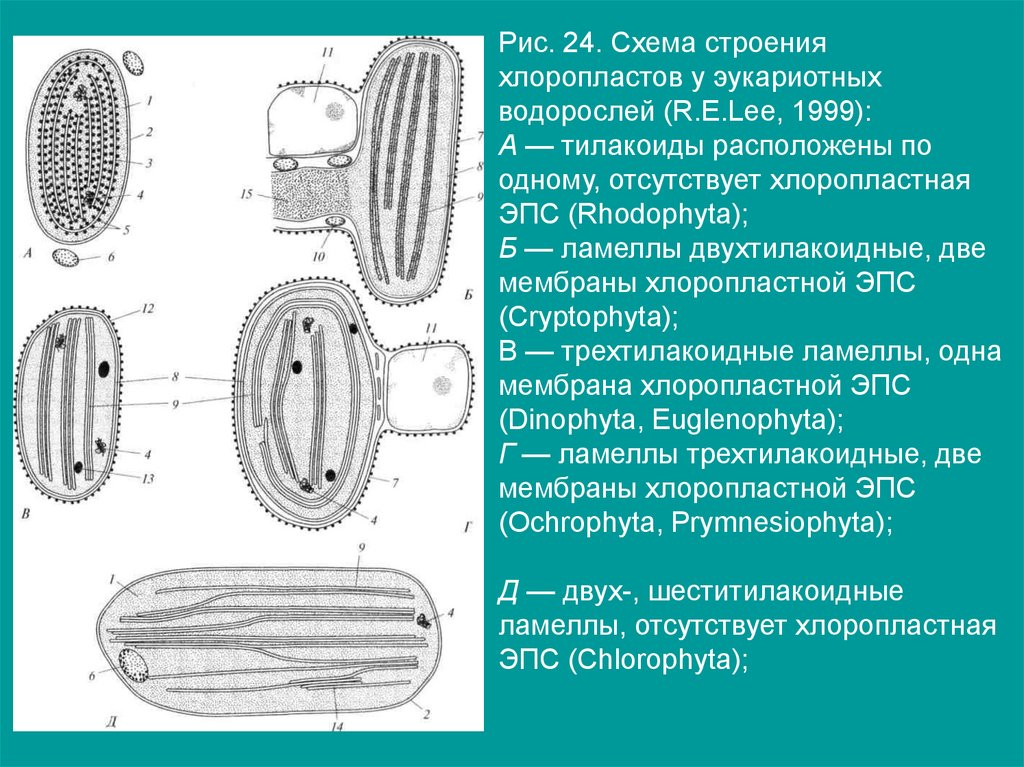
Рис. 24. Схема строенияхлоропластов у эукариотных
водорослей (R.E.Lee, 1999):
А — тилакоиды расположены по
одному, отсутствует хлоропластная
ЭПС (Rhodophyta);
Б — ламеллы двухтилакоидные, две
мембраны хлоропластной ЭПС
(Cryptophyta);
В — трехтилакоидные ламеллы, одна
мембрана хлоропластной ЭПС
(Dinophyta, Euglenophyta);
Г — ламеллы трехтилакоидные, две
мембраны хлоропластной ЭПС
(Ochrophyta, Prymnesiophyta);
Д — двух-, шеститилакоидные
ламеллы, отсутствует хлоропластная
ЭПС (Chlorophyta);
76. Клетки красных водорослей
Запасные вещества – в цитоплазмебагрянковый крахмал и масло.
Зерна багрянкового
крахмала
(3 – в поляризованном свете)
Железистые клетки (а)
красных водорослей
77. Клетки красных водорослей
Митоз полузакрытый, без центриолей.Цитокинез с помощью кольцевой борозды.
Схема образования первичных (1-3) и вторичных (4-8) пор в
клетках талломов красных водорослей
78. Клетки красных водорослей
Иридирующие тельца в клетках:a — крупные иридирующие тельца;
б — хлоропласты;
в — мелкие иридирующие тельца
Стадии развития (1—4) волоска
у Cystoclonium purpureum
79. Размножение красных водорослей
Моноспорангии(а) флоридеевых
водорослей:
1 — Chantransia
pygmaea;
2 — Chantransia
sinensis;
3 — Balbiania
investiens на
междоузлии (б)
Batrachospermum
4 — Thorea
ramosissima
80.
Споры и гаметы не имеют жгутиков.Половой процесс оогамный, но протекает специфически. Женский половой
орган карпогон, состоящий из расширенной части – брюшка и нитевидной –
трихогины. В брюшке карпогона образуется одна яйцеклетка. В антеридиях,
собранных группами, образуются неподвижные половые элементы –
спермации. Переносимые водой спермации попадают через трихогину в
карпогон и затем сливаются с яйцеклеткой. После оплодотворения из него
вырастают гонимобластные нити со спорангиями на концах, в которых
формируются карпоспоры, а группы карпоспор образуют цистокарпии.
Смена ядерных фаз происходит у красных водорослей по-разному: цикл
полового воспроизведения может состоять из чередования самостоятельно
живущих гаметофита и спорофита или включать две диплоидные фазы –
карпоспорофита и тетраспорофита, из которых первый развивается на
гаметофите.
Из красных водорослей агарофитов добывают агар, карраген и агароиды.
Агар применяют в микробиологии (как добавки в питательную среду), в
пищевой промышленности (для желе, мармеладов, консервирования мяса и
рыбы), в фармакологии для приготовления капсул. Многие водоросли
используют для кормовой муки и удобрений.
81. Размножение красных водорослей
Тетраспорангиифлоридеевых
водорослей:
/ — Callithamnion
corymbosum
2 — Cruoria pellita
3 — Corallina
mediterranea
4 — Peyssonnelia
dubia
5 — Gracilaria sp.:
a — выводное
отверстие;
б — концептакул;
в — нити нематеция
82. Размножение красных водорослей
Сперматангии (а)флоридеевых водорослей:
a – сперматангии;
б - антеридиальные нити;
в — базальные клетки,
г – сорус
1 — участок таллома
Batrachospermum;
2 — участок таллома Nemalion;
3, 4 — сперматангии Gelldiura sp.
в поперечном разрезе;
5 — нематеций;
6 — колосоьидный сорус
Polysiphonia sp.;
7 — сорус Lemanea
в поперечном разрезе
8— 11 — сорусы на талломах
Lemanea
83. Размножение красных водорослей
Стадииразвития (1—5) карпогона Batrachospermum:
а — ядро карпогона;
б — трихогина; в — спермий при
слиянии с трихогиной; г — слияние спермия и карпогона;
д — образование перегородки между трихогиной и карпогоном
84. Размножение красных водорослей
Стадииразвития
гонимобласта
Batrachospermum:
1 — 4 — первые клетки
гонимобласта,
образовавшиеся из
оплодотворенного
карпогона;
5 — молодой карпогон;
6 — карпогон с
карпоспорами;
а — стерильные веточки
карпогона;
б — карпоспора;
в — пустой
карпоспорангий
85. Размножение красных водорослей
Схема развития ообластемной нити (1) из карпогона (2) иее последовательное слияние с ауксилярными клетками (3)
с образованием гонимобласта (4) у рода Dumontia
86. Размножение красных водорослей
3 б типАуксилярные клетки
дифференцируются
только после
оплодотворения
аскогона вблизи от
него, т. е. имеется
прокарпий.
Callithamnion.
А —общий вид
таллома;
Б— прокарпий;
В — цистокарпии
87. Классификация
• Класс Бангиофициевые(Bangiophyceae)
88. Род порфиридиум (Porphyridium)
Наиболее примитивный представитель классабангиевых.
Клетки шаровидные одиночные или образующие
бесформенные слизистые скопления бурокрасного или сине-зеленого цвета.
Встречаются на влажных затененных стенах, в
прудах и т. п.
Клетки размножаются делением. В каждой клетке
со звездчатым хлоропластом и пиреноидом
возможно образование одной моноспоры.
89. Порядок: ПОРФИРИДИАЛЬНЫЕ (PORPHYRIDIALES) Семейство: ПОРФИРИДИЕВЫЕ (PORPHYRIDIACEAE)Вид: ПОРФИРИДИУМ БАГРЯНЫЙ (ПАРФIРЫДЫУМ
БАРВОВЫ)PORPHYRIDIUM PURPUREUM (BORY) DREW ET ROSS
Категория охраны: 3
Международная значимость:
Впервые включен в Красную книгу Беларуси.
Описание:
Колонии кроваво-красные или красно-бурые,
корковидные, до 10 см в диаметре,
неопределенных очертаний. Клетки в колониях
шаровидные, 7-15 мкм в диаметре, одетые
неслоистой слизистой обверткой, беспорядочно
расположенные в общей гомогенной слизи.
Хлоропласт осевой, звездчатый, с одним
центральным пиреноидом. Окраска определяется
количественным соотношением фикоэритрина и
фикоцианина .
Распространиение:
Распространен преимущественно в Европе, но
встречается спорадически. В республике отмечены
единичные находки в Мядельском и Минском р-нах
90.
Местообитания:
В наземных условиях: на поверхности почвы, камнях, деревянных крышах и
стенах зданий, на влажных затененных поверхностях, часто в присутствии
мочевины .
Биология:
Размножение вегетативное простым делением клетки на две или несколько
дочерних клеток, иногда акинетами. Дочерние клетки сразу после деления
расходятся или некоторое время остаются заключенными в материнскую
оболочку. Половое размножение отсутствует .
Численность и тенденция ее изменения:
Колонии встречаются небольшими группами в виде корковидных пленок
неопределенных очертаний.
Основные факторы угрозы:
Хозяйственная деятельность человека.
Меры охраны:
Необходимо выявление новых мест роста.
Составители: Михеева Т.М.
91. Porphyra – в северных и южных морях в прибрежной зоне листовидная пластинка, основанием прикрепляется к субстрату:
Длина –до 0,5 редко до 2 м.
Толщина –
1-2 слоя клеток.
В клетках –
1 звездчатый хлоропласт
В оболочке - фибриллы
– ксиланы (полимер
ксилозы); аморфная
фракция – маннаны
Окраска синеватозеленого цвета с
буроватым или пурпурнокрасным оттенком.
92.
Порфира (Porphyra) морская водоросль достигающая 20-30 см в длину.Имеет вид широкой, тонкой пластинки овальной, округлой или ланцетной
формы, с цельными либо лопастными, или надорванными краями. Короткой
ножкой пластинка прикрепляется к подводным предметам. Таллом состоит из
одного слоя клеток. Порфира обитает в Балтийском, Белом, Черном и
Средиземном морях. Окраска синевато-зеленого цвета с буроватым или
пурпурно-красным оттенком.
Бесполое размножение осуществляется моноспорами, возникающими из
вегетативных клеток.
Половые органы карпогоны и антеридии располагаются на талломе
группами. В антеридиях формируются спермации, которые переносятся водой
и попадают на карпогон. Карпогон содержит одну яйцеклетку. Зигота
претерпевает редукционное деление и дает начало карпоспорам, из которых
образуются новые особи.
В Японии порфира культивируется как морская с.-х. культура.
Батрахоспермум (Batrachospermum) - лягушечник, пресноводная
водоросль из группы багрянок, представляется в виде разветвленных
кустиков, окутанных бесцветной студенистой слизью, придающей им
отдаленное сходство с яйцами лягушек.
93. Porphyra - гаметофит
PorphyraA — внешний вид; Б — разрез таллома с антеридиями;
В — разрез таллома с карпогонами, содержимое которых после
оплодотворения делится с образованием карпоспор
94.
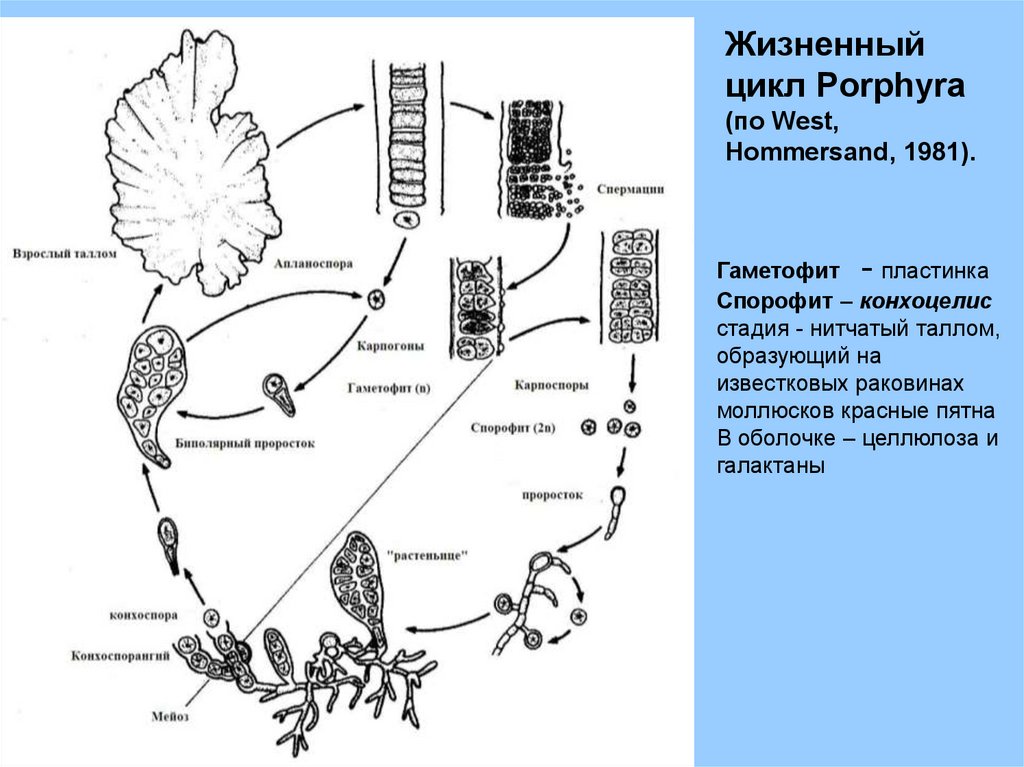
Жизненныйцикл Porphyra
(по West,
Hommersand, 1981).
Гаметофит - пластинка
Спорофит – конхоцелис
стадия - нитчатый таллом,
образующий на
известковых раковинах
моллюсков красные пятна
В оболочке – целлюлоза и
галактаны
95. Класс Флоридеофициевые (Florideophyceae)
• Порядок батрахоспермовые(Batrachospermales)
- гонимобласты развиваются из оплодотворенного карпогона,
нет ауксилярных клеток;
• - уникальный тип жизненного цикла - гаметофит вырастает на
диплоидном спорофите в результате соматической редукции в
его верхушечных клетках (соматическая редукция).
96. Разнообразие красных водорослей
Пресноводныефлоридеевые
водоросли
порядка Nemaliales:
1 — Chantransia
sinensia;
2 — Batrachospermum
moniliforme;
3 — Lemanea nodosa;
4 — Thorea
ramosissima
97. Batrachospermum - лягушечник
Batrachospermum лягушечникСильно разветвленный
студенисто-слизистый
кустик 3-8 см оливкового,
или стального цвета
(избыток фикоцианина)
Напоминает яйца
лягушек
В ручьях, реках, болотах,
озерах с чистой
прозрачной водой на
сучках, камнях
Кустики – из
побегов –
нитей с как бы
нанизанными
на них
бусинами
98. Общий вид батрахоспермума в р. Поплав
99. Batrachospermum
Под микроскопом: нить из 1 ряда крупныхбесцветных клеток. От границы двух соседних
клеток под поперечной перегородкой, т.е. от
верхней части клетки отрастает мутовка
боковых разветвленных веточек из коротких
мелких бочонковидных или каплевидных
клеток со многими хлоропластами –
ассимиляторы.
Из базальных клеток боковых ветвей
растут тонкие многоклеточные коровые нити
вдоль главной оси не плотно друг к другу,
формируя кору
На ассимиляторах формируются, антеридии и
карпогоны
Псевдопаренхиматозный таллом с
одноосевым типом строения
a — инициальная клетка;
б — клетки центральной нити;
в — кольца веточек ограниченного роста;
г — образование побегов междоузлий;
д — нити коры на междоузлиях
100. Batrachospermum
101. Разнообразие красных водорослей
Строение Batrachospermum:1— внешний вид части растения; 2 — ювенильная стадия (шантранзия); 3 —
начало образования взрослого слоевища; 4 — в — стадии развития
гонимобласта. а — моноспорангии, б — трихогина, в—оплодотворенный
карпогон, г — клетки гонимобласта, д — карпоспоры
102. Стадии развития (1—5) карпогона Batrachospermum:
а — ядро карпогона;б — трихогина; в — спермий при
слиянии с трихогиной; г — слияние спермия и карпогона;
д — образование перегородки между трихогиной и карпогоном
103.
Стадии развитиягонимобласта
Batrachospermum:
1 — 4 — первые клетки
гонимобласта,
образовавшиеся из
оплодотворенного
карпогона;
5 — молодой карпогон;
6 — карпогон с
карпоспорами;
а — стерильные
веточки карпогона;
б — карпоспора;
в — пустой
карпоспорангий
После оплодотворения из брюшка карпогона вырастают ветвящиеся
нити – гонимобласты, конечные клетки которых дают карпоспоры.
Совокупность карпоспорангиев напоминает плод малины и
«цистокарпий»
104.
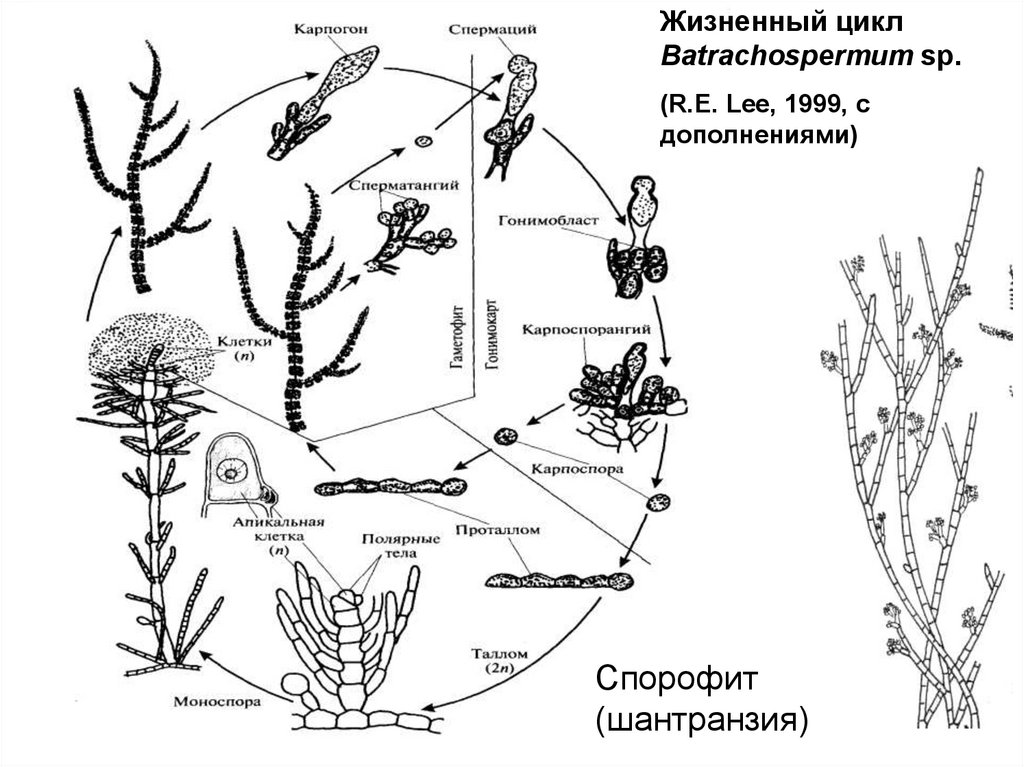
Жизненный циклBatrachospermum sp.
(R.E. Lee, 1999, c
дополнениями)
Спорофит
(шантранзия)
105. Порядок немалиальные (Nemaliales)
• Многоосевой псевдопаренхиматозныйталлом. Гетероморфная смена
поколений: шнуровидые слизистые
слабо ветвящиеся розовые кустики –
гаметофит и нитчатый ветвящийся
тэтраспорофит
106.
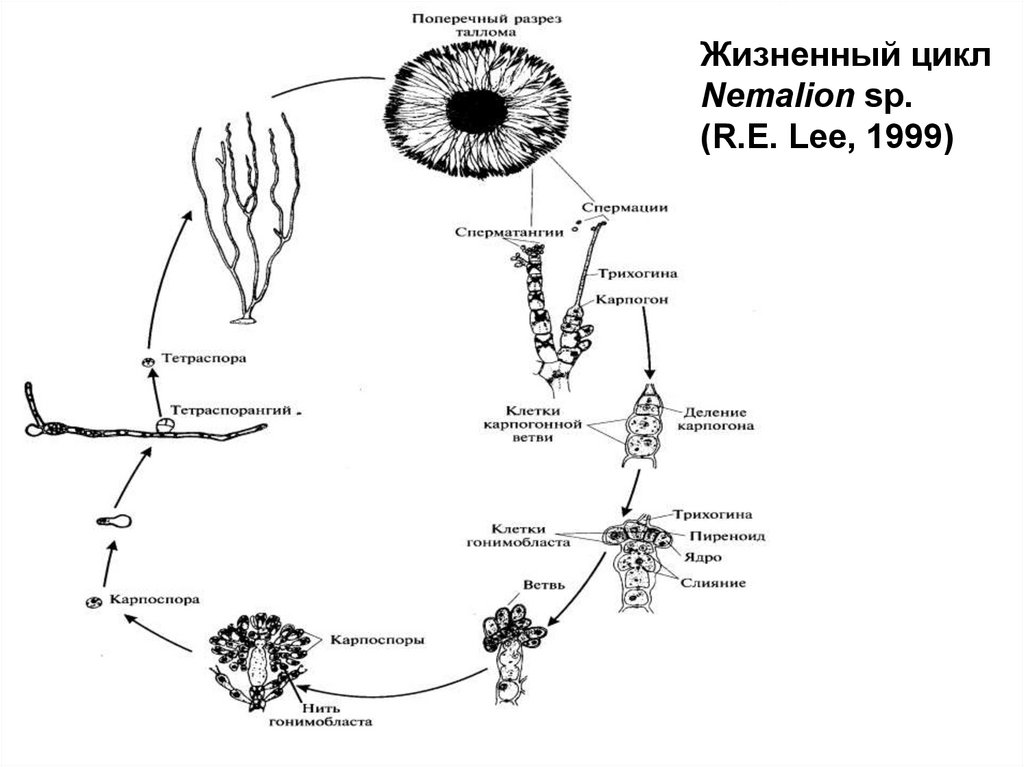
Жизненный циклNemalion sp.
(R.E. Lee, 1999)
107. Разнообразие красных водорослей
Polysiphonia.A—схема
строения
ветви гаметофита с
однослойной корой;
Б —поперечный разрез
ветви с многослойной
корой;
В—тетраспорофит с
тетраспорангиями;
Г
—
собрание
антеридиев;
Д
—
цистокарпий
зрелый
108. Polysiphonia. Схема развития
1 — центральная клетка. 2— перицентральные клетки, 3— мужской гаметофит,4— собрание антеридиев, 5 — центральная клетка, 6 — материнская клетка
антеридиев, 7— антеридии, 8— карпогониальная ветвь, 9 — карпогон с
яйцеклеткой, 10 — трихогина, 11 — спермаций; 12 — ауксилярная клетка,
отчленяющаяся от поддерживающей клетки карпогониальной ветви после того,
как произошло оплодотворение брюшка карпогона, 13 — слияние ауксилярной
клетки с брюшком оплодотворенного картогона, 14 — женский гаметофит,
15 — цистокарпий, одетый оболочкой, 16 — карпоспора, 17 — тетраспорофит,
18 — тетраспорангий с тетраспорами, 19 — тетраспора
109. Polysiphonia. Схема развития
1 — центральная клетка. 2— перицентральныеклетки, 3— мужской гаметофит,
4— собрание антеридиев, 5 — центральная клетка,
6 — материнская клетка антеридиев, 7— антеридии,
8— карпогониальная ветвь, 9 — карпогон с
яйцеклеткой,
10 — трихогина, 11 — спермаций; 12 — ауксилярная
клетка, отчленяющаяся от поддерживающей клетки
карпогониальной ветви после того, как произошло
оплодотворение брюшка карпогона,
13 — слияние ауксилярной клетки с брюшком
оплодотворенного картогона,
14 — женский гаметофит, 15 — цистокарпий,
одетый оболочкой,
16 — карпоспора,
17 — тетраспорофит, 18 — тетраспорангий с
тетраспорами, 19 — тетраспора
110. Polysiphonia. Схема развития
111. Размножение красных водорослей
112. Использование
• Из красных водорослей агарофитов добываютагар, карраген и агароиды.
• Агар применяют в микробиологии (как добавки
в питательную среду), в пищевой
промышленности (для желе, мармеладов,
консервирования мяса и рыбы), в
фармакологии для приготовления капсул.
• Многие водоросли используют для кормовой
муки и удобрений.
113.
114.
Красные водоросли это не толькомикроорганизмы,
благодаря которым
свое название
получило Красное
море. Это еще и
виновники "кровавого
прибоя" на пляжах
китайской провинции
Ляонин.