























 biology
biologySimilar presentations:


 нервная система")
 нервная система")

")
")

")

Вегетативная НС
1.
Вегетативная НСВсе функции организма подразделяются на соматические (анимальные)
и вегетативные (автономные). Вегетативные функции обеспечивают
обмен веществ, терморегуляцию, работу сердечно-сосудистой,
дыхательной, пищеварительной, выделительной и др. систем, рост и
размножение. Вегетативные реакции, как правило, не контролируются
сознанием.
Физиологическое значение:
1. Поддержание гомеостаза - постоянства внутренней среды организма.
2. Участие в вегетативном обеспечении различных форм психической и
физической деятельности.
2.
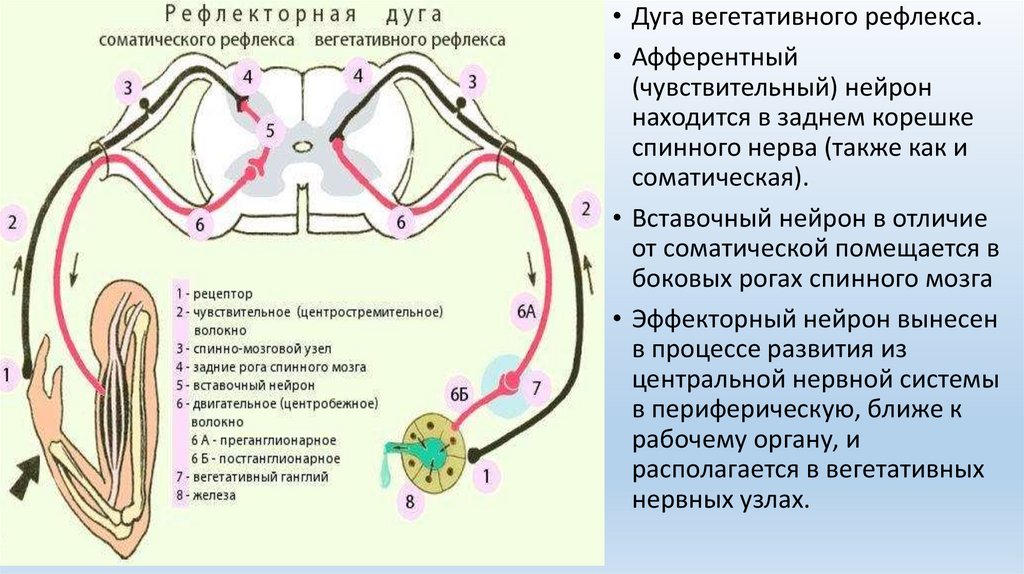
• Дуга вегетативного рефлекса.• Афферентный
(чувствительный) нейрон
находится в заднем корешке
спинного нерва (также как и
соматическая).
• Вставочный нейрон в отличие
от соматической помещается в
боковых рогах спинного мозга
• Эффекторный нейрон вынесен
в процессе развития из
центральной нервной системы
в периферическую, ближе к
рабочему органу, и
располагается в вегетативных
нервных узлах.
3.
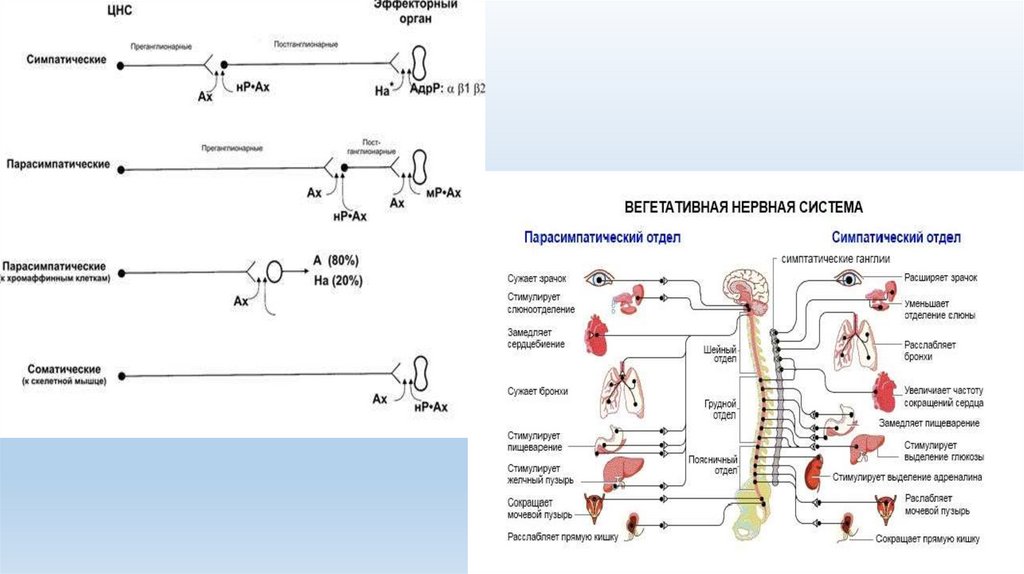
Симпатический отдел. Тела преганглионарных нейронов располагаются в сером веществе боковыхрогов спинного мозга, начиная от 1-го грудного сегмента до 2—3-го поясничного сегмента, и в
двигательных ядрах черепных нервов. Миелиновые аксоны этих нейронов представлены
медленнопроводящими возбуждение В-волокнами. Аксоны преганглионарных нейронов через синапсы
взаимодействуют с нейронами, расположенными в периферических ганглиях.
Околопозвоночные ганглии расположены по обе стороны вдоль позвоночника в виде цепочек,
составляющих правый и левый симпатические стволы.
Особенности строения симпатических и парасимпатических нервов. Симпатические нервы
сформированы в основном преганглионарными, а парасимпатические — постганглионарными
нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной
системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими
безмиелиновыми волокнами типа С.
Локализация ядер – заднее ядро гипоталамуса, нейроны боковых рогов грудных и поясничных
сегментов спинного мозга.
Характеристика отростков – прерываются в ганглиях, преганглионарные короткие, тип В,
постганглионарные длинные, тип С.
Зона иннервации – иннервирует все внутренние органы
Локализация – паравертебральные (пограничный столб), превертебральные (чревное, солнечное
сплетение, брыжеечные узлы).
Медиатор
Преганглионарный синапс – ацетилхолин. Рецептор: Н-холинорецептор
Постганглионарный синапс – норадреналин. Рецептор: Адренорецепторы
4.
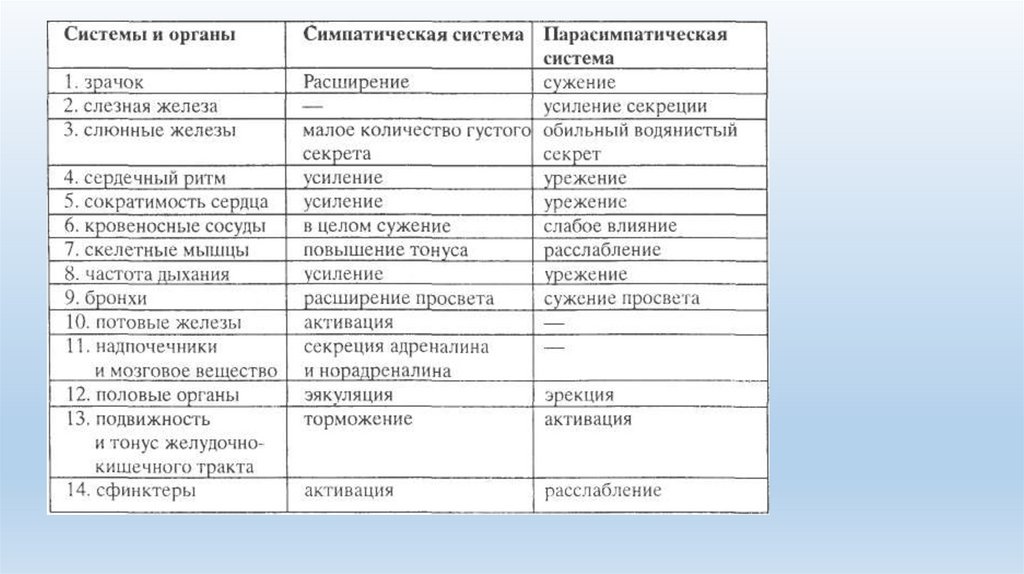
Физиологические свойства симпатической нервной системы:1.благодаря мультипликации в симпатических ганглиях распространяющееся из них возбуждение широко охватывает сразу несколько
различных органов, т.е. является генерализованным. Эти влияния наиболее отчетливо прослеживаются при эмоциональных реакциях.
2.оказывает активирующие влияния на функции иннервируемых органов: усиливает катаболические реакции, силу и частоту
сокращений сердца, повышает артериальное давление, улучшает оксигенацию тканей, увеличивает содержания глюкозы в крови,
скорость проведения возбуждения в скелетных мышцах и их тонус, расширяет бронхи, увеличивает объем легочной вентиляции;
расширяет зрачки, увеличивает секрецию катехоламинов надпочечниками. При этом одновременно снижается тонус
пищеварительного тракта, ослабляются процессы всасывания и ферментативного расщепления в кишечнике.
3. участвует в формировании целостных состояний, как агрессия, стресс, болевые реакции.
4. передача с пре- на ганглионарные нейроны осуществляется с помощью ацетилхолина, а на эффекторы — норадреналина.
5. представлена на периферии на значительном протяжении постганглионарными волокнами типа С, возбуждение на пути от
ганглиев к органам распространяется медленнее, чем в парасимпатической системе.
6. Эффекты действия более продолжительны.
7. Электрические потенциалы в симпатических ганглиях характеризуются продолжительными следовыми явлениями. При стимуляции
преганглионарных волокон регистрируется быстрый возбуждающий постсинаптический потенциал, который сменяется медленным
тормозным постсинаптическим потенциалом.
8. начальная деполяризация обусловлена действием ацетилхолина на никотиновые рецепторы.
9. Для того, чтобы проявился эффект норадреналина, он должен вступить во взаимодействие с адренорецепторами. Выделяют альфа
и бета адренорецепторы. При взаимодействия с альфа-адренорецептором меняется проницаемость мембраны для ионов натрия,
происходит деполяризация и, как следствие - возбуждение и усиление функции органа. При взаимодействии с бетаадренорецепторами происходит увеличение потока калия, гиперполяризация и соответственно торможение и снижение функции
органа. Исключение - взаимодействие норадреналина с бета-АР сердца вызывет усиление деятельности сердца. Помимо этого НА при
взаимодействии с адренорецептором может повышать активность аденилатциклазы, что приводит к образованию цАМФ
(внутриклеточного месенджера - посредника). Это приводит к активации протеинкиназ, являющимися внутриклеточными
регуляторами синтеза различных белков.
5.
Парасимпатический отдел Центральные нейроны расположены в передних отделах гипоталамуса,среднем и продолговатом мозге, во 2-4-м сегментах крестцового отдела спинного мозга.
Периферическая (исполнительная) часть ВНС представлена преганглионарными и
постганглионарными нейронами.
Парасимпатический отдел. Тела преганглионарных нейронов расположены в среднем и
продолговатом мозге среди двигательных нейронов черепных нервов: глазодвигательного, лицевого,
языкоглоточного и блуждающего, а также в сером веществе боковых рогов крестцового отдела
спинного мозга.
Особенности строения симпатических и парасимпатических нервов. Симпатические нервы
сформированы в основном преганглионарными, а парасимпатические — постганглионарными
нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной
системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими
безмиелиновыми волокнами типа С.
Локализация ядер – передний гипоталамус, средний мозг, мост, продолговатый мозг, боковые рога IV крестцовых сегментов спинного мозга.
Характеристика отростков - прерываются в ганглиях, преганглионарные длинные, тип В,
постганглионарные короткие, тип С.
Зона иннервации – ограничена (нет в надпочечниках и стенках большинства сосудов)
Локализация –в иннервируемых органах (интрамурально) или рядом с ними.
Медиатор
Преганглионарный синапс – ацетилхолин. Рецептор: Н-холинорецептор
Постганглионарный синапс – ацетилхолин. Рецептор: М-холинорецептор
6.
• Физиологические свойства парасимпатической нервной системы:• 1. влияния направлены и локальны. Оказывает ограниченное воздействие в
пределах иннервируемого органа.
• 2. оказывает успокаивающее, расслабляющее действие на большинство функций
организма; снижается возбудимость ЦНС и миокарда, уменьшаются интенсивность
метаболизма, сила и частота сердечных сокращений, кровяное давление, объем
легочной вентиляции, температура тела; увеличивается секреция инсулина и как
следствие снижается концентрация глюкозы в крови и увеличивается
внутриклеточное депонирование ее в виде гликогена. При этом одновременно
усиливаются моторная, секреторная и всасывательная функции желудочнокишечного тракта.
• 3. усиливает анаболические реакции.
• 4.Парасимпатические влияния доминируют в формировании сна и психологического
субъективного чувства удовлетворения.
• 5.Медиатором в преганглионарных и постганглионарных волокнах служит
ацетилхолин.
• 6.Парасимпатическая нервная система представлена на значительном расстоянии
преганглионарными волокнами типа В. Вследствие этого возбуждения от центра до
органа доходят быстрее, чем по симпатическим нервам.
• 7.Эффекты действия парасимпатических нервов менее продолжительны.
7.
8.
9.
Виды рефлексов ВНС1. Висцеро-висцеральный рефлекс включает пути, в которых
возбуждение возникает и заканчивается во внутренних органах.
Например, повышение или понижение давления в аорте вызывает
изменение деятельности сердца и тонуса сосудов.
2. Висцеро-дермальные. Возникают при раздражении внутренних
органов и проявляются в изменении потоотделении, изменении тонуса
кожных сосудов, повышение тактильной и болевой чувствительности
определенных областей кожи.
3. Дермо-висцеральные. При раздражении некоторых областей
поверхности тела возникают сосудистые реакции и изменения
функционирования определенных внутренних органов. На основе этих
рефлексов базируются методы рефлексотерапии (прогревание, массаж,
иглоукалывание и др.).
10.
Метасимпатическая нервная система• Метасимпатическая нервная система (МНС) - это комплекс микроганглионарных образований, расположенных в стенках
внутренних органов, обладающих двигательной автоматией - сердце, желудке, кишечнике, мочевом пузыре, бронхах и др.
• Структурно состоит из трех типов клеток (классификация по Догелю на примере Ауэрбахова и Мейснервого сплетений ЖКТ):
• 1-й тип - эфферентные нейроны с многочисленными короткими дендритами, длинные аксоны этих клеток заканчиваются на
мышечных клетках этого органа.
• 2-й тип - крупные, овальные или грушевидной формы афферентные нейроны с 4-5 нервными отростками, выходящими за
пределы ганглия. Аксоны заканчиваются на нейронах первого типа или идут к пара- и превертебральным ганглиям или
заканчиваются на нейронах спинного мозга. Т.е. афферентная импульсация от внутренних органов может замыкаться на разных
уровнях.
• 3-й тип - редко встречающиеся ассоциативные нейроны, расположены в ганглиях, аксоны заканчиваются на дендритах нейронов
1 и 2 типа. Эти нейроны обеспечивают замыкание рефлекса внутри органа.
В метасимпатическом отделе АНС наряду с холинергическими и адренергическими нейронами широко представлены
нехолинергические и неадренергические нейроны (отсюда название - нехолинергический и неадренергический отдел). В качестве
медиаторов в них могут находиться пептиды (динорфин, холецис- токинин, нейропептид Y, соматостатин, ВИП), биогенные амины
(серотонин, гистамин, мелатонин), пурины (АТФ) и др. Некоторые моторные нейроны содержат одновременно до пяти различных
медиаторов. Вероятно, что это необходимо для очень тонкой регуляции работы эффекторных клеток (например, для управления
моторикой кишечника, приуроченной к процессам гидролиза и всасывания питательных веществ).
АТФ связывается в синапсах с пуринорецепторами. Пуринорецепторы разделяют на два подтипа - Р-1-пуринорецепторы и Р-2пуринорецепторы. Р-1-рецепторы чувствительны к аденозину-АМФ-АДФ-АТФ, блокируются метилксантином. Р-2-пуринорецепторы
чувствительны к АТФ-АДФ-АМФ-аденозину, блокируются хинидином.
+Активация пуринорецепторов приводит к расслаблению гладких мышц ЖКТ, ССС, мочеполовой системы
11.
Свойства и функции метасимпатическойнервной системы.
1. Иннервирует только внутренние органы с моторной активностью, содержащие внутреннюю
полость (пищевод, желудок, кишечник, сердце, матка, мочевой и желчный пузыри, сосуды).
2. Имеет также собственную афферентную часть. Не имеет прямых контактов с эфферентной
частью соматической нервной системы. Может выполнять роль самостоятельного
интегрирующего образования, так как в ней имеются готовые рефлекторные дуги
(афферентные - вставочные - эфферентные нейроны). Например, Г.И. Косицкий показал, что в
сердце имеются внутрисердечные рефлексы МНС - растяжение правого предсердия
увеличивает работу левого желудочка, а растяжение левого предсердия повышает работу
правого желудочка. Этот эффект может тормозиться или блокироваться ганглиоблокаторами.
4. Метасимпатическая нервная система не находится в антагонистических отношениях с
симпатической и парасимпатической нервной системой и более независима от ЦНС.
5. Участвует в регуляции локального кровотока и проницаемости сосудистой стенки.
6. Регулирует функции местных эндокринных клеток и секреторной, экскреторной,
всасывательной деятельности ЖКТ.
7. Имеет собственные медиаторы.
12.
13.
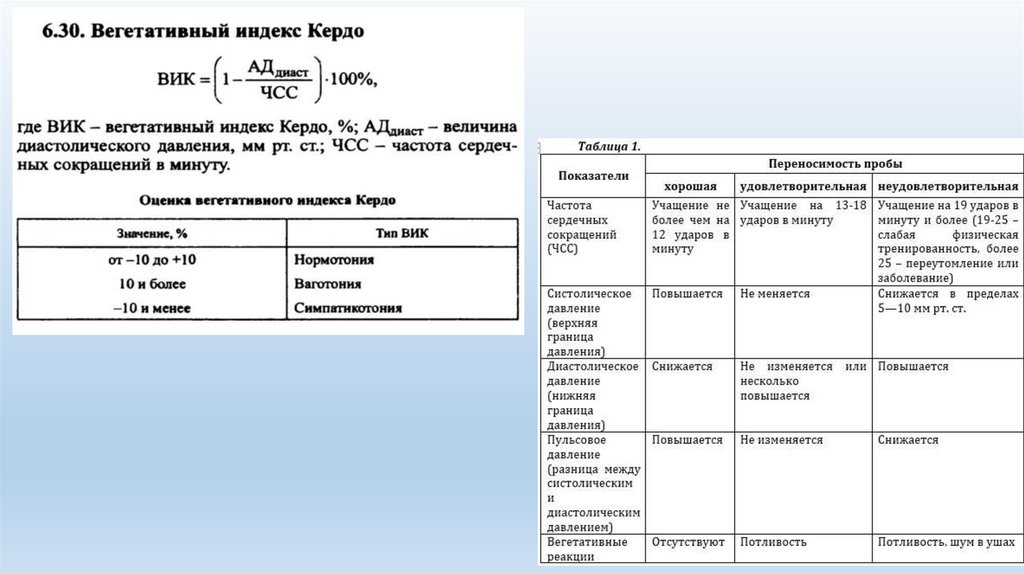
Оценивать вегетативный статус человека (индекс Кердо). Вегетативная реактивность: холодовая проба, ортопроба.
Индекс Кердо — показатель, использующийся для оценки состояния вегетативного тонуса по параметрам, характеризующим состояние сердечно-сосудистой
системы – АД и ЧСС.
Вычисляется по формуле
ВИК = (1-АДд/ЧСС) х 100%
АДд – диастолическое давление, ЧСС – частота сердечных сокращений.
При нормотонии – ВИК от -10 до +10%
При симпатикотонии – ВИК более +10%
При ваготонии – ВИК менее -10%
Холодовая проба. Сущность холодовой пробы заключается в том, что при опускании предплечья в холодную воду (+4°С) происходит рефлекторное сужение
артериол и артериальное давление повышается, причем, тем больше, чем больше возбудимость сосудодвигательных центров.
В состоянии покоя у испытуемого на плечевой артерии трижды до получения стабильных цифр измеряют АД и ЧСС. Затем ему предлагают на 1 мин погрузить
кисть правой руки (немного выше лучезапястного сустава) в воду температуры +4°С. АД и ЧСС измеряют через 30 сек после начала пробы и через 1 минуту, а
также через каждую минуту, после того как рука извлечена из сосуда.
1.Для нормальной вегетативной реактивности характерны следующие изменения. Систолическое АД повышается на 15-20 мм рт.ст., диастолическое АД
повышается на 10-15 мм рт.ст., максимальный подъем АД происходит через 30 сек от начала охлаждения. ЧСС увеличивается на 10-15 уд/мин. Возврат АД к
исходному уровню происходит через 2-3 мин.
2. Гиперреактивность внс характеризуется более значительным повышением АД и ЧСС, что называется гиперсимпатической реакцией.
3. Гипореактивность внс характеризуется менее значительным увеличением АД и ЧСС, что называется слабой симпатической реакцией.
4. Извращенная реактивность внс характеризуется снижением АД и ЧСС, что называется парасимпатической реакцией.
Ортопроба, Принцип метода:
Пассивная ортостатическая проба позволяет выявить нарушения вегетативной нервной регуляции работы сердца, а именно барорецепторного контроля
артериального давления (АД), приводящие к головокружениям и обморочным состояниям и иным проявлениям вегетативной дисфункции.
Описание метода: При проведении пассивной ортостатической пробы сначала измеряют исходный уровень АД и частоту сердечных сокращений (ЧСС) в
положении больного лежа на спине (около 10 минут), после чего ортостатический стол резко переводят в полувертикальное положение, проводя
повторные измерения АД и ЧСС. Рассчитывается степень отклонения АД и ЧСС от исходных показателей в (%).
Нормальная реакция: увеличение ЧСС (до 30% от фона) при незначительном снижении систолического АД (не более 2-3% от исходного).
Снижение АД более 10-15% от исходного: Нарушение вегетативной регуляции по ваготоническому типу.
14.
Важные вегетативные рефлексы• Рефлекс глазосердечный (Даньини-Ашнера) : замедление пульса на 4 —8 ударов в 1 мин. и снижение артериального давления при
надавливании на глазные яблоки; большее замедление указывает на повышенную возбудимость парасимпатической части
вегетативной нервной системы.
• Рефлекс солярный : замедление пульса и снижение артериального давления при надавливании на брюшную стенку между пупком и
мечевидным отростком (область чревного сплетения).
• Рефлекс синокаротидный : надавливание на сонную артерию на шее в течение 20-30 сек и одновременрное измерение пульса.
• Прессорный синокаротидный рефлекс сопровождается повышением периферического сопротивления, давления в аорте, желудочках
сердца, но частота сокращений сердца может не изменяться и сопротивление коронарных сосудов несколько возрастает.
• Геринга рефлекс : замедление пульса при задержке дыхания на стадии глубокого вдоха; если в положении сидя это замедление
превышает 6 ударов в 1 мин., то оно свидетельствует о повышенной возбудимости блуждающего нерва.
• Клиностатический рефлекс: обследуемый плавно переходит из вертикального положения в горизонтальное. При этом пульс
замедляется на 4-6 уд/мин, что считается положительной реакцией, или на 8-12 уд/мин, что считается резко положительной реакцией.
• Ортостатический рефлекс: обследуемый в положении лежа плавно встает. При этом в норме происходит ускорение пульса на 6-24
уд/мин, что считается положительной реакцией, или более, чем на 24 уд/мин, что считается резко положительной реакцией.
• Волосковый рефлекс: пощипывание кожи, трение, прикосновение льда, раздражение эфиром и хлорэтидом, в ответ возникает
«гусиная» кожа в области раздражения.
• Потовые рефлексы: вызывают различными способами: согреванием тела, введением потогонных средств, местным кожным
раздражением. Оценку степени ответного потоотделения производят с помощью различных объективных методик. При раздражении
парасимпатической нс отделяется жидкий, водянисто-обильный пот, при возбуждении симпатической – скудный, вязкий.
15.
• Нервный центр — это совокупность нейронов, необходимых дляосуществления определенного рефлекса или регуляции определенной
функции.
• Классификация нервных центров.
• По локализации в структурах нервной системы различают корковые,
подкорковые и спинальные центры. В головном мозге также выделяют
центры диэнцефальные, мезэнцефальные, бульбарные, гипоталамические,
таламические.
• На функциональной основе центры нервной системы разделяют по
регулируемой функции (например, сосудодвигательный центр, центр
теплообразования, дыхательный центр и др.) или по афферентному
восприятию (например, центры зрения, слуха, обоняния и др.). Выделяют
также центры нервной системы, которые формируют мотивационные
состояния организма, являясь пейсмекерами мотивационных возбуждений
(центры голода, жажды, насыщения и др.).
• Существуют центры нервной системы, которые на основе интеграции
возбуждений формируют целостные реакции организма (например, центры
глотания, чиханья, дефекации, половой центр и т.п.).
16.
Свойства нервных центров• Одностороннее проведение возбуждение в нервном центре. Возбуждение в ЦНС
проводится в одном направлении с аксона на дендрит или тело клетки следующего
нейрона. Основу этого свойства составляют особенности морфологической связи между
нейронами.
• Одностороннее проведение возбуждения зависит от строения синапса и гуморальной
природы передачи в нем импульса: медиатор, осуществляющий передачу возбуждения,
выделяется только в пресинаптическом окончании, а рецепторы, воспринимающие
медиатор, расположены на постсинаптической мембране;
• Замедление проведения возбуждения (центральная задержка). В системе рефлекторной
дуги медленнее всего проводится возбуждение в синапсах ЦНС. В связи с этим центральное
время рефлекса зависит от количества вставочных нейронов.
• Чем сложнее рефлекторная реакция, тем больше центральное время рефлекса. Его
величина связана со сравнительно медленным проведением возбуждения через
последовательно включенные синапсы. Замедление проведения возбуждения создается
вследствие относительной длительности осуществляющихся в синапсах процессов:
выделения медиатора через пресинаптическую мембрану, его диффузии через
синаптическую щель, возбуждения постсинаптической мембраны, возникновения
возбуждающего постсинаптического потенциала и его перехода в потенциал действия;
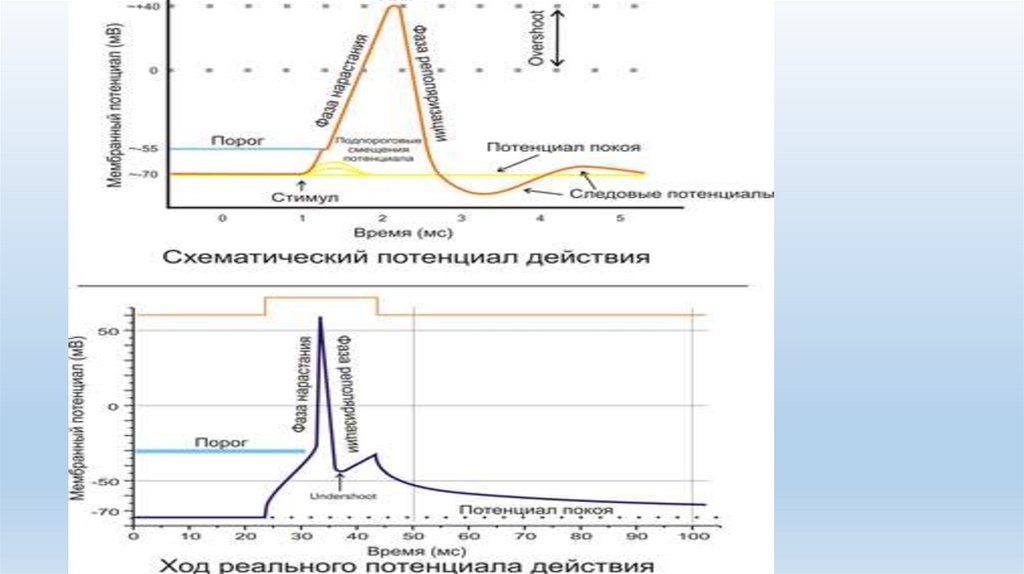
17.
• Последействие — способность сохранять возбуждение послеокончания действия раздражителя, т.е. афферентных импульсов нет, а
эфферентные продолжают действовать еще некоторое время.
• Последействие объясняется наличием следовой деполяризации. Если
следовая деполяризация длительна, то на ее фоне в течение
нескольких миллисекунд могут возникать потенциалы действия
(ритмическая активность нейрона), вследствие чего сохраняется
ответная реакция. Но это дает сравнительно короткий эффект
последействия.
• Более длительное последействие связано с наличием кольцевых
связей между нейронами. В них возбуждение как бы само себя
поддерживает, возвращаясь по коллатералям к первоначально
возбужденному нейрону
18.
19.
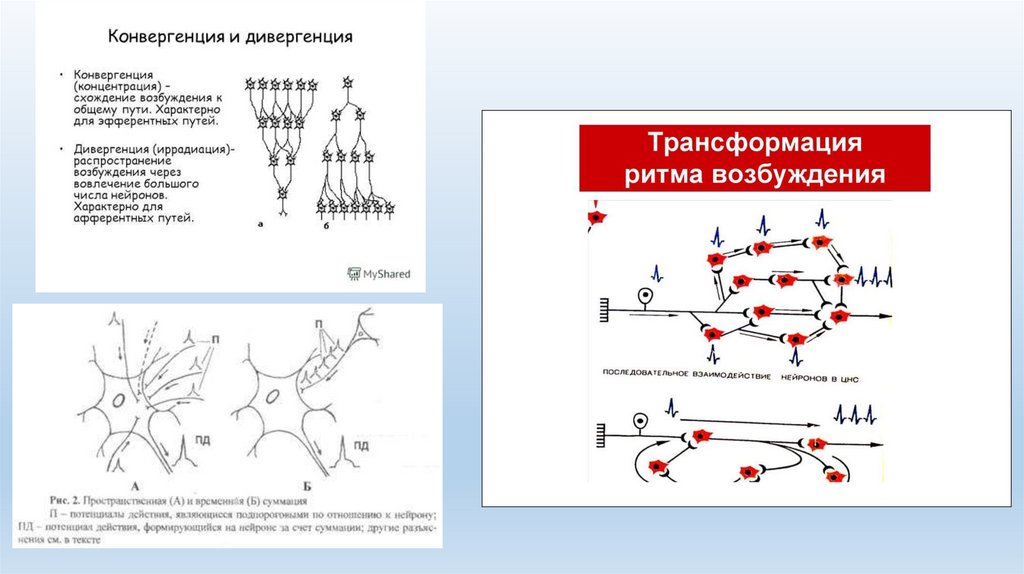
• Трансформация ритма возбуждения. Нервные центры способныизменять ритм поступающих к ним импульсов. Они могут
отвечать на одиночные раздражители серией импульсов или на
раздражители небольшой частоты — возникновением более
частых потенциалов действия. В результате ЦНС посылает к
рабочему органу количество импульсов, относительно
независимое от частоты раздражений.
20.
21.
• Облегчение проведения или проторение пути. Установлено, что послевозбуждения, возникшего в ответ на ритмическое раздражение,
следующий стимул вызывает больший эффект, или для поддержания
прежнего уровня ответной реакции требуется меньшая сила
последующего раздражения. Это явление получило название
«облегчение».
• Его можно объяснить тем, что при первых стимулах ритмического
раздражителя происходит перемещение пузырьков медиатора ближе
к пресинаптической мембране и при последующем раздражении
медиатор быстрее выделяется в синаптическую щель. Это, в свою
очередь, приводит к тому, что вследствие суммации возбуждающего
постсинаптического потенциала быстрее достигается критический
уровень деполяризации и возникает распространяющийся потенциал
действия
22.
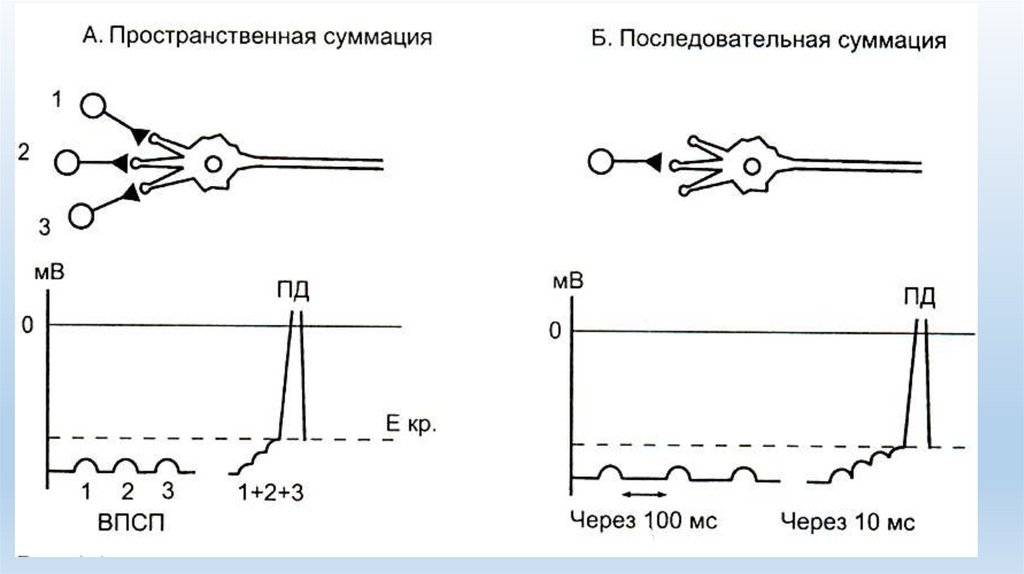
• Суммация, впервые описанная И.М. Сеченовым (1863) и заключающаяся в том, что слабыепо силе раздражители, не вызывающие видимой реакции, при частом повторении могут
суммироваться, создавать надпороговую силу и вызывать эффект возбуждения. Различают
два вида суммации — последовательную и пространственную.
• Последовательная суммация в синапсах возникает в том случае, когда по одному и тому же
афферентному пути к центрам поступает несколько подпороговых импульсов. В результате
суммации местного возбуждения, вызванного каждым подпороговым стимулом, возникает
ответная реакция.
• Пространственная суммация заключается в появлении рефлекторной реакции в ответ на
два или несколько подпороговых стимулов, приходящих в нервный центр по разным
афферентным путям
• Пространственную суммацию, как и последовательную, можно объяснить тем, что при
подпороговом раздражении, пришедшем по одному афферентному пути, выделяется
недостаточное количество медиатора для того, чтобы вызвать деполяризацию мембраны
до критического уровня. Если же импульсы приходят одновременно несколькими
афферентными путями к одному и тому же нейрону, в синапсах выделяется достаточное
количество медиатора, необходимое для пороговой деполяризации и возникновения
потенциала действия;
23.
24.
• Иррадиация. При возбуждении нервного центра нервные импульсыраспространяются на соседние центры и приводят их в деятельное
состояние. Это явление получило название иррадиации. Степень
иррадиации зависит от количества вставочных нейронов, степени их
миелинизации, силы раздражителя. Со временем в результате афферентной
стимуляции только одного нервного центра зона иррадиации уменьшается,
происходит переход к процессу концентрации, т.е. ограничению
возбуждения только в одном нервном центре.
• Это является следствием уменьшения синтеза медиаторов во вставочных
нейронах, в результате чего биотоки не передаются из данного нервного
центра на соседние Выражением данного процесса является точная
координированная двигательная реакция в ответ на раздражение
рецептивного поля. Формирование любых навыков (трудовых, спортивных и
т.д.) обусловлено тренировкой двигательных центров, основу которых
составляет переход от процесса иррадиации к концентрации;
25.
• Индукция. Основой взаимосвязи между нервными центрами является процесс индукции —наведение (индуцирование) противоположного процесса. Сильный процесс возбуждения в нервном
центре вызывает (наводит) торможение в соседних нервных центрах (пространственная
отрицательная индукция), а сильный тормозной процесс индуцирует в соседних нервных центрах
возбуждение (пространственная положительная индукция). При смене этих процессов в пределах
одного центра говорят о последовательной отрицательной или положительной индукции. Индукция
ограничивает распространение (иррадиацию) нервных процессов и обеспечивает концентрацию.
Способность к индукции в значительной степени зависит от функционирования тормозных
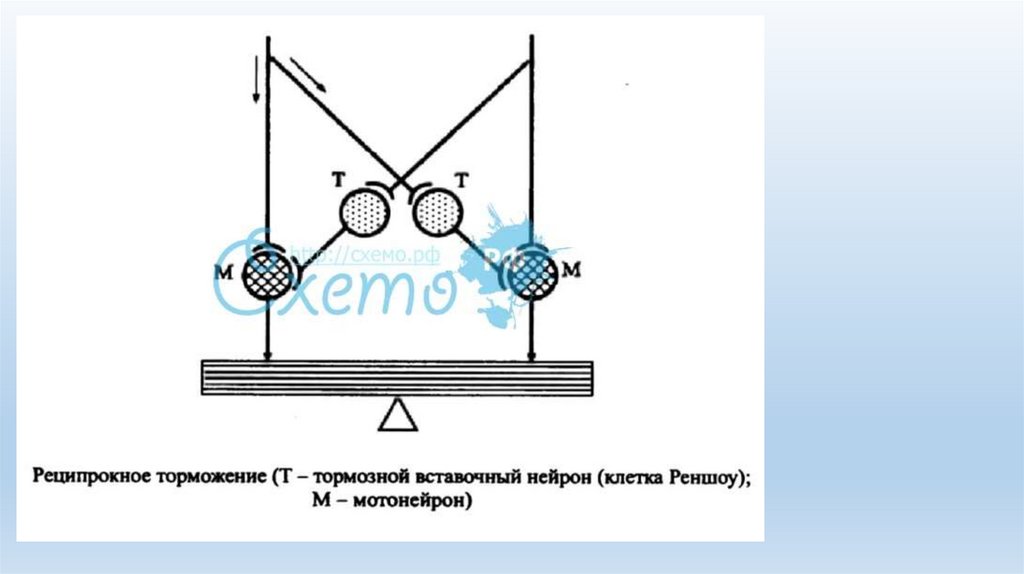
вставочных нейронов — клеток Реншоу.
• От степени развития индукции зависят подвижность нервных процессов, возможность выполнения
движений скоростного характера, требующих быстрой смены возбуждения и торможения.
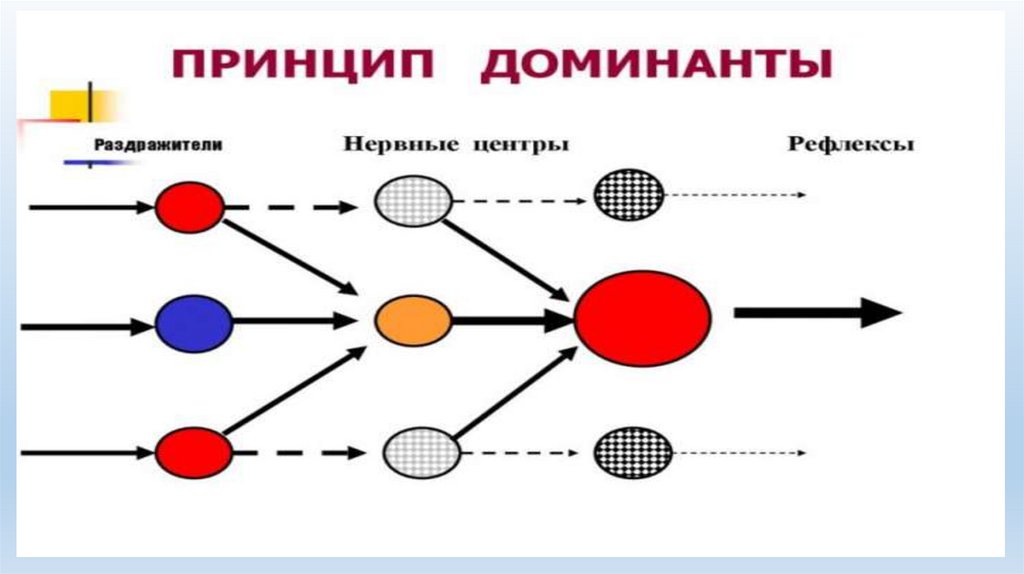
• Индукция является основой доминанты — образования нервного центра повышенной возбудимости.
Это явление впервые было описано А.А. Ухтомским. Доминантный нервный центр подчиняет себе
более слабые нервные центры, притягивает их энергию и за счет этого еще более усиливается. В
результате этого раздражение различных рецепторных полей начинает вызывать рефлекторный ответ,
характерный для деятельности этого доминантного центра. Доминантный очаг в ЦНС может возникать
под влиянием разных факторов, в частности сильной афферентной стимуляции, гормональных
воздействий, мотиваций и т.д.;
26.
• Свойства доминанты1) Повышенная возбудимость, т.е. чувствительность к возбуждению.
2) Устойчивость во времени.
3) Торможение других очагов возбуждения.
4) Использование "чужого" возбуждения из других очагов для усиления собственного.
• Законы формирования доминанты и её динамика (развитие во времени)
• 1. Генерализация доминанты - это распространение повышенной возбудимости на большие территории головного мозга.
Доминантный очаг в этот период очень большой. Какое бы раздражение не давалось, ответ будет только один - доминантный рефлекс.
• 2. Концентрация доминанты (локализация) - это "стягивание" большой зоны возбудимости в один локальный и концентрированный
доминантный очаг.
• 3. Дифференциация доминанты - выбор предпочитаемого, приоритетного варианта раздражения для осуществления доминантного
рефлекса. Формируется механизм различения сходных раздражителей, и лишь один из них начинает играть роль пускового
раздражителя, запускающего деятельность доминанты.
• 4. Консолидация доминанты - укрепление очага возбуждения, приобретение им повышенной устойчивости, подчинение ему других
очагов возбуждения.
• 5. Констелляция доминанты - это создание целого "созвездия" из отдельных доминантных очагов на разных уровнях мозга, связанных
друг с другом и работающих совместно. Происходит вовлечение в структуру доминанты дополнительных очагов, разположенных уже в
разных отделах и областях мозга, а не в соседних участков, как в случае генерализации. Это понятие доминанты в виде констелляции
возбуждённых очагов, взаимодействующих друг с другом, соответствует расширенному понятию нервного центра как управляющей
системы для регуляции определённой функции (например, "дыхательный центр", "пищевой центр" и т.п.). Но при этом считается, что
констелляция имеет всё же временный, функциональный характер, что сближает её с понятием "функциональная система", которое
значительно позже было введено в физиологию П.К. Анохиным.
• 6. Изоляция доминанты - это частичное отделение и обособление доминантного очага от других возбуждённых участков
мозга, приобретение им автономности и пониженной реактивности. На этом этапе происходит ограничение количества входов в
данный доминантный очаг. Образно говоря, можно сказать, что доминанта начинает жить своей собственной жизнью, отдельно от
остального мозга. Это позволяет мозгу начать формировать другие доминанты параллельно с возникшей ранее.
• 7. Разрушение доминанты (затухание доминанты) - прекращение деятельности данного доминантного очага возбуждения, которое
может закончиться прекращением его существования.
27.
28.
29.
• Дивергенция и конвергенция. Способность нейрона устанавливать многочисленныесинаптические связи с различными нервными клетками в пределах одного или разных
нервных центров называется дивергенциеи. Например, центральные окончания аксонов
первичного афферентного нейрона образуют синапсы на многих вставочных нейронах.
Благодаря этому одна и та же нервная клетка может участвовать в различных нервных
реакциях и контролировать большое число других нейронов, что приводит к иррадиации
возбуждения.
• Схождение различных путей проведения нервных импульсов к одному и тому же нейрону
получило название конвергенции. Простейшим примером конвергенции является
замыкание на одном двигательном нейроне импульсов от нескольких афферентных
(чувствительных) нейронов. В ЦНС большинство нейронов получают информацию от разных
источников благодаря конвергенции. Это обеспечивает пространственную суммацию
импульсов и усиление конечного эффекта.
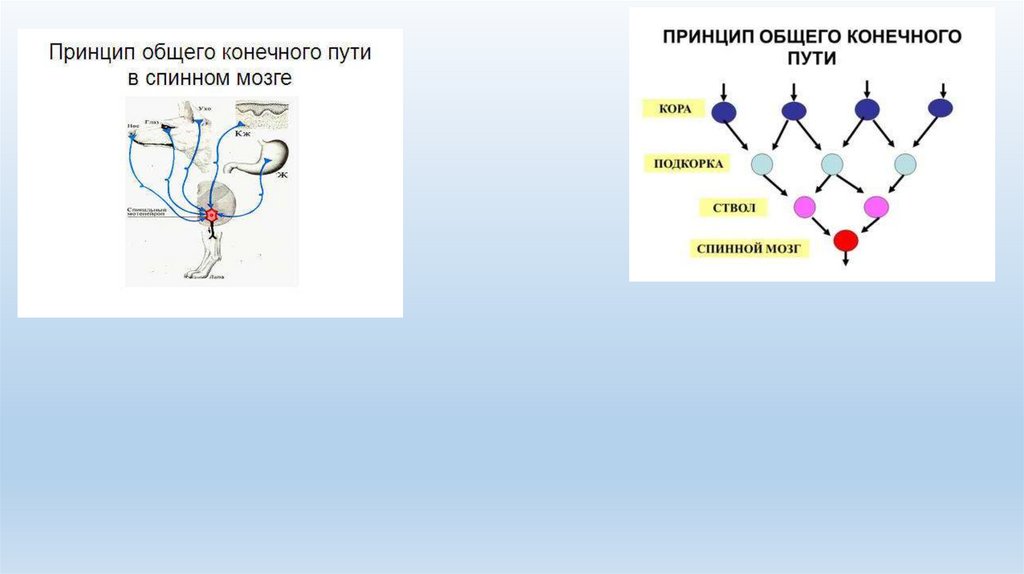
• Явление конвергенции было описано Ч. Шеррингтоном и получило название воронки
Шеррингтона, или эффекта общего конечного пути. Данный принцип показывает, как при
активации различных нервных структур формируется конечная реакция, что имеет
первостепенное значение для анализа рефлекторной деятельности;
30.
• Утомляемость нервного центра. Нервный центр обладает малойлабильностью. Он постоянно получает от множества
высоколабильных нервных волокон большое количество
стимулов, превышающих его лабильность. Поэтому нервный
центр работает с максимальной загрузкой и легко утомляется.
• Исходя из синаптических механизмов передачи возбуждения
утомление в нервных центрах может объясняться тем, что но
мере работы нейрона истощаются запасы медиатора и
становится невозможной передача импульсов в синапсах. Кроме
того, в процессе деятельности нейрона наступает постепенное
снижение чувствительности его рецепторов к медиатору, что
называется десенситизацией;
31.
• Чувствительность нервных центров к кислороду и некоторым фармакологическим веществам. Внервных клетках осуществляется интенсивный обмен веществ, для чего необходимы энергия и
постоянный приток нужного количества кислорода.
• Пластичность нервных центров. Под пластичностью понимают функциональную изменчивость и
приспособляемость нервных центров. Это особенно ярко проявляется при удалении разных отделов
мозга. Нарушенная функция может восстанавливаться, если были частично удалены какие-то отделы
мозжечка или коры больших полушарий. О возможности полной перестройки центров
свидетельствуют опыты по сшиванию функционально различных нервов. Если перерезать
двигательный нерв, иннервирующий мышцы конечностей, и его периферический конец сшить с
центральным концом перерезанного блуждающего нерва, регулирующего внутренние органы, то
через некоторое время периферические волокна двигательного нерва перерождаются (вследствие их
отделения от тела клетки), а волокна блуждающего нерва прорастают к мышце. Последние образуют в
мышце синапсы, свойственные соматическому нерву, что приводит к постепенному восстановлению
двигательной функции. В первое время после восстановления иннервации конечности раздражение
кожи вызывает свойственную блуждающему нерву реакцию — рвоту, гак как возбуждение от кожи по
блуждающему нерву поступает в соответствующие центры продолговатого мозга. Через некоторое
время раздражение кожи начинает вызывать обычную двигательную реакцию, поскольку происходит
полная перестройка деятельности центра.
32.
• Принцип общего конечного пути. Эффекторные нейроны ЦНС(прежде всего мотонейроны спинного мозга), являясь конечными в
цепочке состоящей из афферентных, промежуточных и эффекторных
нейронов, могут вовлекаться в осуществление различных реакций
организма возбуждениями, приходящими к ним от большого числа
афферентных и промежуточных нейронов, для которых они являются
конечным путем (путем от ЦНС к эффектору). Например, на
мотонейронах передних рогов спинного мозга, иннервирующих
мускулатуру конечности, оканчиваются волокна афферентных
нейронов, нейронов пирамидного тракта и экстрапирамидной
системы (ядер мозжечка, ретикулярной формации и многих других
структур). Поэтому эти мотонейроны, обеспечивающие рефлекторную
деятельность конечности, рассматриваются как конечный путь для
общей реализации на конечность многих нервных влияний.
33.
34.
• Основные принципы в работе нервных центров• Принцип общего конечного пути ("воронка Шеррингтона"). Как правило, центры имеют больше афферентных входов, чем
эфферентных выходов. Поэтому входящие потоки возбуждения конкурируют за выход, имея общий конечный путь. В итоге
количество афферентных входов превышает количество эфферентных выходов.
Принцип обратной связи. Это означает, что последующий элемент (нейрон или центр) в последовательной цепи
взаимосвязанных элементов влияет на состояние предыдущего элемента. Обратная связь позволяет произвести отладку
взаимодействия между элементами и добиться их оптимального взаимодействия для достижения предельно возможного
положительного результата в работе системы, состоящей из этих элементов.
Принцип доминанты. Это означает, что нервный путь или нервный центр наиболее активный получает преимущество по
отношению к другим путям или центрам и начинает доминировать, господствовать над ними. Он тормозит их деятельность
и перехватывает их возбуждение, чтобы усилить своё.
Принцип иерархии (соподчинения). Это означает, что одни элементы (нейроны и/или центры) подчиняются влиянию
других элементов. Как правило, вышерасположенные центры подчиняют себе нижерасположенные центры.
• Принцип пластичности. Это означает, что нервный центр перестраивает свою деятельность, приспосабливаясь к
наилучшему выполнению своей функции для достижения общего конечного системного результата. Пластичность является
важнейшей отличительной особенностью биосистем по сравнению с техническими системами.