










 biology
biologySimilar presentations:




")





Регуляция экспрессии генов у прокариот и эукариот
1.
РЕГУЛЯЦИЯЭКСПРЕССИИ ГЕНОВ У
ПРОКАРИОТ и ЭУКАРИОТ
План:
1. Регуляция работы генов у прокариот на
примере лактозного оперона.
2. Регуляция работы генов у эукариот
Специфическая
Неспецифическая
2.
Ф.Жакоб и Ж.Моно 1961: общая теориярегуляции генов
• Сущность
теории
сводится
к
«выключению» или «включению» генов
как
функциональных
единиц,
к
возможности
или
невозможности
проявления их способности передавать
информацию о структуре белка.
• У прокариот гены, контролирующие
синтез
белков-ферментов,
катализирующих ход последовательных
биохимических реакций, объединяются
в структурно-функциональную единицу
– оперон.
3.
Виды опероновИндуцибельный - регуляция по типу
индукции, прямой положительной связи.
Ииндуктором является исходный продукт
(субстрат). Субстрат стимулирует реакции
своего метаболизма.
Репрессибельный - регулятором является
конечный продукт
(корепрессор). Он
тормозит
реакции,
ведущие
к
его
образованию. В отсутствии корепрессора
белок – репрессор имеет низкое сродство к
оператору
и
РНК-полимераза
транскрибирует гены.
4.
Состав индуцибельного оперона• Промотор – участок молекулы ДНК, к
которому присоединяется РНК-полимераза
• Оператор – участок молекулы ДНК, место
связывания
с
регуляторным
белкомрепрессором.
• Структурные гены, кодирующие белкиферменты
• Терминатор – стоп-кодон
5.
Принцип регуляции активности индуцибельногооперона заключается в следующем: сродство
белка-репрессора
с
геном-оператором
определяется наличием или отсутствием
метаболитов той цепи реакций, ферменты
которой кодируются данным опероном.
За синтез белка – репрессора отвечает ген регулятор. Белок-репрессор
обладает
аллостерическим эффектом, т.е. может
связываться
и
с
оператором,
и
с
метаболитом.
6.
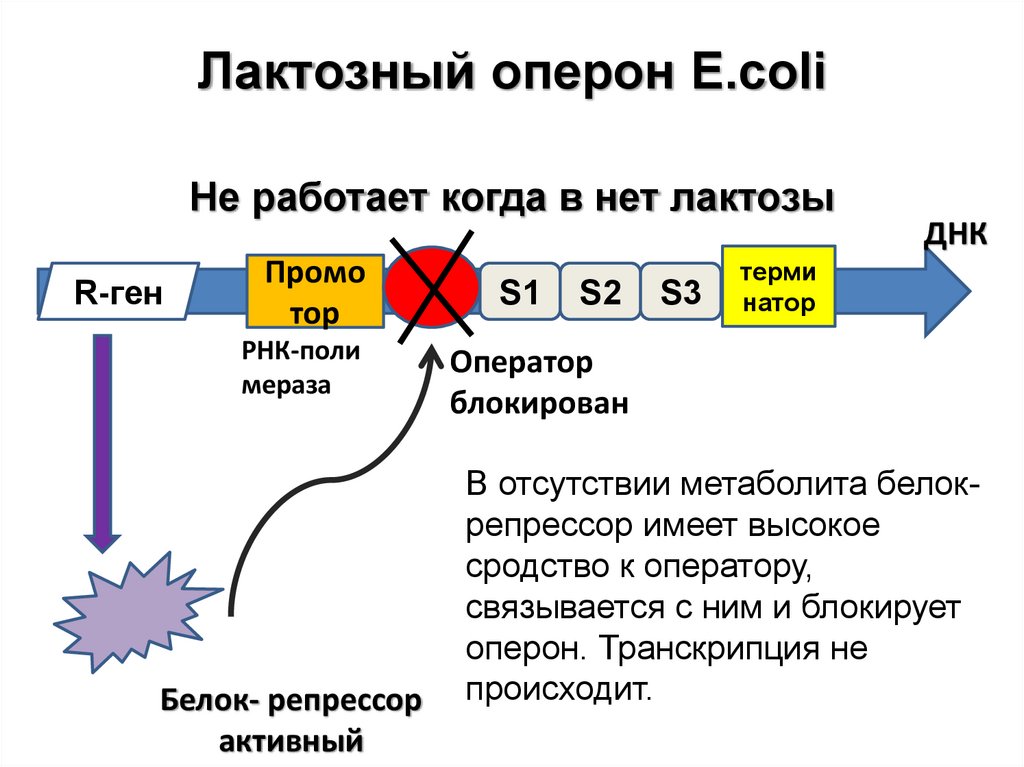
Лактозный оперон E.coliНе работает когда в нет лактозы
R-ген
Промо
тор
РНК-поли
мераза
Белок- репрессор
активный
S1
S2
S3
ДНК
терми
натор
Оператор
блокирован
В отсутствии метаболита белокрепрессор имеет высокое
сродство к оператору,
связывается с ним и блокирует
оперон. Транскрипция не
происходит.
7.
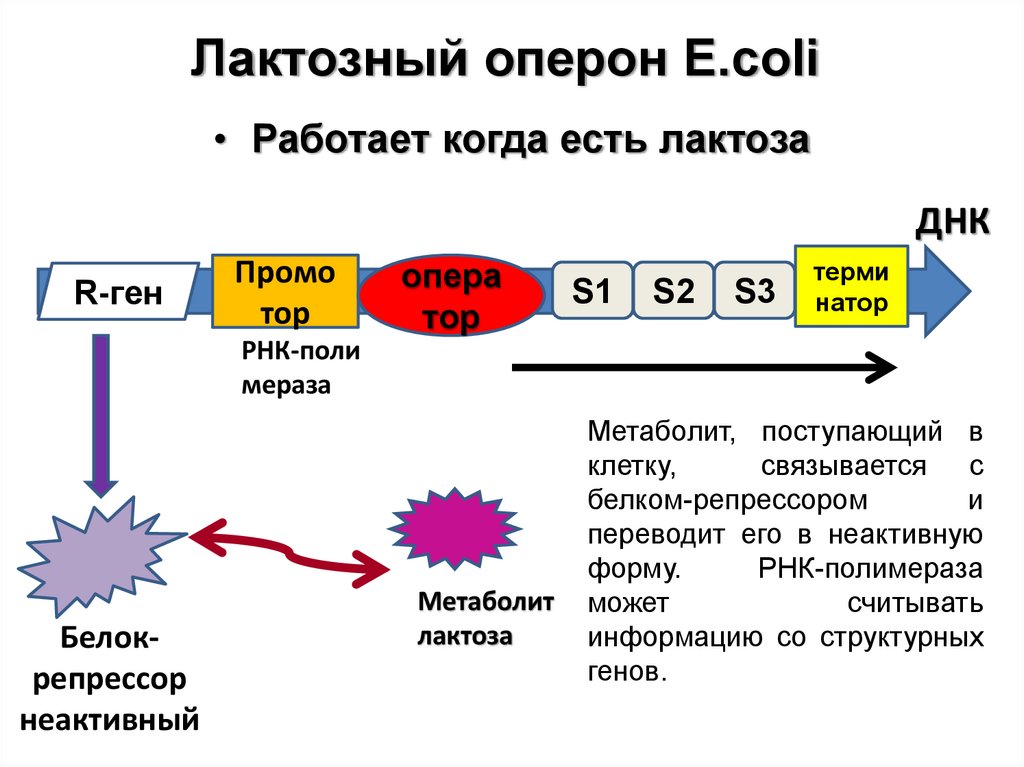
Лактозный оперон E.coli• Работает когда есть лактоза
ДНК
R-ген
Промо
тор
опера
тор
S1
S2
S3
терми
натор
РНК-поли
мераза
Белокрепрессор
неактивный
Метаболит
лактоза
Метаболит, поступающий в
клетку,
связывается с
белком-репрессором
и
переводит его в неактивную
форму.
РНК-полимераза
может
считывать
информацию со структурных
генов.
8.
Регуляция экспрессии генов уэукариот
• Регуляция активности генов у эукариот
проходит на всех этапах реализации
генетической информации, что связано с
особенностями генома и организацией
хроматина,
а
также
разделением
транскрипции и трансляции во времени и в
пространстве.
• На процессы включения и выключения генов
влияют
различные
хромосомные
перестройки, МГЭ, изменяющие эффект
положения гена.
9.
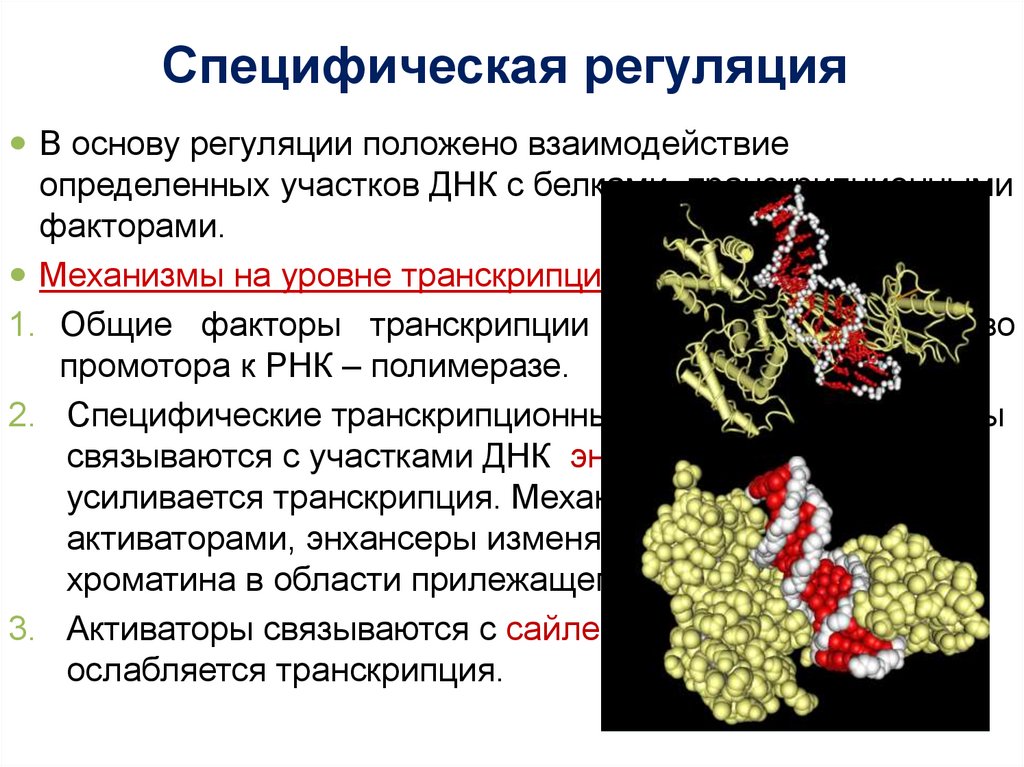
Специфическая регуляцияВ основу регуляции положено взаимодействие
определенных участков ДНК с белками- транскрипционными
факторами.
Механизмы на уровне транскрипции:
1. Общие факторы транскрипции обеспечивают сродство
промотора к РНК – полимеразе.
2. Специфические транскрипционные факторы – активаторы
связываются с участками ДНК энхансерами - при этом
усиливается транскрипция. Механизм: связываясь с
активаторами, энхансеры изменяют конформацию
хроматина в области прилежащего гена.
3. Активаторы связываются с сайленсерами - тогда
ослабляется транскрипция.
10.
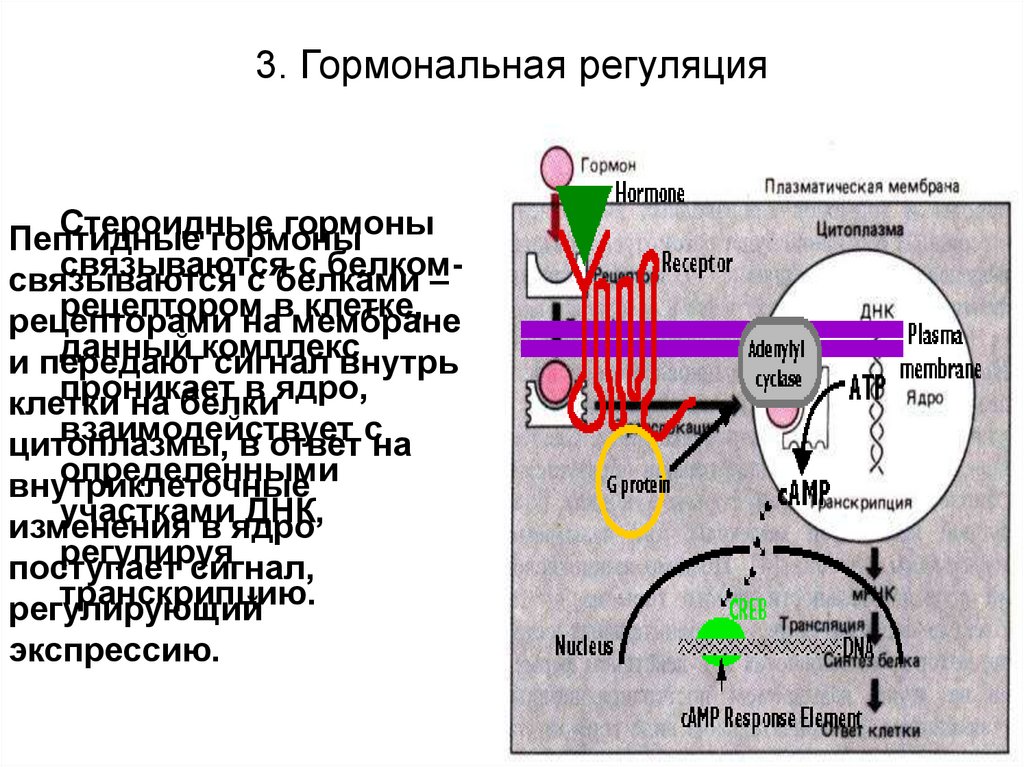
3. Гормональная регуляцияСтероидные
гормоны
Пептидные
гормоны
связываются
с белкомсвязываются
с белками
–
рецептором
клетке,
рецепторами
навмембране
данный комплекс
и передают
сигнал внутрь
проникает
в ядро,
клетки
на белки
взаимодействует
цитоплазмы,
в ответ сна
определенными
внутриклеточные
участками
ДНК,
изменения
в ядро
регулируя
поступает
сигнал,
транскрипцию.
регулирующий
экспрессию.
11.
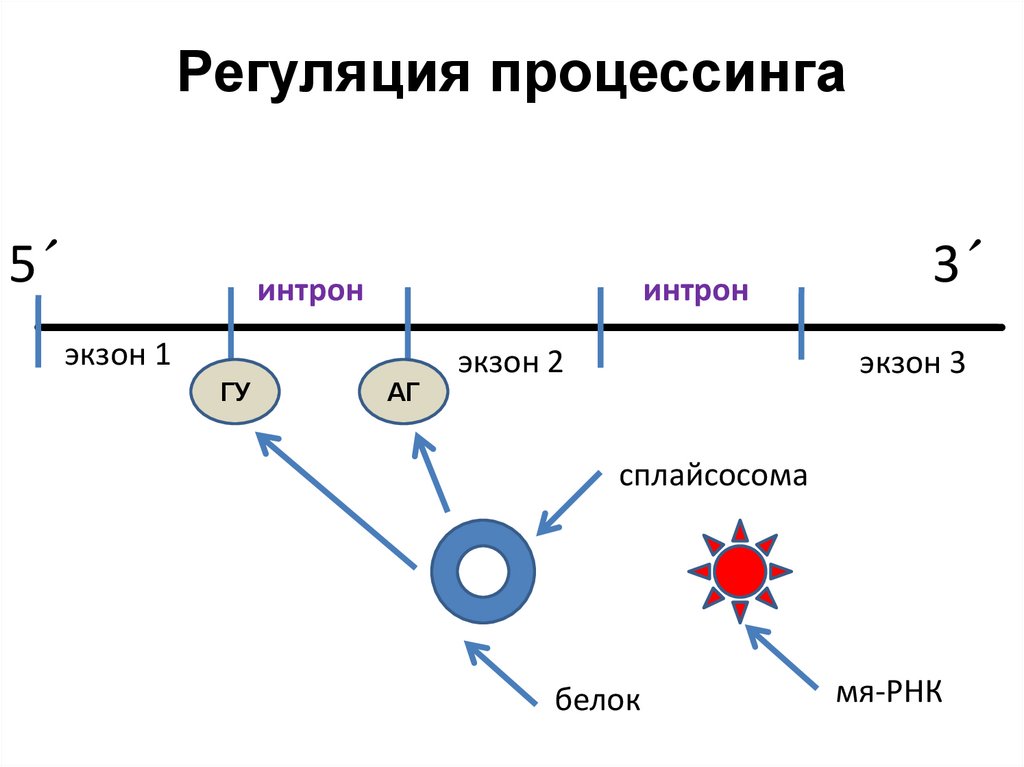
На уровне процессинга
• Точность сплайсинга обеспечивается
взаимодействием белков-сплайсинга (SR) и мяРНК (комплекс сплайсосома).
• Сплайсосома связывается
с концевыми
участками интрона ( 5′ -конец интрона почти
всегда содержит ГУ, а 3′- конец интрона
содержит АГ), что способствует точному
вырезанию
интронов
ферментами
рестриктазами.
12.
Регуляция процессинга5´
интрон
интрон
экзон 1
ГУ
АГ
экзон 2
3´
экзон 3
сплайсосома
белок
мя-РНК
13.
На уровне трансляции1. Редактирование РНК. Отбор определенных молекул и-РНК
и блокировка доступа
несоответствия.
к
рибосомам
в
случае
их
2. Общий контроль: факторы инициации соединяются с КЭП
структурой на 5-конце и-РНК, в результате происходит
соединение с малой субъединицей рибосомы, другой набор
белков
присоединяется
к
полиаденилатной
последовательности на 3-конце. В этом случае и-РНК является
активно транслируемой.
3. Фосфорилирование
белков
(факторов
инициации)
ферментами, что приводит к нарушению связывания
метионин-тРНК с малой субъединицей рибосомы.
4. Негативная
регуляция:
синтезируемый
полипептид
связывается с собственной м-РНК и блокирует дальнейший
синтез.
5. Участие микро-РНК
6. Гормональная регуляция
14.
Посттрансляционная регуляцияИзменение конформации белков – важнейший способ
изменения их биологической активности!
Обеспечение правильного фолдинга и рефолдинга
принадлежит белкам - шаперонам.
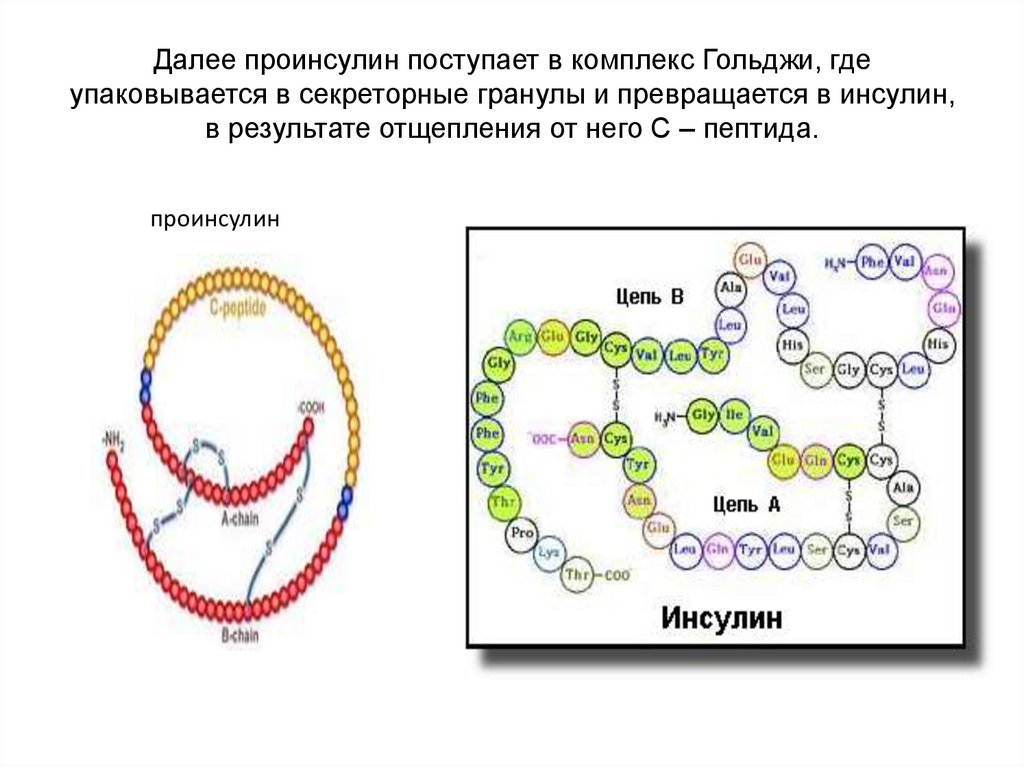
• Рассмотрим на примере синтеза белка инсулина. Нативная
структура инсулина образована двумя полипептидными
цепочками, соединенными дисульфидными мостиками: α цепь –
состоит из 21 аминокислоты, β – цепь – из 30 аминокислот.
Инсулин образуется из препроинсулина в результате
посттрансляционной
модификации.
Ген
препроинсулина
расположен в 11 хромосоме и представлен в геноме одной
копией. После синтеза препроинсулин поступает в ЭПР, где от
него отщепляется лидирующая последовательность — Nконцевой фрагмент, содержащий 24 аминокислотных остатка и
образуется проинсулин. Проинсулин состоит из α- цепи и β –
цепи и С- пептида (31 аминокислотный остаток). См. рис. на
след. слайде.
15.
Далее проинсулин поступает в комплекс Гольджи, гдеупаковывается в секреторные гранулы и превращается в инсулин,
в результате отщепления от него С – пептида.
проинсулин
16.
Неспецифическая регуляция геннойактивности
На генном уровне: потеря активности гена в
результате переноса его при хромосомных
перестройках в участки гетерохроматина.
Пример: эффект положения гена W+ (красные
глаза) при инверсии у гетерозиготной самки (W+
/ w) приводит к белоглазости.
На хромосомном уровне: потеря активности
гена в факультативном гетерохроматине.
На геномном уровне: Изменение структуры
хроматина, связанное с деконденсацией.
17.
• Для экспрессии гена необходимадеконденсация хроматина и освобождение
нуклеосомных белков от ДНК. Этому
способствуют:
1) процессы ремоделирование гистонов Н3 и
Н4 (реакции метилирования,
ацетилирования, фосфорилирования под
действием ферментов: метилазы,
ацетилазы, киназы фосфорилирования
соответственно).
2) Метилирование ДНК с участием
ферментов: ДНК-метилтрансферазы,
метил-цитозин-связывающих белков.
18.
Благодарюза
внимание!