







 – группа генов кодирующих белки, участвующие в общем метаболическом пути. Транскрипция")




























 biology
biologySimilar presentations:

")


")





Регуляция экспрессии генов у про- и эукариот
1.
Тема: Регуляция экспрессиигенов у про- и эукариот
2. План лекции:
1.Регуляция активности генов, понятие, сущность,значение вжизнедеятельности организмов (клеток).
2. Типы регуляции активности генов
3. Оперонная модель регуляции активности генов у
прокариот.
4. Особенности регуляции активности эукариотических генов.
5. Нарушения регуляции активности генов и их связь с
патологическими процессами.
3.
• Известно, что гены определяют структуру всехмолекул, из которых состоят клетки живых
организмов, контролируют все метаболические
процессы и содержат программу развития
организма.
• В каждый момент времени любая клетка, от
бактериальной до человеческой, использует лишь
часть своих генов для синтеза определенных
продуктов.
• Невозможна ситуация, когда все гены клетки
работают одновременно. Мы говорим, что те гены,
которые экспрессируются - включены, а те,
которые не экспресируются – выключены. Это
означает, что экспрессия генов регулируется.
4.
• В то же время известно, что в ходеиндивидуального
развития
многоклеточного
организма из оплодотворенной
яйцеклетки
образуются
разнообразные
типы
клеток,
входящих в состав определенных тканей. Но все
клетки, как правило, несут один и тот же набор
генов. В основе этого лежит выборочное
использование генов, то есть регуляция генов.
• Но разных стадиях дифференцировки клетки,
руководствуясь
лишь
отчасти
внешними
сигналами, избирательно используют тот или иной
набор генов, что определяет пути их развития.
5.
Экспрессия гена регулируется не только в ходеонтогенеза, но также и в течении жизни
дифференцированной клетки. Например, клетки
кожи под действием солнечного ультрафиолетового
облучения
вырабатывают
пигмент
меланин.
Структура гена, отвечающего за синтез пигмента, не
изменяется в ответ на обучение,
просто
внеклеточный сигнал – ультрафиолетовые лучи
включает этот ген.
6.
7.
РЕГУЛЯЦИЯЭКСПРЕССИИ ГЕНОВ
У ПРОКАРИОТ
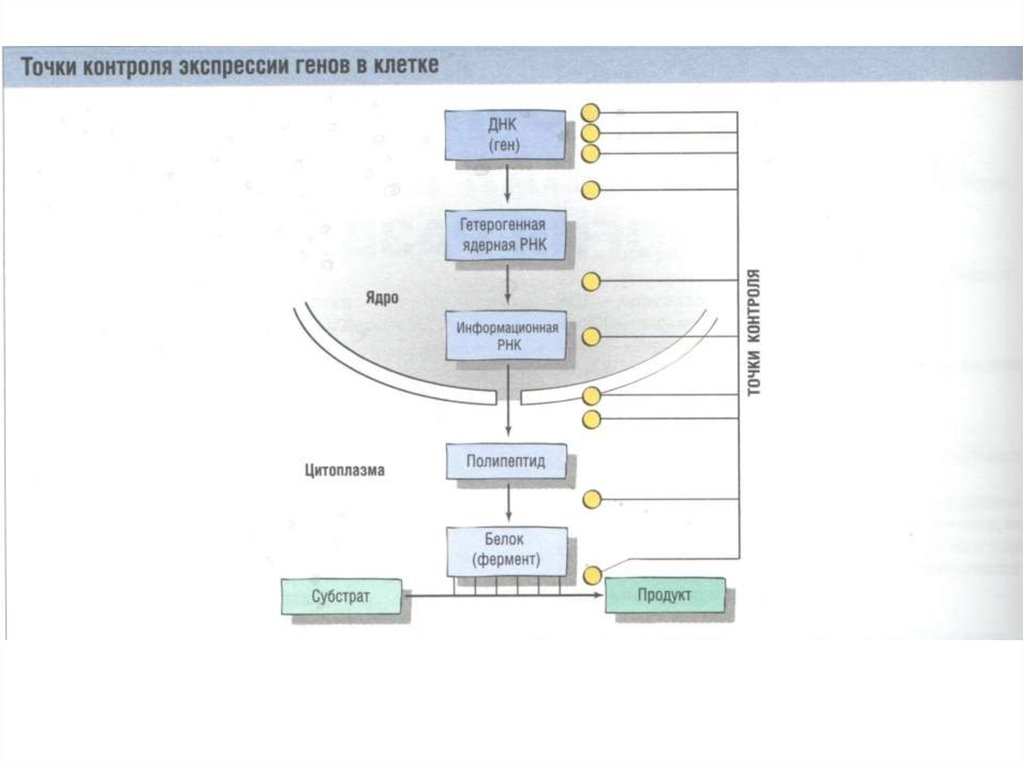
8. Регуляция экспрессии генов осуществляется на различных уровнях:
ТранскрипционныйТрансляционный
Посттрансляционный
9. Регуляция экспрессии генов прокариот
У бактерий гены ферментов,катализирующих ряд
последовательных реакций,
объединяются в одну структурнофункциональную единицу – оперон.
10. Оперон (Ф. Жакоб, Ж. Моно, 1961 г.) – группа генов кодирующих белки, участвующие в общем метаболическом пути. Транскрипция
генов оперона осуществляетсяс общего промотора и регулируется общим
сигналом. В результате образуется единая
полицистронная мРНК
Ж.Моно
Ф.Жакоб
11. В состав оперона входят
Группа сцепленных структурных генов, кодирующихсинтез ферментов для единого метаболического
процесса
Регуляторные участки:
Промотор
Оператор
Терминатор
12. Согласно модели оперона структурные гены делятся на две группы:
Конститутивные гены – транскрипция такихгенов происходит постоянно и не требует
регуляцию
Индуцибельные гены – транскрипция этих
генов регулируется с помощью
специальных белков-регуляторов
(репрессоров или активаторов)
13. Типы оперона
Индуцибельные опероныРепрессибельные опероны
14. Индуцибельные опероны
- регулятором является исходный субстрат (So)цепи контролируемых реакций
- в отсутствие этого субстрата белок-репрессор
имеет высокое сродство к оператору, отчего
РНК-П не может транскрибировать гены
оперона (оперон выключен)
- При накоплении метаболита So в клетке
некоторое количество его связывается с
белкомрепрессором, понижая сродство
последнего к оператору; оперон
«включается» и синтезируются ферменты,
обеспечивающие превращения вещества So.
15. Репрессибельные опероны
- Регулятором служит конечный продукт (Pn) цепиконтролируемых реакций
- В отсутствие этого продукта белок-репрессор имеет
низкое сродство к оператору; поэтому РНК-П
транскрибирует гены оперона – оперон «включен»
и синтезируются ферменты, способствующие
образованию вещества Pn
- При накоплении же данного вещества некоторое
его количество связывается с белком-репрессором
и повышает сродство последнего к оператору –
оперон «выключается», синтез соответствущих
ферментов и образование метаболита Pn
прекращаются
16.
Таким образом, у бактерий используются двапринципиальных способа регуляции
экспрессии генов:
• Регуляция связывания РНК-П с промоторами
(за счет природы промотора, природа сигмафактора РНК-полимеразы, а также, как мы
увидим на примере лактозного оперона,
специального белка САР)
- Регуляция перемещения связавшейся РНКполимеразы от промотора к собственно генам
(при чисто оперонном механизме регуляции)
17. Лактозный оперон
Регуляциянегативная
(белок репрессор)
позитивная
(САР-белок)
18. Механизм негативной регуляция
Лактозный оперон состоит из трехструктурных генов – lacZ, lacY и lacA,
продукты которых необходимы для
использования лактозы в качестве
источника углерода, промотора, с
которым связывается РНК-полимераза, и
операторного участка (оператора), с
которым связывается белок-репрессор –
продукт гена lacI.
19. Механизм негативной регуляция
Ген-регулятор лактозного оперона (lacI)кодирует белок-репрессор. В активной
форме белок-репрессор связывается с
оператором. Оператор – это участок ДНК,
с которым связывается белок-репрессор.
Сам ген lacI в состав оперона не входит,
поскольку его экспрессия осуществляется
с собственного промотора. Это
конститутивный ген, т.е. нерегулируемый.
20. Механизм негативной регуляция
Присоединившись к оператору, репрессорпрепятствует транскрипции структурных
генов Z, Y и А. Таким образом, репрессор
является негативным регулятором; в его
присутствии подавляется экспрессия Z, Y
и А - генов
21.
При избытке лактозы её молекулы связываются ссубъединицами репрессора с образованием
репрессор-индукторного комплекса, в котором
индуктор (лактоза) выступает в роли
аллостерического регулятора, изменяющего
конформацию белка- репрессора, что ведёт к
инактивации последнего. У инактивированного
репрессора резко снижается сродство к зоне
оператора, в результате чего репрессор отсоединяется
от промотора, открывая «вход» для РНК-полимеразы.
Вслед за этим начинается транскрипция оперона, а
затем и синтез ферментов, метаболизирующих
лактозу.
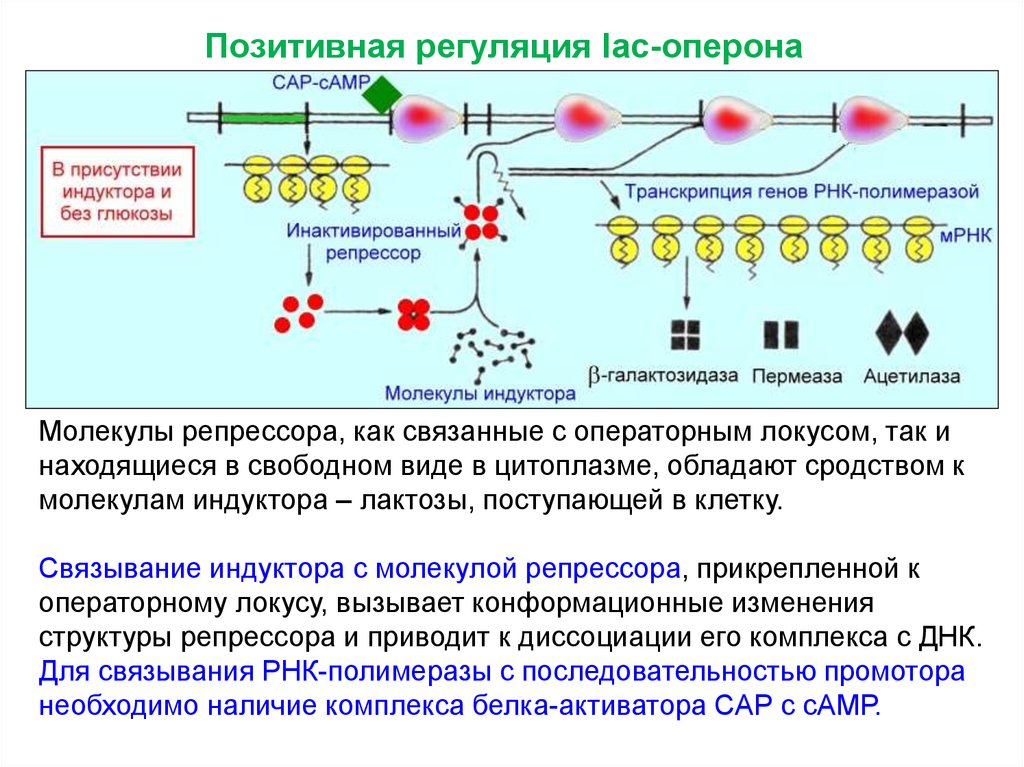
22. Позитивная регуляция
• Почему lac-оперон не транскрибируется(т.е. репрессирован), если в среде находятся
одновременно лактоза и глюкоза?
• Почему в этих условиях не работает индуктор
(лактоза)?
• Почему транскрипция оперона начинается только после
того, как клетки полностью утилизируют глюкозу?
Это пример катаболитной репрессии или глюкозного эффекта,
выражающийся в неспособности клеток E.coli катаболизировать
различные углеводы в присутствии глюкозы, как наиболее
эффективного источника энергии.
23.
Позитивная регуляция lac-оперонаМолекулы репрессора, как связанные с операторным локусом, так и
находящиеся в свободном виде в цитоплазме, обладают сродством к
молекулам индуктора – лактозы, поступающей в клетку.
Связывание индуктора с молекулой репрессора, прикрепленной к
операторному локусу, вызывает конформационные изменения
структуры репрессора и приводит к диссоциации его комплекса с ДНК.
Для связывания РНК-полимеразы с последовательностью промотора
необходимо наличие комплекса белка-активатора САР с сАМР.
24. Позитивная регуляция
• БАК (САР –англ.) – Белок – АктиваторКатаболитных оперонов, который
активируется при соединении с цАМФ
• Образуется комплекс:
цАМФ + белок-активатор (БАК)
• Комплекс цАМФ+БАК связывается со
специфической последовательностью и
усиливает транскрипцию в 20-50 раз.
25. Позитивная регуляция
Комплекс цАМФ+БАК действует какпозитивный регулятор, поскольку его
присутствие необходимо для обеспечения
экспрессии Lac-генов. Таким образом, Lacоперон является объектом как позитивной,
так и негативной регуляции.
26. Репрессибельный оперон
Триптофановый оперонРегуляция
негативная
(белок репрессор)
аттенюация
(окончание
транскрипции
на аттенюаторе)
27. Схема регуляции триптофанового оперона у E.coli
Триптофановый оперонВ триптофановом опероне, как и в лактозном, тоже
имеется двойной механизм регуляции. Во-первых, как
обычно, регулируется перемещение РНК-полимеразы по
оператору. Вторым же объектом регуляции является не
связывание РНК-полимеразы с промотором, а окончание
транскрипции на аттенюаторе.
Как правило, опероны, имеющие аттенюатор, являются
репрессибельными и контролируют синтез того или
иного необходимого компонента – например, редкой
аминокислоты: триптофана, гистидина, фенилаланина.
28.
Триптофановый оперонТриптофановый оперон включает 5 цистронов, кодирующих
четыре фермента заключительного этапа образования
триптофана. При этом последний фермент содержит
субъединицы двух видов, отчего кодируется двумя цистронами.
Вместе с тем, ген предпоследнего фермента цепочки, повидимому, находится где-то вне данного оперона.
Вслед за промотором и оператором в оперонах этого типа
находится т.н. лидерный отдел; именно он оканчивается
аттенюатором.
В процессе транскрипции этого отдела образуется лидерный
участок мРНК. Последний тут же связывает рибосому и начинает
трансляцию с образованием лидерного пептида (ЛП). Ключевая
особенность последнего – среди его 14 аминокислотных
остатков содержатся 2 остатка триптофана, т.е. той самой
аминокислоты, синтез который контролируется опероном.
29.
Триптофановый оперонАналогично, в ЛП фенилаланинового оперонасреди 15 остатков
– 7 остатков фенилаланина, а в ЛП гистидинового оперона – 7
подряд остатков гистидина. Так что механизм аттенюаторной
регуляции во всех этих случаях одинаков.
Когда в клетке достаточно триптофана, то синтез лидерного
пептида идет без задержки: образующая его рибосома не
отстает от РНК-полимеразы. В этих условиях при достижении
РНК-полимеразой аттенюатора с высокой долей вероятности
срабатывает сигнал об окончании транскрипции: РНКполимераза диссоциирует от ДНК и гены не считываются.
30.
Триптофановый оперонТаким образом, триптофан, быстро включаясь в лидерный
пептид блокирует через аттенюаторный механизм синтез
ферментов, необходимых для его образования. Правда,
блокирование это – не полное, т.к. сохраняется небольшая
вероятность того, что РНК-полимераза все же преодолеет
аттенюаторный участок.
Если наоборот, концентрация триптофана в клетке низкая, то
рибосома задерживается с синтезом лидерного пептида и
отстает от РНК-полимеразы. Это так меняет конфигурацию ДНК
или лидерного отдела мРНК, что сигнал об окончании
трансляции на аттенюаторе не срабатывает. Каждая молекула
РНК-полимеразы проходит этот опасный участок и
транскрибирует гены. Т.е. активно синтезируются ферменты,
необходимые для пополнения запаса триптофана в клетке.
31.
Триптофановый оперонЧерез аттенюаторный механизм триптофан может подавлять
активность «своего» оперона, но не до конца.
Более глубокое подавление активности оперона происходит при
очень высокой концентрации триптофана. Тогда реализуется
второй способ регуляции. Триптофан связывается со
специфическим белком-репрессором и повышает его сродство к
оператору триптофанового оперона. Это полностью блокирует
данный оперон.
32.
РЕГУЛЯЦИЯЭКСПРЕССИИ ГЕНОВ
У ЭУКАРИОТ
33.
Регуляция экспрессии генов у эукариотимеет 2 вида регуляции:
1.кратковременная, обеспечивающая
адаптацию
организма
к
возможным
изменениям
окружающей среды
2.длительная,
стабильная,
определяющая дифференцировку
клеток и разный белковый состав
органов и тканей
34.
Оба вида регуляции проявляются наразных
уровнях
реализации
генетической информации:
на уровне ДНК,
на транскрипционном уровне,
на посттранскрипционном уровне
на трансляционном уровне,
на посттрансляционном уровне
35.
Генный или претранскрипционныйуровень.
Регуляция
осуществляется
путем
изменения количества копий конкретного
гена. В гаплоидном наборе один ген
отвечает за синтез одного белка. Вместе с
тем
конкретный
ген
может
быть
представлен
некоторым
количеством
копий. Изменение количества копий гена
может привести к изменению количества
структурного или модуляторного белка в
клетке.
В
результате
нарушается
нормальное течение онтогенеза.
36.
Транскрипционная регуляцияэкспрессии генов эукариот
Транскрипционная регуляция включает в себя
механизмы предотвращающие синтез мРНК.
37. Транскрипционная регуляция экспрессии генов эукариот
Посттранскрипционная регуляцияэкспрессии генов эукариот
Посттранскрипционная регуляция включает в
себя механизмы контролирующие или
регулирующие мРНК после синтеза.
Альтернативный сплайсинг
Скорость транспорта мРНК через ядерную
мембрану
Время жизни мРНК
38. Посттранскрипционная регуляция экспрессии генов эукариот
Трансляционная регуляцияэкспрессии генов эукариот
Трансляционная регуляция включает в себя
механизмы предотвращающие синтез белка.
Как правило, очень часто речь идет о
белковых факторах необходимых для
трансляции
Предотвращение рибосом от связывания с
мРНК
Факторы инициации трансляции
39. Трансляционная регуляция экспрессии генов эукариот
Посттрансляционная регуляцияэкспрессии генов эукариот
Посттрансляционная регуляция включает в
себя механизмы действующие на белок
после его синтеза
Активация белков
Некоторые белки не активны после синтеза,
они должны пройти пострансляционной
модификации
Много белков активируются после
фосфорилирования
Деградация белков