





 ЗФ. Из 44 зубов 3/3I1/1C4/4P3/3M/ Первые Prosimi появились в палеоцене")





. Древнейшие антропоиды: кватрания и")


 и заняла время около 10 млн. лет.")

; - кисть, приспособленная к изготовлению орудий; - высокоразвитый мозг. 1.")














 (2000г.) Кения 5,6-6,2 млн.лет")







;")
")



. 3,9 -4,2 млн. лет")


. Кения, Танзания, Эфиопия. 3,0-3,9-4,4 млн. лет")










")





 Australopithecus africanus — «южная обезьяна")
















 Вост. Африка.")









.")





")






 biology
biologySimilar presentations:










Палеонтология отряда приматов
1. Тема: Палеонтология отряда приматов.
1. Методы датировок палеонтологического материала.2. Основные этапы эволюции приматов.
3. Предшественники рода Homo – человекообезьяны австралопитековые.
Литература:
1. Антропология. Я.Я.Рогинский, М.Г.Левин. М., 1978.
2. Антропология. Е.Н.Хрисанфова, И.В.Перевозчиков. М.,
2002.
3.Антропология. В.М.Харитонов, А.П.Ожигова и др.
М.,2003.
4.Елинек. Я. Большой иллюстрированный атлас
первобытного человека. Прага, 1982.
2.
• 5.Эволюционная антропология . М.А.ДерягинаМ., 1999.
• 6.Введение в теорию антропологии и
археологию палеолита. М.В.Харитонов, М.,
1998.
• 7.Сайт Интернета: Происхождение и эволюция
человека.
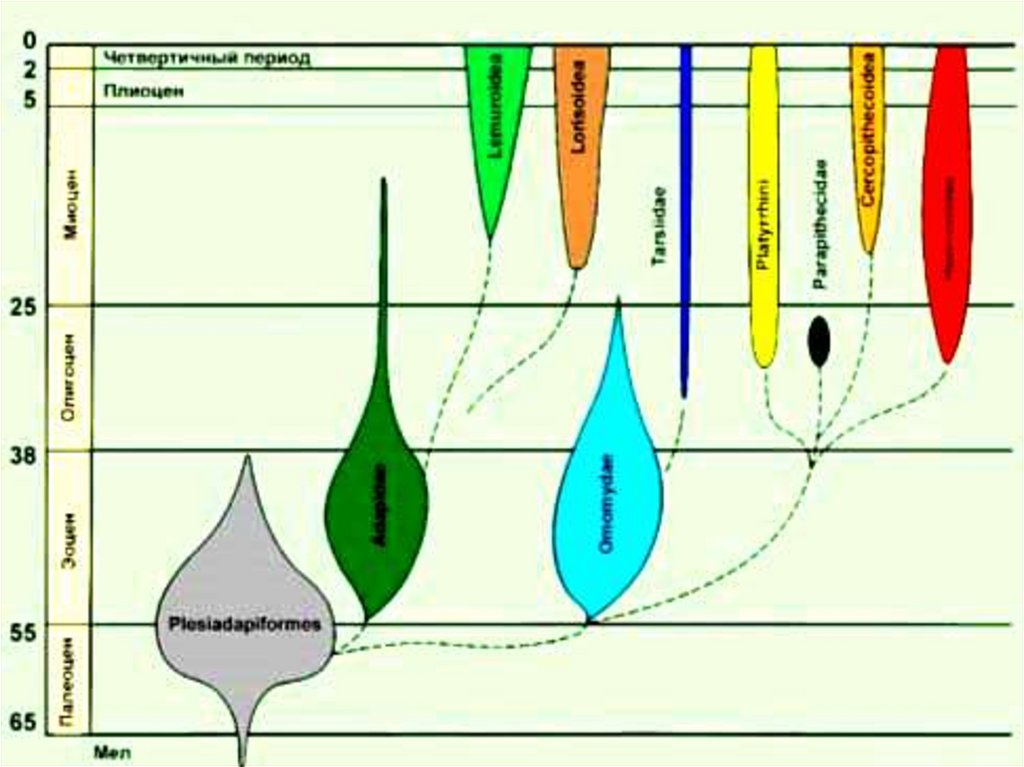
3. 1.Методы датировок палеонтологического материала.
• 1.Геохронологическая шкала временикайнозойской эры.
Третичный период
Палеоген (38 млн. лет)
Неоген (24 млн. лет)
Палеоцен
миоцен
Эоцен
Плиоцен
Олигоцен
4.
Четвертичный периодПлейстоцен (2-3 млн. лет)
неолит
мезолит
палеолит
Голоцен (10 тыс. лет)
Современная эпоха
5.
• 2.Гляциологический метод датировок.• Чередование периодов оледенения (гляциалы) и
отступления ледников межледниковье (интергляциалы)
и влажных периодов (плювиалы) и сухих (аридных).
• 3.Палеонтологический метод датировок основан на
процессе эволюции органического мира.
• 4.Археологический метод датировок применим для
четвертичного периода. Основан на скорости
радиоактивного распада элементов, содержащихся в
породах и органических остатках.
• А) Различают K-Ar часы. Период полураспада 1,3 млн.
лет. Позволяет датировать костные остатки более 1 млн.
лет.
• Б) радиоуглеродный, определяет возраст 30-40000 лет.
6. 2. Основные этапы эволюции приматов. 1.Отряд приматов выделился в конце мелового периода мезозойской эры и эволюция протекала в
кайнозойскую эру.2.Предками были древнейшие насекомоядные
млекопитающие сходные с современными тупайями,
что подтверждается и современными молекулярными
методами.
3. Эволюция была направлена на развитие мозга,
зрительного анализатора, повышенная способность к
хватательной функции кисти-стопы. Все это вело к
появлению типично обезьяньих признаков. Данные
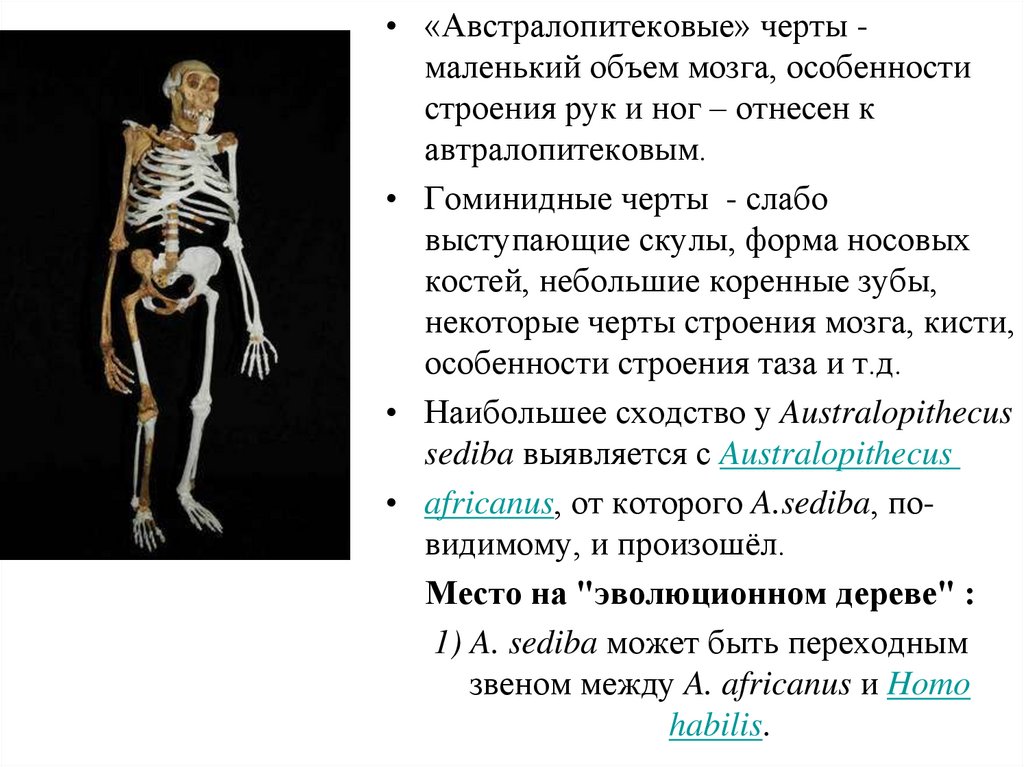
филогенетические тенденции реконструируются как
на палеонтологическом материале, так и на
сравнительно-анатомической.
7. Известна олигоценовая тупайя – анагале - (пустыня Гоби) ЗФ. Из 44 зубов 3/3I1/1C4/4P3/3M/ Первые Prosimi появились в палеоцене
в процессе первойрадиации - плезиодаписовые (известно около 60 видов и 20
родов). Скелет обычного позднепалеоценового примата
Plesiadapis
8. плезиадапис
9. Зубная формула такая же как и у лемуров, напоминали грызунов. Современные тупайи их потомки. Второе расщепление привело к
появлению в Европе иС.Америке эуприматов (начало эоцена), предки
лемуроидов и в это время наблюдается расцвет Prosimii.
Населяли С.Америку, Европу, Азию и Африку.
Эуприматы
Адапиды (предки совр.
лемуров, иск. форма нотарктус)
Омомиды (предки тарзиформ)
10. Скелет среднеэоценового адапида Smilodectes и череп эоценового омомида Tetonius дают хорошее представление о ранней стадии в
эволюциилемуроподобных приматов и предков долгопятов.
11. Smilodectes
12. Третья радиация – рубеж эоцена и олигоцена появились древнейшие высшие приматы – антропоиды. К этому времени большая часть
линий полуобезьян вымирает.Причины: похолодание и не выдержали конкуренции с
грызунами и высшими приматами.
Предпочтительна тарзоидная гипотеза происхождения
высших приматов, генеологическая линия:
плезиапиды
омомиды
антропоиды
Монофилитическая гипотеза (???) происхождения всех
обезьян. (В конце эоцена формируются современные
очертания материков).
Древнейшие формы ископаемых олигоценовых приматов:
амфипитек (Бирма, 40-44 млн. лет), пандаунгия (признаки
полуобезьян и собственно обезьян).
13.
14. Коллекция олигоценовых высших приматов – группа Файюма (Египет, экспедиции Саймонсона). Древнейшие антропоиды: кватрания и
олигопитек. Кромених известны апидиум, парапитек, проплиопитек (близок
гиббонам), египтопитек, мезопитек (Греция, близок
макакам) и др. З.Ф. 2-1-2-3 или 2-1-3-3.
Следующее расщепление привело к разделению узконосых
обезьян на высших и низших. Видимо, это событие
произошло не позднее 23-20 млн. лет назад в Африке и
связано с экологической дивергенцией. Древнейшие
мартышкообразные – викториопитек (20 млн. лет).
С проплиопитековыми связывают эволюцию гоминоидов
– четвертая радиация.
17 млн. лет назад установилась сухопутная связь Африки
и Азии, обезьяны проникли в Азию и Европу.
15. В миоцене различают две формы высших антропоморфных обезьян: дриоформы и рамаформы, разделение их датируют 12-16 млн. лет
назад.Дриопитеки (дриос – дуб) – большая группа
человекообразных ископаемых форм близки шимпанзе и
горилле (Европа, Африка). Ископаемые формы: д.
фантанова, д. дарвинов, проконсул и др.
Рамапитеки (12-7 млн. лет назад, Индия, Пакистан).
Для построения филогенетических ветвей используют
метод электрофореза белков, гемоглобинов (сравнение
последовательности аминокислот в белках у разных
форм); гибридизация ДНК и др. По молекулярным часам
расщепление на узконосых и широконосых произошло 30
млн. лет назад.
16.
• Проконсул- одна из формдриопитековых
17. Эволюция человеческой линии связана с предком типа шимпанзе (бонобо) и заняла время около 10 млн. лет.
18. Согласно молекулярным данным примерно 10 млн лет назад отделилась ветвь оранга, а затем 5-7 млн. лет назад одновременно
разделились ветви человека, шимпанзе игориллы.
Эволюционные линии, ведущие к человеку и шимпанзе,
разделились примерно 5,5 - 6,5 млн. лет назад (или,
возможно, несколько раньше - до 8 млн. лет).
"Человеческая" линия, или семейство Hominidae,
характеризуется важнейшим общим признаком бипедализмом (хождением на двух ногах). Понятно, что
переход к двуногому хождению был связан с
существенными изменениями образа жизни. Поэтому
возникновение нового семейства Hominidae - это
одновременно и формирование новой адаптивной зоны.
19. Гоминидная триада: - прямохождение (бипедия); - кисть, приспособленная к изготовлению орудий; - высокоразвитый мозг. 1.
Прямохождение.Гипотезы:
Миоценовое похолодание: в середине и конце миоцена
в результате глобального похолодания климата
произошло значительное сокращение площадей
тропических лесов и увеличение площади саванн, что
явилась причиной перехода части гоминоидов к
наземному образу жизни.
Однако, известно, что древнейшие известные
прямоходящие приматы жили в тропических лесах.
20.
• Трудовая концепция: согласно широкоизвестной трудовойконцепции Ф. Энгельса и ее позднейшим вариантам,
возникновение прямохождения тесно связано со специализацией
руки обезьяны для трудовой деятельности - переноса предметов,
детенышей, манипулирования пищей и изготовления орудий. В
дальнейшем труд привел к возникновению языка и общества.
Однако, по современным данным, прямохождение возникло
намного раньше изготовления орудий. Прямохождение возникло
не менее 6 миллионов лет назад у Orrorin tugenensis, а
древнейшие орудия из Гоны в Эфиопии имеют датировку лишь
2,7 млн лет назад.
Концепция К. Оуэна Лавджоя, прямохождение возникло в связи
с особой стратегией размножения гоминид. Согласно Лавджою,
прямохождение возникло еще в тропическом лесу, а в саванны
переселились уже двуногие гоминиды.
Вероятнее всего, в эволюции действовала не одна причина,
а целый их комплекс.
21. Модель Лавджоя: «адаптивный комплекс» ранних гоминид
22.
23.
• 2. Происхождение руки, приспособленной кизготовлению орудий.
• Трудовой комплекс:
– Сильное запястье. У австралопитеков, начиная с
Australopithecus anamensis, строение запястья промежуточное
между обезьянами и человеком. Практически современное
строение наблюдается у Homo habilis 1,8 млн лет назад.
– Противопоставление большого пальца кисти. Признак
известен уже 3,2 млн лет назад у Australopithecus afarensis и
Australopithecus africanus. Он был полностью развито у
Paranthropus robustus и Homo habilis 1,8 млн лет назад.
Наконец, было своеобразным или ограниченным у
неандертальцев Европы около 40-100 тыс. лет назад.
– Широкие конечные фаланги пальцев. Очень широкие фаланги
были у Paranthropus robustus, Homo habilis и всех более
поздних гоминид.
– Прикрепление мышц, двигающих пальцы почти современного
типа отмечено у Paranthropus robustus и Homo habilis, но у
них есть и примитивные черты.
24.
• Кости кисти у древнейших прямоходящихгоминоидов (Orrorin tugenensis, Ardipithecus
ramidus kadabba, Australopithecus anamensis и
Australopithecus afarensis) имеют смесь
признаков человекообразных обезьян и
человека. Скорее всего, эти виды могли
использовать предметы как орудия, но не
изготовлять их. Первые изготовители
настоящих орудий - Homo habilis. Вероятно,
орудия изготовляли и южноафриканские
массивные австралопитеки Paranthropus
robustus.
Итак, трудовая кисть в целом
сформировалась около 1,8 млн лет назад
25.
• 3. Высокоразвитый мозг.• Типичные признаки мозга человека следующие:
– Большие общие размеры мозга. У австралопитеков
размер мозга был как у современных шимпанзе.
Бурный рост размеров происходил у Homo habilis
около 2,5-1,8 млн лет назад, а у позднейших гоминид
наблюдается плавное увеличение до современных
значений.
– Специфические поля мозга - зоны Брока и Вернике и
другие поля начали развиваться у Homo habilis и
архантропов, но полностью современного вида
достигли, видимо, только у современного человека.
26.
– Строение долей мозга. У человека значительно развитынижняя теменная и лобная доли, острый угол схождения
височной и лобной долей, височная доля широкая и округлая
спереди, затылочная доля относительно небольшая, нависает
над мозжечком. У австралопитеков строение и размеры
мозга были такие же, как и у человекообразных обезьян.
Строение мозга Homo habilis было примитивным, но сам
мозг имел большие размеры. Для позднейших гоминид
характерно плавное приближение к современному строению
мозга.
• Гоминидный тип мозга возник около 1,8 млн лет назад
или несколько раньше, причем увеличение размеров
предшествовало его усложнению.
• Полностью современный мозг сформировался только
около 25 тыс. лет назад
27. 3. Предшественники рода Homo – человекообезьяны - австралопитековые. Три основных группы, в каждой из которых выделяется
3. Предшественники рода Homo – человекообезьяны австралопитековые.Три основных группы, в каждой из которых выделяется
несколько видов:
Ранние австралопитеки - существовали с 7 до 4 млн лет
назад, обладали наиболее примитивным строением.
Выделяют несколько родов и видов ранних
австралопитеков.
Грацильные австралопитеки - существовали с 4 до 2,5
млн лет назад, имели сравнительно небольшие размеры и
умеренные пропорции. Обычно выделяют один род
Australopithecus c несколькими видами.
28.
• Массивные австралопитеки - существовали с 2,5 до 1млн. лет назад, были очень массивно сложенными
специализированными формами с крайне развитыми
челюстями, маленькими передними и огромными
задними зубами. Массивные австралопитеки
выделяются в самостоятельный род Paranthropus с
тремя видами.
• Первая находка австралопитековых 1924 г. Раймонд
Дарт «Бэби из Таунга» (Трансвааль, ЮАР), детеныш 6
лет, объем мозга 520см3, далее 1936г. (Брум, пещера
Стеркфонтейн) взрослая особь, массивного облика);
1938-1939гг. (Брум) – робустус и т.д.
29. Ранние австралопитеки
Сахелянтроп (Тумай)6-7 млн. лет
2001 г. в месте, называемом ТоросМенелла, в пустыне Дьюраб в
пустынных землях северного Чада,
недалеко от южного края Сахары.
Сахелантроп, по-видимому,
передвигался на двух ногах.
Размер головного мозга очень
небольшой (~ 350 см.куб.), а
черепная коробка удлинена, что
характерно, скорее, для обезьян.
30. Sahelanthropus tchadensis Сахелантроп
31.
• Останки найдены в Республике Чад в ТоросМеналла.• Шесть находок гоминид включили :
полный череп, получивший имя Тумай (Toumai),
два фрагмента нижних челюстей
три изолированных зуба.
• Находки характеризуются большим количеством
специфических особенностей и потому были
выделены в новый род и вид Sahelanthropus
tchadensis
• Череп Сахелянтропа – самый древний целый
череп прямоходящего существа.
32.
• О прямохождении говорит:расположение большого затылочного отверстия,
которое смещено вперед;
базион находится на линии между отверстиями
сонных артерий. Такое положение
промежуточно между вариантами, характерными
для четвероногих человекообразных обезьян и
для более поздних прямоходящих гоминид.
33.
34. Оррорин (Orrorin tugenensis) (2000г.) Кения 5,6-6,2 млн.лет
35.
36.
• В 2001 г. Б. Сеню и М. Пикфорд обнаружила вТуген Хиллс 12 костей гоминид, в том числе
правую и левую бедренные кости; находки
представляли по меньшей мере 5 индивидов
обоего пола.
• Большая древность находки в сочетании с
архаичной морфологией дали основание авторам
открытия для описания нового рода и вида
гоминид Orrorin tugenensis.
• В популярной литературе этот австралопитек
известен как "Человек Тысячелетия"
("Millennium Man").
37.
• Фрагменты левой и правой бедренныхкостей имеют множество признаков,
сближающих O. tugenensis с гоминидами и
отличающих их от человекообразных обезьян.
• В целом, признаки бедренных
костей достаточно явно указывают на двуногий
способ передвижения их обладателей.
• В то же время, строение верхних конечностей
указывает на элементы древесной локомоции
(впрочем, как и у более поздних
австралопитеков).
• Зубы в некоторых признаках сближаются с
зубами шимпанзе.
38.
• Ардипитеки (лат. Ardipithecus) — древнийрод гоминид, который жил в раннем плиоцене.
• Отличительные особенности рода ардипитеков - много
общего с африканскими
обезьянами, шимпанзе и гориллами:
• 1) мнение – предки скорее шимпанзе, чем человека.
• 2) мнение – древнейшие предки человека из-за
сходства зубов с зубами австралопитека.
• Сходство с человеком - в стопе ардипитека
имеется добавочная малая берцовая кость (os
peroneum) - присутствует в стопе человека, но
отсутствует в стопе современных человекообразных
обезьян.
• Иногда ардипитеков относят к ранним австралопитекам.
39. Ardipithecus ramidus Ардипитек рамидус
• 4 млн.л.н.• 1995 Эфиопия
40.
• В Арамисе:• 1995 гг. остатки гоминид включили фрагменты
черепов, нижней челюсти детеныша, большое
количество изолированных зубов, а также
фрагменты костей верхних конечностей по
меньшей мере от 17 индивидов.
• В 1995 г. были обнаружены более 90
окаменевших фрагментов, составивших около
45% скелета взрослого индивида, в том числе
части черепа, позвоночника, рук, таза, ног и
стопы (скелет, получивший прозвище "Арди").
41.
• Это древнейший скелет прямоходящегогоминида с большим количеством обезьяньих
признаков.
• Сначала находкам дали наименование:
• Australopithecus ramidus,
• а позже выделили в новый род –Ardipithecus.
42. Ардипитек кадабба (A. kadabba 5,2-5,8 млн.лет 1997-2000 гг. в долине Аваш);
43. Ардипитек кадабба (Ardipithecus ramidus kadabba)
44.
• Ардипитек кадабба• (Ardipithecus ramidus kadabba) представляет ещё
один род поздномиоценовых гоминид
• Его останки были найдены совместной группой
эфиопских и американских антропологов во
главе с американским исследователем
Йоханессом Хайле Селассие, взявшего такую
экзотическую фамилию в честь известного
эфиопского императора.
• Ранее, в 1995 г. были найдены останки
подобных гоминид. Их отнесли не только к
новому виду, но даже роду гоминид и присвоили
имя Ardipithecus ramidus .
45.
• Морфология останков довольно близкоприближалась к морфологии найденных ранее
представителей этого же рода, но всё же имелись
некоторые различия.
• После тщательного анализа и изучения вновь
найденным останкам ардипетека было присвоено
ещё и имя кадабба ( кадабба в переводе с
афарского означает основатель рода), а новый
гоминид выделен в новый вид Ardipethecus
kadabba.
46.
• Ardipithecus kadabba одно из древнейших прямоходящихсуществ, от которого сохранились кости ног и рук.
• Находки включают:
правую половину нижней челюсти,
изолированные зубы,
ключицу,
фрагменты двух плечевых и локтевой костей,
две фаланги кисти, проксимальную фалангу стопы – кости по
меньшей мере от 5 индивидов;
строение зубов (особенно - клыков) промежуточное между
человекообразными обезьянами и человеком, но ближе всё же к
человеку;
строение длинных костей указывает на вертикальное положение
тела при ходьбе.
47. Южная озёрная обезьяна Австралопитек анамский (A. anamensis). 3,9 -4,2 млн. лет
По некоторым признакам зубов этот вид являетсяпромежуточным между Ardipithecus ramidus и
Australopithecus afarensis. Жили в сухих лесах. Их останки
найдены в Кении
48.
• Австралопитек анамский (лат. Australopithecusanamensis) — ископаемый вид рода австралопитеков,
(Восточная Африка ок. 4 млн. лет назад) - наиболее
древний из видов австралопитеков, известных науке.
1965 г. - первая находка данного вида дистальный фрагмент левой плечевой кости,
обнаружен в Канапои в процессе работ Гарвардской
экспедиции под руководством Б. Паттерсона.
• Спустя почти 30 лет, в 1994 г. - под руководством М.Г.
Лики здесь был найден ряд окаменелостей,
позволивших описать новый вид ранних
австралопитеков.
• Остатки от ок.100 окаменелостей >20 особей.
• Сочетание понгидных и гоминидных признаков
примерно в равной пропорции.
49.
• Положение вида:• 1) потомок ардипитека;
• 2) предок афарских австралопитеков.
• При развитом прямохождении Australopithecus
anamensis иногда, вероятно, ходил на
четвереньках с опорой на согнутые пальцы рук
(о чем говорит строение локтевой кости).
50. Грацильные австралопитеки: Австралопитек афарский (Australopithecus afarensis). Кения, Танзания, Эфиопия. 3,0-3,9-4,4 млн. лет
Найдены остатки более 300особей (в том числе известная
"Люси").
51.
• Австралопитек афарский (лат. Australopithecusafarensis) — вымерший вид австралопитека,
семейство гоминид, (ок. 4 млн. лет назад).
• Вымерли афарские австралопитеки ок. 2,5 млн. лет
назад
Положение:
предки прямо или косвенно други австралопитецины и
род Homo.
• Афарские австралопитеки хорошо известны по
раскопкам в Хадаре, проводившимся в 1973-1977гг.
Международной Афарской Исследовательской
Экспедицией под руководством И. Коппенса, М. Тайеба
и Д. Джохансона.
• Обнаружено более 240 остатков гоминид,
принадлежавших как минимум 35 особям.
52.
• Морфологические особенности:• Наиболее известная находка AL 288-1 - «Люси» (40%
скелета Люси, 30.11.1974 г.) - Д. Джохансоном и Т.
Грэем в Када Хадар (с датой 3,18-3,2 млн.л.)
• Строение черепа:
низкий, с покатым лбом;
надбровье мощное;
височные линии лобной кости образуют
низкий сагиттальный гребень;
затылок преломленный, хорошо выражен затылочный
гребень;
основание черепа изогнуто очень слабо,
подобно понгидам.
в затылочной норме череп A. afarensis более низкий
53.
расширенный в нижней части, чем у A. africanus, чтоявляется более примитивным вариантом;
в этом отношении A. afarensis больше похож на
массивных австралопитеков.
эндокран внешне очень похож на Э. человекообразных
обезьян, однако на нем нет отпечатка полулунной
(«обезьяньей») борозды, обычно хорошо заметного на
эндокранах понгид, у которых данная борозда
ограничивает затылочную долю, тогда как у
современного человека ее нет;
теменная и височная ассоциативные зоны увеличены,
что, несомненно, является прогрессивным признаком.
54.
• Строение зубной системы:значительные размеры резцов и большая разница
размеров резцов I1 и I2, из которых первый больше;
верхние клыки сильно выступают над соседними
зубами, их размеры и морфология промежуточны
между понгидным и гоминидным вариантами,
приближаясь все же к последнему; клыки достаточно
заостренные на вершине, их корни наклонены назад;
верхние клыки самцов A. afarensis больше, чем у самок;
моляры A. afarensis достаточно большие, иногда их
размеры даже описываются как умеренно мегадонтные;
но все же намного меньше, чем у массивных
австралопитеков.
55.
• Комплекс строения конечностей :• Строение "проксимальной части» части
лучевой кости свидетельствует о том, что
афарские австралопитеки, либо их
непосредственные предки, могли ходить на
фалангах пальцев рук.
• Строение таза Australopithecus afarensis - крайне
широкий и короткий.
• Пропорции таза "Люси" - «ультрагоминидны» специализация к двуногой ходьбе у A.
afarensis зашла уже очень далеко.
56.
• Признаки, наблюдающиеся в скелете A. afarensis,свидетельствующие о иных способах
передвижения, могут быть либо наследием,
доставшимся от предков, либо говорят о
широком спектре типов передвижения,
использовавшихся этими австралопитеками,
хотя прямохождение, видимо, было среди них
преобладающим типом.
• Морфология стопы также свидетельствует о
разнообразии видов локомоции,
использовавшихся этим видом; в этом
отношении грацильные австралопитеки весьма
своеобразны и отличаются как от понгид, так и
57.
• В целом руки A. afarensis были несколько удлиненыотносительно ног, соответствующий индекс имеет
значение, промежуточное между значениями,
известными для человекообразных обезьян и
современного человека; также, ноги относительно роста
были несколько короче, чем у современного человека.
• Такие пропорции, видимо, являются следствием
незавершенного перехода к двуногому прямохождению,
что, как уже было сказано, отражено во многих деталях
морфологии костей конечностей.
• Видимо, ходьба афарских австралопитеков была
медленнее, а величина дневного перехода была намного
меньше, чем у современного человека.
58.
• Размеры тела хадарских A. afarensis чрезвычайноварьируют.
• Разница отражает, по-видимому, половой
диморфизм, который оказывается равным
таковому орангутанов и горилл и большим, чем
у шимпанзе и человека.
• Рост маленьких особей едва превышал метр,
тогда как у больших достигал более полутора
метров;
• вес варьировал соответственно от примерно 30
до 45 кг, возможно даже – до 55 кг.
• Australopithecus afarensis - наиболее вероятный
предок человеческой линии эволюции.
59.
Афарский австралопитекД. Джохансон
Сейчас хорошо изучены остатки одного из ранних австралопитеков
– афарского, найденного Д. Джохансоном в Эфиопии.
Размеры: 110-120 см, с двуногой походкой и зубами, похожими на
человеческие, жила 3,5-4 млн. лет назад.
60. Австралопитек афарский
Обезьяньи признаки:вытянутое
(прогнатное) лицо, U
- образное нёбо (с
параллельными друг
другу рядами
коренных зубов, как
у высших обезьян,
"параболическое"
нёбо совр. человека);
объем мозгой
коробки (430 куб. см
Сапиентный
признак- хождение
на двух ногах.
61. Kenyanthropus platyops (плосколицый)
Kenyanthropus platyops (плосколицый)• 1999 г. Кения, западный берег Турканы, 3,5 млн. лет.
• М.Лики: этот вид мог жить в одном биотопе с
Australopithecus afarensis и при этом не конкурировать с
ним, занимая другую нишу (австралопитеки, видимо,
ели что-то более жесткое).
62. Kenyanthropus platyops
63.
• Кениантропы (лат. Kenyanthropus) — древнийрод гоминид, живший в плиоцене около 3,53,2 млн лет назад.
• Найден череп и другие окаменелости;
вероятно, Kenyanthropus platyops был
специализированным видом, не входившим в
прямую родословную рода Homo.
• На черепе мозаика морфологических признаков,
в отдельности встречающихся и у других
австралопитеков, но в определенном сочетании
характеризующая именно K. platyops.
64.
• Мнение учены - форма лицевого скелетапозволяет сближать Kenyanthropus platyops с
более поздним Homo rudolfensis.
• Характерная черта - значительная ширина и
уплощенность лица
65.
• A. africanus Юж. Африка(Трансвааль), 3.3 (или даже 3.5) 2.5 млн. лет назад.
• Имеет ряд прогрессивных черт по
сравнению с предыдущим видом:
более округлый череп, больший
объем мозга, менее примитивные
зубы и лицевые кости . Однако
строение конечностей, повидимому, примитивнее, чем у
afarensis.
• Пока нет единого мнения о том,
какой из видов, afarensis или
africanus, дал начало первым людям
(Homo).
66. Графическая реконструкция африканского австралопитека
Таз африканскогоавстралопитека
67. Первая находка «Бэби из Таунга» Раймонд Дарт, 1924 г. (Южная Африка) Australopithecus africanus — «южная обезьяна
Первая находка «Бэби из Таунга»Раймонд Дарт, 1924 г.
(Южная Африка)
Australopithecus africanus —
«южная обезьяна африканская».
68.
• Отличие от афарского австралопитека:имел более обезьяноподобный скелет,
более объёмный череп;
наклон лба сравнительно небольшой;
затылок тоже достаточно вертикальный и плавно
закруглен;
череп при сравнении с A. afarensis выглядит более
округлым, коротким и высоким..
• A. africanus имел более прогрессивный по строению
череп, чем A.afarensis, но в целом более архаичный
скелет.
• Общие размеры тела A. africanus варьировали примерно
от метра до полутора, вес – от 20 до 40 кг.
69.
• Кости ног A. africanus сочетают архаичные ипрогрессивные особенности.
• Большой палец стопы, вероятно, обладал большой
подвижностью.
• Вместе с тем тазовые кости (известные по ряду
находок) по подавляющему большинству признаков
ближе к человеческому варианту, чем к тазу
человекообразных обезьян.
• Форма таза является наиболее сильным аргументом,
подтверждающим двуногую локомоцию
австралопитеков.
70.
• Строение зубов:• наклон резцов и клыков и их небольшие размеры отличают A. africanus от человекообразных обезьян,
делая его больше похожим на человека.
• Строение мозга Australopithecus africanus, судя
по эндокранам,
• близко к шимпанзе и достаточно далеко от
современного человека, однако, все же, мозг этих
австралопитеков более похож на мозг человека, чем
мозг всех видов Paranthropus.
• Рост мозгового отдела черепа в сравнении с лицевым по
темпам был промежуточным между
человекообразными обезьянами и человеком.
• Объем мозга такой же, как и человекообразных обезьян
71. Australopithecus bahrelghazali Австралопитек бахр-эль-газальский
• 3.5 млн.л.н. - 3.0 млн.л.н• 1996 Чад
72.
• В 1996 г. французский палеонтолог Мишель Бруне натерритории древнего речного русла Бахр-эльГазаль в Чаде.
• Находка - собой верхнюю челюсть с семью зубами.
Данная находка самая западная - стала свидетельством
довольного широкого распространения рода.
• Находки мало отличаются от останков афарских
австралопитеков.
• Зубная система - три корня у P1 и P2, тогда как у A.
afarensis имеется 1-2 корня на P1 и 2 корня на P2;
трехкорневые нижние премоляры нехарактерны ни для
одного другого вида австралопитековых.
73. Australopithecus garhi Австралопитек гари
• 2.5 млн.л.н.• 1999 Эфиопия
74.
• А.Garhi («удивительный»).• Представители этого вида являются возможным
«переходным звеном» от рода австралопитеков к
роду людей (Homo).
• Относятся к группе грацильных австралопитеков, то есть
обладают хрупким, не массивным телосложением
• A. garhi - eдинственный австралопитек, с останками
которого были найдены каменные орудия труда и
расколотые кости антилоп;
• судя по датировке и специализированным чертам, нашим
прямым предком не являлся.
• Представлен череп, зубы, кости конечностей.
• Рост 1,2-1,5 м., объем мозга около 440 см3.
• Размеры передних зубов A. garhi превосходят зубы др. А..
75. Australopithecus sediba Австралопитек седиба
• 2.0 млн.л.н. - 1.8 млн.л.н., описан в 2010, ЮАР76.
• Австралопитек седиба (Australopithecus sediba) —вид австралопитеков, известный по останкам двух
неполных скелета при раскопках в пещере Малапа
(Malapa) на территории объекта Всемирного
наследия ЮНЕСКО, известного как «Колыбель
человечества», в Южной Африке.
один скелет — подростка мужского пола (голотип),
другой — взрослой особи женского пола.
На сегодняшний день найдено более 130 фрагментов
этих скелетов.
описаны в двух статьях в журнале «Science»
южноафриканским палеоантропологом Ли Бергером и
его коллегами как новый вид ранних предков человека.
новый вид назван Australopithecus sediba («sediba»
означает «источник» или «колодец» в языке сото).
77.
• В 2008 г. в карстовой пещере, на глубине несколькихдесятков метров были обнаружены два разрушенных
скелета (взрослой самки и подростка) и отдельные
кости третьего (младенца).
• Находка останков Australopithecus sediba сделана с
использованием популярного сервиса Google Earth,
позволившего выявить местонахождения
многочисленных карстовых пещер в Южной Африке.
• Дискуссия по поводу положения этой формы:
• Черты Australopithecus sediba – промежуточные между
австралопитеками и Homo (по многим признакам этот
вид оказывается даже более гоминидным, чем хабилисы
Восточной Африки).
78.
• «Австралопитековые» черты маленький объем мозга, особенностистроения рук и ног – отнесен к
автралопитековым.
• Гоминидные черты - слабо
выступающие скулы, форма носовых
костей, небольшие коренные зубы,
некоторые черты строения мозга, кисти,
особенности строения таза и т.д.
• Наибольшее сходство у Australopithecus
sediba выявляется с Australopithecus
• africanus, от которого A.sediba, повидимому, и произошёл.
Место на "эволюционном дереве" :
1) A. sediba может быть переходным
звеном между A. africanus и Homo
habilis.
79.
• 2) A. sediba может быть представителем собственнойюжноафриканской линии гоминизации, параллельной
восточноафриканской, но чуть более поздней;
• (в связи с этим можно вспомнить A. garhi – ещё одну
альтернативную ветвь, дошедшую до уровня
изготовления орудий).
• 3) останки, описанные как A. sediba, могли
принадлежать метисам между поздними грацильными
австралопитеками и уже сложившимися
"ранними Homo", хотя эта версия маловероятна.
80. Массивные австралопитеки: Парантропы, или "робустные" австралопитеки - боковая тупиковая ветвь древних гоминид. Раньше
Массивные австралопитеки:Парантропы, или "робустные" австралопитеки боковая тупиковая ветвь древних гоминид. Раньше
считалось, что они перешли к чисто растительной
диете (поэтому у них такие мощные коренные зубы)
и отказались от охотничьих повадок своих
плотоядных предков – «грацильных»
австралопитеков A.afarensis и A.africanus. В
последнее время, однако, получены данные о том,
что как австралопитеки, так и парантропы были
всеядными формами.
P. robustus (2.0-1.5 млн. лет),
81. Paranthropus aethiopicus Парантроп эфиопский
• 2.5 млн.л.н.• 1968 Кения, Эфиопия
82. P. aethiopicus
83.
• Парантроп эфиопский (лат. Paranthropus aethiopicus) —ископаемый вид рода парантропов («массивных
австралопитеков», Восточная Африка ок. 2,5 млн. лет назад)
• Это наиболее древний из парантропов, известных науке.
• В 1985 г. А. Волкером несколько ниже туфа Локалеи был
обнаружен почти целый череп KNM-WT 17000, известный также
как “Черный череп”, позже были сделаны и другие, менее
комплектные находки, например, фрагмент нижней челюсти
KNM-WT 16005.
• Череп KNM-WT 17000 послужил голотипом для описания нового
вида.
• Считается, что P.aethiopicus произошел от A.afarensis и был
предком P.boisei.
84.
• Есть путаница в определении находок этого вида:• 1) род и вид Paraustralopithecus aethiopicus был описан на
основании морфологии нижней челюсти Omo 18-1967-18 из
Омо (2,5 млн.л.н.);
• 2) череп KNM-WT 17000 и описан как Australopithecus
aethiopicus ;
• Древнейший из массивных австралопитеков и единственный
среди этой группы, имевший резко выступавшие вперёд
челюсти.
• Череп сочетает в себе примитивные признаки, сближающие
его с ранними и грацильными австралопитеками, и
специализированные черты массивных австралопитеков.
• P. aethiopicus - вероятный предок более поздних восточноафриканских массивных австралопитеков
• Paranthropus boisei.
85. P. boisei (2.3 - 1.0 млн. лет) Вост. Африка.
86. Paranthropus boisei Парантроп Бойса
• 2.5 млн.л.н. - 1.0 млн.л.н., 1959 г. Кения,Танзания
87.
• Парантроп Бойса (Paranthropus boisei Mary Leakey,1959)
• Известен:
1) зинджантроп (Zinjanthropus boisei), обнаружен в
1959 г. супругами Лики в восточноафриканских
ущельях Олдувай, Кооби-Фора, Локалеи и ряде др.;
• 2) Paranthropus boisei
• Специфика:
максимального развития достиг челюстной аппарат
("Щелкунчик«);
тенденции "робустной" эволюционной линии
трактуются, как черты крайней специализации к
пережевыванию грубой растительной пищи
88.
• Череп P. boisei хорошо известен по множеству находокхорошей сохранности.
• Черепа очень похожи, их небольшие отличия могут
быть отнесены за счет возраста и пола.
• Череп массивный, с хорошо развитым рельефом.
Конфигурация мозговой коробки своеобразная, при
взгляде сверху напоминает кувшин:
• спереди череп очень расширен благодаря мощному
развитию лицевой области, далее следует сильное
позадиглазничное сужение, лоб сильно скошен и сужен,
потом черепная коробка резко расширяется.
• Огромные скуловые дуги служат мощными
укрепляющими элементами, благодаря своей
винтообразной изогнутости. В затылочной норме череп
невысокий и широкий в нижней части.
89.
• Надглазничный рельеф P. boisei развитдостаточно сильно.
• Размер сагиттального гребня, как и у понгид,
различается у особей разного пола: большой у
самцов и низкий у самок.
• Лицевой скелет P. boisei, как и мозговой, очень
своеобразный.
• Бросается в глаза чрезвычайная расширенность
лица, обусловленная огромными размерами
верхней челюсти и скуловой кости.
• Средняя лицевая и скуловая ширина намного
превышают как остальные широтные, так и
высотные размеры.
90.
• Строение мозга P. boisei довольнопримитивное.
• Эндокраны характеризуются
сильно суженной лобной долей
даже по сравнению с более
древними грацильными
австралопитеками.
• Верхняя челюсть P. boisei очень
большая и массивная, очень
высокая.
• Нижняя челюсть, как и верхняя,
отличается огромными размерами
и массивностью.
91. Paranthropus robustus Парантроп массивный
• 2.5 млн.л.н. - 900 тыс.л.н. 1939 ЮАР92.
• Находка - 1938 г. Роберт Брум в пещере Стеркфонтейн.• (школьник Герт Тербланш нашел в местечке Кромдрай,
расположенном неподалеку от Стеркфонтейна,
вмурованный в породу череп).
• Череп, а также ряд обнаруженных в этом
местонахождении окаменелостей были описаны под
новым родовым и видовым названием Paranthropus
robustus. Впоследствии в Южной Африки были
сделаны сотни находок, относящихся к этому виду
гоминид.
• Череп P. Robustus:
массивный, с толстыми стенками и выраженным
рельефом.
Лоб низкий, покатый;
93.
надбровные дуги расположены почтигоризонтально, отличаясь от покатых
у Paranthropus boisei;
лицевой скелет плоский, даже вогнутый;
скулы массивные, высокие;
верхняя челюсть P. robustus массивная,
укороченная;
нижняя челюсть также чрезвычайно массивна;
клыки большие, мегадонтные (хотя не в такой
степени, как у Paranthropus boisei);
затылочное отверстие сдвинуто вперед, что
является одним из свидетельств прямохождения
94.
Эндокран - имеет примитивную форму, мало отличающуюся отформы человекообразных обезьян;
верхние конечности , как и других австралопитеков,
представляют собой мозаику понгидных и гоминидных
признаков;
морфология тазовых костей, несомненно, ближе к человеческой,
нежели к обезьяньей - очень широкий и развернутый;
стопа имеет гоминидный комплекс;
Находка в Сварткранс останки P. Robustus и множество костяных
фрагментов со следами стертости на конце.
Доказано, что аналогичные следы образуются при использовании
костей для вылавливания термитов;
таким образом, возможно, P. robustus могли использовать
костяные орудия.
95. Paranthropus robustus (2.0-1.5 млн. лет назад, Юж. Африка).
Вывод: для всех "робустных"форм характерен костяной гребень
на темени, служивший для
прикрепления жевательных мышц.
У robustus было крупное лицо,
плоское и круглое, без лба, с
большими надбровными дугами и
очень мелкими передними зубами.
Головной мозг у этого вида имел
объем в среднем около 520 куб.
см.
96. Локализация основных находок австралопитековых
97. Древнейшие находки австралопитековых найдены в области Восточно-африканского рифта: Олдувай; Гаруси; Летали; Кооби Фора;
Илерат; Канапои; Зап. Туркана;Хадар; долина р.Омо и др. Открытые стоянки в лесостепи.
Кроме африканских форм есть азиатские: гигантопитек
(Индия); мегапитек (о.Ява). Австралопитековые жили во
временном промежутке: 4-1 млн. лет назад, возможно
появились раньше.
Физический тип: бипеды (двуногие формы); с малым
мозгом и крупными зубами, на которых толстый слой
эмали; вполне человеческий тип разгибательного аппарата
тазобедренного сустава. Тазовая кость расширена.
Относительная малорослость – 100-156см. Четко выражен
половой диморфизм. Объем мозга 300-800см3. (афарские 440-450; африканские – 510-650; парантропы – 650-800).
98. Понгидные черты – лицо крупное, прогнатное, подбородок отсутствует; нос плоский и широкий, основание черепа слабо изогнуто –
свидетельствопримитивных механизмов дыхания, глотания и
голосообразования; динамика роста и созревания, стадии
развития зубной системы сходны с понгидами.
По современным научным данным афарский морфотип
наиболее соответствует переходному звену от понгид к
гоминидам. Массивные австралопитековые по всей
видимости представляют собой тупиковую ветвь и
вымерли около 1 млн. лет назад, для них характерна была
растительная диета, что не стимулировала развитию
орудийной деятельности и явилось причиной вымирания.
99. Физический тип австралопитековых
100. Ученые Р.Дарт и Л.Лики
101. Плио-плейстоценовые гоминиды (A.afarensis, A.robustus, A.afrikanus, A.boisei, Early Homo)
• Возможная филогенетическая преемственность линийA.afarensis
Homo
A.africanus
A.robustus
A.boisei
102.
103.
• Современныепредставления о времени
существования различных
предков человека
(предполагаемых).
104.
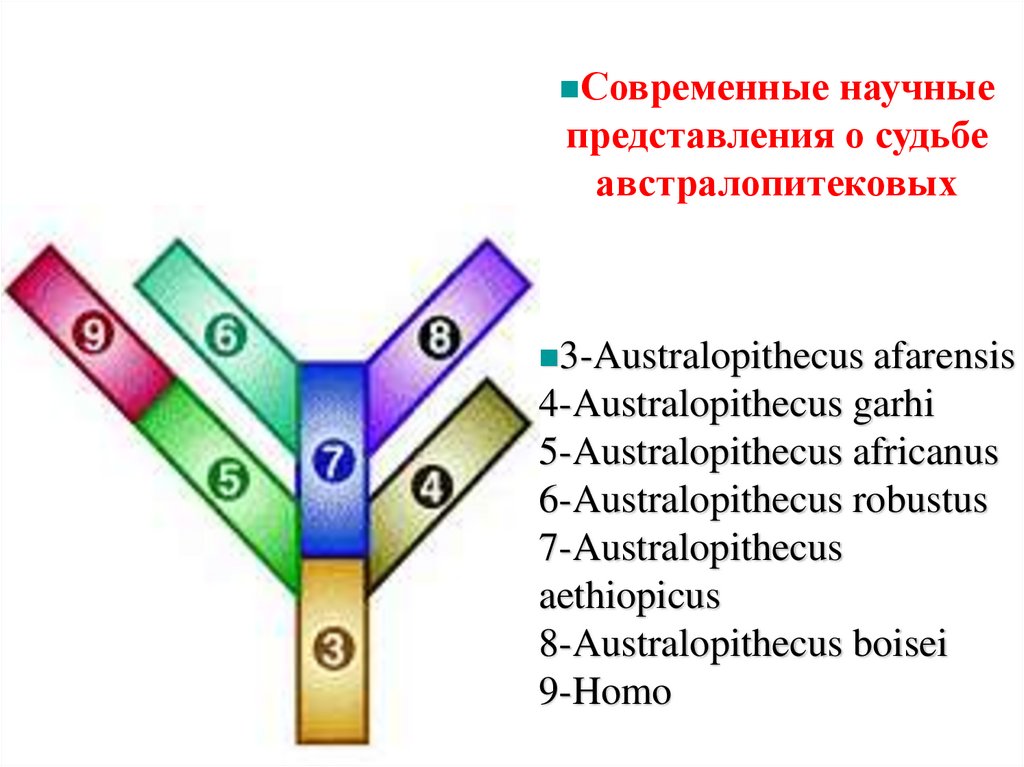
Современные научныепредставления о судьбе
австралопитековых
3-Australopithecus afarensis
4-Australopithecus garhi
5-Australopithecus africanus
6-Australopithecus robustus
7-Australopithecus
aethiopicus
8-Australopithecus boisei
9-Homo
105.
• Ранние австралопитеки включают в себяроды:
• Australopithecus — Australopithecus
anamensis (Leakey, Feibel, McDougal et Walker,
1995)
• Sahelanthropus tchadensis (Brunet at al., 2002)
• Orrorin tugenensis (Senut, Pickford, Gommery,
Mein, Cheboi et Coppens, 2001)
• Ardipithecus ramidus (White, Suwa et Asfaw,
1995).
106.
• Грацильные австралопитеки включают всебя следующие виды:
• Australopithecus afarensis (Johanson, White et
Coppens, 1978)
• Australopithecus bahrelghazali (Brunet, Beauvilain,
Coppens, Heintz, Moutaye et Pilbeam, 1996)
• Australopithecus africanus (Dart, 1925)
• Australopithecus garhi (Asfaw, White, Lovejoy,
Latimer, Simpson et Suwa, 1999)
• Australopithecus sediba (Berger, 2010)
• Kenyanthropus platyops (Leakey, Spoor, Brown,
Gathogo, Kiarie, Leakey et McDougalls, 2001)
107.
• Последняя группа из-за своей специфическойанатомии выделяется в отдельный род —
Paranthropus, насчитывающий три вида:
• Paranthropus aethiopicus (Arambourg et Coppens,
1968)
• Paranthropus boisei (Leakey, 1959)
• Paranthropus robustus (Broom, 1939).