

























































































 biology
biologySimilar presentations:









")
Антропогенез эволюция человека
1. Антропогенез эволюция человека
2.
Царство: Животные
Тип: Хордовые
Класс: Млекопитающие
Отряд: Приматы
Семейство: Гоминиды
3.
• подсемейство Понгины Ponginae– орангутаны, Pongo
• подсемейство Гоминиды Homininae
– триба Гоминини Hominini
• гориллы, Gorilla
• шимпанзе, Pan
– обыкновенный шимпанзе, Pan troglodytes
– бонобо или карликовый шимпанзе, Pan paniscus
• люди, Homo
– человек разумный, Homo sapiens.
4. семейство гоминид
• исторически были выделены триважнейших системы, считающиеся
истинно гоминидными. Эти системы
были названы гоминидной триадой:
• -прямохождение (бипедия);
- кисть, приспособленная к
изготовлению орудий;
- высокоразвитый мозг.
5. Эволюция мозга
• Мозг грацильных австралопитеков, 4 2,5 м. назад, отличался от мозгашимпанзе большей длиной, - вероятно,
обладали несколько большей
чувствительностью и подвижностью
рук, чем современные
человекообразные обезьяны.
6.
• В сравнении же с массивнымиавстралопитеками, грацильные,
вероятно, имели более развитый
зрительный анализатор, что, возможно,
в некоторой мере отражает специфику
пищевого поведения (всеядность?)
7.
• массивные австралопитеки, всравнении с грацильными, обладали
лучшей координацией движений, но
меньшим контролем за эмоциями и
возбудимостью, меньшей
эмоциональностью и пониженными
способностями к прогнозированию
своих действий. Кроме того, массивные
австралопитеки, вероятно, обладали
большими возможностями
8. Архантроп Homo erectus
• Объём мозга достигал значенийсовременного человека, В увеличении
мозга -резко возросшая ширина мозга.
Лобная доля мозга была низкая и узкая,
но с очень сильно развитой
нижнелобной областью, что
проявлялось, в частности, в
формировании рельефа в области зоны
Брока – моторной зоны речи
9.
В теменной доле бурно расширялась надкраевая часть. Височная доля
расширялась в задней части и удлинялась по нижнему краю.
Увеличивались также затылочная доля и мозжечок.
• Обращает на себя внимание бурное
развитие зрительных центров
затылочной доли, происходившее при
возникновении H.erectus.
• Оно может быть связано с изменением
экологической ниши – началом выхода
в открытые саванны.
10.
• У архантропов в сравнении с "ранними Homo" наверняка былусилен сознательный контроль эмоций и, по всей видимости,
выросли возможности памяти. Судя по развитию теменной
доли, значительно развились тактильные возможности и
намного улучшилась координация движений. Вероятно,
прогрессивно развивались способности к речи. Все эти выводы,
полученные на основании изучения формы эндокранов, вполне
согласуются с данными археологии, ведь H. Erectus
пользовались ашельской индустрией, гораздо более
продвинутой сравнительно с галечной культурой
"ранних Homo", использовали огонь, охотились на крупных
животных. Впрочем, они ещё не покидали областей с
тропическим климатом и, скажем, искусство строительства
жилищ у них не было значительно развито (Jablonski et al.,
2000).
11. Homo heidelbergensis
• Форма мозга H. Heidelbergensis свидетельствуе о резком прогрессе вобласти контроля за движениями, в том
числе способностей к прогнозированию и
планированию своих будущих действий.
• развитие области, обеспечивающей
согласование речи и движений рук, а
также рельефное выступание зоны
Брока, свидетельствующее о начале
использования речи.
12.
• Все указанные прогрессивныеизменения развития мозга проявились в
культуре и особенностях образа жизни
пре-палеоантропов. Они заселили
умеренные области Старого Света, что
потребовало нового уровня
приспособленности, более активного
использования огня, строения жилищ,
изготовления новых видов орудий
13. Homo neanderthalensis или палеоантропы
Homo neanderthalensis илипалеоантропы
• У неандертальцев ширина мозга крайне
велика, максимальна для всех групп
гоминид. Очень характерны
относительно малые размеры лобной и
теменной долей при очень больших –
затылочной. В орбитной области (на
месте зоны Брока) были развиты
рельефные бугры.
14.
• Мозг H. neanderthalensis отличался отмозга современного человека, вероятно,
большим развитием подкорковых центров
подсознательного контроля за эмоциями
и памятью, но, вместе с тем, меньшим
сознательным контролем за этими же
функциями.
15.
По-видимому, можно констатировать существенное отставание
палеоантропов от современного человека в плане возможностей
контроля и координации движений. При этом существование речи у
неандертальцев, по данным палеоневрологии, весьма вероятно и
даже почти несомненно. Все эти выводы находят подтверждение в
данных археологии. Неандертальцы были первыми людьми,
заселившими крайне неблагоприятные для жизни приледниковые
территории Северной Евразии. Они жили в основном за счёт охоты,
строили достаточно сложные жилища, погребали умерших.
Впрочем, европейские неандертальцы не знали украшений,
наскальной живописи и обработки кости, они крайне медленно
перенимали новшества и неохотно обменивались культурными
достижениями. Вероятно, разные группы европейских и азиатских
неандертальцев мало общались друг с другом, даже живя в соседних
местностях.
16.
• пропорции древнейших прямоходящихсуществ – Ardipithecus
ramidus из Арамиса – ещё настолько
напоминают обезьяньи, что
закономерности их сложения имеют
мало отношения к таковым
современного человека [Lovejoy et al.,
2009a,b].
• Как можно видеть, пропорции тела и
более поздних австралопитеков не
17.
18. Sahelanthropus tchadensis Сахелантроп
• 7.0 млн.л.н. - 6.0млн.л.н. 2002 Чад
19.
• Останки найдены в Республике Чад в ТоросМеналла. Шесть находок гоминид включили полныйчереп, получивший имя Тумай (Toumai), два
фрагмента нижних челюстей и три изолированных
зуба.
• Находки характеризуются большим количеством
специфических особенностей и потому были
выделены в новый род и вид Sahelanthropus
tchadensis
• Череп Сахелянтропа – самый древний целый череп
прямоходящего существа.
• О прямохождении говорит расположение большого
затылочного отверстия, которое смещено
вперед; базион находится на линии между
отверстиями сонных артерий. Такое положение
промежуточно между вариантами, характерными для
четвероногих человекообразных обезьян и для
более поздних прямоходящих гоминид.
20.
21. Orrorin tugenensis Оррорин
6.0 млн.л.н.2001 Кения
22.
23.
• В 2001 г. Б. Сеню и М. Пикфорд обнаружила в Туген Хиллс 12костей гоминид, в том числе правую и левую бедренные кости;
находки представляли по меньшей мере 5 индивидов обоего
пола.
• Большая древность находки вкупе с архаичной морфологией
дали основание авторам открытия для описания нового рода и
вида гоминид Orrorin tugenensis. В популярной литературе этот
австралопитек известен как "Человек Тысячелетия" ("Millennium
Man").
• Фрагменты левой и правой бедренных костей имеют множество
признаков, сближающих O. tugenensis с гоминидами и
отличающих их от человекообразных обезьян. В целом,
признаки бедренных костей достаточно явно указывают на
двуногий способ передвижения их обладателей. В то же время,
строение верхних конечностей указывает на элементы
древесной локомоции (впрочем, как и у более поздних
австралопитеков). Зубы в некоторых признаках сближаются с
зубами шимпанзе.
24. Ardipithecus kadabba Ардипитек кадабба
• 5.5 млн.л.н.• 2001 Эфиопия
25.
Ардипитек кадабба (Ardipithecus ramidus kadabba)
представляет ещё один род поздномиоценовых гоминид,
населявших тогда нашу Землю. Его останки были найдены
совместной группой эфиопских и американских антропологов во
главе с американским исследователем Йоханессом Хайле
Селассие, взявшего такую экзотическую фамилию в честь
известного эфиопского императора.
• Ранее, в 1995 г. были найдены останки подобных гоминид. Их
отнесли не только к новому виду, но даже роду гоминид и
присвоили имя Ardipithecus ramidus .
• Морфология останков довольно близко приближалась к
морфологии найденных ранее представителей этого же рода,
но всё же имелись некоторые различия.После тщательного
анализа и изучения вновь найденным останкам ардипетека
было присвоено ещё и имя кадабба ( кадабба в переводе с
афарского означает основатель рода), а новый гоминид
выделен в новый вид Ardipethecus kadabba.
26.
• Ardipithecus kadabba одно из древнейшихпрямоходящих существ, от которого сохранились
кости ног и рук.
• Находки включают правую половину нижней
челюсти, изолированные зубы, ключицу, фрагменты
двух плечевых и локтевой костей, две фаланги
кисти, проксимальную фалангу стопы – кости по
меньшей мере от 5 индивидов. Строение зубов
(особенно - клыков) промежуточно между
человекообразными обезьянами и человеком, но
ближе всё же к человеку; строение длинных костей
указывает на вертикальное положение тела при
ходьбе.
27. Ardipithecus ramidus Ардипитек рамидус
• 4 млн.л.н.• 1995 Эфиопия
28.
• Ардипитеки (лат. Ardipithecus) — древнийрод гоминид, который жил в раннем плиоцене.
Поскольку род ардипитеки имеет много общего с
африканскими обезьянами, шимпанзе и гориллами,
некоторые авторы считали его предком скорее
шимпанзе, чем человека. Тем не менее,
большинство считает его древнейшим предком
человека из-за сходства зубов с
зубами австралопитека. Кроме того, в стопе
ардипитека имеется добавочная малая берцовая
кость (os peroneum), которая присутствует в стопе
человека, но отсутствует в стопе современных
человекообразных обезьян. Иногда ардипитеков
относят к ранним австралопитекам.
29.
• Найденные в Арамисе в 1995 гг.остатки гоминид включили фрагменты черепов,
нижней челюсти детеныша, большое количество
изолированных зубов, а также фрагменты костей
верхних конечностей по меньшей мере от 17
индивидов. В 1995 г. были обнаружены более 90
окаменевших фрагментов, составивших около 45%
скелета взрослого индивида, в том числе части
черепа, позвоночника, рук, таза, ног и стопы (скелет,
получивший прозвище "Арди"). Это древнейший
скелет прямоходящего гоминида с большим
количеством обезьяньих признаков.
• Сначала находкам дали
наименование Australopithecus ramidus, а позже
выделили в новый род –Ardipithecus.
30. Australopithecus anamensis Австралопитек анамский
• 4.2 млн.л.н. 3.9 млн.л.н.• 1994 Кения
31.
• Австралопитек анамский (лат. Australopithecus anamensis) —ископаемый вид рода австралопитеков, обитавший в Восточной
Африке около 4 миллионов лет назад. Это наиболее древний из
видов австралопитеков, известных науке.
• Первой находкой данного вида гоминид
был дистальный фрагмент левой плечевой кости,
обнаруженный в Канапои в 1965 г. в процессе работ
Гарвардской экспедиции под руководством Б. Паттерсона.
Спустя почти 30 лет, в 1994 г. группой под руководством М.Г.
Лики здесь был найден ряд окаменелостей, позволивших
описать новый вид ранних австралопитеков.
• Всего найдено около 100 окаменелостей от более чем 20
особей. Все останки сочетают понгидные и
гоминидные признаки примерно в равной пропорции.
• Вероятно, данный вид является потомком ардипитека и
предком афарских австралопитеков.
• При развитом прямохождении Australopithecus anamensis
иногда, вероятно, ходил на четвереньках с опорой на согнутые
пальцы рук (о чем говорит строение локтевой кости).
32. Australopithecus afarensis Австралопитек афарский
• 4.0 млн.л.н. - 2.5млн.л.н.
• 1978 Кения,
Эфиопия,
Танзания
33.
• Австралопитек афарский (лат. Australopithecusafarensis) — вымерший вид австралопитека,
семейство гоминид, живший около 4 миллионов
лет назад. Почти нет сомнений, что до того, как
афарские австралопитеки вымерли 2,5 миллиона
лет назад, от них прямо или косвенно произошли
другие австралопитецины и род Homo.
• Афарские австралопитеки хорошо известны по
раскопкам в Хадаре, проводившимся в 19731977гг. Международной Афарской
Исследовательской Экспедицией под
руководством И. Коппенса, М. Тайеба и Д.
Джохансона. Были обнаружены более 240
остатков гоминид, принадлежавших как минимум
35 особям.
34.
Морфологические особенности хадарских австралопитеков, благодаря
представительности материала, изучены очень хорошо и могут
служить своеобразным эталоном для сравнения при описании других
видов австралопитековых. Среди многочисленных находок
наибольшей известностью пользуются останки самки AL 288-1. Эта
находка получила популярное имя «Люси». 40% скелета Люси были
найдены 30 ноября 1974 г. Д. Джохансоном и Т. Грэем в отложениях
пачки Када Хадар с наиболее вероятной датировкой 3,18-3,2 млн.л.
Череп афарских австралопитеков отличается низким, покатым лбом;
надбровье мощное. Височные линии лобной кости образуют
низкий сагиттальный гребень. Затылок преломленный, хорошо
выражен затылочный гребень. Основание черепа изогнуто очень
слабо, подобно понгидам.
В затылочной норме череп A. afarensis более низкий и расширенный в
нижней части, чем у A. africanus, что является более примитивным
вариантом; в этом отношении A. afarensis больше похож на массивных
австралопитеков.
Эндокран внешне очень похож на таковой человекообразных обезьян,
однако на нем нет отпечатка полулунной («обезьяньей») борозды,
обычно хорошо заметного на эндокранах понгид, у которых данная
борозда ограничивает затылочную долю, тогда как у современного
человека ее нет. Теменная и височная ассоциативные зоны
увеличены, что, несомненно, является прогрессивным признаком.
35.
Из особенностей строения зубов замечательны значительные размеры резцов и
большая разница размеров резцов I1 и I2, из которых первый больше. Верхние
клыки сильно выступают над соседними зубами, их размеры и морфология
промежуточны между понгидным и гоминидным вариантами, приближаясь все же
к последнему; клыки достаточно заостренные на вершине, их корни наклонены
назад. Характерно, что верхние клыки самцов A. afarensis больше, чем у самок.
Моляры A. afarensis достаточно большие, иногда их размеры даже описываются
как умеренномегадонтные; однако относительно передних зубов моляры были
все же намного меньше, чем у массивных австралопитеков.
Комплекс строения дистальной Часть (например, скелета), наиболее удаленная
от центра тела или органа. Противоположность "проксимальной части". части
лучевой кости свидетельствует о том, что афарские австралопитеки, либо их
непосредственные предки, могли ходить на фалангах пальцев рук.
Замечательно строение таза Australopithecus afarensis, который резко отличается
от высокого суженного таза человекообразных обезьян и, напротив, мало
отличим от таза современного человека. Таз крайне широкий и короткий.
Пропорции таза "Люси" в некотором отношении даже «ультрагоминидны».
Видимо, специализация к двуногой ходьбе у A. afarensis зашла уже очень далеко.
Признаки, наблюдающиеся в скелете A. afarensis, свидетельствующие о иных
способах передвижения, могут быть либо наследием, доставшимся от предков,
либо говорят о широком спектре типов передвижения, использовавшихся этими
австралопитеками, хотя прямохождение, видимо, было среди них
преобладающим типом.
36.
Морфология стопы A. afarensis также свидетельствует о разнообразии
видов локомоции, использовавшихся этим видом; в этом отношении
грацильные австралопитеки весьма своеобразны и отличаются как от
понгид, так и от более поздних гоминид.
В целом руки A. afarensis были несколько удлинены относительно ног,
соответствующий индекс имеет значение, промежуточное между
значениями, известными для человекообразных обезьян и
современного человека; также, ноги относительно роста были
несколько короче, чем у современного человека. Такие пропорции,
видимо, являются следствием незавершенного перехода к двуногому
прямохождению, что, как уже было сказано, отражено во многих
деталях морфологии костей конечностей. Видимо, ходьба афарских
австралопитеков была медленнее, а величина дневного перехода
была намного меньше, чем у современного человека.
Размеры тела хадарских A. afarensis чрезвычайно варьируют. Разница
отражает, по-видимому, половой диморфизм, который оказывается
равным таковому орангутанов и горилл и большим, чем у шимпанзе и
человека. Рост маленьких особей едва превышал метр, тогда как у
больших достигал более полутора метров; вес варьировал
соответственно от примерно 30 до 45 кг, возможно даже – до 55 кг.
Australopithecus afarensis - наиболее вероятный предок человеческой
линии эволюции.
37. Kenyanthropus platyops
• 3.5 млн.л.н. 3.2 млн.л.н.• 2001 Кения
38.
• Кениантропы (лат. Kenyanthropus) — древний род гоминид,живший в плиоцене около 3,5-3,2 млн лет назад.
• Найден череп и другие окаменелости; вероятно, Kenyanthropus
platyops был специализированным видом, не входившим в нашу
прямую родословную.
• Основанием для описания вида явилась мозаика
морфологических признаков, в отдельности встречающихся и у
других австралопитеков, но в определенном сочетании
характеризующая именно K. platyops.
• По мнению авторов описания вида, форма лицевого скелета
позволяет сближать Kenyanthropus platyops с более
поздним Homo rudolfensis.
• Характернейшей чертой авторы первоописания считают
значительную ширину и уплощенность лица кениантропа,
однако она, во-первых, может быть значительно преувеличена
деформацией, а, во-вторых, не намного более сильно
выражена, чем у многих A. africanus и, тем более, массивных
австралопитеков; отличия по этому признаку от синхронного и
территориально идентичного видаA.afarensis также вовсе не
очевидны.
39. Australopithecus africanu Австралопитек африканский
• 3.5 млн.л.н. - 2.4млн.л.н.
• 1925 ЮАР
40.
• Австралопитек африканский (лат. Australopithecusafricanus) — вымерший вид австралопитека,
семейство гоминид, живший около 3,5—2,4 миллиона
лет назад.В отличие от афарского
австралопитека имел более обезьяноподобный
скелет, но более объёмный череп.
• К этому виду принадлежала первая находка
австралопитека – череп детёныша, известного как
"Бэби из Таунга", описанный Р.А. Дартом в 1925 году.
Именно этот череп, а именно относительно
переднее расположение затылочного отверстия,
позволили Дарту предположить, что австралопитеки
были прямоходящими.
• A. africanus имел более прогрессивный по строению
череп, чем A.afarensis, но в целом более архаичный
скелет.
41.
• Пропорции, видимо, были промежуточными междупропорциями шимпанзе и современного человека.
• Общие размеры тела A. africanus варьировали
примерно от метра до полутора, вес – от 20 до 40 кг.
• Кости ног A. africanus сочетают архаичные и
прогрессивные особенности. Большой палец стопы,
вероятно, обладал большой подвижностью.
• Вместе с тем тазовые кости (известные по ряду
находок) по подавляющему большинству признаков
ближе к человеческому варианту, чем к тазу
человекообразных обезьян. Форма таза является
наиболее сильным аргументом, подтверждающим
двуногую локомоцию австралопитеков.
42.
• Череп A. africanus в среднем больше, чем у A. afarensis; наклонлба сравнительно небольшой, затылок тоже достаточно
вертикальный и плавно закруглен; череп при сравнении с A.
afarensis выглядит более округлым, коротким и высоким.
• Строение зубов: наклон резцов и клыков и их небольшие
размеры - отличают A. africanus от человекообразных обезьян,
делая его больше похожим на человека.
• Строение мозга Australopithecus africanus, судя по эндокранам,
близко к шимпанзе и достаточно далеко от современного
человека, однако, все же, мозг этих австралопитеков более
похож на мозг человека, чем мозг всех видов Paranthropus.
• Рост мозгового отдела черепа в сравнении с лицевым по
темпам был промежуточен между человекообразными
обезьянами и человеком. Хотя во взрослом состоянии эти
австралопитеки имели объем мозга примерно такой же, как и
человекобразные обезьяны, по-видимому, объем этот
достигался в онтогенезе раньше.
43. Australopithecus bahrelghazali Австралопитек бахр-эль-газальский
Australopithecus bahrelghazaliАвстралопитек бахр-эльгазальский
• 3.5 млн.л.н. - 3.0
млн.л.н
• 1996 Чад
44.
• Австралопитек бахр-эль-газальский (лат. Australopithecusbahrelghazali) — предположительно, самостоятельный
вид австралопитека, семейство гоминид, живший около 3,5—
3,0 миллионов лет назад.
• Это самая западная находка австралопитека. Обнаружена
в 1996 году французским палеонтологом Мишелем Бруне на
территории древнего речного русла Бахр-эль-Газаль в Чаде.
Она представляет собой верхнюю челюсть с семью зубами.
Поскольку ранее останки австралопитеков обнаруживались
только в Восточной и Южной Африке, данная находка стала
свидетельством довольного широкого распространения рода.
• Передняя часть нижней челюсти с 7 зубами гоминида была
найдена М. Брюнет с сотрудниками и послужила голотипом
для описания нового вида Australopithecus bahrelghazali.
45.
• Всего обнаружены один фрагмент верхней и один –нижней челюстей, а также изолированный
зуб. В целом данные находки мало отличаются от
останков афарских австралопитеков.
• Своеобразие зубной системы состоит в наличии трех
корней у P1 и P2, тогда как у A. afarensis имеется 1-2 корня
на P1 и 2 корня на P2; трехкорневые нижние премоляры
нехарактерны ни для одного другого вида
австралопитековых.
• Важность находки заключается в ее географическом
положении: большинство австралопитеков найдено в
Восточной и Южной Африке, а Австралопитек бахр-эльгазальский - в Северной, что свидетельствует о очень
широком распространении группы.
46. Australopithecus garhi Австралопитек гари
• 2.5 млн.л.н.• 1999 Эфиопия
47.
• Австралопитек гари (лат. Australopithecusgarhi) — вымерший вид австралопитека,
семейство гоминид, живший около 2,5
миллионов лет назад. Представители этого
вида являются возможным «переходным
звеном» от рода австралопитеков к
роду людей (Homo). Относятся к группе
грацильных австралопитеков, то есть
обладают хрупким, не массивным
телосложением
48.
• «Garhi» в переводе с афарского означает«удивительный» или «сюрприз».
• Это поздний грацильный австралопитек с датировкой
около 2,5 млн. лет назад.
• A. garhi - eдинственный австралопитек, с останками
которого были найдены каменные орудия труда и
расколотые кости антилоп; судя по датировке и
специализированным чертам, нашим прямым
предком не являлся.
• Представлен череп, зубы, кости конечностей. Рост
1,2-1,5 м. Объем мозга около 440 см3. Размеры
передних зубов A. garhi превосходят таковые у
любых других видов австралопитеков.
49. Australopithecus sediba Австралопитек седиба
• 2.0 млн.л.н. - 1.8млн.л.н.
• 2010 ЮАР
50.
Австралопитек седиба (Australopithecus sediba) —
вид австралопитеков, известный по останкам, возраст которых
оценивается в 2.0 - 1.8 млн лет (плейстоцен). Два неполных скелета
этого вида были найдены при раскопках в пещере Малапа (Malapa) на
территории объекта Всемирного наследия ЮНЕСКО, известного как
«Колыбель человечества», в Южной Африке. Один скелет —
подростка мужского пола (голотип), другой — взрослой особи женского
пола. На сегодняшний день найдено более 130 фрагментов этих
скелетов. Эти находки первоначально были описаны в двух статьях в
журнале «Science» южноафриканским палеоантропологом Ли
Бергером и его коллегами как новый вид ранних предков человека. Там
же новый вид назван Australopithecus sediba («sediba» означает
«источник» или «колодец» в языке сото).
В 2008 году в карстовой пещере, на глубине нескольких десятков
метров были обнаружены два разрушенных скелета (взрослой самки и
подростка) и отдельные кости третьего (младенца). По этим находкам
в 2010 году описан новый вид. Слово "sediba" на местном африканском
языке сесото означает«родник».
51.
• Интересно, что находкаостанков Australopithecus sediba сделана с
использованием популярного сервиса Google Earth,
позволившего выявить местонахождения
многочисленных карстовых пещер в Южной Африке.
• По большинству черт Australopithecus sediba –
промежуточен между австралопитеками и Homo (по
многим признакам этот вид оказывается даже более
гоминидным, чем хабилисы Восточной Африки). Изза ряда "австралопитековых" черт (таких, как
маленький объем мозга, особенности строения рук и
ног).
52.
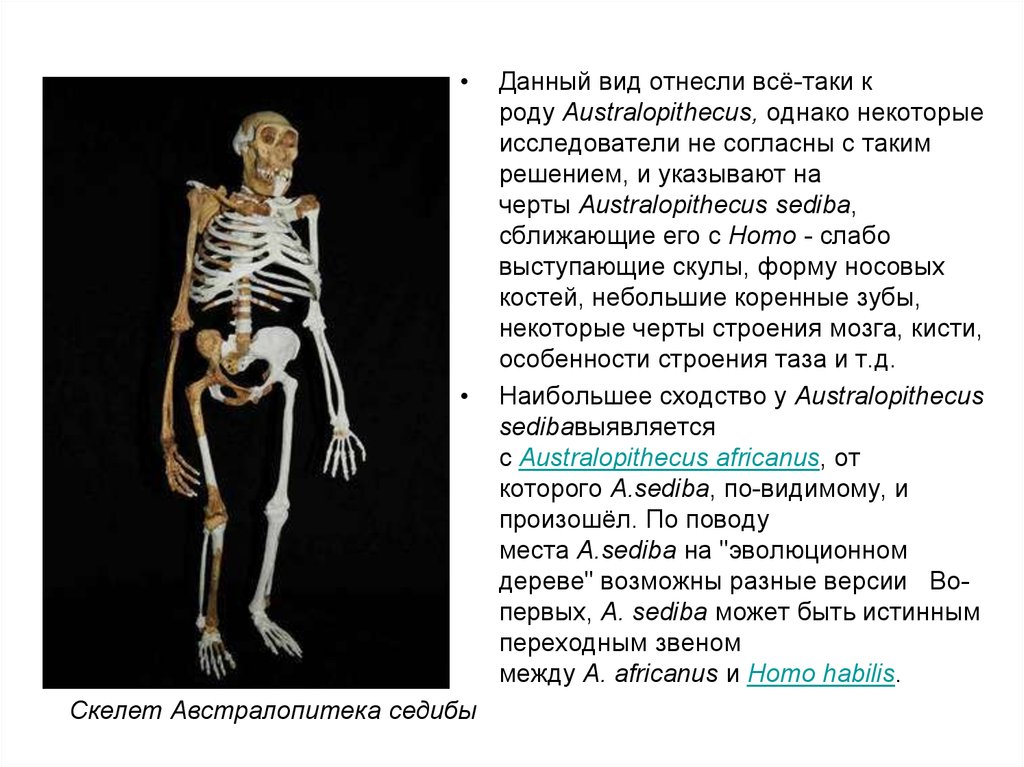
Скелет Австралопитека седибы
Данный вид отнесли всё-таки к
роду Australopithecus, однако некоторые
исследователи не согласны с таким
решением, и указывают на
черты Australopithecus sediba,
сближающие его с Homo - слабо
выступающие скулы, форму носовых
костей, небольшие коренные зубы,
некоторые черты строения мозга, кисти,
особенности строения таза и т.д.
Наибольшее сходство у Australopithecus
sedibaвыявляется
с Australopithecus africanus, от
которого A.sediba, по-видимому, и
произошёл. По поводу
места A.sediba на "эволюционном
дереве" возможны разные версии Вопервых, A. sediba может быть истинным
переходным звеном
между A. africanus и Homo habilis.
53.
• Однако, древнейшие "ранние Homo" и в Южной, и в ВосточнойАфрике имеют гораздо большие – как минимум на 300 тысяч
лет – датировки; впрочем, эти находки крайне фрагментарны и
в будущем, возможно, окажется возможным объединить их
с A. sediba в один таксон. Во-вторых, A. sediba может быть
представителем собственной южноафриканской линии
гоминизации, параллельной восточноафриканской, но чуть
более поздней; в связи с этим можно вспомнить A. garhi – ещё
одну альтернативную ветвь, дошедшую до уровня изготовления
орудий. В-третьих, останки, описанные как A. sediba, могли
принадлежать метисам между поздними грацильными
австралопитеками и уже сложившимися "ранними Homo", хотя
эта версия маловероятна.
54. Paranthropus aethiopicus Парантроп эфиопский
• 2.5 млн.л.н.• 1968 Кения,
Эфиопия
55.
• Парантроп эфиопский (лат. Paranthropusaethiopicus) — ископаемый вид
рода парантропов («массивных австралопитеков»),
обитавший в Восточной Африке около 2,5 миллионов
лет назад. Это наиболее древний из парантропов,
известных науке.
• В 1985 А. Волкером несколько ниже туфа Локалеи
был обнаружен почти целый череп KNM-WT 17000,
известный также как “Черный череп”, позже были
сделаны и другие, менее комплектные находки,
например, фрагмент нижней челюсти KNM-WT
16005. Череп KNM-WT 17000
послужил голотипом для описания нового вида.
56.
(Если быть точным, история данного вида довольно запутанная.
Первоначально род и видParaustralopithecus aethiopicus Arambourg et
Coppens, 1968 были описаны на основании морфологии
нижней челюсти Omo 18-1967-18 из Омо (2,5 млн.л.н.). Позже, когда был
найден череп KNM-WT 17000, он был описан как Australopithecus
aethiopicus Walker, Leakey, Harris et Brown, 1986, но прямо не мог быть
ассоциирован с Paraustralopithecus aethiopicus Arambourg et Coppens,
1968 (так как
там голотип - нижняя челюсть). Позже этот же череп был описан ещё и
как Australopithecus walkeriFerguson, 1989 (тут была хитрая
таксономическая игра с родовыми и видовыми названиями). В итоге
договорились называть данный вид Paranthropus aethiopicus, к которому
относят и челюсть, и череп, хотя челюсть - всегда с некоторыми
оговорками).
Это древнейший из массивных австралопитеков и единственный среди
этой группы, имевший резко выступавшие вперёд челюсти.
Череп парантропа эфиопского сочетает в себе примитивные признаки,
сближающие его с ранними и грацильными австралопитеками, и
специализированные черты массивных австралопитеков.
Paranthropus aethiopicus - вероятный предок более поздних восточноафриканских массивных австралопитеков Paranthropus boisei.
57. Paranthropus boisei Парантроп Бойса
• 2.5 млн.л.н. - 1.0млн.л.н.
• 1959 Кения,
Танзания
58.
• Парантроп Бойса (Paranthropus boisei Mary Leakey,1959), прежде также зинджантроп (Zinjanthropus
boisei), — самый массивный вид парантропов,
обнаруженный в 1959 году супругами Лики в
восточноафриканских ущельях Олдувай, КообиФора, Локалеи и ряде других. Они датированы от 2,5
до 1 млн лет назад. Вплоть до 1959 года считался
«несомненным австралопитеком»
• У этих австралопитеков максимального развития
достиг челюстной аппарат. Первый открытый череп
этого вида из-за размеров зубов даже получил
прозвище "Щелкунчик".
59.
Череп P. boisei хорошо известен по множеству находок хорошей
сохранности, с датировками в пределах 1,7-1,9 млн.л. Эти черепа
очень похожи, их небольшие отличия могут быть отнесены за счет
возраста и пола. Череп массивный, с хорошо развитым мускульным
рельефом. Конфигурация мозговой коробки своеобразная, при взгляде
сверху напоминает кувшин: спереди череп очень расширен благодаря
мощному развитию лицевой области, далее следует сильное
позадиглазничное сужение, лоб сильно скошен и сужен, потом
черепная коробка резко расширяется. Огромные скуловые дуги служат
мощными укрепляющими элементами, благодаря своей винтообразной
изогнутости. В затылочной норме череп невысокий и широкий в нижней
части.
Надглазничный рельеф P. boisei развит достаточно сильно.
Размер сагиттального гребня, как и упонгид, различается у особей
разного пола: большой у самцов и низкий у самок.
Лицевой скелет P. boisei, как и мозговой, очень своеобразный.
Бросается в глаза чрезвычайная расширенность лица, обусловленная
огромными размерами верхней челюсти и скуловой кости. Средняя
лицевая и скуловая ширина намного превышают как остальные
широтные, так и высотные размеры.
60.
• Строение мозга P.boisei довольно
примитивное.Эндокраны х
арактеризуются сильно
суженной лобной долей
даже по сравнению с
более древними
грацильными
австралопитеками.
• Верхняя челюсть P.
boisei очень большая и
массивная, очень
высокая. Нижняя
челюсть, как и верхняя,
отличается огромными
размерами и
массивностью.
61.
• Относительные размеры передних и заклыковых зубов P.boisei резко отличаются от пропорций, характерных для
грацильных австралопитеков. Это классический вариант
заклыковой мегадонтии, когда клыки и резцы редуцированы, а
премоляры и моляры, напротив, гипертрофированы в размерах.
При этом размер моляров увеличивается от первого к третьему.
Эмаль зубов очень толстая, возможно, пища массивных
австралопитеков состояла из грубоволокнистой, жесткой
растительности.
• Всё строение черепа P. boisei говорит о больших нагрузках,
приходившихся на него; это выражается в большой толщине
черепных стенок, развитии рельефа, общей архитектуре с
массивными укрепляющими элементами, гипертрофированных
челюстях. Жевательный аппарат P. boisei был чрезвычайно
развит, все развитие костного рельефа и морфология
нижнечелюстного сустава говорят о большой мощности
жевательных мышц. Очевидно, массивные австралопитеки
были адаптированы к питанию растительной пищей, причем,
судя по всему, количество этой пищи было весьма
значительным.
62. Paranthropus robustus Парантроп массивный
• 2.5 млн.л.н. 900 тыс.л.н.• 1939 ЮАР
63.
В 1938 г. Роберт Брум проводил раскопки в пещерах Стеркфонтейна. 8
июня 1938 г. школьник Герт Тербланш нашел в местечке Кромдрай,
расположенном неподалеку от Стеркфонтейна, вмурованный в породу
череп. Р. Брум выменял череп у мальчика за 5 плиток шоколада.
Череп, а также ряд обнаруженных в этом местонахождении
окаменелостей были описаны под новым родовым и видовым
названием Paranthropus robustus. Впоследствии в ряде
местонахождений Южной Африки были сделаны сотни находок,
относящихся к этому виду гоминид.
Таким образом Paranthropus robustus является вторым открытым
видом австралопитеков.
Череп P. robustus массивный, с толстыми стенками и выраженным
рельефом. Лоб низкий, покатый; надбровный валик мощный,
надбровные дуги расположены почти горизонтально, отличаясь от
покатых у Paranthropus boisei. Лицевой скелет плоский, даже вогнутый.
Скулы массивные, высокие.
Верхняя челюсть P. robustus массивная, укороченная. Нижняя
челюсть также чрезвычайно массивна. Заклыковые зубы
большие, мегадонтные (хотя не в такой степени, как у Paranthropus
boisei).
Затылочное отверстие сдвинуто вперед, что является одним из
свидетельств прямохождения массивных австралопитеков.
64.
• Эндокран P. robustus имеет примитивную форму, малоотличающуюся от формы человекообразных обезьян.
• Верхние конечности P. robustus, как и других австралопитеков,
представляют собой
мозаику понгидныхи гоминидных признаков.
• Морфология тазовых костей P. robustus, несомненно, ближе к
человеческой, нежели к обезьяньей. Таз очень широкий и
развернутый. Общий комплекс черт говорит о двуногой
походке P. robustus, хотя эта походка, возможно, слегка
отличалась от походки представителей рода Homo.
• Стопа этих массивных австралопитеков, видимо, была
достаточно человекоподобной.
• В местонахождении Сварткранс, где также обнаружены останки
P. robustus, было найдено множество костяных фрагментов со
следами стертости на конце. Опытным путем было показано,
что аналогичные следы образуются при использовании костей
для вылавливания термитов; таким образом, возможно, P.
robustus могли использовать костяные орудия.
65. Homo habilis Человек умелый
• 2.3 млн.л.н. - 1.5млн.л.н.
• 1964 Кения, ЮАР
66.
• Челове́к уме́лый (лат. Homo habilis) —высокоразвитый австралопитек или первый представитель
рода Homo.
• Обнаружен археологами Лики (Мэри и Джонатаном)
в 1960 году и описан в 1964 году по сенсационной находке из
ущелья Олдувай вТанзании. В Олдувайском ущелье они вместе
с костями вымершего саблезубого тигра смилодона нашли
стопу, пяточную кость,ключицу и обломки черепа
нового гоминида. Возможно, он пал жертвой хищника. Череп,
как установили потом, принадлежал ребёнку 11—12 лет. Судя
по строению стопы новый гоминид был прямоходящим.
• Нового гоминида сначала
назвали презинджантроп (Prezinjanthropus), а затем в 1964 году
по предложению Луиса Лики переименовали в Homo
habilis (Человек умелый). Позже аналогичные находки были
сделаны в Кооби-Фора, Сварткрансе и других
местах Восточной иЮжной Африки.
67.
Человек умелый был первым из наших предков, который стал
регулярно изготавливать каменные орудия труда и перешёл на
всеядность; с человека умелого начинается бурный рост размеров
головного мозга. Homo habilis был создателем ранней галечной (так
называемой «олдувайской» или «олдованской») культуры.
Homo habilis иногда определяется как
поздний грацильный австралопитек, однако ряд прогрессивных
признаков в строении приближает его к более поздним гоминидам.
Череп Homo habilis хорошо известен по множеству находок хорошей
сохранности. Череп во многом схож с австралопитеками,
однако обладал более округлым и высоким сводом, а мозговая
полость - имела в среднем более крупные размеры; костный рельеф
черепа был довольно слабо развит. Боковые стенки черепа H.
habilis почти вертикальные, относительно высокие. Лобная кость
убегающая назад, однако выпуклая в большей степени, чем у
австралопитеков; надбровье мощное, выступающее. Затылочная
кость высокая, округлая, рельеф ее очень слабый, у H. habilis нет
характерных для австралопитеков затылочных гребней. Затылочное
отверстие вытянутое, смещено вперед на основании черепа, что
свидетельствует о прямохождении.
68.
• Лицевой скелет H. habilis более прогрессивный, чем уграцильных австралопитеков. Горизонтальная уплощенность
лица в большинстве случаев слабая, носовая область
выступает вперед, тогда как скулы сдвинуты назад.
• Челюсти H. habilis имеют меньшие размеры и выступают
слабее, нежели у австралопитеков.
• Зубы H. habilis в среднем меньше, чем у грацильных и
массивных австралопитеков, но превосходят размерами зубы
более поздних архантропов. Таким образом, отчетливо
проявляется эволюционная тенденция к общему постепенному
уменьшению размеров зубов.
• Пропорциями конечностей H.habilis не отличались от
австралопитековых.
• Стопа H.habilis была уплощённой, без выраженного свода, но
пальцы были прямыми, короткими, абольшой палец полностью
приведён к остальным.
69. Homo rudolfensis Человек рудольфийский
• 2.3 млн.л.н. - 1.5млн.л.н.
• 1978 Кения
70.
• Человек рудольфский (лат. Homo rudolfensis) — вымерший«старейший» вид людей, представляющий (согласно традиционным
представлениям) переходную ступень от человека
умелого к эректусам. Жили 2,3—1,5 млн лет назад. Рост составлял
1,5—1,8 м, вес 45—80 кг. Согласно антропологическим
исследованиям 2012 года, человек рудольфский был не вариацией
человека умелого, а представителем параллельной линии в
эволюции древних людей; от кого из них произошёл современный
человек, неизвестно.
• Второй из устоявшихся видов "ранних Homo", сосуществовавший
с Homo habilis (т.к. останки обнаружены в тех же местонахождениях
и в тех же слоях).
• Этот вид «ранних Homo», обладавший крупным мозгом, впервые
был описан В.П. Алексеевым какPithecanthropus rudolfensis в 1978 (в
зарубежной литературе всегда фигурирует другой год описания –
1986, являющийся годом выхода перевода книги В.П. Алексеева на
английский язык). В последующем взгляд на систематическое
положение этого типа гоминид неоднократно пересматривался,
наиболее часто используемым стало наименование Homo
rudolfensis.
71.
Наиболее характерным отличием от Homo habilis является большая
массивность и большие размеры зубов у Homo rudolfensis (хотя некоторые
исследователи полагают, что эти отличия могут быть объяснены
половым диморфизмом).
Верхняя хронологическая граница существования этого вида довольно
расплывчата. Постепенность морфологического перехода между H.
habilis и H. rudolfensis, с одной стороны, и первыми архантропами – с
другой, не позволяет четко определить момент перехода одного вида в
другой.
Голотипом вида H. rudolfensis явился широко известный череп KNM-ER
1470, имеющий датировку около 2 млн.л.
Лицо H. rudolfensis заметно уплощенное, а скуловая область развернута
вперед. Форма лицевого скелета позволяет сближать H. rudolfensis с
более древним Kenyanthropus platyops; некоторые исследователи даже
предлагают отнести этот вид «ранних Homo» Кооби Фора к
роду Kenyanthropus с образованием названия Kenyanthropus
rudolfensis (Collard et Wood, 2001).
Однако в целом H. rudolfensis довольно слабо отличаются от Homo habilis.
Для H. rudolfensis характерна умеренно выраженная
заклыковая мегадонтия и легкая редукция передних зубов.
Объем мозга H. rudolfensis – более 700 см3 (это гораздо больше, чем у
австралопитеков).
72. Homo ergaster Человек работающий
• 1.8 млн.л.н. - 1.4млн.л.н.
• 1975 Кения,
Танзания,
Грузия
73.
• Homo ergaster (Человек работающий) —ископаемый вид людей, появившийся
в Африке 1,8 млн лет назад в результате
эволюцииHomo habilis или Homo rudolfensis.
Рассматривается как промежуточное звено
между австралопитеками и Homo erectus,
иногда описывается
как африканский подвид питекантропов (лат.
Homo erectus ergaster), однако всё большее
число исследователей склонны отличать их
от неафриканских представителей
человеческого рода, живших в промежуток
1,8—1,4 млн.л.н. — время существования
Homo ergaster.
74.
• Обычно под термином Homo ergaster понимаются ранниесравнительно массивные восточноафриканские (точнее – из
области озера Туркана) гоминиды, не принадлежащие к
австралопитекам и несущие почти полный комплекс признаков
архантропов.
• Homo ergaster выделились среди ранних Homo и, судя по
существенному изменению морфологии (в том числе,
пропорций скелета - высокий рост, длинные ноги и
относительно небольшие широтные размеры), освоили новую
экологическую нишу - открытые вторичные саванны. Видимо,
окончательный переход к жизни на открытых пространствах
привел в быстрому расселению за пределы африканского
континента.
• Эргастеры были первыми гоминидами, вышедшими за пределы
Африки (Находки в Дманиси, Грузия, датируются 1,8 млн. лет
назад).
• Строение эргастеров хорошо изучено по множеству хорошо
сохранившихся находок (в том числе, знаменитый
скелет "Мальчик из Турканы" KNM-WT 15000).
75.
Черепа H.ergaster низкие и массивные, кости свода толстые. Лобная
кость убегающая назад, слабо изогнутая. Надглазничные дуги мощные,
резко выступают вперед. Заглазничное сужение выражено сильно.
Затылочная кость сильно преломлена, присутствует мощный округлый
затылочный валик. Мозговой отдел черепа относительно небольшой, а
лицевой отдел - крупный, высокий, с массивной выступающей вперед
верхней челюстью. Нижняя челюсть тяжелая, крупная. В целом, лицо
очень прогнатное и вместе с тем уплощенное в горизонтальной
плоскости.
Зубы по абсолютным размерам не уступают зубам "ранних Homo". При
этом намечается тенденция к относительному увеличению передних
зубов, и уменьшению задних.
Посткраниальный скелет в целом уже близок к скелету современного
человека.
Объем мозга примерно 800-900 см3, для некоторых экземпляров
(найденных в Дманиси) - менее 700 см3.
По мнению ряда исследователей, жизненный цикл Homo ergaster был
не таким, как у современных людей: они быстро развивались и рано
взрослели - полностью формируясь уже к 12 годам; при этом уHomo
ergaster отсутствовал выраженный ростовой спурт (скачок) - резкое
ускорение роста, происходящее у современных подростков во время
полового созревания.
76. Homo erectus Человек прямоходящий
• 1.5 млн.л.н. - 400тыс.л.н.
• 1894 Ява, Китай,
Греция, Испания, и
т.д.
77.
• Эректус (лат. Homo erectus — человекпрямоходящий; устар. архантропы) —
ископаемый вид людей, который
рассматривают как непосредственного
предшественника современных людей.
• Исследование ДНК X-хромосомы в 2008 году
привело к выводу, что азиатский вид Homo
erectus вполне мог скрещиваться с Homo
sapiens и быть предком современных людей
по смешанным линиям (не прямой мужской и
не прямой женской).
78.
Наиболее массивные представители древнейших людей. Являясь уже
несомненными людьми, Homo erectus всё же сильно отличались от
современного человека, так что многие антропологи склоняются к
выделению для них особого рода Питекантропов (Pithecanthropus).
Иногда среди этого типа гоминид выделяют множество
видов: Pithecanthropus pekinensis (синантроп) в Китае, Pithecanthropus
leakeyi в Восточной Африке, Pithecanthropus palaeojavanicus на
Яве,Pithecanthropus mauritanicus в Северной Африке и т.д. Видимо,
существовало как минимум две основных географических ветви этих
гоминид – западная или афро-европейская и восточная или азиатская.
Рядом исследователей Homo erectus объединяется в один вид с более
древними Homo ergaster, однако отличия между ними достаточно
велики.
От более поздних гоминид Homo erectus отличались в основном
заметно более низким черепом и деталями лицевой морфологии.
Эти гоминиды продолжали расселение по планете, достигнув на
востоке Индонезии, а на западе - Испании. При этом население, судя
по всему, концентрировалось в южных областях и не выходило за
пределы субтропического пояса.
79. Homo heidelbergensis Человек гейдельбергский
• 800 тыс.л.н. - 130тыс.л.н.
• 1908 Кения,
Эфиопия,
Германия, и т.д.
80.
• Гейдельбергский человек (лат. Homoheidelbergensis) — ископаемый вид людей,
европейская разновидность человека
прямоходящего (родственный
восточноазиатскому синантропу и
индонезийскому питекантропу), обитавший в Европе
(от Испании и Британии до Белоруссии) 800—130
тыс. лет назад. По-видимому, является потомком
европейского Homo antecessor (к переходной форме
можно отнести Homo cepranensis) и
непосредственным
предшественником неандертальца.
81.
• Относительно поздняя форма, близкая Homo erectus. Отархантропов отличались некоторыми прогрессивными (и
иногда специализированно-неандертальскими) признаками, в
частности большим мозгом и округлым затылком; от
палеоантропов же архаично-низким черепом с толстыми
стенками и очень массивными челюстями с большими зубами.
• Название происходит от грота Гейдельберг в западной
Германии, где в 1907 г. при разработке карьера была найдена
огромная и массивная челюсть гоминида,
послужившая голотипом для описания вида.
• Промежуточность облика здесь, как и в случае "ранних Homo",
приводила к тому, что в различных публикациях гоминиды этого
типа рассматривались в составе как поздних архантропов, так и
ранних палеоантропов.
• Иногда гоминид данного типа разделяют на несколько видов например, Homo antecessor для относительно древних находок
в Испании, Homo rhodesiensis для находок из Южной и
Восточной Африки.
82. Homo helmei
• 500 тыс.л.н. – 130 тыс.л.н.• 1935 Африка
83.
• Африканские формы палеоантропов, неимеющие специализированных признаков
европейских неандертальцев. Часто их
относят в "ранним архаичным Homo sapiens"
или "поздним архаичным Homo sapiens".
• Голотипом Типовой экземпляр
биологического вида. В палеонтологии эталонная находка, окаменелость, по которой
описан ископаемый вид. для вида послужил
своеобразный череп "Флорисбад",
найденный Т.Ф.Дрейером в Южной Африке в
1932 году. В строении черепа Homo
helmei мозаично сочетаются архаичные и
сапиентные черты.
84. Homo neanderthalensis Неандерталец
• 130 тыс.л.н. - 28тыс.л.н.
• 1864 Африка,
Европа, Ближний
Восток
85.
• Неандерта́лец, человекнеандертальский (лат. Homo
neanderthalensis или Homo
sapiens neanderthalensis; в
советской литературе также
именовался палеоантропом)
— ископаемый вид человека,
обитавший 130—28 тыс. лет
назад, и который, согласно
современным научным
данным, частично является
предком современного
человека.
86.
Специализированный к хищничеству вариант человека. Обладали
множеством вполне человеческих черт строения и поведения, но всё
же заметно отличались от нас - в том числе, значительной
массивностью скелета и черепа. Вероятно, многие особенности
неандертальцев Европы сформировались под влиянием тяжелейших
условий ледникового периода около 70-60 тысяч лет назад. Интересно,
что у некоторых представителей Homo neanderthalensis объем
головного мозга превышал значения, типичные для современного
человека.
Кости неандертальцев были исторически первыми обнаруженными
останками ископаемых . К настоящему времени неандертальцы наиболее тщательно изученный вид ископаемых людей.
Термин "неандерталец" имеет не вполне определенные границы.
Ввиду обширности и неоднородности этой группы гоминид,
используется также ряд терминов: "атипичные неандертальцы" для
ранних неандертальцев (периода 130-70 т.л.н.),
"классические неандертальцы" (для европейских форм периода 70-40
т.л.н.), "пережиточные неандертальцы" и т. д.
По новейшим данным, неандертальцы могли скрещиваться с людьми
современного типа, и современные неафриканские популяции Homo
sapiens имеют примерно 2.5% неандертальских генов.
87. Homo floresiensis Человек флоресский
• 95 тыс.л.н. - 12тыс.л.н.
• 2004 Флорес
88.
• Человек флоресский (Homo floresiensis BROWN ETAL., 2004) — возможно, карликовый ископаемый вид
людей. Именуются также «хоббитами» по аналогии с
существами, придуманными Дж. Р. Р. Толкином.
Останки Homo floresiensis обнаружены
в 2003 вИндонезии (остров Флорес, пещера Лианг
Буа), где найдены несколько скелетов, разной
степени сохранности возрастом приблизительно в
17—95 тысяч лет.
• Основой для описания этого сильно "уклонившегося"
вида гоминид послужили сенсационные находки на
острове Флорес. В пещере обнаружено довольно
много костей, из которых набольший интерес
представляют находки с номерами от LB1 до LB16.
89.
• Наилучшим образом сохранился почти полный скелет Лянг Буа1 (LB1), найденный в сентябре 2003 г. и названный "хоббитом"
(а также «Фло»). Индивид Лянг Буа 1 обладает крайне низким
ростом, сопоставимым с минимальными размерами
современных пигмеев – от 1,04 до 1,09 м. Отношение длины
рук к длине ног у Лянг Буа 1 чрезвычайно большое, далеко
выходящее за пределы индивидуальной изменчивости
современного человека и примерно равное значениям
австралопитеков. Мозг крайне маленький - 400 см3, меньше чем
у шимпанзе.
• Карликовый вид, предположительно возник на
основе Homo erectus или другого вида Homo в условиях
островной изоляции. По другой версии, специфический или
патологический представитель Homo sapiens – пигмей или
микроцефал.
• Изначально был описан как Sundanthropus floresianus.
90. Homo sapiens sapiens Человек разумный
• 100 тыс.л.н.• 1758 Планета
Земля
91.
Челове́к разу́мный (лат. Homo sapiens; в русскоязычных текстах
встречается также написание Хомо Сапиенс илиГомо Сапиенс —
вид рода Люди (Homo) из семейства гоминид в отряде приматов,
единственный живущий в настоящее время. От
современных человекообразных, помимо ряда анатомических
особенностей, отличается значительной степенью
развития материальнойи нематериальной культуры (включая
изготовление и использование орудий труда), способностью к
членораздельной речи иабстрактному мышлению. Человек как
биологический вид является предметом исследования физической
антропологии. Природа и сущность человека является предметом
как философского, так и религиозного диспута.
Человечество — совокупность человеческих индивидов.
Разнообразие культур, форм общественной жизни и социальной
организацииявляется предметом изучения социальных и гуманитарных
наук (социальной и культурной антропологии, социологии, экономики,и
стории и др.).
Неоантропы (др.-греч. νέος — новый и ἄνθρωπος — человек) —
обобщённое название людей современного вида, ископаемых и ныне
живущих.
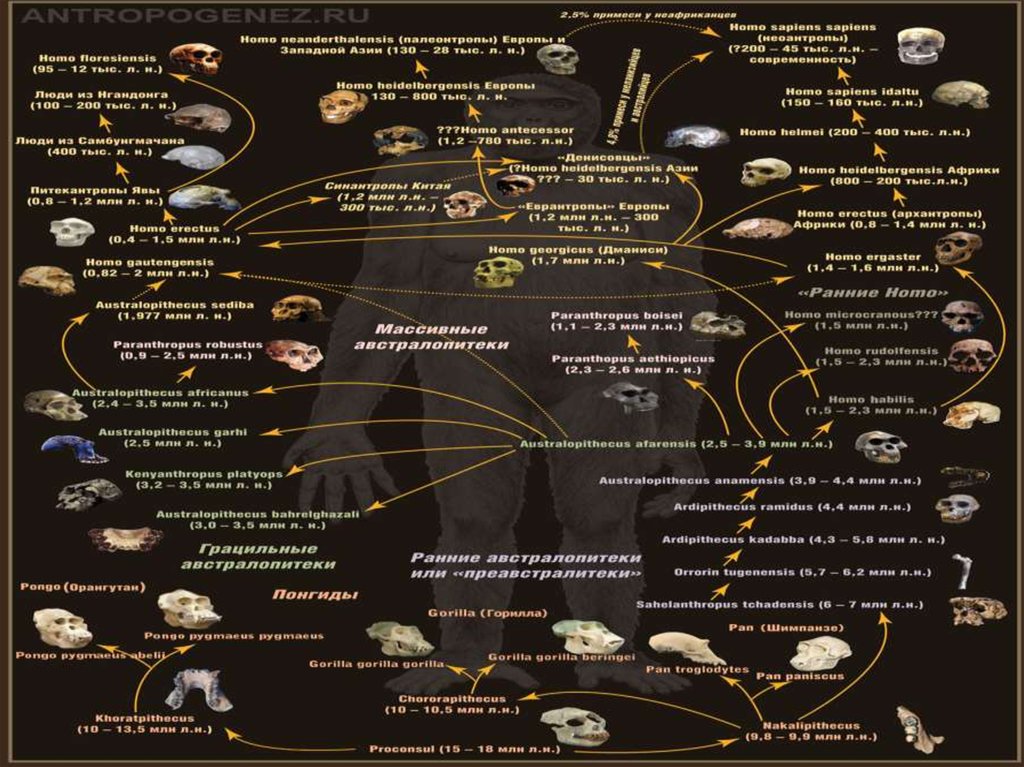
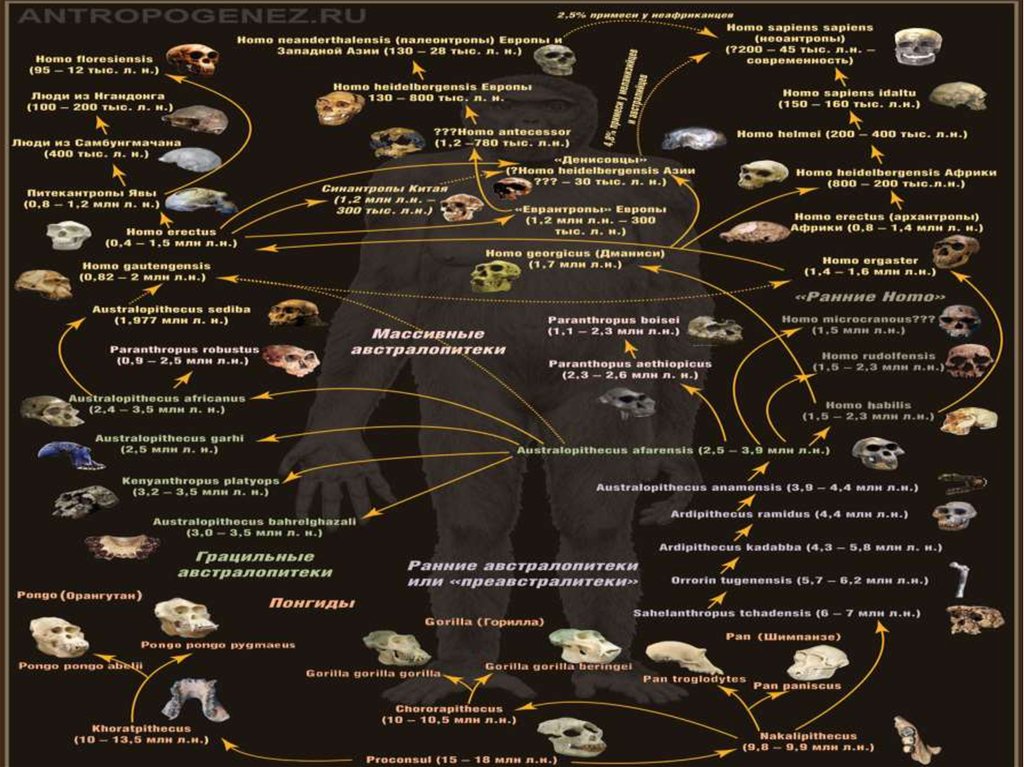
92. Эволюционное дерево человека
• Следующая схема была подготовлена нашимНаучным редактором Станиславом
Дробышевским совместно с журналом
"Техника - Молодежи" .
• Смотреть схему нужно снизу вверх (начиная
от древних миоценовых обезьян).