



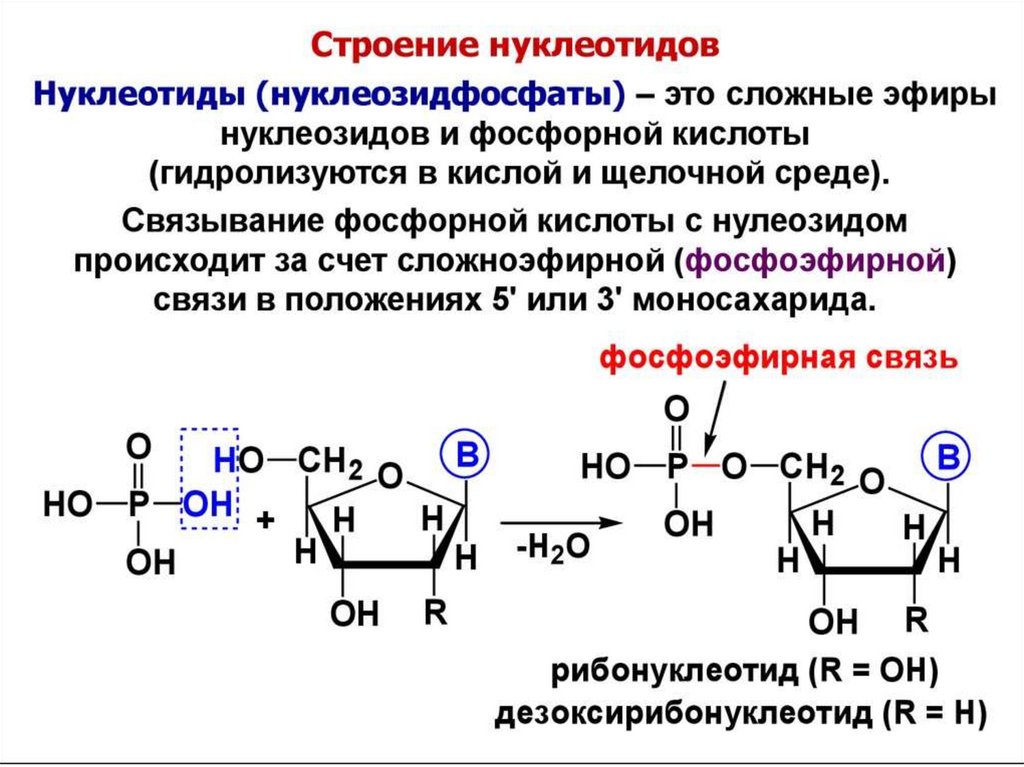
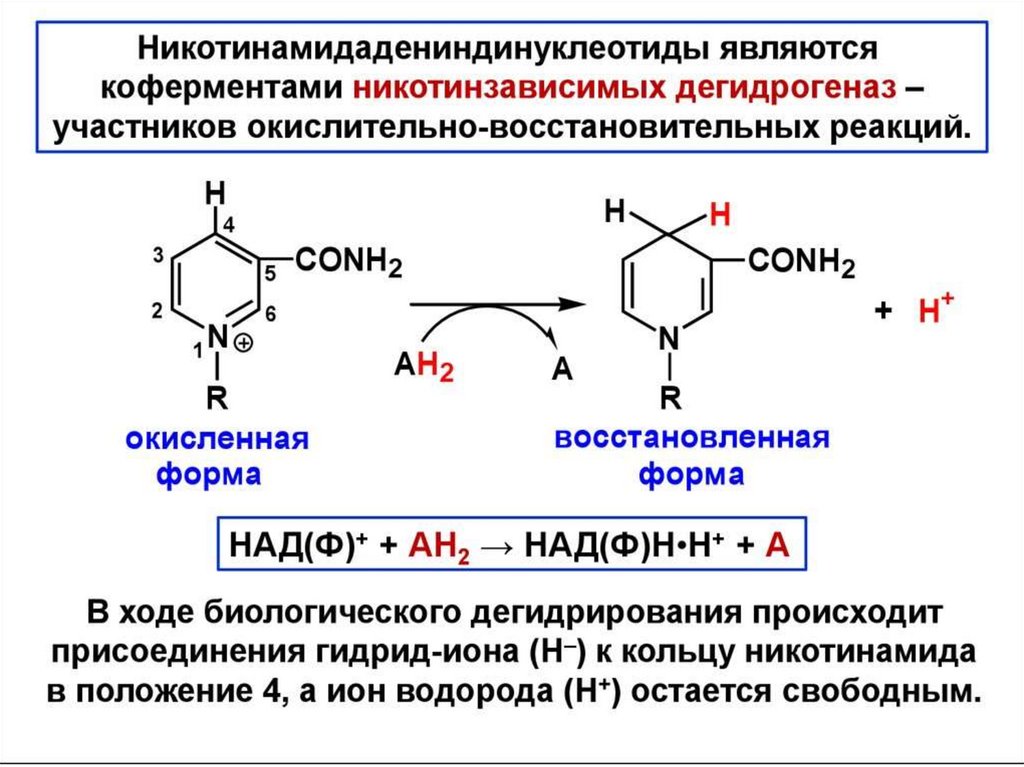


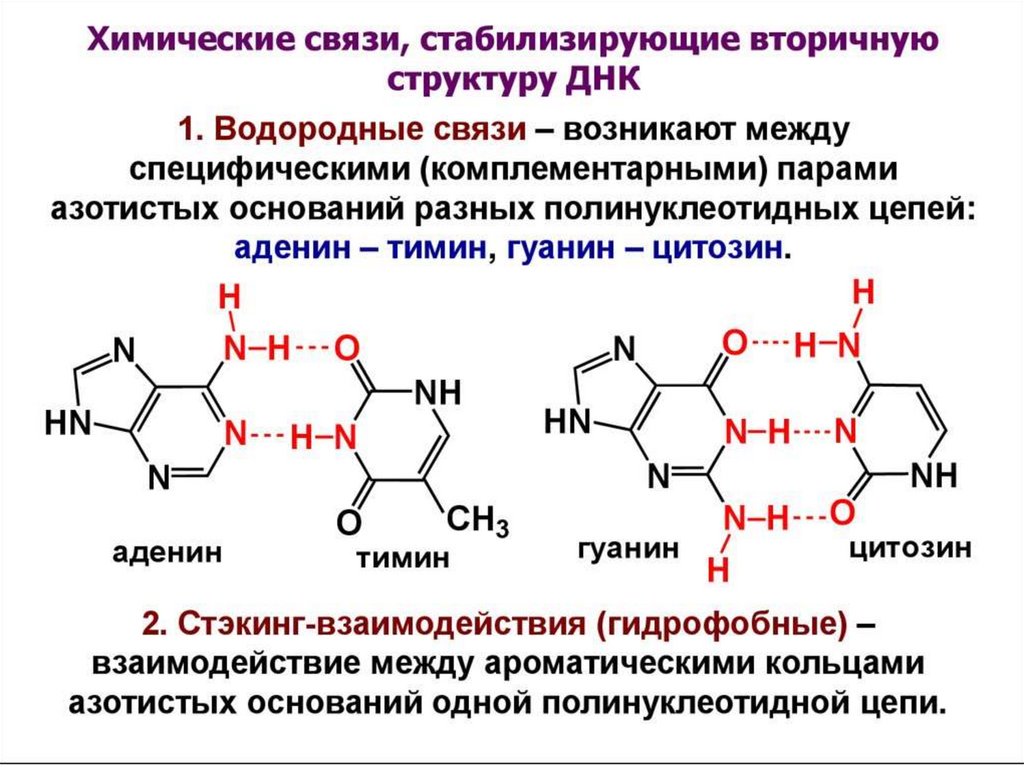

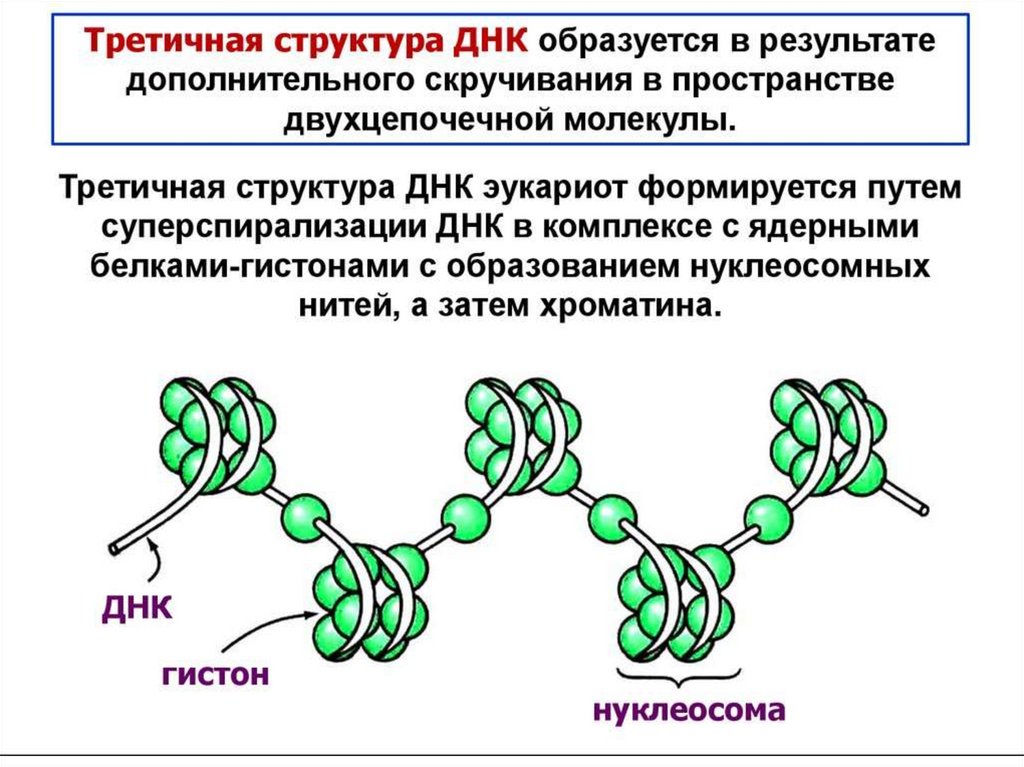
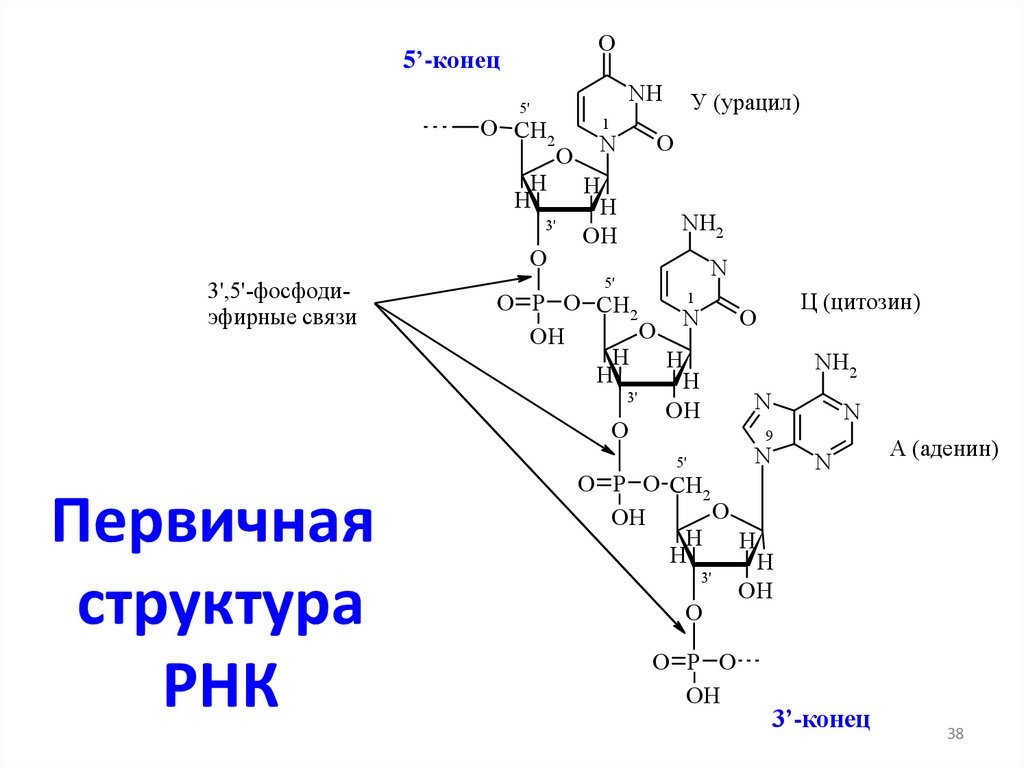
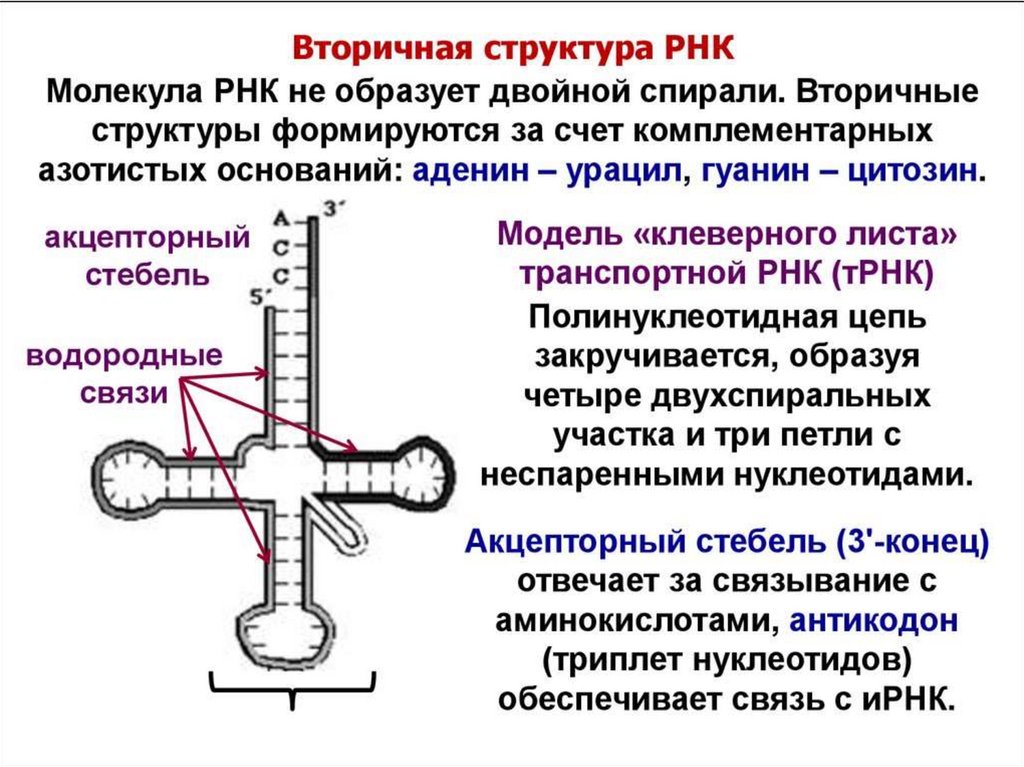




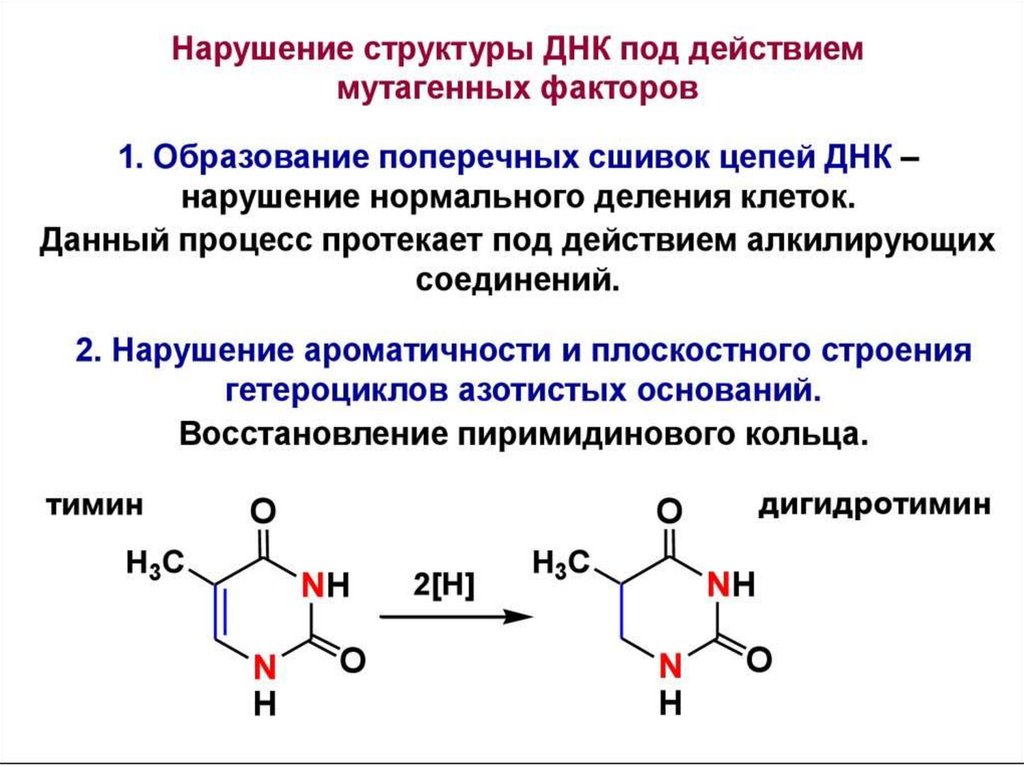
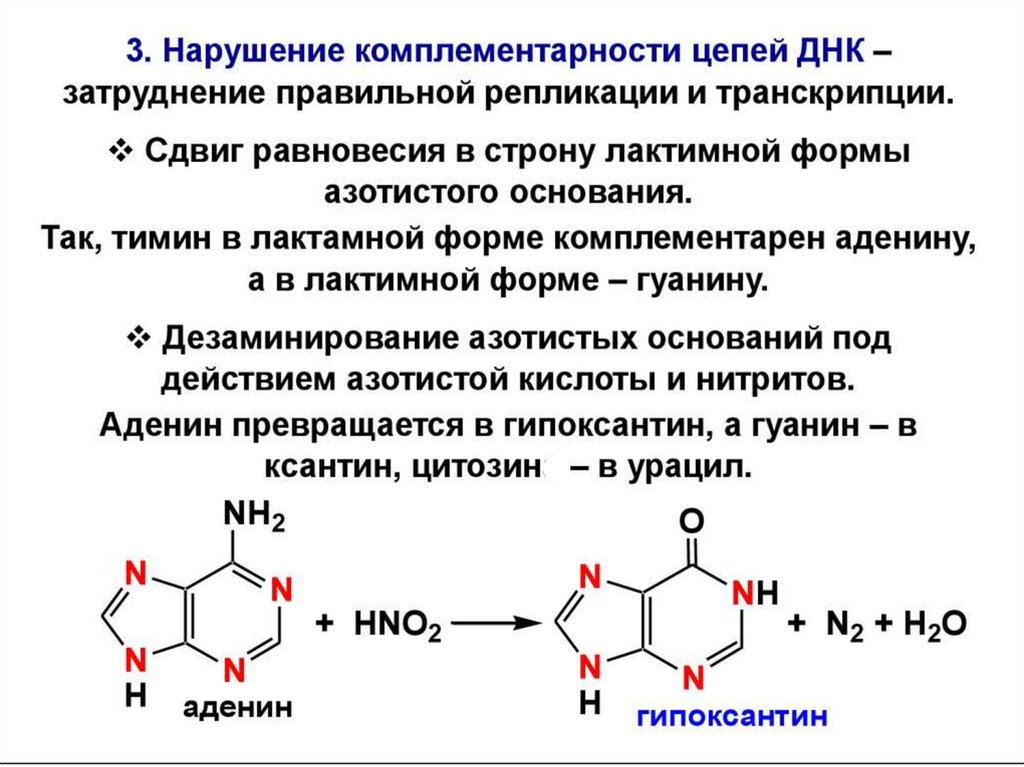
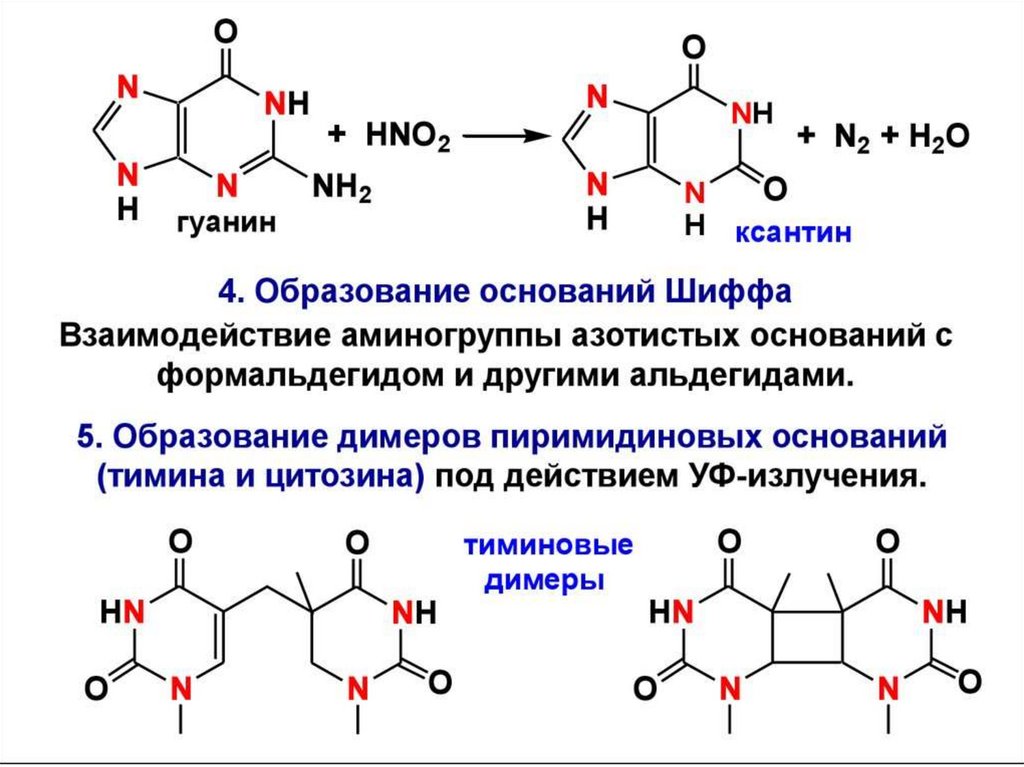




 цепь ДНК выступает в качестве матрицы для синтеза дочерней цепи")















")




 chemistry
chemistrySimilar presentations:









")
Нуклеиновые кислоты
1. Ставропольский государственный медицинский университет Кафедра общей и биологической химии
Нуклеиновыекислоты
NH2
N
O
O
O
N
HO P O P O P O CH2
O
OH
OH OH
H
H
H
H
OH OH
N
N
1
2.
ПЛАН ЛЕКЦИИ:1. Нуклеиновые основания – производные пурина и
пиримидина.
2. Строение мононуклеотидов.
3. Нуклеиновые кислоты: рибонуклеиновая (РНК)
и дезоксирибонуклеиновая (ДНК) кислоты.
4. Механизм передачи генетической информации.
2
3. Нуклеиновые кислоты-
Нуклеиновые кислотыБИОПОЛИМЕРЫ, СОСТОЯЩИЕ ИЗ НУКЛЕОТИДОВИ ОСУЩЕСТВЛЯЮЩИЕ ХРАНЕНИЕ И ПЕРЕДАЧУ
ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ВО ВСЕХ ЖИВЫХ
СИСТЕМАХ
3
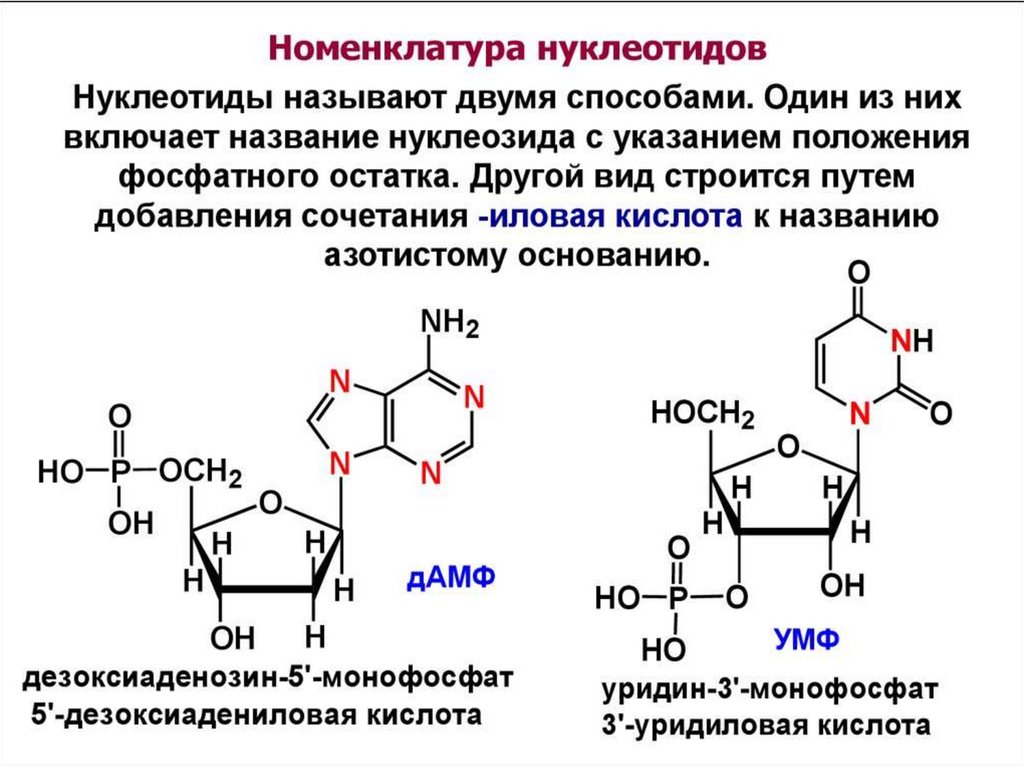
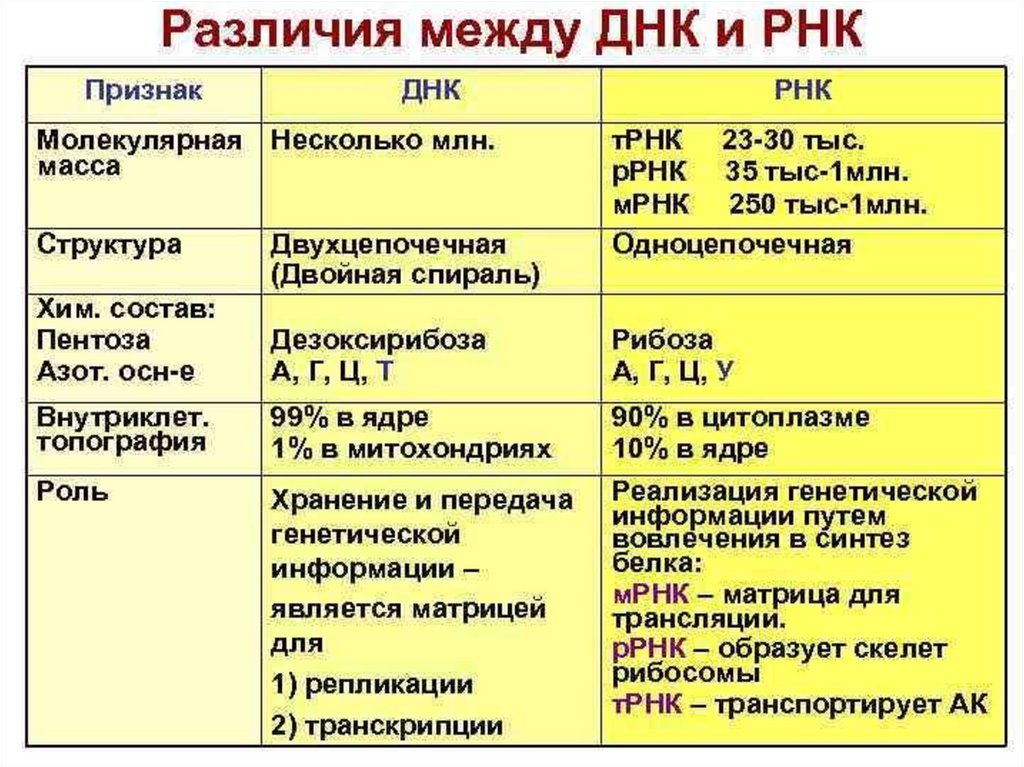
4. Известны два класса НК:
Рибонуклеиновые кислоты (РНК);Дезоксирибонуклеиновые кислоты (ДНК)
РНК
ДНК
β- рибоза
β- дезоксирибоза
урацил
аденин, гуанин, цитозин
фосфаты
тимин
аденин, гуанин,
цитозин, фосфаты
4
5.
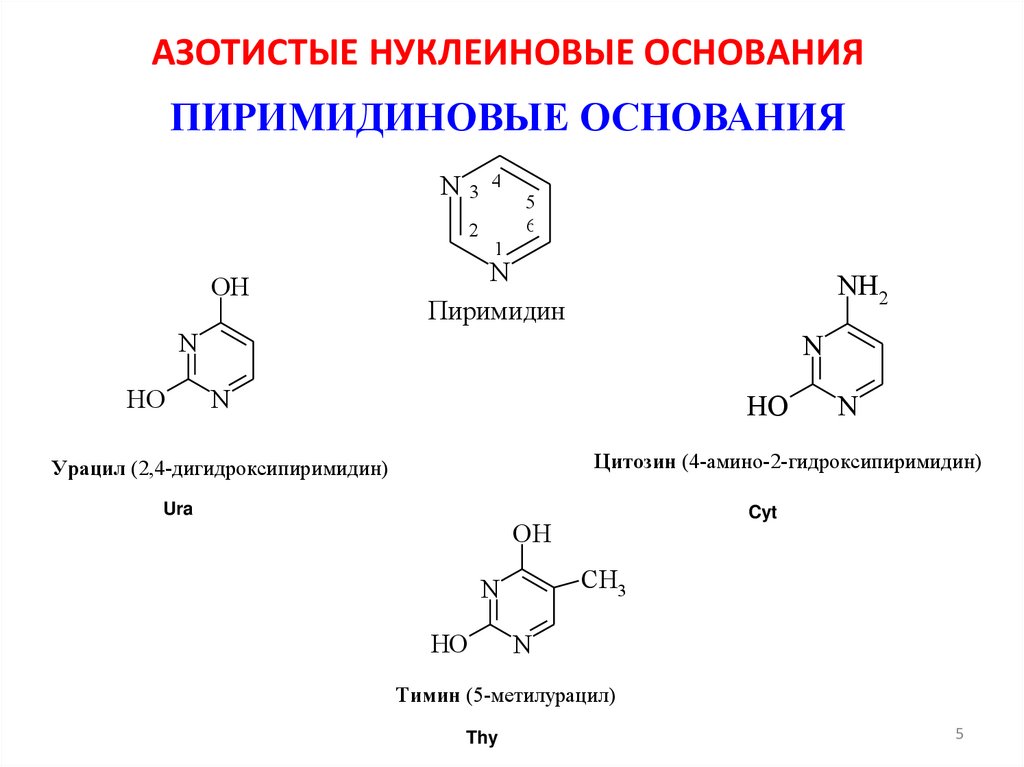
АЗОТИСТЫЕ НУКЛЕИНОВЫЕ ОСНОВАНИЯПИРИМИДИНОВЫЕ ОСНОВАНИЯ
N3 4
2
OH
5
6
1
N
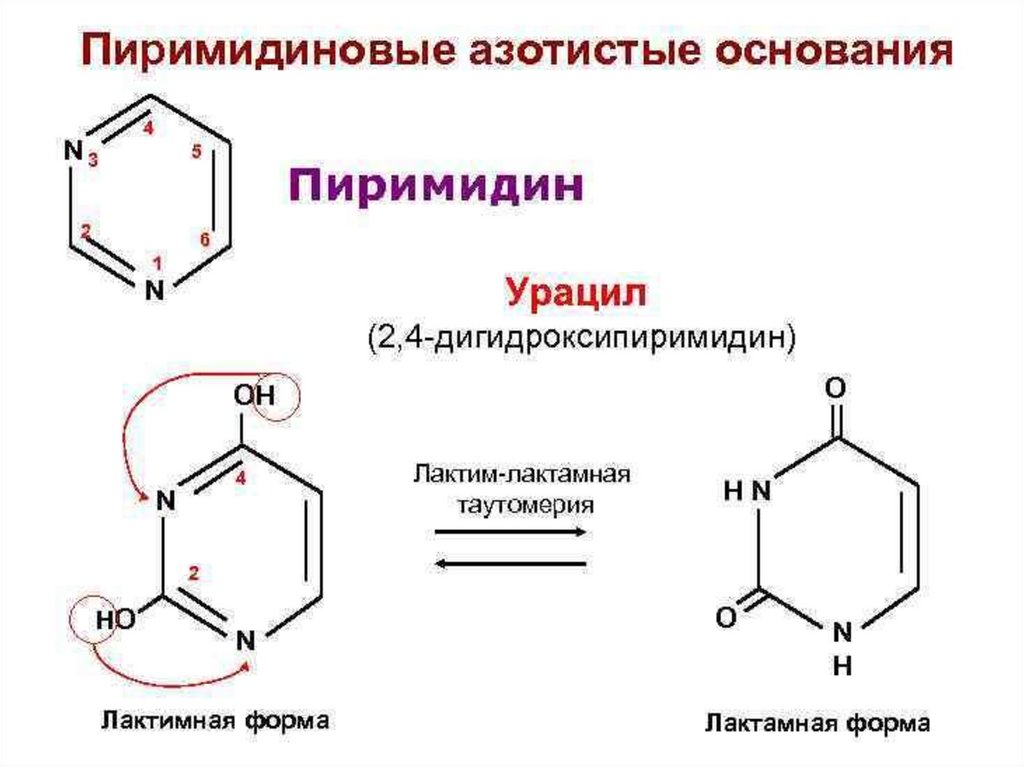
Пиримидин
NH2
N
HO
N
N
HO
N
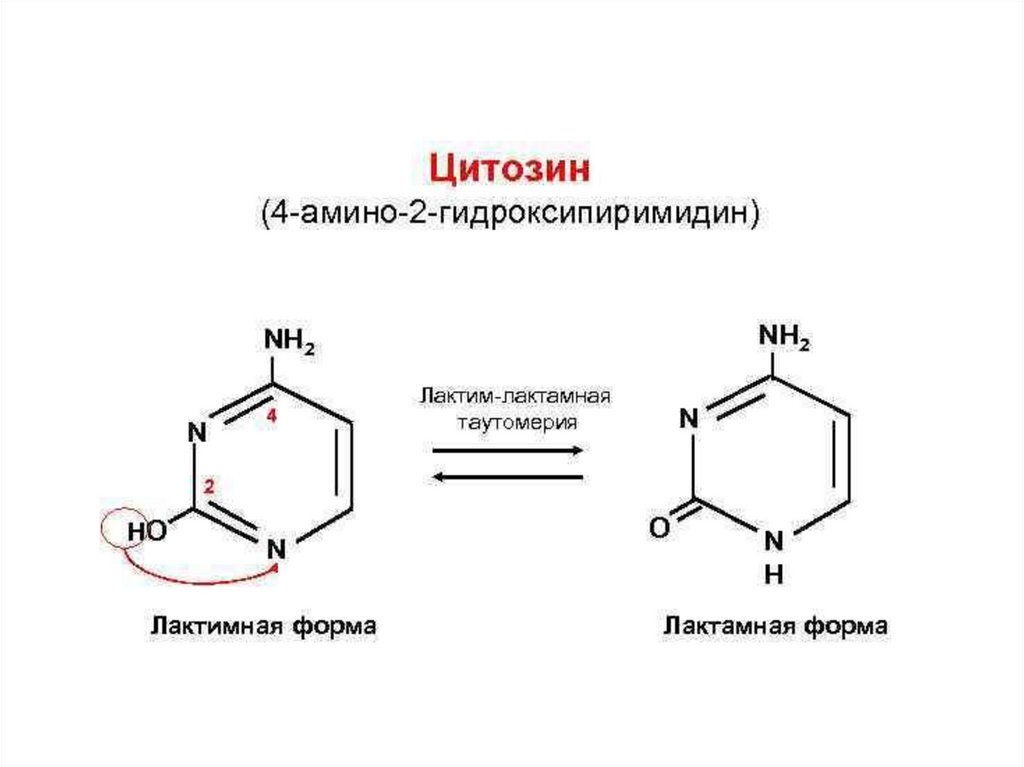
Цитозин (4-амино-2-гидроксипиримидин)
Урацил (2,4-дигидроксипиримидин)
Ura
Cyt
OH
CH3
N
HO
N
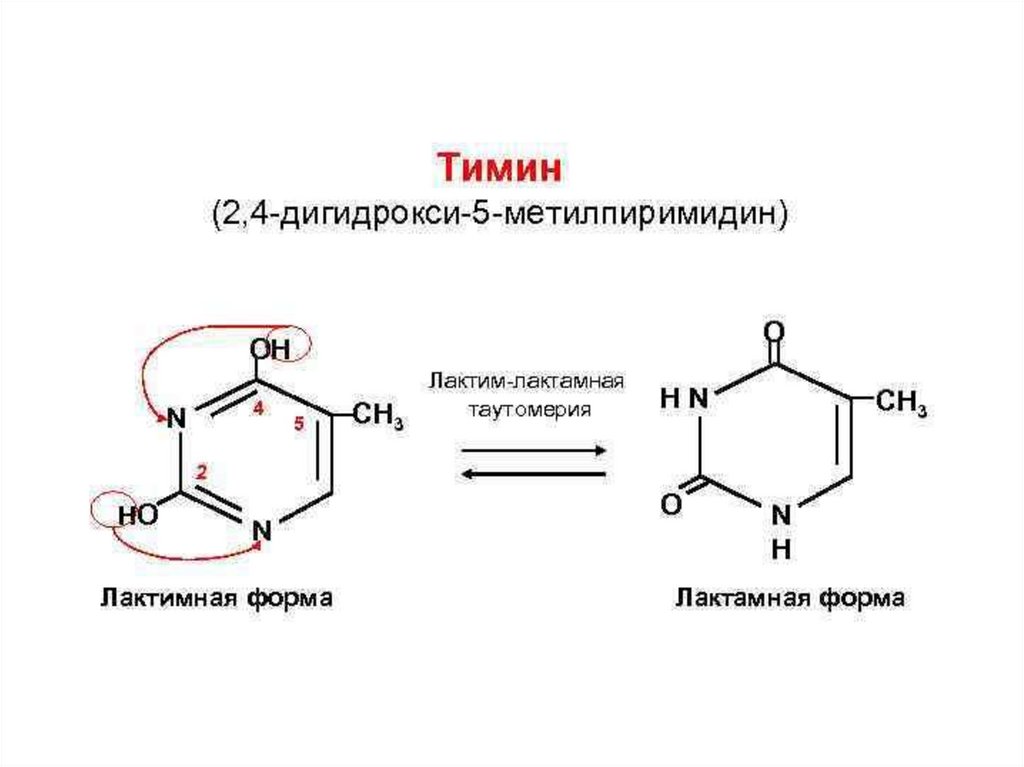
Тимин (5-метилурацил)
Thy
5
6.
67.
78.
89.
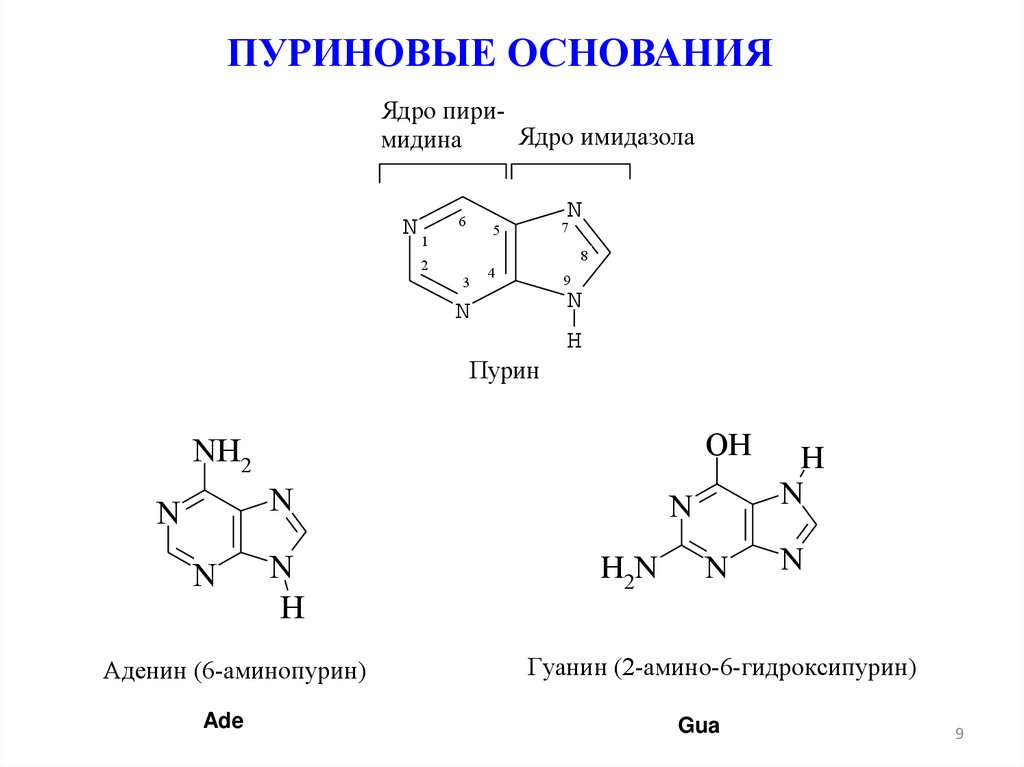
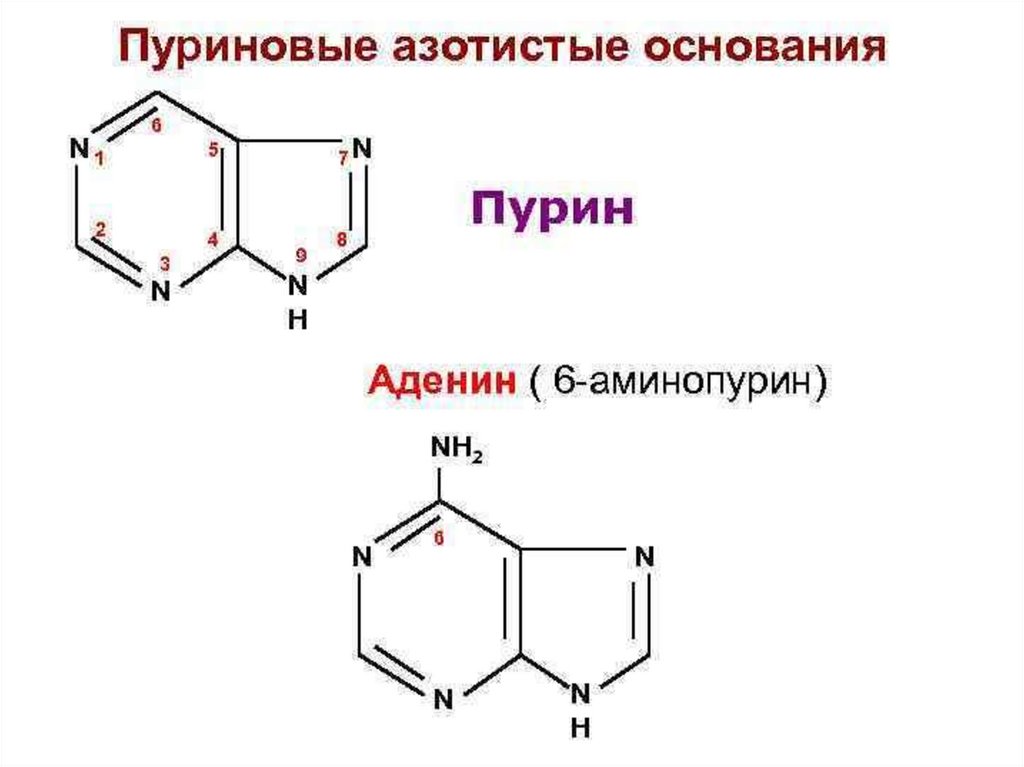
ПУРИНОВЫЕ ОСНОВАНИЯЯдро пириЯдро имидазола
мидина
N1
N
6
7
5
8
2
3
4
9
N
N
H
Пурин
NH2
N
N
N
N
H
Аденин (6-аминопурин)
Ade
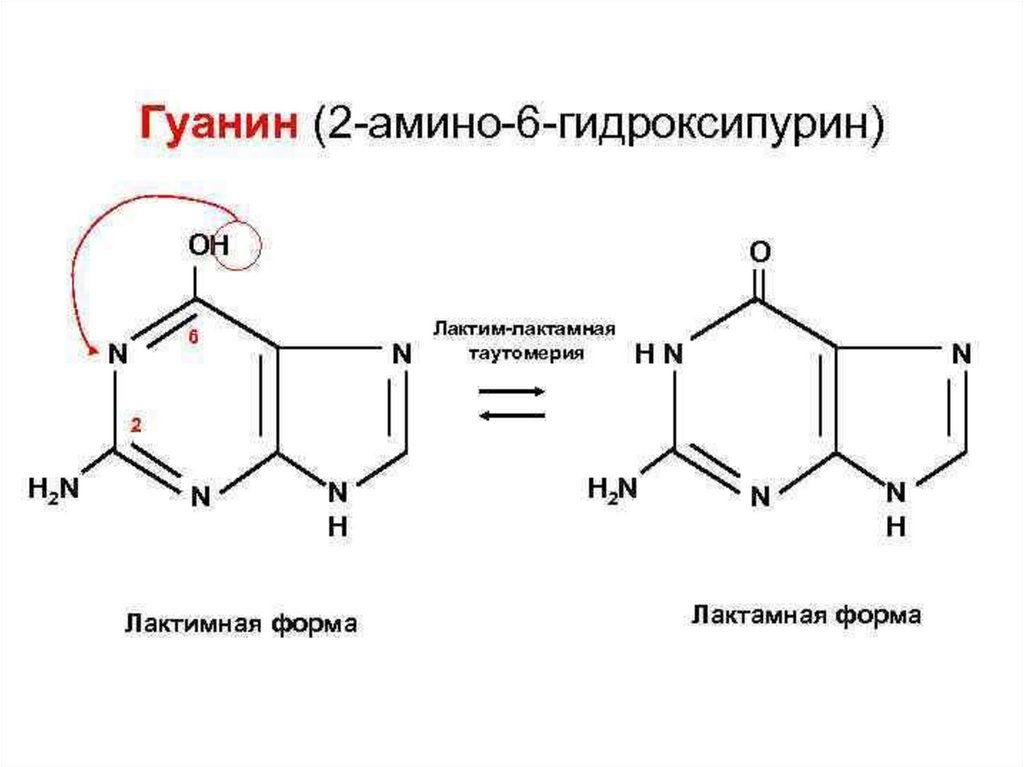
OH
H
N
N
N
N
H2N
Гуанин (2-амино-6-гидроксипурин)
Gua
9
10.
1011.
1112.
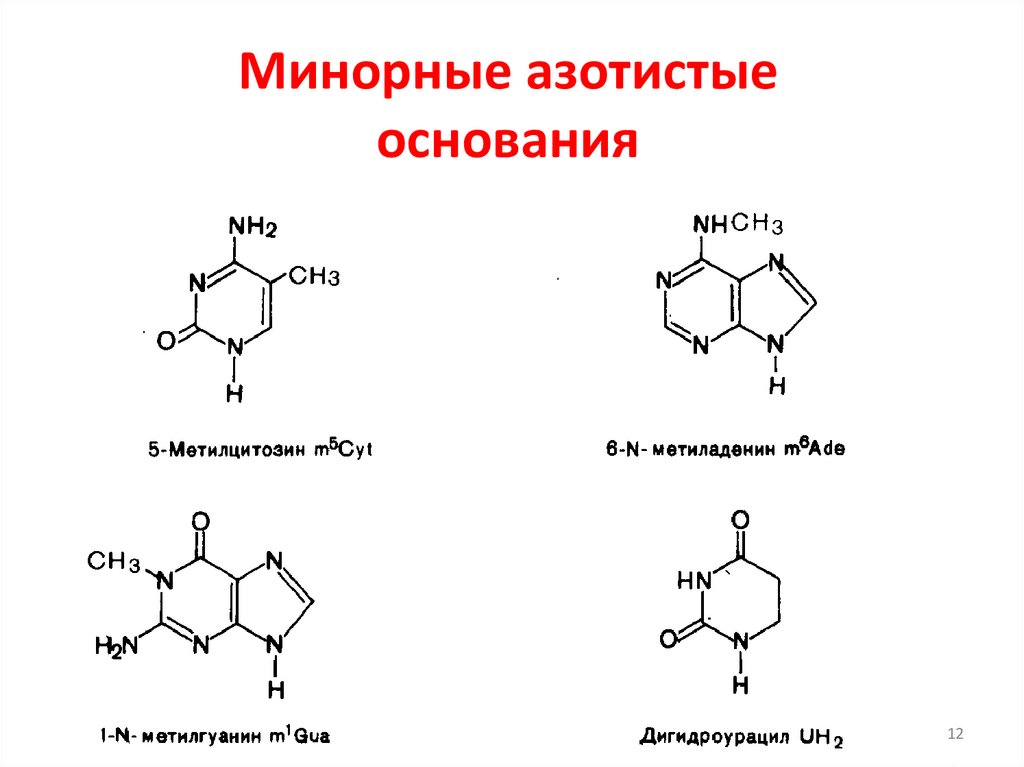
Минорные азотистыеоснования
12
13.
1314.
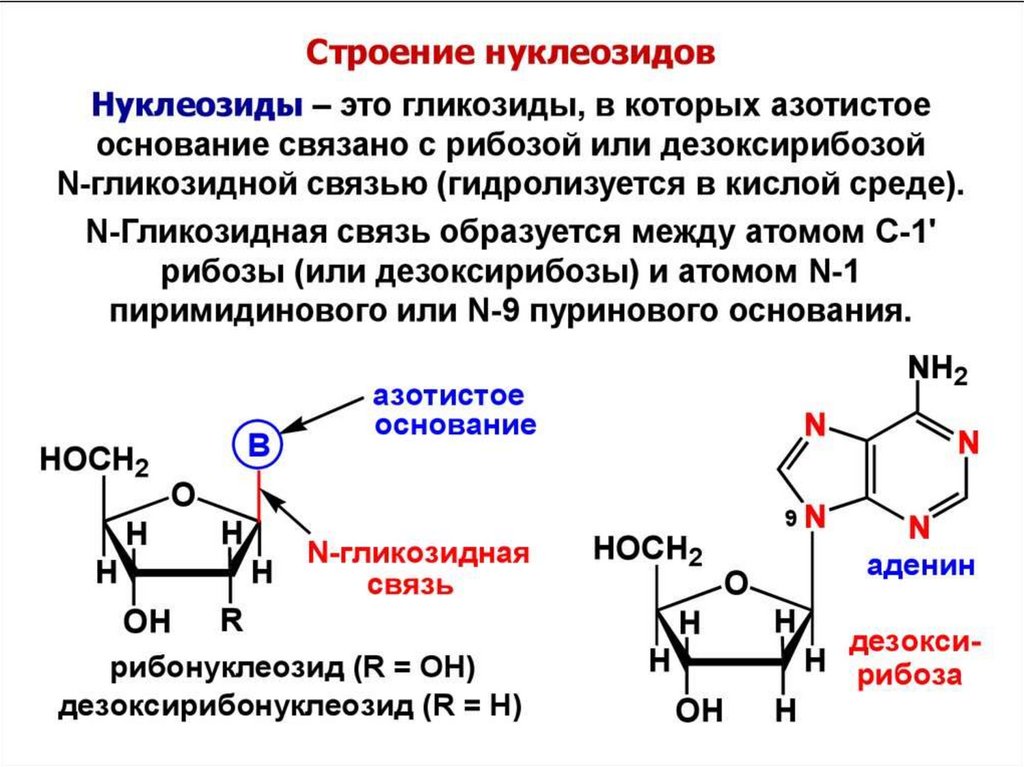
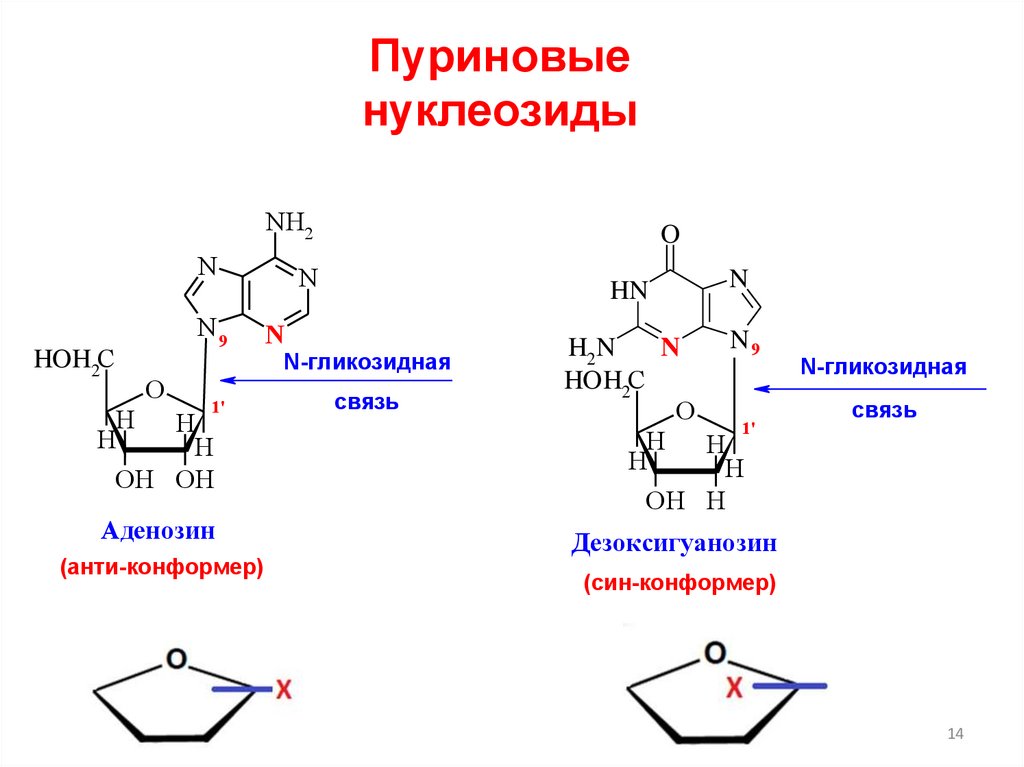
Пуриновыенуклеозиды
NH2
N
N9
HOH2C
O
H
H
1'
H
H
OH OH
Аденозин
(анти-конформер)
O
N
HN
N
N-гликозидная
связь
N
N9
N
H2N
HOH2C
O
1'
H
H
H
H
OH H
N-гликозидная
связь
Дезоксигуанозин
(син-конформер)
14
15.
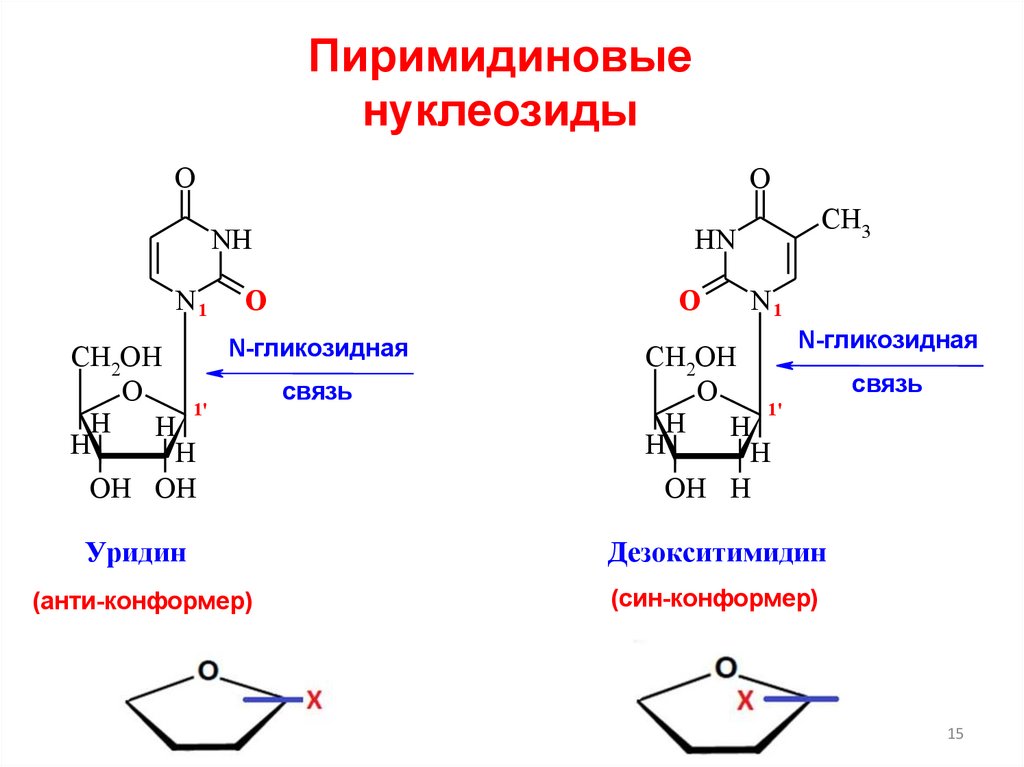
Пиримидиновыенуклеозиды
O
O
NH
N1
O
N-гликозидная
CH2OH
связь
O
1'
H
H
H
H
OH OH
CH3
HN
O
N1
CH2OH
O
1'
H
H
H
H
OH H
N-гликозидная
Уридин
Дезокситимидин
(анти-конформер)
(син-конформер)
связь
15
16.
1617.
1718.
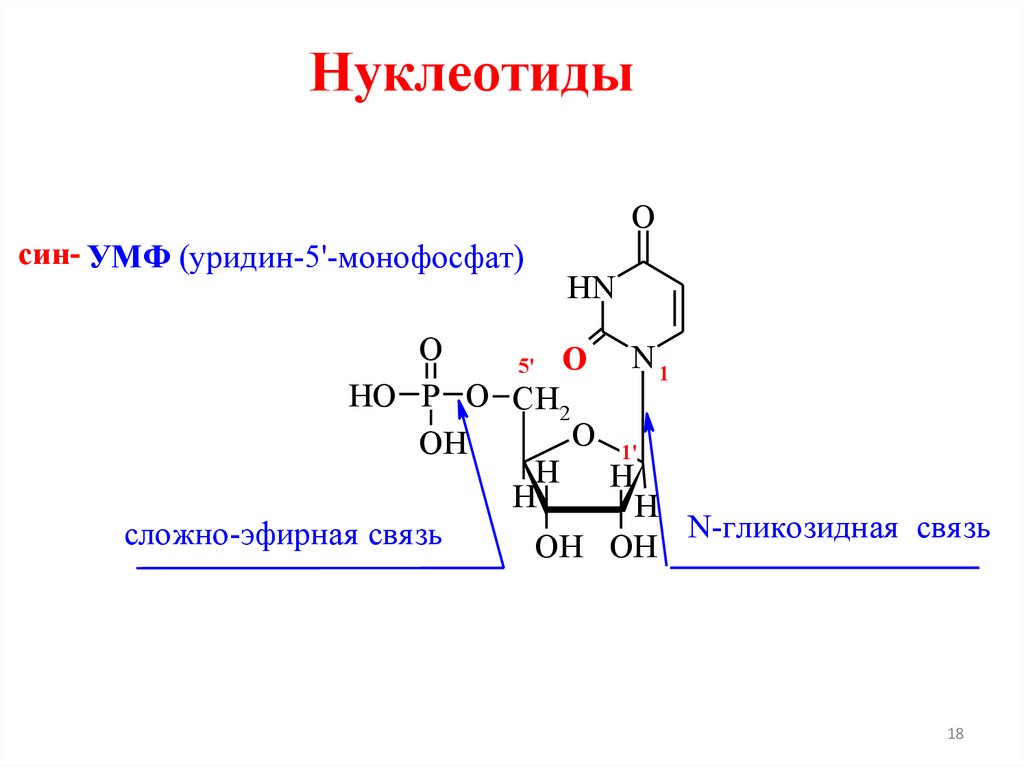
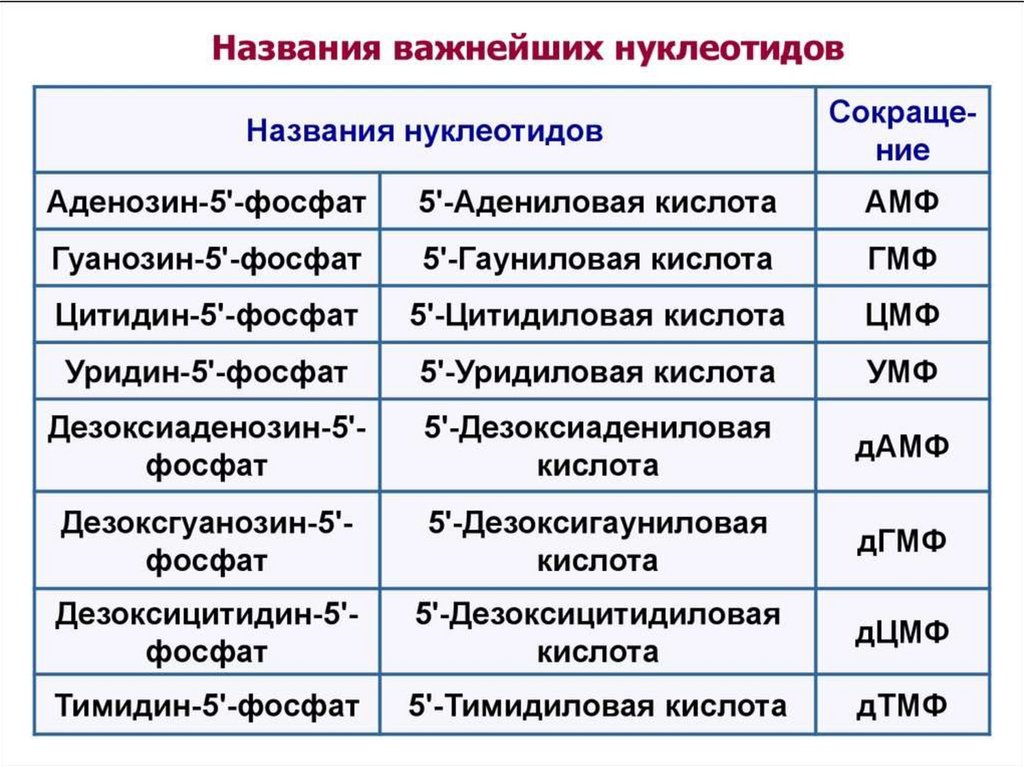
Нуклеотидысин- УМФ (уридин-5'-монофосфат)
O
HN
O
N1
5' O
HO P O CH2
O 1'
OH
H
H
H
H
N-гликозидная свя зь
сложно-эфирная свя зь
OH OH
18
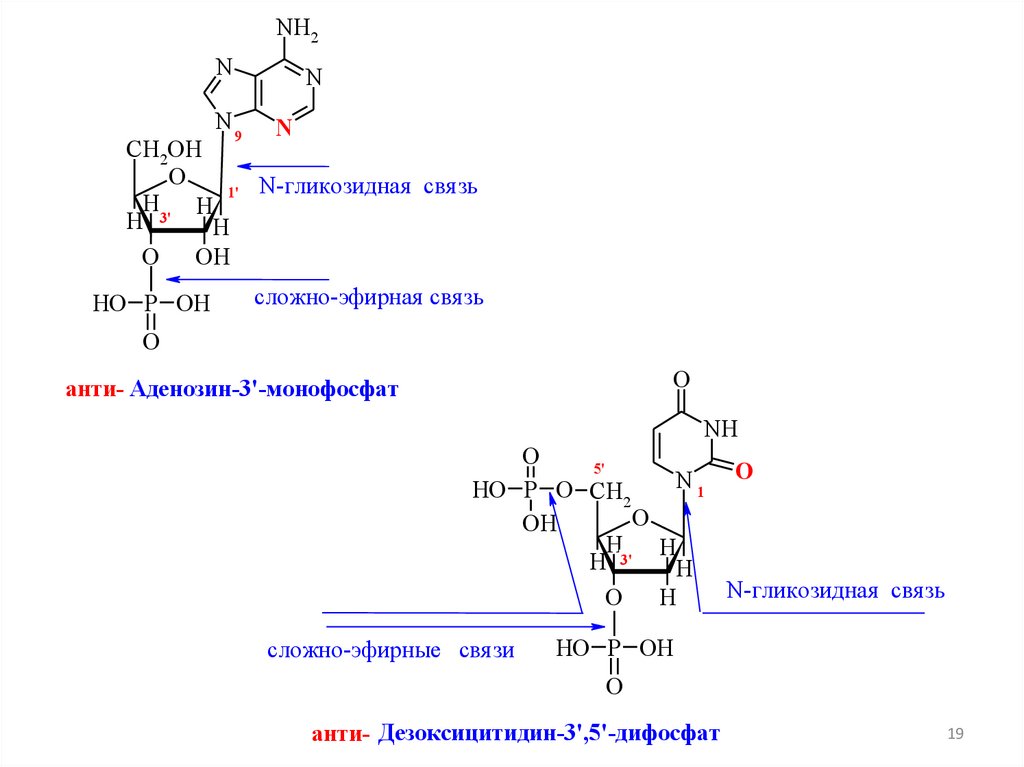
19.
NH2N
N
N
N
9
CH2OH
O
1' N-гликозидная свя зь
H3' H
H
H
O
OH
HO P OH
сложно-эфирная свя зь
O
O
анти- Аденозин-3'-монофосфат
NH
O
5'
N1 O
HO P O CH2
O
OH
H
H
H 3'
H
N-гликозидная свя зь
O
H
сложно-эфирные свя зи
HO P OH
O
анти- Дезоксицитидин-3',5'-дифосфат
19
20.
2021.
2122.
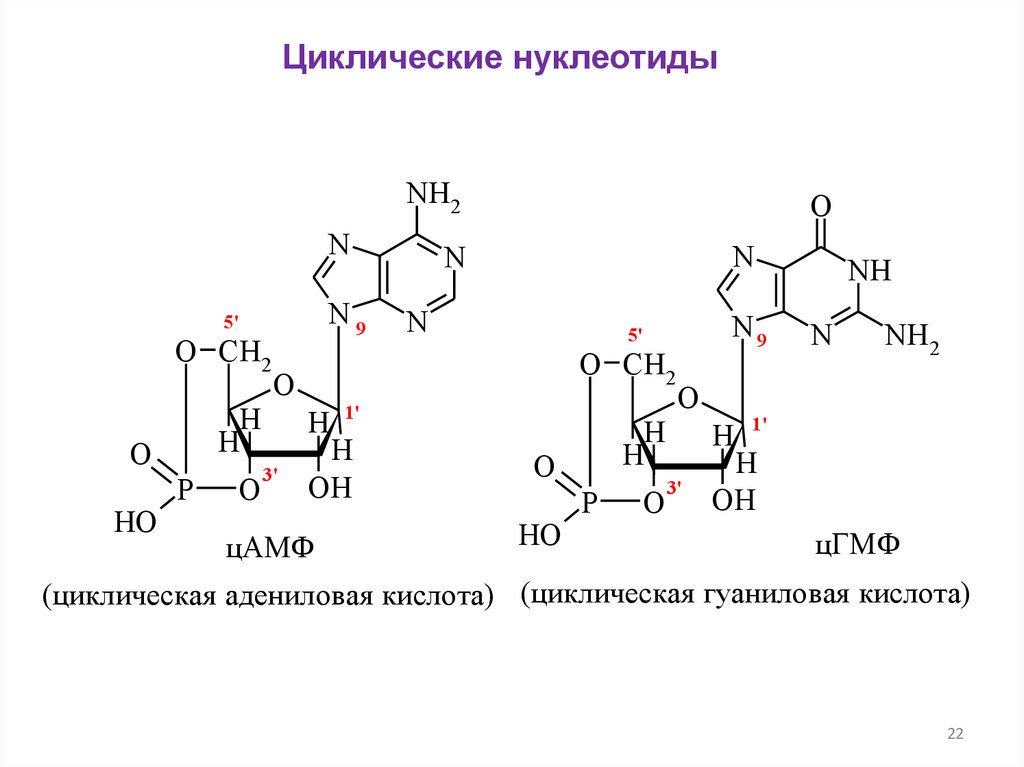
Циклические нуклеотидыNH2
N
N9
5'
O CH2
O
HO
P
O
H
3'
N
N
N
1'
H
OH
цАМФ
N9
5'
O CH2
O
H
H
O
H
H
O
HO
O
P
O
3'
NH
N
NH2
H 1'
H
OH
цГМФ
(циклическая адениловая кислота) (циклическая гуаниловая кислота)
22
23.
2324.
2425.
2526.
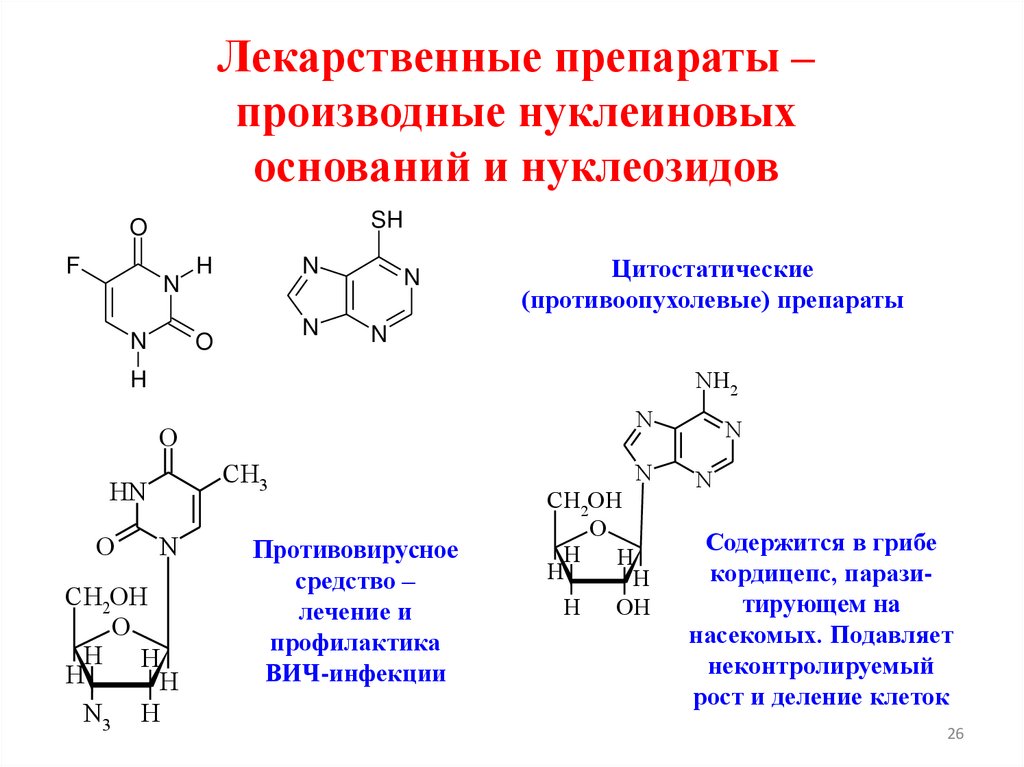
Лекарственные препараты –производные нуклеиновых
оснований и нуклеозидов
SH
O
F
N
N
H
N
N
O
N
Цитостатические
(противоопухолевые) препараты
N
NH2
H
N
O
CH3
HN
O
N
CH2OH
O
H
H
H
H
N3 H
Противовирусное
средство –
лечение и
профилактика
ВИЧ-инфекции
N
CH2OH
O
H
H
H
H
H
OH
N
N
Содержится в грибе
кордицепс, паразитирующем на
насекомых. Подавляет
неконтролируемый
рост и деление клеток
26
27.
2728.
2829.
2930.
3031.
3132.
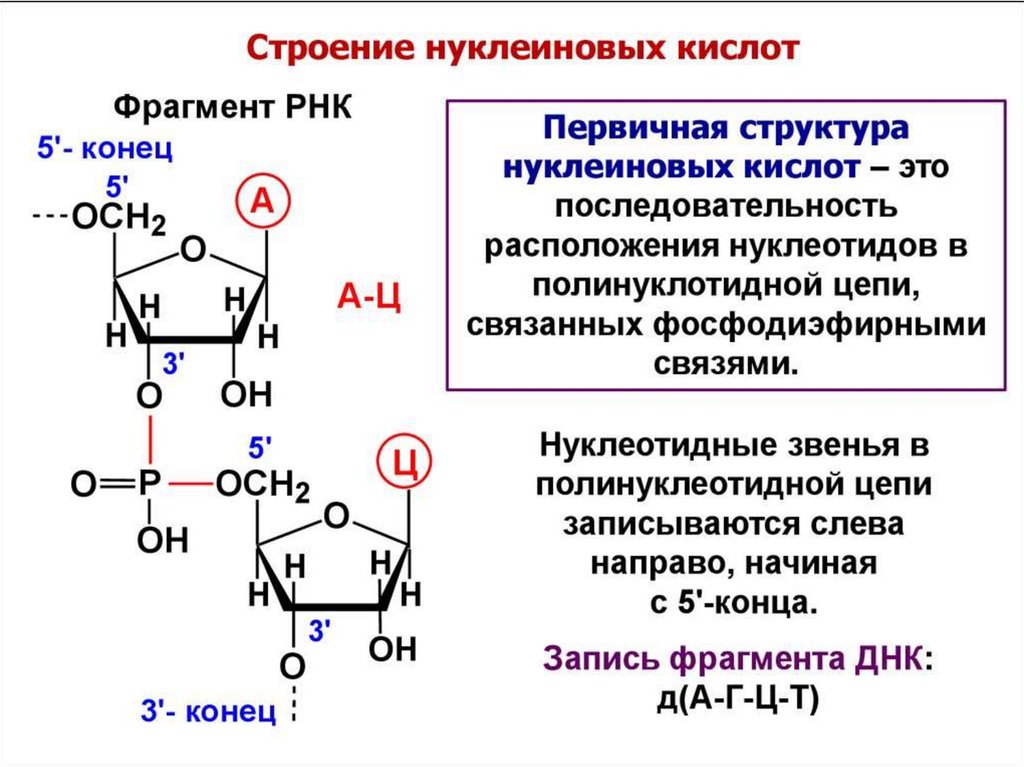
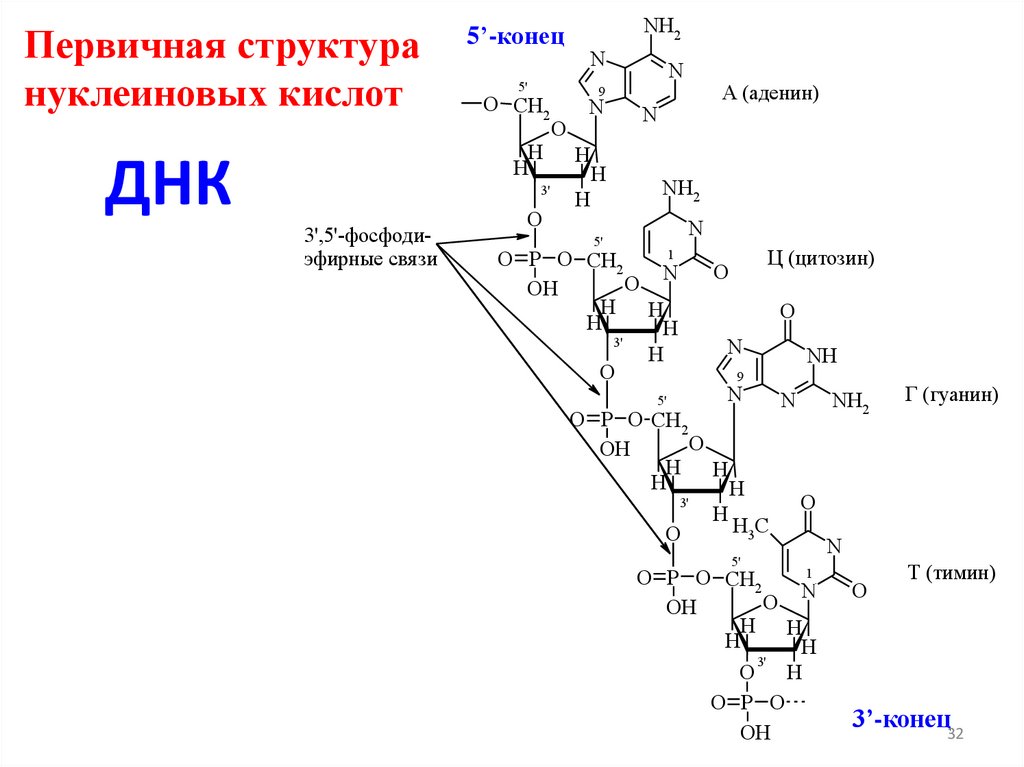
Первичная структурануклеиновых кислот
ДНК
3',5'-фосфодиэфирные свя зи
NH2
5’-конец
N
5'
O CH2
А (аденин)
9
O
N
H
H
H
3'
H
O
N
H
N
NH2
N
5'
1
O P O CH2
N
O
OH
H
H
H
H
3'
H
O
Ц (цитозин)
O
O
N
NH
9
N
5'
O P O CH2
O
OH
H
H
H
H
3'
O
H
N
NH2
O
H3C
5'
Г (гуанин)
1
O P O CH2
N
O
OH
H
H
H
H
3'
O
H
O P O
OH
N
O
Т (тимин)
3’-конец32
33.
3334.
3435.
3536. Правила Чаргаффа
1.Количество пуринов равно количеству
пиримидинов: А+Г=Т+Ц.
2. Количество аденина равно количеству тимина, а
гуанина — цитозину: А=Т, Г=Ц.
3. Количество оснований, содержащих аминогруппу в
положениях 4 пиримидинового и 6 пуринового
ядер, равно количеству оснований, содержащих в
этих же положениях оксогруппу: А+Ц=Г+Т.
Для РНК правила Чаргаффа либо не выполняются,
либо выполняются с некоторым приближением, т.к.
в них содержится много других оснований.
36
37.
3738.
O5’-конец
NH
5'
O CH2
H
H
3'
O
3',5'-фосфодиэфирные свя зи
1
O
N
H
H
OH
У (урацил)
O
NH2
N
5'
1
O P O CH2
N
O
OH
H
H
H
H
3'
OH
O
NH2
N
N
9
N
5'
Первичная
структура
РНК
Ц (цитозин)
O
O P O CH2
O
OH
H
H
H
H
3'
O
O P O
OH
N
А (аденин)
OH
3’-конец
38
39.
3940.
4041.
4142.
4243.
4344.
4445.
4546.
4647.
4748.
• Универсальный биологический процесспередачи генетической информации в
поколениях клеток и организмов,
благодаря созданию точных копий
ДНК.
• ДНК – единственная молекула клетки,
способная к самоудвоению.
49. Скорость репликации ДНК
• У прокариот – 1000 нуклеотидов /сек• У эукариот – 100 нуклеотидов /сек
(медленнее, потому что ДНК сложно упакована –
нуклеосомы и другие уровни упаковки)
50. Место репликации в клеточном цикле
• Репликация ДНК всегда предшествуетделению клетки.
Интерфаза
Деление
Репликация
S-период
(Synthesis)
Каждая дочерняя клетка получает точную копию всей ДНК
51. Принципы репликации
1. Полуконсервативность2. Комплементарность
3. Антипараллельность
4. Униполярность
5. Прерывистость
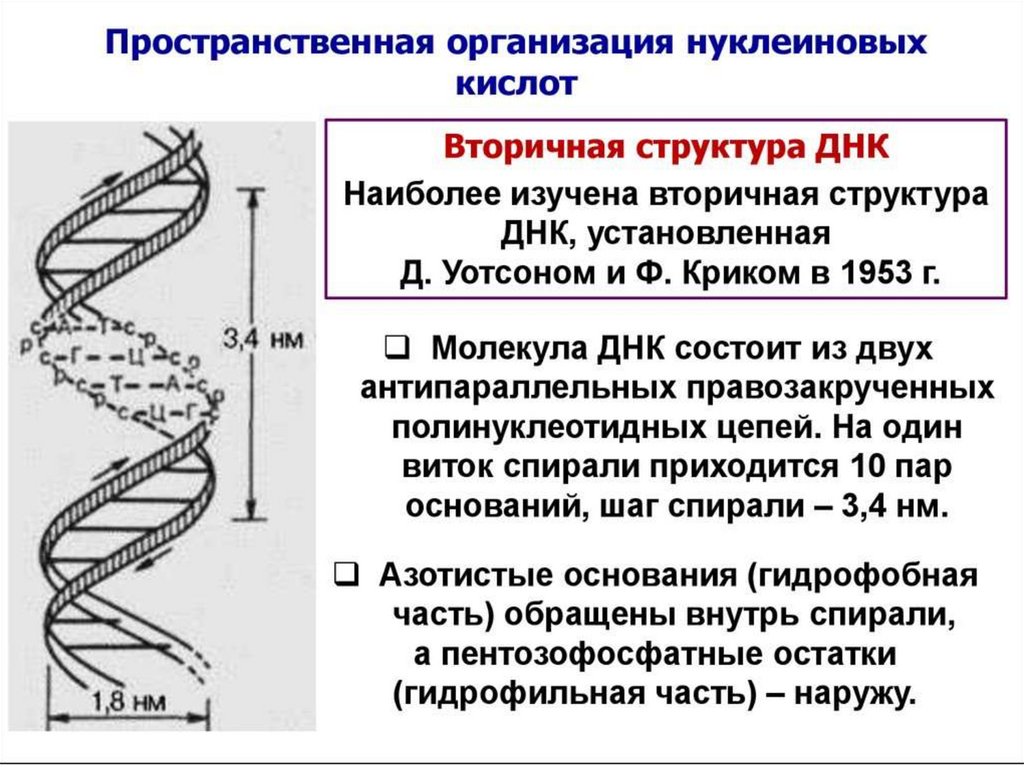
52. Полуконсервативность – каждая исходная (материнская) цепь ДНК выступает в качестве матрицы для синтеза дочерней цепи
Старые цепочки ДНКВновь синтезированные
Полуконсервативный
Консервативный
Дисперсионный
53. Комплементарность
Вновь синтезируемая ( дочерняя) цепь ДНК строитсяпо принципу комплементарности. В состав
растущей цепи включается тот нуклеотид , который
комплементарен нуклеотиду родительской цепи
(аденин с тимином, гуанин с цитозином).
54. Антипараллельность – синтез дочерней цепи ДНК происходит в противоположном от материнской цепи направлении
55. Униполярность:
Удвоение цепи ДНК идет в направлении от 5` концак 3` концу, следовательно новый нуклеотид
присоединяется
к 3 ` концу растущей цепи.
5'
3'
56. Прерывистость репликации
Репликация может идти одновременно в нескольких местахмолекулы ДНК.
ori
ori
ДНК одной
хромосомы
Репликон
Репликон – расстояние между двумя сайтами
начала репликации ori ~ 100 тыс. н.п.
У прокариот вся кольцевая молекула –
один репликон
57. ДНК ГеликазА – фермент, разделяющий цепи двухцепочечной ДНК на одинарные ДНК ТОПОИЗОМЕРАЗА – фермент, изменяющий степень
Основные ферменты репликацииДНК ГЕЛИКАЗА – фермент, разделяющий цепи двухцепочечной
ДНК на одинарные
ДНК ТОПОИЗОМЕРАЗА – фермент, изменяющий степень
сверхспиральности, возникающее при раскручивании двух
цепей в репликативной вилке
ПРАЙМАЗА – фермент, обладающий РНК – полимеразной
активностью; служит для образования РНК-праймеров,
необходимых для инициации синтеза ДНК
ДНК ПОЛИМЕРАЗА – синтезирует новую цепь ДНК по принципу
комплементарности
ДНК ЛИГАЗА – фермент, образующий фосфодиэфирную связь
между двумя полинуклеотидами
SSB (single-strand binding protein)-белки –связывающиеся с
одноцепочечными нитями ДНК и предотвращают
комплементарное спаривание
58. Свойства ДНК-полимеразы
1. Присоединяет по одномунуклеотиду с 3‘ конца растущей
цепочки.
3'
2. Требует для начала работы
спаренного 3‘ конца.
3. Отщепляет один нуклеотид назад,
если он не спарен – т.е.
исправляет свои ошибки.
Логически
связанные
свойства !
59. ДНК-полимераза исправляет ошибки
Если новый нуклеотид не спарен – фермент не можетдвигаться дальше.
Тогда он выедает неверный
нуклеотид и ставит другой.
60. Выводы по репликации ДНК
• В результате репликации каждая дочерняяклетка получает точную копию всей ДНК
содержавшейся в материнской клетке.
• ДНК всех клеток одного организма –
одинаковая, как по количеству молекул,
т.е. хромосом, так и по их нуклеотидному
составу.
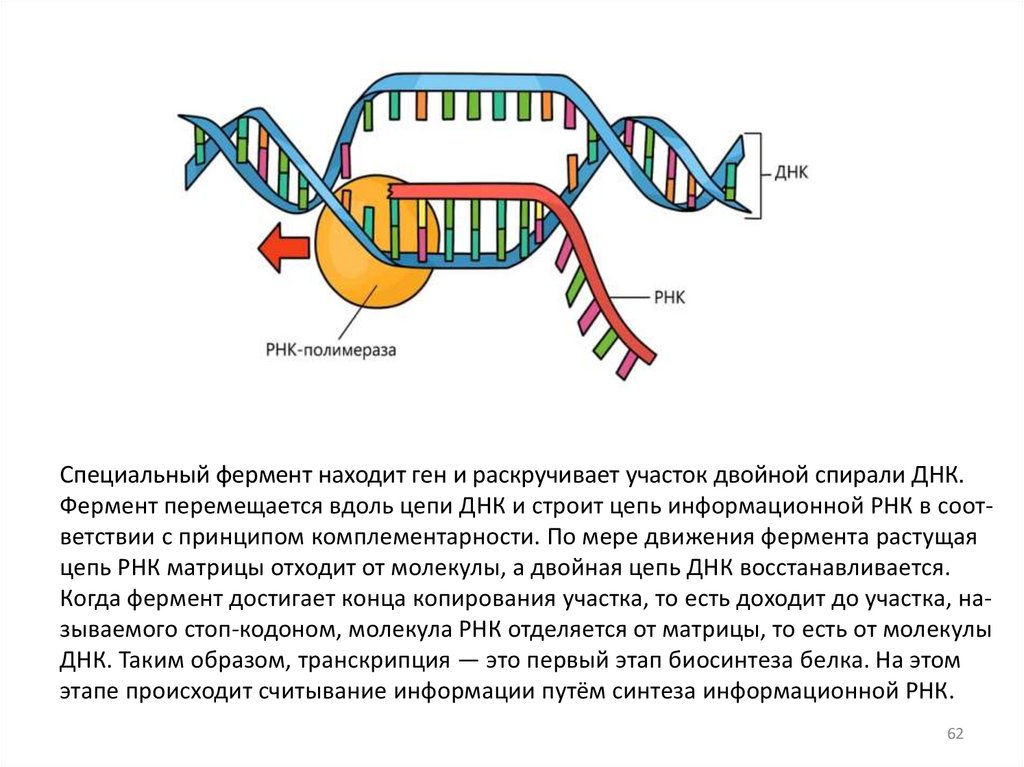
61. Транскрипция
Транскрипция - это первый этап реализациигенетической информации, при котором в клетках
осуществляется биосинтез РНК на матрице ДНК, т.е.
переписывание информации о структуре белка с ДНК
на специальный посредник – м РНК.
62.
Специальный фермент находит ген и раскручивает участок двойной спирали ДНК.Фермент перемещается вдоль цепи ДНК и строит цепь информационной РНК в соответствии с принципом комплементарности. По мере движения фермента растущая
цепь РНК матрицы отходит от молекулы, а двойная цепь ДНК восстанавливается.
Когда фермент достигает конца копирования участка, то есть доходит до участка, называемого стоп-кодоном, молекула РНК отделяется от матрицы, то есть от молекулы
ДНК. Таким образом, транскрипция — это первый этап биосинтеза белка. На этом
этапе происходит считывание информации путём синтеза информационной РНК.
62
63. Общие параметры транскрипции
• Скорость – около 30 нуклеотидов / сек• Частота ошибок – 1 на 104 нуклеотидов, т.е.
на пять порядков выше, чем при
репликации.
• Синтез РНК – гораздо менее точный
процесс, чем синтез ДНК.
64.
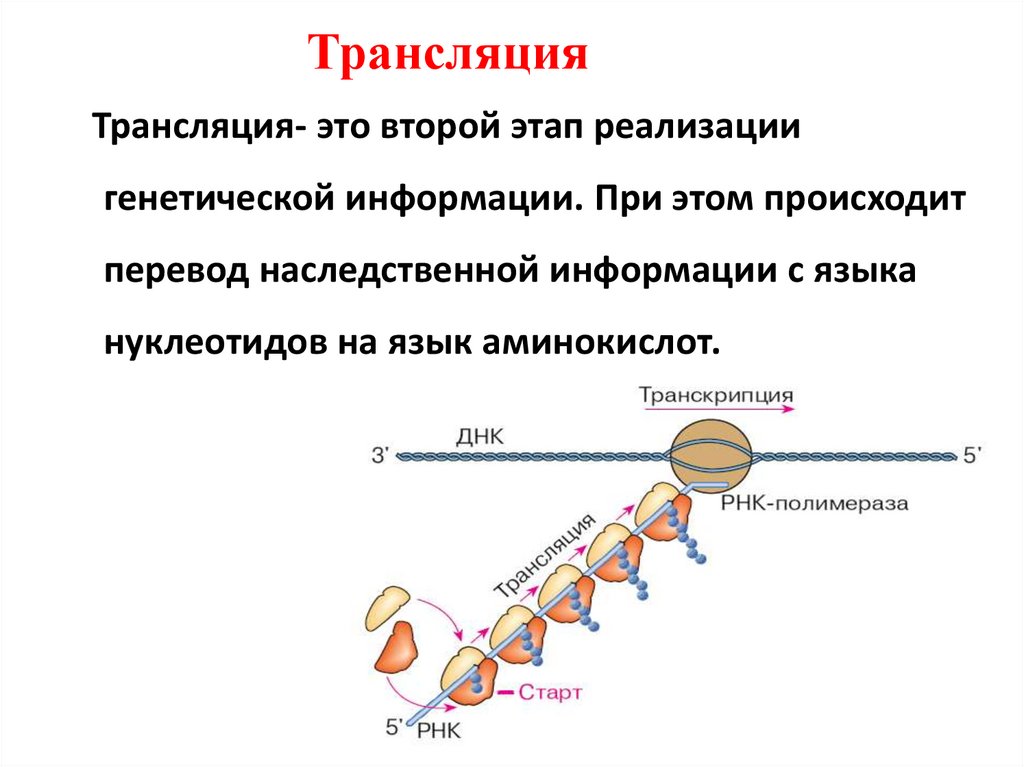
ТрансляцияТрансляция- это второй этап реализации
генетической информации. При этом происходит
перевод наследственной информации с языка
нуклеотидов на язык аминокислот.
65. и-РНК → БЕЛОК
• Синтез белка по матрице и-РНК,осуществляемый на рибосомах
• Самый сложный из матричных синтезов
– не просто копирование, а перевод с
языка нуклеиновых кислот на язык
белков (генетический код).
66.
РибосомыРибосомы — молекулярные машины биосинтеза
белка. Они представляют собой сложный комплекс
молекул рибосомальных РНК (рРНК) и белков.
Каждая рибосома состоит из двух неравных частей,
называемых большой и малой субъединицами.
Большая субъединица примерно в 2 раза больше
малой.
66
67. Участники трансляции:
Аминокислоты;т – РНК;
Аминоацил- т – РНК синтетазы;
м – РНК;
Рибосомы;
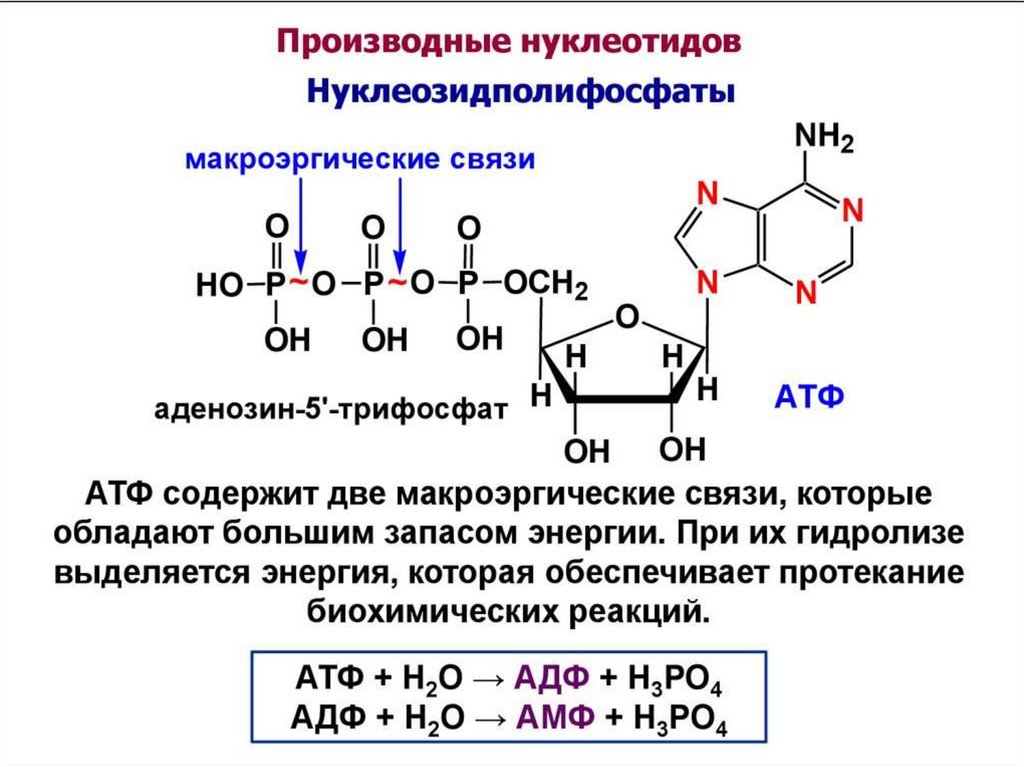
АТФ, ГТФ
Белковые факторы инициации, элонгации, терминации
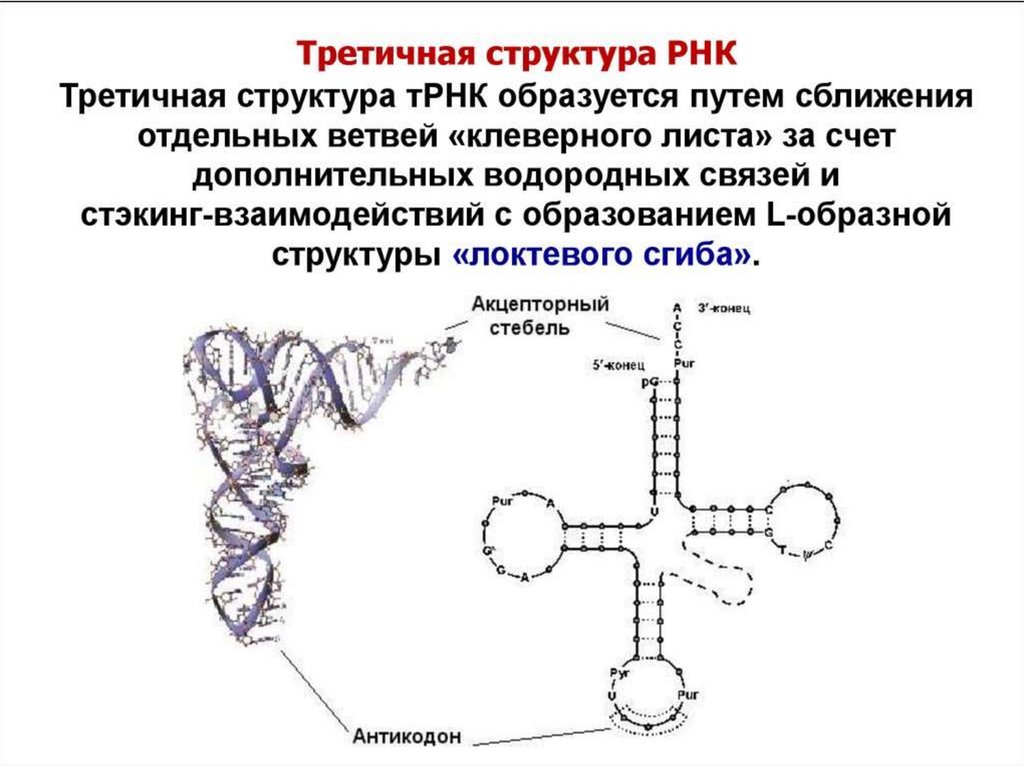
Ионы
68.
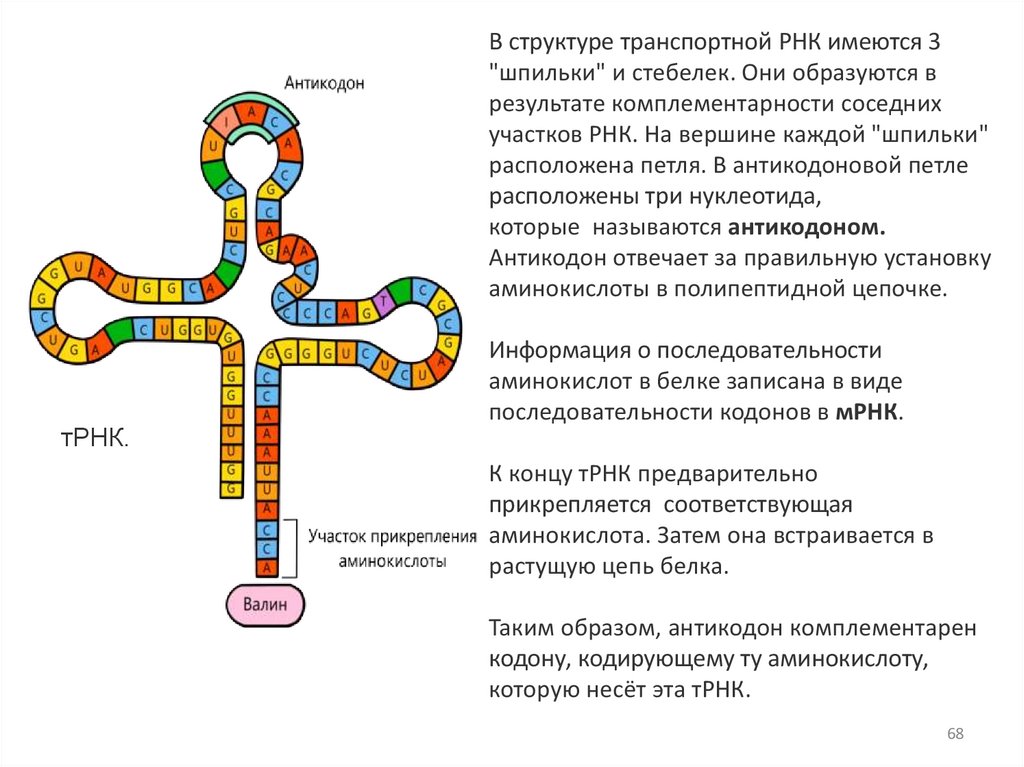
В структуре транспортной РНК имеются 3"шпильки" и стебелек. Они образуются в
результате комплементарности соседних
участков РНК. На вершине каждой "шпильки"
расположена петля. В антикодоновой петле
расположены три нуклеотида,
которые называются антикодоном.
Антикодон отвечает за правильную установку
аминокислоты в полипептидной цепочке.
тРНК.
Информация о последовательности
аминокислот в белке записана в виде
последовательности кодонов в мРНК.
К концу тРНК предварительно
прикрепляется соответствующая
аминокислота. Затем она встраивается в
растущую цепь белка.
Таким образом, антикодон комплементарен
кодону, кодирующему ту аминокислоту,
которую несёт эта тРНК.
68
69.
Этапы трансляции1. Инициация (начало)
2. Элонгация (удлинение)
3. Терминация (окончание)
70.
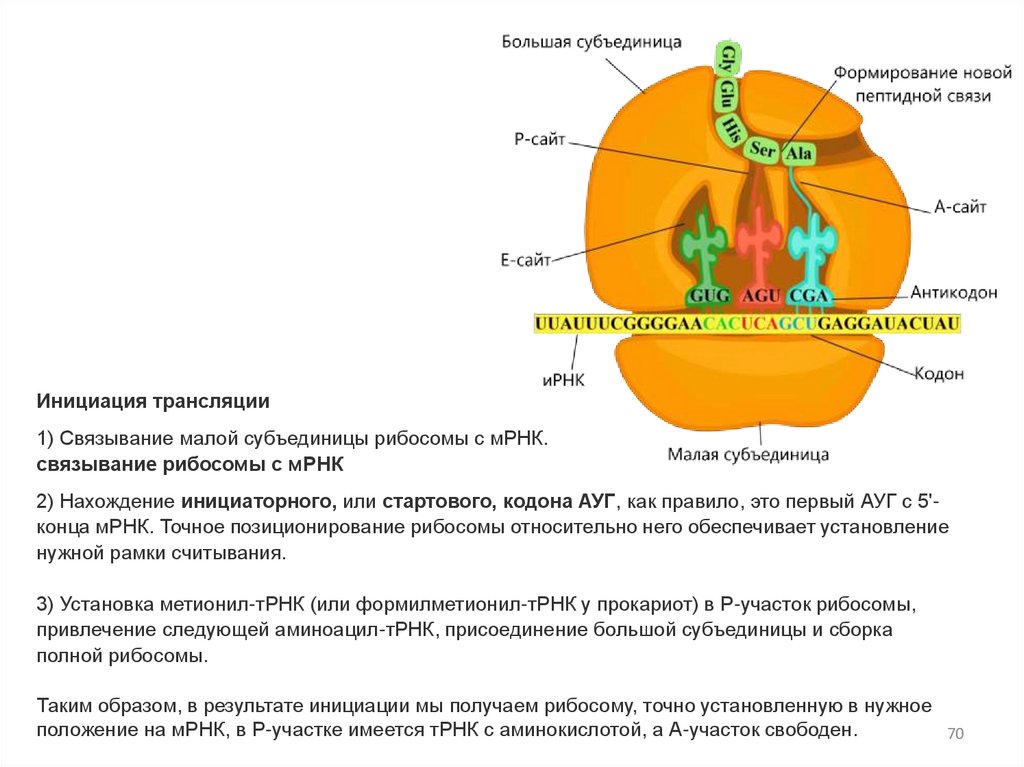
Инициация трансляции1) Связывание малой субъединицы рибосомы с мРНК.
связывание рибосомы с мРНК
2) Нахождение инициаторного, или стартового, кодона АУГ, как правило, это первый АУГ с 5'конца мРНК. Точное позиционирование рибосомы относительно него обеспечивает установление
нужной рамки считывания.
3) Установка метионил-тРНК (или формилметионил-тРНК у прокариот) в Р-участок рибосомы,
привлечение следующей аминоацил-тРНК, присоединение большой субъединицы и сборка
полной рибосомы.
Таким образом, в результате инициации мы получаем рибосому, точно установленную в нужное
положение на мРНК, в Р-участке имеется тРНК с аминокислотой, а А-участок свободен.
70
71.
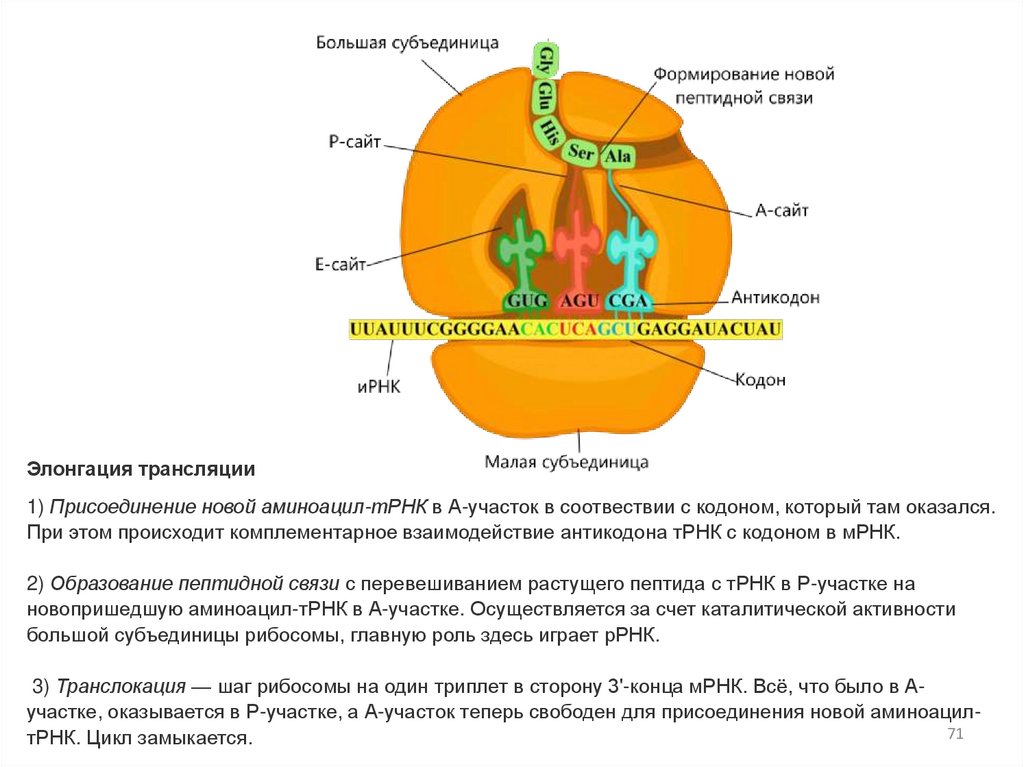
Элонгация трансляции1) Присоединение новой аминоацил-тРНК в А-участок в соотвествии с кодоном, который там оказался.
При этом происходит комплементарное взаимодействие антикодона тРНК с кодоном в мРНК.
2) Образование пептидной связи с перевешиванием растущего пептида с тРНК в Р-участке на
новопришедшую аминоацил-тРНК в А-участке. Осуществляется за счет каталитической активности
большой субъединицы рибосомы, главную роль здесь играет рРНК.
3) Транслокация — шаг рибосомы на один триплет в сторону 3'-конца мРНК. Всё, что было в Аучастке, оказывается в Р-участке, а А-участок теперь свободен для присоединения новой аминоацил71
тРНК. Цикл замыкается.
72.
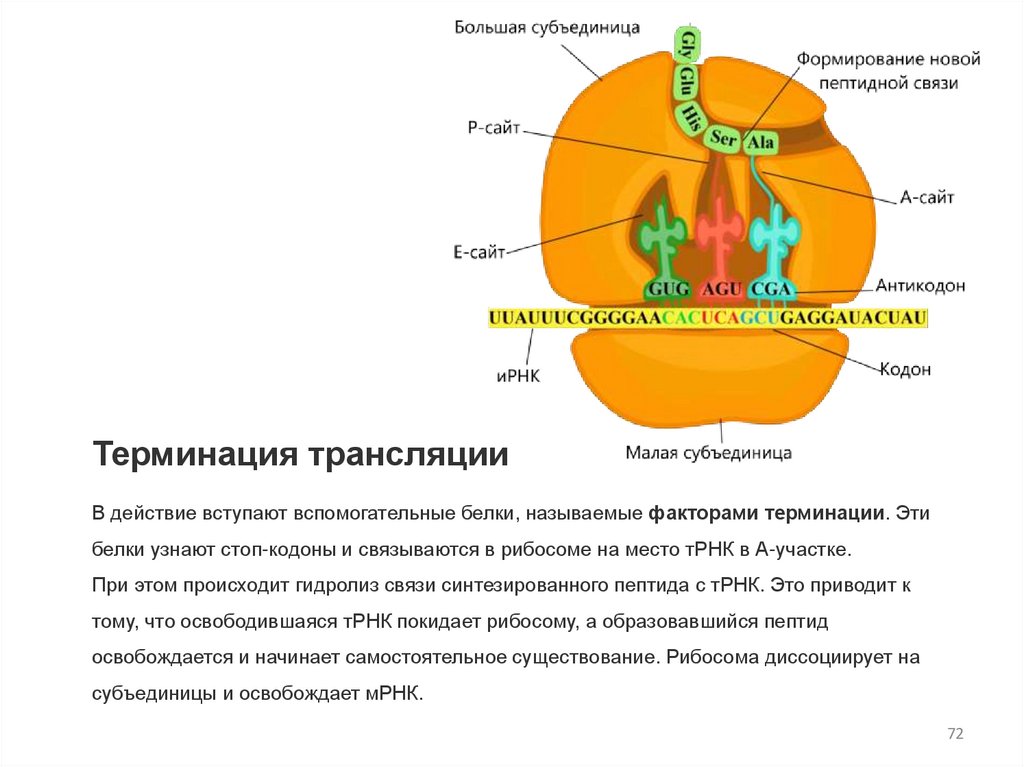
Терминация трансляцииВ действие вступают вспомогательные белки, называемые факторами терминации. Эти
белки узнают стоп-кодоны и связываются в рибосоме на место тРНК в А-участке.
При этом происходит гидролиз связи синтезированного пептида с тРНК. Это приводит к
тому, что освободившаяся тРНК покидает рибосому, а образовавшийся пептид
освобождается и начинает самостоятельное существование. Рибосома диссоциирует на
субъединицы и освобождает мРНК.
72
73. Генетический код
• Способ записи информации о первичнойструктуре белков через последовательность
нуклеотидов ДНК и РНК.
• «Словарь» перевода с языка нуклеиновых
кислот на язык белков.
• Полностью расшифрован к 1966
74. Георгий Антонович Гамов (1904-1968)
История открытия генетического кодаГеоргий Антонович Гамов
(1904-1968)
Физик-теоретик
1954
Сформулировал
проблему кода и
предположил его
триплетность.
75. Проблема
Алфавит белков20 а.к.
Алфавит ДНК
и РНК
4 нуклеотида
76. Обоснование триплетности кода Гамовым
н. а.к.Сколько а.к. можно
закодировать
Моноплетный
1→1
4
Дуплетный
2→1
16
Триплетный
3→1
64
77. Свойства генетического кода
• 1. Триплетность - каждая аминокислота кодируетсяпоследовательностью из 3-х нуклеотидов.
Определение: триплет или кодон - последовательность из трех
нуклеотидов, кодирующая одну аминокислоту.
• 2. Неперекрываемость - нуклеотид, входящий в состав кодона не
входит в состав другого кодона
• 3. Коллинеарность- порядок расположения аминокислот в
полипептидной цепи соответствует порядку расположения
кодонов в полинуклеотидной цепи
• 4. Вырожденность - все аминокислоты, за исключением
метионина и триптофана, кодируются более чем одним
триплетом:
• 5. Униполярность 5→3
• 6. Непрерывность - считывание информации происходит
непрерывно, без знаков препинания.
• 7. Специфичность - один кодон кодирует одну аминокислоту.
Например, кодон АУГ кодирует метионин.
• 8. Универсальность - Генетический код един для всех живущих на
Земле существ.
78.
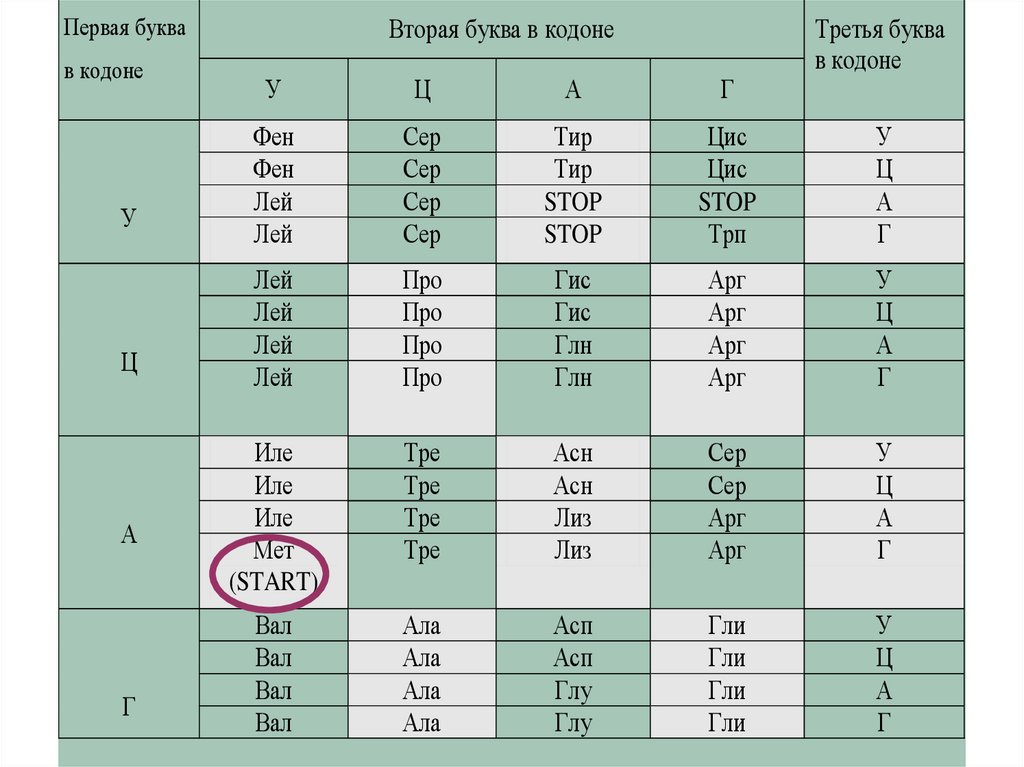
Первая буквав кодоне
У
Ц
А
Г
Вторая буква в кодоне
Третья буква
в кодоне
У
Ц
А
Г
Фен
Фен
Лей
Лей
Сер
Сер
Сер
Сер
Тир
Тир
STOP
STOP
Цис
Цис
STOP
Трп
У
Ц
А
Г
Лей
Лей
Лей
Лей
Про
Про
Про
Про
Гис
Гис
Глн
Глн
Арг
Арг
Арг
Арг
У
Ц
А
Г
Иле
Иле
Иле
Мет
(START)
Тре
Тре
Тре
Тре
Асн
Асн
Лиз
Лиз
Сер
Сер
Арг
Арг
У
Ц
А
Г
Вал
Вал
Вал
Вал
Ала
Ала
Ала
Ала
Асп
Асп
Глу
Глу
Гли
Гли
Гли
Гли
У
Ц
А
Г
79.
Спасибо завнимание!
79