





















































 biology
biologySimilar presentations:







")


Физиология, питание, размножение микроорганизмов. Лекция 2
1.
Лекция 2.Физиология, питание,
размножение
микроорганизмов
ТулГУ, Медицинский институт,
кафедра
«Санитарно-гигиенических
и
профилактических дисциплин»
Д.б.н., профессор Честнова Татьяна Викторовна
2.
МЕТАБОЛИЗМ БАКТЕРИАЛЬНОЙ КЛЕТКИВ процессе метаболизма выделяют два вида обмена:
1)пластический (конструктивный): анаболизм (с
затратами энергии), катаболизм (с выделением
энергии);
2)энергетический обмен (протекает в дыхательных
мезосомах): дыхание; брожение.
3.
Конструктивныйметаболизм
4.
ОСНОВНЫЕ ВИДЫ ПЛАСТИЧЕСКОГО ОБМЕНАБелковый
Углеводный
Липидный
Нуклеиновый
Белковый
обмен
характеризуется
катаболизмом
и
анаболизмом.
В
процессе
катаболизма
бактерии разлагают
белки под действием
протеаз
с
образованием
пептидов.
Под
действием пептидаз
из
пептидов
образуются
аминокислоты.
Углеводы представлены в
бактериальной клетке в
виде
моно-,
дии
полисахаридов,
а также
комплексных
соединений.
Полисахариды
входят
в
состав некоторых капсул,
крахмал
и
гликоген
являются
запасными
питательными веществами.
Жиры или липиды являются важными
компонентами ЦПМ и клеточной стенки
грамотрицательных бактерий, а также
служат
запасными
веществами.
В
бактериальных
жирах
преобладают
длинно-цепочечные насыщенные жирные
кислоты и не-насыщенные жирные
кислоты, содержащие одну двойную связь.
Сложные липиды представлены фосфатидилинозитом,фосфатидилглицерином
и
фосфатидилэтаноламином.Ключевым
промежуточным
продуктом
для
биосинтеза жирных кислот является
ацетилкоэнзим
А.
Ключевыми
промежуточными продуктами для синтеза
фосфолипидов является продукт ФДФпути:
диоксиацетилфосфат,
восстанавливающийся
в
глицерол-3фосфат, который соединяется с остатками
жирных кислот.
Пуриновые
и
пиримидиновые
нуклеотиды — это те строительные
блоки, из которых синтезируются
нуклеиновые кислоты. Кроме того,
Пуриновые
и
пиримидиновые
нуклеотиды входят в состав многих
коферментов
и
служат
для
активации и переноса аминокислот,
сахаров,
липидов
в
реакциях
полимеризации.
Синтез глюкозы происходит
из
пирувата,
за
счет
обратных реакций, путей
распада
глюкозы.
Для
обхода реакций, идущих
только
в
одном
направлении,
имеются
обходные пути, например
глиоксилатный цикл.
Исходным
соединением
для
образования
пентозной
части
нуклеотидов
служит
рибозо-5фосфат, образующийся в ПФ-пути.
Углеродный скелет пиримидинов
происходит из аспартата, который
образуется в цикле трикарбоновых
кислот. Атомы азота и аминогруппы
пуринов
и
аминосодержащих
пиримидинов
происходят
из
аспартата и глутамина.
5.
Распад белков в аэробных условиях –анаэробных – гниение.
тление, в
В результате распада аминокислот клетка получает ионы аммония,
необходимые для формирования собственных аминокислот.
процессы синтеза преобладают над распадом
6.
Углеродныескелеты
аминокислот
образуются
из
промежуточных продуктов обмена. Исходным материалом
служат промежуточные продукты фруктозодифосфатного (ФДФ)
и пентозофосфатного (ПФ) путей, цикл трикарбоновых кислот:
пируват, кетоглутаровая кислота, оксалоацетат, фумарат,
эритрозо-4-фосфат, рибозо-4-фосфат.
Аминогруппы вводятся в результате непосредственного
аминирования или переаминирования.
Перевод неорганического азота в органические соединения
происходит всегда через аммиак.
Нитраты и нитриты и молекулярный азот предварительно
восстанавливаются в аммиак и только лишь после этого
включаются в состав органических соединений.
7.
УГЛЕВОДНЫЙ ОБМЕНВ углеводном обмене у бактерий катаболизм преобладает над
анаболизмом.
Сложные углеводы внешней среды могут расщеплять только те
бактерии, которые выделяют ферменты – полисахаридазы.
Полисахариды расщепляются до дисахаров, которые под действием
олигосахаридаз распадаются до моносахаров, причем внутрь клетки
может поступать только глюкоза.
Часть ее идет на синтез собственных полисахаридов в клетке,
другая часть подвергается дальнейшему расщеплению, который
может идти по двум путям: по пути анаэробного распада углеводовброжению (гликолизу) и в аэробных условиях – по пути горения.
8.
В зависимости от конечных продуктов выделяютследующие виды брожения:
1) спиртовое (характерно для грибов);
2) пропионионово-кислое
(характерно
для
клостридий, пропионобактерий);
3) молочнокислое
(характерно
для
стрептококков);
4) маслянокислое (характерно для сарцин);
5) бутиленгликолевое (характерно для бацилл).
9.
ЛИПИДНЫЙ ОБМЕНОсуществляется с помощью ферментов –
липопротеиназ, летициназ, липаз, фосфолипаз.
Липазы катализируют распад нейтральных
жирных кислот, т.е. ответственны за отщепление
этих кислот от глицерина. При распаде жирных
кислот клетка запасает энергию. Конечным
продуктом распада является ацетил-КоА фермент.
10.
Биосинтез липидов осуществляется за счетацетилпереносящих белков.
При этом ацетильный остаток переходит на
глицерофосфат с образованием фосфатидных кислот, а
они уже вступают в химические реакции с образованием
сложных эфиров со спиртами.
Эти превращения лежат в основе синтеза
фосфолипидов.
Бактерии способны синтезировать как насыщенные,
так и ненасыщенные жирные кислоты, но синтез
последних более характерен для аэробов, так как
требует наличие кислорода.
11.
НУКЛЕИНОВЫЙ ОБМЕН БАКТЕРИЙСинтез осуществляется с помощью ферментов: рестриктазы,
лигазы, ДНК-полимеразы, ДНК-зависимой-РНК-полимеразы.
Рестриктазы
вырезают
участки
ДНК,
убирая
нежелательные вставки.
Лигазы обеспечивают сшивку фрагментов нуклеиновой
кислоты.
ДНК-полимеразы ответственны за репликацию дочерней
ДНК по материнской.
ДНК-зависимые-РНК-полимеразы
отвечают
за
транскрипцию, осуществляют построение РНК на матрице
ДНК.
12.
Энергетическийобмен
13.
Энергия в бактериальной клетке накапливается вформе молекул АТФ.
У хемоорганотрофных бактерий реакции, связанные с
получением энергии в форме АТФ, — это реакции
окисления-восстановления, сопряженные с реакциями
фосфорилирования.
Энергия аккумулируется
в молекулах АТФ.
14.
При использовании в качестве источника углерода иэнергии глюкозы или других гексоз начальные этапы
окисления глюкозы являются общими, как при
оксидативном, так и при бродильном метаболизмах.
К ним относятся пути превращения глюкозы в пируват
(при использовании в качестве источника энергии
отличных от глюкозы гексоз, или дисахаридов, они в
результате химических превращений вступают в цепь
реакций, превращающих глюкозу в пируват).
15.
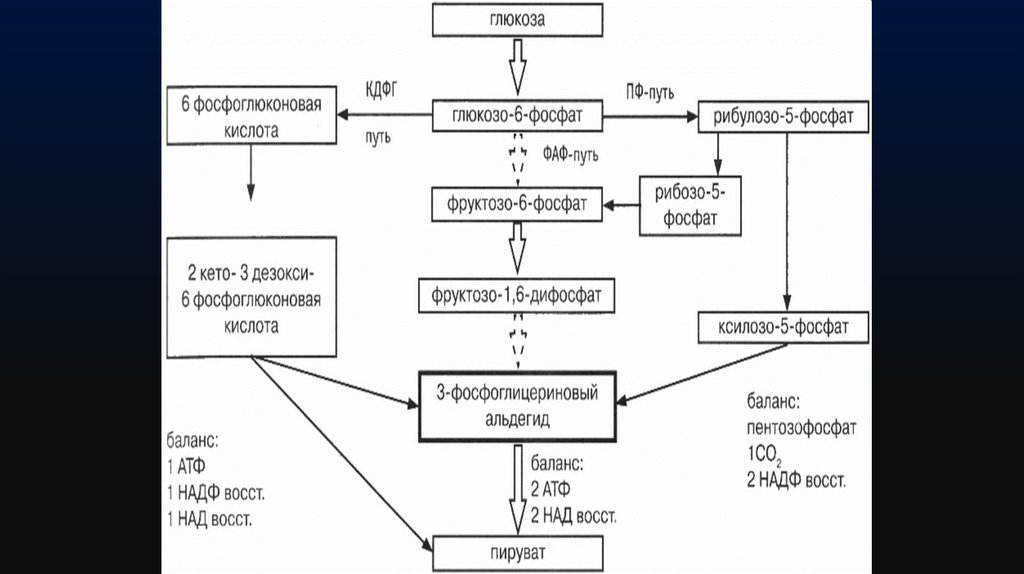
16.
Расщепление глюкозы до пировиноградной кислоты, одномуиз важнейших промежуточных продуктов обмена веществ, у
бактерий происходит 3 путями
1) через образование фруктозо-1,6-дифосфата (ФДФ-путем,
или гликолитическим распадом, или, по имени изучавших
его исследователей, путем Эмбдена—Мейергофа—
Парнаса);
2) через пентозофосфатный путь (ПФ-путь);
3) через путь Энтнера—Дудорова, или КДФГ-путь (путь 2кето-3-дезокси-6-фосфоглюконовая кислоты).
17.
Глюкозав
бактериальной
клетке
сначала
фосфорилируется при участии АТФ и фермента
гексокиназы до метаболически активной формы
глюкозо-6-фосфата (Г-6-Ф), которая служит исходным
соединением для любого из трех указанных выше путей.
Глюкозо-6-фосфат: вид молекулы
18.
1. ФДФ-путь.Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под
действием фосфофруктокиназы превращается во фруктозо1,6-дифосфат, который в дальнейшем через образование Зфосфоглицеринового альдегида окисляется до
пировиноградной кислоты.
Баланс окисления глюкозы по ФДФ-пути слагается из
образования 2 молекул пирувата, 2 молекул АТФ и 2 молекул
восстановленного НАД.
19.
2. ПФ-путь.В этом случае глюкозо-6-фосфат через реакции
дегидрирования
и
декарбоксилирования
превращается в рибулезо-5-фосфат (Ри-5-Ф), который
находится в равновесии с рибозо-5-фосфатом и
ксилулозо-5-фосфатом. Ри-5-Ф расщепляется до Зфосфоглицеринового альдегида, промежуточного
продукта превращения глюкозы в пируват.
20.
Образовавшиесяпентозофосфаты
превращаются
в
результате транскетолазных и трансальдолазных реакций
во фруктозо-6-фосфат, замыкая реакции в цикл, и в 3фосфоглицериновый альдегид, промежуточный продукт
превращения глюкозы в пируват по ФДФ-пути.
При одном обороте цикла образуется 1 молекула Зфосфоглицеринового альдегида, 3 молекулы С02 и 2
молекулы восстановленного НАДФ.
21.
3. КДФГ-путь (путь Этнера—Дудорова)Этот путь расщепления глюкозы специфичен только для
бактерий. Встречается у бактерий, потерявших фермент
фосфофруктокиназу, например у бактерий рода Pseudomonas.
Процесс начинается с дегидрирования глюкозо-6-фосфата до 6фосфоглюконовой
кислоты.
От
нее
под
действием
дегидрогеназы отщепляется вода и образуется 2-кето-3-дезокси6-фосфоглюконовая кислота (КДФГ), которая расщепляется
альдолазой на пируват и 3-фосфоглицериновый альдегид.
Последний окисляется до пировиноградной кислоты так же, как
и по ФДФ-пути.
22.
На каждую молекулу глюкозы образуется:• молекула АТФ,
• 1 молекула восстановленного НАД
• 1 молекула восстановленного НАДФ, которая
эквивалентна 1 молекуле АТФ и 1 молекуле
восстановленного НАД.
бактерии рода Pseudomonas
23.
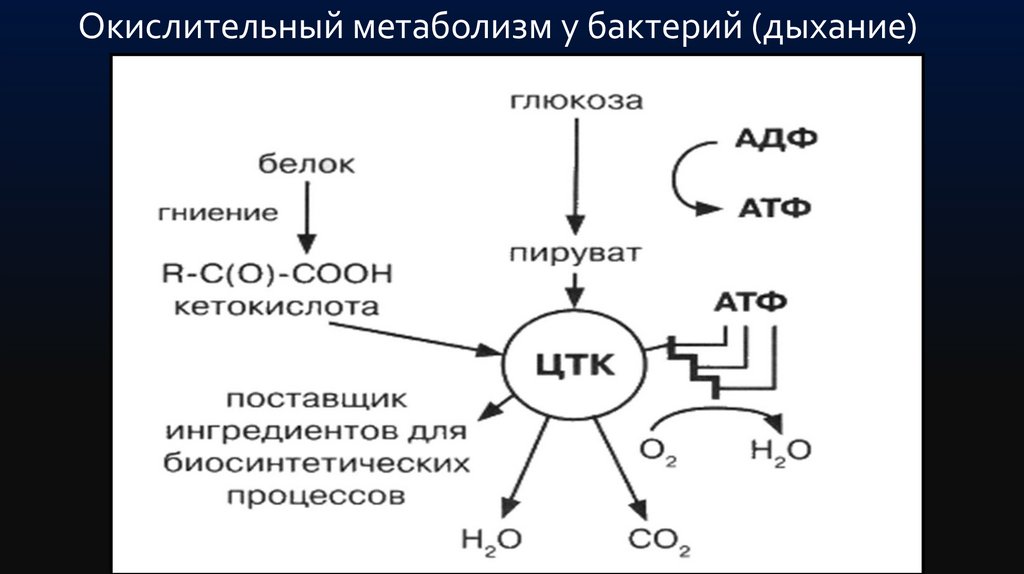
Окислительный метаболизм у бактерий (дыхание)24.
Бактерии, обладающие окислительным метаболизмом,энергию получают путем дыхания.
Дыхание — процесс получения энергии в реакциях
окисления-восстановления, сопряженных с реакциями
окислительного фосфорилирования, при котором
донорами электронов могут быть органические (у
органотрофов) и неорганические (у литотрофов)
соединения, а акцептором — только неорганические
соединения.
25.
В зависимости от акцепторов протонов и электронов среди бактерийразличают аэробы, факультативные анаэробы и облигатные
анаэробы.
Для аэробов акцептором является кислород.
Факультативные анаэробы в кислородных условиях используют
процесс дыхания, в бескислородных – брожение.
Для облигатных анаэробов характерно только брожение, в
кислородных условиях наступает гибель микроорганизмов из-за
образования перекисей, идет отравление клетки.
26.
Облигатные аэробы (бруцеллы, легионеллы, псевдомонады,микобактерии, возбудитель сибирской язвы) растут и
размножаются только в присутствии кислорода. Используют
кислород для получения энергии путем кислородного дыхания.
Они подразделяются на:
1) строгие аэробы (менингококки, бордетеллы), которые растут
при парциальном давлении атмосферы воздуха;
2) микроаэрофилы (листерии) растут при
парциальном давлении атмосферного воздуха.
пониженном
27.
Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии)не используют кислород для получения энергии. Тип метаболизма у них
бродильный. Они подразделяются на:
1) строгие анаэробы – микроорганизмы для которых молекулярный
кислород токсичен; он либо убивает микроорганизмы, либо
ограничивает их рост. Энергию строгие анаэробы получают
маслянокислым брожением;
2) аэротолерантные микроорганизмы (молочнокислые бактерии)
используют кислород для получения энергии, но могут существовать в
его
атмосфере.
Энергию
получают
гетероферментативным
молочнокислым брожением
28.
Факультативные анаэробы (пневмококки, энтерококки,энтеробактерии, коринебактерии, франциселлы) способны
расти и размножаться как в присутствии кислорода, так и в
отсутствии его. Они обладают смешанным типом
метаболизма.
Процесс получения энергии у них может происходить
кислородным дыханием в присутствии кислорода, а в его
отсутствии переключаться на брожение. Различное
физиологическое
отношение
микроорганизмов
к
кислороду связано с наличием у них ферментных систем,
позволяющих существовать в атмосфере кислорода.
29.
В окислительных процессах, протекающих в атмосферекислорода образуются токсические продукты:
• перекись водорода Н2О2
• закисный радикал кислорода О2Для
нейтрализации
токсичных
форм
кислорода,
микроорганизмы, способные существовать в его атмосфере,
имеют защитные механизмы:
1. У облигатных аэробов и факультативных анаэробов
накоплению закисного радикала кислорода препятствует
фермент супероксиддисмутаза.
30.
2. У аэротолерантных микроорганизмов накоплению закисного радикалакислорода препятствует высокая концентрация ионов марганца, перекись
водорода разрушается ферментом пероксидазой.
3. У строгих анаэробов наличие фермента супероксиддисмутазы
коррелирует с их устойчивостью к кислороду. Для культивирования
строгих анаэробов создаются условия, позволяющие удалять
атмосферный кислород: использование анаэростатов, добавление в
питательные среды редуцирующих кислород веществ, например
тиогликолята натрия, использование поглотителей кислорода.
У
бактерий,
обладающих
окислительным
метаболизмом,
акцептором электронов (или водорода (Н+)) является молекулярный
кислород.
В этом случае пируват полностью окисляется в цикле трикарбоновых
кислот до С2.
31.
32.
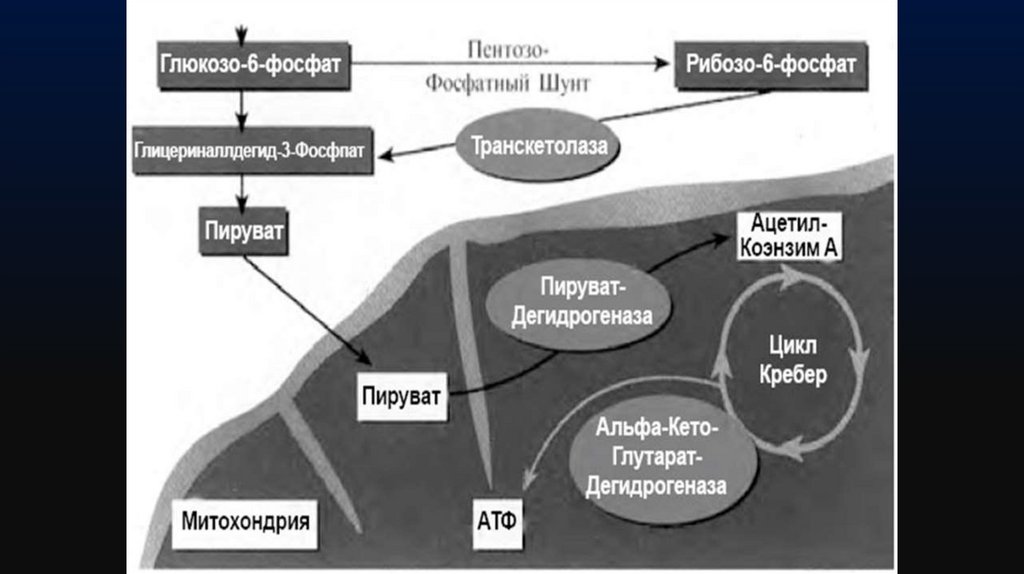
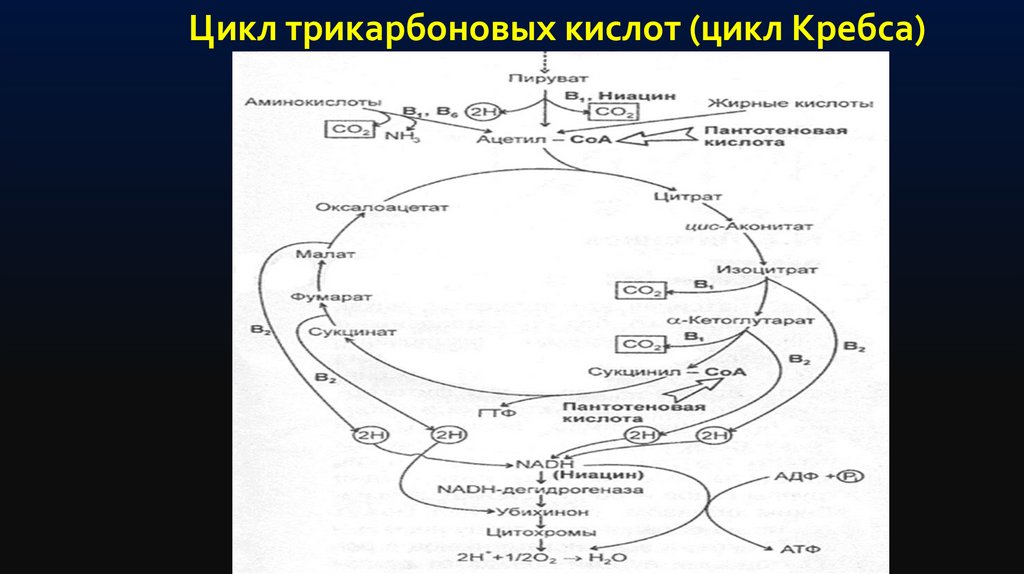
Цикл трикарбоновых кислот (цикл Кребса)33.
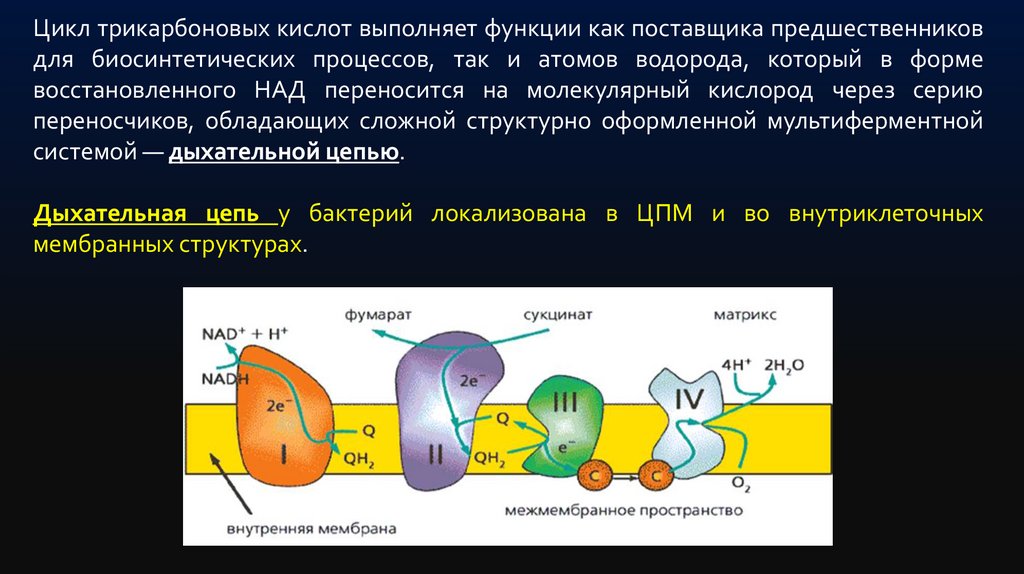
Цикл трикарбоновых кислот выполняет функции как поставщика предшественниковдля биосинтетических процессов, так и атомов водорода, который в форме
восстановленного НАД переносится на молекулярный кислород через серию
переносчиков, обладающих сложной структурно оформленной мультиферментной
системой — дыхательной цепью.
Дыхательная цепь у бактерий локализована в ЦПМ и во внутриклеточных
мембранных структурах.
34.
Типичная цепь выглядит следующим образом:ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2
Среди бактериальных цитохромов различают цитохромы в, с, а и а3.
Конечным этапом переноса электронов (протонов) по дыхательной
цепи является восстановление цитохромов а - а3 (цитохромоксидазы).
Цитохромоксидаза является конечной оксидазой, передающей
электроны на кислород. Образующиеся при окислении ФАД или
хинонов протоны связываются ионами О2- с образованием воды.
35.
Среди бактериальных цитохромов различают цитохромы в, с, а иа3. Конечным этапом переноса электронов (протонов) по
дыхательной цепи является восстановление цитохромов а - а3
(цитохромоксидазы).
Цитохромоксидаза является конечной оксидазой, передающей
электроны на кислород. Образующиеся при окислении ФАД или
хинонов протоны связываются ионами О2- с образованием воды.
цитохром С
36.
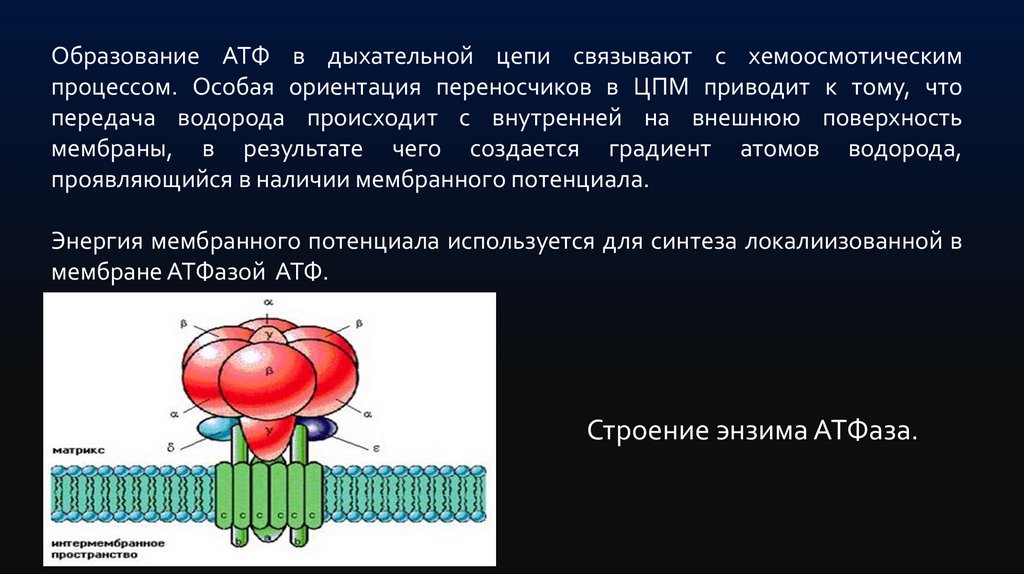
Образование АТФ в дыхательной цепи связывают с хемоосмотическимпроцессом. Особая ориентация переносчиков в ЦПМ приводит к тому, что
передача водорода происходит с внутренней на внешнюю поверхность
мембраны, в результате чего создается градиент атомов водорода,
проявляющийся в наличии мембранного потенциала.
Энергия мембранного потенциала используется для синтеза локалиизованной в
мембране АТФазой АТФ.
Строение энзима АТФаза.
37.
У некоторых бактерий цитохромы отсутствуют, и приконтакте с кислородом происходит непосредственный
перенос водорода на кислород с помощью
флавопротеидов, конечным продуктом при этом
оказывается перекись водорода — Н2О2.
38.
39.
ФЕРМЕНТЫ МИКРООРГАНИЗМОВВ микробной клетке ферменты являются
биологическими катализаторами.
Набор ферментов в клетке строго индивидуален
для вида.
Способность микроорганизма утилизировать
субстраты за счет своего набора ферментов
определяет его биохимические свойства.
40.
ПО МЕСТУ ДЕЙСТВИЯ ВЫДЕЛЯЮТ:1) экзоферменты - действуют вне клетки,
принимают участие в процессе распада крупных
молекул, которые не могут проникнуть внутрь
бактериальной
клетки;
характерны
для
грамположительных бактерий);
2) эндоферменты - действуют в самой клетке,
обеспечивают синтез и распад различных веществ)
41.
В зависимости от катализируемых химическихреакций все ферменты делят на шесть классов:
№ п/п
Класс ферментов
Функции
1.
Оксидоредуктазы
Катализируют окислительно-восстановительные реакции между
двумя субстратами
2.
Трансферазы
Осуществляют межмолекулярный перенос химических групп
3.
Гидролазы
Осуществляют гидролитическое расщепление
внутримолекулярных связей
4.
Лиазы
Присоединяют химические группы по двум связям, А также
осуществляют обратные реакции
5.
Изомеразы
Осуществляют процессы изомеризации, обеспечивают
внутреннюю конверсию С образованием различных изомеров
6.
Лигазы (синтетазы) Соединяют две молекулы, вследствие чего происходит
расщепление пирофосфатных связей в молекуле АТФ
42.
Питание бактерий43.
Под питанием понимают процессы поступления и выведенияпитательных веществ в клетку и из клетки. Питание в первую
очередь обеспечивает размножение и метаболизм клетки.
Среди необходимых питательных веществ выделяют:
углерод, кислород, водород, азот, фосфор, калий, магний,
кальций.
Кроме органогенов, необходимы микроэлементы. Они
обеспечивают активность ферментов. Это цинк, марганец,
молибден, кобальт, медь, никель, вольфрам, натрий, хлор.
44.
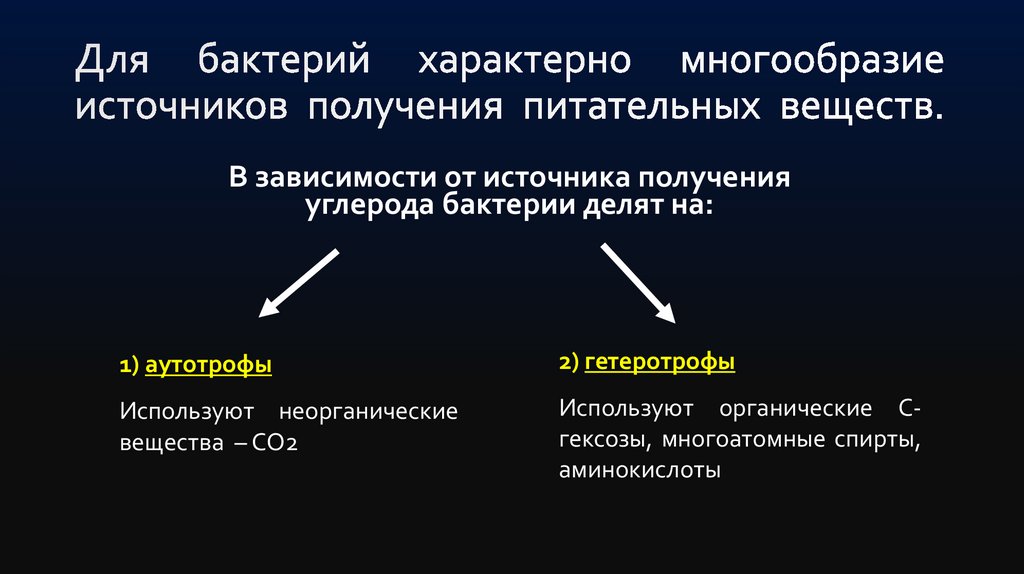
В зависимости от источника полученияуглерода бактерии делят на:
1) аутотрофы
2) гетеротрофы
Используют неорганические
вещества – СО2
Используют органические Сгексозы, многоатомные спирты,
аминокислоты
45.
Процессы питания должны обеспечивать энергетическиепотребности бактериальной клетки.
По источникам энергии микроорганизмы делят на:
1) фототрофы – источник солнечная энергия;
2) хемотрофы – получают энергию за счет окислительновосстановительных реакций;
3) хемолитотрофы – используют неорганические соединения;
4) хемоорганотрофы – используют органические вещества.
Медицинская микробиология
гетерохемоорганотрофами
46.
По степени гетеротрофности микроорганизмы делятся на:1) сапрофиты – питаются мертвым органическим материалом;
2) паразиты – питаются за счет макроорганизма.
Облигатные
47.
Факторами роста бактерий являются витамины,аминокислоты, пуриновые и пиримидиновые основания,
присутствие которых ускоряет рост. Среди бактерий
выделяют:
1)
прототрофы
(способны
сами
синтезировать
необходимые вещества);
2) ауксотрофы
(нуждаются
Микроорганизмы ассимилируют питательные вещества в
виде небольших молекул, поэтому белки, полисахариды и
другие биополимеры могут служить источниками питания
только после расщепления их экзоферментами до более
простых соединений.
48.
Транспорт веществДля того, чтобы питательные вещества могли подвергнуться
превращениям в цитоплазме клетки, они должны проникнуть в
клетку через пограничные слои, отделяющие клетку от
окружающей среды.
Ответственность за поступление в клетку питательных веществ
лежит на ЦПМ.
49.
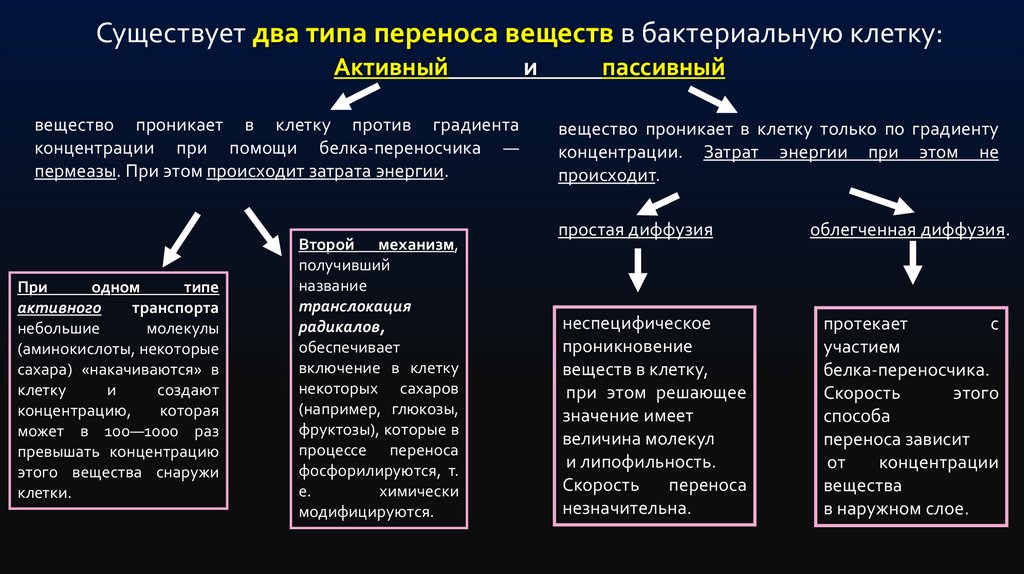
Существует два типа переноса веществ в бактериальную клетку:Активный
вещество проникает в клетку против градиента
концентрации при помощи белка-переносчика —
пермеазы. При этом происходит затрата энергии.
При
одном
типе
активного
транспорта
небольшие
молекулы
(аминокислоты, некоторые
сахара) «накачиваются» в
клетку
и
создают
концентрацию,
которая
может в 100—1000 раз
превышать концентрацию
этого вещества снаружи
клетки.
Второй механизм,
получивший
название
транслокация
радикалов,
обеспечивает
включение в клетку
некоторых сахаров
(например, глюкозы,
фруктозы), которые в
процессе переноса
фосфорилируются, т.
е.
химически
модифицируются.
и
пассивный
вещество проникает в клетку только по градиенту
концентрации. Затрат энергии при этом не
происходит.
простая диффузия
облегченная диффузия.
неспецифическое
проникновение
веществ в клетку,
при этом решающее
значение имеет
величина молекул
и липофильность.
Скорость
переноса
незначительна.
протекает
с
участием
белка-переносчика.
Скорость
этого
способа
переноса зависит
от
концентрации
вещества
в наружном слое.
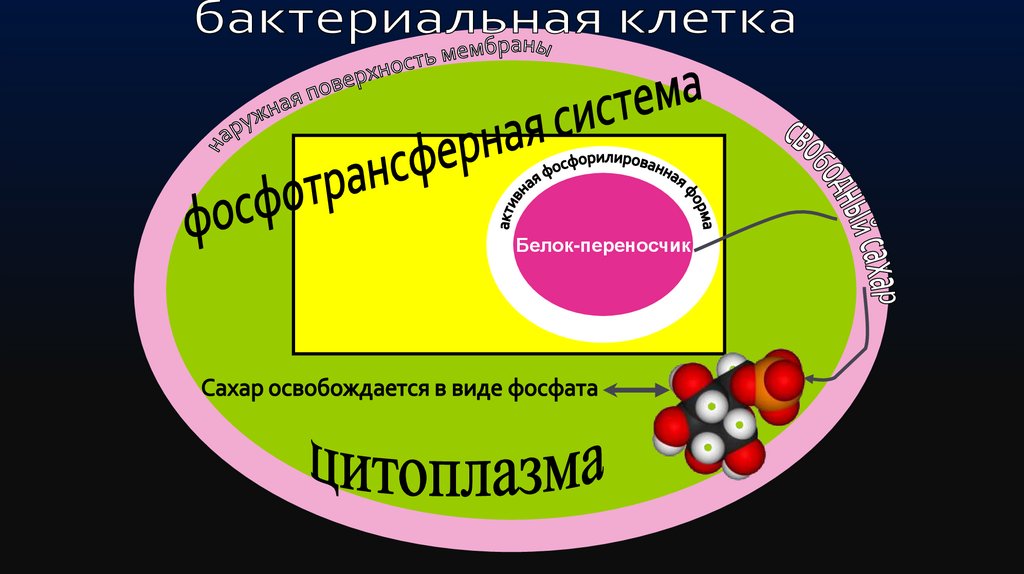
50.
Для осуществления этих процессов в бактериальнойклетке локализуется специальная фосфотрансферная
система, составной частью которой является белокпереносчик,
находящийся
в
активной
фосфорилированной форме.
Фосфорилированный белок связывает свободный
сахар на наружной поверхности мембраны и
транспортирует его в цитоплазму, где сахар
освобождается в виде фосфата.
51.
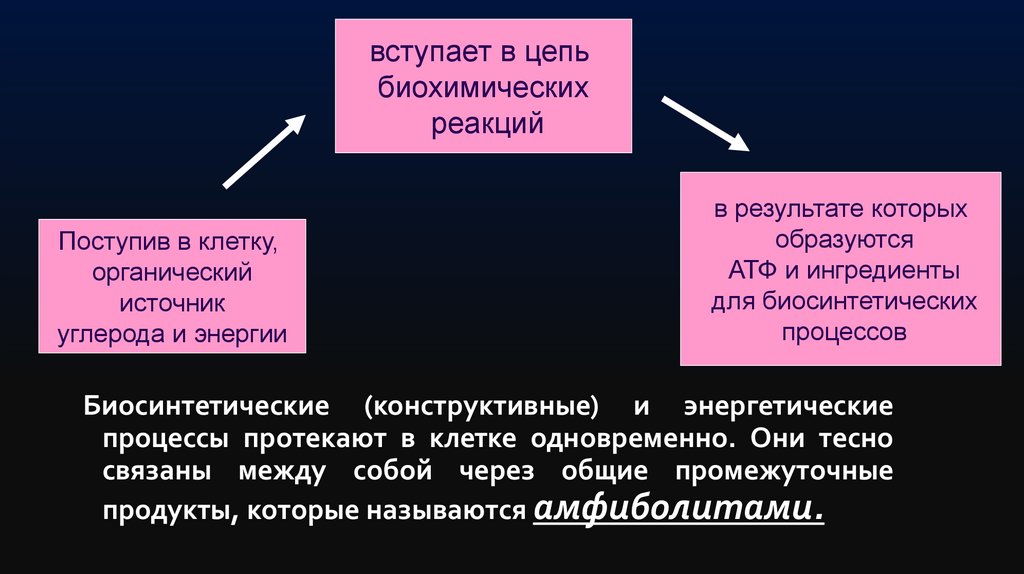
Белок-переносчик52.
вступает в цепьбиохимических
реакций
Поступив в клетку,
органический
источник
углерода и энергии
в результате которых
образуются
АТФ и ингредиенты
для биосинтетических
процессов
Биосинтетические (конструктивные) и энергетические
процессы протекают в клетке одновременно. Они тесно
связаны между собой через общие промежуточные
продукты, которые называются амфиболитами.
53.
ФАЗЫ РАЗМНОЖЕНИЯ БАКТЕРИЙ1. Начальная (лаг-фаза) охватывает промежуток времени от момента посева бактерий
до начала их роста. Ее продолжительность составляет в среднем 2-5 часов и зависит от
состава питательной среды.
2. Экспоненциальная (логарифмическая) фаза. Характеризуется постоянной
максимальной скоростью деления клеток. Это время варьирует от нескольких минут до
нескольких часов.
3. Стационарная фаза наступает когда число клеток перестает увеличиваться. При
уменьшении в питательной среде концентрации питательных веществ, снижении
парциального давления кислорода, накоплении токсических продуктов обмена,
уменьшается скорость роста бактерий. Продолжительность стационарной фазы
составляет несколько часов и зависит от вида бактерий.
4. Фаза отмирания наступает вследствие накопления кислых продуктов обмена или в
результате автолиза под влиянием собственных ферментов. Продолжительность этой
фазы колеблется от десятка часов до нескольких недель.
54.
ВИРУСЫ.ТИПЫ ВЗАИМОДЕЙСТВИЯ ВИРУСА С
КЛЕТКОЙ.
СТАДИИ РЕПРОДУКЦИИ ВИРУСОВ.
55.
ТИПЫ ВЗАИМОДЕЙСТВИЯ ВИРУСА С КЛЕТКОЙРазличают три типа взаимодействия вируса с клеткой:
1. Продуктивный тип - завершается образованием нового поколения
вирионов и гибелью зараженных клеток (цитолитическая форма).
2. Абортивный тип- не завершается образованием новых вирионов,
поскольку инфекционный процесс в клетке прерывается на одном из
этапов.
3. Интегративный тип, или вирогения— характеризуется встраиванием
(интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их
совместным сосуществованием (совместная репликация).
56.
СТАДИИ РЕПРОДУКЦИИ ВИРУСОВРепродукция вирусов осуществляется в несколько
стадий:
• адсорбция вируса на клетке;
• проникновение вируса в клетку;
• «раздевание» вируса;
• биосинтез вирусных компонентов в клетке;
• формирование вирусов;
• выход вирусов из клетки.
57.
СТАДИИ РЕПРОДУКЦИИ ВИРУСОВ• Адсорбция. Вирус адсорбируется на определенных участках клеточной
мембраны - так называемых рецепторах. Число специфических рецепторов на поверхности одной клетки колеблется от 104 до 105.
Следовательно, на клетке могут адсорбироваться десятки и даже сотни
вирусных частиц.
• Проникновение в клетку. Существует два способа проникновения
вирусов животных в клетку: виропексис и слияние вирусной оболочки с
клеточной мембраной. При виропексисе после адсорбции вирусов
происходят инвагинация (впячивание) участка клеточной мембраны и
образование внутриклеточной вакуоли, которая содержит вирусную
частицу. Процесс слияния осуществляется одним из поверхностных
вирусных белков капсидной или суперкапсидной оболочки с клеточной
мембраной.
58.
СТАДИИ РЕПРОДУКЦИИ ВИРУСОВ• «Раздевание». Процесс «раздевания» заключается в удалении
защитных вирусных оболочек и освобождении нуклеиновой
кислоты.
• Биосинтез компонентов вируса. Проникшая в клетку
вирусная нуклеиновая кислота дезорганизует работу
клеточных систем и заставляет клетку синтезировать новые
вирусные белки и нуклеиновые кислоты.
• Формирование (сборка) вирусов. Вирусные нуклеиновые
кислоты и белки обладают способностью специфически
«узнавать» друг друга и самопроизвольно соединяться.
59.
СТАДИИ РЕПРОДУКЦИИ ВИРУСОВ• Выход вирусов из клетки. Различают два основных типа
выхода вирусного потомства из клетки.
Первый тип — взрывной — характеризуется одновременным
выходом большого количества вирусов. При этом клетка
быстро погибает. Такой способ выхода характерен для
вирусов, не имеющих суперкапсидной оболочки.
Второй тип — почкование. Он присущ вирусам, имеющим
суперкапсидную оболочку.
Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.).
60.
БАКТЕРИОФАГИ. ВЗАИМОДЕЙСТВИЕ ФАГАС БАКТЕРИАЛЬНОЙ КЛЕТКОЙ
Бактериофаги – вирусы бактерий. Бактериофаги широко распространены: они
выявлены у большинства бактерий, а также у других микроорганизмов, например, у
грибов и поэтому их часто называют фагами. Наиболее детально изучена структура
крупных фагов, к которым относятся фаги E. Coli.
Они состоят из головки икосаэдрического типа, в которой заключена или ДНК, или
РНК.
Большинство фагов содержат двунитевую ДНК, замкнутую в кольцо. Хвостовой
отросток имеет внутри полый цилиндрический стержень, сообщающийся с головкой, а
снаружи – чехол, способный к сокращению наподобие мышцы.
Чехол присоединен к воротничку, окружающему стержень около головки.
Хвостовой отросток заканчивается шестиугольной базальной пластинкой с шипами, от
которых отходят нитевидные структуры – фибриллы.
61.
По морфологии фаги подразделяются на 6 групп:1) фаги с длинным отростком, чехол которого
сокращается;
2) фаги с длинным отростком, чехол которого не
сокращается;
3) фаги с короткими отростками;
4) фаги с аналогом отростка;
5)фаги без отростка;
6) нитевидные фаги.
62.
По специфичности взаимодействия различаютследующие бактериофаги:
1)
поливалентные
–
взаимодействуют с
родственными видами бактерий;
2)
моновалентные
–
взаимодействуют
с
бактериями определенного вида;
3) типовые – взаимодействуют с отдельными
типами бактерий данного вида.
63.
ВЗАИМОДЕЙСТВИЕ ФАГОВ С БАКТЕРИЯМИМОЖЕТ ПРОТЕКАТЬ:
1) по продуктивному типу – образуется фаговое
потомство и бактерии лизируются;
2) 2) по абортивному типу – фаговое потомство не
образуется
и
бактерии
сохраняют
свою
жизнедеятельность;
3) 3) по интегративному типу – геном фага
встраивается в хромосому бактерии и сосуществует с
ней.
64.
В ЗАВИСИМОСТИ ОТ ТИПА ВЗАИМОДЕЙСТВИЯРАЗЛИЧАЮТ:
• Вирулентные бактериофаги - взаимодействуют с бактерией
по продуктивному типу.
• Умеренные
бактериофаги
взаимодействуют
с
чувствительными бактериями по интегративному типу. При
интегративном типе взаимодействия ДНК умеренного фага
встраивается в хромосому бактерии, реплицируется синхронно
с геном размножающейся бактерии, не вызывая ее лизиса.
ДНК бактериофага, встроенная в хромосому бактерии,
называется профагом, а культура бактерий – лизогенной. Такое
сосуществование бактерии и умеренного бактериофага
называется лизогенией.
65.
БАКТЕРИОФАГИ ИСПОЛЬЗУЮТ:1) в лабораторной диагностике инфекций при внутривидовой
идентификации бактерий, т.е. определения фаговара. Для этого
применяют метод фаготипирования;
2) фаги применяют также для лечения и профилактики ряда бактериальных
инфекций.
Производят
брюшнотифозный,
сальмонеллезный,
дизентерийный, синегнойный, стафилококковый, стрептококковый фаги;
комбинированные: колипротейный, пиобактериофаги.
Бактериофаги
назначают по показаниям перорально, парентерально или местно в виде
жидких, таблетированных форм, свечей, аэрозолей.
3) бактериофаги широко применяют в генной инженерии в качестве
векторов для получения рекомбинантных ДНК.