










 бактерий")



")
")




.")













")











 метаболизм")

























 питательные среды")
















 biology
biologySimilar presentations:










Физиология микроорганизмов
1. Физиология микроорганизмов
Зав.кафедройд.м.н., профессор
Г.И.Чубенко
2.
Микроорганизмам, как и другимбиологическим объектам присущи все
физиологические функции.
Они питаются, растут,
дышат, размножаются.
3.
Совокупность биохимических реакций умикроорганизмов подчиняется
принципу биохимического единства, т.е.
в биохимическом отношении все живые
существа на Земле сходны.
4. Химический состав бактерий.
Вода основной компонент бактериальной клетки,она находится в свободном и связанном
состоянии.
Гидролитические процессы расщепления белков,
углеводов и липидов происходят в результате
присоединения к ним воды.
При недостатке воды нарушается и размножение
бактерий.
Связанная вода определяет устойчивость к
физическим факторам
5. В микробной клетке содержатся
углерод (45%), азот (8-15%), кислород (30%), водород(6-8%) и минеральные вещества от 3 до 10 % (фосфор,
сера, магний, железо, кальций, калий, цинк, кобальт,
медь и др.), Вода.
6.
7. Минеральные вещества
стимулируют процессы роста и размножениябактерий,
определяют рН среды,
окислительно-восстановительный потенциал,
поддерживают осмотическое давление,
активность ферментативных процессов.
8.
В составе бактерий имеютсябелки,
углеводы,
липиды,
витамины.
9. Белки
составляют 50-80% сухого вещества микробной клетки. Всостав белков прокариот входят 20 аминокислот.
Белки входят в состав различных морфологических
структур,
являются составными частями ферментов, токсинов,
антигенов,
определяют отношение к красителям, лекарственным и
дезинфицирующим веществам.
Они могут быть простыми и сложными
Простые белки при гидролизе распадаются на аминокислоты
(лейцин, триптофан и др.).
Сложные белки (протеиды)- это соединения простых белков
с небелковыми группами: нуклеиновыми кислотами,
полисахаридами, липидами и др.
10. Нуклеиновые кислоты бактерий представлены РНК и ДНК
Содержание нуклеиновых кислот вмикробной клетке зависит
от вида бактерий,
состава питательной среды,
фазы развития микробной популяции.
11.
РНК содержится преимущественно вцитоплазме, обеспечивает транскрипцию и
трансляцию на рибосомах.
ДНК- преимущественно в нуклеоиде носитель генетической информации.
12. Углеводы (12-18%) бактерий
Представлены:многоатомными спиртами (сорбит, дульцит,
манит);
полисахаридами (гексозы, пентозы, гликоген,
декстрин);
моносахаридами (глюкоза, глюкуроновая кислота
и др. )
Углеводы выполняют главную энергетическую
функцию и определяют антигенную
специфичность микроорганизмов.
13. Липиды
Липиды (истинные жиры)встречаются у риккетсий,дрожжей, микобактерий, грибов и др.
Бактериальные липиды играют роль резервных веществ.
Представлены свободными жирными кислотами,
нейтральными жирами, восками, фосфолипидами.
Могут быть использованы как исходные компоненты для
синтеза белков.
Входят в состав основной токсической фракции многих
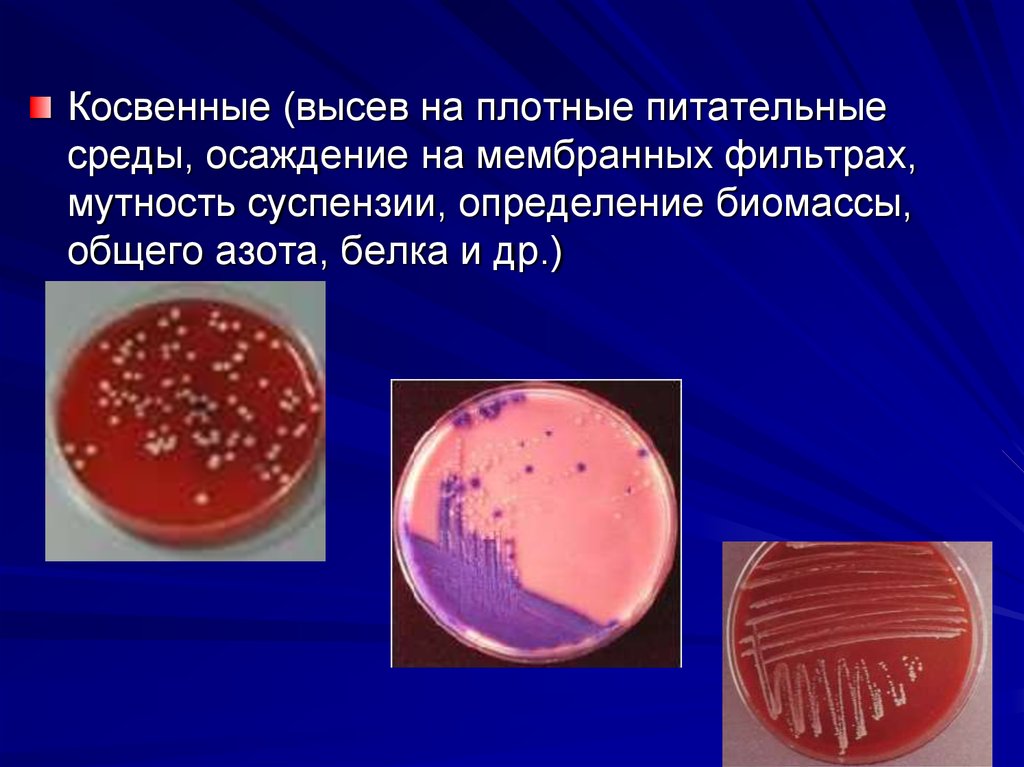
микроорганизмов.
Определяют проницаемость клеточных мембран, их
стабильность
устойчивость к кислотам и щелочам, заряд клетки.
14. Метаболизм
- совокупность ферментативных реакцийнаправленных на получение энергии и превращение
простых соединений в макромолекулы.
- совокупность двух противоположных процессов:
катаболизма (энергетического метаболизма)
анаболизма (пластического, конструктивного
метаболизма).
Питательные вещества,
поступающие в клетку, служат
источником энергии и
строительным материалом для
синтеза клеточных структур.
15. Этапы ферментативных реакций катаболизма и анаболизма:
Начальный (периферический метаболизм)- ферментыоказывают воздействие на исходный субстрат
Ферментативное образование промежуточных
продуктов (амфиболитов)
Образование конечных продуктов и выделение их в
среду
16. Конструктивный метаболизм (анаболизм)
Поток реакций, в результате которых за счетпоступающих извне веществ строится вещество
клетки, сопровождается потреблением свободной
энергии, запасенной в макроэргах.
В процессе анаболических реакций эта энергия
расходуется на синтез многочисленных макромолекул
органических соединений.
17. Питание (бактерий)
По способу питания бактерии относятся кголофитным объектам (нет специализированных
органов для принятия пищи).
Особенности питания бактерий:
• питательные вещества поступают через всю
поверхность микробной клетки в растворенном виде.
• высокая скорость процессов метаболизма
• пластичность к меняющимся условиям окружающей
среды.
18. Механизмы питания у бактерий
Пассивная диффузия (поградиенту концентрации)
Облегченная диффузия
(посредством
системы транслоказ и
пермеаз по градиенту
концентрации)
Активный транспорт (против
градиента концентрации с
помощью переносчиков и с
затратами энергии)
Транслокация (модификация
химического вещества) с
затратами энергии
19.
Пермеазы катализируют присоединение веществасубстрата к активному центру на своей поверхностии проводят это вещество с наружной поверхности
ЦПМ на внутреннюю.
Здесь пермеаза освобождается от вещества, а сама
вновь вступает во взаимодействие с новой порцией
субстрата.
Пермеазы составляют значительную часть белков
ЦПМ.
20. Выход веществ из микробной клетки
осуществляется:фосфотрансферазной реакцией, при фосфорилировании
переносимой молекулы;
котрансляционной секрецией,
21. Типы секреции: I тип секреции
Требует наличия 3 белков:• Транспортной аденозинфосфатсинтетазы ЦПМ;
• Белка ЦПМ, формирующего канал в периплазме;
• Белка-секретина, образующего канал в
пептидогликане.
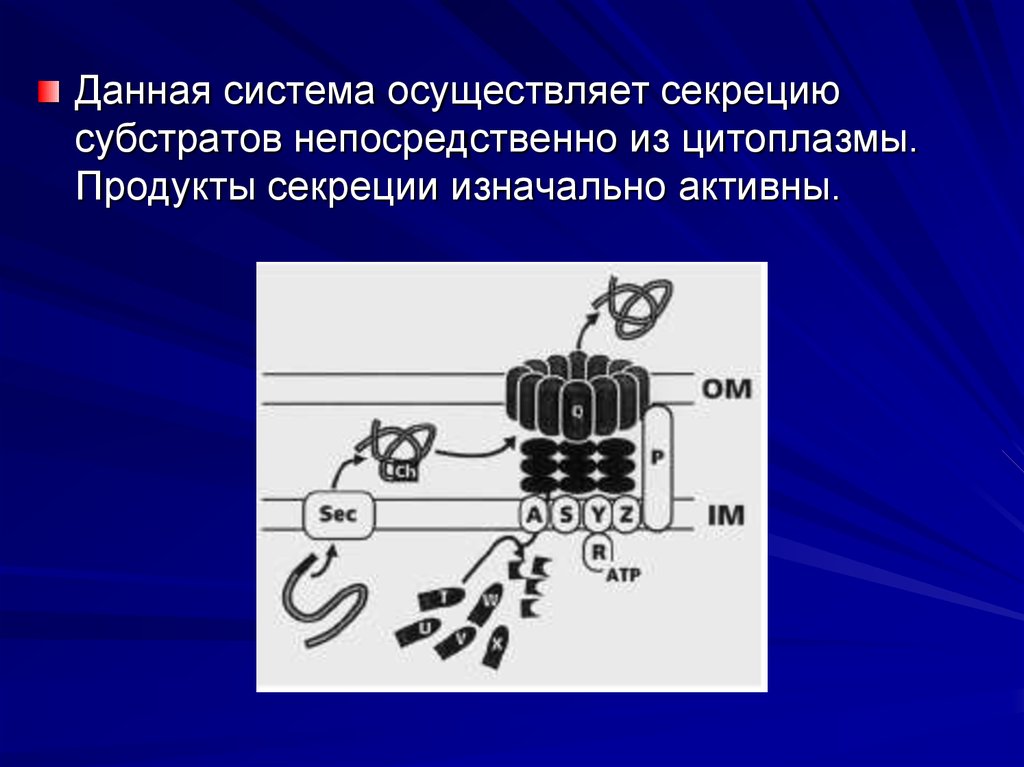
22.
Данная система осуществляет секрециюсубстратов непосредственно из цитоплазмы.
Продукты секреции изначально активны.
23. II тип секреции «общий секреторный путь» (GSP).
Секреторный аппарат формирует 12-14 белков, основнаячасть которых расположена в ЦПМ.
Секретируемые белки первоначально накапливаются в
периплазме, где формируют молекулы четвертичной
структуры, а затем происходит их удаление через
особый канал, образованный в клеточной стенке
белком секретином.
24.
Система секреции состоит из двух частей иосуществляется в две стадии.
Sec-система направляет предшественников к
транслокационному комплексу ЦПМ;
Через транслоказу пресекретируемый белок
высвобождается в периплазматичекую щель, где
принимает свою нативную конформацию и затем
секретируется через клеточную стенку.
Белковые молекулы
переходят через secаппарат в полностью
развернутом виде.
25. III тип секреции
Обеспечивает не только перенос секреторных продуктовчерез мембранные структуры бактериальной клетки, но
и доставку их внутрь эукариотических клеток.
Структурно представляет собой «молекулярный шприц»,
образованный двадцатью белками, преимущественно
расположенными в ЦПМ.
26. Система III типа
Транспортируемые молекулы находятся в интактнойформе и активируются только после попадания в
цитоплазму эукариотической клетки-мишени.
Кроме интегральных мембранных белков в состав
аппарата секреции входит и несколько
цитоплазматических белков, участвующих в доставке
секреторных продуктов к локусу секреции.
27. IV тип секреции
Обеспечивает выделение белков- аутотранспортёров.У каждой молекулы существует терминальный N-участок с
сигнальной последовательностью, обеспечивающей
транспорт через ЦПМ, и терминальный С- участок,
формирующий пору в клеточной стенке, через которую
проходит остальная часть молекулы (сериновые протеазы
энтеробактерий -SPATE) .
28. Типы питания
В зависимости от источников углеродамикроорганизмы делятся:
Автотрофы- синтезируют все компоненты из
неорганического углерода (СО2)
Гетеротрофы- источник углерода органические
соединения
29.
Олиготрофы- достаточно низкойконцентрации органических веществ.
Копиотрофы- нуждаются в высокой
концентрации органических веществ.
30.
Строгие (облигатные) паразиты- живут только внутриклетки-хозяина;
Факультативные паразиты- существуют как внутри,
так и вне клетки-хозяина;
Сапрофиты – используют органические остатки
(растений и животных);
31. В зависимости от источников азота
микроорганизмы делятся:Прототрофы- способны синтезировать
азотсодержащие соединения из солей аммония,
нитратов, нитритов и глюкозы.
Ауксотрофы- ассимилируют только готовые
азотсодержащие органические соединения
32. Факторы роста микроорганизмов
макроэлементы (Ca, Mg, Fe, К, Mn)микроэлементы (Co, Ni, Cu, Zn, Mo и др.)
Пурины и пиримидины
витамины
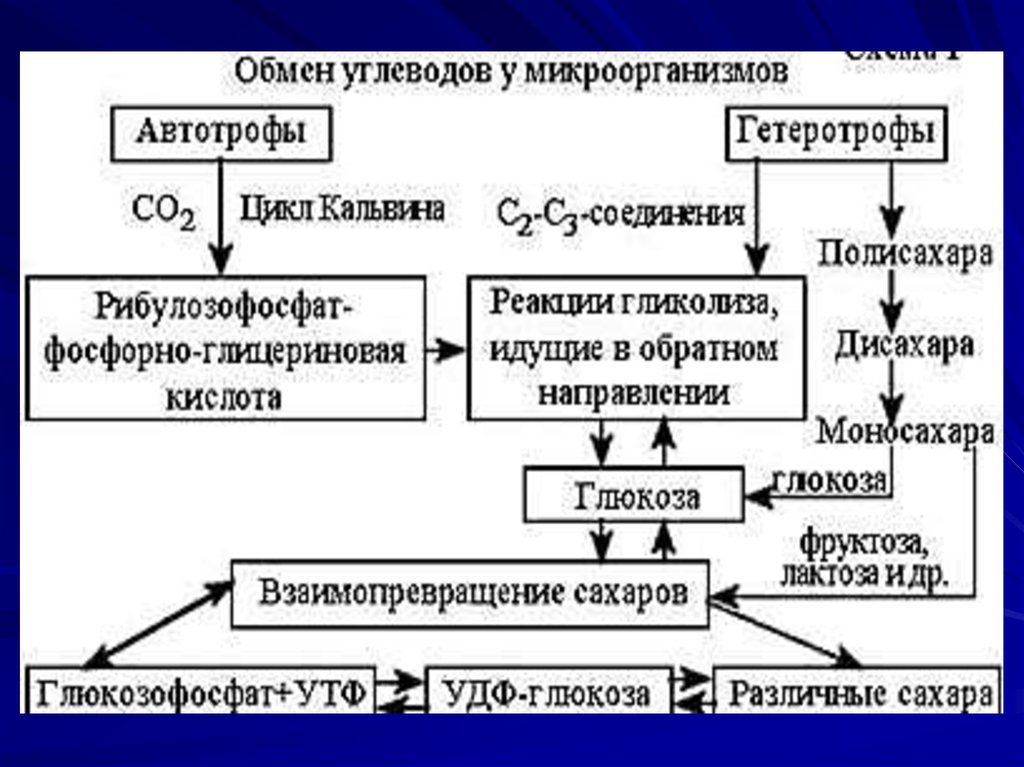
33. Синтез углеводов
Углеводы представлены в виде моно-, ди-,полисахаридов, а также комплексных
соединений
Автотрофы синтезируют углеводы в реакциях
восстановления пентозофосфатного цикла
Гетеротрофы - в гликолитическом пути и
путем глюконеогенеза (из неуглеводных
предшественников)
34.
35. Получение аминокислот прокариотами
Осуществляется из:Пирувата, альфакетоглутората, фумарата (из
цикла трикарбоновых кислот) в реакциях
аминирования (ионами NH4) и
переаминирования;
Из молекул белка (протеазы, пептидазы);
В готовом виде из клетки-хозяина
36.
37. Синтез липидов
У бактерий преобладаютдлиноцепочечные
(С14-С18) насыщенные
жирные кислоты и
ненасыщенные жирные
кислоты с одной
двойной связью.
Из промежуточных
продуктов:
ацетилкоэнзима А и
диоксиацетилфосфата.
38. Энергетический метаболизм (катаболизм)
- Поток химических реакций, сопровождающийсямобилизацией энергии и преобразованием ее в форму,
которая затем может использоваться во всех
энергозависимых процессах.
- В процессе катаболических реакций происходит
выделение энергии, которая накапливается в молекулах
макроэргов.
39.
40. В зависимости от источников энергии
Микроорганизмы делятся:Фототрофы- способные использовать энергию
света (фотосинтезирующие)
Хемотрофы- получают энергию за счет
окислительных и восстановительных реакций
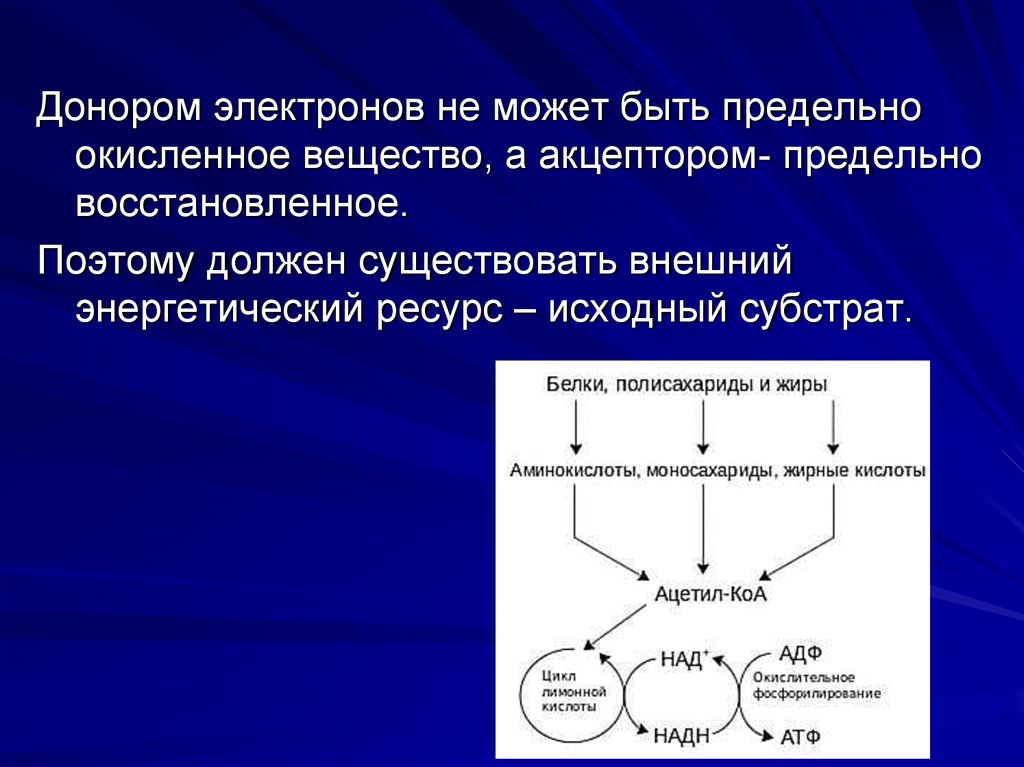
41.
Донором электронов не может быть предельноокисленное вещество, а акцептором- предельно
восстановленное.
Поэтому должен существовать внешний
энергетический ресурс – исходный субстрат.
42. В зависимости от природы доноров электронов
Микроорганизмы делятся:Хемолитотрофы (хемоавтотрофы)- Н2, Fe, NH3, CH3
и др.
Хемоорганотрофы (хемогетеротрофы)-органические
соединения.
Большинство прокариот являются
хемоорганогетеротрофами.
43.
44. Механизмы получения энергии у бактерий:
Окислительный метаболизм (дыхание);Бродильный (ферментативный) метаболизм
Смешанный метаболизм
фотосинтез
45. При окислительном метаболизме
Энергия образуется в реакциях окислениявосстановления, при которых донорами электроновмогут быть органические и неорганические
соединения, а акцептором- только молекулярный
кислород.
46.
Окислениепроисходит в
результате переноса
электронов через
локализованную в
мембране
дыхательную
электронтранспортную цепь
(мембранное
фосфорилирование).
47.
Дыхательная цепь у бактерий- сложнаямультиферментная система, локализована на ЦПМ и
мезосомах.
Переносчики располагаются в мембране
асимметрично, по разные стороны.
Они осуществляют транспорт водорода (электронов),
используя коферменты (НАД, флавопротеины,
хиноны и цитохромы).
48. Организация дыхательной цепи
49.
У микроорганизмов существует несколькотипов богатых энергией соединений. Самые
многочисленные:
ацилфосфаты,
нуклеотидди- и трифосфаты,
аденозинфосфосульфат
ацилтиоэфиры
50. При анаэробном дыхании
происходит перенос высокоэнергетической фосфатнойгруппы от молекулы-донора на АДФ с образованием
АТФ.
Субстратное фосфорилирование
S- Ф + АДФ= S + АТФ
51. Ферментативный (бродильный) метаболизм
Процесс получения энергии при которомотщепленный от субстрата водород
переносится на органические соединения.
Брожение разновидность анаэробного дыхания.
52. В зависимости от типа конечных продуктов различают:
Спиртовое брожениеМаслянокислое
Молочнокислое
Муравьинокислое
Уксуснокислое
Пропионовокислое и др.
53.
54. Сбраживание углеводов
Осуществляется:Гликолитическим путем (Эмбдена-МейергофаПарнаса);
В окислительном пентозофосфатном (ВарбургаДиккенса-Хореккера) пути;
2-кето-3- дезокси-6-фосфоглюконатном пути
(Энтнера-Дудорова)
55.
56. фотосинтез
Типы фотосинтеза:Бескислородный фотосинтез, наблюдается у
зеленых и пурпурных бактерий и гелиобактерий;
Кислородный фотосинтез, зависимый от
хлорофила (цианобактерии, прохлорофиты)
Бескислородный фотосинтез, зависимый от
бактериородопсина (галофильные
архебактерии)
57. По способу дыхания
Анаэробы (облигатные –для них кислород токсичен;факультативные - растут и размножаются как при
присутствии О2, так и без него);
Аэробы (облигатные)-содержание кислорода не ниже
парциального давления воздуха до 40%
Микроаэрофилы- при пониженной концентрации О2
(ниже 2%).
Капнофилы- пониженное содержание О2 и
повышенное СО2.
58.
Прокариоты способны синтезировать энергиюв большем количестве, чем требуется.
Они способны консервировать ее путем синтеза
восстановленных высокополимерных молекул
(полисахаридов) упакованных в гранулы,
окруженные белковой оболочкой.
59. Ферменты бактерий
Все метаболические процессы протекающие вмикроорганизмах являются ферментозависимыми.
Набор ферментов конкретных микроорганизмов
определяется их генетической информацией.
60. По своей природе
ферменты-белки.Ферменты распознают соответствующие субстраты,
вступают с ними во взаимодействие,
ускоряют протекание химических реакций.
Могут быть связаны с конкретными структурами
микробной клетки.
Ферменты могут функционировать самостоятельно, или
образовывать взаимосвязанные комплексы.
61. Классификация ферментов
(International Union of Biochemistry)класс – подкласс- субподкласс.
62.
Оксидоредуктазыокислительно-восстановительные ферменты:
дегидрогеназы, оксидазы.
лиазы
отщепляют от субстратов группы
негидролитическим путем (карбоксилазы)
гидролазы
расщепляют вещества на более простые с
присоединением воды (эстеразы, фосфатазы,
глюкозидазы и др.)
изомеразы
превращают органические соединения в их
изомеры (фосфо- гексоизомераза)
ускоряют синтез сложных соединений из более
простых (аспарагинсинтетаза,
глутаминсинтетаза)
переносят отдельные радикалы и атомы от одних
соединений к другим.
Лигазы,
синтетазы
Трансферазы
63. По месту действия
различают:Эндоферменты, катализируют метаболизм, проходящий
внутри клетки;
Экзоферменты- выделяются клеткой в окружающую
среду, расщепляя макромолекулы питательных
субстратов до простых соединений, усваиваемых
клеткой.
64.
Регуляция метаболизма микробной клеткисводится к регуляции активности
ферментативных реакций.
Скорость ферментативных реакций регулируется:
Путем изменения количества ферментов;
Путем изменения их активности
65. активность ферментов
Характеристикой активности ферментовявляется скорость, с которой они
катализируют ту или иную реакцию.
Она измеряется скоростью превращения
субстрата или скоростью накопления продуктов
реакции.
66. Активность ферментов зависит
от температуры среды,рН,
Присутствия ионов металлов,
конечных продуктов метаболизма,
электрических импульсов
Фермент
Оптимальное значение рН
света и др.
Пепсин
Пируват-карбоксилаза
1,5-2
4,8
Каталаза
6,8-7
Фумараза
6,5
Уреаза
Кабоксипептидаза
6,8-7,2
7,5
Трипсин
6,5-7,5
Аргиназа
9,5-9,9
67. Количество фермента
Регулируется:на этапе транскрипции,
трансляции,
в процессе сборки и разрушения ферментного белка
68.
Конститутивные ферменты- постоянносинтезируются в микробных клетках в определенных
концентрациях (ферменты гликолиза).
Индуцибельные (адаптивные) ферменты
- их концентрация резко возрастает в зависимости от
наличия соответствующего субстрата
69.
Аллостерическиеферменты – их активность
меняется в зависимости от
взаимодействия с
метаболитами или
субстратами. Активатором
выступает субстрат.
Накопление конечных
продуктов метаболизма
приводит к их
ингибированию.
70.
В зависимости от химической природысубстрата различают ферменты:
сахаролитические,
протеолитические,
липолитические.
71. Методы определения ферментативной активности
Химический метод – количественное определение субстратаили продуктов с помощью химических реагентов
(гликозилгидролазы – по образованию восстанавливающих
сахаров).
Спектрофотометрический метод – измерение скорости
ферментативной реакции по изменению поглощения субстрата
при характеристической длине волны (лиазы – по
образованию двойной связи).
72.
Манометрический метод – определение количества газа,выделяющегося в процессе реакции (оксидазы – по
поглощению О2, декарбоксилазы – по выделению СО2).
Поляриметрический метод – фиксируется изменение
оптического вращения (β-фруктофуранозидаза).
Хроматографический – количественное определение
субстрата или продуктов с помощью различных видов
хроматографии: бумажной (анализ сахаров),
тонкослойной (гликозидов со сложными агликонами),
ВЭЖХ (аминокислотный анализ и др.).
73. Вещества ауторегуляторы
Регулируют межклеточные взаимодействия. Онихарактеризуются строгой родо- и видоспецифичностью.
Большинство таких ауторегуляторов- вещества
липидной природы, которые воздействуют на
клеточные ферменты.
Они обеспечивают популяционные взаимодействия в
составе биопленок.
74. Питательные среды
Используют для выращивания микроорганизмов вискусственных условиях.
Они могут быть по консистенции: плотными, жидкими,
полужидкими.
В зависимости от набора питательных веществпростыми и сложными.
75. Питательные среды
По происхождению:Животного или растительного происхождения
Синтетические среды - готовят из определенных
химически чистых органических и неорганических
соединений, взятых в точно указанных концентрациях.
Эти среды легко воспроизводимы.
76. По целевому назначению питательные среды
Делят:Универсальные (МПА, МПБ);
Элективные (ЖСА, желчный бульон);
Дифференциально-диагностические (Эндо,
Плоскирева);
Специальные: транспортные, обогащения,
с повышенной питательной ценностью и др.
77. Элективные (избирательные) питательные среды
Обеспечивают преимущественный ростопределенной группы бактерий (желточно-солевой
агар, желчный бульон)
78. Дифференциально-диагностические среды
Позволяют дифференцироватьгруппы или виды бактерий по
ферментативной активности
Среды Эндо, Левина,
Плоскирева – по способности
и неспособности
ферментировать лактозу
Среды Гисса – используются
на 3 этапе выделения чистой
культуры для определения
спектра сахаролитической
активности.
79. Требования к условиям культивирования
Среда должна быть полноценнойопределенной рН
Определенного осмотического давления и
редокс-потенциала
Оптимальная температура
Вязкости
Изотоничности, прозрачности
Стерильности
Длительность культивирования
освещение
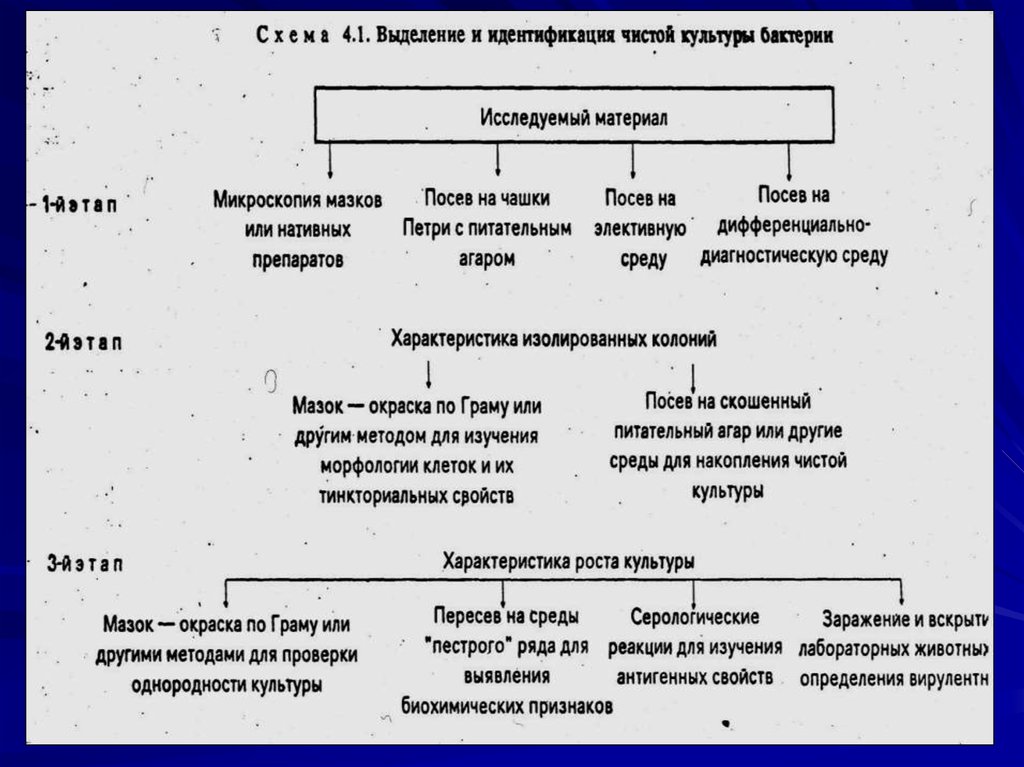
80. Этапы выделения чистой культуры бактерий
1 этап:микроскопия мазков или нативного материала;
посев на чашки с набором питательных сред (ЭС, ДДС,
СО).
2 этап:
Изучение морфологии клеток и их тинкториальных
свойств;
Изучение культуральных свойств изолированных колоний
(Пересев на скошенный агар)
3 этап:
Проверка однородности выделенной культуры;
Идентификация культуры по биохимическим и
антигенным свойствам,
определение вирулентных свойств (лабораторных
животных)
определение чувствительности в антибиотикам и
бактериофагам
81.
82. Рост бактерий
Координированное увеличение количествавсех компонентов микробной клетки (массы
клетки).
83. Размножение бактерий
Увеличение количества клеток в популяцииПоперечное деление:
Синтеза поперечной перегородки ( у грам+ бактерий)
Образования перетяжки ( у грам «-» бактерий)
84. Эффективность размножения микроорганизмов оценивается:
- Концентрацией клеток культуры в мл.- Временем генерации- промежутком времени за
который число клеток удваивается
- Константой скорости деления- число удвоений в
час
- Константой скорости роста
85. Стадии развития микроорганизмов в жидкой питательной среде:
Стационарная фага (отсутствия роста) длится 1 -2 часа.Лаг-фаза- приспособления. Размножение бактерий идет
медленно в следствие адаптации к условиям среды
Лог-фаза- экспоненциального роста. Увеличение
количества бактерий в геометрической прогрессии.
Фаза отрицательного ускорения - уменьшения скорости
деления бактерий
Стационарная фаза. Количество погибающих и
образующихся клеток равно.
Фага гибели- включает в себя гибель микроорганизмов
вначале в логарифмической прогрессии, а затем
скорость отмирании снижается. Бактерии могут
сохраняться в виде покоящихся форм и спор.
86.
87. В жидкой среде бактерии
Образуют:Помутнение;
Поверхностную пленку;
Осадок;
Пристенный или придонный рост
88.
На плотной средебактерии образуют
колонии, которые
различаются по
своим свойствам
89. Методы оценки состояния микробной культуры
Прямые (непосредственный подсчетклеток под микроскопом в счетных камерах
или фиксированных мазках)
90.
Косвенные (высев на плотные питательныесреды, осаждение на мембранных фильтрах,
мутность суспензии, определение биомассы,
общего азота, белка и др.)
91.
Появление методапроточной
цитометрии
позволило изучать
физиологическое
состояние
отдельных клеток.
92. Пигменты бактерий
По химическому составу и свойствам неоднородны.растворимые в воде;
растворимые в спирте;
нерастворимые в воде;
нерастворимые в воде
и спирте.
93.
Каротиноиды- жирорастворимые пигментыкрасного, желтого и оранжевого цвета
(микобактерии)
Пирроловые- спирторастворимые: продигиозин
(Serratia marcescens);
Фенозиновые- водорастворимые пигменты
синие или зеленые:
пиоцианин (синегнойная палочка)
Меланины- нерастворимые пигменты черного и
коричневого цвета (порфиромонасы)
94. Функции пигментов
защищают бактерии от природнойультрафиолетовой радиации,
участвуют в процессах дыхания, реакциях
синтеза,
обладают антибиотическим действием