














 biology
biologySimilar presentations:










Бактериофаги
1. Бактериофаги
ФаготипированиеБактериофа́ги (фаги) (от др.-греч. φᾰγω —
«пожираю») — вирусы, избирательно
поражающие бактериальные клетки.
2.
• Фаги, бактериальные вирусы, вызывающиеразрушение (лизис) бактерий и других
микроорганизмов.
• Б/ф. размножаются в клетках, лизируют их и
переходят в др., как правило, молодые, растущие
клетки.
• Впервые перевиваемый лизис бактерий
(сибиреязвенной палочки) наблюдал в 1898
русский микробиолог Н. Ф.Гамалея.
• В 1915 английский учёный Ф. Туорт описал это же
явление у гнойного стафилококка,
• в 1917 французский учёный Ф. Д'Эрелль назвал
литический агент, проходящий через
бактериальные фильтры, «Бактериофагом».
3.
4.
5.
Подробная схема Т-четного фага; справа показано, как он заражаетбактерию. Сначала фаг прикрепляется базальной пластинкой,
опираясь при этом на хвостовые нити. Молекула лизоцима на
базальной пластинке разъедает стенку бактериальной клетки, хвост
сокращается, и нить ДНК выстреливается в бактерию. Фенотип фага
после этого отбрасывается, он уже не нужен.
6. Взаимодействие бактериофагов с клеткой
• специфично, так как они, как правило, инфицируютбактерии только определённого вида. Подобно вирусам
животных, репродуктивный цикл литических бактериофагов
включает адсорбцию свободного фага на клетке, инъекцию
ДНК, репродукцию фага, выход дочерних популяций.
• Прикрепление фага к бактерии (адсорбция) происходит
при помощи поверхностных структур бактериальной стенки,
служащих рецепторами для вирусов (в
липополисахаридном слое, в наружной мембране). На
бактериях без клеточной стенки (протопласты, L-формы)
бактериофаги не адсорбируются. Некоторые фаги в
качестве рецепторов используют F-пили. Помимо
рецепторов, адсорбция фага зависит от рН среды,
температуры, наличия катионов и некоторых соединений.
При избытке фага на одной клетке может
адсорбироваться до 200-300 вирусных частиц.
7.
• После адсорбции происходит ферментативноерасщепление клеточной стенки лизоцимом,
находящимся в дистальной части отростка. Базальная
пластина хвоста лизирует прилегающий фрагмент
клеточной стенки, выделяя присутствующий в отростке
лизоцим. Одновременно в чехле высвобождаются ионы
Са2+, активизирующие АТФазу, что вызывает сокращение
чехла и вталкивание стержня хвоста через ЦПМ в клетку.
Затем вирусная ДНК впрыскивается в цитоплазму
(внедрение вирусной ДНК). Поскольку диаметр канала
лишь немного превышает диаметр молекулы ДНК (около 20
нм), то ДНК способна попадать в цитоплазму только в
форме нити.
• Проникнув в клетку, ДНК фага «исчезает»; уже через
несколько минут обнаружить вирус не удаётся. В этот, так
называемый скрытый период (эклипс) вирус берёт на себя
генетическое управление клеткой, осуществляя полный
цикл репродукции фага. К его окончанию составляющие
фага соединяются в зрелый вирион.
• Синтез фаговых белков. В первую очередь
синтезируются ферменты, необходимые для образования
копий фаговой ДНК. К ним относятся
8. Взаимодействие фага с бактериальной клеткой.
9.
• ДНК-полимераза, киназы (для образованиянуклеозидтрифосфатов) и тимидилат синтетаза. Они
появляются в клетке через 5-7 мин после её заражения.
Клеточная РНК-полимераза транскрибирует вирусную ДНК
в мРНК, которая транслируется бактериальными
рибосомами в «ранние» белки фага, включая вирусную
РНК-полимеразу и белки, способные посредством
различных механизмов ограничивать экспрессию
бактериальных генов. Вирусная РНК-полимераза
осуществляет транскрипцию «поздних» белков (например,
белков оболочки и эндолизина), необходимых для сборки
фаговых частиц дочернего поколения. Некоторые вирусы
расщепляют ДНК клетки-хозяина до нуклеотидов, чтобы
использовать их для синтеза собственных нуклеиновых
кислот.
• Репликация нуклеиновых кислот реализуется за счёт
активности вновь синтезированных вирусных ДНКполимераз, производящих множественные копии вирусных
нуклеиновых кислот.
• Вновь синтезированные белки формируют в цитоплазме
пул предшественников, входящих в состав головок и
хвостов дочерних
10.
• вирусных частиц. Другой пул содержит ДНК потомства.Специальные аффинные области в вирусной ДНК
индуцируют объединение предшественников головок
вокруг агрегатов нуклеиновой кислоты и образование ДНКсодержащих головок. Заполненная головка затем
взаимодействует с хвостовой частью, образуя
функциональный фаг. Весь процесс (от адсорбции до
появления вновь синтезированных вирусов) занимает
около 40 мин. После образования потомства клетка
хозяина лизируется, высвобождая дочернюю
популяцию. В разрушении клеточной стенки участвуют
различные факторы: фаговый лизоцим, увеличенное
внутриклеточное давление. Такой лизис бактериальной
клетки называется лизисом изнутри (внутренний), но
может быть и лизис извне (наружный) – происходит, когда
на клетку адсорбируется сразу очень много бактериофагов,
они проделывают многочисленные отверстия в клеточной
стенке, через которые содержимое бактериальной клетки
вытекает и она гибнет. В этом случае бактериофаг не
размножается.
11.
• Если фаги вызывают лизис зараженной клетки с выходом вокружающую среду большого количества фаговых частиц, спрособных
поражать новые клетки, то такие фаги называют вирулентными.
• В некоторых случаях вирулентных свойств фага оказывается
недостаточно для разрушения бактерии. Подобные вирусы —
умеренные фаги — претерпевают любопытные превращения,
известные как редукция фага.
• При этом процессе ДНК вируса не вызывает образования
вирусспецифических белков и нуклеиновых кислот, но включается в
бактериальную хромосому. С практической точки зрения бактерия
приобретает новый набор генов (встроенного вируса), а также
становится «иммунной» к повторному заражению (интерференция
вирусов).
• Подобный феномен известен как лизогения, а популяции бактерий —
как лизогенные культуры. ДНК умеренного вируса реплицирует
синхронно с размножением лизогенной бактерии, а иногда фаг
начинает спонтанно размножаться, а клетка подвергается лизису.
• Некоторые умеренные фаги не способны образовывать дочерние
популяции, то есть являются дефектными вирусами. Дефектные фаги
используют как векторы в генной инженерии.
• Вирусная ДНК может длительно сохраняться в бактериальном
потомстве. Такие латентные бактериофаги известны как провирусы,
или профаги.
12. Пути развития умеренного фага
13.
• Электроннаямикрофотография
одного из сложных
бактериальных
вирусов — фага Т2.
Хорошо видны
молекулы белка,
слагающие хвост и
голову, а также
хвостовые нити
(слева). Размеры
здесь также в
ангстремах.
.
14.
• Относительные размеры некоторых вирусов(электронная микрофотография с увеличением
в 200000 раз). Слева направо — один из
маленьких фагов φX174, вирус табачной
мозаики, сложный бактериофаг Т4 (из группы
так называемых Т-четных фагов бактерии
кишечной палочки).
15.
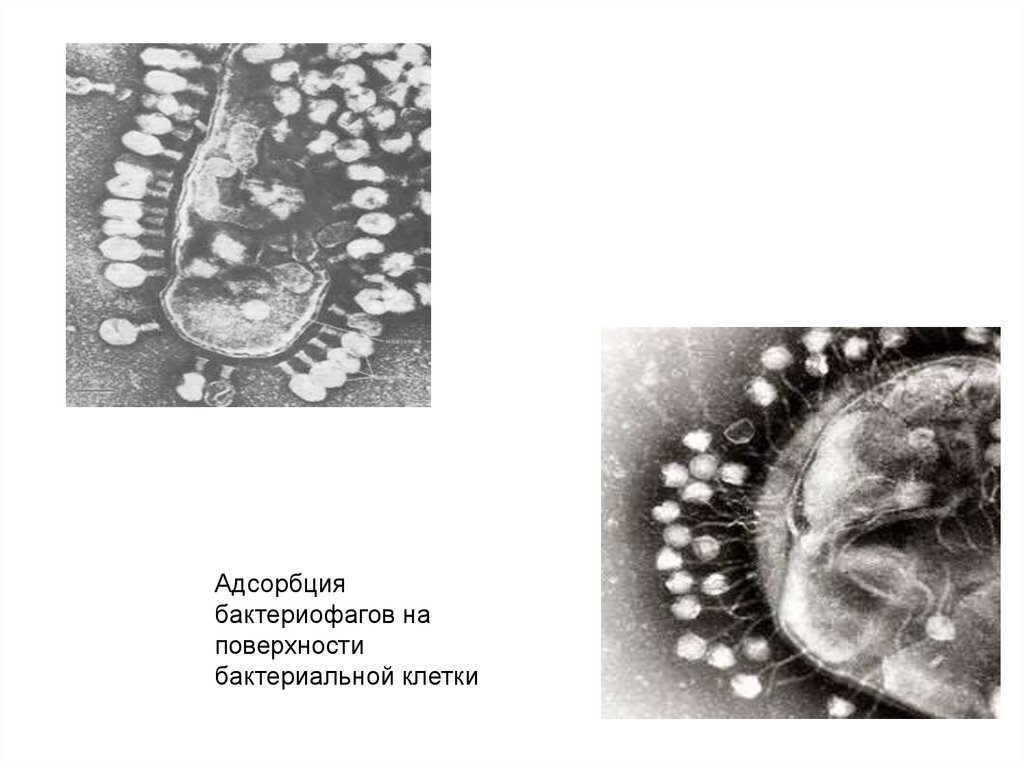
Адсорбциябактериофагов на
поверхности
бактериальной клетки
16.
• Б. делят на вирулентные, вызывающие лизис клетки собразованием новых частиц, и умеренные
(симбиотические), которые адсорбируются клеткой и
проникают в неё, но лизиса не вызывают, а остаются в
клетке в латентной (скрытой) неинфекционной форме
(Профаг). Культуры, содержащие латентный фаг,
называются лизогенными. Лизогения передаётся потомству
бактерии. Лизогенная культура может содержать 2—3 и
более фагов; она, как правило, устойчива против
находящихся в ней фагов (лишь небольшая часть клеток
лизируется и освобождает зрелые фаги). Воздействуя на
лизогенную культуру ультрафиолетовыми или
рентгеновскими лучами, перекисью водорода и некоторыми
другими веществами, можно значительно увеличить
количество клеток, освобождающих фаг (т. н. индукция Б.).
Лизогения широко распространена среди всех видов
бактерий и актиномицетов. В ряде случаев многие свойства
лизогенной культуры (токсичность, подвижность бактерий и
др.) зависят от наличия в ней определённых профагов.