


































































 biology
biologySimilar presentations:










Вирусы бактерий. Бактериофаги
1. вирусы бактерий
БАКТЕРИОФАГИ2. БАКТЕРИОФАГИ
• «пожирающие бактерии» (от бактерия +греч. phagos – пожирающий)
• вирусы бактерий, специфически
проникающие в бактериальные клетки и
поражающие их.
• Для обозначения используют:
- название м/о, из которых они выделены:
н-р, колифаги, стафилофаги,
- буквы латинского алфавита:
н-р, Т-бактериофаги
3.
4.
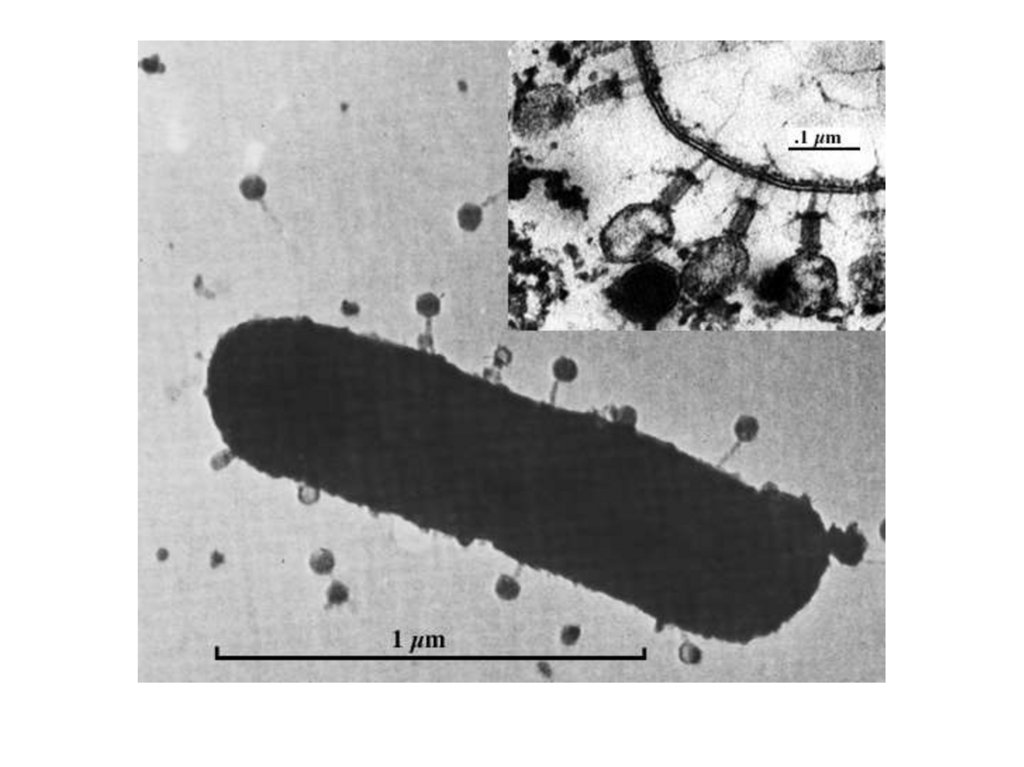
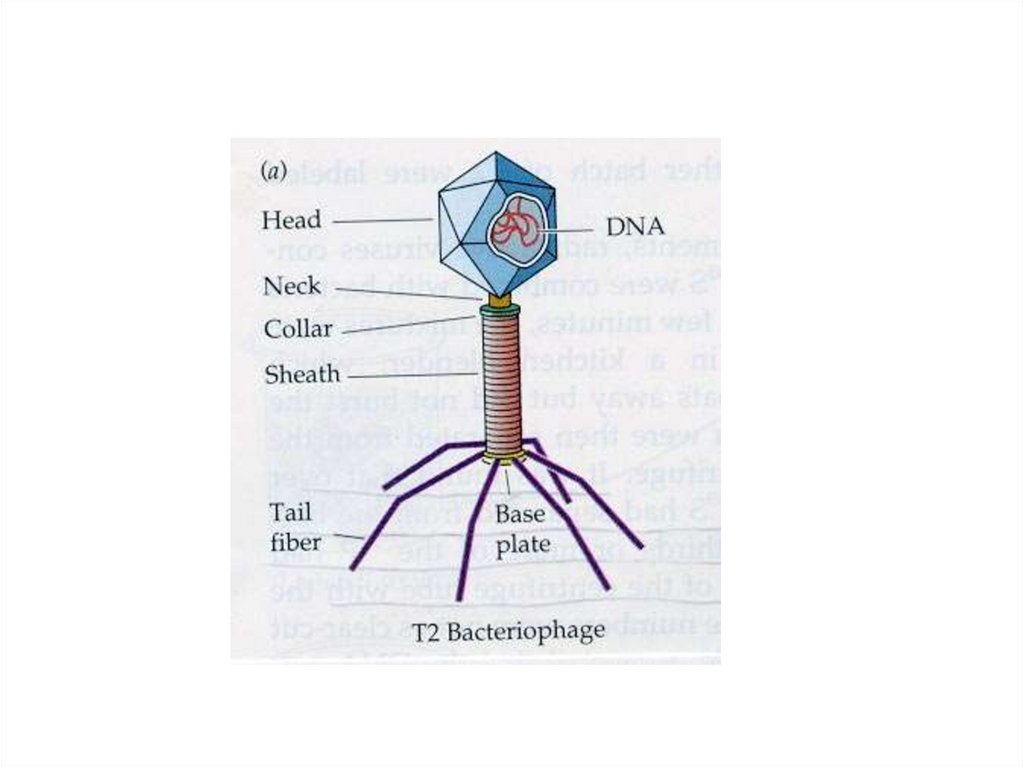
5. Строение бактериофагов
• икосаэдрическая головка,• хвостовой отросток:
• внутри – полый цилиндрический стержень,
сообщающийся с головкой,
• снаружи - чехол отростка, заканчивающийся
шестиугольной базальной пластинкой с шипами,
• от шипов отходят фибриллы (нити),
• капсид головки и чехол хвостового отростка
бактериофага состоят из полипептидных
субъединиц, уложенных:
• - по икосаэдрическому (головка),
• или спиральному (отросток) типу симметрии.
6.
7. Нуклеиновая кислота фага
• Бактериофаги (фаги) содержат ДНК илиРНК:
- Двунитевые или однонитевые,
- Линейные или кольцевые.
• Большинство – двунитевую ДНК,
замкнутую в кольцо
8. В состав головки входит:
• полипептид, состоящий из:• аспарагиновой и глутаминовой кислот,
• и лизина,
• - у некоторых – гистоноподобный белок →
суперспирализация ДНК.
9. В состав сокращающегося чехла входит:
• у некоторых фагов входит АТФ и ионыкальция.
• В дистальной части отростка – лизоцим.
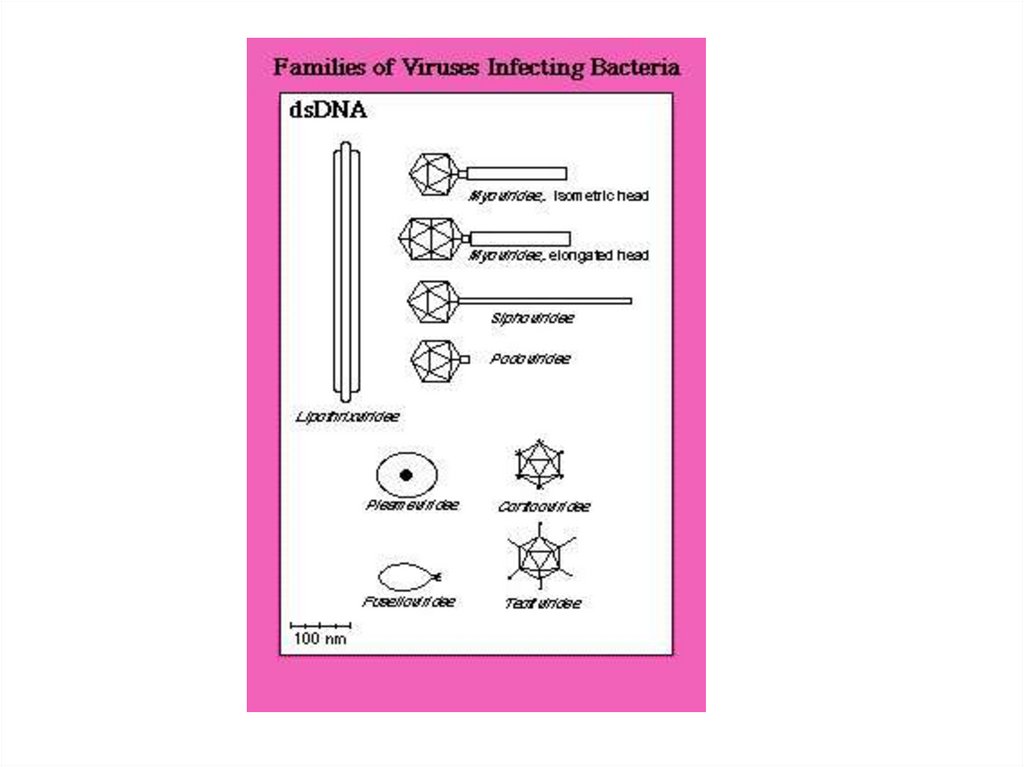
10. Морфологические типы бактериофагов
• I тип (нитчатые)– без головки (только отросток)
• II тип
– без отростка (только головка)
• III тип
– головка и отросток, короткий без чехла
• IV тип
– головка и отросток, длинный с чехлом, не
сократительный
• V тип
– головка и отросток, длинный с чехлом, сократительный
11.
12. Классификация бактериофагов по спектру действия
• полифаги– поражают несколько видов
• монофаги (видовые)
– поражают один вид
• типовые фаги
– поражают часть вида (фаговар)
13. Классификация фагов в зависимости от эффекта действия на бактериальную клетку
• вирулентные• умеренные
14. Классификация фагов в зависимости от эффекта действия на бактериальную клетку
вирулентный фаглизис
вирулентный фаг
дефектный фаг
общая
абортивная
трансдукция
15. Классификация фагов в зависимости от эффекта действия на бактериальную клетку
умеренный фагЛизогения:
• без изменения фенотипа бактерии
• с изменением фенотипа бактерии (фаговая конверсия)
лизис
умеренный
дефектный
специализированная трансдукция
16. Взаимодействие фагов с бактериями
• 3 типа:- продуктивный – вирулентные фаги→фаговое
потомство, бактерии лизируются;
- интегративный – умеренные →встраиваются
в геном клетки и сосуществуют с ней;
- абортивный → фаговое потомство не
образуется, бактерии сохраняют свою
жизнедеятельность.
17. Вирулентные бактериофаги
• попав в бактерию, реплицируются,формируя 200-300 фаговых частиц, и
вызывают гибель (лизис) бактерии = это
продуктивный тип взаимодействия
18. Взаимодействие вирулентного фага с бактериальной клеткой
адсорбция фага на специальных рецепторах КС(на протопластах не происходит)
проникновение НК
(депротеинизация)
репликация фаговой НК и синтез фаговых белков
сборка фаговых частиц
выход зрелых фагов
лизис бактерии
(«взрыв»)
бактерия не погибает
(некоторые нитчатые фаги)
ПРОДУКТИВНАЯ ИНФЕКЦИЯ
19. Этапы взаимодействия вирулентного фага с бактериальной клеткой =продуктивный тип взаимодействия
• 1. Бактериофаги с сокращающимся чехломадсорбируются на клеточной стенке с помощью
фибрилл хвостового отростка.
• 2. Чехол хвостового отростка сокращается, и
стержень с помощью ферментов (лизоцима)
просверливает оболочку клетки.
• 3. Через канал стержня бактериофага
нуклеиновая кислота инъецируется из головки
в бактериальную клетку, а капсид бактериофага
остается снаружи бактерии.
20. Взаимодействие бактериофага с оболочкой клетки
21. Этапы взаимодействия вирулентного фага с бактериальной клеткой =продуктивный тип взаимодействия
• 4. Инъецированная внутрь клетки нуклеиноваякислота подавляет биосинтез компонентов
клетки, заставляя ее синтезировать
нуклеиновую кислоту и белки бактериофага:
- происходит полный распад ДНК бактерии и ее
утилизация.
- если ДНК бактерии не хватает для образования
фаговой ДНК – она синтезируется из компонентов
среды.
22. Этапы взаимодействия вирулентного фага с бактериальной клеткой =продуктивный тип взаимодействия
• 5. Образовавшиеся в разных частях клеткикомпоненты бактериофага собираются в
фаговые частицы путем заполнения фаговой
нуклеиновой кислотой пустотелых капсидов
головки
• 6. Сформированная головка соединяется с
хвостовой частью, образуя новый фаг.
• Затем в результате лизиса клетки бактериофаги
выходят из нее.
23. Взаимодействие умеренного фага с бактериальной клеткой
адсорбция фага на специальных рецепторах КС(на протопластах не происходит)
проникновение НК
(депротеинизация)
интеграция фаговой НК в геном бактерии
профаг
(фаговый репрессор блокирует транскрипцию)
лизогенная культура
ЛИЗОГЕНИЗАЦИЯ
в дальнейшем – может произойти индукция профага
продуктивная инфекция
24. Взаимодействие умеренного фага с бактериальной клеткой
• Умеренные бактериофагивзаимодействуют с бактериями:
- либо по продуктивному,
- либо по интегративному типу.
• Продуктивный цикл умеренного фага идет
как и у вирулентных фагов, и заканчивается
лизисом бактерий.
25. Взаимодействие умеренного фага с бактериальной клеткой
• При интегративном типе ДНК умеренногофага встраивается в хромосому бактерии:
- приобретает форму кольца,
- интегрируется в гомологичную область,
- реплицируется синхронно с геномом
бактерии, не вызывая ее лизиса
(передается при делении бактерии).
26. Взаимодействие умеренного фага с бактериальной клеткой
• ДНК фага, встроенная в хромосомубактерии, называется профагом,
• культура бактерий — лизогенной;
• сам процесс – лизогенией
(от греч. lysis – разложение, genea –
происхождение).
27. Взаимодействие умеренного фага с бактериальной клеткой
• При лизогении фаги не образуются в результате“выключения“ фаговых генов репрессором
(=низкомолекулярный белок), кодируемым одним
геном фага.
• Профаги могут спонтанно или под действием
индуцирующих агентов (УФ-лучи, митомицин С и
др.) дерепрессироваться, исключаться из
хромосомы. Этот процесс заканчивается
продукцией фагов (индукция профага) и лизисом
бактерий.
28. Взаимодействие умеренного фага с бактериальной клеткой
• Профаг придает бактерии новые свойства, чтополучило название фаговой конверсии (лат.
conversio – превращение).
• Конвертироваться могут:
- морфологические,
- культуральные,
- биохимические,
- антигенные и другие свойства бактерий.
• Например, наличие профага в дифтерийной
палочке обусловливает ее способность
продуцировать дифтерийный экзотоксин.
29. Применение фагов
• для профилактики,• для лечения инфекций,
• в генной инженерии в качестве векторов для
получения рекомбинантной ДНК,
• для диагностики (например, для
фаготипирования с целью выявления
источника инфекции или внутривидовой
идентификации).
30. Практическое применение бактериофагов
Фагопрофилактика• брюшной тиф
• дизентерия
31. Практическое применение бактериофагов
ФаготерапияФаг применяется в том случае, когда
антибиотики применять нельзя,
Чаще всего местно
Н-р, стафилококковые инфекции
32. Практическое применение бактериофагов
Фагодиагностика1. Выявление определённого вида бактерий в
патологическом материале
–
реакция нарастания титра фага
2. Идентификация чистой культуры
–
определение вида
–
фагоиндикация
определение фаговара
фаготипирование
33. Выделение бактериофага
Материал:• объект внешней среды
• бактериальная культура
бактериальный фильтр
фильтрат
МПБ + чувствительная бактерия
Инкубация 24 час
роста нет – фаг присутствует (очищают фильтрованием)
рост есть – фаг отсутствует
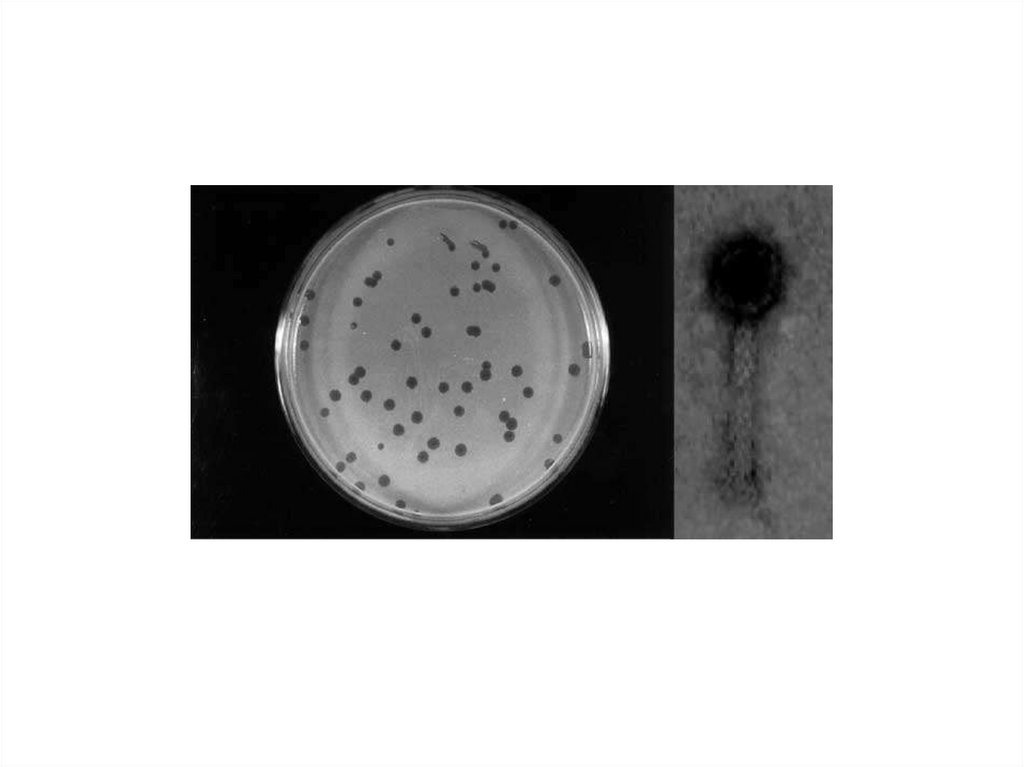
34. Определение активности бактериофагов = фагоиндикация
–Качественный метод:метод «стерильной дорожки»
–Количественные методы:
• А) Метод Грациа
• Б) Метод Аппельмана
35. Определение активности бактериофагов = фагоиндикация
– Качественный метод:На чашку газоном засевают культуру
микроорганизмов, наносят каплю
бактериофага и дают ей стечь. Чашки
инкубируют при 37 градусах 24 час и
учитывают результат:
• там, где стекал бактериофаг,
образовалась «стерильная дорожка»
36. Метод фагоиндикации = «стекающая капля» = «стерильная дорожка»
стекающая капля по газонузасеянной культуры
регистрация роста бактерий в
месте стекания капли
рост есть – роста нет – +
«стерильная дорожка»
37. Определение активности бактериофагов = фагоиндикация
–Количественные методы:• А) Метод Грациа: готовят десятикратные разведения
фага в хлориде натрия от 10-2 до 10-7
• Затем по 0,5 мл из каждого разведения смешивают с
таким же объемом бульонной культуры и 4 мл
расплавленного и остуженного до 45 град. агара и
выливают на чашки Петри.
• Когда агар застынет чашки помещают в термостат при
37 градусах на 24 часа и затем учитывают результаты:
-одна фаговая частица образует одно «стерильное пятно»
- Величина, показывающая концентрацию фага называется
титром.
38. Титрование фага по Грациа
• МПА + разведениефагосодержащего
материала +
чувствительная
культура
• МПА (подложка)
чашка Петри со средой
(в разрезе)
39.
40. Определение активности бактериофагов = фагоиндикация
–Количественные методы:Б) Метод Аппельмана: готовят десятикратные
разведения фага в питательном бульоне
от 10-2 до 10-8.
• Затем в каждую пробирку добавляют по 0,2мл
бульонной культуры и ряды ставят в термостат.
• После инкубации в термостате учитывают результаты:
= в положительном случае наблюдается
просветление среды.
Разведение в последней пробирке, где произошел
полный лизис культуры, называется титром фага.
41. Титрование фага по Аппельману
42. Фаготипирование бактерий
1. засев газоном на чашку с питательным агаромизучаемого штамма
2. чашку делят на квадратики и на каждый наносят
бактериофаг
3. инкубация
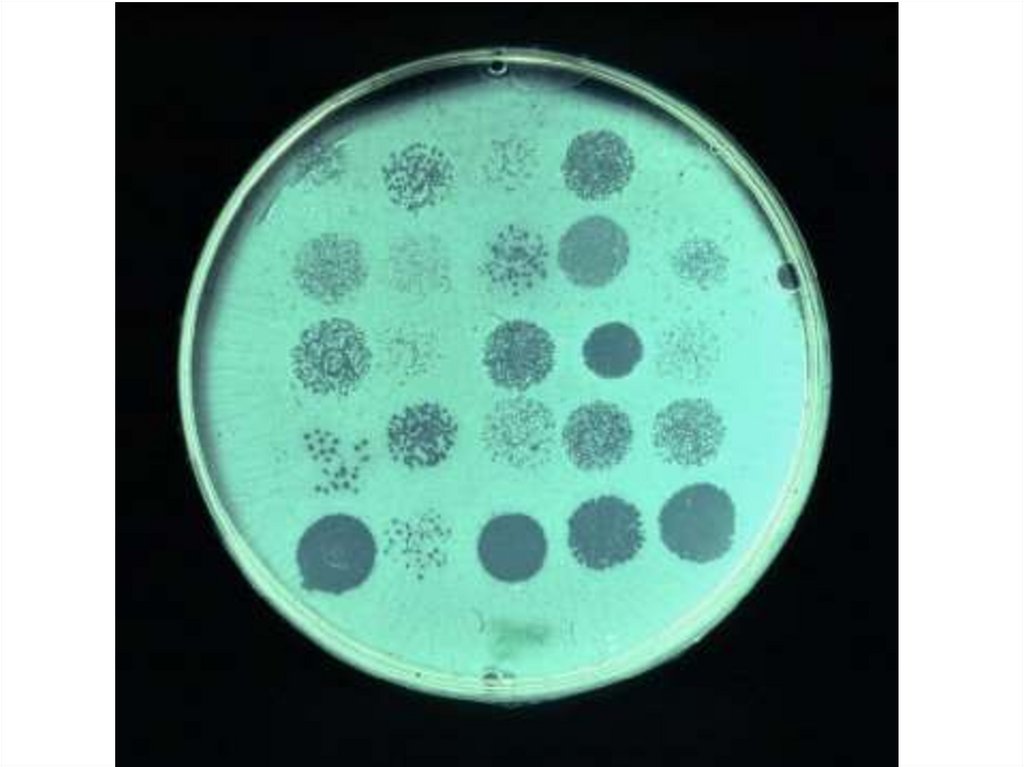
4. регистрация «стерильных пятен» («бляшек»)
5. фаготип (фаговар) = перечень типовых фагов,
лизирующих данный вариант
43.
44. Фаготипирование стафилококков
45. Определение спектра литического действия фага
• Чашку делят на квадратики и на каждыйгазоном засевают испытуемые штаммы,
• затем на каждый квадратик петлей или
пипеткой наносят каплю фага
• после инкубации в термостате в течение 24 час
определяют наличие «стерильных пятен».
• Количество культур, которые лизирует
бактериофаг – спектр его литического
действия.
46. Прионы
• – белковые молекулы, способныевызывать разрушение клеток организма
человека и животных.
• Они характеризуются устойчивостью:
- к высоким температурам,
- ионизирующей радиации,
- ультрафиолету.
47. Прионы
Прионный белок может существовать в двухформах:
нормальная клеточная форма(РrPc) обнаруживается в организме всех
млекопитающих.
Ген, кодирующий этот белок, расположен
в коротком плече 20 хромосомы.
РrPc участвует в передаче нервных
импульсов, в поддержании циркадных
ритмов клетки,
48. Прионы
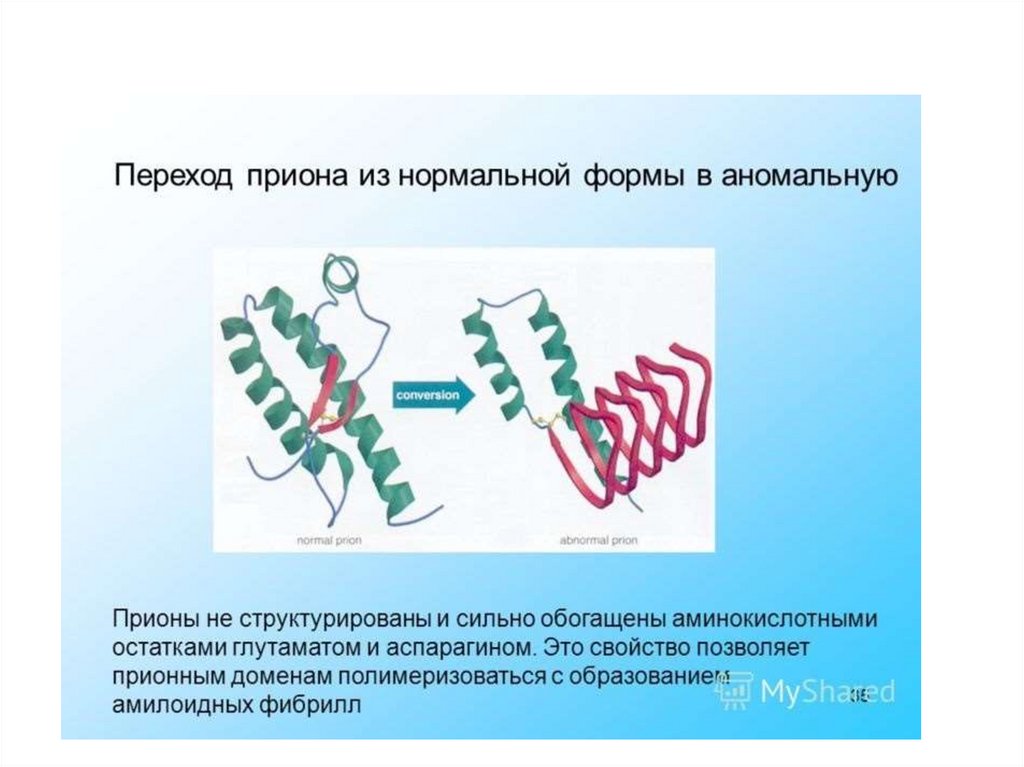
инфекционная форма (PrPs) –характеризуется:
- измененной вторичной и третичной
структурой молекулы,
- высокой устойчивостью к нагреванию,
ультрафиолетовому свету, проникающей
радиации и переваривающему действию
протеаз.
49.
50.
51.
52. Схема «размножения» прионов
53. ГЕНЕТИКА БАКТЕРИЙ
54. ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА У БАКТЕРИЙ
• 1. Нуклеоид = кольцевая молекула ДНК =бактериальная «хромосома» – кодирует
жизненно важные признаки;
• 2. внехромосомные факторы наследственности:
- Плазмиды,
- Транспозоны,
- IS-последовательности.
– кодируют признаки, дающие преимущество.
55.
• Единицей наследственности является ГЕН =участок ДНК, в котором зашифрована
последовательность аминокислот в
полипептидной цепочке, контролирующая
отдельный признак особи.
56. Гены:
• Структурные = обуславливают синтезопределенного белка (фермента), при мутации
образуется белок измененного состава,
• Ген-регулятор = определяет синтез белковой
молекулы-репрессора, подавляющего деятельность
структурных генов в отсутствии субстрата,
= при наличии субстрата репрессор временно
инактивируется и структурные гены,
освобожденные от его влияния, начинают
функционировать.
57.
• Совокупность генов, сосредоточенных внуклеоиде («Хромосоме») бактерий называется
генотип.
• Фенотип – совокупность всех признаков
микроорганизма, сформировавшаяся в результате
взаимодействия генотипа с внешней средой.
• Репликоны = генетические элементы, способные
самостоятельно реплицироваться: ДНК и
плазмиды.
58. Бактериальная«хромосома» = ДНК
двухцепочечная кольцевая молекула,
сод-т до 5 тыс. генов,
имеет молекулярную массу 1,7х109-2,8х109 дальтон,
включает 3х106-5х106 пар оснований,
имеет гаплоидный набор генов,
• расположена в цитоплазме клетки в многократно
свернутом и плотно упакованном виде,
• содержит гены, обуславливающие жизненно-важные
для бактерий признаки.
59. ДНК
60. Генетическая карта
• = это схематическоеизображение всех генов
микроорганизма.
• Гены, отвечающие за
определенный признак,
обозначают строчными
буквами латинского
алфавита со знаком «+»
(например, гистидиновый
ген – his+),
• отсутствие гена - знак «–»
61. ВНЕХРОМОСОМНЫЕ ФАКТОРЫ НАСЛЕДСТВЕННОСТИ
• 1.автономные – являются репликоном– плазмиды
• 2.неавтономные - реплицируются только в составе
репликона (нуклеоида или плазмиды):
– Транспозоны,
– IS-последовательности,
– Умеренные фаги.
62. ВСТРАИВАНИЕ ВНЕХРОМОСОМНЫХ ФАКТОРОВ НАСЛЕДСТВЕННОСТИ В НУКЛЕОИД
• в гомологичных участках– Плазмиды,
– Умеренные фаги.
• в любых участках
– Транспозоны,
– IS-последовательности.
63. ПЛАЗМИДЫ
– внехромосомные факторы наследственности у бактерий,• двухцепочечные молекулы ДНК,
• несут 40-50 генов,
• не являются жизненно-важными для бактерии,
• обусловливают признаки, позволяющие лучше
приспособиться к условиям обитания.
возможные состояния:
–
–
автономное (в цитоплазме),
интегрированное (в нуклеоиде).
В этом случае плазмида называется ЭПИСОМА.
64. контроль репликации плазмид со стороны нуклеоида
– строгий (делятся синхронно с нуклеоидом)1-2 копии на клетку (большие плазмиды),
– ослабленный (делятся чаще нуклеоида)
10-30 копий на клетку (малые плазмиды).
65. ПЛАЗМИДЫ
функции
1. регуляторная – компенсирует нарушение функции ДНК
нуклеоида,
2. кодирующая – вносит в генотип новую информацию.
– Могут быть:
– Трансмиссивные (конъюгативные) – содержат tra-оперон,
– Нетрансмиссивные (неконъюгативные) - не содержат.
66. ФУНКЦИИ TRA-ОПЕРОНА
• детерминирует образованиеконъюгативных пилей,
• мобилизирует на перенос:
– саму конъюгативную плазмиду (F+),
– другую, неконъюгативную, плазмиду (RTF),
– участок нуклеоида (Hfr).
67. Фенотипические признаки, сообщаемые бактерии плазмидами
• устойчивость к антибиотикам,• образование бактериоцинов,
• продукция факторов патогенности,
• способность к синтезу антибиотиков,
• расщепление сложных органических веществ,
• образование ферментов рестрикции и
модификации.
68. Наиболее изучены плазмиды:
• F- плазмида = половой фактор – контролирует синтезполовых ворсинок,
= бывает: - автономной→ бактерия наз-ся F+ штаммом,
- интегрированной → Hfr – штамм,
= конъюгативная.
R-плазмида (resistance - устойчивость) –
обусловливает синтез ферментов, разрушающих
антибиотики, сульфаниламиды и др., в результате
бактериальная клетка становится устойчивой к
лекарственным препаратам,
- в 1 плазмиде м.б. 3-10 детерминант устойчивости.
69. Наиболее изучены плазмиды:
• Col-плазмиды - обусловливают синтез бактериоцинов( = белки, задерживающие рост других штаммов бактерий того
же вида). Бактерии, несущие такие плазмиды, обладают
преимуществом при заселении биотопа.
Плазмиды патогенности – определяют:
• синтез энтеротоксинов (Ent-)
• ферментов патогенности (Hly-),
• поверхностного антигена вирулентности ( Vir-).
Плазмиды биодеградации – несут информацию
об утилизации органических соединений, которые бактерии
используют в качестве источника углерода и энергии.
70. ТРАНСПОЗОНЫ
определение
= нуклеотидные последовательности (от 2 000 до 20000 пар
нуклеотидов), способные менять место своей локализации в
молекуле ДНК и мигрировать из одной молекулы ДНК в
другую.
состояние в бактериальной клетке
1. интегрированное в репликон (реплицируется вместе с
ним),
2. автономное (замыкается в кольцо и не реплицируется).
71. ТРАНСПОЗОНЫ
Состав:
– маркеры транспозона = особые концевые структуры,
которые отличают транспозон от др. фрагментов ДНК ,
–гены транспозиции,
–гены, детерминирующие синтез:
- токсинов,
- ферментов, обеспечивающих устойчивость к антибиотику,
- белков, обеспечивающих др. признаки.
72. IS-ПОСЛЕДОВАТЕЛЬНОСТИ
= вставки нуклеотидных последовательностей (порядка
1 000 пар нуклеотидов),
• содержат только гены, необходимые для собственного
перемещения:
= ген, кодирующий фермент транспозазу – обеспечивает
исключение IS-элемента из ДНК и его интеграцию в новый
локус,
= ген, обуславливающий синтез репрессора,
регулирующего весь процесс перемещения,
• -не способны реплицироваться самостоятельно.
отличия от транспозонов:
1. содержат только гены транспозиции,
2. не обнаружены в свободном состоянии.
73. Функции IS-ПОСЛЕДОВАТЕЛЬНОСТЕЙ
1. координация взаимодействия внехромосомныхфакторов наследственности между собой и с
бактериальной хромосомой для обеспечения их
рекомбинации
2. регуляторная - регуляция транскрипции генов путём их
«включения/выключения»
3. индукция мутаций - инверсии, дупликации на
протяжении 5-9 пар нуклеотидов