








































")


 biology
biologySimilar presentations:





")

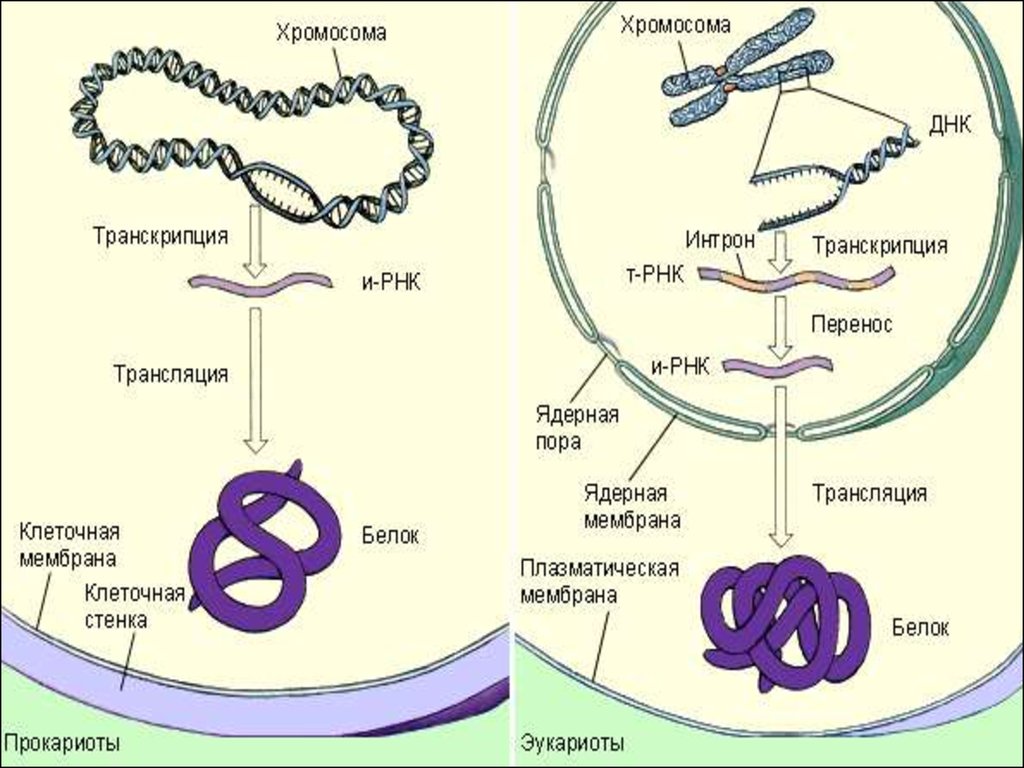
. Функции ДНК транскрипция")


Лекция 8. Молекулярная генетика. Транскрипция РНК эукариот
1. Транскрипция у эукариот Лекция 8
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА2. Словарик
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ3. Словарик
– процесс переноса остатка фосфорнойкислоты от фосфорилирующего агента-донора к субстрату,
катализируется киназами
– замещение атомов водорода в органических
соединениях остатком уксусной кислоты CH3CO (ацетильной группой),
катализируется ацетилтрансферазами
– последовательности ДНК, усиливающие транскрипцию
при взаимодействии со специфическими белками
– последовательности ДНК репрессирующие активность
генов, ослабляющие транскрипцию при взаимодействии с белками
– последовательности ДНК, определяющие
специфичность действия энхансеров и сайленсеров
4.
5.
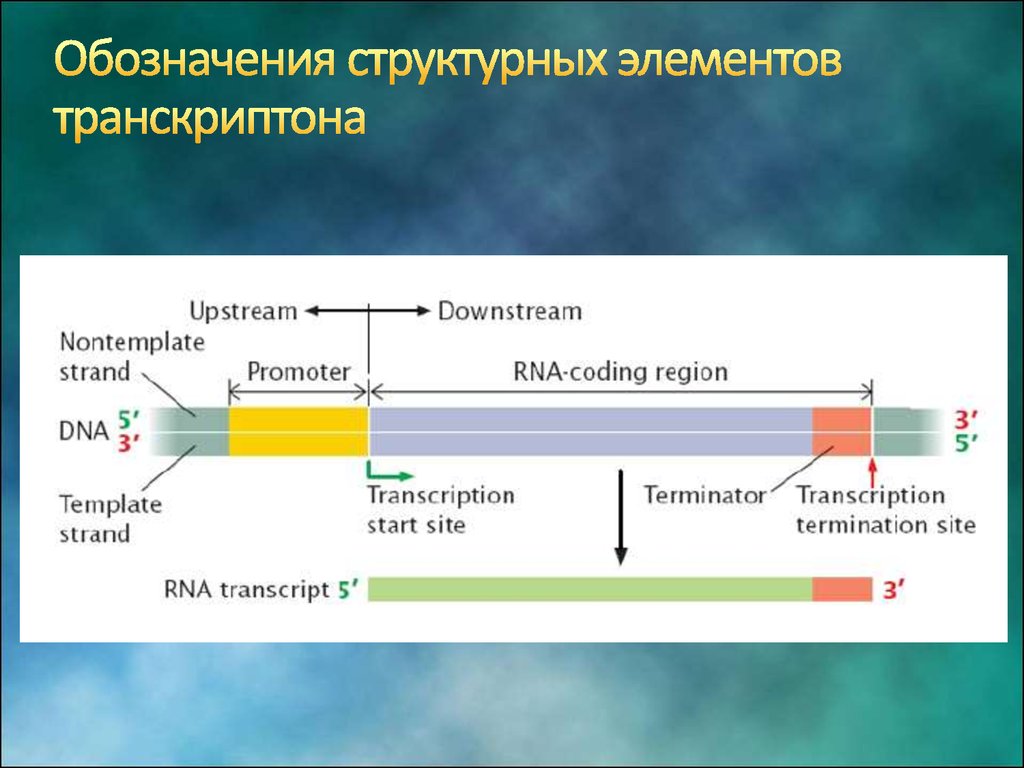
6. Особенности транскриптона эукариот
1• единица транскрипции – ген
2
• оператор отсутствует
3
• иная структура промотора
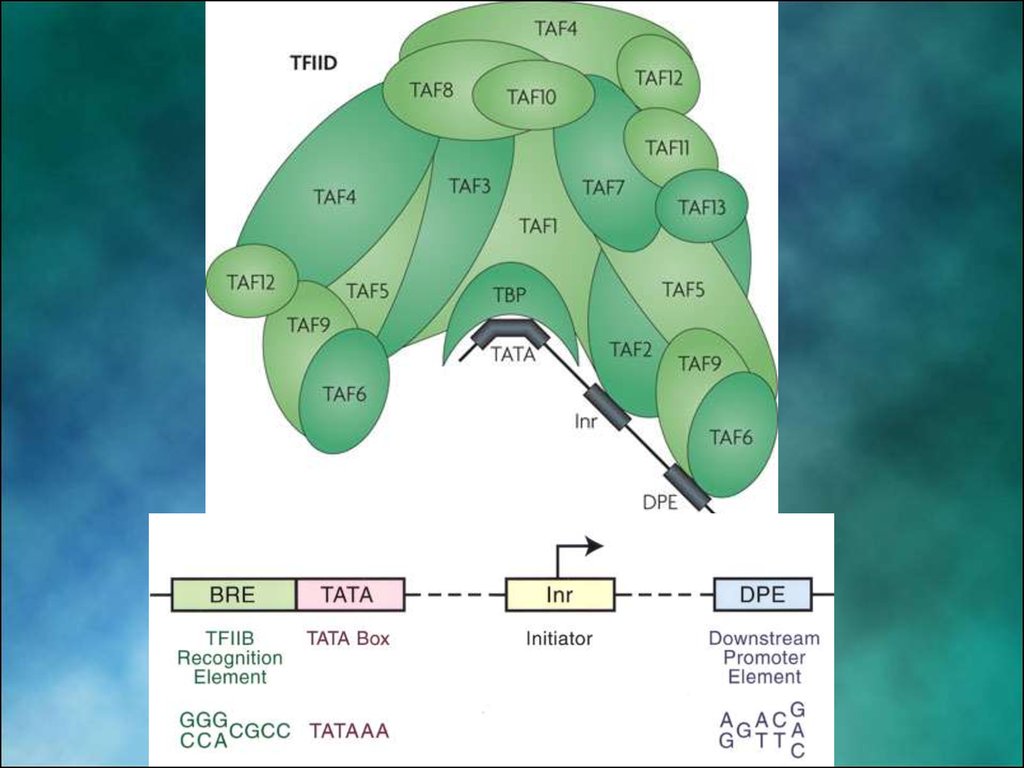
7. Структура промотора эукариот
На расстоянии -25 п.н. от +1 нукл. находится ТАТА-бокс, егопозиция определяет точку инициации транскрипции
На расстоянии -60-80 п.н. находится ЦААТ-бокс
Расстояние между ЦААТ и ТАТА большое и РНК-полимераза не
способна накрыть всю эту область. ЦААТ опознается своим
белком, а ТАТА - своим.
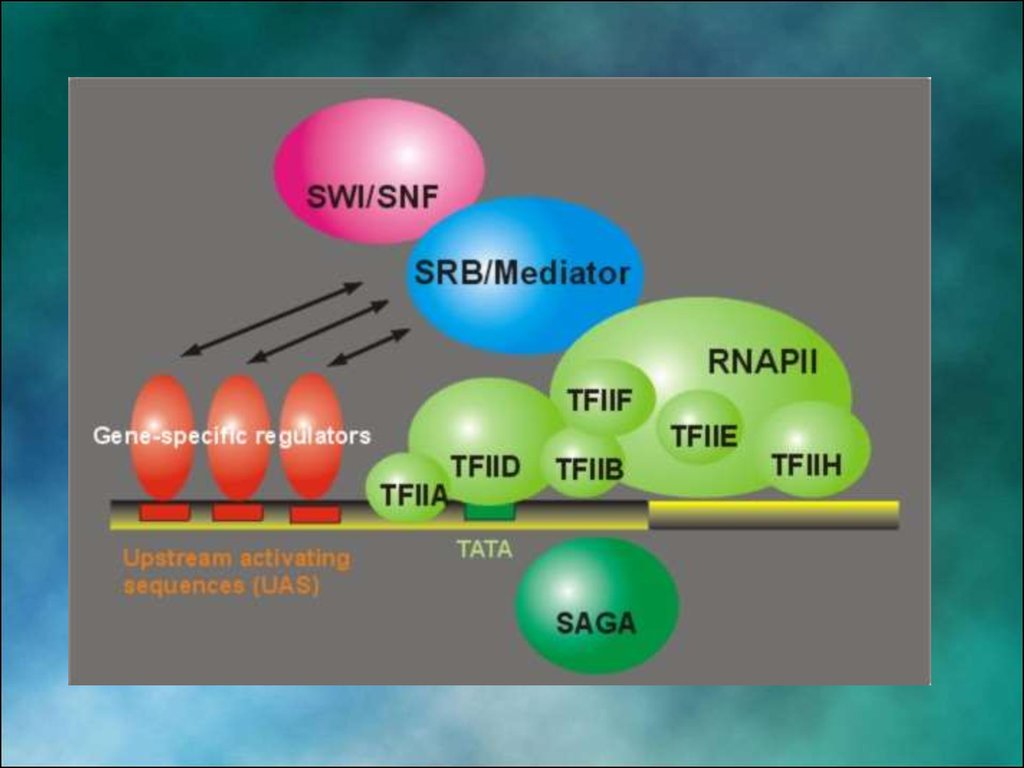
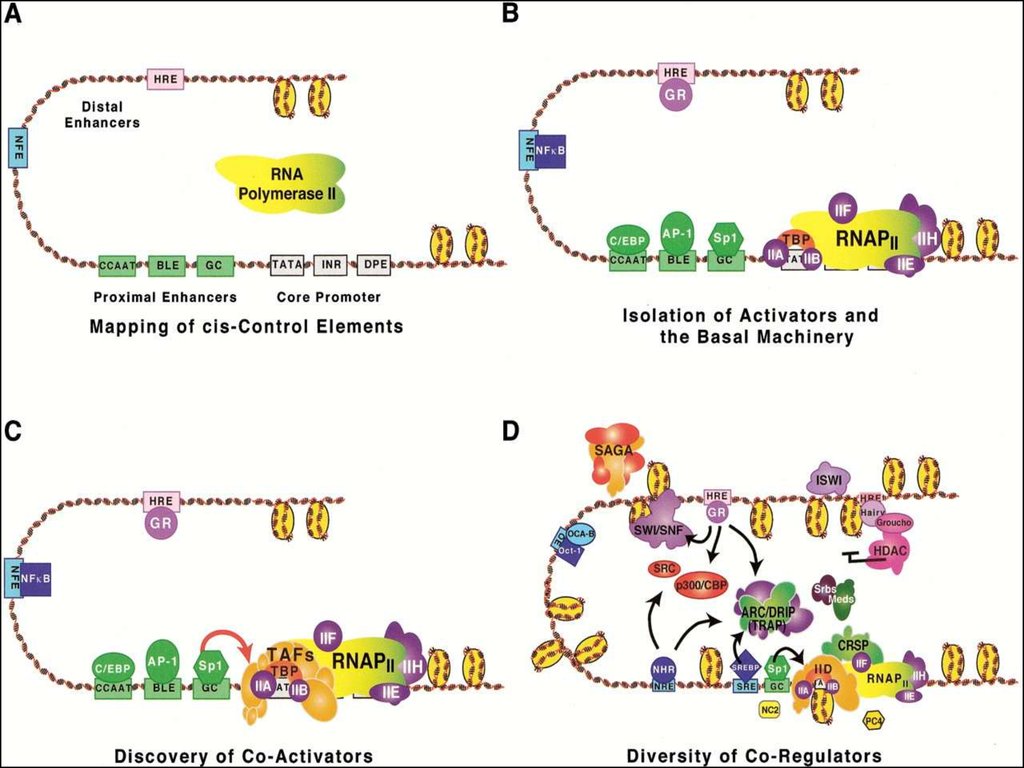
8. Факторы транскрипции эукариот
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ9.
10. Ядерные РНК-полимеразы
РНК-полимераза IРНК-полимераза II
РНК-полимераза III
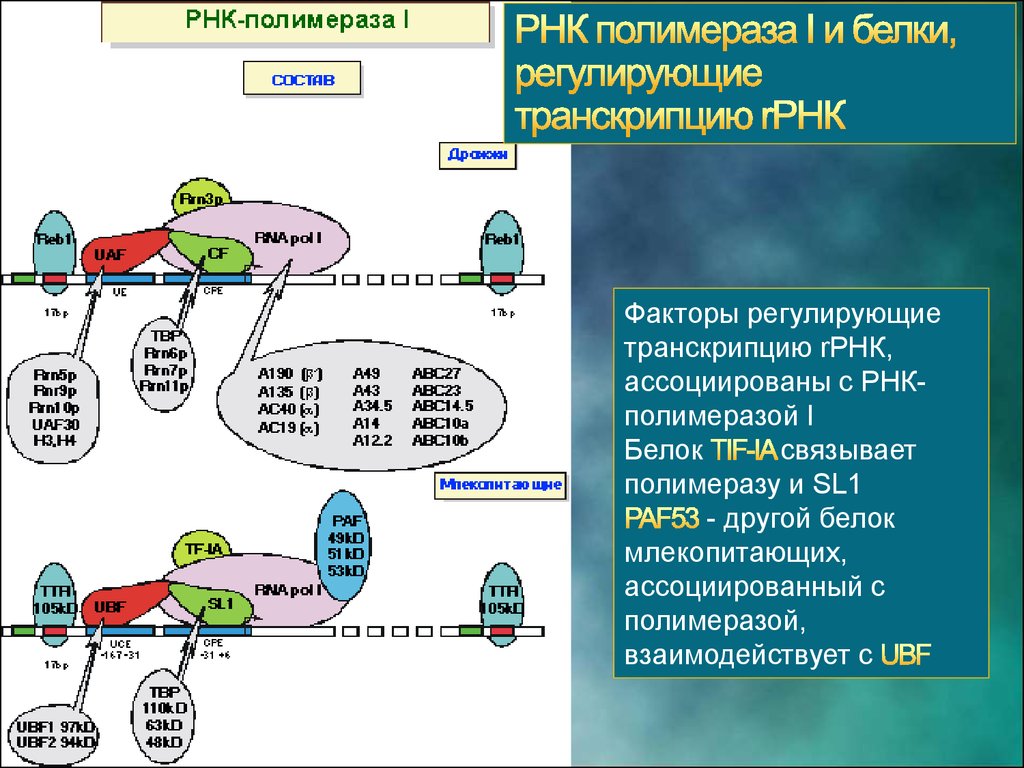
11. РНК-полимераза I
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ12. РНК-полимераза I. Функции
РНК полимераза I транскрибирует гены 18S, 5.8S и 28SТранскрипция генов rРНК и сборка рибосом происходит в
ядрышке
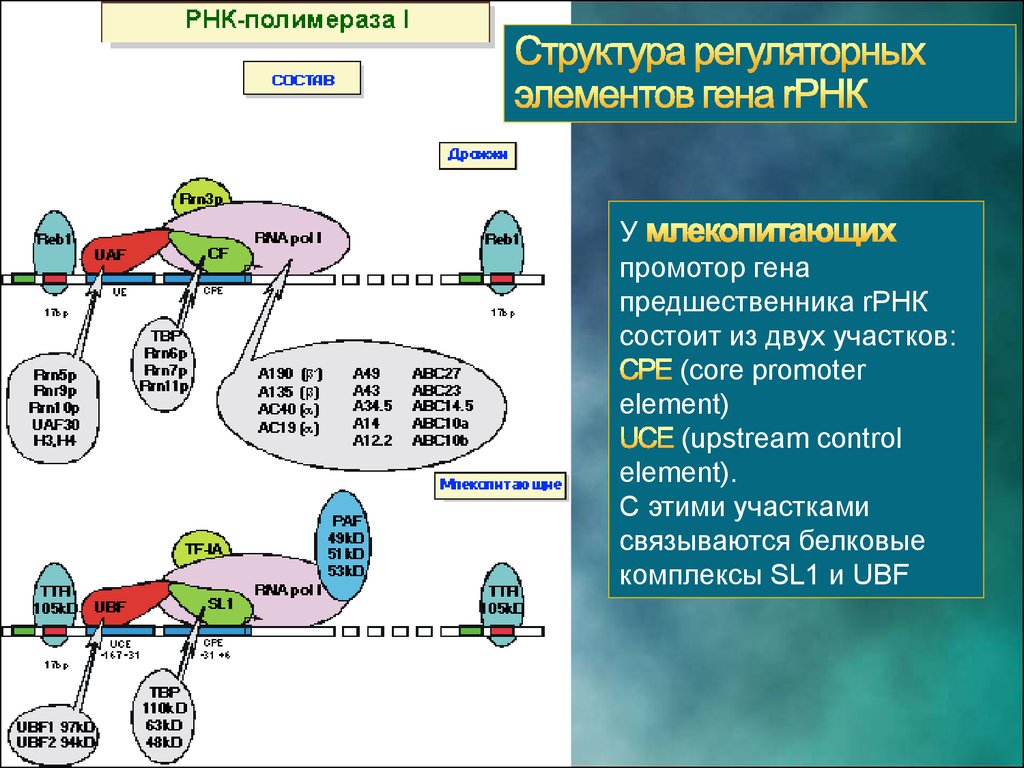
13.
Упромотор гена
предшественника rРНК
состоит из двух участков:
(core promoter
element)
(upstream control
element).
С этими участками
связываются белковые
комплексы SL1 и UBF
14.
В состав SL1 входитбелок
(TATA-box
binding protein), хотя в
промоторе rРНК нет ТАТА
последовательности.
В SL1 входят также три
дополнительных белка,
называемых
,
и
.
15.
Факторы регулирующиетранскрипцию rРНК,
ассоциированы с РНКполимеразой I
Белок
связывает
полимеразу и SL1
- другой белок
млекопитающих,
ассоциированный с
полимеразой,
взаимодействует с
16.
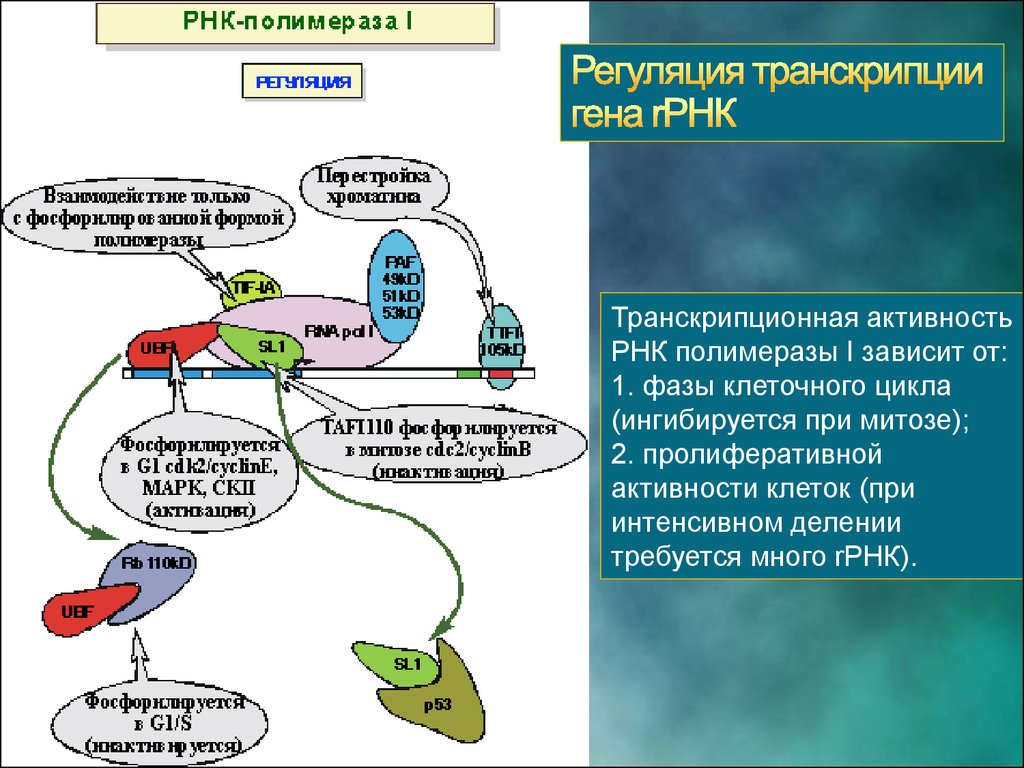
Транскрипционная активностьРНК полимеразы I зависит от:
1. фазы клеточного цикла
(ингибируется при митозе);
2. пролиферативной
активности клеток (при
интенсивном делении
требуется много rРНК).
17.
РНК-полимераза I должнабыть фосфорилирована для
успешного взаимодействия с
18.
Активность основныхфакторов инициации
(
и
) регулируется в
зависимости от фазы
клеточного цикла:
1.
инактивируется в
митозе при помощи
фосфорилирования.
2.
активируется
фосфорилированием в G1
фазе.
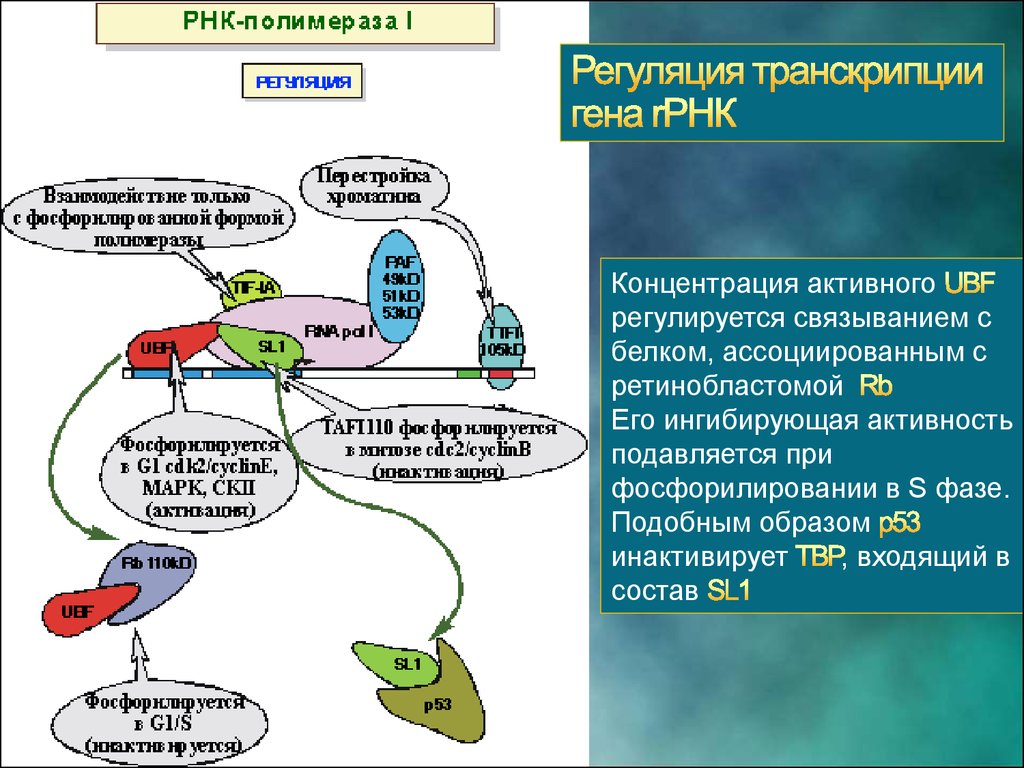
19.
Концентрация активногорегулируется связыванием с
белком, ассоциированным с
ретинобластомой
Его ингибирующая активность
подавляется при
фосфорилировании в S фазе.
Подобным образом
инактивирует
, входящий в
состав
20.
Белокспособствует
передвижению нуклеосом по
ДНК для эффективной
транскрипции РНК
полимеразой I.
21. РНК-полимераза III
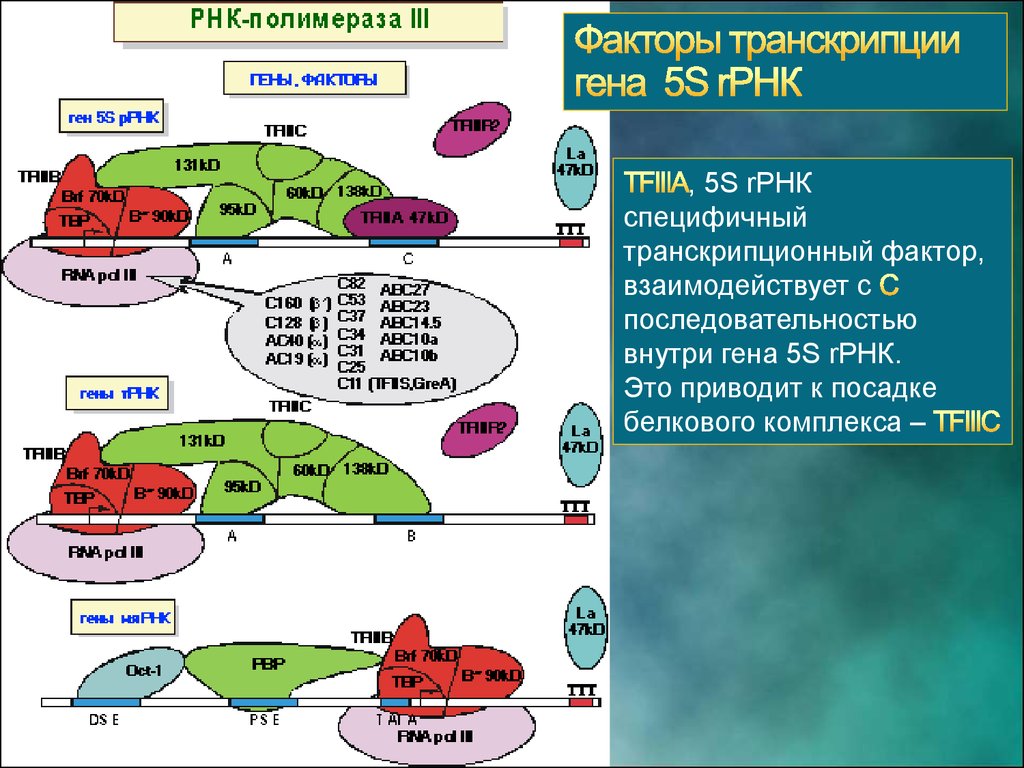
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ22. РНК-полимераза III. Функции.
РНК полимераза III транскрибирует гены 5S,
и
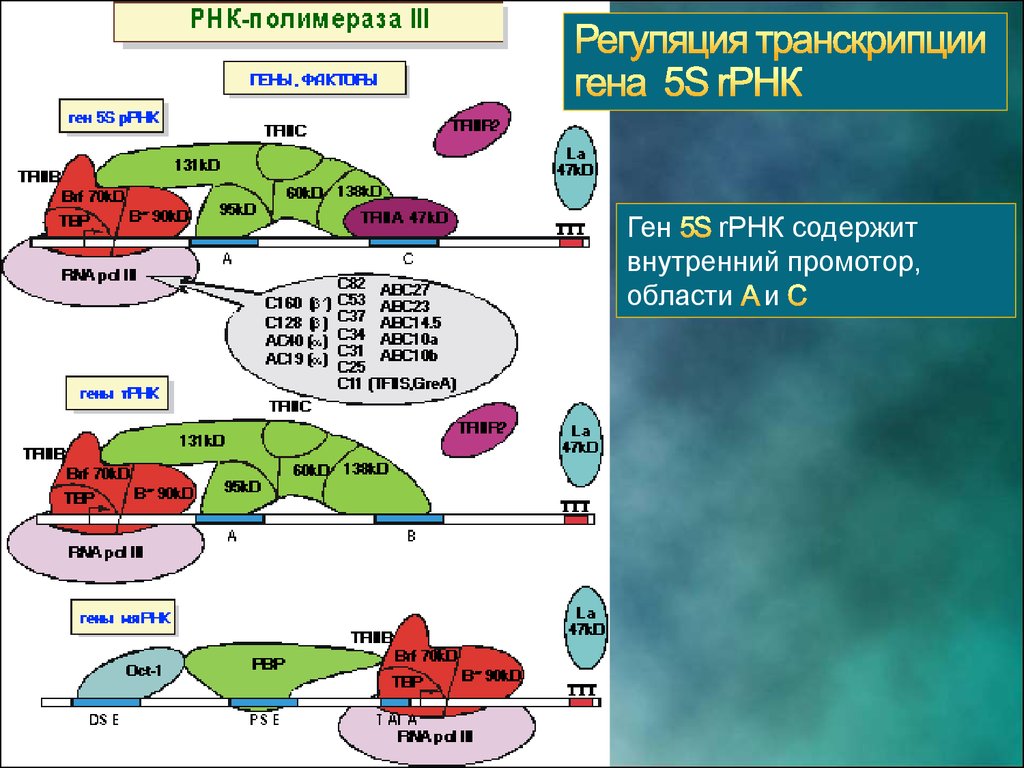
23.
ГенrРНК содержит
внутренний промотор,
области и
24.
, 5S rРНКспецифичный
транскрипционный фактор,
взаимодействует с
последовательностью
внутри гена 5S rРНК.
Это приводит к посадке
белкового комплекса –
25.
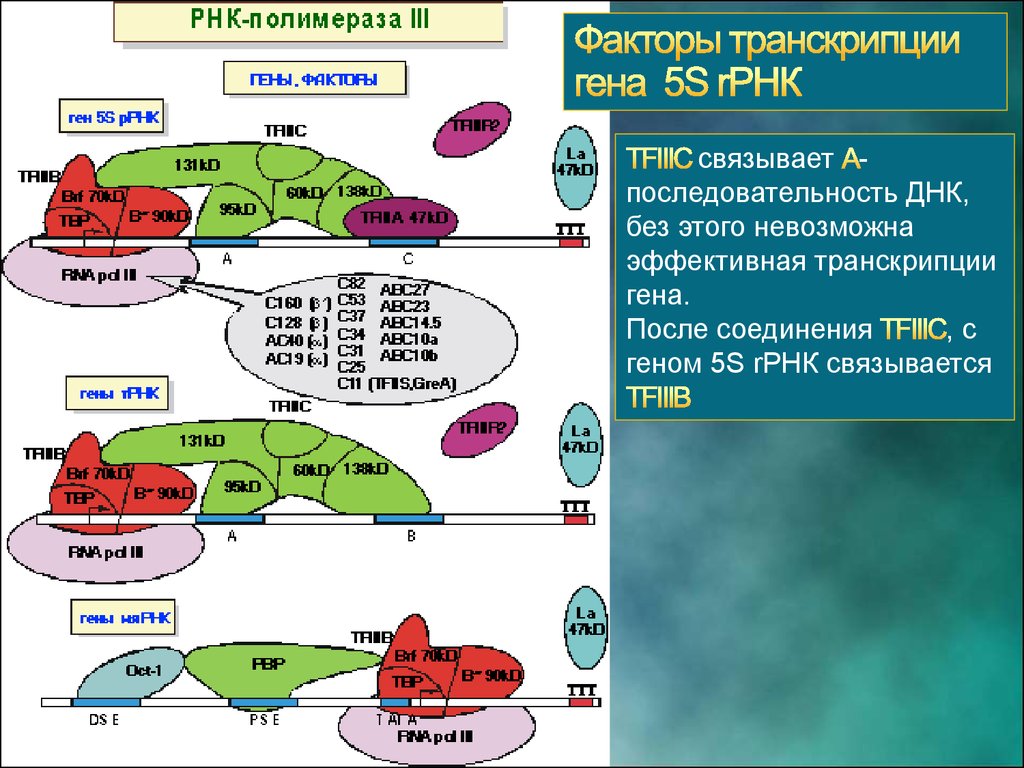
связывает последовательность ДНК,без этого невозможна
эффективная транскрипции
гена.
После соединения
,с
геном 5S rРНК связывается
26.
В составвходит
фактор способствует
присоединению РНК
полимеразы III и началу
транскрипции
27.
Генысодержат
дополнительный участок
ДНК – , способствующий
связыванию
.
Белок-белковое
взаимодействие
и
заменяется ДНКбелковым.
Далее,
присоединяет
и
28.
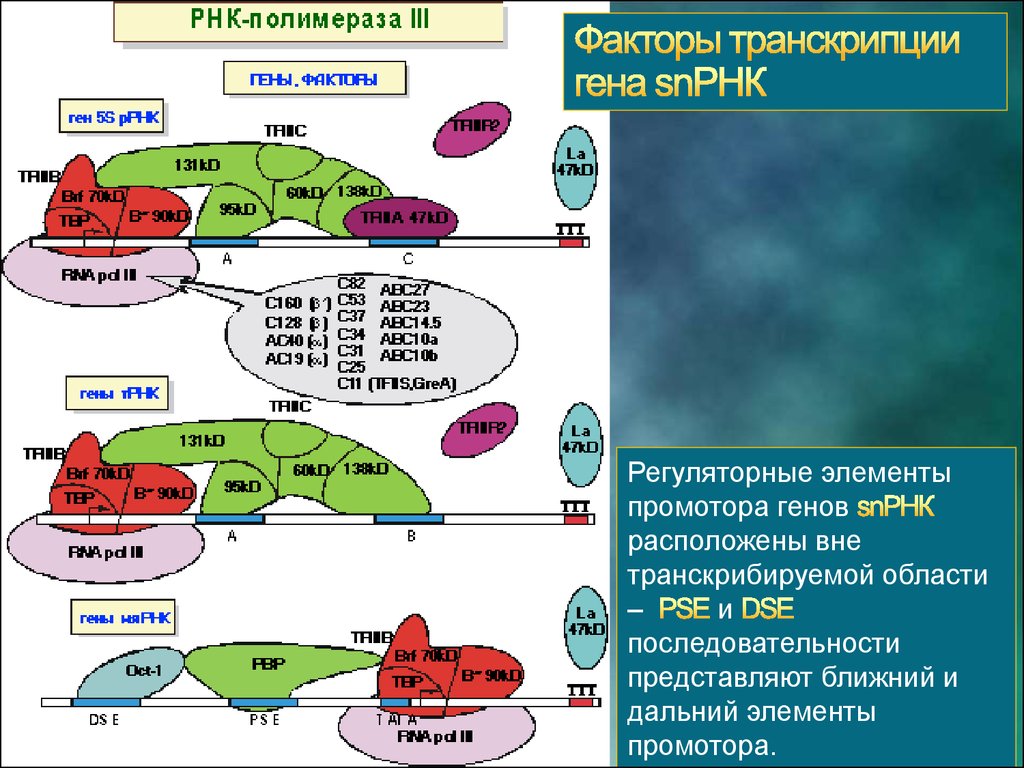
Регуляторные элементыпромотора генов
расположены вне
транскрибируемой области
–
и
последовательности
представляют ближний и
дальний элементы
промотора.
29.
Ссвязывается белок
С
– фактор
.
Связывание
и
приводит к присоединению
30.
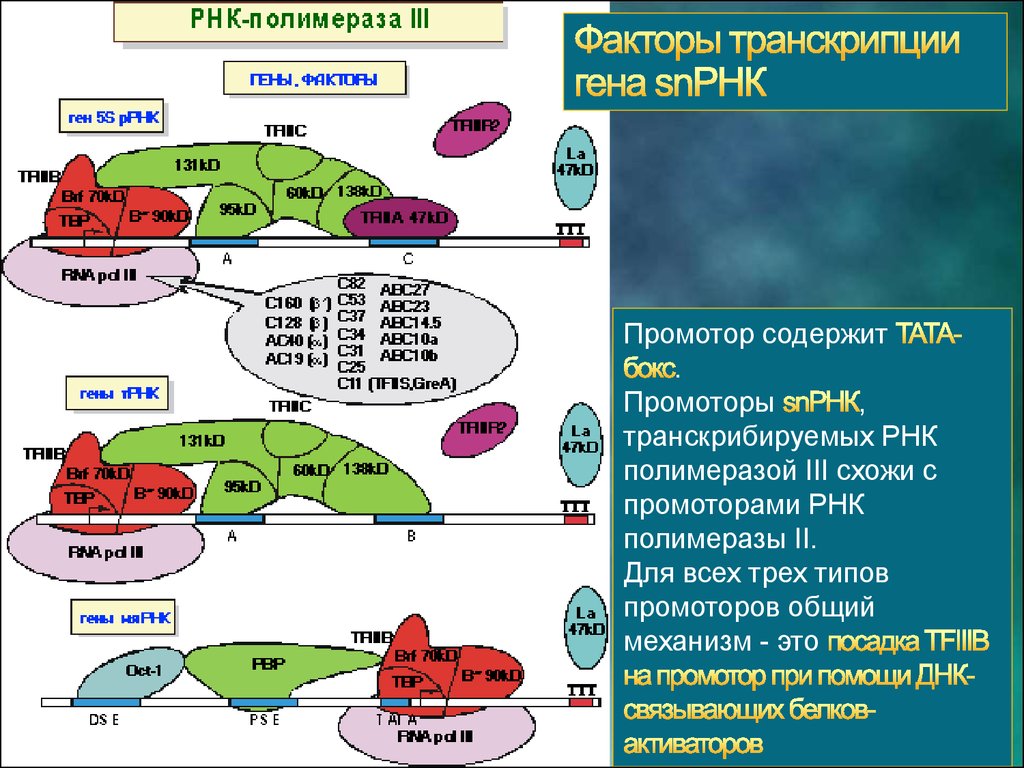
Промотор содержит.
Промоторы
,
транскрибируемых РНК
полимеразой III схожи с
промоторами РНК
полимеразы II.
Для всех трех типов
промоторов общий
механизм - это
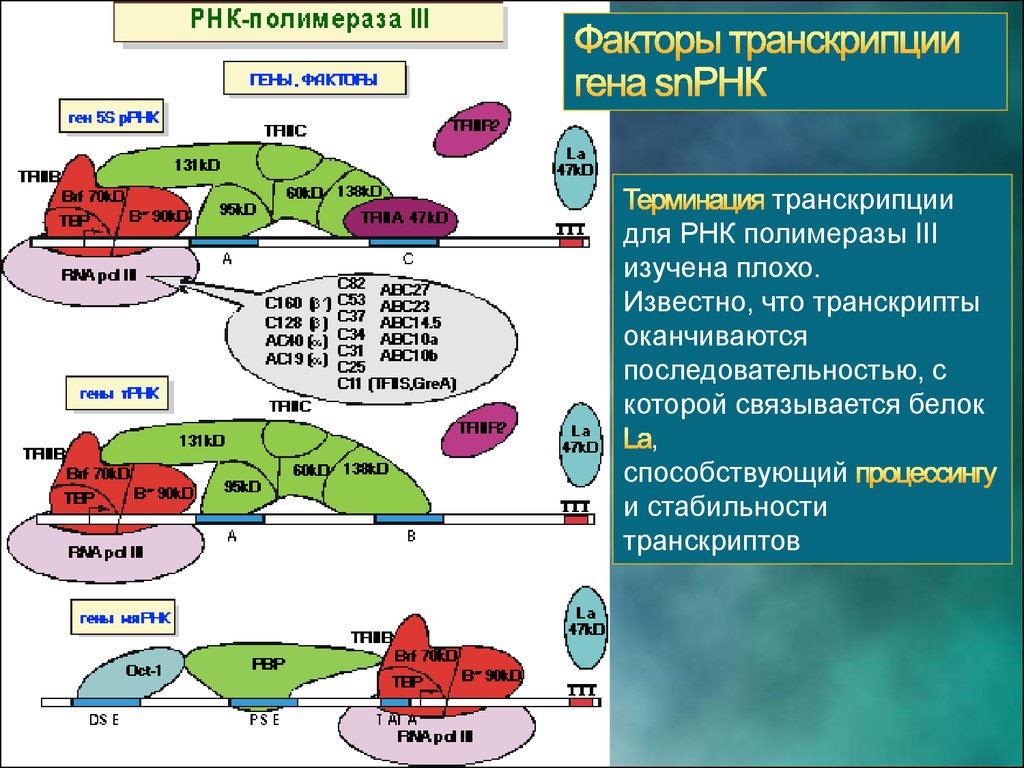
31.
транскрипциидля РНК полимеразы III
изучена плохо.
Известно, что транскрипты
оканчиваются
последовательностью, с
которой связывается белок
,
способствующий
и стабильности
транскриптов
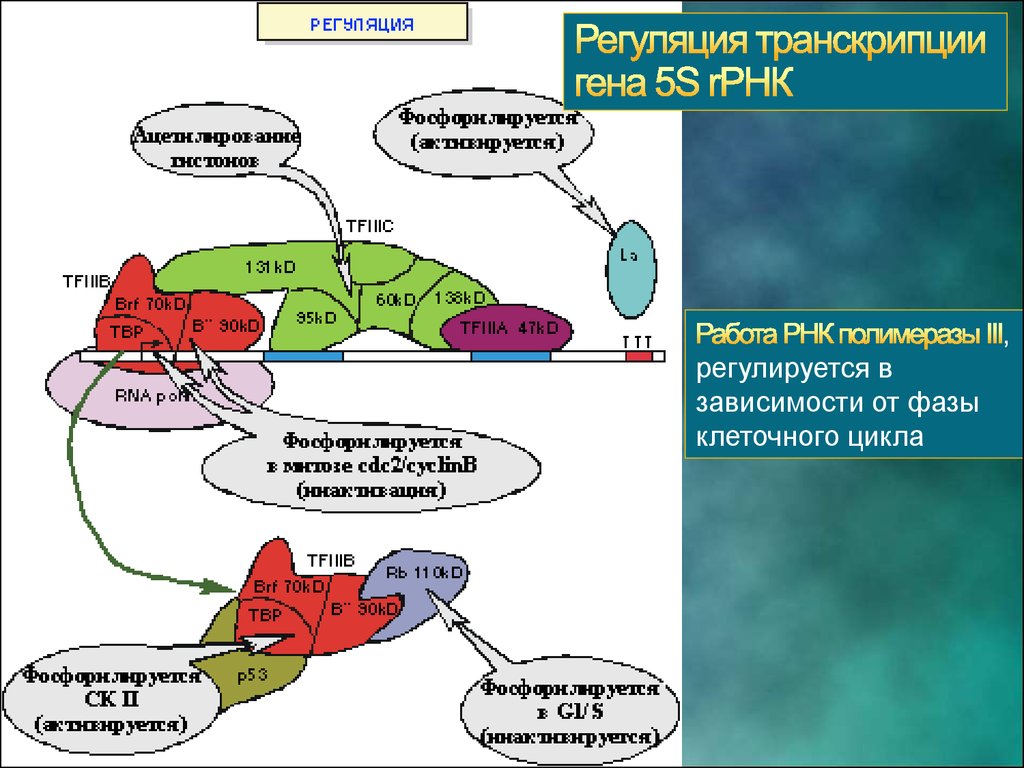
32.
,регулируется в
зависимости от фазы
клеточного цикла
33.
направлено наподавление
транскрипции во время
митоза.
La белок активируется
при фосфорилировании
34.
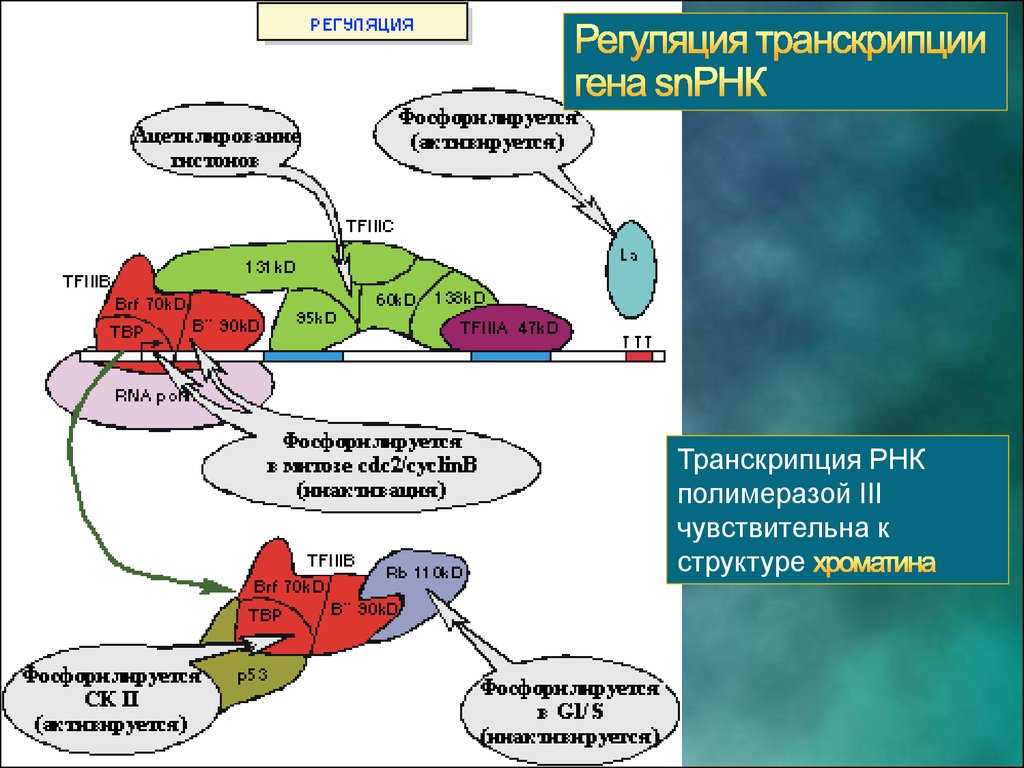
Транскрипция РНКполимеразой III
чувствительна к
структуре
35.
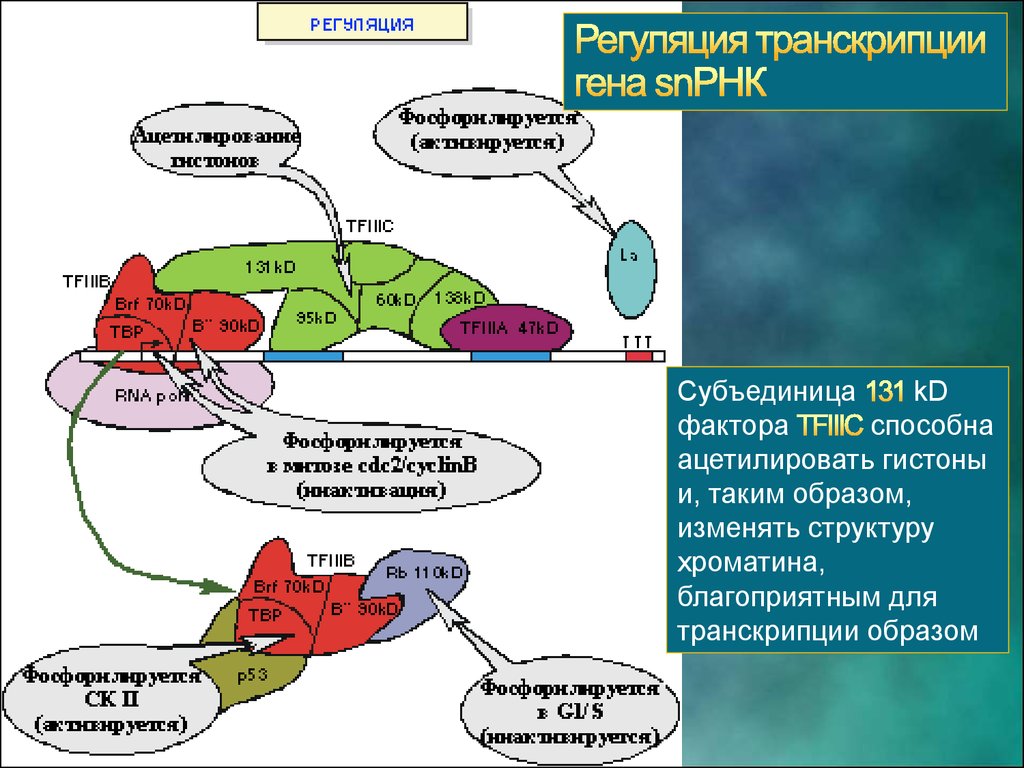
СубъединицаkD
фактора
способна
ацетилировать гистоны
и, таким образом,
изменять структуру
хроматина,
благоприятным для
транскрипции образом
36. РНК-полимераза II
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ37. РНК-полимераза II. Функции
транскрибирует, а также некоторые
наиболее подверженный регуляции фермент из трех РНК
полимераз
состоит из 17 субъединиц, одной из особенностей строения
наибольшей субчастицы РНК полимеразы II является ее Сконцевой домен
, построенный из повторяющихся
гептамеров. Количество этих повторов варьирует для разных
организмов, составляя в среднем несколько десятков.
Последовательное укорочение С-концевого домена приводит
к гибели клеток.
38.
39.
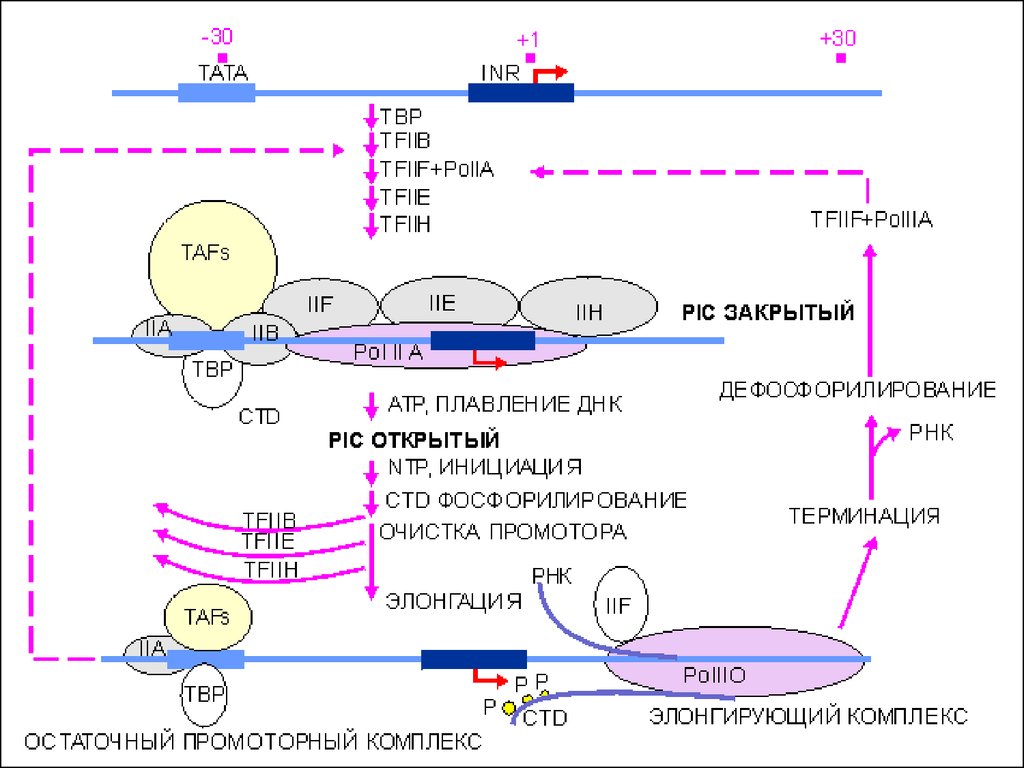
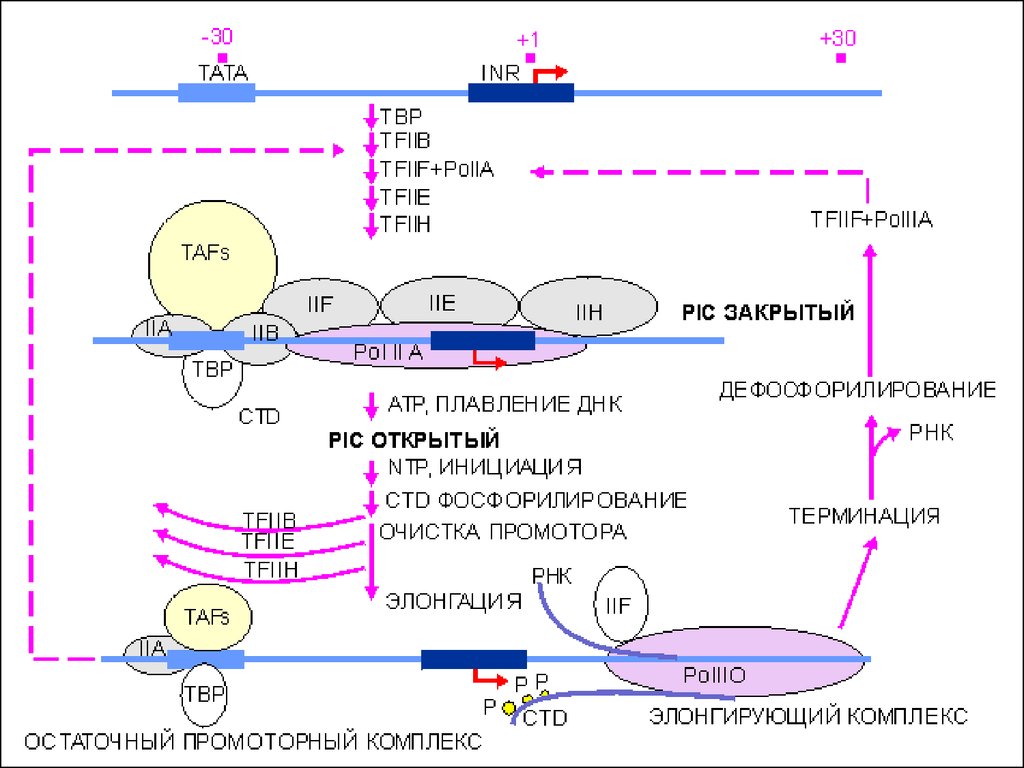
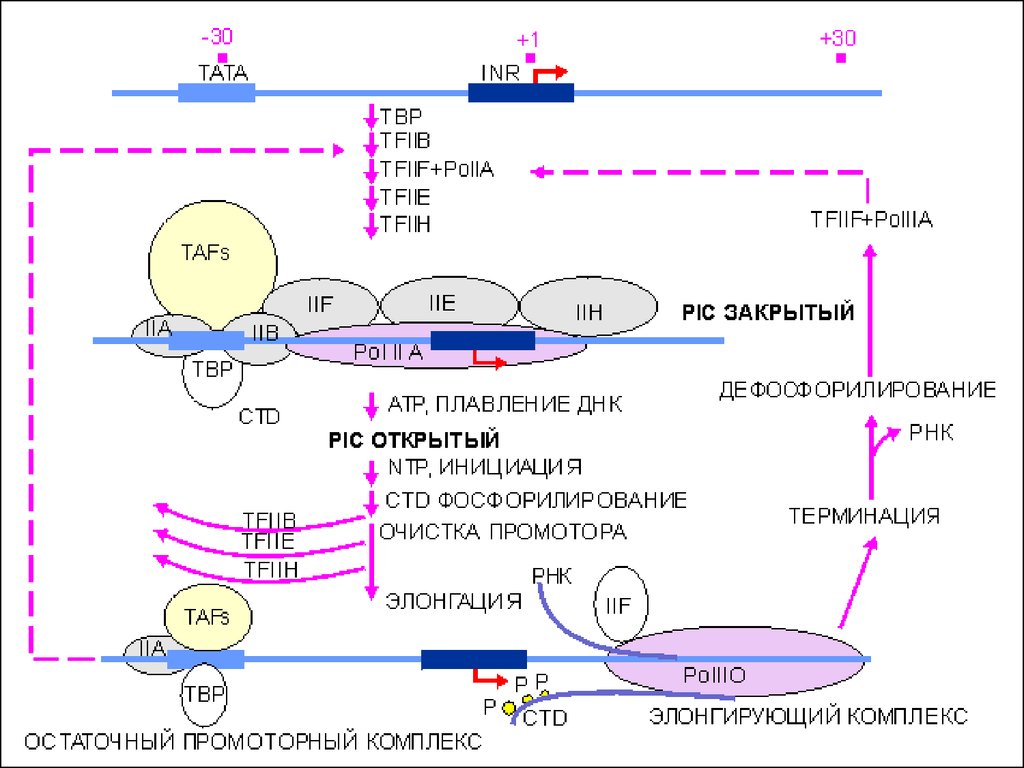
40. Основные стадии транскрипции
Инициация• Образование базального комплекса
• Начало синтеза цепи РНК
Элонгация
• Продолжение процесса транскрипции,
рост дочерней цепи РНК
Терминация
• Завершение процесса транскрипции
41.
42.
43. Инициация транскрипции «в комиксах»
ДНКК цепи подходит
фактор
транскрипции TFIID
самый крупный
фактор транскрипции.
Желтый участок -
44.
45. Инициация транскрипции «в комиксах»
связывается сбоксом и располагается
рядом с сайтом
инициации
Это присоединение
вызывает формирование
ДНК
46. Инициация транскрипции + Pol II «в комиксах»
Затем подходятфакторы
(оранжевый) и
(красный)
присоединяется к
рядом с
подходит к
.
собирается
дистанционно
47. Инициация транскрипции + Pol II «в комиксах»
связывается си
и
способствует
присоединению
собран и
подходит к точке
инициации
транскрипции
К сформировавшемуся
комплексу подходит
фактор
48. Инициация транскрипции + Pol II «в комиксах»
TFIIETFIIH
связывается,
подходит
Все готово и для
инициации
транскрипции
требуется энергия
49.
50. Элонгация транскрипции «в комиксах»
В двухцепочечной ДНКформируется «
»,
обеспечивающий доступ
Pol II к единице
транскрипции и
начинается синтез mРНК
Начинается элонгация
цепи и факторы
,
,
,
отсоединяются
действует
дистанционно и
mРНК быстро
растет
51. Элонгация транскрипции «в комиксах»
Элонгациязавершается когда
достигается конец
транскрипта
Закрывается «
в ДНК
»
Pol II освобождается
вместе с
полученными от АТФ
фосфатами
52. Терминация транскрипции «в комиксах»
Комплекс Pol II отходит,mРНК освобождается.
Фактор TFIID отходит,
позволяя ДНК вернутся в
.
Первичный
транскрипт готов и
после процессинга
будет
транспортирован из
ядра клетки
53.
54.
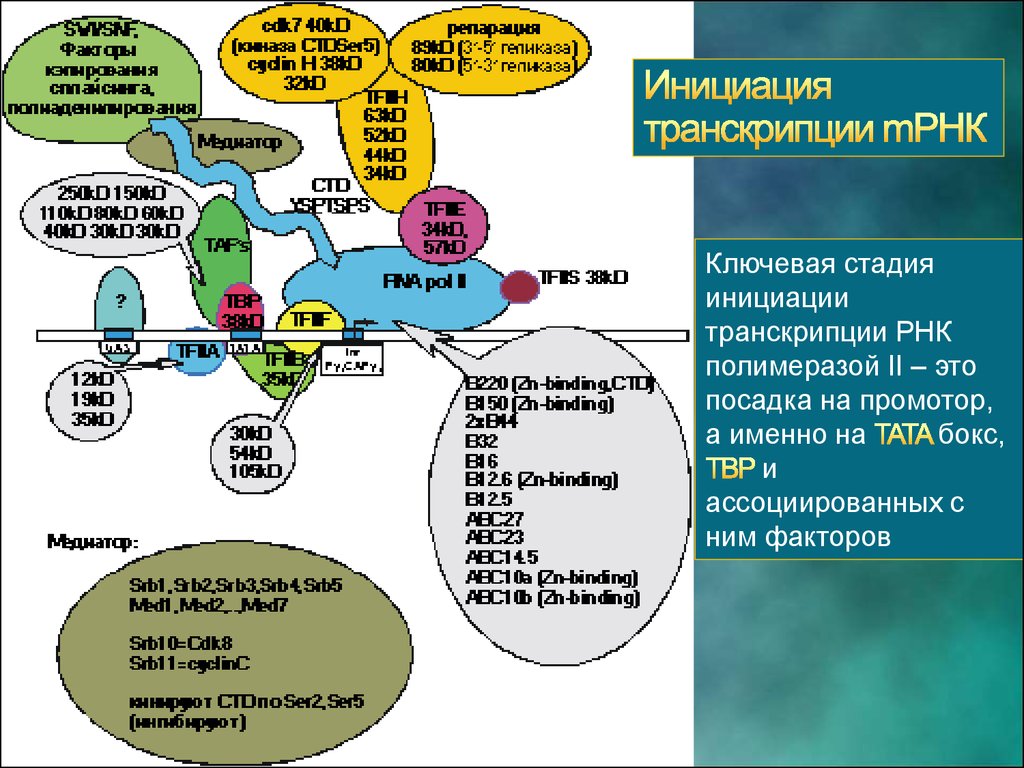
Ключевая стадияинициации
транскрипции РНК
полимеразой II – это
посадка на промотор,
а именно на
бокс,
и
ассоциированных с
ним факторов
55.
Большая субъединицабелка
фосфорилируется.
Эта субъединица
является ATPзависимой ДНК
56.
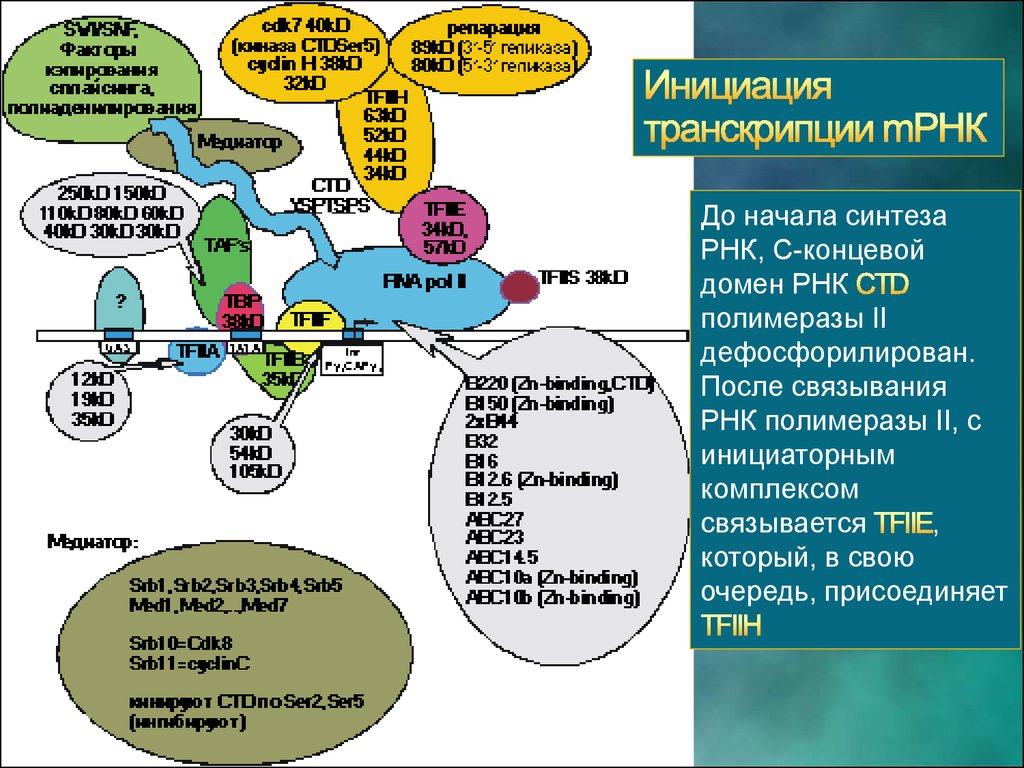
До начала синтезаРНК, С-концевой
домен РНК
полимеразы II
дефосфорилирован.
После связывания
РНК полимеразы II, с
инициаторным
комплексом
связывается
,
который, в свою
очередь, присоединяет
57.
- комплекс,важный для репарации
и транскрипции.
В его состав входят
, локально
расплетающие ДНК в
двух направлениях (5’
и 3’) и
РНК
полимеразы
58.
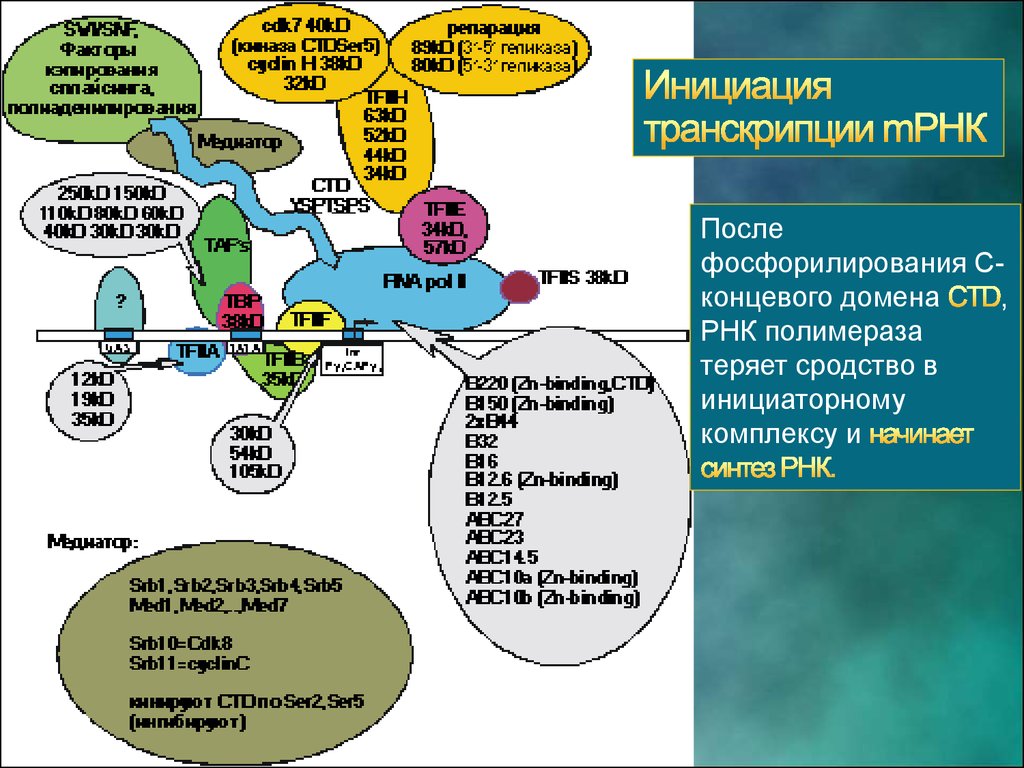
Послефосфорилирования Сконцевого домена
,
РНК полимераза
теряет сродство в
инициаторному
комплексу и
59.
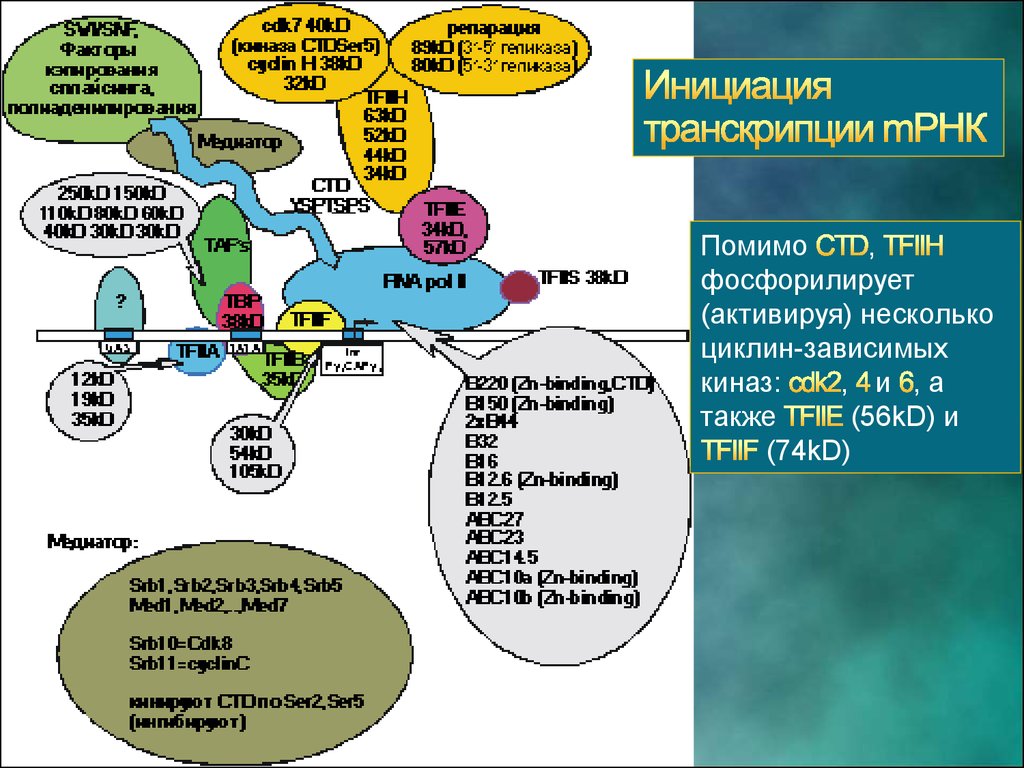
Помимо,
фосфорилирует
(активируя) несколько
циклин-зависимых
киназ:
, и ,а
также
(56kD) и
(74kD)
60.
Кроме описанных факторов в инициации транскрипцииРНК полимеразой II участвует множество других.
Они обеспечивают регуляцию транскрипции различных
генов.
61.
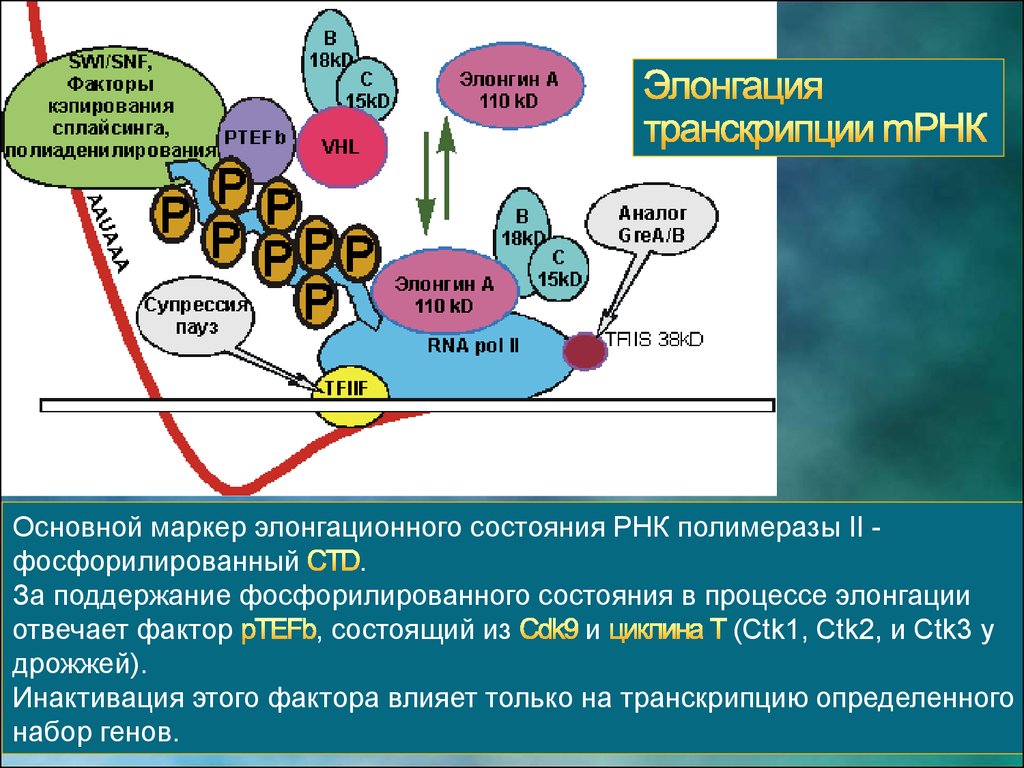
Основной маркер элонгационного состояния РНК полимеразы II фосфорилированный.
За поддержание фосфорилированного состояния в процессе элонгации
отвечает фактор
, состоящий из
и
(Ctk1, Ctk2, и Ctk3 у
дрожжей).
Инактивация этого фактора влияет только на транскрипцию определенного
набор генов.
62.
Кроме, фактором элонгации РНК полимеразы II является
гетеротример
.
Негативный регулятор этого фактора –
. Он связывает комплекс
и и предотвращает их ассоциацию с
63.
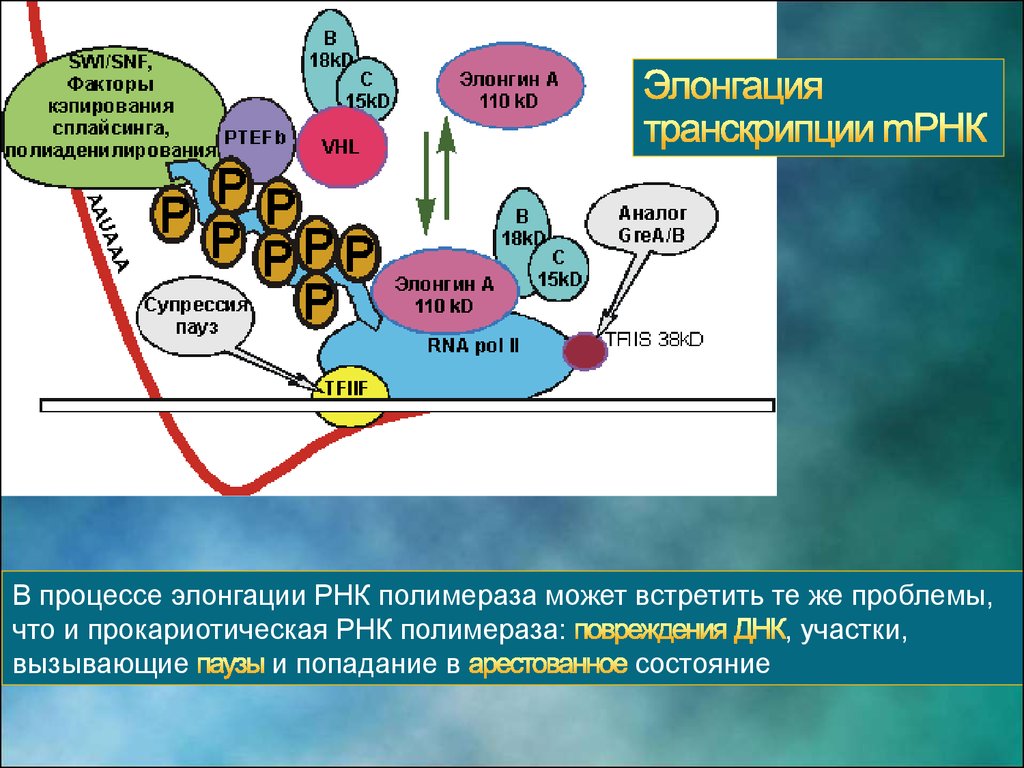
В процессе элонгации РНК полимераза может встретить те же проблемы,что и прокариотическая РНК полимераза:
, участки,
вызывающие
и попадание в
состояние
64.
Если работающая РНК-полимераза встретит повреждение ДНК,нуждающееся в репарации, то с таким остановленным комплексом
связываются белки
и
.
Эти белки опосредуют посадку факторов репарации на повреждение, а
также играют важную роль для восстановления транскрипции после
репарации.
65.
За супрессию пауз РНК-полимеразы отвечаетарестованного состояния –
.
, а за выход из
66.
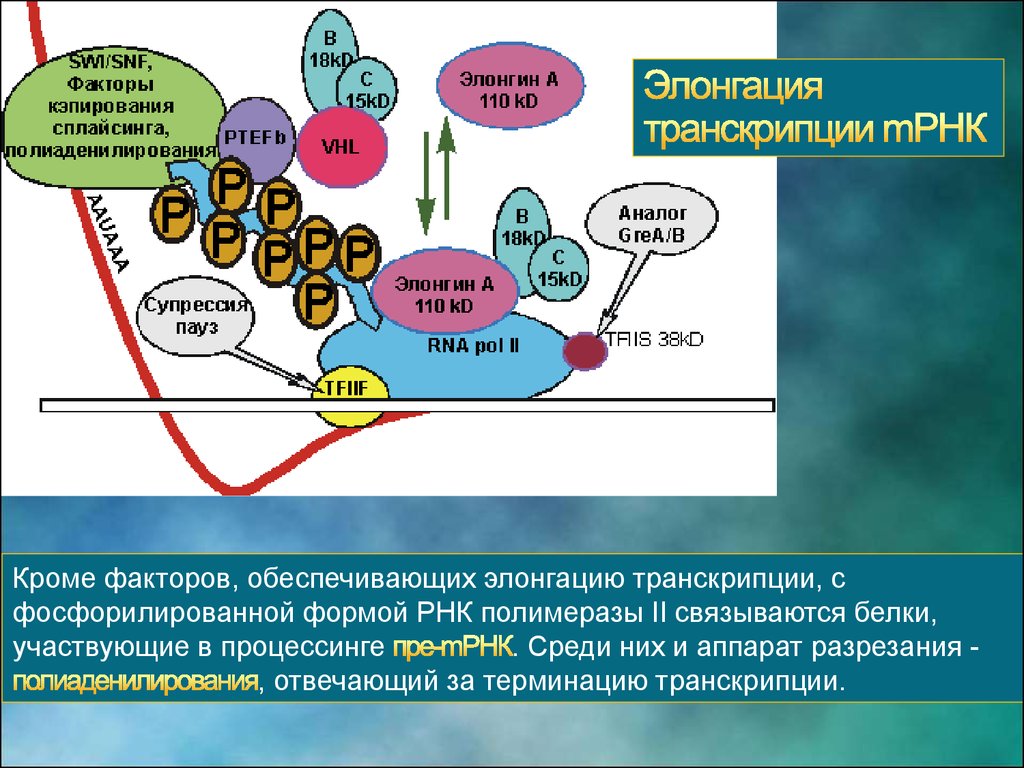
Кроме факторов, обеспечивающих элонгацию транскрипции, сфосфорилированной формой РНК полимеразы II связываются белки,
участвующие в процессинге
. Среди них и аппарат разрезания , отвечающий за терминацию транскрипции.
67. Регуляция транскрипции
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. ТРАНСКРИПЦИЯ РНК ЭУКАРИОТ68. Некодирующие регуляторные элементы
Геном животных и человека – сложнейшая система смногоуровневой системой регуляции.
Важную роль в процессах регуляции играют некодирующие
последовательности:
,
,
. Они
определяют уровень транскрипции генов.
69. Энхансеры. Структура.
- это последовательности ДНК, формирующие модули.Модули - это отдельные части энхансеров.
Например, М1+М2+М3+М4 - один энхансер, но он состоит из 4-х
модулей.
Все 4 модуля узнаются своими белками, а они, сидя на ДНК,
взаимодействуют друг с другом.
Если в клетке присутствуют все соответствующие белки, то участку ДНК
придается определенная конформация и начинается синтез РНК.
70. Энхансеры: «петлевая модель» действия
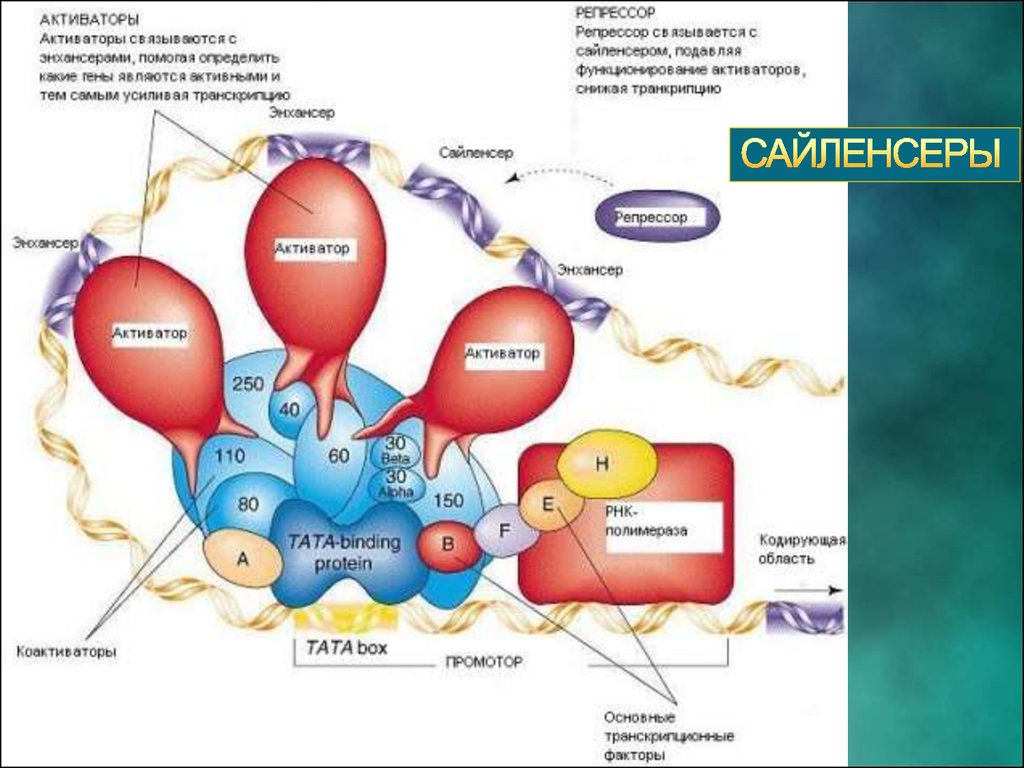
(Activators) - белки,связывающиеся с энхансерами,
которые помогают РНКполимеразе правильно начать
транскрипцию.
(Repressor)- белки,
которые связывают активаторы,
чем снижают или прекращают
транскрипцию.
(Basal factors)белки, которые ориентируют РНКполимеразу на начало
структурной части гена.
(или Pribnow box)- часть
промотора, являющаяся сайтом
связывания для белковых
факторов.
(Transcription factors)- помогают
занять правильную позицию
активаторам и РНК- полимеразе.
71.
72. Инсуляторы (англ. insulate - изолировать )
Последовательности нуклеотидов, которые могут подавлятьпозитивное и негативное влияние эухроматина и гетерохроматина на
экспрессию генов, интегрированных в этот хроматин и
фланкированных указанными последовательностями в новом сайте
интеграции.
Такие участки ДНК как бы изолируют ген, находящийся между ними,
способствуя сохранению его обычной пространственной структуры.
73. Механизм действия инсуляторов
А Схема функционирования двух генов (желтый и голубой),локализованных в домене, контролируемом двумя инсуляторами
(зеленый) и белками, которые связываются с инсуляторами (желтый).
Энхансеры локализуются между двумя генами (вместе с
транскрипционными факторами, обозначенными бирюзовым и
оранжевым), они могут активировать транскрипцию с промотора каждого
гена.
74. Механизм действия инсуляторов
В Если пограничный элемент, инсулятор (красный), расположен междудвумя энхансерами, формируется новый хромосомный домен, в
результате голубой ген остается в одном, а темно-желтый – в другом
домене.
Бирюзовый транскрипционный фактор не может действовать на промотор
темно-желтого гена в соответствии с локализацией второго инсулятора.
Энхансер может активировать транскрипцию с промотора голубого гена,
который остался в том же домене.