
 под действием группового (А) и индивидуального (В) отбора")


")












 альтруизм")

















 biology
biologySimilar presentations:


")




")


Групповой отбор vs индивидуальный отбор
1. Групповой отбор vs индивидуальный отбор
2. Изменение состава популяции (по признаку альтруистического поведения) под действием группового (А) и индивидуального (В) отбора
Остались только «альтруисты»Не осталось «альтруистов»
3. Каким же образом распространяются признаки выгодные для группы и не выгодные для особи?
• Английский генетик истатистик Рональд Э.
Фишер (1890-1962)
первым попытался
ответить на этот вопрос на
примере
предупреждающей
окраски.
4.
5. Устойчивая эволюционная или эволюционно-стабильная стратегия (ЭСС)
• Что это такое? ЭСС – это такая стратегия, прикоторой в популяции, где почти все особи
действуют в соответствии с ЭСС,
отклоняющаяся стратегия не получает
преимущества.
• Предположим, есть самцы «ястребы» и
«голуби». Выигрыш - +50, но проигрыш
ястреба - –100, проигрыш от потери времени
при столкновении голубя с голубем - –10, а с
ястребом - 0.
6.
• Ситуация 1. В популяции одни «голуби».Выигрыш +50–10= +40; проигрыш = –10.
Средний итог равен +15. Если появляется
ястреб он все время выигрывает (+50).
• Ситуация 2. В популяции одни «ястребы».
Выигрыш = +50; проигрыш = –100.
Средний итог равен –25. Если появляется
голубь, он все время убегает, т.е. его очки
всегда равны 0, но больше, чем у ястребов, и
он оказывается в преимуществе.
• Равновесие устанавливается при соотношении
голуби/ ястребы, как 5:7. При этом средний
успех 6,25 и для голубей, и для ястребов.
7.
• Заметим, что это не групповой отбор,если бы все были бы «голубями», то
выигрыш популяции (15 очк./ос.) был бы
большим.
• Будет ли устойчива система (5 г – 7 я) по
отношению к другим стратегиям?
• Стратегия «отпорщика». Если ястреб –
как ястреб, если голубь – как голубь,
если отпорщик – тоже как голубь
• Стратегия «задира»: как ястреб до
отпора, а тогда как голубь
8.
• Из всех четырех стратегий только «отпорщик»устойчив. Почему?
• «Голуби» неустойчивы против «ястребов», «задир».
• Популяцию «ястребов» бы заполонили «голуби» и
«задиры».
• Популяцию «задир» бы наводнили бы «ястребы».
• Ни одна стратегия не может вытеснить «отпорщиков»,
но «отпорщики» не могут вытеснить «голубей».
• То есть, если по случайным причинам возрастает
число «голубей», сразу получат другие стратегии
преимущество.
9.
10.
11.
12.
13.
• Joop Jukema, Theunis Piersma Permanent female mimics in a lekkingshorebird // Biol. Lett. 22 June 2006 vol. 2 no. 2 161-164
14. Альтруизм на примере общественных перепончатокрылых
15.
• УильямГамильтон 1964.
Hamilton W.D. 1964.
The genetical
evolution of social
behaviour // J. of
Theor. Biol. 7 p.152
1936-2000
16.
• Итоговая (результирующая, совокупная )приспособленность inclusive fitness
• Дельта ИП= (Na-na)ra+Σ(Nr-nr)rr
Na – число детей альтруиста
na– число детей альтруиста при эгоист. пов
Nr – число детей родственника альтруиста при
поддержке
nr – число детей родственника альтруиста без
поддержки
r – коэффициенты родства
17.
• Гаплодиплодия (самки –диплойдны,самцы – гаплойдны). Коэффициент
родства с дочерью 1/2 , а с сестрой – 3/4
• Организм – это лишь средство,
инструмент распространения генов.
18. Реципрокный (взаимный) альтруизм
• Trivers R.L. 1971. Theevolution of reciprocal
altruism. // Quarterly
Rev. of Biol. 46 p. 3557
• Роберт Трайверс
19.
• Допустим у нас есть вид птиц, которыйпоражен клещами. Они могут снять их
отовсюду кроме головы.
• «Простак» снимает клещей с любого
• «Плут» дает снимать с себя, но сам не снимает
никогда
• Простаки неумолимо вымирают, и плуты в
наказание страдают пораженные клещами.
• То есть плуты – ЭСС.
• Но пусть у нас есть еще одна стратегия –
«злопамятные».
• При низкой численности – не ЭСС, но выше
критической – ЭСС.
20. Признаки реципрокного альтруизма
• Особи не родственныдруг другу.
• Выигрыш существенно
больше потери.
• Необходимо
индивидуальное
опознавание.
21. Родительский вклад
• Trivers R.L. 1972. Parental investment andsexual selection // Sexual Selection and the
Descent of Man (ed. B. Campbell )
Chicago, Aldine p. 136-179.
• Любой вклад родителя в потомка,
повышающий шансы выживания за счет
возможностей вклад в другого потомка
22.
• Вклад ограничен (число отложенныхяиц, собранного корма)
• Должен ли он распределяться
равномерно или неравномерно?
• Если условия жесткие, то поддерживают
старших, если мягкие то младших. В
какой-то момент прерывает вклад
(оставляя для других), но если
последний потомок, может продолжать.
23. Могут ли потомки манипулировать вкладом?
• Trivers R.L. 1974. Parent – offspring conflict //American Zoologist 14 p. 249-264.
• Родители, дети и дети между собой имеют
коэффициент родства 1/2, но потомок по
отношению к самому себе 1.
• Мать стремится кормить до тех пор пока вклад
не достигнет среднего вклада будущего
потомка, а детеныш до тех пор пока проигрыш
будущих потомков не будет выше его
удвоенного выигрыша.
• 20 видов муравьев 3:1, рабовладельцы 1:1
24. Существует ли конфликт между полами?
• Каждый пол заинтересован сэкономить свойвклад для того, чтобы вложить его в
следующие поколения. Вклад самки
изначально больше – самки заботятся чаще.
• Какова должна быть стратегия покинутой
самки? «Обмануть» другого самца, «убедив»
что он отец.
• Поэтому естественный отбор поддерживает
«инфантицид» у самцов. (Брюс-эффект,
поведение львов)
• Или другая стратегия – бросить самца раньше.
25.
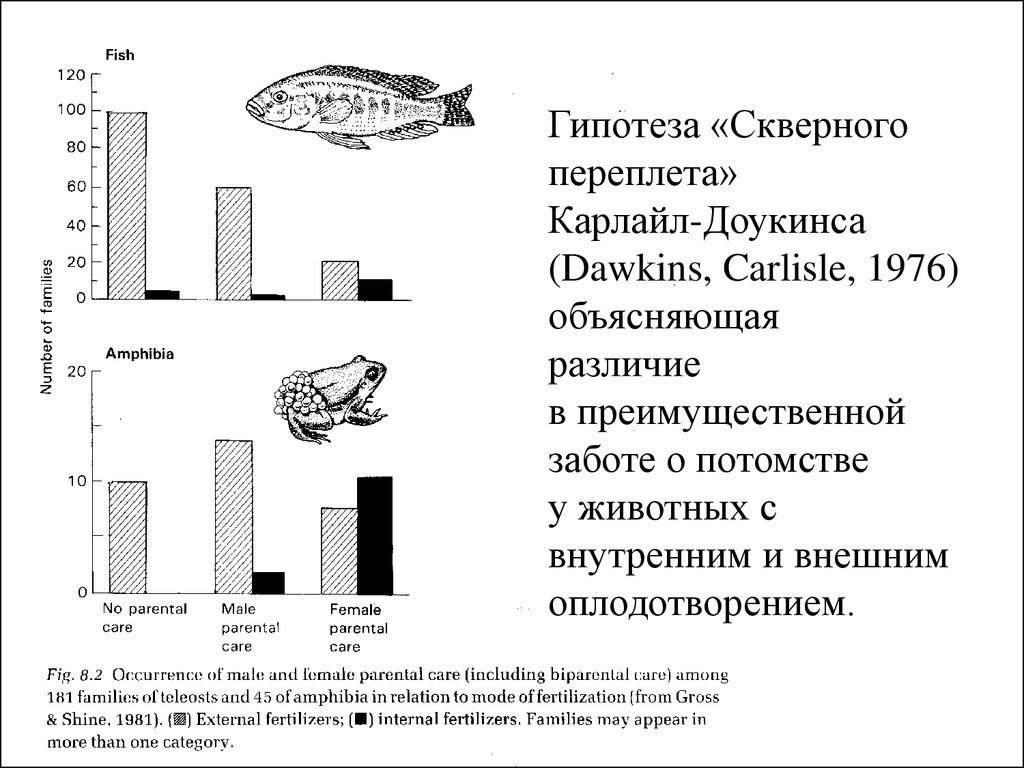
Гипотеза «Скверногопереплета»
Карлайл-Доукинса
(Dawkins, Carlisle, 1976)
объясняющая
различие
в преимущественной
заботе о потомстве
у животных с
внутренним и внешним
оплодотворением.
26.
• Таким образом, самка заинтересовананайти самца, который будет вкладывать в
потомство
• Самка «изучает» самцов. Один из
способов – это долгое ухаживание. С
другой стороны самец уверен в
отцовстве. Это т.н. стратегия «домашнего
уюта» (domestic-bliss strategy).
• По Трайверсу вклад самца привязывает к
самке (но для этого важно, чтобы все
самки себя вели также).
27.
• Генетический выигрыш за успешновыращенного детеныша +15
• Расходы на одного детеныша – 20.
• Длительное ухаживание – 3.
28.
• Если мы введем 4 стратегии:• «скромница» не спаривается, пока
самец не предпринял долгого
ухаживания,
• «распутница» быстро соглашается
спариваться с любым самцом,
• «верный» готов к длительному
ухаживанию,
• «гуляка» не ухаживает долго, а
отправляется искать других самок.
29.
• Ситуация 1. Все самки скромницы, все самцыверные супруги.
Средний выигрыш 15 – 10 – 3=+2
• Ситуация 2. Появилась распутница.
Ее выигрыш 15 – 10=+5
• Ситуация 3. В популяции распутниц
появляется гуляка.
Гуляка в выигрыше +15,
распутница
проигрывает 15–20= –5 скромницы получают
преимущество,
число гуляк начинает
сокращаться
• Точка стабильности – 5/6 скромницы, 5/8
верные
30. Связь приспособленности и соотношения полов потомства
• Trivers R.L.,Willard D.E.,
1973. Natural
selection of
parental ability
to vary the sex
ratio of
offspring //
Science 179, p.
90-92.
31.
Эдвард Уилсон,Гарвардский Университет
1975
32.
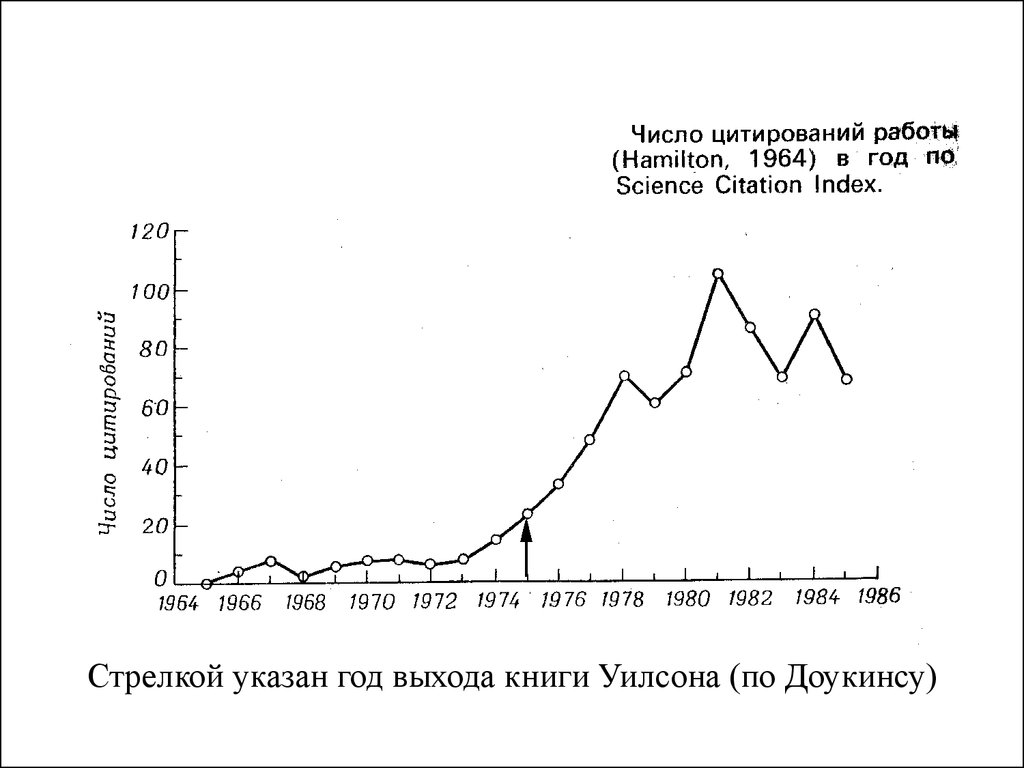
Стрелкой указан год выхода книги Уилсона (по Доукинсу)33. Какие существуют возражения социобиологическому подходу
1. Обратный ход времени (будущийрепродуктивный успех)
• 2. Противоречия с генетикой поведения
(Селекция генов, а не селекция особей)
• 3. Существует противоречие между
распределением тех факторов и ответа, то есть
почему у остальных не так.
• 4. «Нефальсифицируемость». Наблюдения
Бертрама 1976 за эгоистическим поведением
молодых львов.
34. Ответы на возражения
• Речь идет не об эволюции, а об устойчивости• Работа Ротенбьюлера (Rothenbuhler, 1964) по
санитарному поведению пчел. Заболевание
«гнильца пчел», выбрасывание зараженных
личинок определяется двумя генами (?).
• Например, гаплодиплоидия распространена
шире – но можно предположить, что она
возникла позже, это механизм устойчивости, а
не эволюции.
• Далеко все теории фальсифицируемы, иногда
полезны те теории, которые хоть мобилизуют
на исследования.
35. Возможен ли групповой отбор? Парадокс Симпсона
p — доля альтруистов, w — численность популяции.John S. Chuang, Olivier Rivoire, Stanislas Leibler. Simpson’s Paradox in a Synthetic Microbial System // Science.
2009. V. 323. P. 272–275.
36. Межгрупповая конкуренция способствует внутригрупповой кооперации
«Внутригрупповые перетягивания каната (tag-of-war) вложены вмежгрупповые». Члены группы соревнуются за свою долю общественного
пирога. Размер пирога зависит от успешности группы в соревновании с другими
группами. Чем больше сил тратят особи на внутригрупповую борьбу, тем
меньше их остается на общественно-полезную деятельность.
Анализ этой модели показал, что межгрупповая конкуренция может быть
мощным стимулом для развития внутригрупповой кооперации даже при
низком уровне внутригруппового родства.
Reeve H.K., Hölldobler B. The emergence of a superorganism through intergroup competition // PNAS, 2007.
37. Какова альтернатива?
• Генотип (гены) не могут контролироватьповедение в непрерывном режиме. Системе
нужны внутренние положительные стимулы.
«Бунт фенотипа».
• Суть социального поведения в формировании
связей или привязанностей. Объект
концентрации внимания или атрактор
организует поведение. (Структура внимания,
бюджет времени, пространственные
отношения).
• Нет изначальной полезности
• Социальность, как эволюционный фактор