














 biology
biologySimilar presentations:
 клетки")






 в медицинской практике. Проблемы и последствия")


Перепрограммирование геномов дифференцированных клеток с использованием экзогенной экспрессии транскрипционных факторов
1.
Перепрограммирование геномовдифференцированных клеток
с использованием экзогенной экспрессии
транскрипционных факторов
2.
Способы восстановления потенциала соматических клеток(Yamanaka 2007)
3.
Экспериментальные подходы восстановленияпотенций в клетках утративших их в процессе
дифференцировки
Перенос ядер в энуклеированный ооцит или яйцеклетку – технология
клонирования животных.
1952 – Briggs and King; 1997 - Wilmut et al., 1997;
Слияние эмбриональных стволовых клеток с дифференцированными –
технология гибридных клеток.
1996 – Матвеева и др.
Трансфекция дифференцированных клеток векторами экспрессирующие
транскрипционные факторы и факторы регулирующие пролиферацию.
2006 – Takahashi and Yamanaka
4.
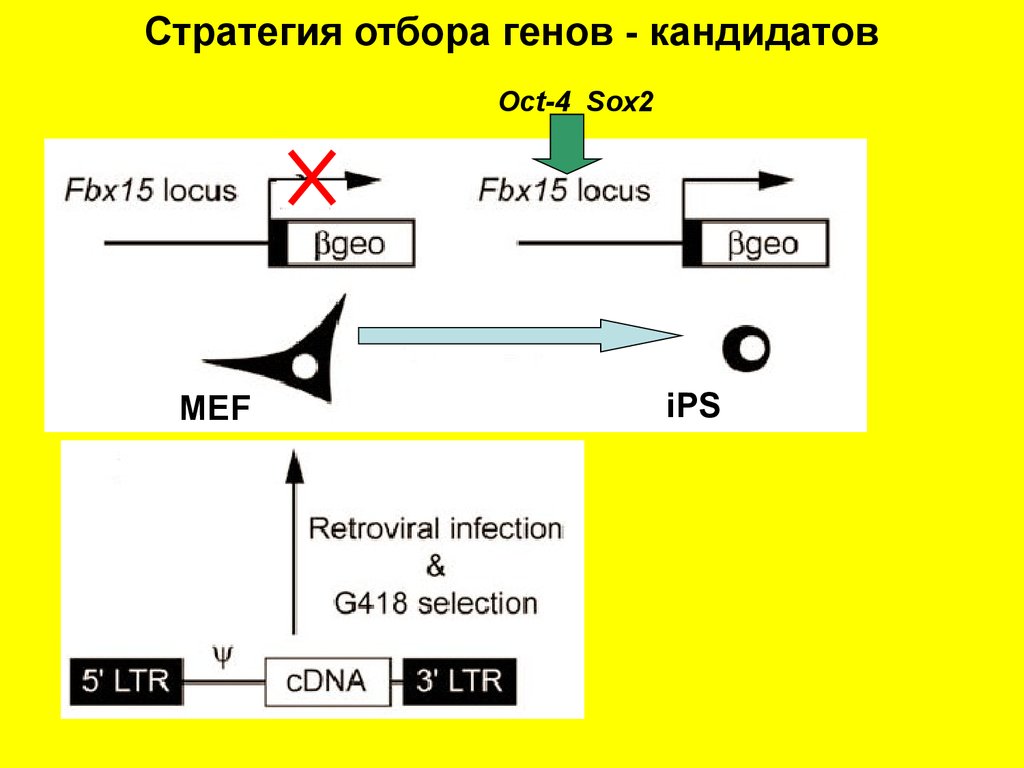
Стратегия отбора генов - кандидатовOct-4 Sox2
MEF
iPS
5.
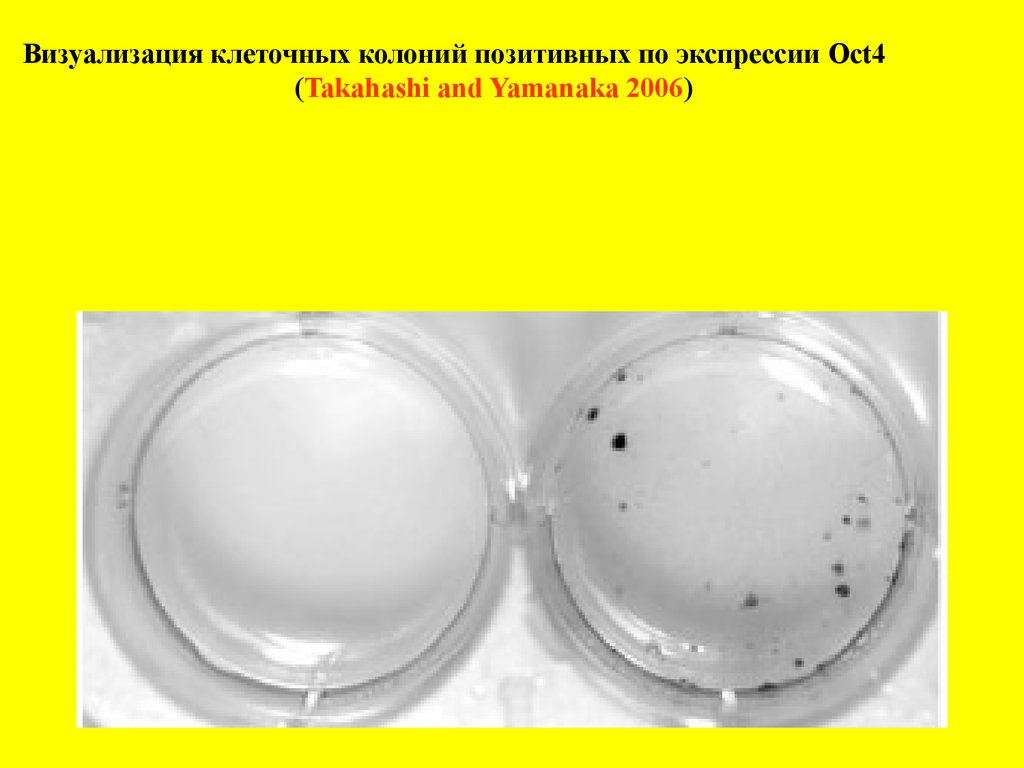
Визуализация клеточных колоний позитивных по экспрессии Oct4(Takahashi and Yamanaka 2006)
6.
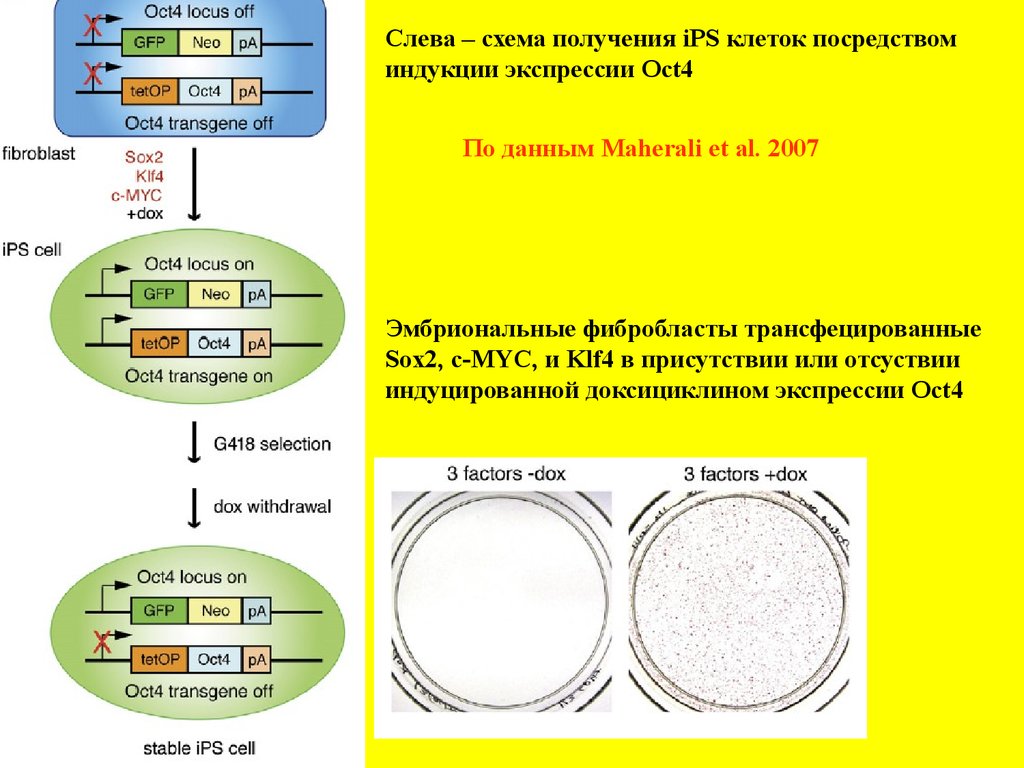
Слева – схема получения iPS клеток посредствоминдукции экспрессии Oct4
По данным Maherali et al. 2007
Эмбриональные фибробласты трансфецированные
Sox2, c-MYC, и Klf4 в присутствии или отсуствии
индуцированной доксициклином экспрессии Oct4
7.
8.
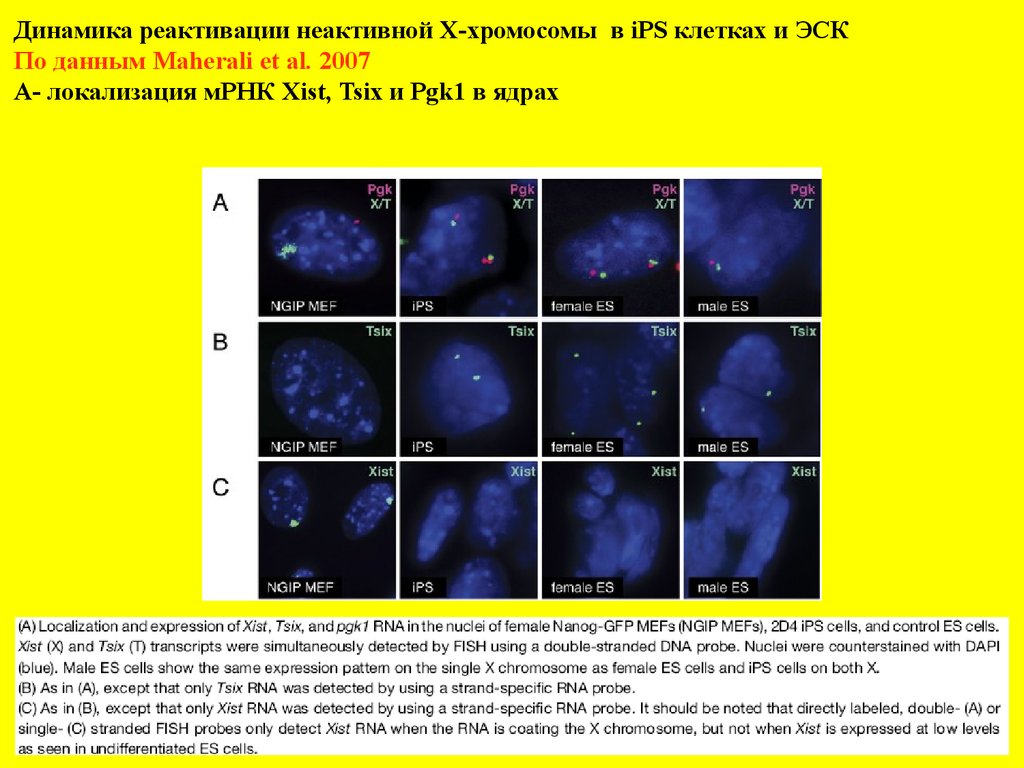
Динамика реактивации неактивной Х-хромосомы в iPS клетках и ЭСКПо данным Maherali et al. 2007
А- локализация мРНК Xist, Tsix и Pgk1 в ядрах
9.
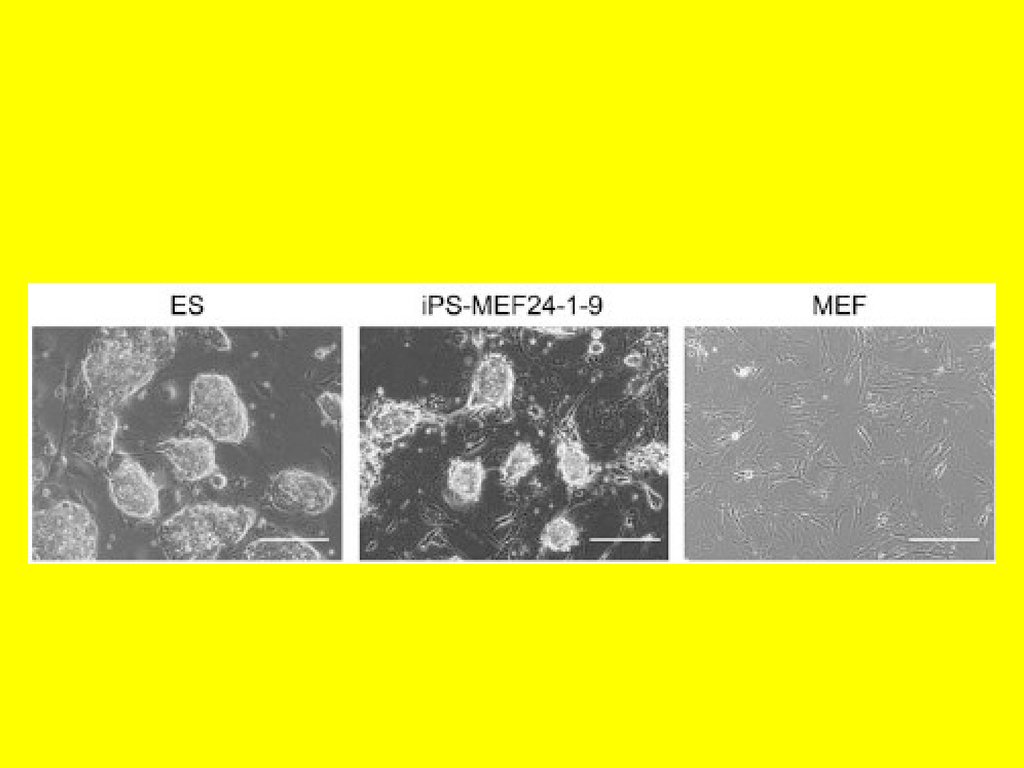
Динамика образования колоний клеток, включая iPS (Wernig et al., 2007)10.
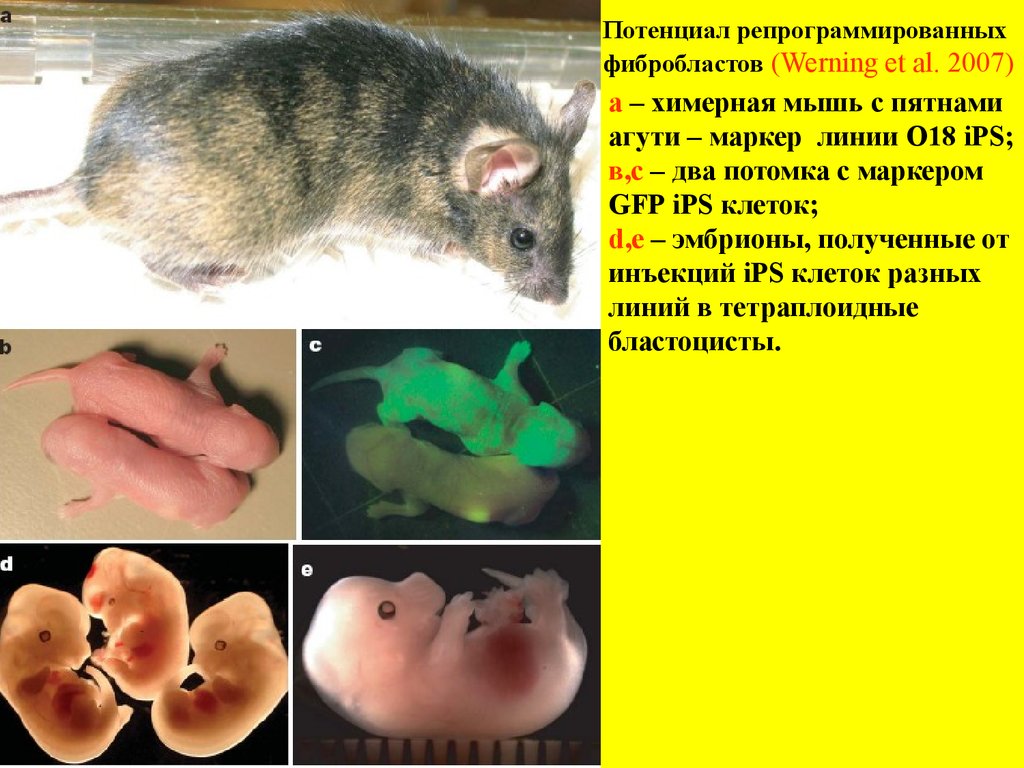
Потенциал репрограммированныхфибробластов (Werning et al. 2007)
а – химерная мышь с пятнами
агути – маркер линии O18 iPS;
в,с – два потомка с маркером
GFP iPS клеток;
d,e – эмбрионы, полученные от
инъекций iPS клеток разных
линий в тетраплоидные
бластоцисты.
11.
Идентификация генотипов iPS средипотомков F1 в скрещиваниях
между химерными самцами с
мышами C57BL.
Окраска потомков F2 в
скрещивании iPS мышей с C57BL
(Okita et al., 2007)
12.
Роль 4-х факторов в превращении фибробластов в эмбриональныестволовые клетки (Yamanaka 2007).
ЭС клетки иммортальны и имеют активный открытый хроматин. с-Myc
индуцирует эти важные свойства в фибробластах, Klf4 препятствует
апоптозу, а Oct4 вместе с Sox2 необходимы для поддержания
плюрипотентности.
13.
Конструкция лентивирусного вектора в экспериментах порепрограммированию соматических клеток человека (Yu et al., 2007)
14.
Список тестированных генов по их способности трансформироватьсоматические клетки человека в IPS клетки (Yu et al., 2007)
15.
Оптимизация репрограммирования мезенхимальных клеток человека, полученныхпосле дифференцировки из ЭСК человека, маркированных GFP и NeoR (Yu et al.,
20 November 2007)
16.
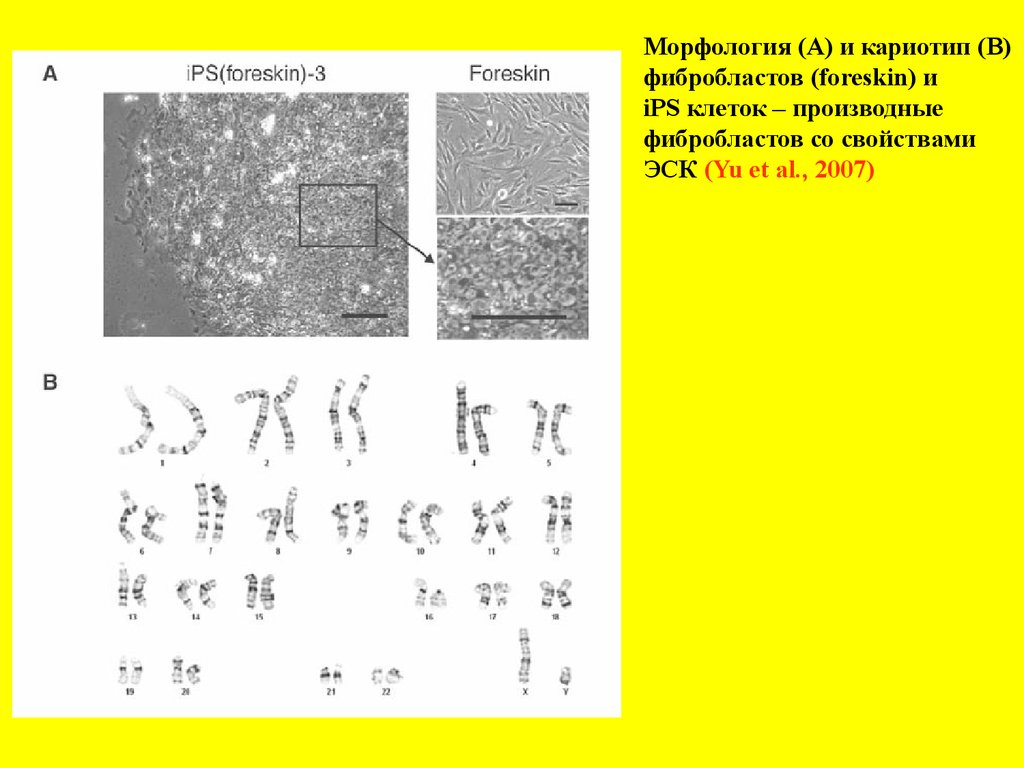
Морфология (А) и кариотип (В)фибробластов (foreskin) и
iPS клеток – производные
фибробластов со свойствами
ЭСК (Yu et al., 2007)
17.
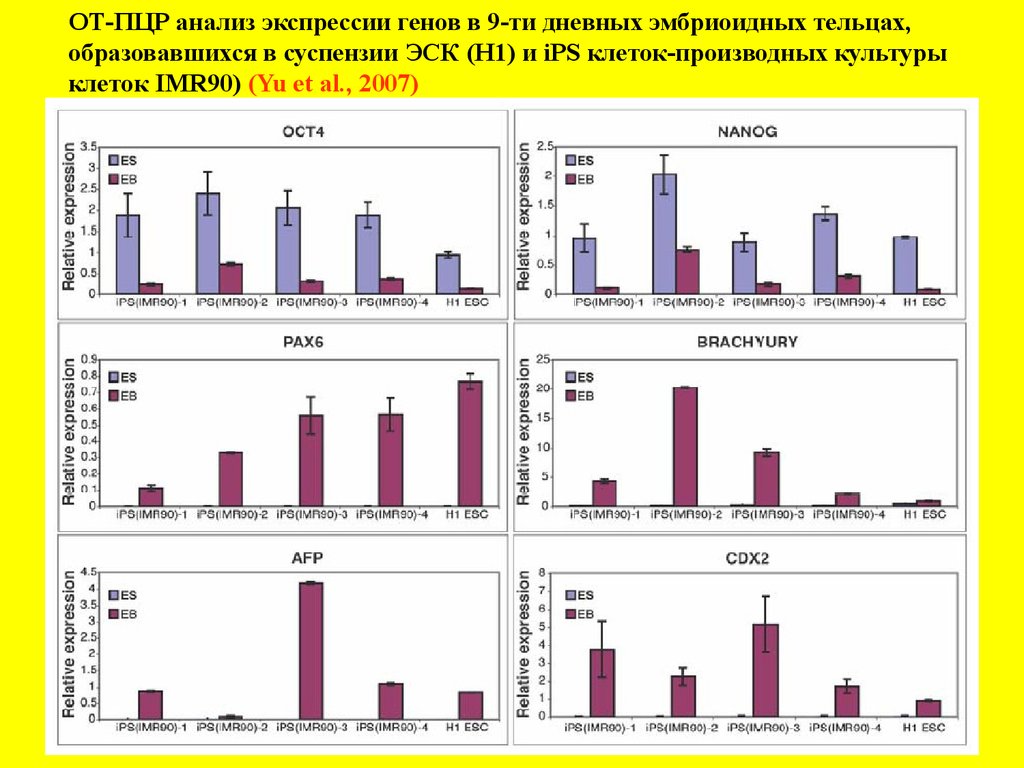
ОТ-ПЦР анализ экспрессии генов в 9-ти дневных эмбриоидных тельцах,образовавшихся в суспензии ЭСК (Н1) и iPS клеток-производных культуры
клеток IMR90) (Yu et al., 2007)
18.
Профили метилирования промотора гена Oct4 в клетках IMR90 (фетальныефибробласты) , фибробластов, ЭСК–Н1 и серии клонов iPS клеток. Темные кружки –
метилированные CpG-сайты, светлые неметилированные (Yu et al., 2007)
19.
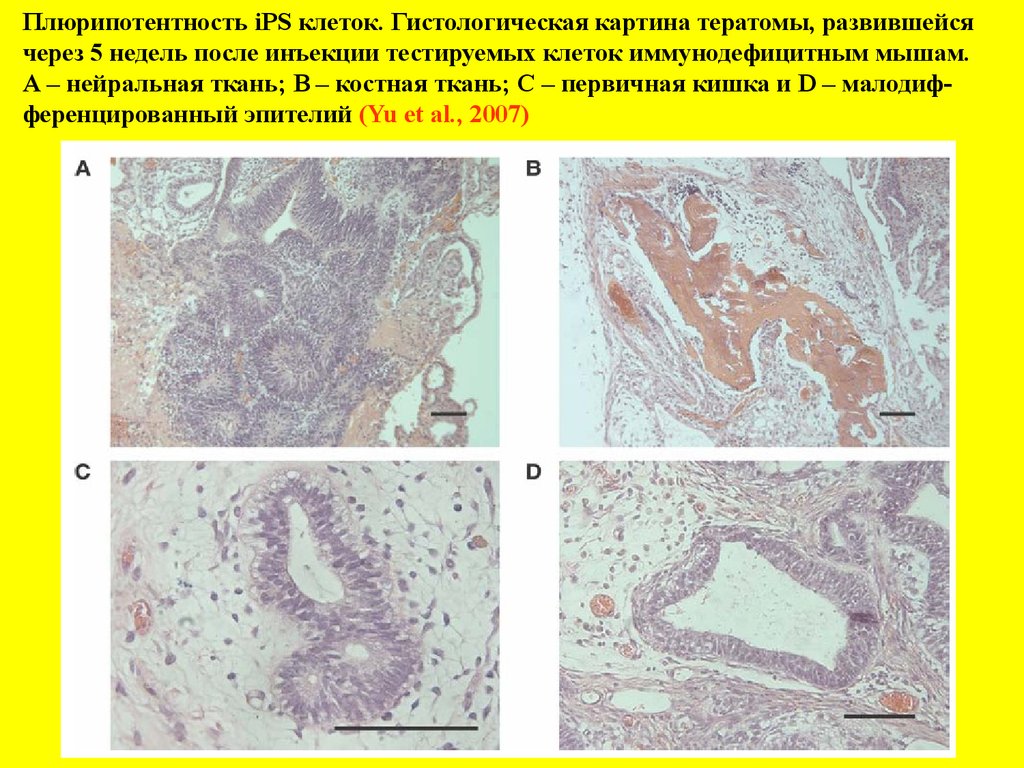
Плюрипотентность iPS клеток. Гистологическая картина тератомы, развившейсячерез 5 недель после инъекции тестируемых клеток иммунодефицитным мышам.
А – нейральная ткань; В – костная ткань; С – первичная кишка и D – малодифференцированный эпителий (Yu et al., 2007)
20.
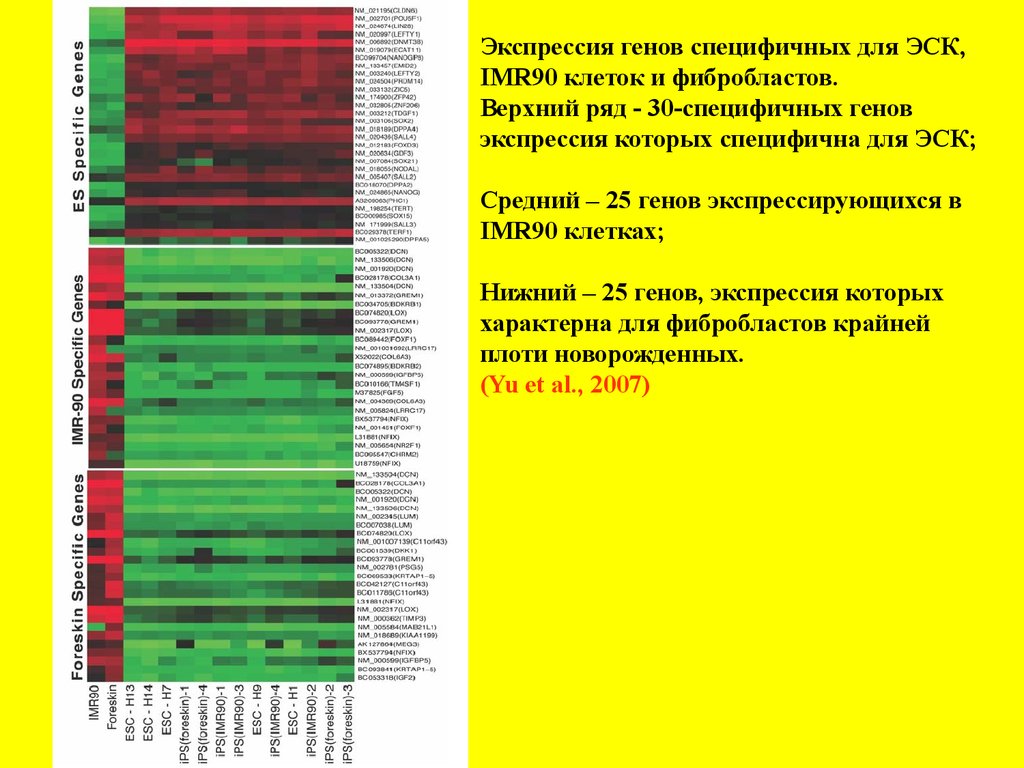
Экспрессия генов специфичных для ЭСК,IMR90 клеток и фибробластов.
Верхний ряд - 30-специфичных генов
экспрессия которых специфична для ЭСК;
Средний – 25 генов экспрессирующихся в
IMR90 клетках;
Нижний – 25 генов, экспрессия которых
характерна для фибробластов крайней
плоти новорожденных.
(Yu et al., 2007)
21.
Индукция плюрипотентности вфибробластах взрослого человека.
А - временная схема индукции;
В – морфология фибробластов;
С – вид колонии не ЭС клеток;
D – вид колонии ЭСК человека;
Е – вид колонии iPS клеток (пассаж 6);
F – колония iPS клеток при
большем увеличении;
G – спонтанная дифференцировка в
центре колонии iPS клеток;
Н – N – иммуногистология: SSEA1 (H),
SSEA3 (I), SSEA4 (J), TRA-1-60 (K),
TRA-1-61 (L), TRA-2-49-AP (M),
Nanog (N). (Takahashi et al., 2007)
22.
iPS клетки изсиновиоцитов и
фибробластов BJ
(Takahashi et al., 2007)
23.
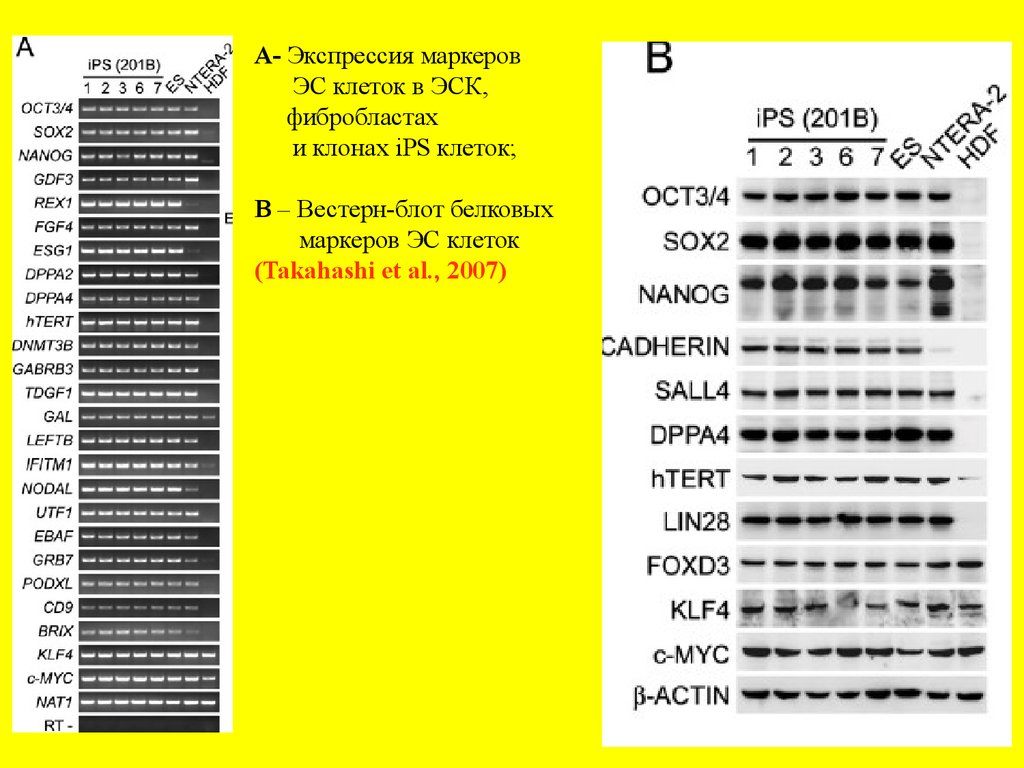
A- Экспрессия маркеровЭС клеток в ЭСК,
фибробластах
и клонах iPS клеток;
В – Вестерн-блот белковых
маркеров ЭС клеток
(Takahashi et al., 2007)
24.
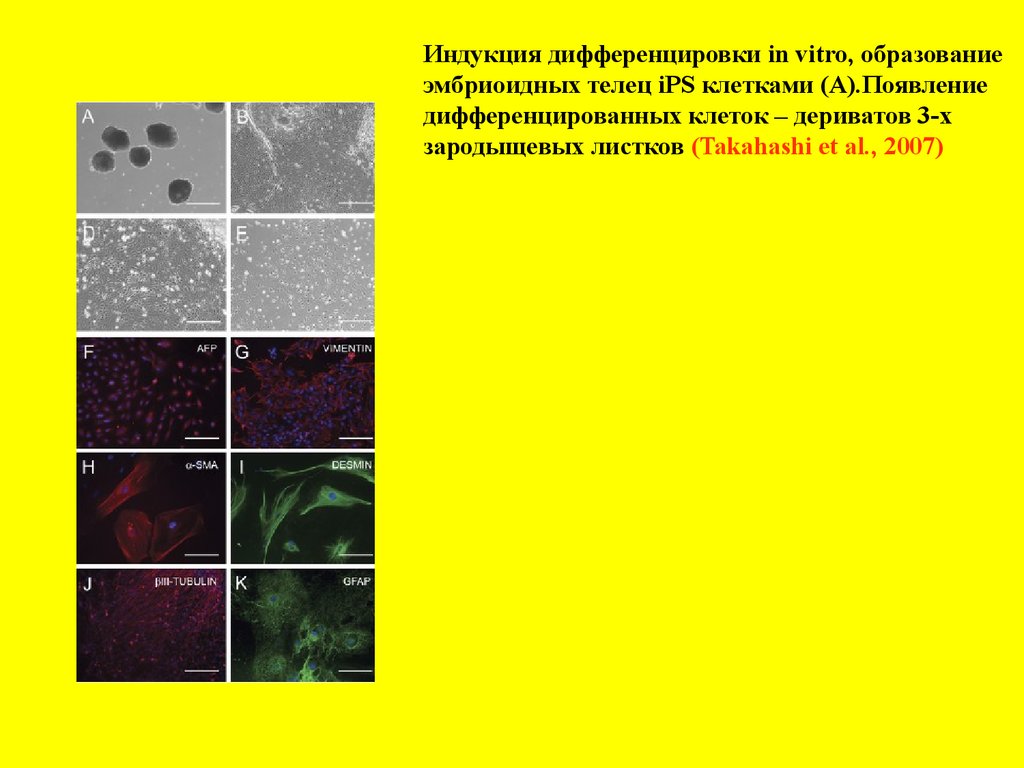
Индукция дифференцировки in vitro, образованиеэмбриоидных телец iPS клетками (А).Появление
дифференцированных клеток – дериватов 3-х
зародыщевых листков (Takahashi et al., 2007)
25.
Экспрессия генов-маркеровкардиомиоцитов в эмбриоидных
тельцах трех клонов iPS клеток
(Takahashi et al., 2007)
26.
Тератома развившаяся из iPS клеток (Takahashi et al., 2007)27.
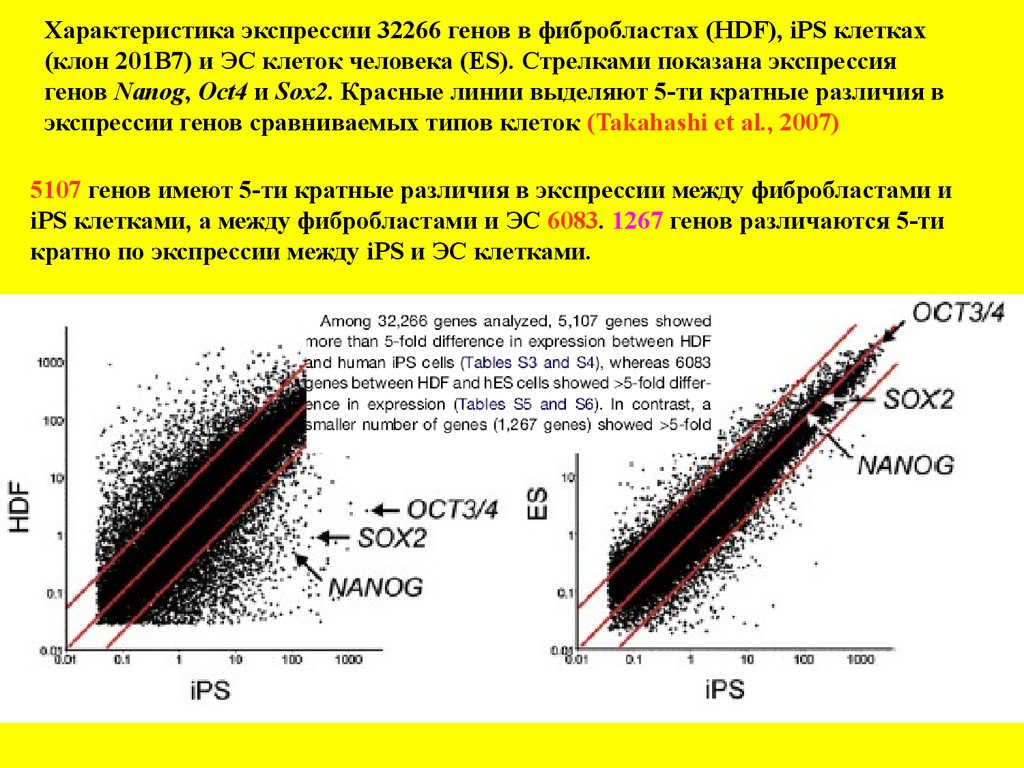
Характеристика экспрессии 32266 генов в фибробластах (HDF), iPS клетках(клон 201В7) и ЭС клеток человека (ES). Стрелками показана экспрессия
генов Nanog, Oct4 и Sox2. Красные линии выделяют 5-ти кратные различия в
экспрессии генов сравниваемых типов клеток (Takahashi et al., 2007)
5107 генов имеют 5-ти кратные различия в экспрессии между фибробластами и
iPS клетками, а между фибробластами и ЭС 6083. 1267 генов различаются 5-ти
кратно по экспрессии между iPS и ЭС клетками.
28.
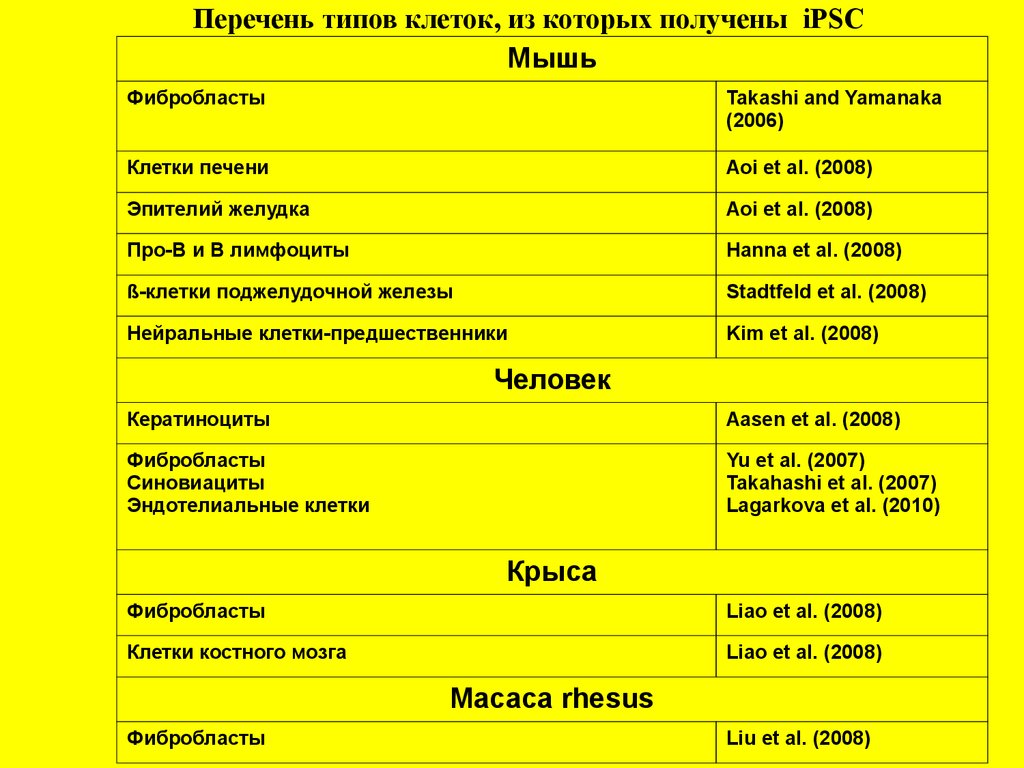
Перечень типов клеток, из которых получены iPSCМышь
Фибробласты
Takashi and Yamanaka
(2006)
Клетки печени
Aoi et al. (2008)
Эпителий желудка
Aoi et al. (2008)
Про-В и В лимфоциты
Hanna et al. (2008)
ß-клетки поджелудочной железы
Stadtfeld et al. (2008)
Нейральные клетки-предшественники
Kim et al. (2008)
Человек
Кератиноциты
Aasen et al. (2008)
Фибробласты
Синовиациты
Эндотелиальные клетки
Yu et al. (2007)
Takahashi et al. (2007)
Lagarkova et al. (2010)
Крыса
Фибробласты
Liao et al. (2008)
Клетки костного мозга
Liao et al. (2008)
Macaca rhesus
Фибробласты
Liu et al. (2008)
29.
«Лабиринт к плюрипотентности» представляет транскрипционные иморфологические события в ходе репрограммирования соматического генома в iPSC.
(Papp and Plath Cell Research 2011online:doi:10.1038/cr.2011.28)