












 biology
biologySimilar presentations:










Спектроскопия комбинационного рассеяния света при изучении липидов в модельных мембранах
1. Спектроскопия комбинационного рассеяния света при изучении липидов в модельных мембранах и биологических клетках
Н.В. СуровцевИнститут автоматики и электрометрии
СО РАН
2020 г.
2. Липиды – широкий класс веществ, включающий жиры. В докладе будет идти речь преимущественно о фосфолипидах, холестерине,
триглицеридахЛипиды
Особенность фосфолипидов заключается в
образовании квазиплоского бислоя в воде
ВОДА
ВОДА
2
3.
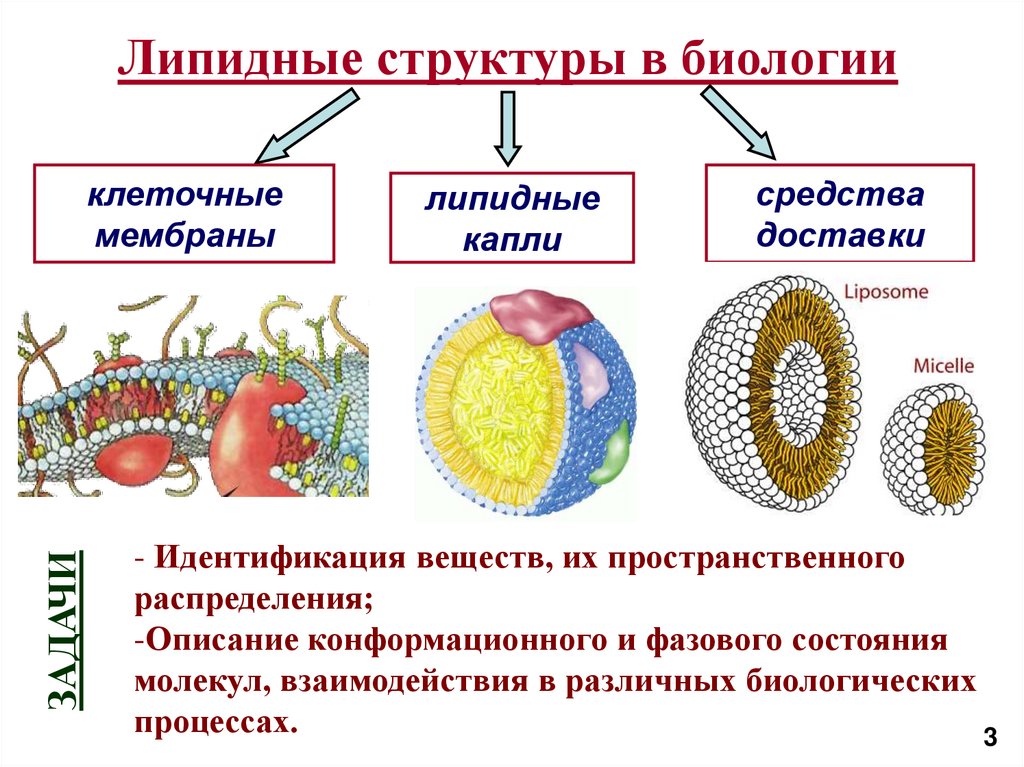
Липидные структуры в биологииЗАДАЧИ
клеточные
мембраны
липидные
капли
средства
доставки
- Идентификация веществ, их пространственного
распределения;
-Описание конформационного и фазового состояния
молекул, взаимодействия в различных биологических
процессах.
3
4.
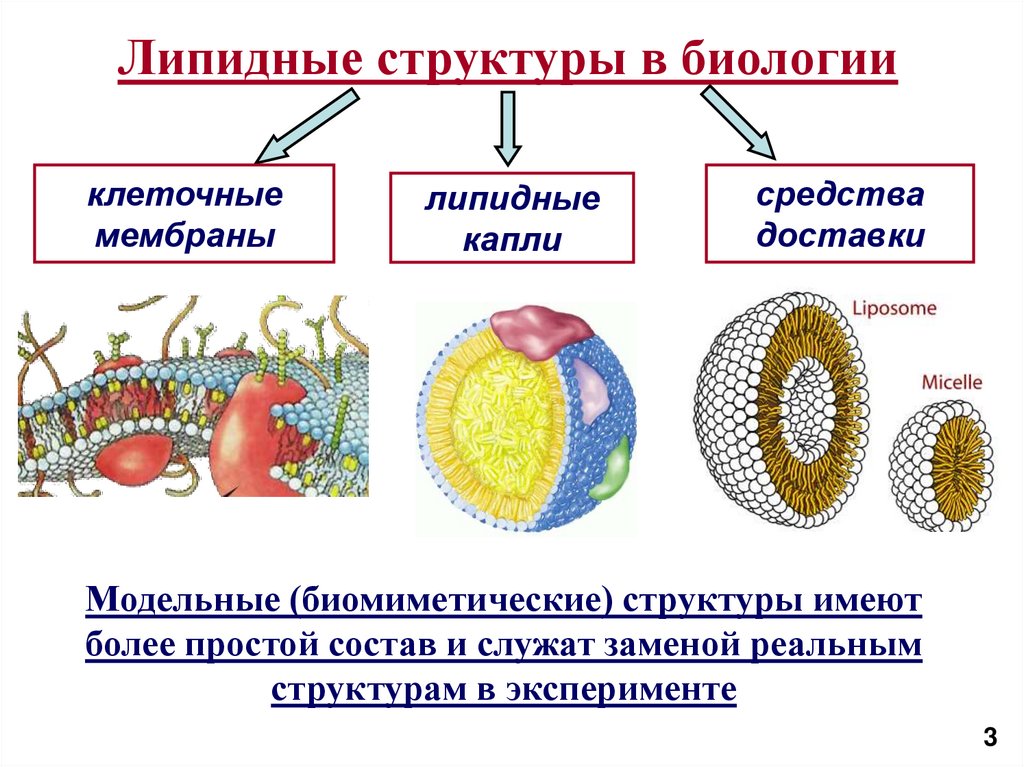
Липидные структуры в биологииклеточные
мембраны
липидные
капли
средства
доставки
Модельные (биомиметические) структуры имеют
более простой состав и служат заменой реальным
структурам в эксперименте
3
5.
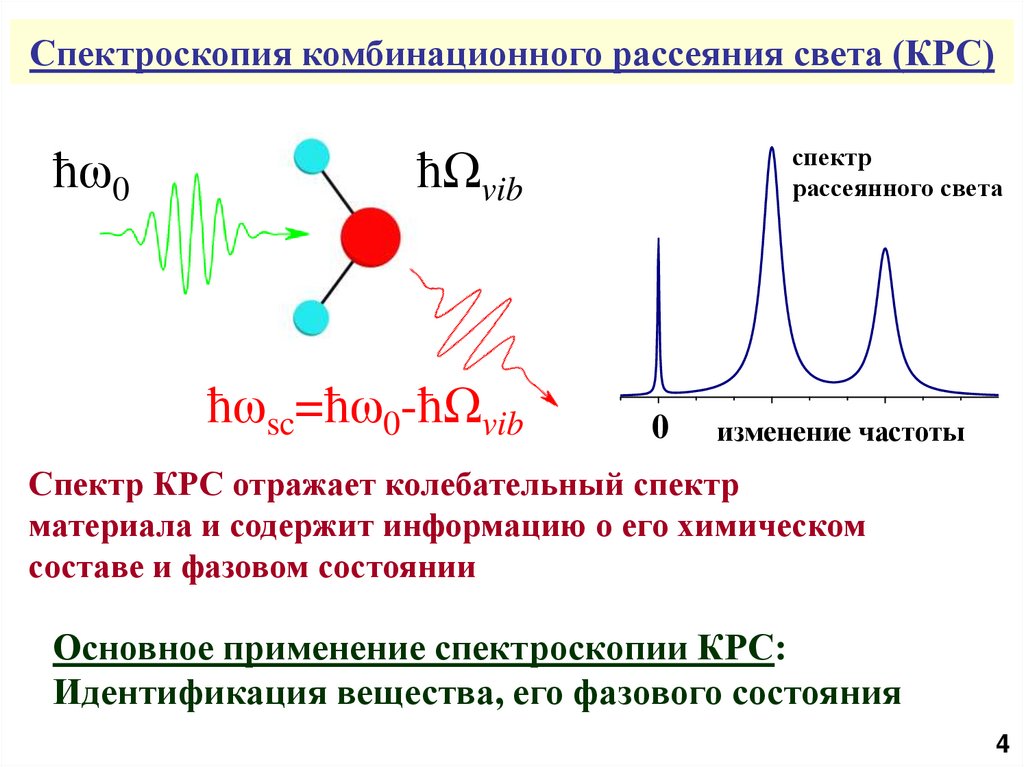
Спектроскопия комбинационного рассеяния света (КРС)ħω0
ħΩvib
ħωsc=ħω0-ħΩvib
спектр
рассеянного света
0
изменение частоты
Спектр КРС отражает колебательный спектр
материала и содержит информацию о его химическом
составе и фазовом состоянии
Основное применение спектроскопии КРС:
Идентификация вещества, его фазового состояния
4
6.
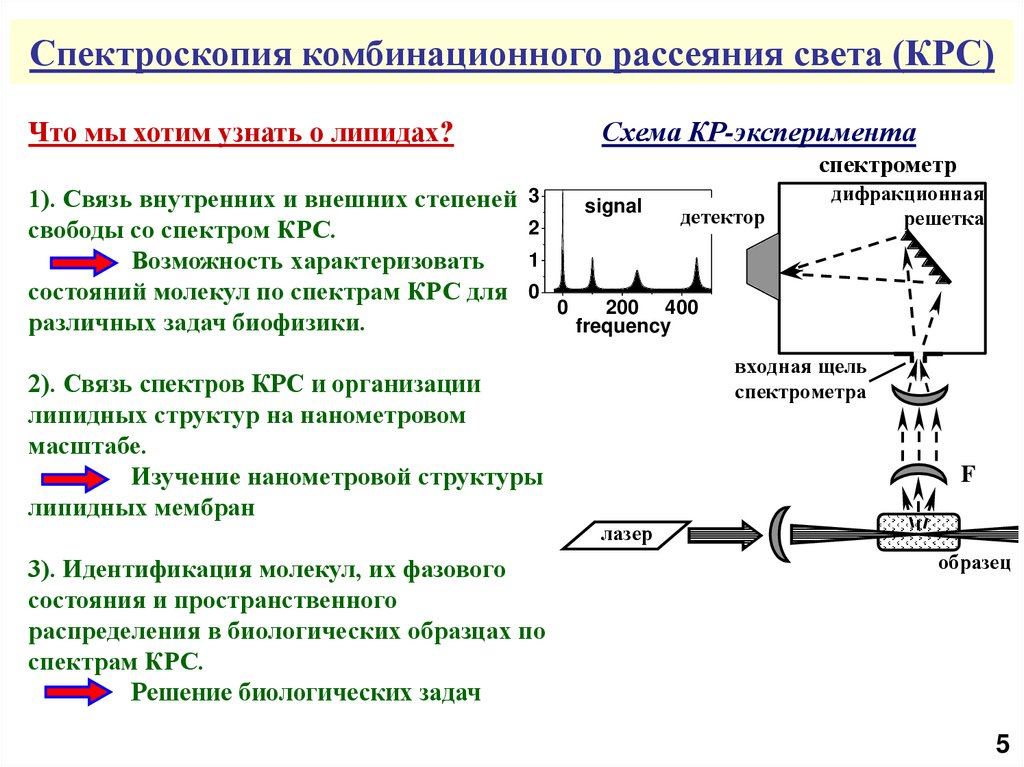
Спектроскопия комбинационного рассеяния света (КРС)Что мы хотим узнать о липидах?
Схема КР-эксперимента
спектрометр
1). Связь внутренних и внешних степеней
свободы со спектром КРС.
Возможность характеризовать
состояний молекул по спектрам КРС для
различных задач биофизики.
3
signal
2
детектор
дифракционная
решетка
1
0
0
200 400
frequency
входная щель
спектрометра
2). Связь спектров КРС и организации
липидных структур на нанометровом
масштабе.
Изучение нанометровой структуры
липидных мембран
F
лазер
3). Идентификация молекул, их фазового
состояния и пространственного
распределения в биологических образцах по
спектрам КРС.
Решение биологических задач
образец
5
7. Особенности колебательных спектров липидов
Сложные и длинные молекулы, состоящие изнебольшого набора характерных связей:
C-H, C-C, C=O, N-H, O-H …
O
O H H H H H H H H H
HO P O С С С С С С С С С С H
H H H H H H H H H
O- Проблема «тонкой» идентификации вещества, его
конформационного состояния;
- Характеризация «мезомасштаба»
6
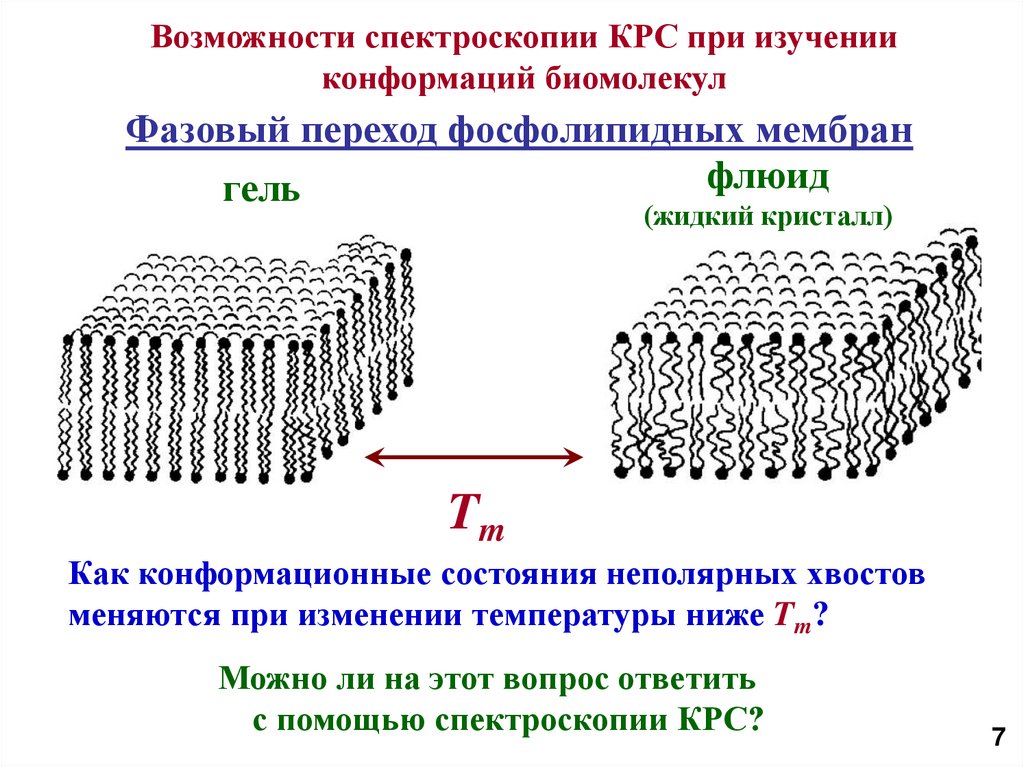
8.
Возможности спектроскопии КРС при изученииконформаций биомолекул
Фазовый переход фосфолипидных мембран
флюид
гель
(жидкий кристалл)
Tm
Как конформационные состояния неполярных хвостов
меняются при изменении температуры ниже Tm?
Можно ли на этот вопрос ответить
с помощью спектроскопии КРС?
7
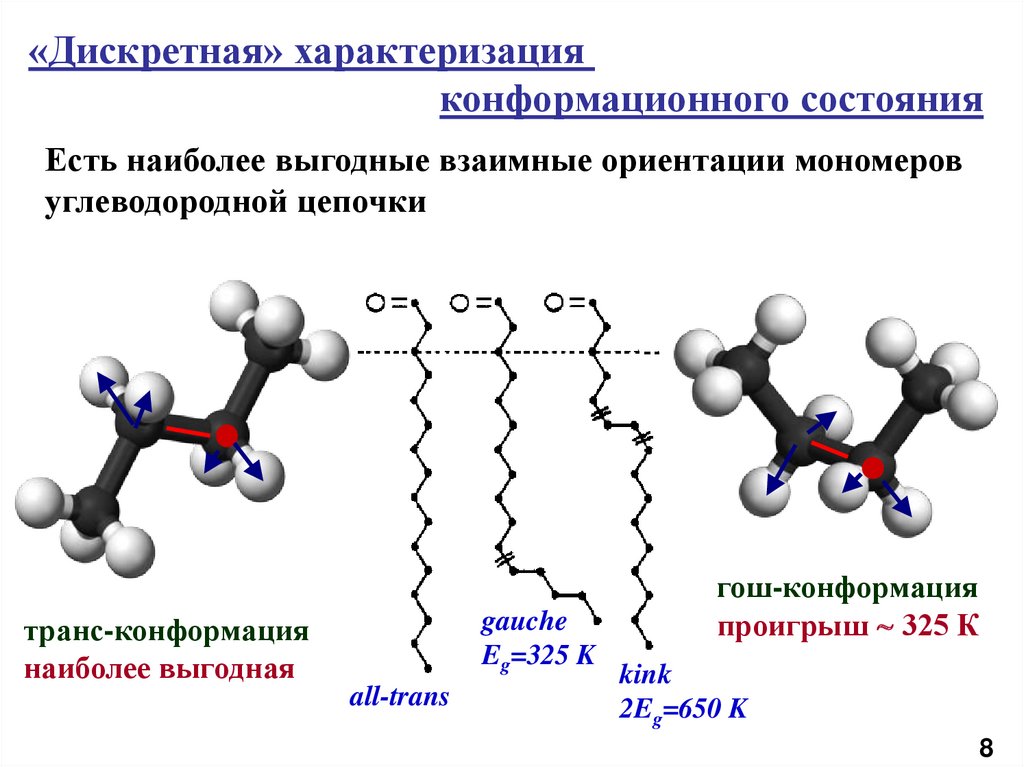
9.
«Дискретная» характеризацияконформационного состояния
Есть наиболее выгодные взаимные ориентации мономеров
углеводородной цепочки
gauche
Eg=325 K
транс-конформация
наиболее выгодная
all-trans
гош-конформация
проигрыш 325 К
kink
2Eg=650 K
8
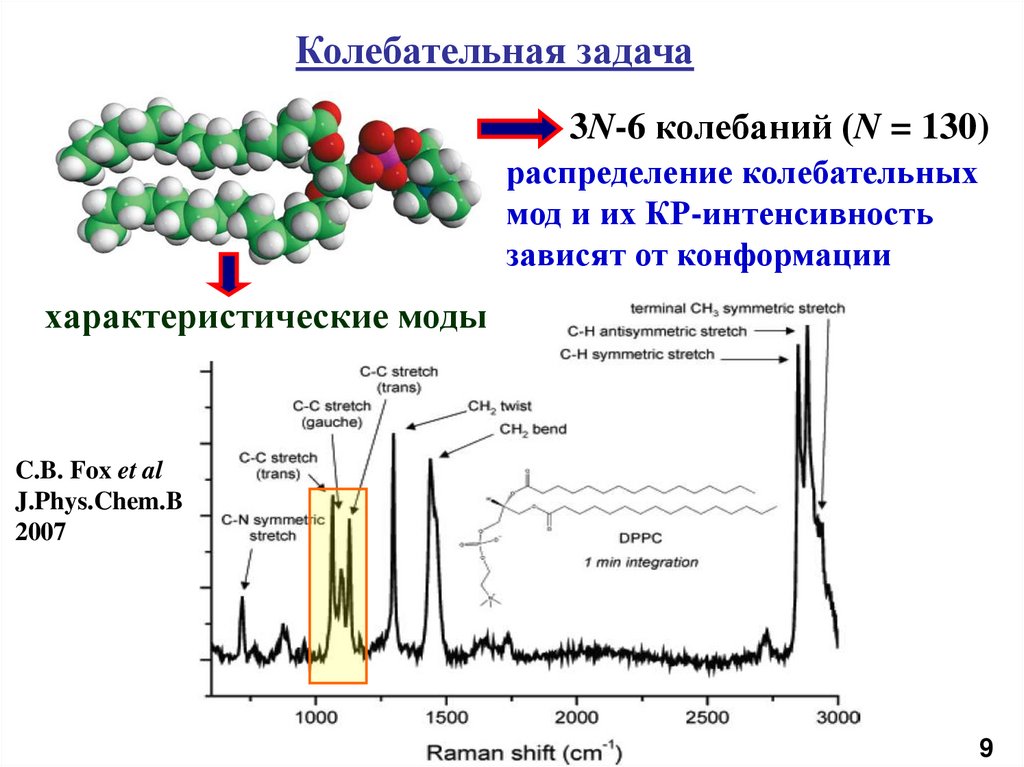
10.
Колебательная задача3N-6 колебаний (N = 130)
распределение колебательных
мод и их КР-интенсивность
зависят от конформации
характеристические моды
C.B. Fox et al
J.Phys.Chem.B
2007
9
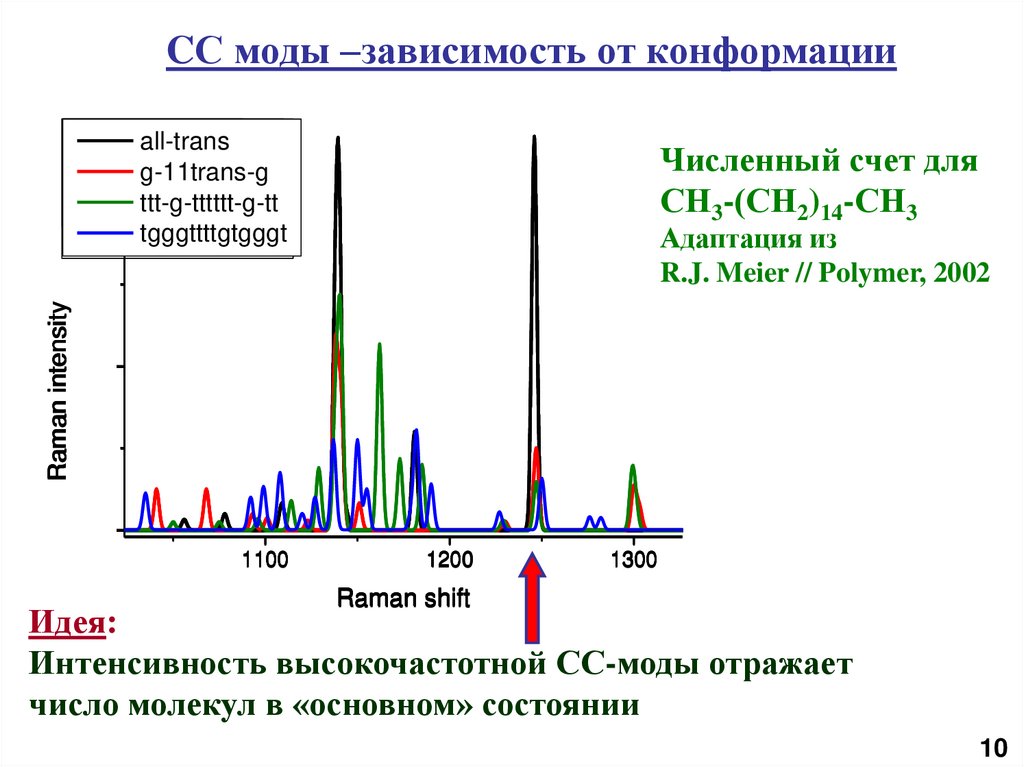
11.
CС моды –зависимость от конформацииall-trans
g-11trans-g
ttt-g-tttttt-g-tt
tgggttttgtgggt
Численный счет для
СН3-(СН2)14-СН3
Raman intensity
Адаптация из
R.J. Meier // Polymer, 2002
1100
1200
1300
Raman shift
Идея:
Интенсивность высокочастотной СС-моды отражает
число молекул в «основном» состоянии
10
12.
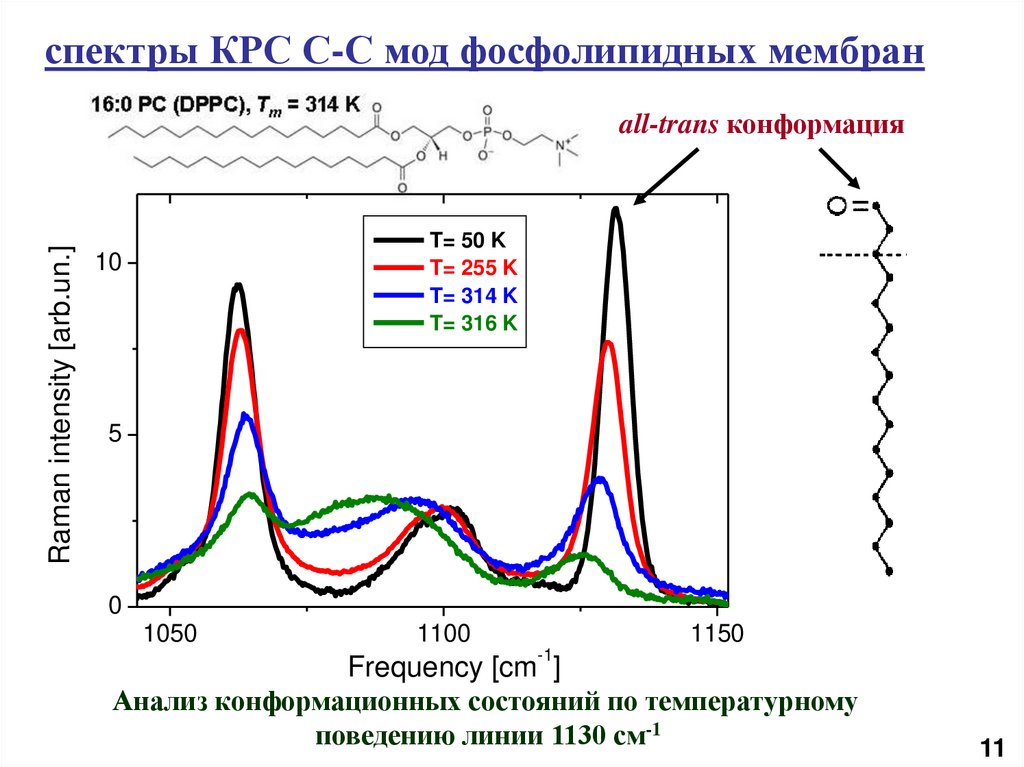
спектры КРС С-С мод фосфолипидных мембранRaman intensity [arb.un.]
all-trans конформация
T= 50 K
T= 255 K
T= 314 K
T= 316 K
10
5
0
1050
1100
1150
-1
Frequency [cm ]
Анализ конформационных состояний по температурному
поведению линии 1130 см-1
11
13.
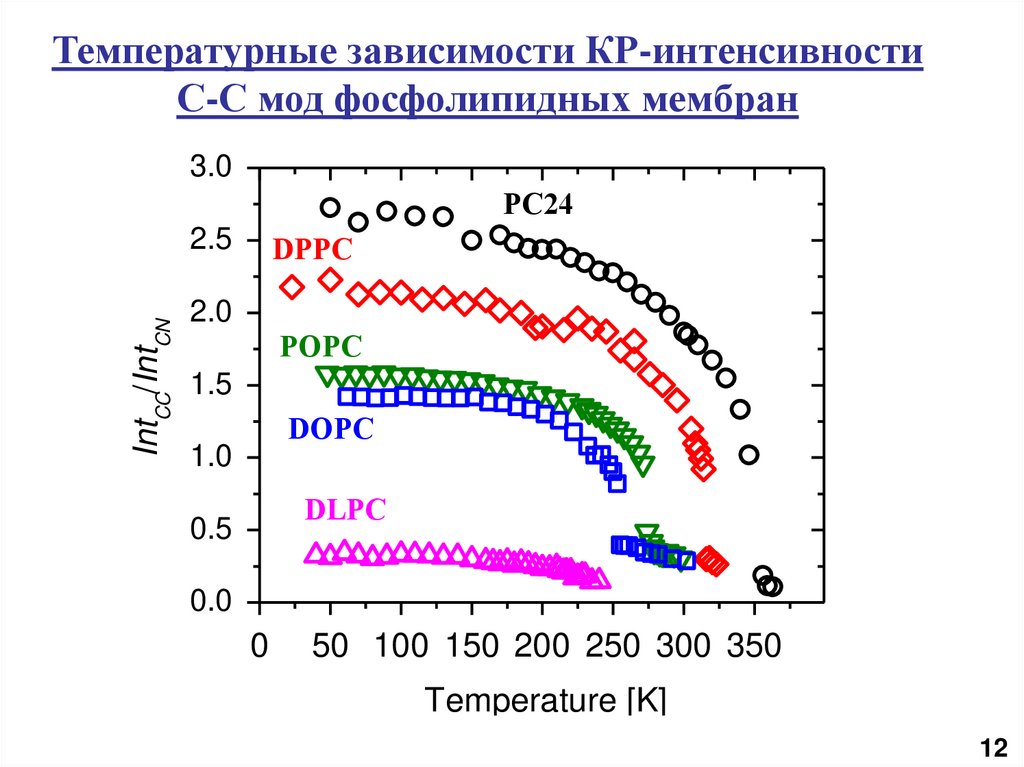
Температурные зависимости КР-интенсивностиС-С мод фосфолипидных мембран
3.0
PC24
IntCC/IntCN
2.5
DPPC
2.0
POPC
1.5
DOPC
1.0
DLPC
0.5
0.0
0
50 100 150 200 250 300 350
Temperature [K]
12
14.
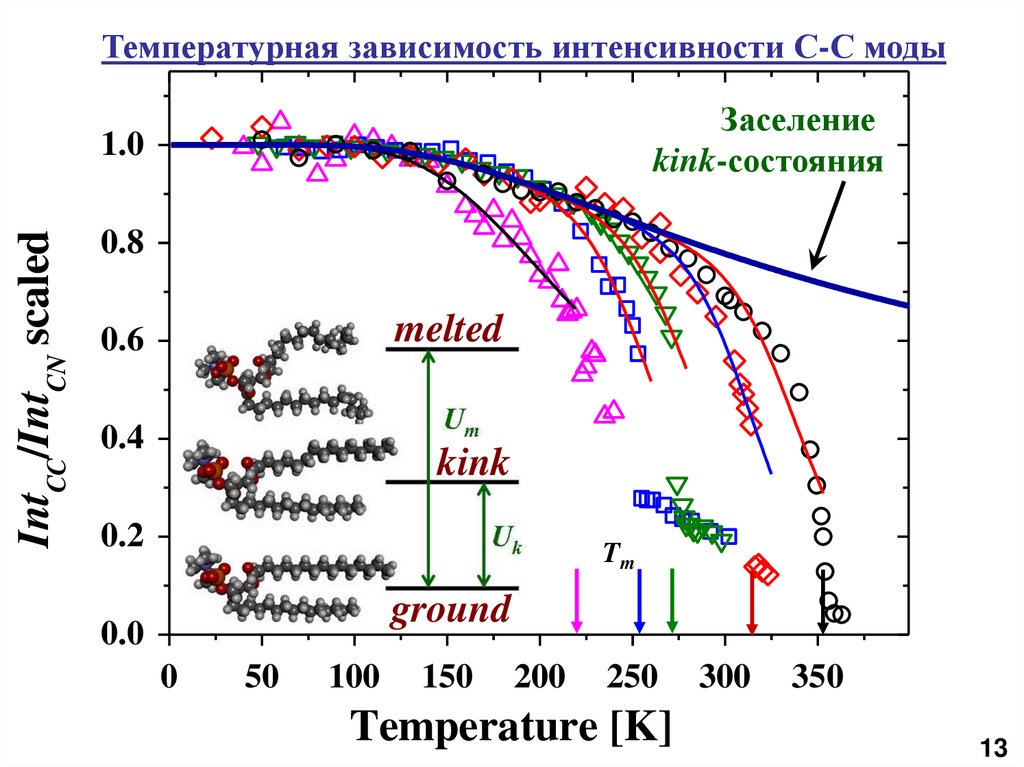
Температурная зависимость интенсивности С-С модыЗаселение
kink-состояния
IntCC/IntCN scaled
1.0
0.8
melted
0.6
0.4
kink
0.2
Tm
ground
0.0
0
50
100
150
200
250
Temperature [K]
300
350
13
15.
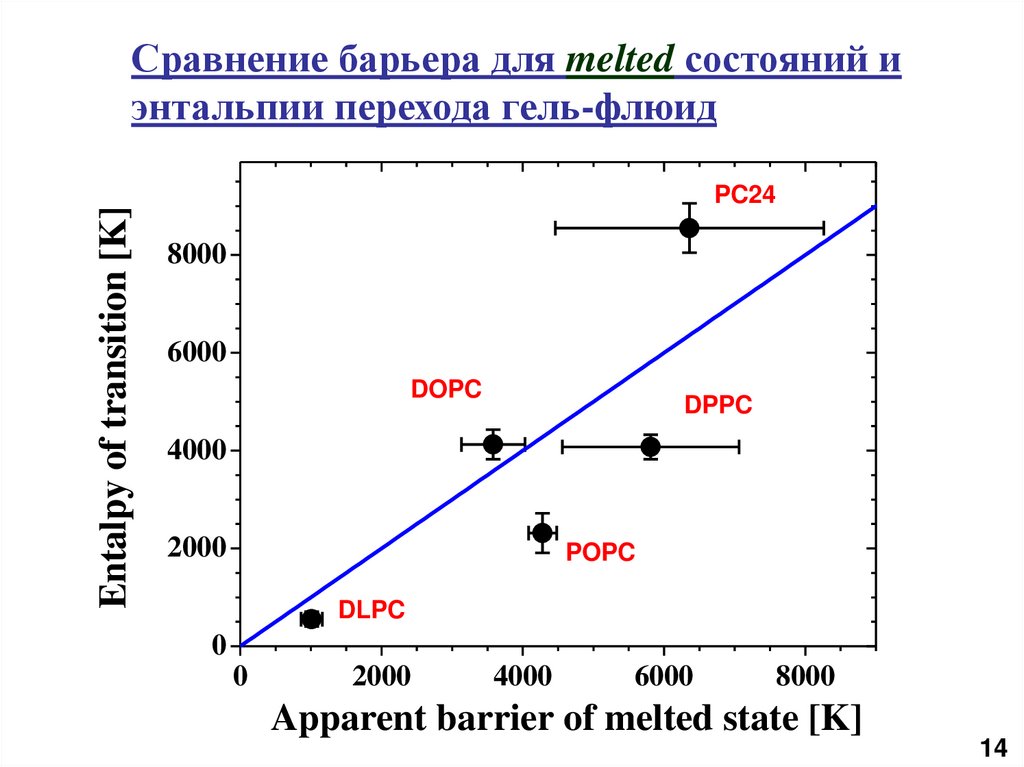
Entalpy of transition [K]Сравнение барьера для melted состояний и
энтальпии перехода гель-флюид
PC24
8000
6000
DOPC
DPPC
4000
2000
POPC
DLPC
0
0
2000
4000
6000
8000
Apparent barrier of melted state [K]
14
16.
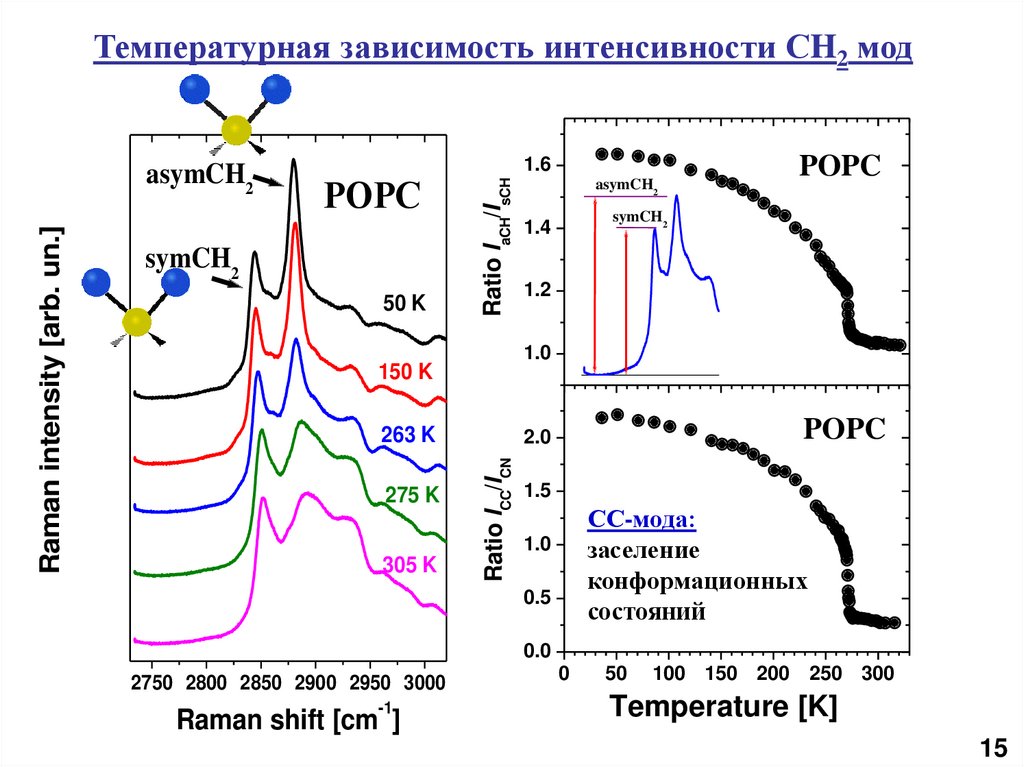
Температурная зависимость интенсивности СH2 модsymCH2
50 K
Ratio IaCH/IsCH
POPC
1.6
263 K
275 K
305 K
asymCH2
POPC
symCH2
1.4
1.2
1.0
150 K
POPC
2.0
Ratio ICC/ICN
Raman intensity [arb. un.]
asymCH2
1.5
СС-мода:
заселение
конформационных
состояний
1.0
0.5
0.0
2750 2800 2850 2900 2950 3000
-1
Raman shift [cm ]
0
50
100 150 200 250 300
Temperature [K]
15
17.
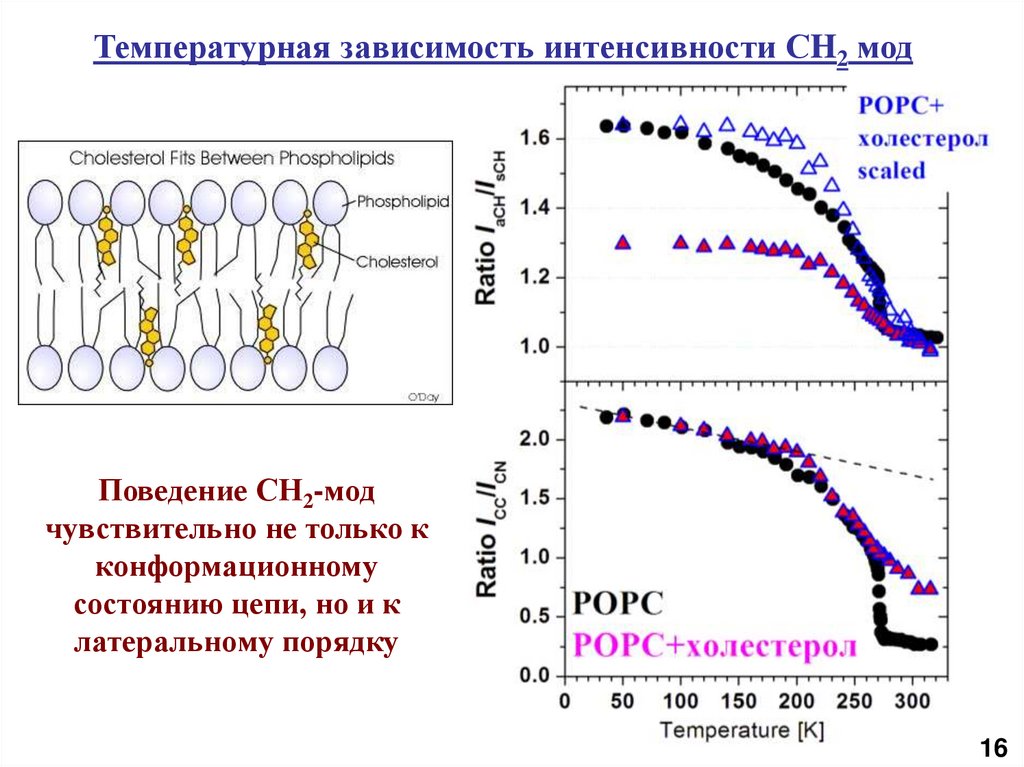
Температурная зависимость интенсивности СH2 модПоведение СН2-мод
чувствительно не только к
конформационному
состоянию цепи, но и к
латеральному порядку
16
18.
-Поведение СС мод в спектре КРС позволяет описатьконформационное состояние неполярных хвостов
фосфолипидных мембран.
- Сравнение поведения СС и СН2 мод в спектре КРС
позволяет делать выводы о характере латерального
порядка.
Перспективы применения этих знаний:
-для многокомпонентных мембран и к проблеме
латеральных неоднородностей в этих мембранах (проблема
«рафтов»);
-для исследования лекарственных средств с модельными
мембранами;
-для описания фазовых изменений в биологических
объектах при замораживании.
17
19.
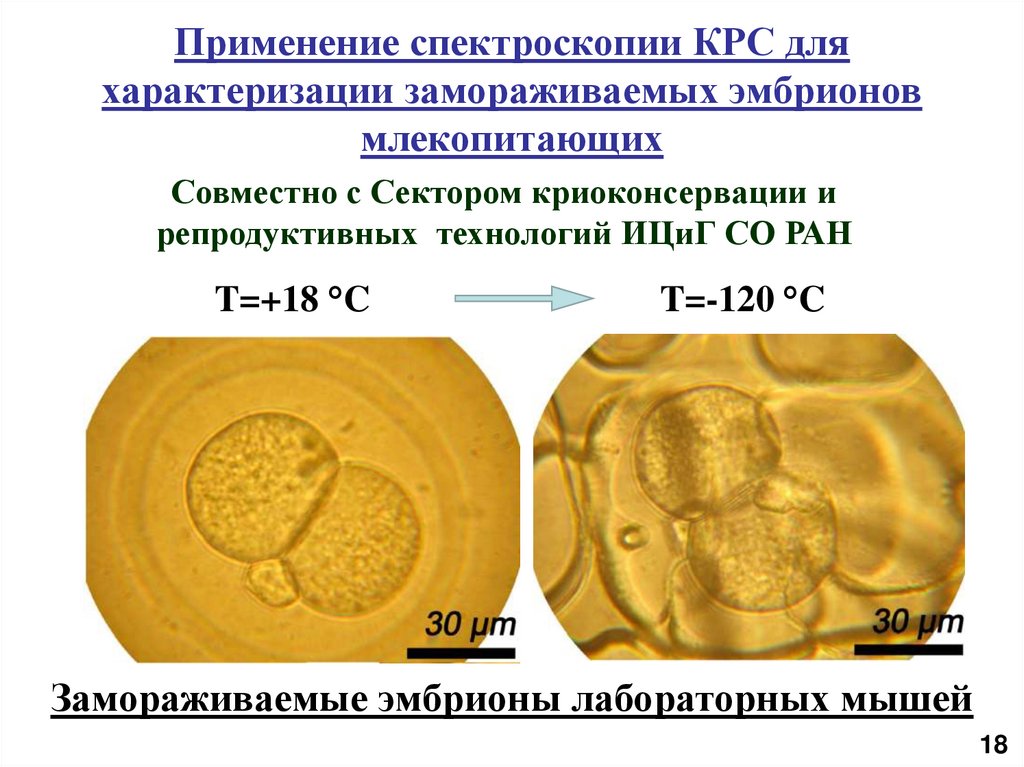
Применение спектроскопии КРС дляхарактеризации замораживаемых эмбрионов
млекопитающих
Совместно с Сектором криоконсервации и
репродуктивных технологий ИЦиГ СО РАН
T=+18 °C
T=-120 °C
Замораживаемые эмбрионы лабораторных мышей
18
20.
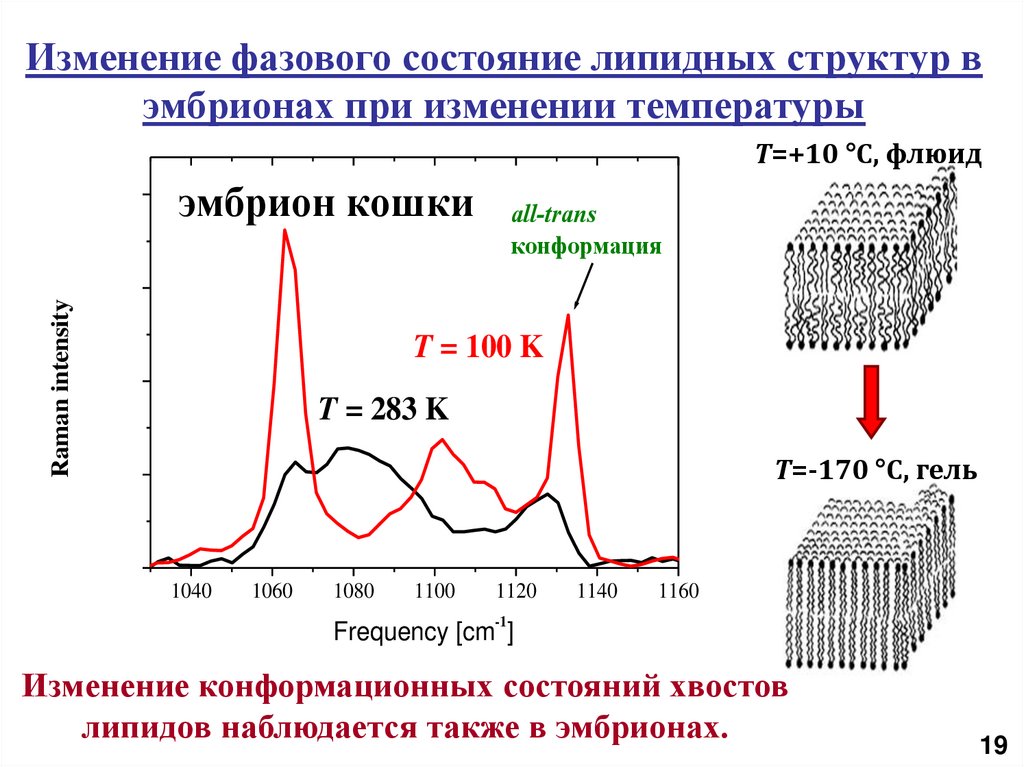
Изменение фазового состояние липидных структур вэмбрионах при изменении температуры
T=+10 °C, флюид
Raman intensity
эмбрион кошки
all-trans
конформация
T = 100 K
T = 283 K
T=-170 °C, гель
1040
1060
1080
1100
1120
1140
1160
-1
Frequency [cm ]
Изменение конформационных состояний хвостов
липидов наблюдается также в эмбрионах.
19
21. Температурная эволюция С-С мод в замораживаемом эмбрионе кошки
ЭмбрионDOPC
Фазовый переход в липидных гранулах может быть
обнаружен по скачку в параметрах “all-trans” моды.
20
22. Температурная эволюция СН2 мод в замораживаемом эмбрионе кошки
Tm = −6 CDOPC
Эмбрион
Фазовый переход в липидных гранулах похож на переход в
синтетических везикулах.
21
23. Выделение в спектры КРС в области СН-колебаний различных компонент
Scaled Intensity [arb.un.]1.0
0.8
Disordered Lipids
Ordered Lipids
Protein
KSOM solution
0.6
0.4
0.2
0.0
2700
2800
2900
Raman shift [cm 1]
3000
3100
22
24. Состояние липидов в ооцитах кошки при T = 0 °C
35 mDisordered
Lipid
Ordered
Lipid
KSOM
Solution
70 m
Protein
23
25. Распределение веществ в эмбрионах мыши
По спектрам диапазона СН-stretch модКартирование
24
26. Собственные колебательные моды фосфолипидных мембран в спектрах КРС
Собственные колебанияпластины толщиной 2d
N cm
1
cS N
4cd
если 2d = 5 nm, cS = 2.4 km/s
1= 8 cm-1
Приближения:
Континуальное описание продольных колебательных мод со
свободными граничными условиями.
Пренебрегаем вкладом латеральных акустических мод в
области первых двух-трех собственных мод бислоя (~ 8, 16, 24
см-1) и распространением по нескольким слоям фосфолипидов.
25
27. Ожидаемые ТГц спектры КРС фосфолипидных мембран
колебательные моды бислоя1
2
4
3
5
Vibration density
N cm
0
10
20
30
40
1
cS N
4cd
mechanical strain
50
-1
frequency [cm ]
1
2
4
3
5
Raman intensity
разрешенные
моды в
спектре КРС
2d
first
harmonic
0
10
20
30
40
-1
frequency [cm ]
50
second
harmonic
Fig. 1
26
28. Ожидаемые ТГц спектры КРС фосфолипидных мембран
12
4
5
КР-моды бислоя
40
50
Raman intensity
3
0
10
20
30
-1
frequency [cm ]
1
2
4
5
Raman intensity
3
А если нет контакта
между слоями?
0
10
20
30
40
-1
frequency [cm ]
50
КР-моды монослоя
27
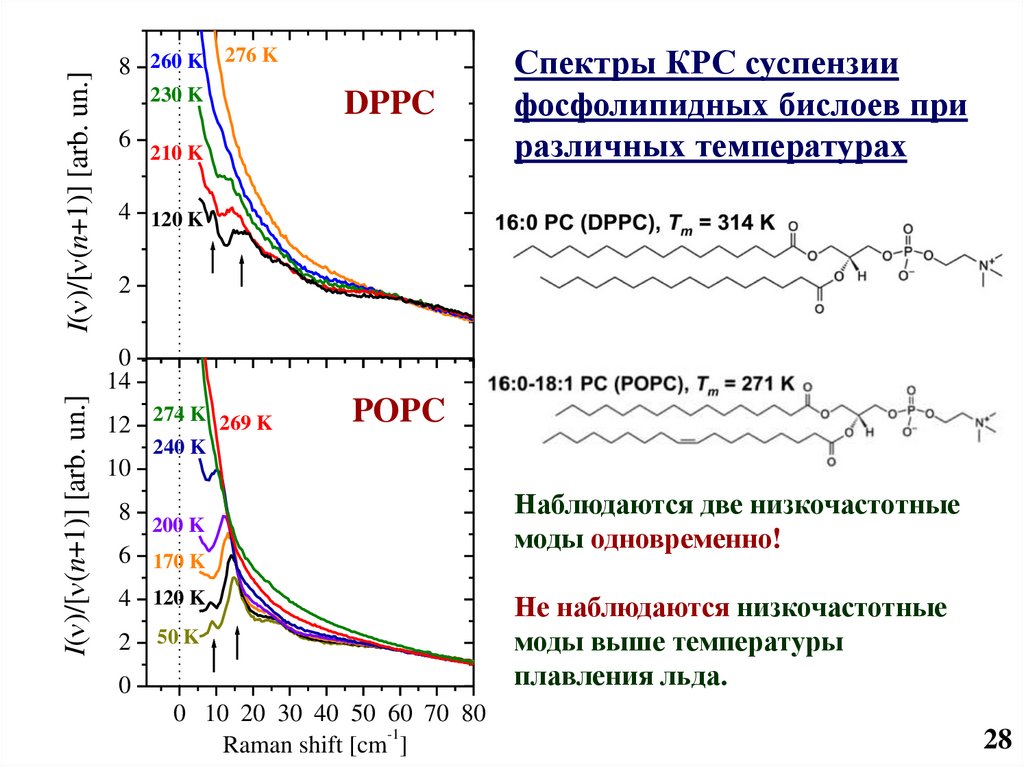
29.
I( )/[ (n+1)] [arb. un.]8 260 K 276 K
230 K
6
a)
DPPC
210 K
Спектры КРС суспензии
фосфолипидных бислоев при
различных температурах
4 120 K
2
I( )/[ (n+1)] [arb. un.]
0
14
12 274 K 269 K
POPC
b)
240 K
10
8
200 K
6 170 K
4 120 K
2
50 K
0
0 10 20 30 40 50 60 70 80
-1
Raman shift [cm ]
Наблюдаются две низкочастотные
моды одновременно!
Не наблюдаются низкочастотные
моды выше температуры
плавления льда.
28
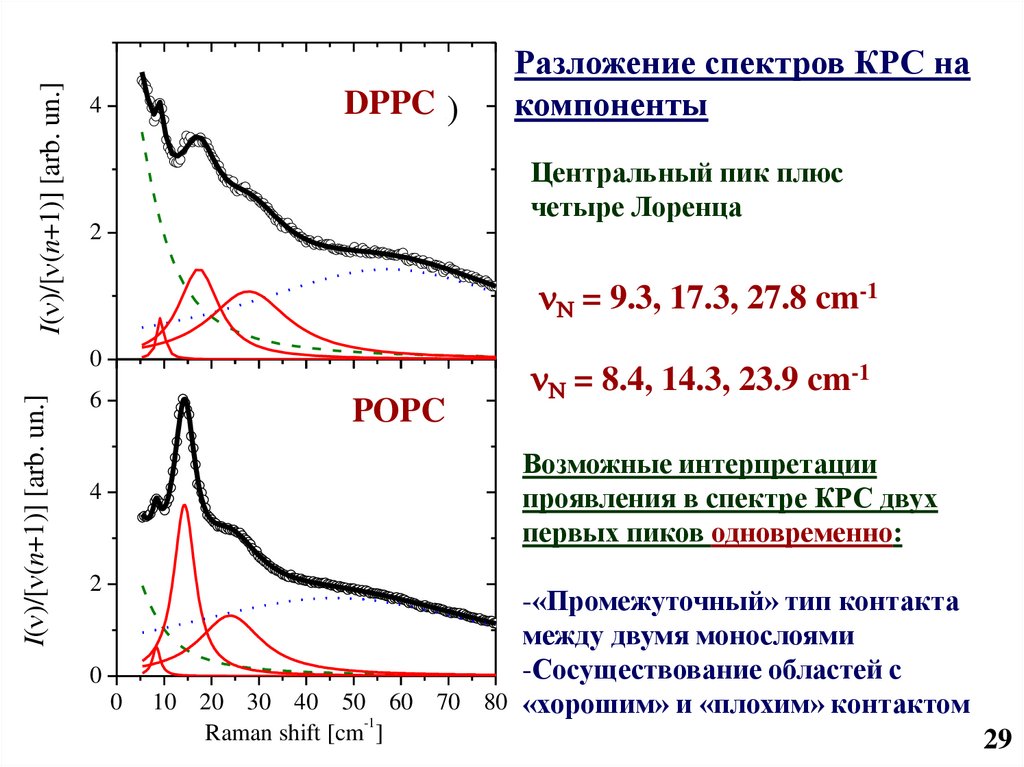
30.
I( )/[ (n+1)] [arb. un.]DPPCa)
4
Центральный пик плюс
четыре Лоренца
2
N = 9.3, 17.3, 27.8 cm-1
0
I( )/[ (n+1)] [arb. un.]
Разложение спектров КРС на
компоненты
6
POPC
b)
N = 8.4, 14.3, 23.9 cm-1
Возможные интерпретации
проявления в спектре КРС двух
первых пиков одновременно:
4
2
0
0
-«Промежуточный» тип контакта
между двумя монослоями
-Сосуществование областей с
10 20 30 40 50 60 70 80 «хорошим» и «плохим» контактом
-1
Raman shift [cm ]
29
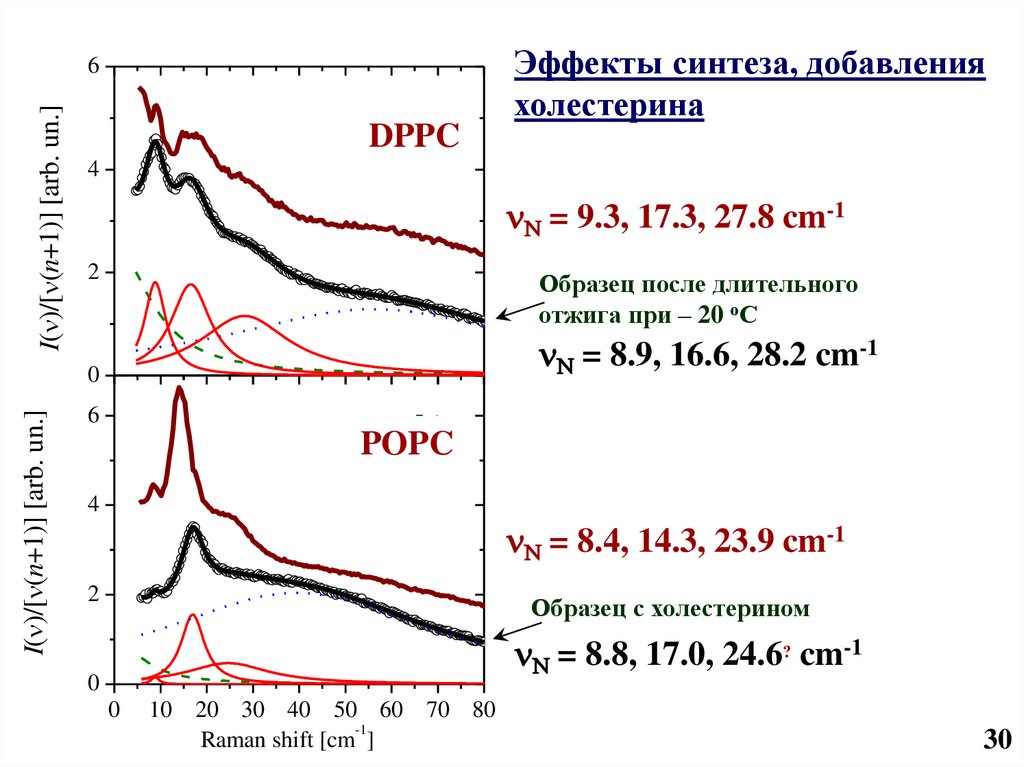
31.
I( )/[ (n+1)] [arb. un.]6
a)
DPPC
4
N = 9.3, 17.3, 27.8 cm-1
2
Образец после длительного
отжига при – 20 оС
N = 8.9, 16.6, 28.2 cm-1
0
I( )/[ (n+1)] [arb. un.]
Эффекты синтеза, добавления
холестерина
6
b)
POPC
4
N = 8.4, 14.3, 23.9 cm-1
2
Образец с холестерином
N = 8.8, 17.0, 24.6? cm-1
0
0
10 20 30 40 50 60 70 80
-1
Raman shift [cm ]
30
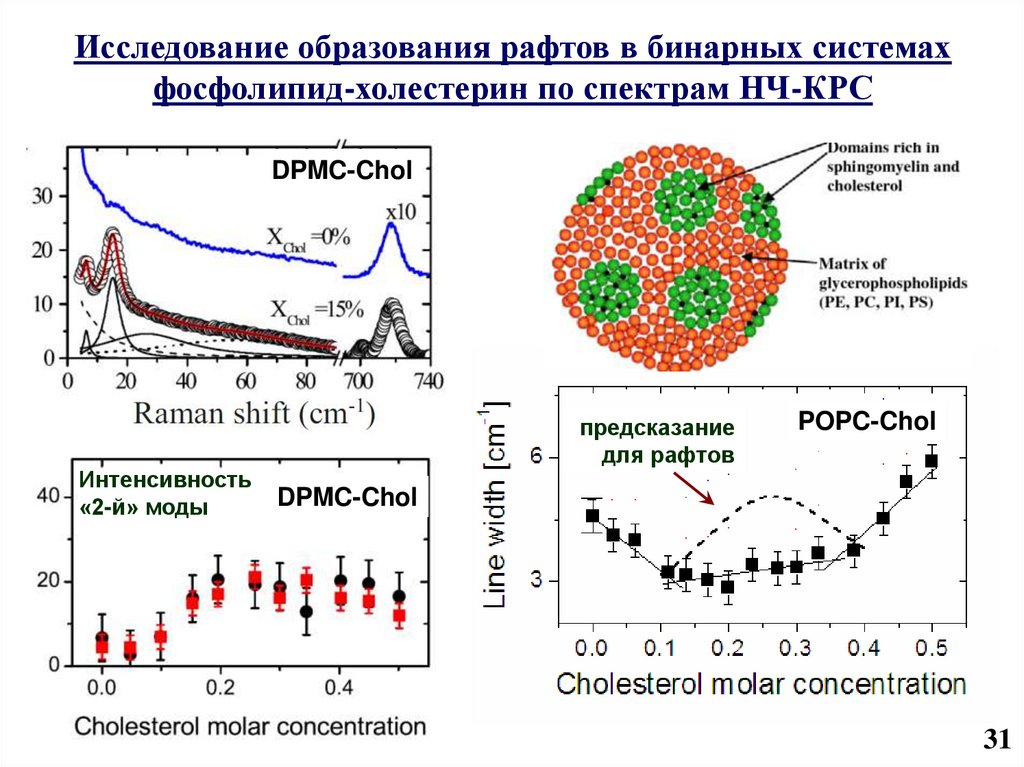
32.
Исследование образования рафтов в бинарных системахфосфолипид-холестерин по спектрам НЧ-КРС
DPMC-Chol
Интенсивность
«2-й» моды
предсказание
для рафтов
POPC-Chol
DPMC-Chol
31
33.
ЗаключениеСпектроскопия комбинационного рассеяния света может
применяться для описания липидных структур, включая:
- Характеризацию конформационного состояния (по модам
СС).
-Характеризацию латерального порядка (по модам СН2 и
сравнению с поведением СС)
-Описание собственных колебательных мод
фосфолипидных слоев
Эти возможности спектроскопии КРС могут быть
эффективно применены при решения различных
биологических задач, включая:
-Проблемы криоконсервации
-Описание образования рафтов в биомембране
-Взаимодействие с лекарственными средствами
32