































. Кольцевое видообразование через цепь расселяющихся подвидов Становление серебристой чайки и клуши-хохотуньи, от Берингова")








































 biology
biologySimilar presentations:
")









Вид и видообразование. Лекция 5
1. Вид и видообразование
Если мы не будем знать,что такое вид - ничто
не удержит нас
от заблуждений
Боэций
Аниций Манлий Торкват
Северин Боэций
480-524
Боэций (слева) на Рафаэлевой фреске «Афинская школа»
2. План:
1.Значение понятие "вид"2. История формирования взглядов
3.Современная трактовка понятия "вид"
4.Критерии вида
5.Пути и способы видообразования
3. 1.Значение понятие "вид"
1.Значение понятие "вид"Вид является основным понятием в биологии:
1. это инструмент для описательной биологии.
разработка систематики-науки о разнообразии
организмов и таксономии - теории и практики
классификации организмов:
2. разработка понятия "вид" необходима для изучения и
описания разнообразия мира;
3. для объяснения причин и изменения данного
разнообразия.
4. изменение вида и его становления это
микроэволюционные процессы
- основа
эволюции.
Длительный путь формирования понятия,
содержание которого менялось в зависимости:
• от мировоззренческих установок
уровня развития науки;
4. Этапы формирования понятия "вид"
Этапы формирования понятия "вид"Критерий - новое в осмыслении
дискретности и разнообразия в природе.
I. вид как логическая категория,
введенная Аристотелем.
Вид - группы сходных предметов,
• а в отношении организмов-
группа сходных животных
или растений,
Аристотель
(384 -322 до н.э.)
• "species".
Платон: вид как конкретное отражение
идеи заложенной творцом.
Платон
(427-347 до н.э.)
5. II. понятие о виде как биологической категории
• отражает органическоеразнообразие и дискретность
• Вид - это реально существующие
морфологически сходные типы
организмов.
Джон Рэй
(1627 –1705)
Это привело к формированию
типологической концепции вида
на основе креационизма.
• Вид = тип сходных организмов,
созданных творцом.
Карл Линней
(1775-1778)
6.
С этой точки зрения:разнообразие - есть существование
нескольких универсалий;
изменчивость - результат
несовершенного проявления идеи,
заложенной в каждом виде творцом;
особи - есть выражение одного и того же
типа признаков;
Основа классификации морфологический и монотипический
стандарт (голотип).
7. III. номиналистическая концепция вида
• развитие трансформизма и идеиизменяемости видов.
Положения:
1. отрицание вида как реальной
категории;
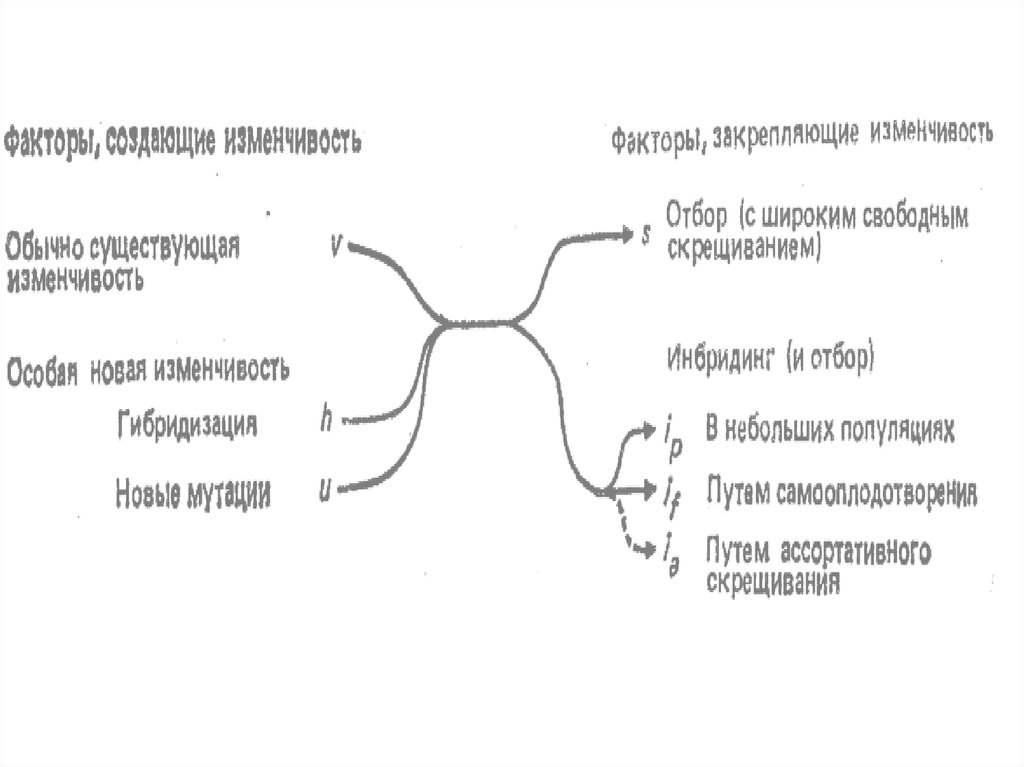
2. утверждение о реальности
только особи;
3. вид - условная категория,
созданная наукой для объединения
сходных особей с целью их
изучения.
ЖоржЛуи
Леклерк,
граф
де
Бюффон
(17071788)
В основе этой концепции лежит
эмпирический подход -
непосредственное изучение
особей.
Жан Батист Ламарк
(1744-1829)
8. IV. 2-я половине 19 века - теория Дарвина и доказательство реальности вида:
1. вид как историческаяреальность;
2. противоречивый характер вида:
устойчивость + динамизм;
3. отделение понятия
реальности вида от его
дискретности:
• изменчивость вида нарушает
четкость границ между
видами (континуум) и
дискретность не обязательно
выражена.
Чарлз Дарвин
(1809 -1882)
9.
• Виды реальны,а могут не реальны границы между
ними
• Вид как временное образование,
т.к. организмы живут во времени пространстве.
• Для эволюции вид реален как
отражение времени в организмах.
10.
• С этого времени начинается формированиебиологической концепции вида,
Вид сочетает в себе:
-дискретность реального вида в данный
момент (описывается общими признаками) и
-эволюционную потенцию к непрерывному
изменению.
Основное внимание было обращено на
устойчивость вида как рез-т эволюции,
выраженной в морфофизиологической
однородности.
Эта целостность вида отразилась в понятии
"монотипического вида"
11. V. В начале 20 века - кризис понятия
• C одной стороны, это связано абсолютизациейтакого признака как целостность вида
• С другой стороны - установлением
внутривидовых группировок: подвидов,
географических, экологических и
физиологических рас.
+
выявлением сложной генетической
структуры и соответственно ей дробление
вида на сотни устойчивых мелких форм.
Известное течение "дробителей" - "жорданизм"
(Жордан). (пшеница - 300 видов).
Результат: отрицание понятия "вид" как
целостной категории.
12. VI. 1930-е вид как сложная динамическая система.
Системный подход ( Вавилов и Клаузен)• Установление генетического
единства вида:
это общий генофонд за счет скрещивания
популяций
+
защита изоляционными барьерами от
проникновения генов других видов.
• Вид как защищенный генофонд и сложная
генетическая система с внутренним
разнообразием, имеющим адаптивное
значение.
Перенос акцента на репродуктивную и
экологическую целостность вида
( а не на морфологическую, как в 19 в).
13.
• Это дальнейшаяразработка
биологической
концепции вида,
• На основе чего
сформировалось понятие
политипического вида
(имеющего сложную
структуру).
Никола́й Ива́нович
Вави́лов
(1887- 1943)
14. VII. Современный этап - дальнейшая разработка понятия политипического вида.
Вид - сложная динамическая система, представленнаясложной структурой, что позволяет сочетать
устойчивость и изменчивость.
Развитие иерархического подхода к систематизации
организмов:
• Альфа-таксономия (описание новых видов каталог);
• Бета- таксономия (классификация);
• Гамма - таксономия (изучение изменчивости видов,
индивидуальная, географическая).
Недопустимость описания вида по 1 голотипу,
вследствие внутривидовой изменчивости, нужна
репрезентативная выборка
(от организменного - к популяционному подходу).
15.
• Проблема единого и адекватного определения понятия"вид".
• Многообразие точек зрения на "вид" - отражение его
многоаспектности.
• С позиций систематики , это объективно определяется
несколькими причинами:
1.Это трудности установления границ вида и его
внутренняя противоречивость
2 . Неравномерность таксономического распределения.
(напр. клювоголов. -1 гаттерия, а чешуйчатые - тыс.
видов.)
3.Огромное разнообразие видов в природе
(1,2 млн. видов животных, более 500 тыс. растений,
по разным оценкам разнообразие от 5-7 млн.)
Вымирают быстрее, чем успеваем описывать.
• Глобальная проблема - сохранения б/р.
16.
Число описанных видов живых организмов насоставляет примерно 1,7 миллиона, в том
числе:
• позвоночные животные — 64 тыс.,
беспозвоночные — 1,3 миллиона,
• растения (включая красные и зелёные
водоросли) — 308 тыс.
• грибы (включая лишайники) — 48 тыс.
(по другим данным — около 100 тысяч видов)
17.
Биологическая концепция(предложена Эрнстом Майром).
Вид признаётся дискретным только в данный момент, с течением
времени вид непрерывно эволюционирует. Наряду с этим в описании
вида используются и «традиционные» признаки, и экологобиологические (популяционная структура вида, способность особей
давать плодовитое потомство). В связи с этим особое значение
приобретают генетические связи внутри вида, а видовой статус
является свойством популяции, а не отдельного индивидуума.
Отсюда биологическое определение вида: вид – группа
особей, сходных по морфолого-анатомическим,
физиолого-экологическим, биохимическим и
генетическим признакам, занимающих естественный
ареал, способных свободно скрещиваться между
собой и давать плодовитое потомство; есть другое
определение: вид – репродуктивно связанная
совокупность популяций.
18.
Репродуктивная концепция(предложена
Р. Мейером (Meier) и Р. Виллманном
(Willmann) на основе взглядов основателя кладистики
Вилли Хеннига):
Здесь основным критерием вида является наличие репродуктивной изоляции
между особями разных видов – то есть репродуктивный барьер определяет
видовой статус. Проигнорирована способность скрещиваться и давать
плодовитое потомство, поскольку эта способность воплощается и у таксонов
более низкого ранга (популяций). В рамках такого подхода процесс
видообразования сводится к формированию репродуктивного разрыва между
сестринскими подвидами.
Отсюда определение вида: виды – репродуктивно
изолированные естественные популяции или группы
популяций, возникающие в результате распада
предкового (стволового) вида в ходе видообразования
и прекращающие своё существование в результате
вымирания или нового акта видообразования.
19.
Эволюционная концепция(предложена
Э. О. Вили (Wiley) и Р. Мейденом (Mayden) на
основе взглядов систематика Дж. Симпсона):
Здесь вид рассматривается как своеобразный индивидуум и в рамках концепции
переживает рождение, существование и гибель. Привлекается и понятие
предкового вида, который рассматривается как «родитель» и сохраняет свой
видовой статус после видообразования. Индивидуальность вида сохраняется
токогенетическими связями.
Отсюда эволюционное определение вида: вид
– биологический
объект, состоящий из организмов, сохраняющий
свою индивидуальность во времени и
пространстве, и имеющий свою собственную
эволюционную судьбу и исторические
тенденции.
20.
Филогенетические концепции:1. Концепция Б. Мишлера (Mishler) и Э. Териота (Theriot).
Она группирует организмы в виды на основании их происхождения от общего
предка (доказательство монофилии), а репродуктивные способности вида отходят
на задний план. В качестве «предка» здесь рассматривается не предковый вид
(как в репродуктивной концепции), а таксон с более низким статусом: популяция
или даже отдельная особь. Исследуемая группа организмов наделяется видовым
статусом на основе методов кладистики и биологических критериев.
Вид – наименьшая монофилетическая группа, которая
заслуживает формального признания.
2. Концепция К. Вилера (Wheeler) и Н. Плетника (Platnick)
напротив отрицает применимость к виду филогенетических критериев. Здесь
логика строится из следующего: поскольку внутри вида отсутствуют
репродуктивные барьеры, то генеалогические связи между особями являются
сетчатыми, или токогенетическими, поэтому описание видообразования в
качестве монофилетического процесса неправильно, нужно для описания вида
применять наиболее общие параметр.
Вид – наименьшая совокупность популяций, где
происходит половое размножение, или размножение
бесполых линий, которые характеризуются уникальной
комбинацией состояний признаков.
21.
• Вид создается процессом эволюции, поэтомусуществует только эволюционный вид как
всеобщее биологическое явление.
• В традиционной трактовке вид - качественный
этап процесса эволюции.
Вид - это совокупность особей с общими
морфофизиологическими признаками,
способными скрещиваться,
представленных системой популяций с общим
ареалом (сплошным или разорванным),
формирующих совокупность экологических
ниш.
(Северная иволга:
подвиды
Балтиморская и Буллокская)
22. Основные признаки вида
1. Политипичность, то есть вид представленразнообразными по признакам группами особей.
2. Иерархичность структуры, что обусловливает
стабильность
3. Сохранение единства вида
(на основе 2 противоположных процессов)
• -дифференциации --приспособления к
микрообитаниям;
• -мобильности -- расселение и освоение ареала
4.Генетическая устойчивость, что обусловливает
независимость эволюционной судьбы
23. КРИТЕРИИ ВИДА
Фенотипическая перекрываемость признаков близкородственныхвидов затрудняет их выделение. Только сумма критериев позволяет
идентифицировать вид.
1. Морфологический критерий (недостаточность из-за видов
двойников : напр. комары, светлячки, есть среди грызунов, птиц)
2. Физиологический (иммунные реакции -иммунологич. метки")
3. Биохимический (ферменты, электрофорез белков, пигменты крови,
ДНК)
(но, внутривидовые флуктуации, синтез ам-кт разными путями у
близких видов)
4. Географический – ареал (но, совпадение ареалов, космополитические
виды и быстроразмножающиеся)
5. Экологический - своя ниша (но, перекрывание ниш)
6. Этологический - поведение жив-х, сигналы, позы, (но,проблема
изучения, аналогии)
7. Цитологический - число и форма хромосом, внутриклеточная
морфология (но, геномные и хромосомные перестройки)
8. Генетический - набор, структура генов; нескрещиваемость,
(но, межвидовые гибриды)
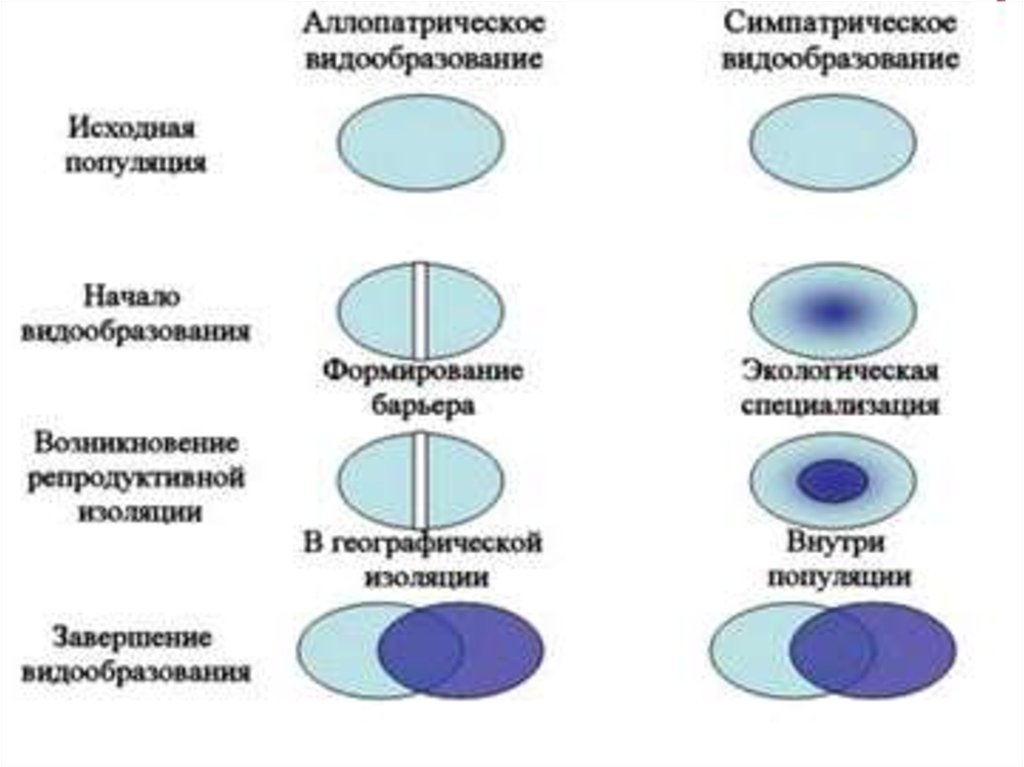
24. ПУТИ И СПОСОБЫ ВИДООБРАЗОВАНИЯ
Видообразование - это разделение вовремени и пространстве вида на 2 или
несколько.
• Или разделение генетически открытой системы
на генетически устойчивые, благодаря
микроэволюционным процессам.
Основные пути в/о:
Филетический
Гибридогенный
Дивергентный:
Аллопатрический,
Симпатрический
25. I. ФИЛЕТИЧЕСКОЕ в/о
• это изменение вида вцелом в чреде
поколений
• доказательство
эволюции видов на
историческом отрезке
времени
• изучено на
палеонтологическом
материале
26.
Филетическое направление в линииплейстоценовых европейских слонов
Ламеллярный индекс служит мерой количества эмали на коренных
зубах.
27. Дивергентный путь I. АЛЛОПАТРИЧЕСКОЕ В/О
-образование новых видов изтерриториально изолированных
разновидностей
Характерные черты:
1. в основе процессов лежат формы
пространственной изоляции
2. в/о связано с историей формирования
ареала
3. медленный путь - через сотни тысяч
поколений
28.
29.
30.
• Способы:1. Фрагментирование ареала,
а). распадение ареала широко
распространенного родительского вида
(ландыш при наступлении ледника)
В четвертичный период ареал
ландыша был разорван ледниками
на несколько самостоятельных,
изолированных частей.
Поэтому ныне ландыш в виде
обособленных форм сохранился в
Южной Европе, Закавказье, на
Дальнем Востоке.
31.
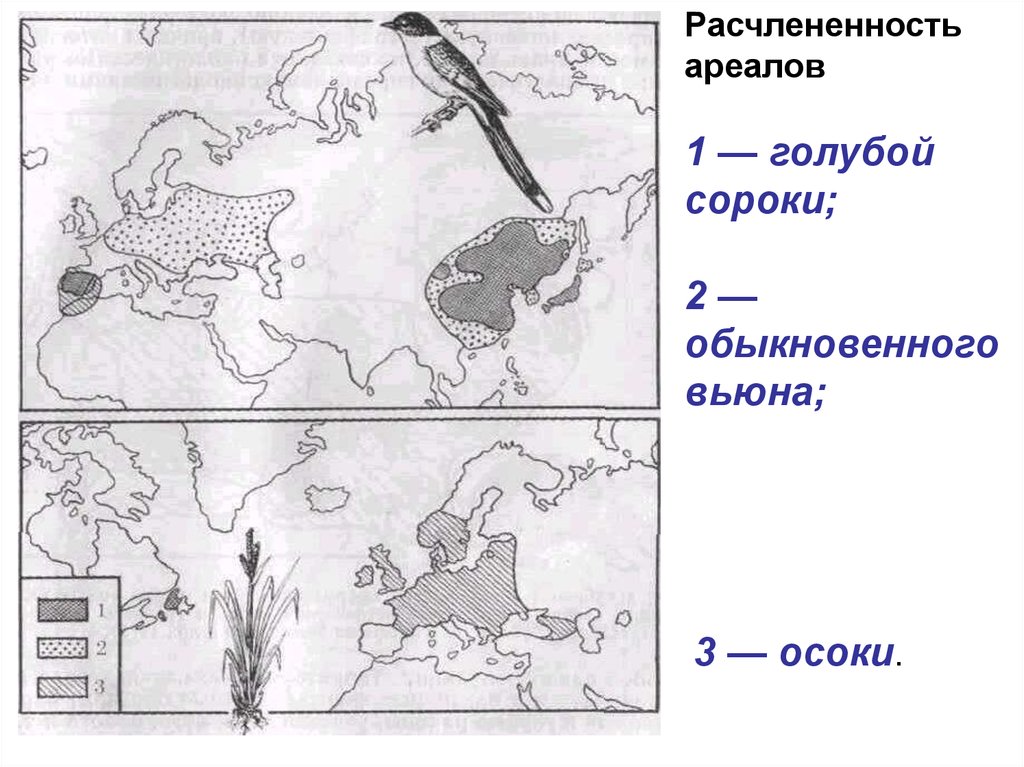
Расчлененностьареалов
1 — голубой
сороки;
2—
обыкновенного
вьюна;
3 — осоки.
32.
б). Распадение вида, при мозаичном ареале.Пример, несколько видов одуванчиков, возникшие из предкового вида
одуванчика, который занимал весь европейский континент.
• Одуванчик обыкновенный сейчас занимает лесные опушки, обочины
дорог, какие-то сорные места.
• Одуванчик – кок-сагыз произрастает в засушливых местах, : его листья
немного тоньше и больше срезаны.
• Одуванчик розовый освоил высокогорье Тян-Шаня. Он очень похож на
одуванчик обыкновенный, но отличается от него окраской лепестков.
33.
2. Расселение исходного видаА). образование новых видов на периферии
ареала и при миграции в новые условия –
периферические изоляты принцип основателя в
малой выборке –
быстрое изменение генофонда за счет дрейфа
генов + действие "катастрофического
отбора" (элиминирующий большинство особей)
на краевые популяции, обитающие
экстремальных условиях.
Выживают немногие с высокой
фенотипической устойчивостью к
неблагоприятным факторам.
Численность может восстановиться после
возвращения к норме, но генофонд уже другой.
34.
35.
Б). Перипатрическое видообразование(peripatric speciation) [греч. peri—вокруг, около,
возле и patris — родина]
Частный случай - образование небольшой
локальной популяции на границе с большой
популяцией,
сформированной несколькими особямиоснователями;
Репродуктивная изоляция возникает в результате
закрепления новых аллелей за счет дрейфа
генов.
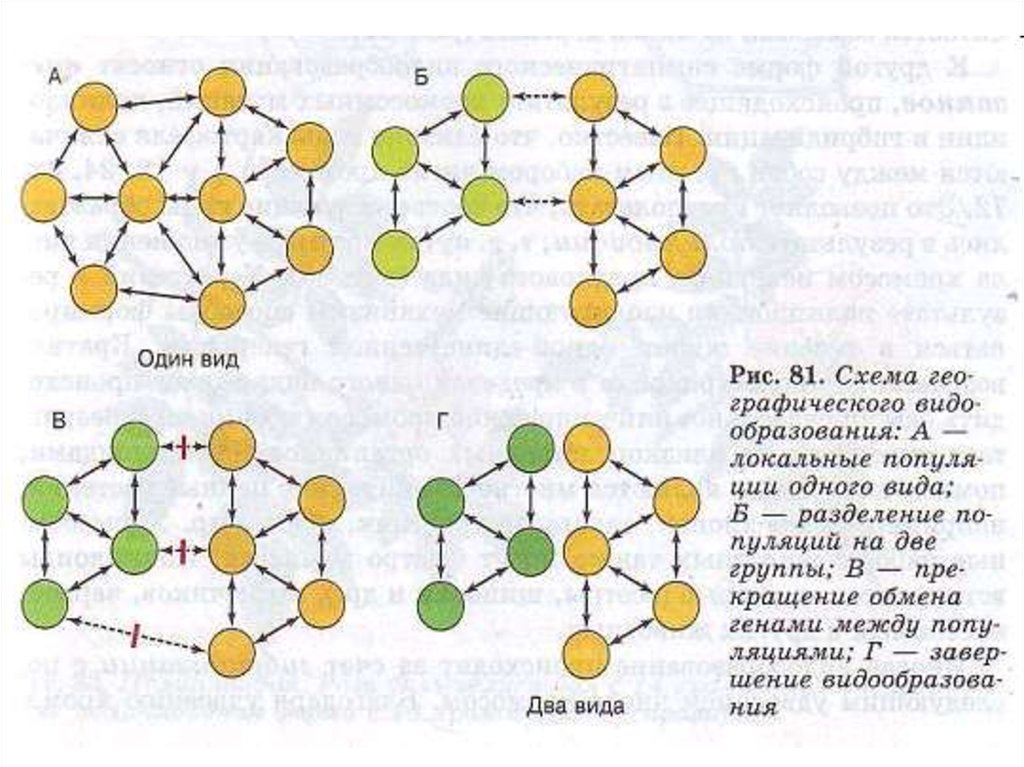
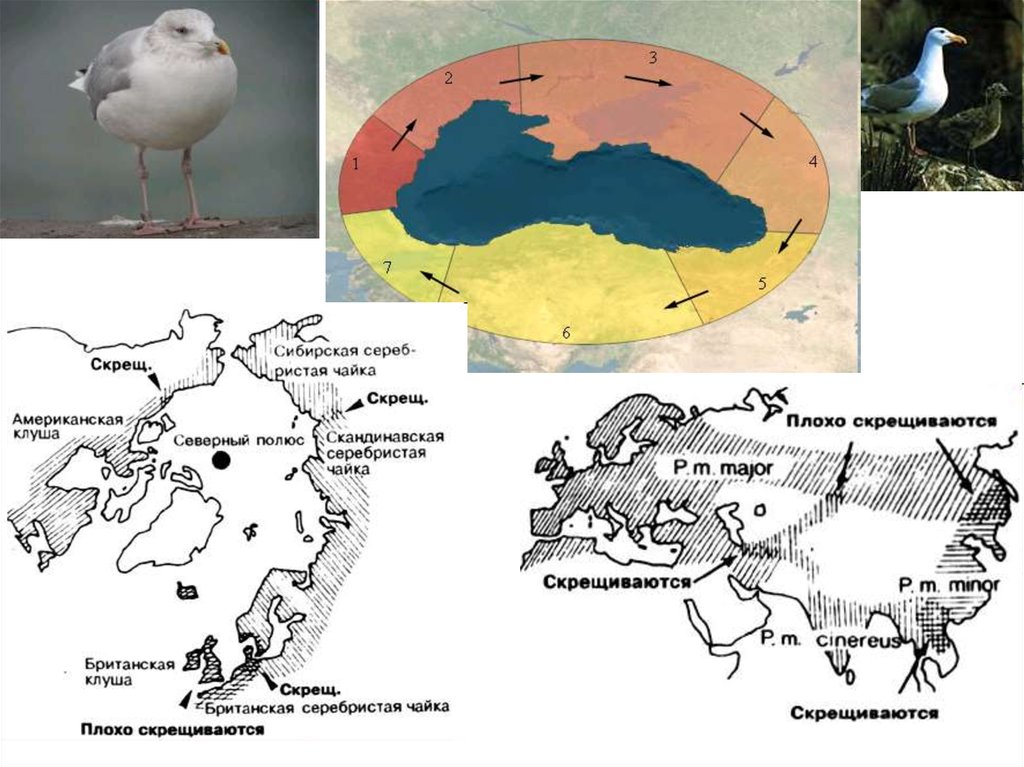
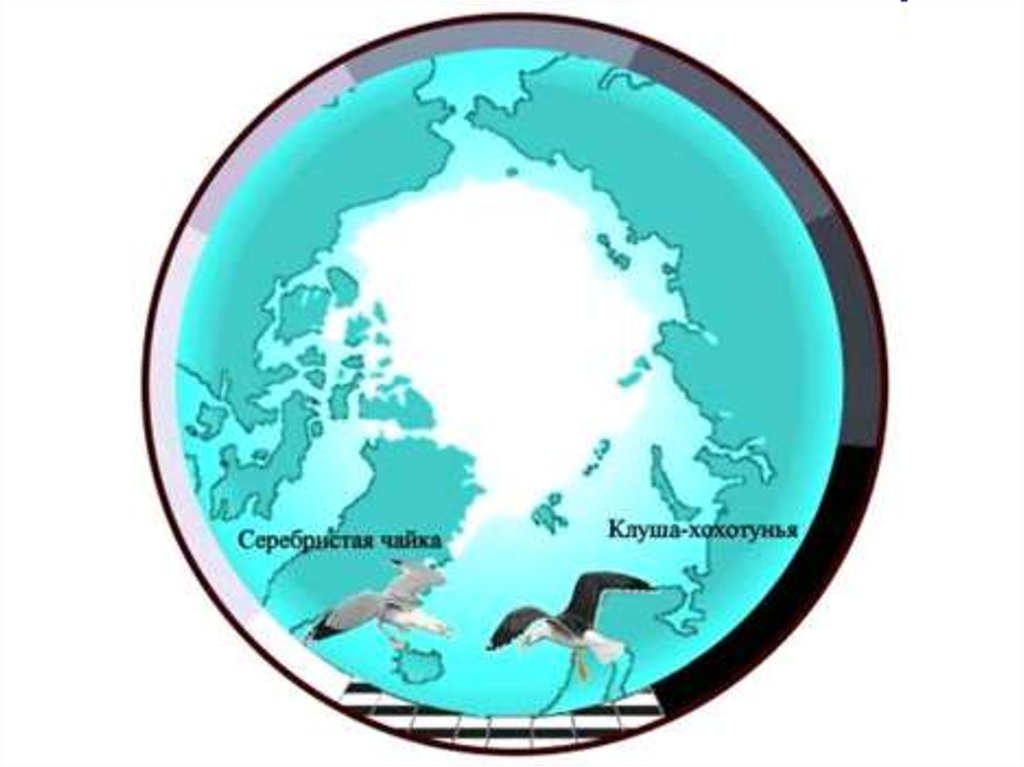
36. В). Кольцевое видообразование через цепь расселяющихся подвидов Становление серебристой чайки и клуши-хохотуньи, от Берингова
В). Кольцевое видообразование через цепьрасселяющихся подвидов
Становление серебристой чайки и клушихохотуньи, от Берингова пролива к Северному
морю)
37.
38.
39.
40.
41.
3) Путь через полиморфизм:А). При широком ареале в/о за счет
полиморфизма в популяции при
разном давлении отбора: центр
ареала К-селекция, а периферия rселекция.
Б). При разнообразии условий в
ареале: изоляция дизруптивный
отбор -> отбор в пользу
адаптивного гена (супергена) в
локальных популяциях за счет
стабильных условий. Это
адаптивная специализация.
42.
4. Специализированный способ у растений- при перенаселении.
Механизм - через обеднение состава популяции
при изреживании
варианты:
• новый генофонд самопыление новый вид
• специализация
• расширение адаптивной реакции (акклимация)
43. Дивергентный путь: II. СИМПАТРИЧЕСКОЕ в/о
- образование новых видов внутриодного вида на одной территории.
• Характерная черта: новые виды
морфологически близки
к родительскому
44.
• Разнообразие рыбцихлид озераВиктория.
• Более 500 видов
цихлид произошли
от общего предка
• в течение 12 тыс.
лет.
• отличаются друг от
друга по
морфологии,
образу жизни,
поведению
и ряду других
признаков.
45.
46.
Способы:1. Генетическая
изоляция и быстрое
обособление дочерней
формы;
а). за счет
автополиплоидии –
т.е. быстрого изменения
генотипа при его кратном
увеличении.
Мех-м: задержка расхождения хромосом под
влиянием какого-либо фактора (пример:
картофель - исх. 12, но могут 24, 48, 72 хромосомы)
47.
48.
б). Стасигенез (Уайт) - обособление прикрупномаштабных хромосомных мутаций
(перицентрические инверсии,
транслокации)
Получение потомства за счет:
- скрещивания с такими же формами
- самоопыления у растений и партеногенеза у
животных (иглокожие, членистоногие,
рыбы, ящерицы)
Такие формы могут сосуществовать
совместно или вытеснить родительский вид
за счет большей устойчивости, гетерозиса
(поэтому распространены в Арктике, в
высокогорных районах).
49.
2. Экологический способ:А) Биотопическая изоляция
Б). Сезонная изоляция
Основа: полиморфизм +
дизруптивный отбор.
Механизм - разделение экониш.
Все механизмы разделения ниш.
Разное использование пространства,
времени, пищевая специализация,
поведенческие реакции и т.д.
50.
• фиалкатрехцветная
обитает на лугах,
среди кустарников,
фиалка полевая —
на полях
фиалка собачья —
в лесах
фиалка болотная —
на лугах,
в сырых лесах,
• фиалка опушенная
— на опушках, в
оврагах, на склонах.
51.
• Речной гравилат инастоящий
гравилат
в одной местности в
природе не
скрещиваются.
• Речной гравилат
предпочитает канавы и
сырые луга,
• Настоящий гравилат, сухие места
расположения у лесных
дорог и в кустарниках.
52.
53.
В). Пищевая специализацияПример, это 5 видов синиц образовались по :
• выбору мест кормежки,
• составу кормов,
• по способам поиска и добычи.
Синица большая долбит ветви и древесные стволы и питается
крупными насекомыми; хохлатая синица питается семенами
хвойных деревьев.
Мелкие виды (лазоревка) —обследует только стебли
травянистых растений. Самые мелкие виды (московки, хохлатые
синицы) чаще обследуют в поисках корма концевые ветви
деревьев. Гаичка и московка добывают мелких насекомых в щелях
коры и в почках.
54.
Г). Занятие новых эконишв/о через расы
Яблонная муха – пример ранней стадии
симпатрического в/о.
В восточной части Сев. Америки до появления
европейцев мух развивались только в плодах
боярышника.
С завозом в Америку яблонь (1647 г.), открылась
Подобное
видообразование
произошло у
наездника - личинки
паразитируют на
личинках мух.
После появление новой
расы мух появилась и
новая раса
наездников,
предпочитающая
паразитировать на
яблонных мухах.
новая экониша . В 1864 году личинки были
обнаружены в яблоках, зафиксирована яблонная
раса этого вида.
Сейчас расы очень сильно разошлись, почти не
скрещиваются друг с другом (уровень гибридизации
не превышает 4—6 %). Яблоневая раса спаривается
почти исключительно на яблонях, а боярышниковая
— на боярышнике, что при разном
времени
созревания плодов, приводит к
репродуктивной изоляции.
В скором времени возможно выделение из этих рас
самостоятельных видов.
55.
Мухи Rhagoletis«боярышникового» вида
спариваются
исключительно на
боярышнике.
Отделившийся 150 лет
назад «яблоневый» вид
спаривается только на
яблонях
Личинки Rhagoletis pomonella в яблоке.
56.
Д). Формирование новой адаптивной нормыОпыты по выращиванию тлей Dysaphis на "стрессовых" субстратах
(малопригодных и непригодных кормовых растениях).
В опытах (Г.Х. Шапошников), популяция тлей Dysaphis anthrisci maicopica
была пересажена с того кормового растения, на котором в природе
развивается данный вид (Anthriscus nemorosa),
на другое, малопригодное кормовое растение Chaerophyllum bulbosum,
а затем на ранее совсем непригодное Ch. maculatum.
После каждого пересаживания наблюдался "кризисный" период, резко
повышалась смертность и росла изменчивость (происходила
морфологическая дестабилизация).
Затем популяция приспосабливалась к новым условиям: смертность и
изменчивость снижались, формировалась новая "адаптивная
норма", отличная от прежней (частоты морфотипов стабилизировались
вокруг новой, смещенной по сравнению с прежней, моды).
57.
Ж). Избирательное спаривание– как способ репродуктивной изоляции
Опыты по "искусственному видообразованию"
свидетельствуют о детерминированном
изменении половых предпочтений в ходе
адаптационного процесса
58.
59.
Иногда сложно вычленить пути в/оНапример, австрийская мухоловка:
распад ареала при изменении климата,
не скрещиваются в одном ареале.
изоляты ,
затем соединение ареала,
60.
61.
62. ГИБРИДОГЕННОЕ в/о
-это образование вида путемскрещивания 2-х родительских форм.
Характерная черта: новый вид
имеет признаки обоих родительских
форм.
Способ:
1. за счет аллополиплоидии,
то есть гибридизации с последующим
удвоением хромосом.
63.
64.
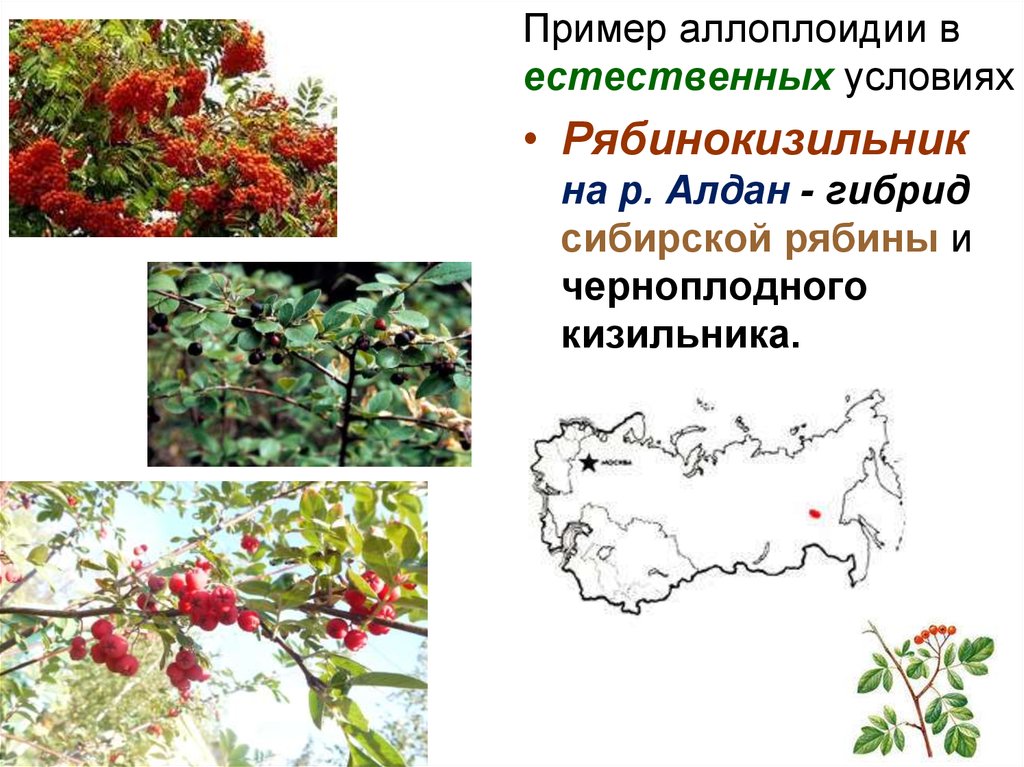
Пример аллоплоидии вестественных условиях
• Рябинокизильник
на р. Алдан - гибрид
сибирской рябины и
черноплодного
кизильника.
65.
Известна гибридизация у скальныхящериц, жаб.
При антропогенном изменении водоемов
усилилась гибридизация рыб, водных
растений (дополнительный фактор инвазии)
Известны гибриды волков и собак.
Гибридизация - основа селекции растений
- до 50% сортов
66.
Сливадомашняя
объединила
хромосомы
терна и алычи
терн
алыча
Очень редко может гибридизация может
самопроизвольно произойти в дикой природе, где
терн и алыча растут вместе.
Человек, заметив необычное растение со
съедобными плодами, ввел его в культуру.
67.
БелугаСтерлядь .
68.
ЧирПелядь
пелчир
69.
2. Амфиплоидные особи (диплоидныенаборы хромосом от каждого из
родительских видов),
• реципрокное скрещивание с
родительским,
триплоид
новый вид при условии вегетивного
размножения или партеногенеза и
гиногенеза у животных
Это партеновиды (напр.ящериц)
70.
3. Отдаленная гибридизациякомплексы взаимосвязанных форм
(саламандры:
• диплоидные бисексуальные виды +
триплоидные самки гиногенез за счет
гаплоидных спермиев родительских
видов).
• - это особая модель в/о, не настоящие
виды - клептоны.
Для процесса видообразования большое
значение имеет
"принцип основателя"
71.
72.
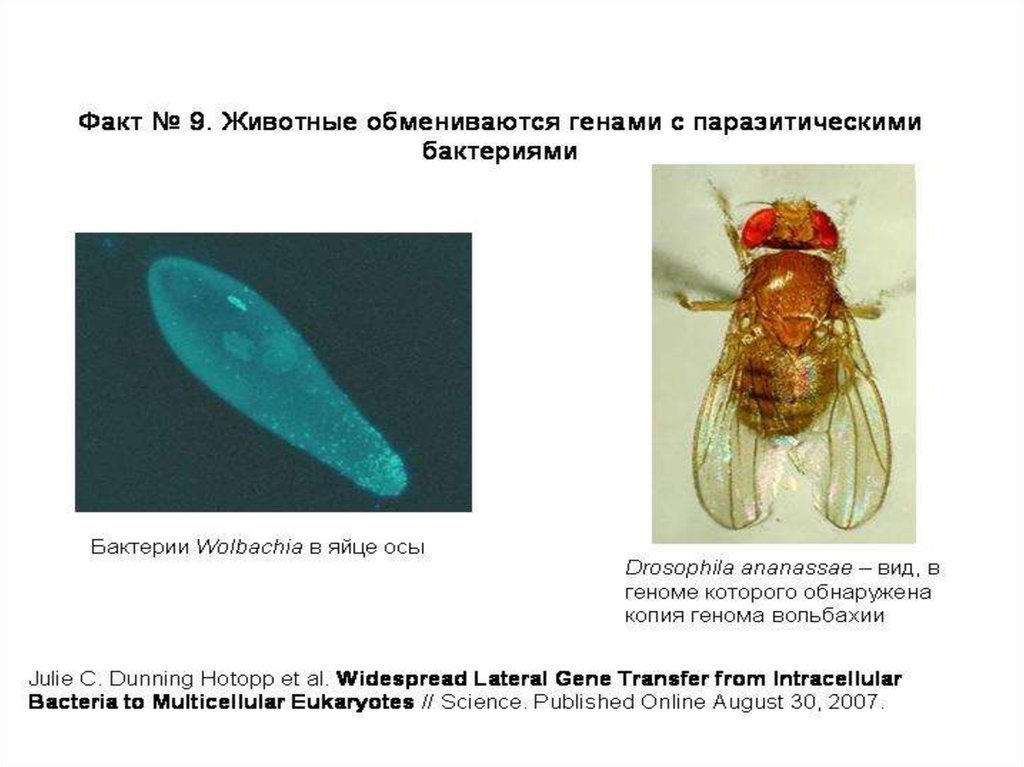
Инфекционное видообразование,которое может вызываться симбиотическими и
паразитическими микроорганизмами.
«Неканонический" изолирующий механизм, тесно связанный
с распознаванием "своих" и "чужих".
Теоретические модели инфекционного видообразования
разработаны для бактерии Wolbachia pipientis –
внутриклеточного симбионта или паразита многих видов
наземных беспозвоночных.
Вольбахия может жить только внутри клеток хозяина и
передается потомству по материнской линии, проникая в
яйцеклетки зараженных самок. Бактерии, попавшие в самца,
погибают вместе с ним и не могут перейти в следующее
поколение хозяев. Вольбахия выработала множество
способов тонкой регуляции размножения и индивидуального
развития своих хозяев. Цель – обеспечить распространение
бактерии в инфицированной популяции.
73.
74.
Наиболее широко распространенный (и, по-видимому,эволюционно самый древний) эффект вольбахии "цитоплазматическая несовместимость". Проявляется он в том,
что когда зараженный самец оплодотворяет незараженную самку,
эмбрионы оказываются нежизнеспособными и гибнут на ранних
стадиях развития.
Происходит это потому, что отцовские хромосомы в
оплодотворенном яйце ведут себя ненормально и в конце концов
разрушаются, и яйцо остается гаплоидным.
Установлено, что вольбахии, живущие в самце, каким-то образом
изменяют ("метят") хромосомы сперматозоидов. Эта "метка" и
является причиной разрушения хромосом после оплодотворения.
Однако если самка заражена тем же самым штаммом вольбахии,
отцовские хромосомы не разрушаются, и из яйца развивается
нормальная особь (естественно, зараженная). По-видимому,
присутствующие в яйцеклетке бактерии "распознают" метку и
спасают хромосомы от разрушения.
Это распознавание и "спасение" (rescue) - обычно является
штаммоспецифичным, т.е. вольбахии спасают только хромосомы,
помеченные тем же самым штаммом (хотя есть и исключения)
75. Проблема определения понятия для видов, у которых отсутствует перекрестное размножение
• Признание универсальности видовойкатегории в органическом мире не означает,
что у бесполых форм должны
существовать виды, идентичные видам,
размножающимся половым путем.
• "Вид — совокупность популяций и
клонов, эволюционирующая независимо
от других таких совокупностей".
76. Проблема определения вида при других способах размножения
Из-за отсутствия обмена генами уагамных, облигатнопартеногенетических и
самооплодотворяющихся
организмов нет генетического
единства — важной характеристики
вида.
• Однако важна не генетическая
замкнутость видов, а
генетическая устойчивость,
поэтому у таких форм вид
представлен системой близких
биотипов — группой
фенотипически сходных
особей, обладающих
близкородственным
генотипом, занимающих
общий ареал и связанных
общностью
эволюционной судьбы
77.
• Агамные (грен. агамос — безбрачный) — этоорганизмы, размножающиеся без участия
гамет. (амебы, эвглены, хлорелла, и др.).
• Клон (греч. клон - побег, отпрыск) — генетически
однородное потомство одного организма,
образовавшееся путем бинарного дробления и
преимущественно путем вегетативного
размножения.
• Облигатнопартеногенетические организмы —
это организмы, которые размножаются только
путем партеногенеза (некоторые тли,
орехотворки, кавказская скальная ящерица).
• Самооплодотворяющиеся организмы
встречаются среди плоских паразитических червей
(печеночный сосальщик, бычий цепень, свиной
солитер), а также среди растений самоопылителей
(фиалки, кислицы, некоторые виды ковыля).
78. ПРОБЛЕМЫ ВИДА И ВИДООБРАЗОВАНИЯ
Смена типологических представлений на популяционные нерешила проблему, усложнила работу систематиков
(из-за большой внутривидовой изменчивости).
В практике пользуются типологическим подходом,
выделяя отличительные признаки.
Аспекты
понятия вид
Представления
типологические
Представления
популяционные
сущность
вида
неизменность как
таксономической
единицы
вероятностный
процесс на основе
наследственности
главный
критерий
сохранение видовых изменчивость
признаков
признаков
скрещивающихся
популяций
79.
Определениевида
Совокупность
Группа
особей,
скрещивающихся
тождественных по популяций,
видовому
репродуктивно
признаку
изолированных от
других групп
Значение
репродуктивной изоляции
Условие
возникновения
вида
Побочный продукт
адаптивной
дивергенции
пространственно
изолированных
групп
Проблемы
применения
понятия
Для выделения
видов-близнецов
Как видовой
категории для
однополых
организмов
80.
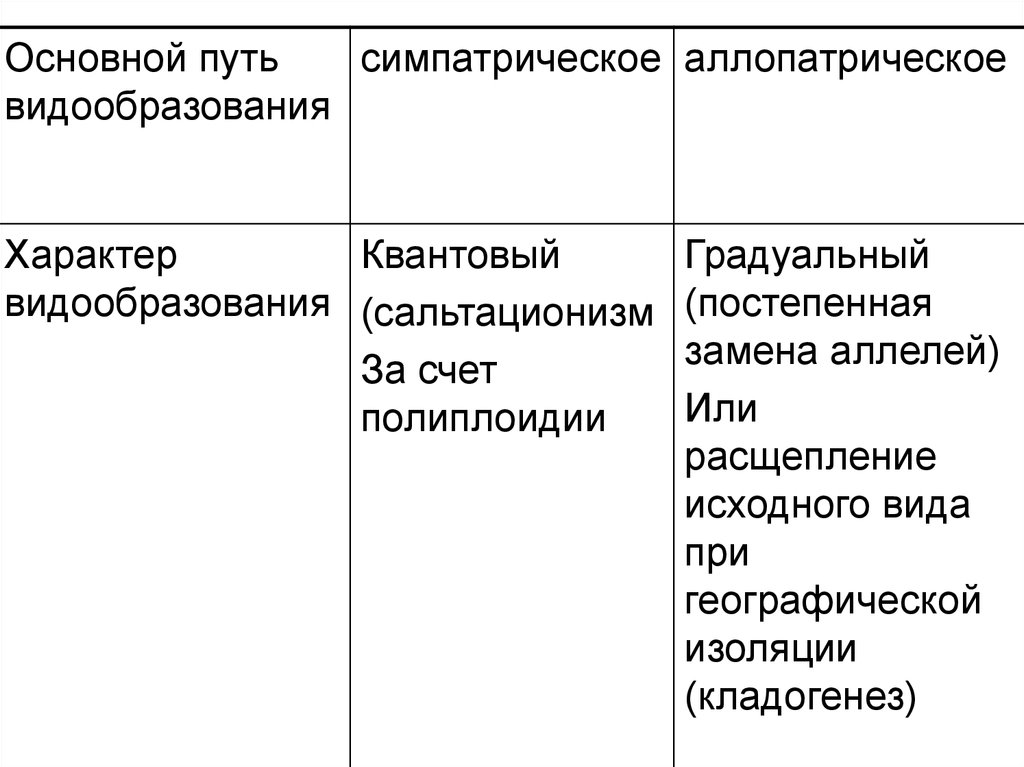
Основной путьсимпатрическое аллопатрическое
видообразования
Характер
Квантовый
видообразования (сальтационизм
За счет
полиплоидии
Градуальный
(постепенная
замена аллелей)
Или
расщепление
исходного вида
при
географической
изоляции
(кладогенез)
81.
• Появляется все больше доказательств, что в/о несводится к простой замене аллелей.
Это не объясняет варьирование генома на порядок
нуклеотидных пар у кл. птиц (10*9) и
млекопитающих (10*10).
• Может быть закрепление аббераций хромосом
(перицентрические инверсии) за счет гомозигот на
периферии ареала и через принцип основателя,
дрейф генов
(миф о Адаме и Еве -научн. осн. как гомозиготы по
мутациям)
• Отсюда, репродуктивная изоляция - не есть
побочный продукт длительной дивергенции вида,
а первичное условие возникновение вида.
(но не пространственная как в типологической
концепции, а за счет генетических
механизмов).
82.
• В/о - это следствие крупных реорганизацийгенома, сопряженных с репродуктивной
изоляцией.
• Внутривидовая изменчивость (по СТЭ материал эволюции) - "адаптивная стратегия"
для сохранения устойчивости и целостности
вида в условиях нормальных колебаний среды.
Две системы генетической изменчивости:
• группы генов, отвечающие за жизненно
важные признаки
• группы генов, отвечающие за адаптивно
менее значимые – популяционные признаки.
83.
Это основано на двойственной структурнофункциональной организации генома:1. внутренне сбалансированные
коадаптивные блоки генов или
супергены, влияющие на жизненно важные
видовые признаки. Блоки могут быть
защищены от кроссинговера инверсией или
сильными эпистатическими взаимодействиями
(их реорганизация отметается отбором).
2. свободно-скрещивающиеся локусы как
открытая генетическая система, не
влияющая существенно на
жизнеспособность и адаптивно менее
значима популяционной
(полиморфизм, клинальная изменчивость).
.
84.
Генетический мономорфизм видов сочетаетсяс системной организацией популяций, сохраняющей
уровни генетического полиморфизма
• В/о идет через реорганизацию
закрытой системы генома, а новые
виды возникают за счет основателя.
В/о можно рассматривать как чередование
периодов:
видовой трансформации при качественной
реорганизации геномов
+ с фазами длительной стабильности видов.