








































 мужская стерильность")



")


 biology
biologySimilar presentations:










Репродуктивные системы растений
1. Репродуктивные системы растений
2.
Тема 4. Репродуктивные системы растенийЗначения способа размножения растений для селекции.
Обзор способов размножения растений. Половое размножение:
циклы полового размножения, продолжительность половых
циклов, структура и морфологические особенности цветка, типы
цветков, гаметогенез, опыление и оплодотворение.
Самоопыление. Перекрестное опыление. Неполовое
размножение: вегетативное размножение, апомиксис.
Особенности биологии цветения растений в селекции:
самонесовместимость, однодомность и двудомность, мужская
стерильность.
3.
Экзаменационные вопросы.Способы размножения растений, значение для селекции.
Половое размножение: циклы полового размножения, продолжительность
половых циклов,
Гаметогенез, опыление и оплодотворение.
Самоопыление. Перекрестное опыление. Неполовое размножение:
вегетативное размножение, апомиксис.
Половая гибридизация в селекции растений.
Структура и морфологические особенности цветка, типы цветков.
Искусственная гибридизация. Цветение в гибридизации: состояние цветка,
синхронизация цветения, подбор материнского растения и подходящего цветка.
Кастрация. Опыление.
Особенности биологии цветения растений в селекции: самонесовместимость,
однодомность и двудомность, мужская стерильность.
4. значение способа размножения для селекции
Генетическая структура растений зависит от способа их размножения. Методы селекцииобычно выбирают так, чтобы природная генетическая структура вида сохранялась в
сорте. В противном случае будет необходимо предпринять ряд усилий для размножения и
культивирования нового сорта.
У цветковых растений искусственная гибридизация необходима для проведения
генетических исследований, чтобы понять наследование интересующих селекционера
признаков, и для передачи нужных генов от одного родителя к другому. Для достижения
этого, селекционер должен хорошо понимать биологию цветения и другие факторы,
связанные с цветением растений.
Искусственная гибридизация требует эффективного контроля опыления, чтобы
только пыльца нужных растений принимала участие в скрещивании. С этой целью,
селекционеру необходимо понимать функционирование репродуктивных систем видов
растений участвующих в селекционном процессе. Контроль опыления имеет решающее
значение для семеноводческой отрасли гибридов первого поколения.
Способ репродуцирования также определяет методику размножения и поддержания
сортов выведенных селекционерами.
5. Обзор репродуктивных механизмов в растениях
1. Гермафродитизм в сравнении с разнополостью. Гермафродитизм способствуетснижению генетической изменчивости, в то время как разнополость, посредством
перекрёстного опыления, способствует генетической изменчивости.
2. Самоопыление в сравнении с перекрестным опылением. Гермафродиты –
самофертильны и могут быть как самоопыляемыми так и перекрестно опыляемыми.
3. Самооплодотворение в сравнении с перекрестным опылением. Механизм
самонесовместимости у некоторых видов препятствует самоопылению, тем самым
способствуя перекрёстному опылению.
4. Половое размножение в сравнении с вегетативным размножением. Виды,
размножающиеся половым путем способны размножаться через семена. Неполовое
размножение проявляется одним из следующих способов - вегетативным размножением
(при котором семена не производятся) или апомиксисом(при котором семена
образуются).
6. Типы размножения.
Растения, как правило, подразделяют на две группы по типу репродуцирования:размножающиеся половым путем и вегетативно.
Термин бесполое размножение также применяется к явлению, когда семена образуются
без слияния гамет (так называемый апомиксис).
7. Половое размножение
Половой жизненный цикл растения (чередование поколений).Нормальный половой жизненный цикл цветкового растения можно описать как состоящую из событий
от образование семян до гибели растения (от семени до семени у видов размножающихся с помощью
семян).
Цветущее растение проходит две основные фазы развития: вегетативную и репродуктивную.
В вегетативной фазе у растения образуются только вегетативные органы (стебель, ветви, листья и т.д).
В репродуктивный период образуются цветки.
8.
У некоторых видов для перехода от вегетативной стадии крепродуктивной требуется воздействие определенных факторов
окружающей среды (например, температуры, фотопериода).
Продолжительность между фазами варьируется в зависимости от
вида и ими можно манипулировать, изменяя условия
культивирования.
Процесс полового размножения у видов, размножающихся половым
путем, состоит из двух основных процессов:
Во время первого процесса, мейоза, уменьшается число хромосом в
клетке с диплоидного (2n) до гаплоидного (n) уровня.
В течение второго процесса, оплодотворения, происходит
объединение содержимого ядер двух половых клеток, каждая из
которых имеет гаплоидный набор хромосом с образованием
диплоидной клетки.
У большинства растений, эти процессы разделены в жизненном
цикле на две отдельные фазы или поколения, между которыми они
чередуются (так называемое чередование поколений)
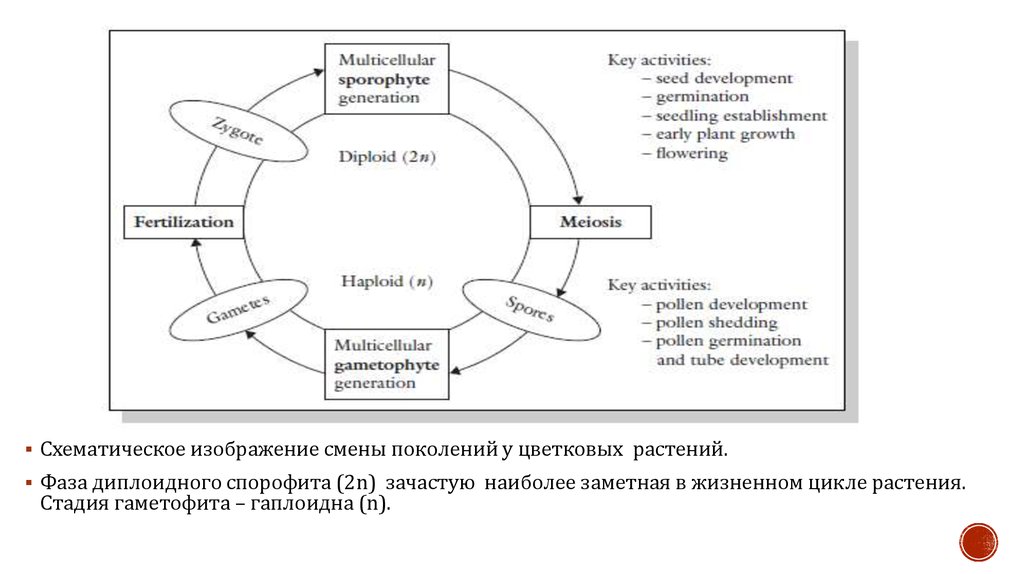
9.
Схематическое изображение смены поколений у цветковых растений.Фаза диплоидного спорофита (2n) зачастую наиболее заметная в жизненном цикле растения.
Стадия гаметофита – гаплоидна (n).
10.
Первый этап или поколение гаметофита начинается с гаплоидныхспор образующихся во время мейоза.
Клетки, полученные из гаметофита путем митоза гаплоидны.
Многоклеточный гаметофит производит гаметы путем митоза.
Во время полового процесса объединяются гаметы, образуется
зигота, с которой начинается фаза диплоидного спорофитного
поколения.
У высших растений (папоротников, голосеменных,
покрытосеменных), поколение мужского гаметофита сводится к
крошечной пыльцевой трубке и трём гаплоидным ядрам (так
называемый микрогаметофит).
Женский гаметофит (так называемый мегагаметофит) состоит из
одной многоядерной клетки, называемой также зародышевым
мешком. Генотип гаметофита или спорофита оказывает влияние на
процесс полового размножения у видов имеющих
самонесовместимость.
11. Продолжительность жизненного цикла растений
Селекционеру необходимо знать жизненный цикл растения, чтобы манипулировать с ним. На стратегиюселекционного процесса оказывает влияние продолжительность жизненного цикла растений.
12.
ОднолетниеОднолетние растения (однолетники) проходят свой жизненный цикла в течение одного
вегетационного сезона.
Примерами таких растений могут служить кукуруза, пшеница и сорго. Однолетники можно
дополнительно разделить на озимые и яровые культуры. Озимые однолетники (например,
пшеница) культивируется в течение двух сезонов. Их сеют осенью и в течение холодного
периода осень – зима - весна они подвергаются критическим физиологическим изменениям,
называемым яровизацией, которая необходима для цветения и плодоношения весной - летом.
В культуре, некоторые не однолетники (например, хлопок, анютины глазки) выращиваются,
как однолетние растения.
Двулетние
Двулетние культуры проходят свой жизненный цикл в течение двух сельскохозяйственных
сезонов.
В первом сезоне, происходит нарастание вегетативной массы корней и листьев, затем
отрастает стебель, на котором образуются цветы и плоды, и после образования семян во
втором сезоне растение погибает. Растению обычно требуются специальные экологические
условия или обработки (например, яровизации), чтобы индуцировать в конечном счёте
репродуктивную фазу. Например, у сахарной свеклы в первом сезоне происходит нарастание
вегетативной массы. Зимой она проходит стадию яровизации и весной начинается цветение с
последующим плодоношением летом второго сезона.
13.
МноголетниеМноголетники - это растения, которые повторяют свой жизненный цикл
неопределенное время в зависимости от условий окружающей среды и
принадлежности к тому или иному виду. Они могут быть травянистыми (хоста,
брунера, гейхера, многолетние кормовые травы), древесными кустарниками
(смородина, крыжовник), лианами (виноград, актинидия), и деревьями (яблоня,
слива, персик и др.).
Монокарпики
Монокарпики характеризуются повторяющимся длительным вегетативным
циклом, который может продолжаться в течение многих лет, без репродуктивного
периода. После того как происходит цветение и плодоношение, растение умирает.
Типичным примером являются бромелиевые (ананас). После цветения и
плодоношения вегетативная масса отмирает, а новые растения возникают из
корневой системы старого растения.
Следует отметить, что некоторые культуры по своей природе являющимися
двулетниками или многолетниками выращиваются как однолетние.
Например, у сахарной свеклы в первый вегетационный сезон получают в
промышленных масштабах корнеплод. В селекционно-семеноводческих целях её
подвергают яровизации и на следующий год производят семена.
14. Строение цветка
Генетические манипуляции с цветковыми растениями обычными инструментами осуществляетсяс помощью техники скрещивания, которая подразумевает под собой работу с цветками.
Структура цветка влияет на способ его кастрации (процедуры удаления пыльников необходимой
для проведения скрещивания). Размер цветка влияет на виды инструментов и методы, которые
могут быть использованы для скрещивания.
15. Морфология репродуктивных частей растения
Цветок, как правило, состоит из четырёх основных частей: лепестков, чашелистиков, тычинок и пестика.Цветки различаются по цвету, размеру, количеству и расположению этих частей. Обычно цветок имеет
цветоложе, к которому эти части крепятся.
В центре цветка располагается пестик, состоящий из рыльца, столбика и завязи (которая содержит семяпочки).
Пестик также называют гинецей.
16. Типы цветков
Обоеполый - когда тычинки и пестик находятся в одном и том же цветке (а)Однополые тычинки или пестик могут отсутствовать. (б) Если несовершенные цветки имеют тычинки они -
называются тычиночными (мужскими) цветками, если только пестик пестичными (женскими) цветками.
У однодомных (моноцийных) растений как кукуруза или огурец есть как тычиночные (мужские) так и пестичные
(женские) цветки на одном растении.
(а)
(б)
17. ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ
• пыльца, попав на рыльце пестика, начинает прорастать, образуяпыльцевую трубку
• содержимое пыльцевого зерна (вегетативная клетка и два спермия)
переходит в пыльцевую трубку
• пыльцевая трубка достигает зародышевого мешка и у большинства
растений врастает через микропиле
• при попадании в зародышевый мешок пыльцевая трубка лопается,
синергиды разрушаются и спермии попадают внутрь зародышевого
мешка, один из спермиев сливается с ядром яйцеклетки - образуется
зигота, из которой в дальнейшем развивается зародыш семени; второй
спермий сливается с диплоидным центральным ядром - образуется
триплоидное ядро, из которого затем развивается эндосперм
18. СХЕМА ДВОЙНОГО ОПЛОДОТВОРЕНИЯ
19.
Двойное оплодотворение(a) Микрогаметогенез
(b) Макрогаметогенез
20.
Микроспорогенезна ранних этапах развития генеративных почек происходит закладка тычиночных
бугорков
из верхней части бугорков развивается пыльник, а из нижней части - тычиночная нить
в пыльниках образуются крупные клетки мужского археспория (материнские клетки
пыльцы)
из археспориальных клеток пыльцы в результате двух делений мейоза возникают 4
гаплоидные микроспоры, окруженные общей оболочкой (тетрада микроспор)
Микрогаметогенез
оболочка, окружающая тетраду микроспор, распадается
вокруг каждой микроспоры образуются по две оболочки: наружная (экзина) и внутренняя
(интина)
в каждой микроспоре происходит первый митоз, в результате чего образуются
вегетативная (большая) и генеративная (малая) клетки
в последующем вегетативная клетка больше не делится и обеспечивает рост пыльцевой
трубки
генеративная клетка делится еще раз по типу митоза, в результате чего образуются 2
спермия
зрелый мужской гаметофит представляет собой пыльцевое зерно, состоящее из
вегетативной клетки и двух спермиев
21.
Макроспорогенезв завязи пестика появляется небольшой бугорок, из верхней его части формируется тело
семяпочки (нуцеллус), а из нижней части – ножка семяпочки (фуникулюс), по бокам
нуцеллуса возникают покровы семяпочки – интегументы, интегументы на верхушке
нуцеллуса не срастаются и образуют микропиле;
в субэпидермальном слое нуцеллуса закладывается крупная археспориальная клетка
(материнская клетка семяпочки);
из неё в результате 2 делений мейоза возникают 4 гаплоидные макроспоры (тетрада
макроспор), расположенных линейно по отношению друг другу; в дальнейшем 3
макроспоры дегенерируют
Макрогаметогенез
ядро макроспоры делится по типу митоза 3 раза, без цитокинеза и образуется
восьмиядерный зародышевый мешок
у полюсов зародышевого мешка (микропилярного и халазального) располагаются по 4
ядра
от каждого полюса по одному ядру отходит к центру, где они сливаются, образуя
центральное диплоидное ядро
у микропилярного полюса формируется яйцевой аппарат из трех клеток: яйцеклетки и
двух синергид
у халазального полюса – три антиподы
такой тип формирования зародышевого мешка называется нормальный или Polygonum-
тип
22. Гаметогенез.
Гаметогенез в растениях приводит к образованию пыльцы и яйцеклеток. Пыльца попадаетна рыльце женского цветка, и затем спермий через пыльцевую трубку прорастает в
зародышевый мешок, где сливается с яйцеклеткой.
23. опыление
Опыление это перенос пыльцы с пыльника на рыльце цветка.Основываясь на механизме опыления, растения могут быть
самоопыляемыми или перекрестноопыляемыми.
Самоопыляемые виды опыляются пыльцой одного и того же цветка
(автогамия). Цветки должны быть обоеполыми.
Перекрестноопыляемые виды опыляются из различных источников.
На самом деле, виды демонстрируют разную степень перекрестного
опыления, начиная от отсутствия перекрестного опыления до
облигатного перекрестного опыления
24. Таблица 1 Механизмы опыления растений.
Агент опыления (опылитель)Ветер
Насекомые
Пчёлы
Характеристика цветка
Мелкие цветки (свекла, злаки) двудомные виды
Ночные мотыльки
Цветки белые или бледно-окрашенные, хорошо заметные в ночное время, с
сильным резким запахом испускаемым после захода солнца
Жуки
Цветки белые или тусклые, крупные; одиночные или соцветия
Мухи
Бабочки
Цветки тусклые или коричневые
Яркие цвета (часто оранжевые, красные); нектарники расположены у
основания длинных тонких лепестков венчика
Летучие мыши
Большой цветок с крепкой плодо- и цветоножкой; тусклого или бледного
цвета, с сильным фруктовым или мускусным ароматом, густым обильным
нектаром
Птицы
Яркие цвета (красный, желтый) без запаха с густым обильным нектаром
Цветки яркие и эффектные (цвет синий, желтый); сладкий аромат;
уникальное строение цветка; венчик, обеспечивающий удобную посадочную
площадку
25. Самоопыление
Механизмы, способствующие самоопылениюНекоторые естественные механизмы содействуют или обеспечивают самоопыление. Клейстогамия – это состояние,
при котором цветок не открывается. Иногда, рыльце цветка тесно окружено пыльниками, что делает его склонным к
самоопылению.
Очень немногие виды являются облигатными самоопылителями. На уровень самоопыления влияют такие факторы
как, вид и количество насекомых опылителей, воздушного потока и температуры. У некоторых видов, пыльца может
стать стерильной, когда температура опускается ниже нуля. Любой цветок, который открывается до самоопыления в
некоторой степени восприимчив к перекрестному опылению.
Ячмень
Нут
Клевер
Фасоль
Хлопок
Вигна
Баклажан
Лен
Салат
Овёс
Горох
Сорго
Персик
Рис
Томат
Пшеница
соя
Hordeum vulgare
Cicer arietinum
Trifolium spp.
Phaseolus vulgaris
Gossypium spp.
Vigna unguiculata
Solanum melongena
Linum usitatissimum
Lactuca sativa
Avena sativa
Pisum sativum
Sorghum bicolor
Prunus persica
Oryza sativa
Lycopersicon esculentum
Triticum aestivum
Glycine max
26. Генетико-селекционные последствия самоопыления
Самоопыление считается высшей степенью инбридинга которое растение может достичь.Это способствует гомозиготности всех генных локусов и признаков растения. Следовательно,
последствия перекрестного опыления результатом, которого является гетерозиготность быстро
устраняется. У облигатного самоопылителя доля перекрестного опыления не должна
превышать 4%. Генотипы гамет одного растения одни и те же. Кроме того, потомство одного
растения гомозиготное (однородно). Популяция самоопыляемых видов, по существу, состоит из
смеси гомозиготных линий.
Самоопыление ограничивает создание новых комбинаций генов (нет интрогрессий новых генов
путем гибридизации). Другим генетическим следствием самоопыления является то, что
мутации (которые, как правило, рецессивны) легко доступны для селекционера или природы
вследствие гомозиготности, после применения соответствующего отбора.
Повторяющееся самоопыление не имеет генетических последствий у самоопыляемых видов
(без инбредной депрессии). Аналогично, нет такого явления как самонесовместимости.
27. Перекрестноопыляющиеся виды
Механизмы, которые способствуют перекрестному опылениюДвудомность.
Разнесённое во времени созревание тычинок и пестиков.
Самонесовместимость - пыльца цветка не прорастает на собственном рыльце
пестика.
Мужская стерильность, условие, согласно которому пыльца стерильна, что
также способствует перекрёстному опылению.
Гетеростилия тычинки и пестик значительно различаются по длине.
28. Генетико-селекционные последствия перекрестного опыления
По генетической структуре перекрёстноопыляющиеся видыхарактеризуются той или иной степенью гетерозиготности.
У таких видов характерно проявление самонесовместимости. Широкий
генофонд, из которого создаются новые комбинации при формировании
следующего поколения. Инбредная депрессия.
Летальные рецессивные аллели, которые были подавлены из-за
гетерозиготности при самоопылении имеют возможность стать
гомозиготными и, следовательно, проявлять летальные признаки.
Явление гетерозиса.
29.
Бесполое размножениеБесполое размножение подразделяется на вегетативное размножение и
апомиксис.
Вегетативное размножение
Как указывалось ранее, некоторые виды размножаются с помощью
различных вегетативных частей: луковицами, клубнелуковицами,
корневищами, стеблями и почками. Вегетативное размножение
широко используется в садоводстве. Для посадки используют небольшие
отрезки растений - черенки, которые делают из различных частей
растения (например, корень, стебель, листья).
30. Возможности селекции при вегетативном размножении
1. Вегетативно размножаемые (клональные) виды с жизнеспособными семенами и высокой фертильностьюпыльцы могут быть улучшены путем гибридизации.
2. Клональные сорта могут использоваться в производстве сразу же после скрещивания, при условии, что в них
сочетаются желаемые комбинации признаков. Клональная селекция, следовательно, намного быстрее.
3. В селекции видов, товарным продуктом которых являются вегетативные части растений, не важно, чтобы
гибрид был фертильным.
4. Вследствие большого коэффициента размножения вегетативно размножающихся культур (с помощью
методов, таких как черенкование или микроклональное размножение), селекционеру достаточно получить
всего лишь одно растение с комплексом желательных признаков для использования в качестве источника
дальнейшего репродуцирования.
5. Гетерозисный эффект (гибридная сила) сразу же фиксируется в полученном селекционером гибриде. Один
раз, полученный гетерозисный генотип сохраняться неопределенно долго.
6. Достаточно трудно получить большое количество посадочного материала из клонов за короткий промежуток
времени.
7. Некоторые виды растений, которые являются исключительно партенокарпическими (например, банан), не
могут быть улучшены путем гибридизации.
8. Такие виды как манго и цитрусовые продуцируют многозародышевые проростки. Эта репродуктивная
неравномерность усложняет селекционный процесс, так как материнские клоны смешиваются с гибридным
потомством.
9. Клональные культуры являются многолетними перекрёстниками и неиспытывают инбредную депрессию, так
как обладают высокой гетерозиготностью.
31. Апомиксис
- явление, при котором без слияния мужской и женской гамет, образуетсяжизнеспособный зародыш, и развиваются «вегетативные семена».
В результате семена образующиеся в процессе апомиксиса являются клонами
материнского растения. То есть, апомиксис- это разновидность бесполого образования
семян. В отличие от полового размножения, при апомиксисе нет возможности получить
новые рекомбинации генов и следовательно разнообразия в потомстве.
Партеногенез:
генеративный n, Из гаплоидной, неоплодотворенной яйцеклетки.
соматический 2n. Из диплоидной яйцеклетки, возникшей на основе диплоспории, при
которой материнская клетка мегаспоры развивается в зародышевый мешок либо без
мейоза вообще, либо с видоизмененным мейозом, когда уменьшения числа хромосом
при делении клетки не происходит.
Апогамия :
генеративная n, Из синергиды или антиподы гаплоидного зародышевого мешка.
соматическая 2n. Обычно в результате слияния двух клеток зародышевого мешка или
диплоидной синергиды или антиподы, возникших на основе диплоспории.
Адвентивная эмбриония 2n. Из соматической клетки нуцеллуса или интегументов
семяпочки при параллельном развитии нормального зародышевого мешка.
32. Преимущества для селекционера
Путём апомиксиса гибридные семена могут быть получены из растенийвыращенных из гибридных семян.
Селекционеру не нужно проводить скрещивание родительских линий каждый
год, чтобы получить гибридные семена. Это преимущество ускоряет
селекционные программа и снижает затраты на выведение и размножение
новых гибридов.
Существуют некоторые проблемы в селекции растений, связанные с
апомиксисом. Виды, обладающие факультативным апомиксисом, являются
более сложными для использования в селекции, поскольку в их потомстве
присутствуют растения из семян полученных половым путём (амфимиксис) и
апомиксисом. Облигатные апомикты легче использовать в селекции
обычными способами, при условии совместимости родительских линий
(бесполое размножение).
33. Биологические ограничения полового размножения в селекции растений
Некоторые биологические ограничения полового размножения используется вкачестве инструмента в селекции растений. К ним относятся двудомность,
однодомность, самонесовместимость, и мужская стерильность.
Одним из важнейших аспектов селекции и семеноводства является контроль
опыления для обеспечения участия только нужной в данном скрещивании пыльцы.
При производстве гибридных семян, успех зависит от наличия эффективной,
надежной, практичной и экономически оправданной системы контроля опыления.
Опыление может контролироваться тремя основными способами:
1 Механический контроль.
2 Химический контроль.
Примерами химических веществ могут быть Dalapon®, Estrone®, Ethephon®,
Hybrex®, и Generis®. Применение этих агентов вызывает мужскую стерильность у
растений, тем самым, обеспечивая перекрестное опыление.
3 Генетический контроль.
34. Самонесовместимость
(или отсутствие завязывания семян присамоопылении) явление, при котором пыльцу с цветка не
воспринимает рыльце того же цветка и, следовательно, не
образуются семена.
Реакция несовместимости генетически обусловлена локусом S, с
множеством аллелей, которые могут насчитывать более 100 у
некоторых видов, например у клевера лугового (Trifolium pratense).
35. Cистемы самонесовместимости
Системы самонесовместимости можно разделить на два основных типа:гетероморфную и гомоморфную.
1 Гетероморфная несовместимость.
Это связано с различиями в длине тычинок и столбика пестика (так называемый
гетеростилия) (рис.).
Один тип цветка называют контактный, столбики
длинные, а пыльники короткие (томат). Другой тип
цветка, называют тычиночный, столбики короткие, а
пыльники длинные (например, у примулы (Primula)).
36.
2 Гомоморфная несовместимость.Есть два вида гомоморфной несовместимости - гаметофитный и
спорофитный.
При гаметофитной несовместимости способность пыльцы
функционировать определяется генотипом пыльцевого зерна, а не
растения, которое его продуцирует.
Аллели гена (генов) несовместимости действуют самостоятельно в
столбике пестика. Они не проявляют доминирования. Несовместимая
пыльца подавляется в столбике. Пестик диплоидный и, следовательно,
содержит два аллеля несовместимости (например, S1S3, S3S4).
Несовместимость проявляется, если встречаются идентичные аллели
как в пыльце так и в столбике. Только гетерозиготные S аллели
работают в этой системе.
37.
Рисунок 4.6 Виды самонесовместимости: (а) спорофитный и (б) гаметофитный.Спорофитная несовместимость обнаружена у сложноцветных и крестоцветных.
Она связанна с наличием двух генеративных ядер в пыльцевых зернах, тогда как
при гаметофитной несовместимости одно генеративное ядро в пыльцевой
трубке, как например у различных видов клевера.
38.
При спорофитной несовместимости, несовместимость пыльцыопределяется растением (спорофитом), который её производит. Эта
система несовместимости встречается у таких видов, как редис и
капуста.
Спорофитная система отличается от гаметофитной тем, что S аллели
проявляют доминирование.
Кроме того, несовместимость может иметь индивидуальное
действие, как в пыльце, так и в столбике пестика, что делает систему
несовместимости комплексной.
Доминирование определяется опылителем. Несовместимая пыльца
может ингибироваться на рыльце пестика.
Например, растения с генотипом S1S2 где S1 является
доминирующим аллелем над S2 (S1>S2), будет производить пыльцу,
которая будет функционировать как S1. Кроме того, пыльца с
аллелем S1 не будет прорастать в столбике пестика имеющего такой
же S1 аллель, а в столбике с аллелем S2 будет прорастать.
39. Использование самонесовместимости в селекции растений
Бесплодие любого рода препятствует селекции растений. Тем не менее, этотнедостаток может быть использован используя такие методы, как удаление рыльца
пестика (или применения электрошока), раннее опыление (до действия
ингибирующих белков), или инструмента для содействия определенным методам
селекции. Самонесовместимость может быть преодолена понижением температуры
(замедление развития ингибирующего вещества).
Самонесовместимость способствует гетерозиготности. Следовательно,
самоопылением самонесовместимых растений можно создать ряд растений
обладающих значительной изменчивостью, из которых селекционер может отобрать
лучшие рекомбинантные комбинации.
Были установлены системы самонесовместимости для производства гибридных
семян у некоторых культур (например, капуста, редис), которые проявляют
спорофитный тип несовместимости (рис. 4.7). Самонесовместимые инбредные линии
используются в качестве родителей. Эти системы, как правило, используется для
контроля за опылением при коммерческом производстве гибридных семян.
40.
Спорофитная несовместимость широкоиспользуется в селекции капусты и
других видов Brassica.
Простые гибриды более однородны и
проще в производстве семян. Обычно
используется топкросс.
Самонесовместимый родитель
используется в качестве материнского
компонента скрещивания, и
фертильная отцовская линия в
качестве источника пыльцы
(опылителя)
41. Мужская стерильность
- это явление, при котором у растенияпыльники или пыльца нефункциональны.
Может проявляться чаще всего как отсутствие или наличие очень
небольшого количества пыльцы, серьёзного уродства или
отсутствия цветков или тычинок, или невозможности
растрескивания пыльников.
Так же, как и самонесовместимость, мужская стерильность
обеспечивает перекрёстное опыление.
42. Мужская стерильность
1. Истинная мужская стерильность. Связана с однополыми цветкамиу которых отсутствуют тычинки (при двудомности или
однодомности) или если у обоеполых цветков ненормальные или
нефункциональные микроспоры (что приводит к абортированию
пыльцы).
2. Функциональная мужская стерильность. Пыльники не в
состоянии растрескиваться, хотя пыльца является фертильной
(вбрычанский тип, Джон Бер тип у томата).
3. Индуцированная мужская стерильность. Селекционеры могут
использовать химические вещества, чтобы вызвать стерильность
(например гиббереллины у томата).
Истинная мужская стерильность
Существует три вида стерильности пыльцы – ядерная (ЯМС),
цитоплазматическая (ЦМС), и ядерно-цитоплазматическая (ЯЦМС).
43. Генетическая (ядерная, генная) мужская стерильность
Генетическая (ядерная, генная) мужская стерильность широко распространена урастений.
Ген стерильность был обнаружен у ячменя, хлопка, сои, томата, картофеля и фасоли.
Считается, что почти у всех диплоидных и полиплоидных видов растений есть хотя бы
один мужской локус стерильности. Генетическое мужское бесплодие может проявляться
в абортации пыльцы или аномальном развитии пыльников.
Генетическая (ядерная, генная) мужская стерильность зачастую обусловлена одним
рецессивным ядерным геном ms, доминантный аллель, MS, способствует образованию
нормальных пыльников и фертильной пыльцы.
Однако, мужская стерильность у люцерны, как сообщается, находится под контролем
двух независимо наследуемых генов. Экспрессия гена стерильности может меняться с
изменением условий окружающей среды.
Для успешного применения в селекции растений, система мужской стерильности
должна быть стабильной в широком диапазоне условий окружающей среды и
полностью ингибировать образование семян.
44.
Селекционер в случае ядерной стерильности неможет производить и поддерживать популяцию,
имеющую 100% растений с мужской стерильностью.
Растения, обладающие генетической мужской
стерильностью имеют генотип (msms) и могут
размножаться путем скрещивания их с
гетерозиготными (Msms) фертильными растениями.
Этот скрещивание будет давать потомство, в котором
50% растений будет с мужской стерильностью (msms)
и 50% с мужской фертильностью (Msms).
Если скрещивание будет изолировано, то
селекционеры всегда будут получать 50% растений
обладающих мужской стерильностью, собирая семена
только с растений с мужской стерильностью.
45. Цитоплазматическая мужская стерильность
Цитоплазматическая мужская стерильность (ЦМС) контролируетсягенами цитоплазмы (митохондриальным геном).
ЦМС передается только через яйцеклетку (материнский фактор).
ЦМС была обнаружена у таких видов как кукуруза, сорго, сахарная
свекла, морковь, капуста и лён.
Эта система имеет реальные преимущества в селекции
декоративных видов, потому что все потомство получается с
мужской стерильностью и следовательно растения не образуют
плодов и семян.
Без плодов, декоративные растения обладают более
продолжительным цветением и дольше сохраняют свежий вид
(декоративность).
46.
Использование в селекции цитоплазматической мужскойстерильности.
N - нормальная цитоплазма, S - стерильная цитоплазма.
47. Ядерно-цитоплазматическая мужская стерильность (ЯЦМС)
Используется в производстве гибридных семян лука и морковиЭтот тип стерильности определяется взаимодействием генов цитоплазмы и ядра
У стерильной линии стерильная цитоплазма (S) сочетается с гомозиготным по
рецессивному аллелю гена стерильности ядром (msms).
Специально путём многократных возвратных скрещиваний выводят линию закрепитель
стерильности, обладающую точно таким же набором генов, что и стерильная линия за
исключением нормальной цитоплазмы (N).
Третьим компонентом скрещивания при получении семян F1 гибрида является фертильная
родительская линия опылитель имеющая нормальную цитоплазму и доминантные аллели
гена стерильности в ядре (MsMs) или (Msms).
При выращивании гибридных растений F1 культур у которых в пищу используют плоды, к
гибридным семенам необходимо подмешивать семена сорта-опылителя
48.
S msmsN msms
Получение F1 гибридных семян
S msms Материнская стерильная линия
S msms
S msms
N msms
N msms
S msms
N MsMs
S msms
Линия – закрепитель стерильности
N MsMs
Другая линия
N MsMs
Размножение стерильной линии
F1S Msms
Схема получения гибридных
семян на основе ЯЦМС
N msms
49. Использование мужской стерильности в селекции
Мужская стерильность в основном используется в качествеинструмента в селекции растений для замены кастрации при
гибридизации.
Селекционеры используют мужскую стерильностью для защиты
своих авторских прав (особенно гибриды полученные с помощью
ЦМС (полностью стерильные) и ядерной функциональной мужской
стерильностью.