










































 biology
biologySimilar presentations:










Селекция сортов самоопыляющихся культур
1. Селекция сортов самоопыляющихся культур.
2.
• Генетические основы методов создания селекционных популяцийНа втором этапе селекционного процесса проводят отбор. Под
этим термином в селекции понимают, два мероприятия.
• 1. Сам отбор, т.е. оценку растений селекционного материала и
выделение лучших из них в качестве родоначальников будущих
потомств.
• 2. Применение разных методов получения от отобранных растений
семенных или вегетативных потомств.
• В процессе отбора на основе генофонда популяции исходного
материала селекционер создает новые - сортовые популяции.
• Структура сортовых популяций у разных видов зависит от способа
размножения растений.
• Сортовые популяции самоопыляющихся растений представляют
собой чистые линии или смесь очень близких чистых линий.
• Способ размножения растений определяет структуру создаваемой
сортовой популяции.
• При селекции самоопыляющихся растений, необходимо возможно
скорее достичь гомозиготности селектируемого материала, чтобы
иметь возможность определить качество будущего сорта.
3.
4.
• При планировании работ по выполнению второго этапа селекционногопроцесса с самоопыляющимися растениями необходимо учитывать
следующие обстоятельства:
• 1) количество выделяемых в исходном материале ценных растений
зависит от трудоемкости культуры, количества селекционного
материала, физических возможностей селекционера и его помощников
и, наконец, наличия необходимой для выращивания растений площади
и техники.
• Количество выделенных ценных растений также определяется
насыщенностью ими исходного материала. В сортовых и полученных
в результате мутагенеза популяциях их может быть единицы, в
лучшем случае десятки.
• При использовании в качестве исходного материала второго
гибридного потомства количество ценных растений может
исчисляться сотнями и тысячами.
• Все зависит от удачного подбора генотипов родительских сортов и
семенной продуктивности гибридов первого поколения.
• Объем популяции второго гибридного потомства можно получить
практически любой величины, получая семена от нужного количества
растений первого поколения.
5.
• в селекции самоопыляющихся растений методом гибридизацииприменяют скрещивание высококачественных довольно близких
по большинству признаков сортов, так чтобы расщепление шло по
возможно меньшему числу генов.
• 2) в течение какого числа поколений следует проводить пересев
селекционного материала (до проведения отбора) для достижения
полной гомозиготности.
• Гетерозиготность выделенных ценных растений в случае
использования в качестве исходного материала сортов и
индуцированного мутагенеза может быть по небольшому числу
генов, следовательно их гомозиготность может быть достигнута через
несколько поколений пересева.
• Гибридные потомства таких сравнительно мало трудоемких и
требующих небольшую площадь питания культур, как горох и салат
обычно выращивают в количестве нескольких тысяч и выделяют, в
качестве родоначальников будущих потомств, ценных растений
порядка нескольких сотен.
6.
• При самоопылении в инбредных потомствах от гетерозиготырасщепление на гомозиготы и гетерозиготы по одному гену можно
определить по формуле:
(2n-1 - 1)AA : 2Aa : (2 n-1 -1)aa;
• где: n - число поколений.
• Долю гомозигот по обеим аллелям можно определить, преобразовав
формулу суммированием долей гомозигот и отнеся полученную
величину к объему гибридной популяции данного поколения.
• (2 n-1 - 1)AA + (2 n-1 - 1)aa = (2 n -2)AA+aa
• Отсюда доля гомозигот по обоим аллелям гетерозиготного локуса в
потомстве будет равна:
(2n - 2)/2n; где: n - число поколений.
• Для определения доли гомозигот по всем локусам необходимо
полученную для одного гена величину возвести в степень равную
числу пар гомологичных хромосом. Так, для достижения неполной (на
уровне 95% растений) гомозиготности по всем хромосомам для гороха
n = 7, необходимо 8-м поколений пересева. (Вероятность
гомозиготности по одной хромосоме равна 0,992, по 7-и хромосомам 0,9927=0,945).
• Таким образом, пересев логичнее вести до 7-8-го поколения.
7.
• Из полученного в результате пересева селекционного материалавыделяют ценные растения в количестве нескольких десятков и даже
сотен и получают от них семена будущих семей.
• Число выделенных родоначальников семей зависит как от уровня
насыщенности ими селекционного материала, так и возможности
провести объективное их испытание
• Каждая семья представляет группу растений гомозиготных по одним и
тем же аллелям полигенов и точно отражает генетическую природу
будущей сортовой популяции, которая будет от нее получена.
• Лучшие семьи размножают и получают от них линии, которые
испытывают в конкурсном и станционном сортоиспытании.
• Затем лучшую из них или группу линий передают в государственное
сортоиспытание.
• Некоторые селекционеры в качестве исходного материала используют
не второе гибридное потомство, а третье или четвертое.
• Это может быть обусловлено тем, что во втором бывает недостаточное
количество растений, а также меньшей гетерозиготностью более
поздних потомств.
8.
• Часто некоторые селекционеры не выдерживают бездеятельности припересеве селекционного материала в течение 7-8 лет и применяют
семейственный отбор в ранних поколениях.
• В этом случае, степень гетерозиготности некоторых растений
родоначальников семей выявляется при испытании их потомств по
расщеплению.
• При селекции высоко трудоемких культур, например, выращиваемых
через рассаду томата, перца и баклажана, в каждом поколении,
начиная с третьего и кончая седьмым, применяют семейственный
отбор.
• В каждом поколении в лучших семьях отбирают лучшие растения и
используют их в качестве родоначальников будущих семей.
• Такая схема второго этапа в какой-то мере позволяет вести работу
только с лучшими растениями и семьями, но очень затрудняет работу
по исключению гетерозиготности селекционного материала.
• Более того, если первоначально высокое качество гибрида было
обусловлено, в основном, за счет гетерозиготности, то она, благодаря
непрерывному отбору лучших растений в каждом поколении, будет
постоянно сохраняться в селекционном материале.
• В какой-то мере проблема устранения гетерозиготности решается
оценкой семей в последних поколениях отбора при оценке их
выравненности и выбраковке расщепляющихся.
9.
• Наиболее сложно выполнение второго этапа селекционного процессапри использовании исходного материала, полученного в результате
отдаленной гибридизации.
• Как показали исследования Соловьевой Н.А. (1970), при работе с
исходным материалом томата, полученного от скрещивания
культурного томата с L. hirsutum, расщепление по морфологическим,
биологическим и хозяйственным признакам наблюдалось во всех
поколениях отбора вплоть до девятого.
• Отбор лучших, скороспелых и урожайных растений не обеспечивал
получение семей такого же качества.
• В этой ситуации выполнение второго этапа селекционного процесса
также осуществляется методами, аналогичными применяемыми при
селекции перекрестноопыляемых растений.
10.
• Типы самоопыляющихся сортов• С точки зрения генетической структуры, существуют два типа
самоопыляющихся сортов:
• 1. полученные из одного растения.
• 2. полученные из смеси растений.
• Отбор одного растения может или не может предшествовать
запланированному скрещиванию, но часто это имеет место.
• Сорта, полученные из одного растения, гомозиготны и однородны.
• Тем не менее, сорта, полученные из смеси растений могут быть
однородными, но, так как отдельные растения имеют различные
генотипы, и в небольшом количестве происходят скрещивания у
большинства самоопыляющихся видов, позже в популяции появляется
некоторая гетерозиготность.
• Методы селекции самоопыляющихся видов могут быть разделены на
две большие группы - те что предшествует гибридизации и которые
осуществляются после гибридизации.
11.
• Общие обозначения в селекции растений• Селекционеры используют сокращения для облегчения работы в своих
селекционных программах. Некоторые символы являются
стандартными генетическими обозначениями, тогда как другие были
разработаны селекционерами.
• К сожалению, нет ни одной всеобъемлющей и универсальной системы
использования этих обозначений.
• Символы применяемые в скрещиваниях:
• 1. F. Символ F обозначает потомство от скрещивания между двумя
родителями. Индекс (х) представляет собой поколение (Fx) . Если
родители являются гомозиготными, то поколение F1 будет
однородным. Скрещиванием двух растений F1 (или самоопылением
F1) получают поколение F2 (F1 х F1 = F2). Посеяв семена растений F2
получают популяцию F2, самое разнообразное поколение после
скрещивания, в котором селекционеры часто начинают отбор. После
самоопыления растений F2 получим семена F3 растений, и так далее. 2
2. ⊗. Символ ⊗ обозначает самоопыление.
• 3 S. Обозначение S также используется с числовыми индексами. В
одном случае S0 = F1; в другой системе S0 = F2.
12.
• Символы для инбредных линий• Инбредные линии обозначаются символами F, I либо S описываются
двумя системами.
• Система I описывает инбредную линию основываясь на поколении
растений которые в настоящее время растут.
• Система II описывает как поколение растения, из которого возникла
линия, так и поколение растений которые в настоящее время растут.
Чтобы провести различия между этими двумя системами используются
следующие варианты:
• Вариант 1. Базовая популяция F2. Селекционер отбирает растение из
популяции F2 собирает с него семена F3 и высевает их в следующем
сезоне.
• В системе I: из высеянного семени вырастает F3 линия.
• В системе II: из высеянного семени вырастает F3 линия полученная из
линии в F2 или F2: 3 линия.
• Таким же образом используется обозначение S
• Система I: S1 линия.
• Системе II и: S0 даёт линию S1 и S0: 1 линия.
13.
• Обозначения родословной• Знание родословной или происхождение сорта позволяет селекционеру
проследить последовательность операций в селекционной программе.
• Селекционеры для написания родословных растений следуют краткой
системе обозначений. Некоторые родословные простые, другие
сложные. Некоторые из общепринятых обозначений следующие:
• 1. косая черта, /, указывает на скрещивание.
• 2. цифры между косыми чертами, / 2 /, указывают на
последовательность или порядок скрещивания. / 2 / эквивалентно / / и
указывает на второе скрещивание. Аналогично, / первое скрещивание,
и / / / третье скрещивание.
• 3 обратное скрещивание обозначается как *; *3 указывает что генотип
подвергался обратному скрещиванию три раза с другим генотипом.
• Следующие примеры могут быть использованы для иллюстрации эих
обозначений.
• Родословная 1: MSU48-10/3/Pontiac/Laker/2/MS-64.
• Интерпретация:(а) Первое скрещивание было Pontiac (в качестве
женского компонента) х Laker (в качестве мужского компонента).
• (б) Второе скрещивание было [Pontiac / Laker (как женщина)] х MS-64
(как мужчина).(с) третье скрещивание MSU48-10 (как женщина)
×[Pontiac / Laker / / MS-64 (как мужчина)].
• Родословная 2: MK2-57 *3/SV-2. Эквивалентно формуле: MK2-
14.
• Следующие примеры могут быть использованы для иллюстрации этихобозначений:
• Родословная 1: MSU48-10/3/Pontiac/Laker/2/MS-64.
• Интерпретация:
• (а) Первое скрещивание было Pontiac (в качестве женского компонента)
х Laker (в качестве мужского компонента).
• (б) Второе скрещивание было [Pontiac / Laker (в качестве женского
компонента)] х MS-64 (в качестве мужского компонента).
• (с) третье скрещивание MSU48-10 (в качестве женского компонента)
×[Pontiac / Laker / / MS-64 (в качестве мужского компонента)].
• Родословная 2: MK2-57 *3/SV-2.
• Эквивалентно формуле: MK2-57/3/MK2-57/2/MK2-57/SV-2.
Интерпретация: генотип MK2-57 был подвергнут обратному
скрещиванию (беккроссу) три раза с генотипом SV-2.
15.
• Массовый отбор• Целью массового отбора является улучшение популяции за счет
увеличения частоты желательных генов.
• Отбор основан на фенотипе растения и необходимо одно поколение в
каждом цикле. Массовый отбор совершается один или несколько раз
(периодический (рекурентный) массовый отбор).
• Улучшение у самоопыляющихся культур ограничено генетической
изменчивостью в первоначальной популяции (то есть, новая
изменчивость не образуется во время селекционного процесса).
• Целью при создании сорта с помощью массового отбора является
повышение средних показателей по сравнению с базовой популяцией.
• Использование. Как современный метод селекции растений, массовый
отбор применяется в следующих случаях:
• 1. Для поддержания сортовой чистоты, при сортовом засорении или
расщеплении. Нетипичные растения просто выбраковываются из
популяции, а семена собираются с оставшихся растений.
Существующие сорта загрязняются на протяжении многих лет в
результате природных процессов (например, вследствие переопыления,
мутаций) или человеческого фактора (например, случайного
смешивания семян во время уборки или на разных этапах доработки
семян).
16.
• 2. Для выведения сорта из базовой популяции путём гибридизации.• 3. Для сохранения идентичности текущих сортов или кандидатов в сорта.
Селекционер отбирает несколько сотен (200-300) растений и высаживает их
на отдельные делянки для оценки. Делянки, демонстрирующие
значительные фенотипические отличия от других делянок бракуются, а с
остальных производят сбор семян в общую тару. Перед объединением
семян, семена с элитных растений собираются с каждой делянки, и
хранятся для будущего использования при размножении оригинального
сорта.
• 4. Когда новая культура интродуцируется в новую область производства,
селекционер может адаптировать её к новым регионам, проведя отбор по
ключевым признакам, необходимым для успешного производства
(например, скороспелости).
• 5. Массовый подбор можно использовать в селекции сорта на
горизонтальную (полевую) устойчивость к болезням. Селекционер
использует низкую концентрацию инокулиума патогенна, стимулируя
умеренное развитие заболевания, для того чтобы оценить полигенные
генетические эффекты (вместо главных эффектов гена).
• Таким образом, создаётся сорт, обладающий расонеспецифической
средней толерантностью к заболеванию. Кроме того, он будет обладать
стабильной урожайностью и долговечной устойчивостью к болезням.
• 6. Некоторые селекционеры используют частично массовый отбор в своих
селекционных программах для отбраковки нежелательных растений, тем
самым сокращая исходный материал и экономя время и затраты на
17.
• Процедура• Главной процедурой при проведении в массового отбора является
выбраковка нетипичных растений или растений с нежелательными
признаками.
• Такой отбор некоторыми исследователями называется, негативным
массовым отбором.
• Конкретные стратегии по работе с популяцией меняются в
зависимости от вида, интересующих признаков или творческого
подхода селекционера, для того, чтобы найти способы, облегчающие
селекционные программы.
• В то время как браковка и массовый сбор семян, как представляется,
являются основными стратегиями массового отбора, некоторые
селекционеры могут отбирать и продвигать большое количество
растений, которые являются желательными и однородными по
интересующим их признакам (положительный массовый отбор).
• Там где это применимо, можно собирать для посева единичные семена
с каждого растения.
18.
19.
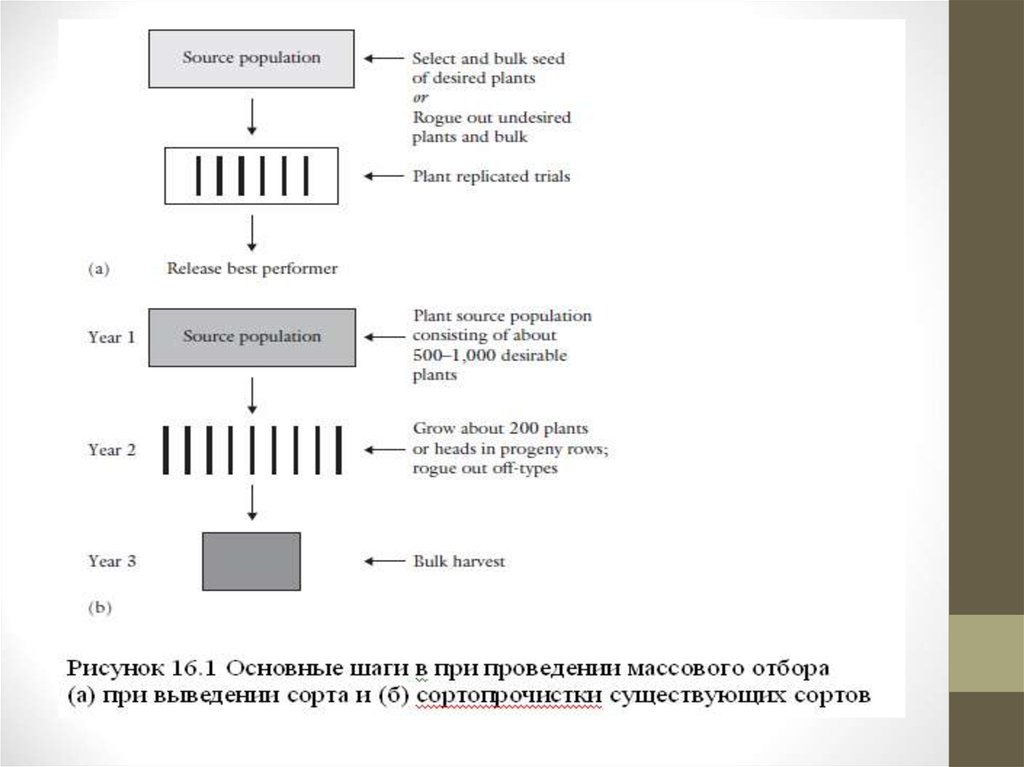
• Селекционер высевает гетерогенную популяцию в поле, и ищетнетипичные растения, чтобы их удалить и уничтожить (рис. 1).
• Таким образом, оригинальная генетическая структура сохраняется в
максимально возможной степени.
• Могут быть использованы механические устройства (например, сито
для определения оптимального размера зерна), или отбор может быть
проведён по визуальной оценке селекционера.
• Кроме того, отбор может быть основан на целевых признаках (прямой
отбор), либо проводится опосредованно путем отбора признаков
коррелирущих с улучшаемым признаком.
• 1 год. Если цель состоит в очищении созданного сорта от примесей,
семена отобранных растений высеваются рядками, чтобы подтвердить
чистоту отобранных растений до сбора семян в общую тару.
Оригинальный сорт необходимо посадить рядом для сравнения.
• 2 год. Проводят оценку семян в повторяющихся испытаниях, с
использованием оригинальных сортов для сравнения. Этот тест может
быть проведен в различных местах и в течение нескольких лет. Семена
собираются в общую тару.
20.
• Генетические последствия• Засорение от переопыление может привести к появлению гетерозигот в
популяции.
• К сожалению, в случае если доминантные эффекты генов участвуют в
экспрессии признака, гетерозиготы неотличимы от доминантных
гомозигот.
• В естественно самоопыляющихся популяциях гетерозиготы
расщепляясь в следующих поколениях способствуют появлению
нетипичных растений.
• Массовый отбор наиболее эффективен, если выражение желаемого
признака обусловлено аддитивным действием генов.
• В самоопыляющихся популяциях , от одного поколения к следующему
с помощью естественного инбридинга изменяется частота гетерозигот
в пользу гомозигот.
• Массовый отбор основан на фенотипе растения. Следовательно, он
наиболее эффективен если интересующий признак имеет высокую
наследуемость.
21.
• Преимущества• 1 Быстро, просто, и понятно. Могут быть оценены большие популяции
в течении одного поколения.
• 2 Относительно недорого.
• 3 Сорт получается фенотипически достаточно однороден и
представляет собой смесь чистых линий, что расширяет его
генетическую основу и делает его более адаптивным и стабильным.
• Недостатки
• 1. Для большей эффективности, желательные признаки должны иметь
высокую наследуемость.
• 2. Поскольку отбор основан на фенотипе, оптимального результат
достигается, если отбор проводится в однородной окружающей среде.
• 3. Фенотипическая однородность меньше, чем у сортов полученных с
использованием чистых линий.
• 4. Вследствие доминированя, гетерозиготы неотличимы от
гомозиготных генотипов. Без тестирования потомства, отобранная
гетерозигота будет расщепляться в следующем поколении.
22.
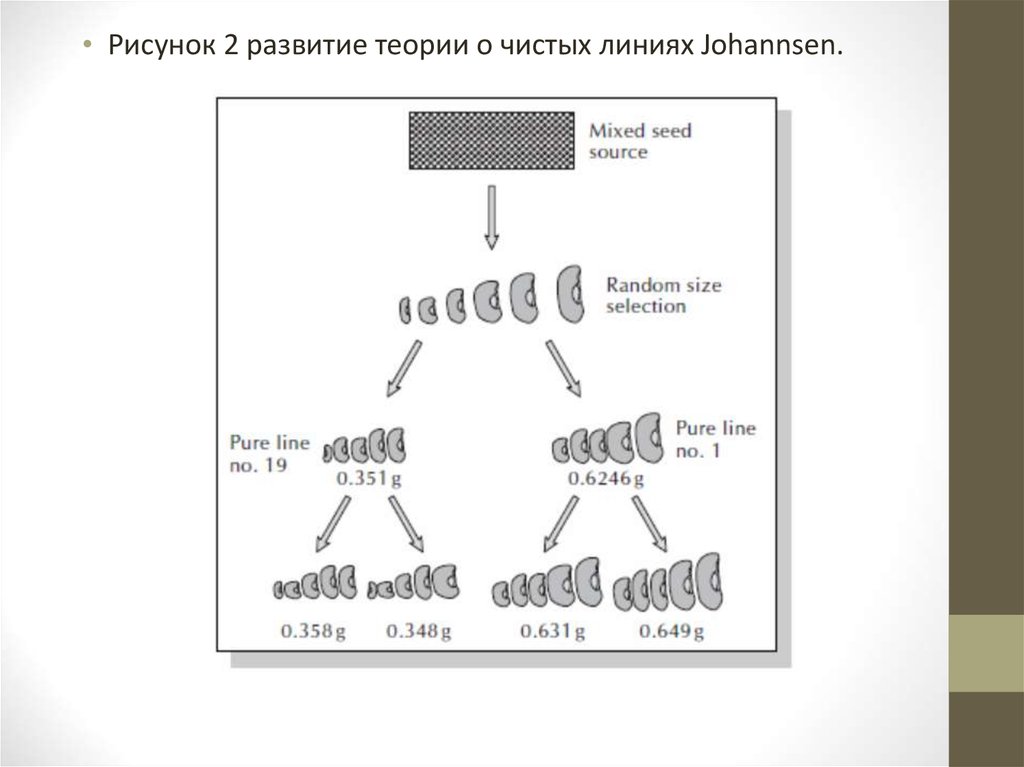
• Селекция чистых линий• Теория создания чистых линий была разработана в 1903 году датским
ботаником Иогансеном. Проводя оценку по массе семян фасоли, он
показал, что смешанные популяции самоопыляющихся видов можно
разобрать на генетически чистые линии. Однако эти линии были
впоследствии оказались нечувствительными к отбору внутри каждой
из них (рис. 2).
• 1. Линии, которые генетически отличаются могут быть успешно
изолированы из популяции смешанных генетических типов.
• 2. Любые изменение, происходящие в чистой линии не наследственны,
а связаны только с изменяющимися факторами окружающей среды.
• Линии имеют важное значение для многих селекционных программ.
• Они используются в качестве сорта или как родители при производстве
гибридных семян (инбредные линии).
• Кроме того, линии используются при создании генетического фонда
(содержат конкретные гены отвечающие за устойчивости к болезням
или содержание питательных веществ), а также и синтетических и
мультилинейных сортов.
23.
• Рисунок 2 развитие теории о чистых линиях Johannsen.24.
• Ключевые особенности• Линейным сорт, по определению считается, если коэффициент родства
составляет, по крайней мере 0,87.
• Определение «чистая линия» предполагает, что сорт имеет одинаковые
аллели во всех локусах.
• Практически такое состояние генотипов недостижимо. По мнению
таких исследователей как K.J. Frey в генотипах сортов состоящих их
различных чистых линий наблюдаются высокие темпы возникновения
мутаций.
• Линейные сорта имеют очень узкую генетическую базу и, как правило,
однородны по интересующим селекционера признакам (например,
высота, скороспелость).
• В случаях возникновения имущественных споров, линии легко
однозначно идентифицировать.
25.
• Использование• Селекция чистых линий желательна при создании сортов для
определенных целей:
• 1. Сорта для механизированного производства, которые должны
соответствовать определенной спецификации для равномерной
эксплуатации сельскохозяйственных машин (например, равномерное
созревание, одинаковая высота растений).
• 2. Сорта создаваемые для товарного рынка, который ставит во главу
угла внешнюю привлекательность продукции (например, равномерная
форма, размер).
• 3. Сорта для рынка переработки (например, спрос на определенные
стандарты консервирования, текстуры).
• 4. Продвижения сортов "спортов", которые появляются в популяции
(например, мутант цветок для декоративных целей).
• 5. Улучшение недавно одомашненных культур, которые имеют
некоторую изменчивость.
• 6. Селекция чистых линий также является неотъемлемой частью
других методов отбора, таких как отбор на основе родословной и
массовый популяционный отбор.
26.
• Процедура• Отбор чистой линии в селекции представляет собой повторяющиеся
циклы самоопыления, после первоначального отбора из смеси
гомозиготных линий. Природные популяции самоопыляющихся видов
состоят из смесей гомозиготных линий с гетерозиготами,
появляющимися вследствие мутаций и скрещиваний.
• Последовательность операций
• 1-й год. Первым шагом является создание вариабельной базовой
популяции (например, интродуцированной популяции,
расщепляющейся популяции от скрещиваний, местных сортов) её
высадки, проведении отбора и сбора семян с лучших растений (рис. 3).
• 2-й год. Выращивание потомств отобранных растений. Выбраковка
любых отклонений. Сбор семян индивидуальных потомств.
• 3-6 года. Проведение предварительных испытаний, выход
экспериментальных линий, включение соответствующих сортов
стандартов для сравнительной оценки.
• 7-10 года. Проведение испытаний лучших потомств в нескольких
местах. Выпуск наиболее продуктивных линий в качестве новых
сортов.
27.
• Рисунок 3 Процедура селекции чистых линий.28.
• Преимущества• 1. Быстрый метод селекции.
• 2. Относительно недорогой. Базовой популяцией могут служить
местные сорта. Выбранный размер популяции может быть
малым или большим, в зависимости от объекта селекции.
• 3. Сорт созданный этим методом имеет высокую
выравненность.
• 4. Он применим при улучшении признаков имеющих низкую
наследуемость, потому что отбор основан на четком проявлении
признака в потомстве.
• 5. При отборе чистой линии, отбираются только лучшие чистые
линии для максимального генетического прогресса.
29.
• Недостатки• 1. Сортовая чистота, может быть нарушена из-за механических
примесей, переопыления с другими сортами и мутаций.
• 2. Сорт-линия имеет узкую генетическую базу и, следовательно,
восприимчив к неблагоприятным внешним факторам.
• 3. Не образуется новых генотипов. Улучшение ограничено
изоляцией от наиболее желательных или лучших генотипов из
смешанной популяции.
• 4. Метод способствует генетической эрозии.
• 5. Эксплуатация делянок с потомствами требует больше
ресурсов (времени, пространства, денег).
30.
• Отбор по родословной• Отбор по родословной является широко используемым методом
селекции самоопыляющихся культур.
• Основное отличие отбора по родословной от массового отбора или
отбора чистых линий состоит в том, что используется гибридизация
для генерации изменчивости (создание базовой популяции. Этот метод
был впервые описан H. H. Lowe в 1927 году.
• Отбор по родословной способ селекции, при котором селекционер
ведет учет происхождение сорта.
• Численность популяции, по необходимости, устанавливается путем
скрещивания отобранных родителей, а затем регулируется
проведением интенсивного отбора.
• Документирование родословной позволяет селекционеру проследить
путь от родителей к потомству и, в конечном счете, к индивидуальному
F2 растению из любого последующего поколения.
• Чтобы добиться успеха, селекционер должен уметь различать
желательные и нежелательные растения основываясь на фенотипе в
расщепляющихся популяциях. Это метод непрерывного
индивидуального отбора после гибридизации и продолжается до
достижения желаемого уровня гомозиготности.
• На этом этапе появляются фенотипически однородные растения.
31.
• Использование• Отбор по родословной применяется в селекции видов, которые
позволяют проводить отбор индивидуальных растений, которые
должны соблюдаться, описал и собирали отдельно. Он используется
при селекции арахиса, табака, томата, некоторых зерновых культур,
особенно там, где легко идентифицируются качественные признаки
нуждающиеся в улучшении.
• Процедура. Ключевыми шагами процедуры отбора по родословной
являются:
• 1. Создание базовой популяции, путём скрещивания отобранных
родительских пар.
• 2. Высадка потомств отобранных растений.
• 3. Проведение тщательной оценки отобранных растений из поколения
в поколение.
32.
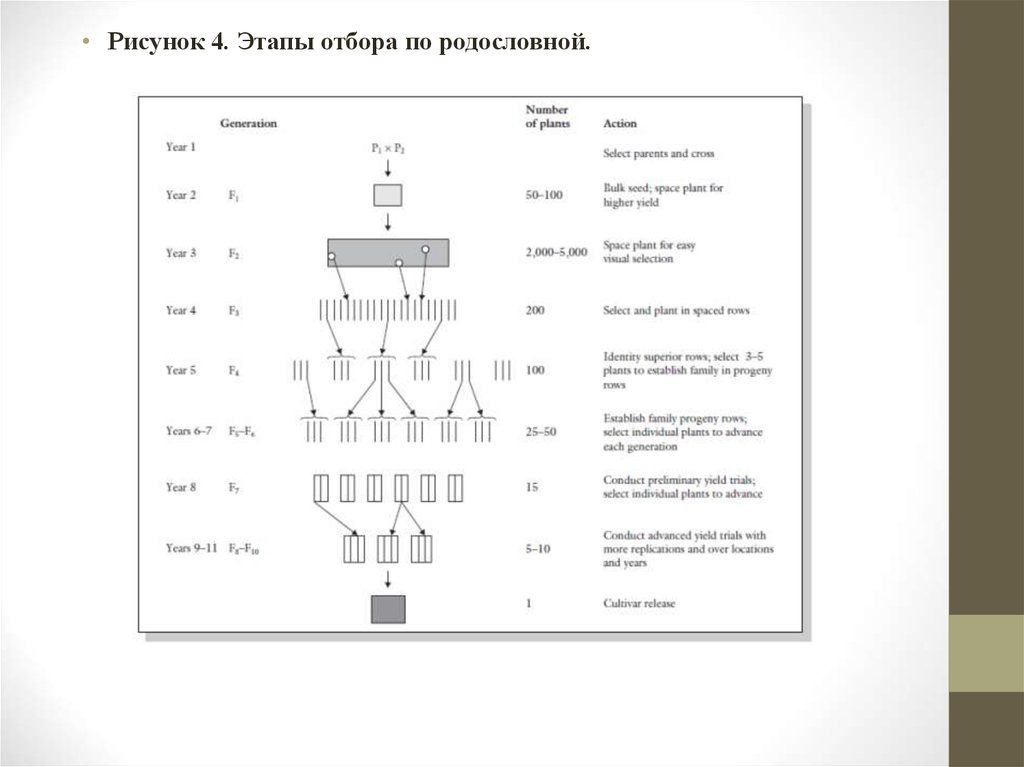
• Последовательность• 1 Год. Определение гомозиготных родителей и проведение около 20200 скрещиваний (рис.4).
• 2 Год. Выращивание 50-100 F1 растений, включая родителей для
сравнения и идентификации гибридов.
• 3-й год Выращивание примерно 2000-5000 F2 растений. Посадка
растений таким образом, чтобы была возможность провести оценку и
описание отдельных растений. Включение сортов стандартов для
сравнения. Лучшие растения отбираются и семена собираются
отдельно учитывая их идентичность.
• 4 Года. Семена от высших растений высеваются отдельными рядками в
F3-F5 поколениях, убедившись, что схема посева удобна для учета.
Отбор на данном этапе внутри и между делянками, определив сначала
лучшие делянки и отбор 3-5 растений из каждого потомства для
посадки следующего поколения.
• 5 Год. К концу F4 поколения, должна быть 25-50 рядков с записями
каждого растения и ряда.
• 6 Год. Отдельные семьи высаживают в F6 для производства
экспериментальных линий для проведения предварительных
сортоиспытаний в F7. Стандартным сортом должен быть
адаптированный к местным условиям сорт или сорта.
33.
• 7 Год. Сортоиспытание в разных местах, регионах, и в разныегоды, проводится в F8-F10 поколениях, оценивая только лучший
экспериментальный материал для следующего поколения.
• В конечном счете, цель состоит в отборе одной или двух линий,
которые превосходят стандартный сорт для выпуска их в
качестве нового сорта.
• Следовательно, оценка в производственных испытаниях должна
выявить улучшенное проявление признаков, которые имеют
агрономических значение для успешного производства
конкретной культуры (например, устойчивость к полеганию,
устойчивость к осыпанию, устойчивость к болезням).
• После того как по результатам сортоиспытания будет отобрана
лучшая линия начинается стандартный процесс по подготовке
допуска сорта к реализации (т.е. размножение семян и
сертификация).
34.
• Рисунок 4. Этапы отбора по родословной.35.
• Генетические вопросы• Подробные записи хранятся от одного поколения к другому в
отношении происхождения и других характеристик растений.
• Метод позволяет селекционеру создать генетическую изменчивость в
ходе процесса. Следовательно, селекционер может влиять на
генетические вариации родителей.
• Этот метод применяется при селекции на вертикальную устойчивость
к болезням, а не на горизонтальную. Продукт (сорт) имеет
относительно узкую генетическую базу, но не исключительную, как у
чистых линий.
• Преимущества. 1. Ведение учета по родословной обеспечивает
генетической информацией о сорте, которая недоступна при
использовании других методов отбора.
• 2. Отбор основан не только на фенотипе, но и на генотипе
(потомственные линии), что делает эффективным отбор улучшенных
линий среди расщепляющихся потомств растений.
• 3. Используя данные учёта, селекционер может вести отбор в
потомствах линий, несущих гены отвечающие за проявление
желательного признака.
• 4. Там где это необходимо создаются сорта с высокой степенью
генетической чистоты (например, для сертификации продукции на
определенных рынках).
36.
• Недостатки• 1. Ведение учёта медленный, утомительный, трудоемкий и дорогой
процесс. Отбор по родословной оказывает давление на ресурсы
(например, например увеличивается площадь земельных участков
необходимая для обеспечения размещения посадок растений удобных
для наблюдения и оценки). Посев и сбор семян трудоемкие операции.
Тем не менее, широко использующееся современное оборудование для
исследования селекционного участка и сбора семян значительно
упрощает оценку, ведение наблюдений и записей. Следовательно
большие популяции растений могут быть обработаны без особого
труда.
• 2. Этот способ отбора не подходит для видов, у которых отдельные
растения трудно выделить и описать.
• 3. Отбор по родословной является длительной процедурой,
требующей около 10-12 и более лет при использовании только одного
вегетационного периода.
• 4. Этот метод больше подходит для селекции на качественную
(вертикальную), чем количественную (горизонтальную) устойчивость
болезням.
• 5. Отбор в F2 ( раннее тестирование поколений) на основе
количественных признаков, таких как продуктивность не эффективен.
Более эффективно проводить отбор из F3 линии посаженной рядами,
чем проводить отбор отдельных растений в F2.
37.
• Массовый популяционный отбор• Массовый популяционный отбор - это метод по улучшению
сельскохозяйственных культур, при использовании которой в ранних
поколениях используется влияние естественного отбора, а
искусственный отбор проводится в более поздних потомствах.
• Шведский учёный H. Nilsson-Ehle впервые разработал процедуру, а H.
V. Harlan со своими коллегами предоставил дополнительное
теоретическое обоснование этого метода, в рамках своей селекционной
работы с ячменём в 1940 году.
• По предложению Харлана, метод основан на тестировании потомств F2
от массовых скрещиваний и отказа от скрещиваний основанных на
фенотипическом проявлении продуктивности.
• Другими словами, основной целью является разделение скрещиваний
по отбору родителей на основе оценки продуктивности.
• В настоящее время этот метод используется для других целей.
38.
• Ключевые особенности• Основанием для отсрочки проведения искусственного отбора является
предоставление давлению естественного отбора (например, абиотических
факторов, таких как засуха, холод) устранить или снизить продуктивность
менее приспособленных генотипов в популяции.
• Так же, как и при отборе по родословной, этот метод позволяет выводить чистолинейные сорта, основываясь на теории чистых линий для расщепляющихся
популяций.
• Генетические рекомбинации в гетерозиготном состоянии не могут быть
использованы у самоопыляющихся видов, так как самоопыление постепенно
увеличивает гомозиготность потомства.
• При проведении самоопыления в F6 гомозиготность составляет уже около
98,9%. Стратегия селекции растений заключается в задержке проведения
искусственного отбора пока не будет достигнут высокий уровень
гомозиготности.
• Использование массового популяционного отбора
• Наиболее подходит для селекции самоопыляющихся видов, которые, как
правило, имеют высокую плотность посадки в производстве (например,
злаковые культуры - пшеница, ячмень).
• Также используется в селекции бобов и сои.
• Тем не менее, он не подходит для улучшения плодовых и многие овощных
культур, у которых конкурентоспособность нежелательна.
39.
• Процедура• После проведения скрещиваний, от нескольких сотен до нескольких
тысяч отобранных в F2 растений плотно высаживают по заданной, как
правило, обычной густоте посадки.
• Семена со всего участка собираются в общую тару.
• Часть семян используется для посева на другом участке поля для
проведения следующего отбора, подвергая посадки давлению
естественного отбора в течение следующих 2-3 поколений.
• В F5, растения высеваются менее густо, чтобы позволить провести
эффективную оценку и обор индивидуальных растений.
• Предварительные сортоиспытания можно проводить в F7 с
последующими производственными сортоиспытаниями и выпуском
сорта для производства.
• Последовательность
• 1-й год Идентификация желательных родителей (сортов, потомств
простых скрещиваний и т.д.) и проведение необходимых количеств
скрещиваний между ними (рис.5).
• 2-й год после проведения скрещиваний соответствующих родителей,
около 50-100 F1 растений высаживают и собирают с них семена в
общую тару, после проведения браковки.
40.
• Рисунок 5. Этапы массового популяционного отбора.41.
• Генетические вопросы• Повторяющиеся из поколения в поколение самоопыления могут
привести к трем основным результатам:
• 1. В продвинутых поколениях, растения будут гомозиготными почти по
всем локусам.
• 2. Значение средней популяционной будет улучшена в результате
естественного отбора.
• 3. Генотипы с высокими показателями хозяйственно-ценных признаков
будут сохранены в популяции.
• Массовый популяционный отбор способствует интергенотипической
конкуренции. Позволяя естественному отбору работать в более ранних
поколениях, частоты генов в популяции в каждом поколении будет
зависеть от:
• 1. Генетического потенциала генотипа по продуктивности.
• 2. Конкурентоспособности генотипа.
• 3. Влияния окружающей среды на экспрессию генотипа.
• 4. Пропорций и типов генотипов в следующем поколении (т.е.
выборки).
42.
• Влияние этих факторов может меняться от одного поколения кдругому. Более того, вполне возможно, что желательные генотипы
могут быть вытеснены из популяции более агрессивными
нежелательными генотипами. Например, высокие растения могут
заглушить низкие желательные растения. Невозможно предсказать,
какие растения F2 будет представлена в следующем поколении, ни
предсказать генетическую изменчивость для каждого признака в
любом поколении.
• Роль естественного отбора в массовом популяционном отборе не
неопровержима. Это, как предполагается, играют важную роль в
генетических сдвигах в сторону конкурентных типов, в основном из-за
высокой плодовитости конкурентоспособных видов. Такое воздействие
приемлемо, когда учитываются признаки, которые придают
преимущество за счет устойчивости к биотическим и абиотическим
стрессам. Например, подвергая базовую популяцию различным
стрессовым ситуациям (например, засолению, холодным
температурам, заболачиванию, засухе, изменению фотопериода)
плодовитость может быть очень низкой у плохо приспособленных
генотипов. Эти факторы, влияют на адаптацию растений. Если два
генотипы конкурируют то, их выживание зависит от количества семян,
произведенных каждым генотипом, а также количество семян их
потомства.
43.
Преимущества
1. Простой и удобный для проведения отбор.
2. Менее трудоемкий и менее дорогой в ранних поколениях.
3. Естественный отбор может увеличить частоту желательных
генотипов к концу отбора.
4. Он совместим с массовым отбором у самоопыляющихся видов.
5. Массовый отбор позволяет проанализировать большое количество
расщепляющихся потомств. Следовательно, селекционер может
сделать и оценить больше скрещиваний.
6. Выведенный сорт будет адаптирован к условиям окружающей среды,
так как был создан на основе материала, который пережил годы
естественного отбора.
7. Индивидуальный отбор проводится, когда растения в популяции
станут гомозиготными, что делает его более эффективным для оценки
и сравнения продуктивности растения
44.
• Недостатки• 1. Улучшенный генотипы могут быть потеряны под давлением
естественного отбора, в то время как нежелательные размножаются на
ранних поколениях.
• 2. Не подходит для видов, которые имеют редкую плотность посадки в
производстве.
• 3. Генетические характеристики популяции трудно установить, от
одного поколения к другому.
• 4. генотипы не в равной степени представлены в каждом поколении,
потому что не все растения в одном поколении представлены в
следующем поколении. Неправильная выборка может привести к
генетическому дрейфу.
• 5. Не рекомендуется проводить отбор в межсезонье в питомниках и
теплицах так как он может благоприятствовать генотипам, которые
нежелательны в регионе где ведется селекция.
• 6. Процедура длительная, нет возможности использовать межсезонные
посадки.