











 biology
biologySimilar presentations:
")









Молекулярная биология прокариот
1.
Молекулярная биология прокариотКаменский Петр Андреевич
peter@protein.bio.msu.ru
Основная литература:
1. L.Snyder. Molecular Genetics of Bacteria. Любое издание после 2010 года.
2. C.Lewin. Genes (или переводные издания). Любое издание после 2010 года.
3. M.Singer, P.Berg. Genes and Genomes (или переводные издания). Любое издание
после 2010 года.
Дополнительная литература:
1. Спирин А. С. Молекулярная биология. Рибосомы и биосинтез белка. Любое издание
после 2010 года.
2. B.Alberts. Molecular Biology of the Cell (или переводные издания). Любое издание
после 2010 года.
2.
Лекция 1.Репликация. Транскрипция. Трансляция.
3.
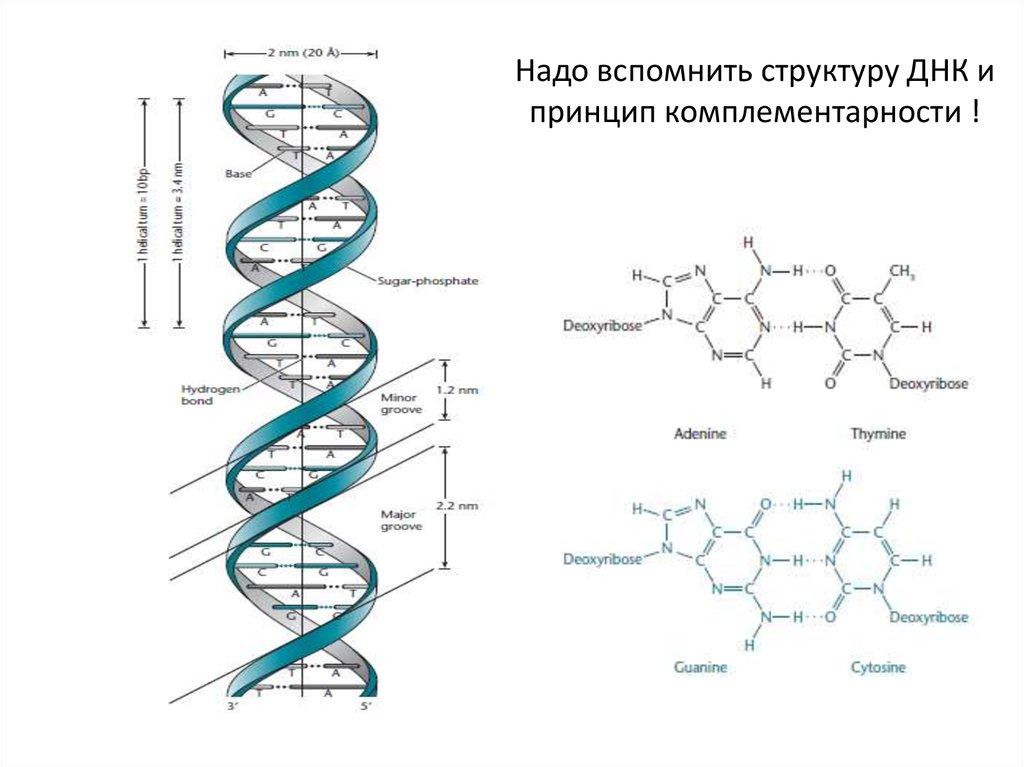
Надо вспомнить структуру ДНК ипринцип комплементарности !
4.
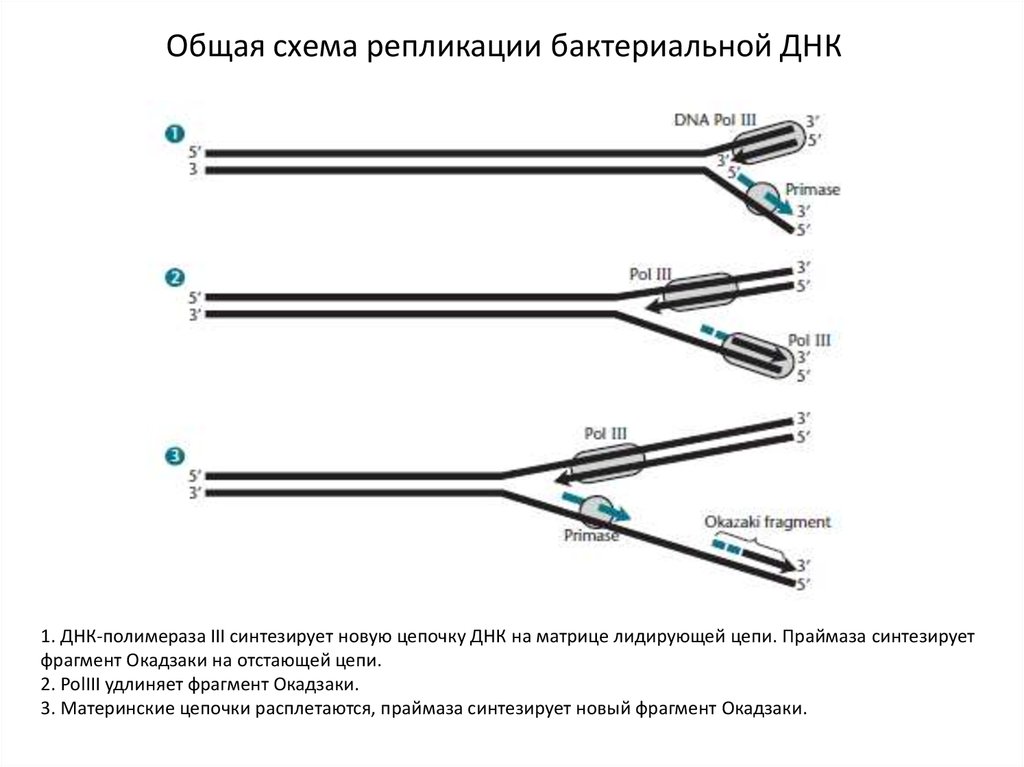
Общая схема репликации бактериальной ДНК1. ДНК-полимераза III синтезирует новую цепочку ДНК на матрице лидирующей цепи. Праймаза синтезирует
фрагмент Окадзаки на отстающей цепи.
2. PolIII удлиняет фрагмент Окадзаки.
3. Материнские цепочки расплетаются, праймаза синтезирует новый фрагмент Окадзаки.
5.
Общая схема репликации бактериальной ДНК4. Pol III удлиняет фрагмент Окадзаки на отстающей цепи и достигает предыдущего фрагмента.
5. Pol I удаляет РНК-часть предыдущего фрагмента Окадзаки и заменяет ее на последовательность ДНК.
6. Лигаза соединяет два фрагмента ДНК в одну непрерывную цепочку. Далее снова начинается п.1.
6.
Два ДНК-полимеразных комплекса, синтезирующие на лидирующей и отстающей цепи,на самом деле никогда не расходятся и работают скоординированно
Тромбонная репликация бактериальной ДНК
7.
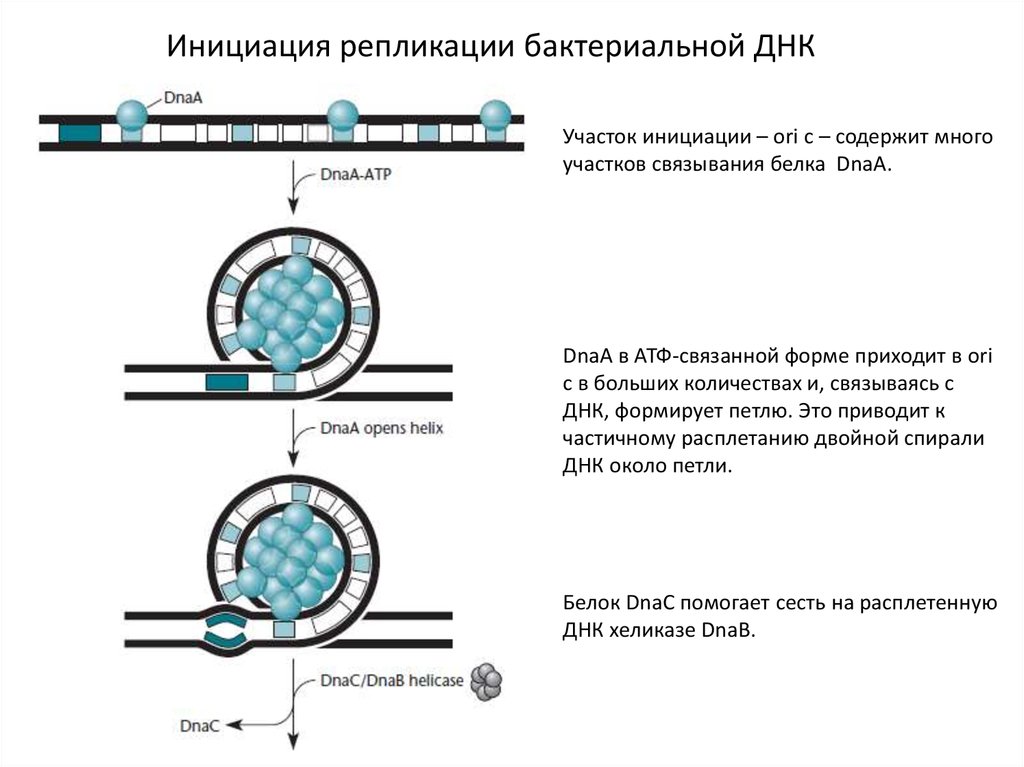
Инициация репликации бактериальной ДНКУчасток инициации – ori c – содержит много
участков связывания белка DnaA.
DnaA в АТФ-связанной форме приходит в ori
c в больших количествах и, связываясь с
ДНК, формирует петлю. Это приводит к
частичному расплетанию двойной спирали
ДНК около петли.
Белок DnaC помогает сесть на расплетенную
ДНК хеликазе DnaB.
8.
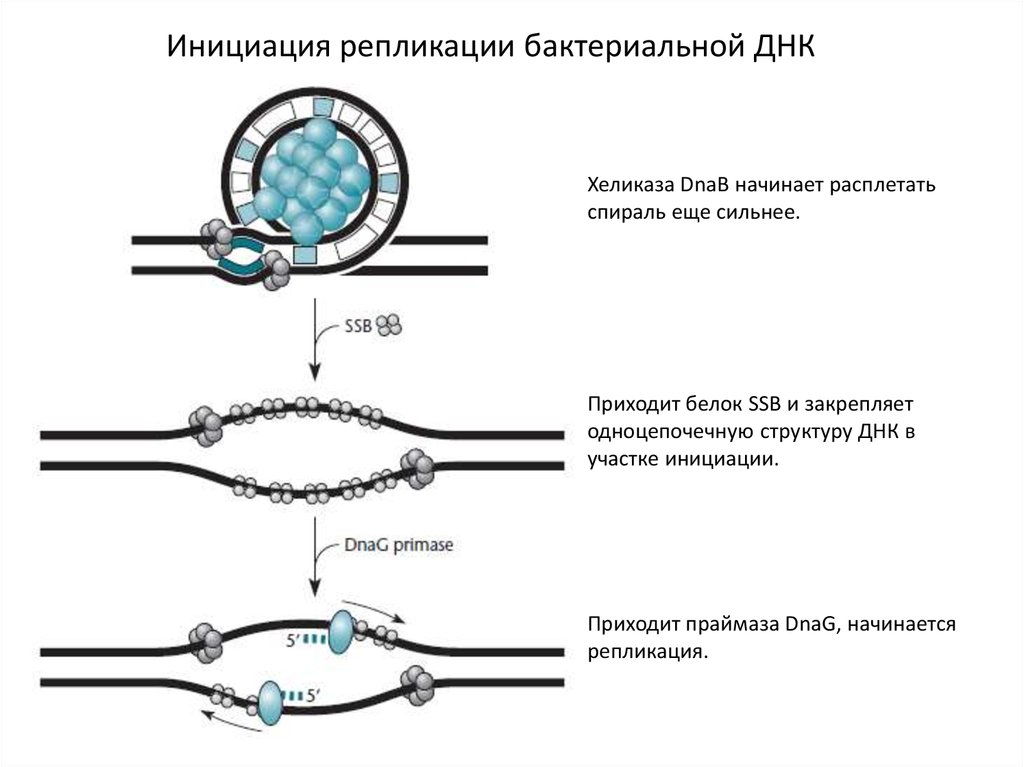
Инициация репликации бактериальной ДНКХеликаза DnaB начинает расплетать
спираль еще сильнее.
Приходит белок SSB и закрепляет
одноцепочечную структуру ДНК в
участке инициации.
Приходит праймаза DnaG, начинается
репликация.
9.
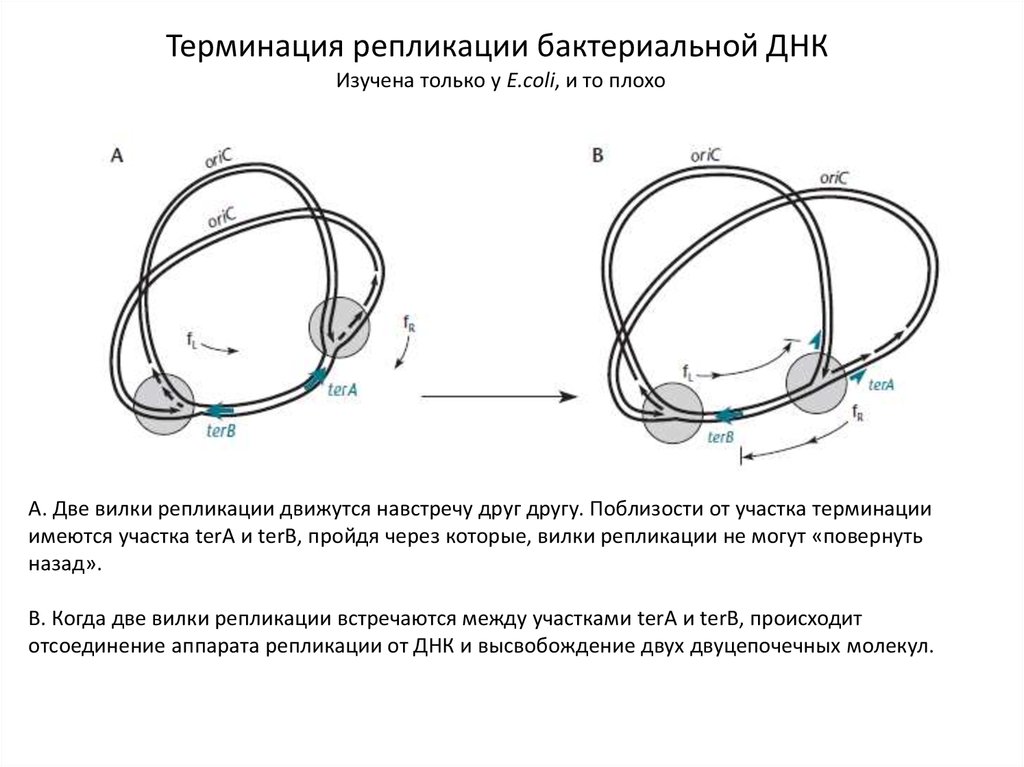
Терминация репликации бактериальной ДНКИзучена только у E.coli, и то плохо
А. Две вилки репликации движутся навстречу друг другу. Поблизости от участка терминации
имеются участка terA и terB, пройдя через которые, вилки репликации не могут «повернуть
назад».
В. Когда две вилки репликации встречаются между участками terA и terB, происходит
отсоединение аппарата репликации от ДНК и высвобождение двух двуцепочечных молекул.
10.
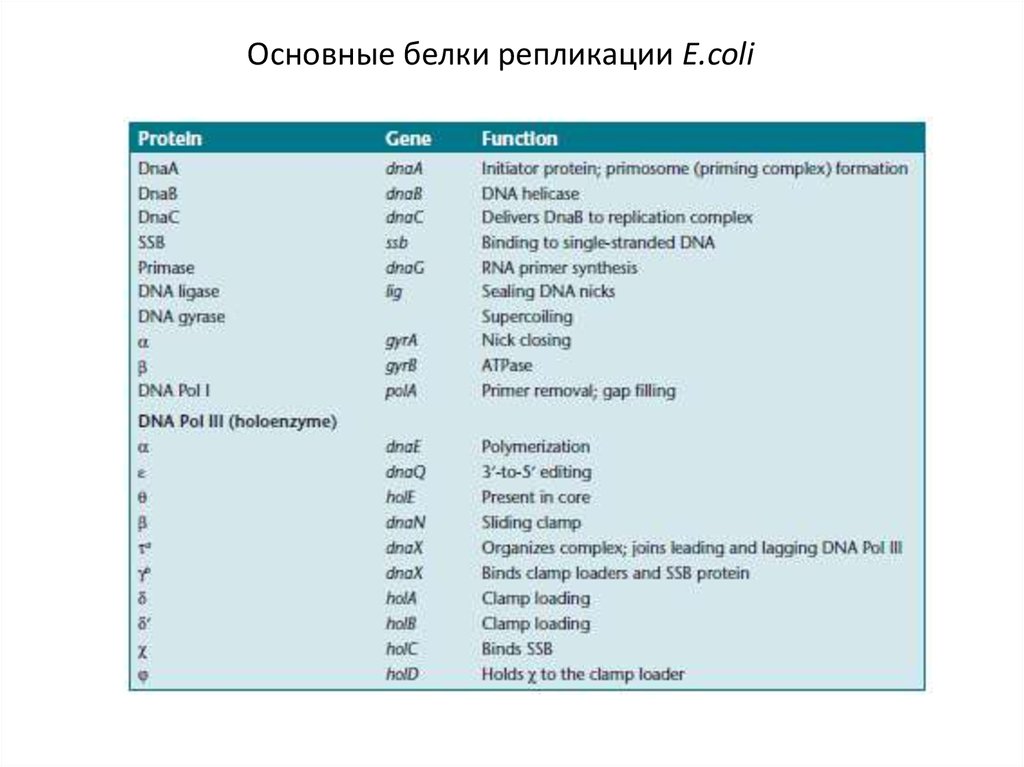
Основные белки репликации E.coli11.
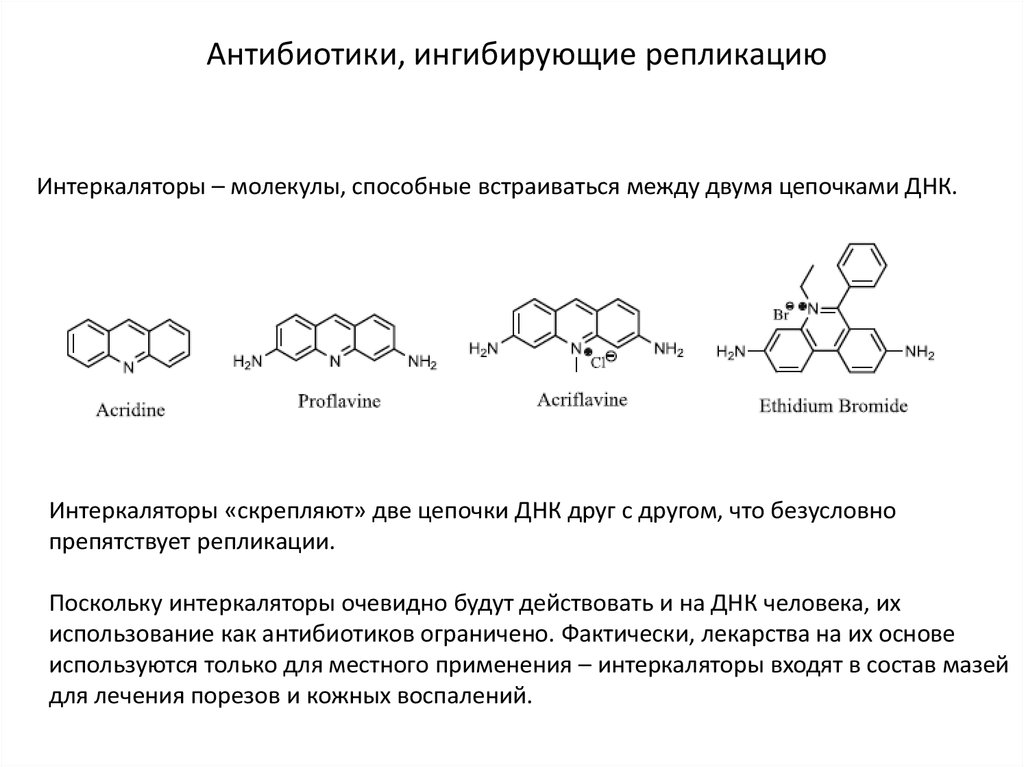
Антибиотики, ингибирующие репликациюИнтеркаляторы – молекулы, способные встраиваться между двумя цепочками ДНК.
Интеркаляторы «скрепляют» две цепочки ДНК друг с другом, что безусловно
препятствует репликации.
Поскольку интеркаляторы очевидно будут действовать и на ДНК человека, их
использование как антибиотиков ограничено. Фактически, лекарства на их основе
используются только для местного применения – интеркаляторы входят в состав мазей
для лечения порезов и кожных воспалений.
12.
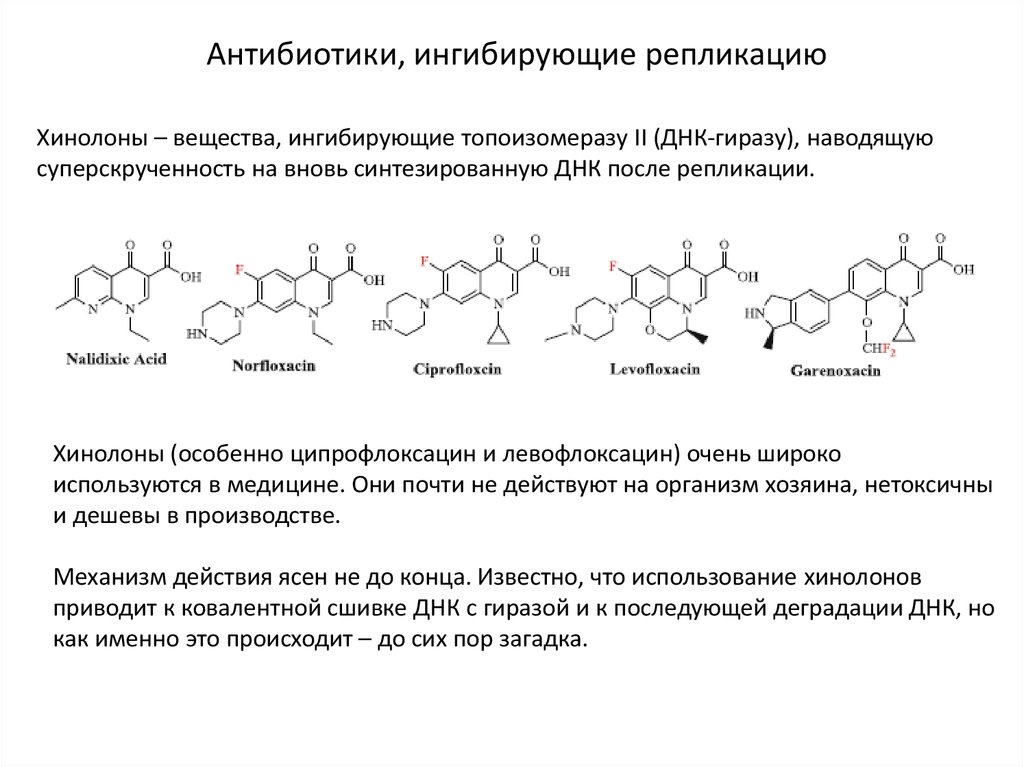
Антибиотики, ингибирующие репликациюХинолоны – вещества, ингибирующие топоизомеразу II (ДНК-гиразу), наводящую
суперскрученность на вновь синтезированную ДНК после репликации.
Хинолоны (особенно ципрофлоксацин и левофлоксацин) очень широко
используются в медицине. Они почти не действуют на организм хозяина, нетоксичны
и дешевы в производстве.
Механизм действия ясен не до конца. Известно, что использование хинолонов
приводит к ковалентной сшивке ДНК с гиразой и к последующей деградации ДНК, но
как именно это происходит – до сих пор загадка.
13.
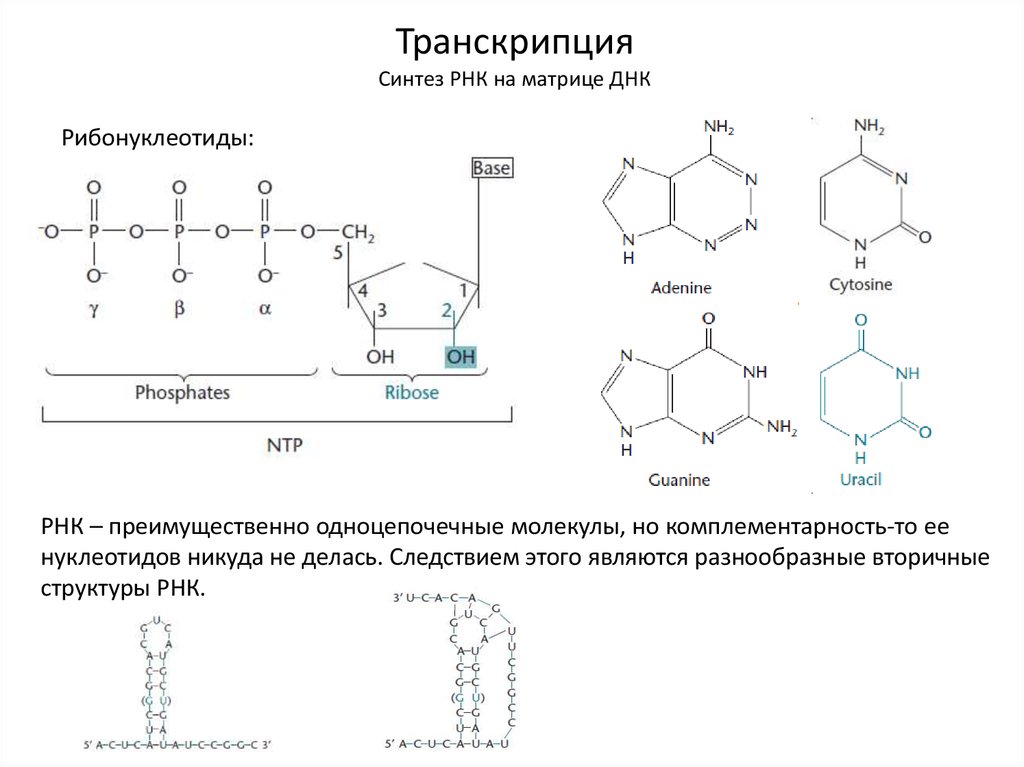
ТранскрипцияСинтез РНК на матрице ДНК
Рибонуклеотиды:
РНК – преимущественно одноцепочечные молекулы, но комплементарность-то ее
нуклеотидов никуда не делась. Следствием этого являются разнообразные вторичные
структуры РНК.
14.
РНК-полимеразаСигма-субъединица придает комплексу функциональность: именно она физически
взаимодействует с транскрибируемой РНК.
15.
РНК-полимераза в комплексе с ДНК16.
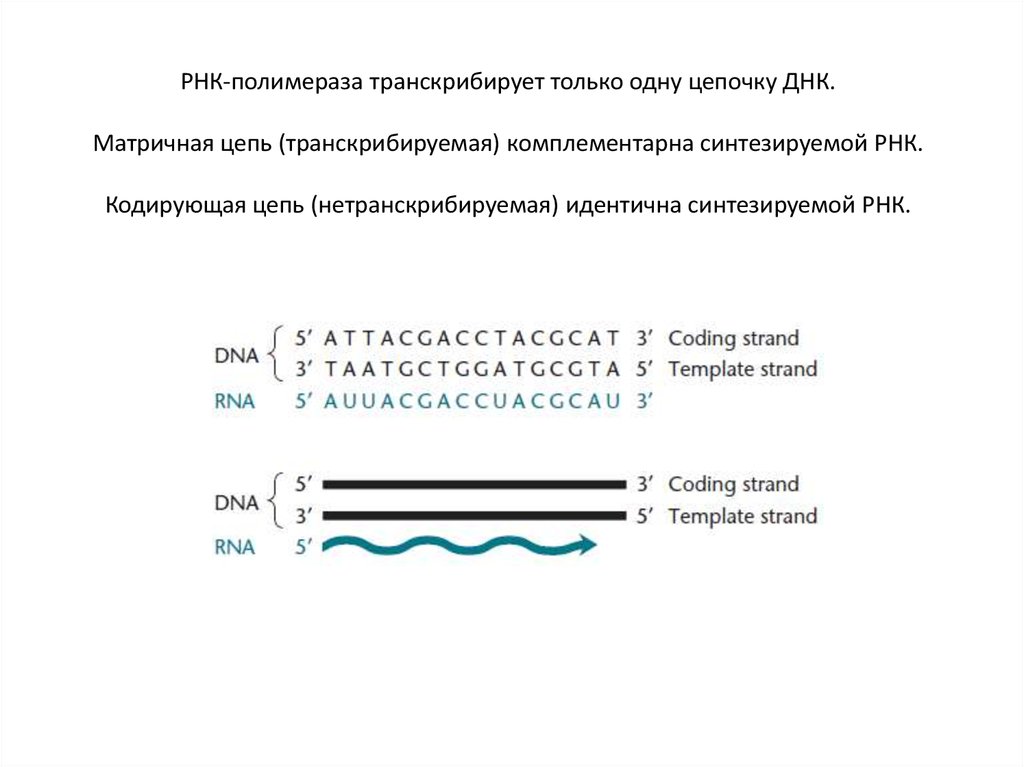
РНК-полимераза транскрибирует только одну цепочку ДНК.Матричная цепь (транскрибируемая) комплементарна синтезируемой РНК.
Кодирующая цепь (нетранскрибируемая) идентична синтезируемой РНК.
17.
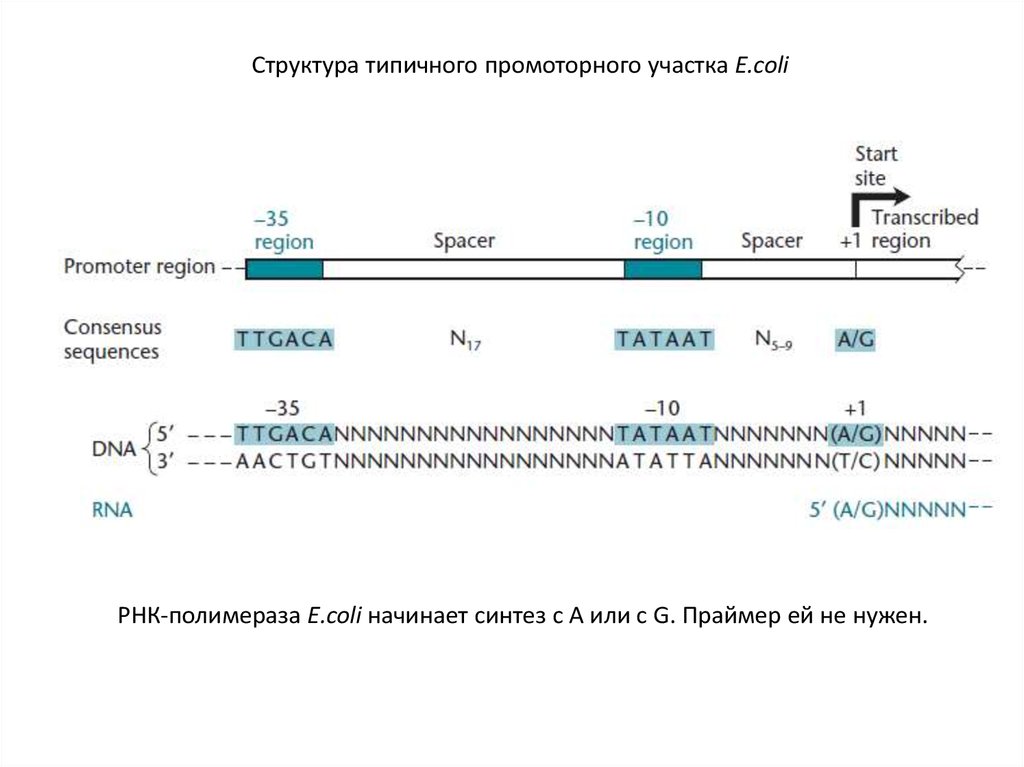
Структура типичного промоторного участка E.coliРНК-полимераза E.coli начинает синтез с А или с G. Праймер ей не нужен.
18.
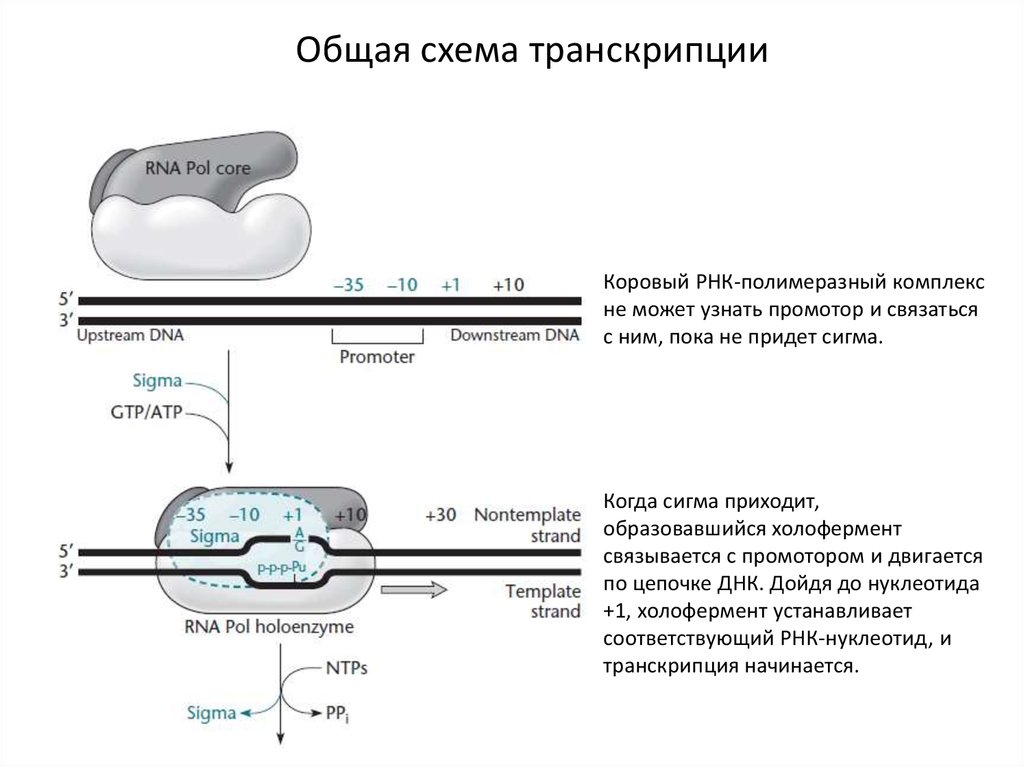
Общая схема транскрипцииКоровый РНК-полимеразный комплекс
не может узнать промотор и связаться
с ним, пока не придет сигма.
Когда сигма приходит,
образовавшийся холофермент
связывается с промотором и двигается
по цепочке ДНК. Дойдя до нуклеотида
+1, холофермент устанавливает
соответствующий РНК-нуклеотид, и
транскрипция начинается.
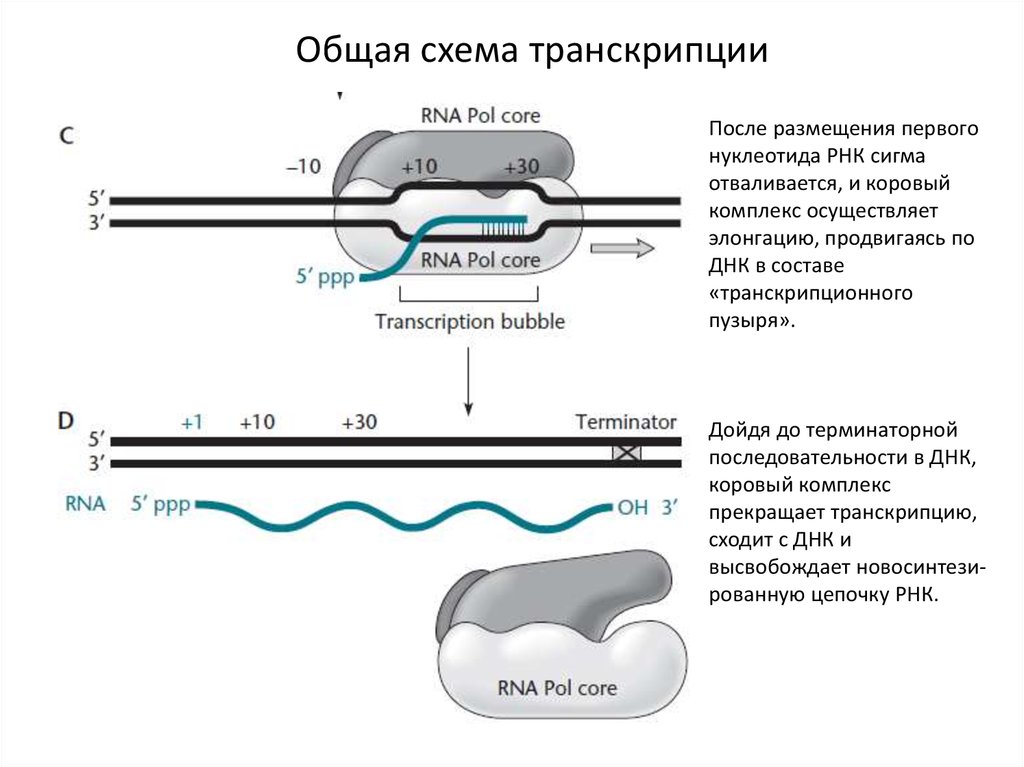
19.
Общая схема транскрипцииПосле размещения первого
нуклеотида РНК сигма
отваливается, и коровый
комплекс осуществляет
элонгацию, продвигаясь по
ДНК в составе
«транскрипционного
пузыря».
Дойдя до терминаторной
последовательности в ДНК,
коровый комплекс
прекращает транскрипцию,
сходит с ДНК и
высвобождает новосинтезированную цепочку РНК.
20.
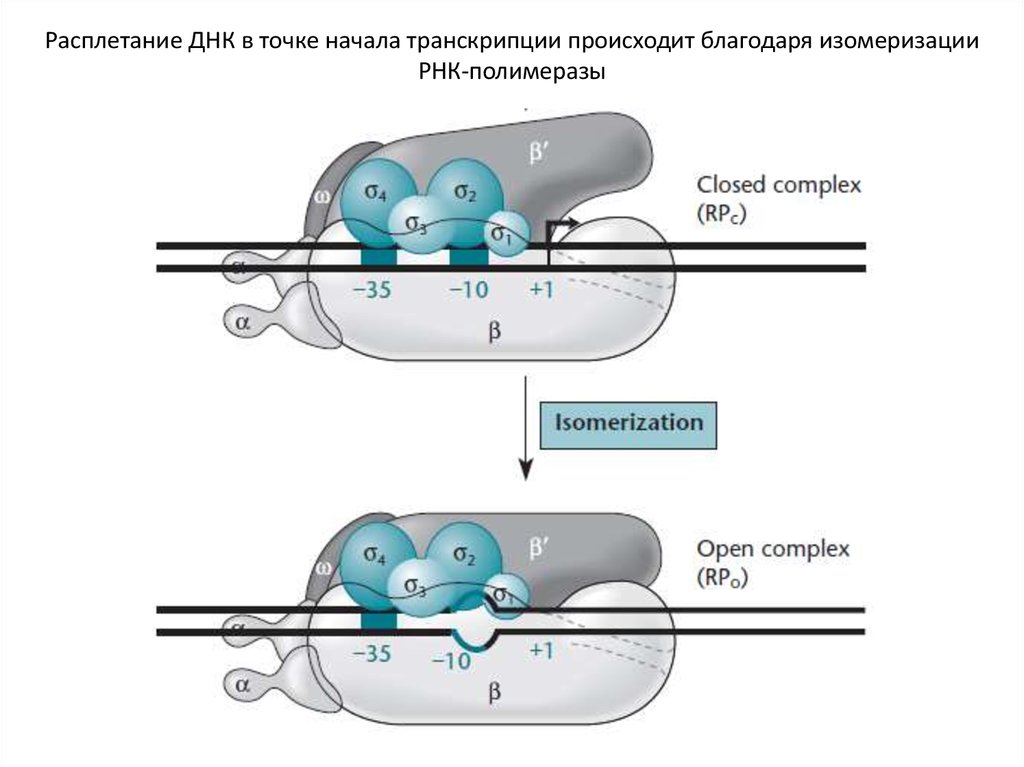
Расплетание ДНК в точке начала транскрипции происходит благодаря изомеризацииРНК-полимеразы
21.
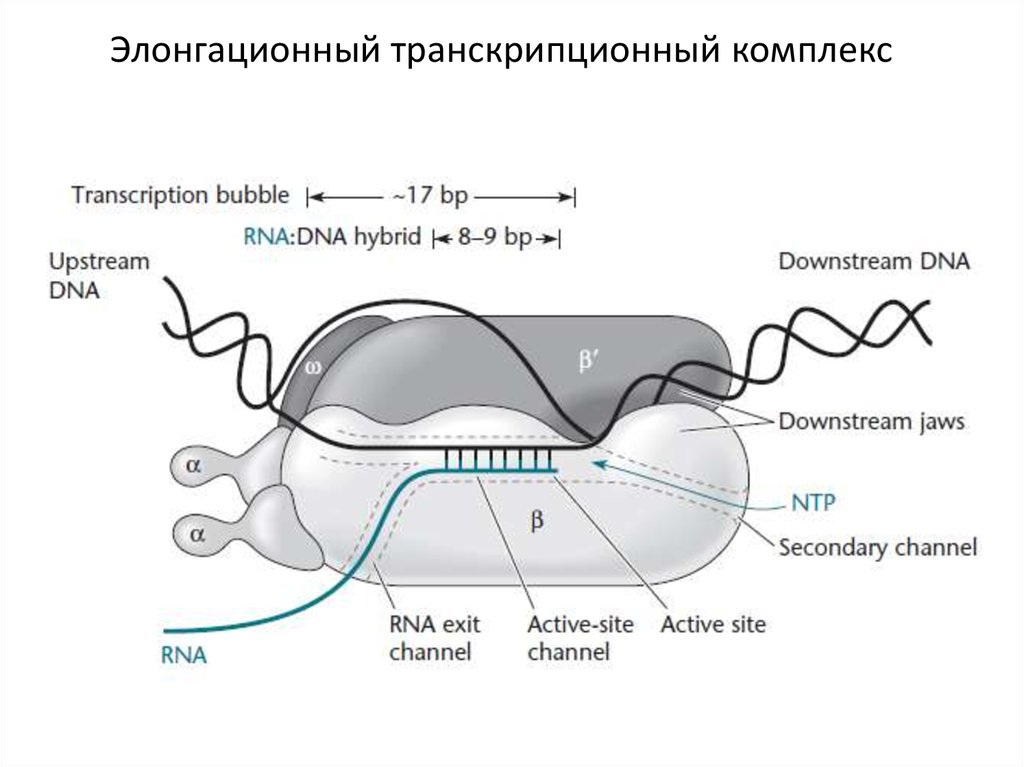
Элонгационный транскрипционный комплекс22.
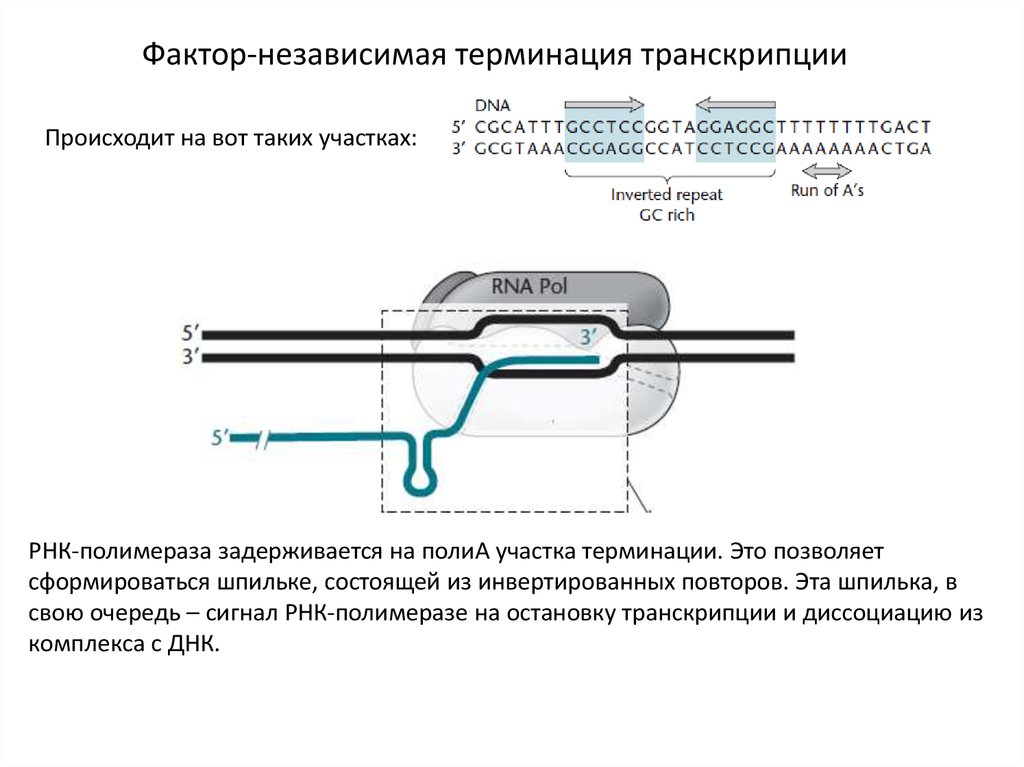
Фактор-независимая терминация транскрипцииПроисходит на вот таких участках:
РНК-полимераза задерживается на полиА участка терминации. Это позволяет
сформироваться шпильке, состоящей из инвертированных повторов. Эта шпилька, в
свою очередь – сигнал РНК-полимеразе на остановку транскрипции и диссоциацию из
комплекса с ДНК.
23.
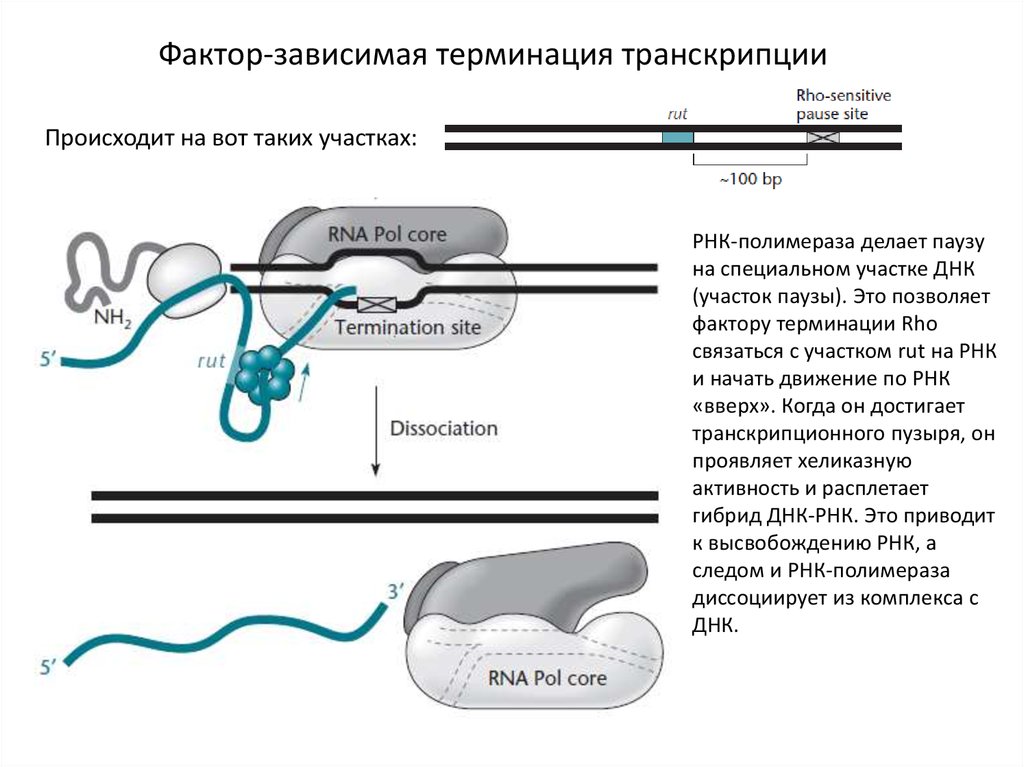
Фактор-зависимая терминация транскрипцииПроисходит на вот таких участках:
РНК-полимераза делает паузу
на специальном участке ДНК
(участок паузы). Это позволяет
фактору терминации Rho
связаться с участком rut на РНК
и начать движение по РНК
«вверх». Когда он достигает
транскрипционного пузыря, он
проявляет хеликазную
активность и расплетает
гибрид ДНК-РНК. Это приводит
к высвобождению РНК, а
следом и РНК-полимераза
диссоциирует из комплекса с
ДНК.
24.
Антибиотики, ингибирующие транскрипциюРифамицины – класс соединений, связывающих бета-субъединицу РНК-полимеразы и
блокирующие присоединение новых нуклеотидов к растущей цепочке РНК.
Самый известный антибиотик – рифампицин, использующийся в медицине с середины
прошлого века и до сих пор являющийся одним из основных средств против
туберкулеза.
25.
Антибиотики, ингибирующие транскрипциюЛассо-пептиды связывают бета-субъединицу РНК-полимеразы и ингибируют ее
активный центр.
В клиническую практику пока не вошли, точный молекулярный механизм действия
изучен плохо.
26.
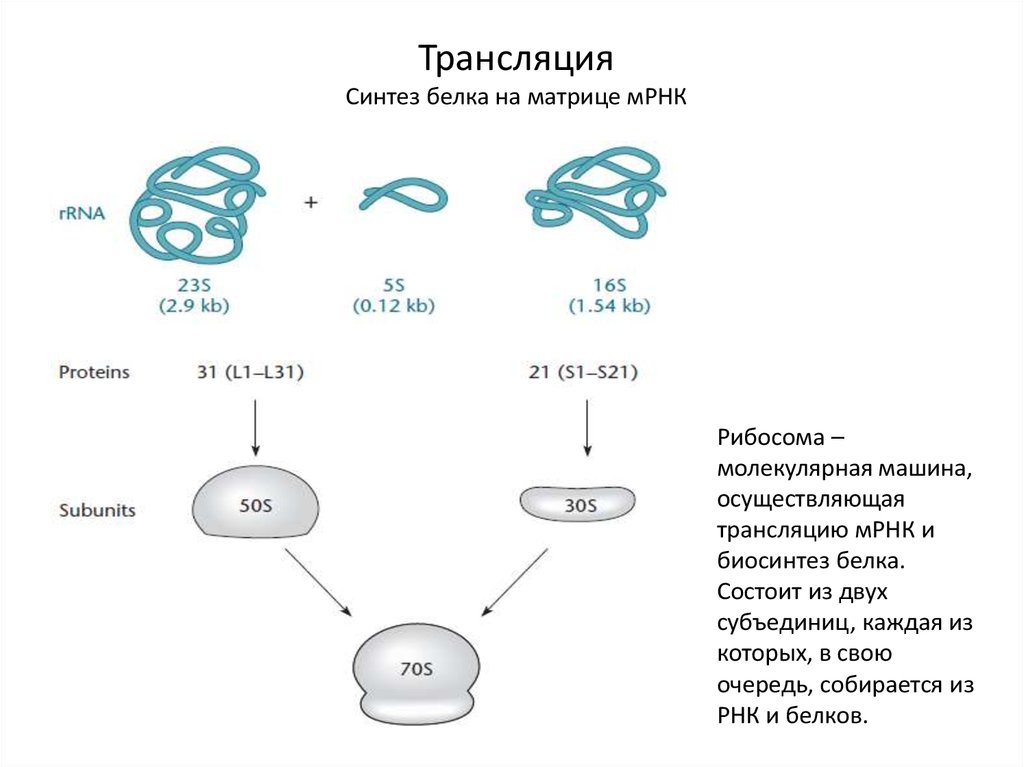
ТрансляцияСинтез белка на матрице мРНК
Рибосома –
молекулярная машина,
осуществляющая
трансляцию мРНК и
биосинтез белка.
Состоит из двух
субъединиц, каждая из
которых, в свою
очередь, собирается из
РНК и белков.
27.
тРНК – адапторная молекула, задающая однозначное соответствие между каждымтриплетом (тремя нуклеотидами) мРНК и аминокислотой.
Но не наоборот: генетический код вырожден, и одна аминокислота может
кодироваться несколькими разными триплетами.
28.
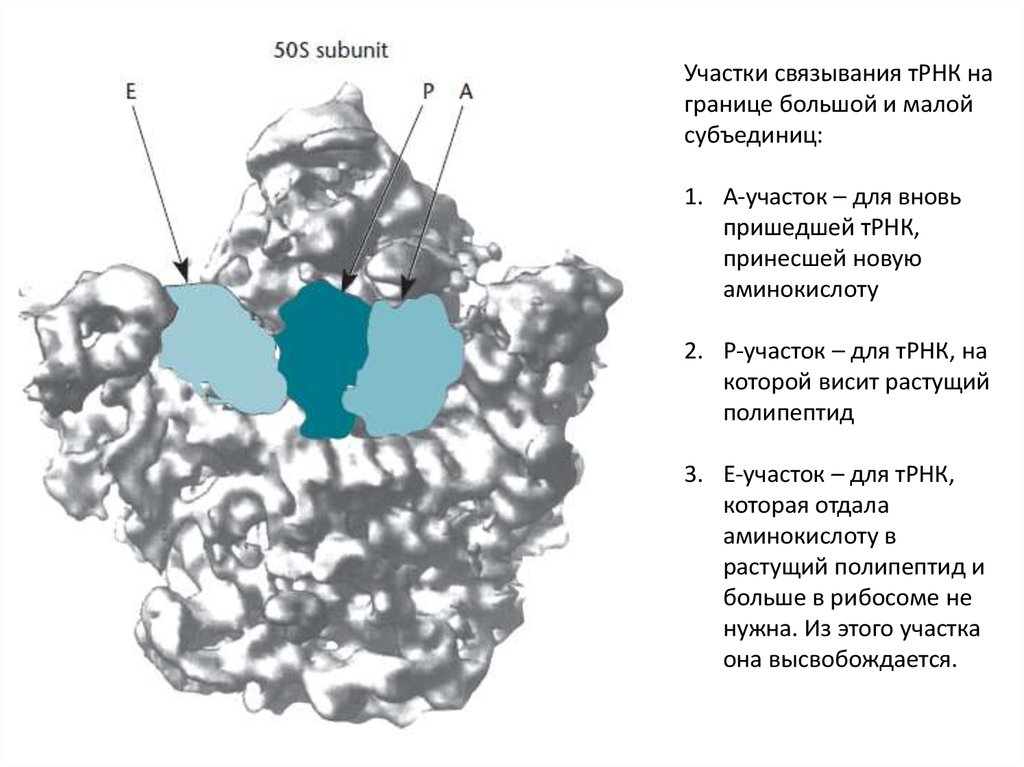
Участки связывания тРНК награнице большой и малой
субъединиц:
1. А-участок – для вновь
пришедшей тРНК,
принесшей новую
аминокислоту
2. Р-участок – для тРНК, на
которой висит растущий
полипептид
3. Е-участок – для тРНК,
которая отдала
аминокислоту в
растущий полипептид и
больше в рибосоме не
нужна. Из этого участка
она высвобождается.
29.
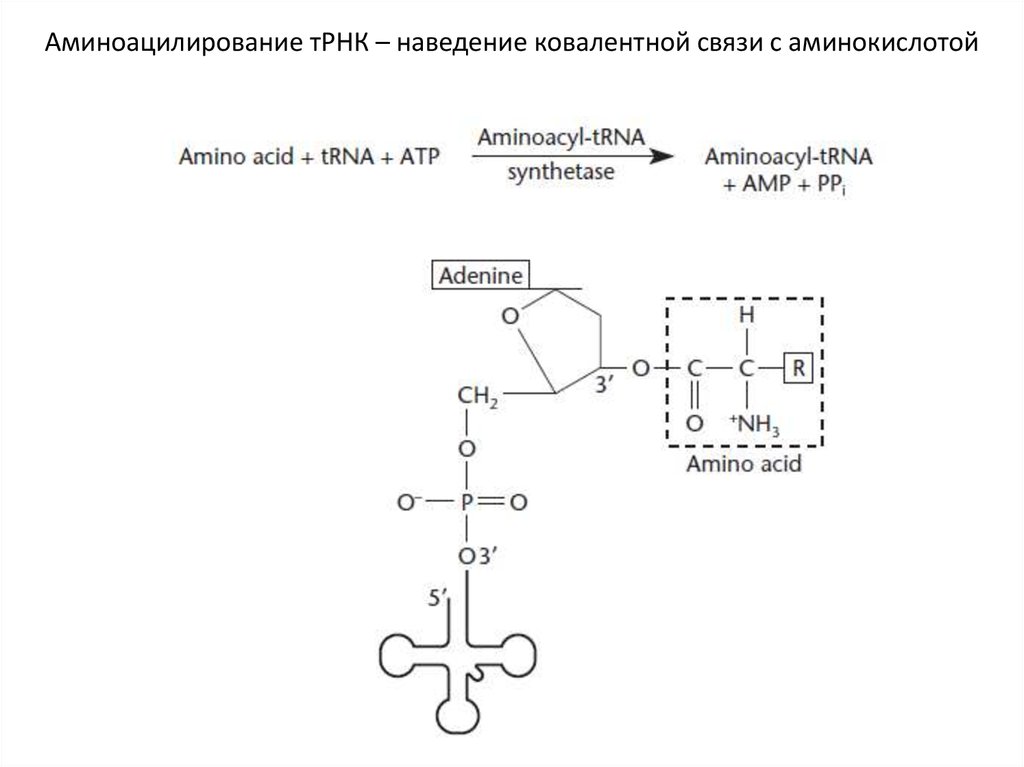
Аминоацилирование тРНК – наведение ковалентной связи с аминокислотой30.
Инициация трансляцииАналог промотора в ДНК – последовательность Шайна-Дальгарно в мРНК
Эту последовательность узнает рибосома (точнее – 3’-конец рРНК малой
субъединицы) и связывается с ней.
31.
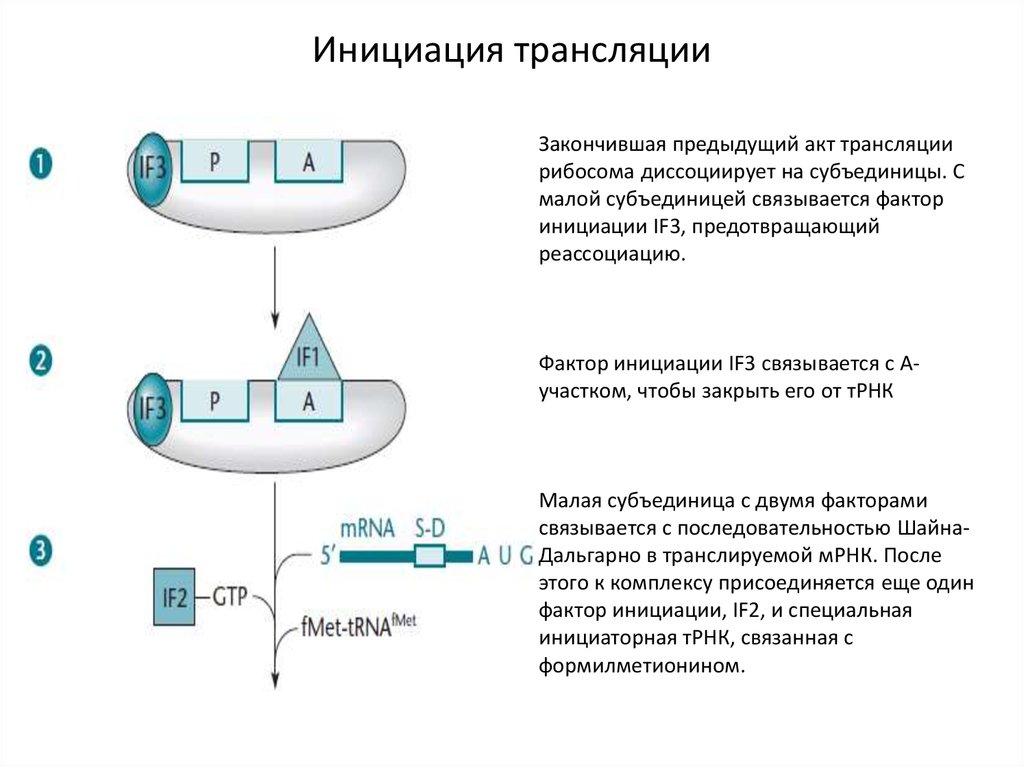
Инициация трансляцииЗакончившая предыдущий акт трансляции
рибосома диссоциирует на субъединицы. С
малой субъединицей связывается фактор
инициации IF3, предотвращающий
реассоциацию.
Фактор инициации IF3 связывается с Аучастком, чтобы закрыть его от тРНК
Малая субъединица с двумя факторами
связывается с последовательностью ШайнаДальгарно в транслируемой мРНК. После
этого к комплексу присоединяется еще один
фактор инициации, IF2, и специальная
инициаторная тРНК, связанная с
формилметионином.
32.
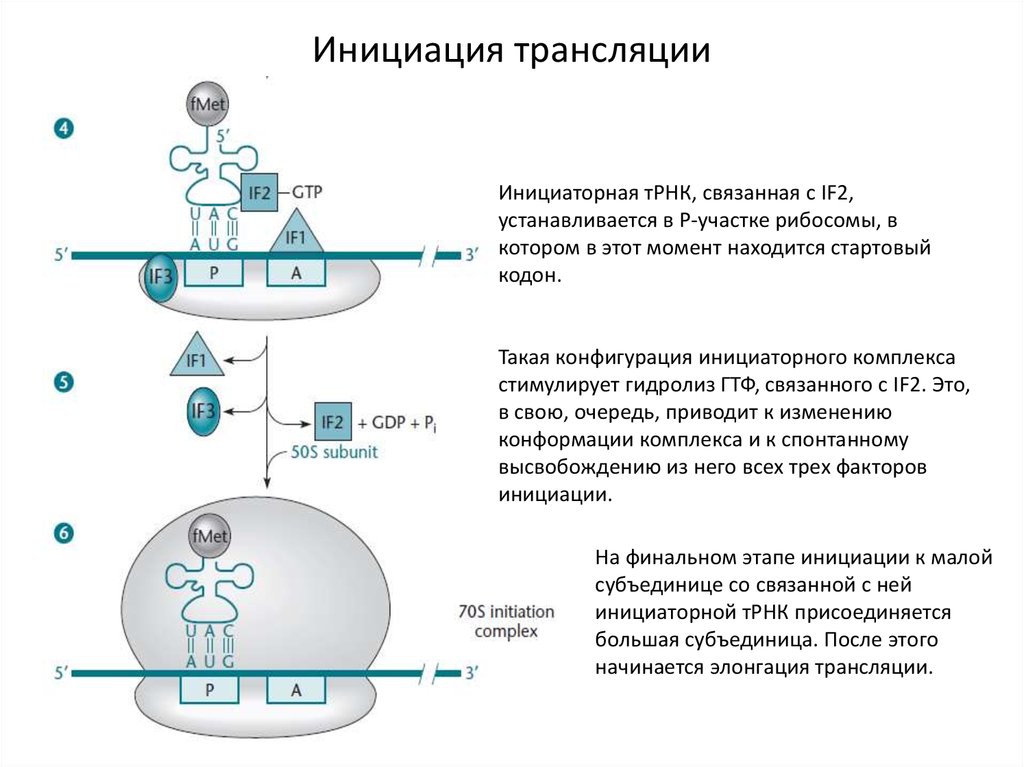
Инициация трансляцииИнициаторная тРНК, связанная с IF2,
устанавливается в Р-участке рибосомы, в
котором в этот момент находится стартовый
кодон.
Такая конфигурация инициаторного комплекса
стимулирует гидролиз ГТФ, связанного с IF2. Это,
в свою, очередь, приводит к изменению
конформации комплекса и к спонтанному
высвобождению из него всех трех факторов
инициации.
На финальном этапе инициации к малой
субъединице со связанной с ней
инициаторной тРНК присоединяется
большая субъединица. После этого
начинается элонгация трансляции.
33.
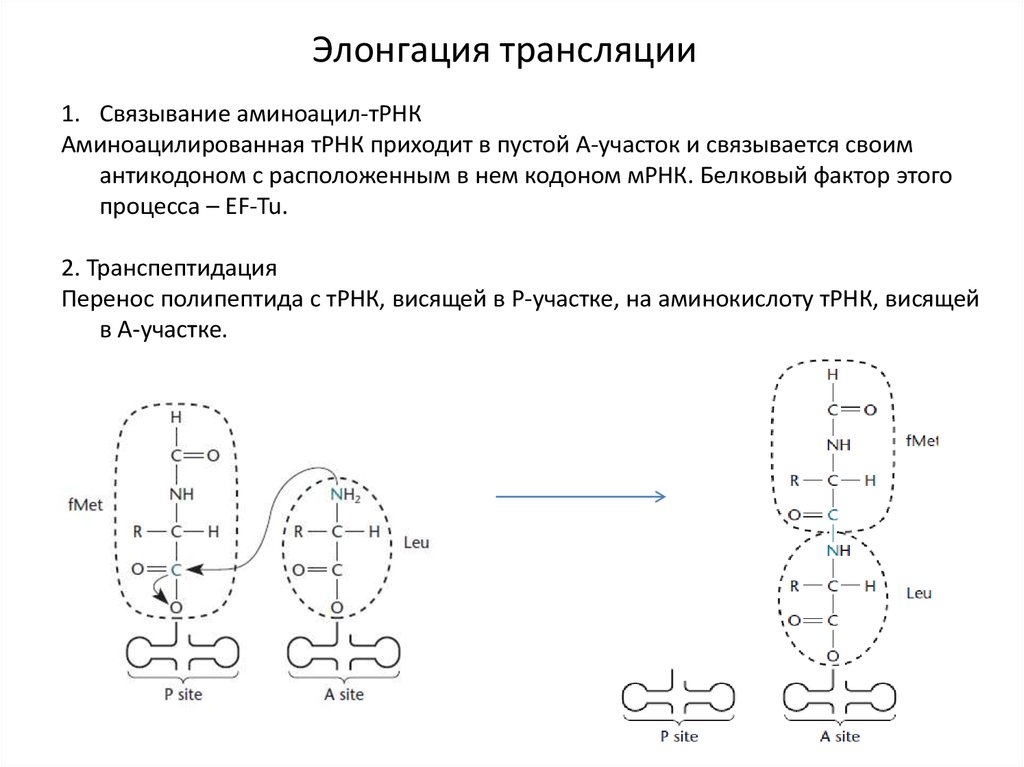
Элонгация трансляции1. Связывание аминоацил-тРНК
Аминоацилированная тРНК приходит в пустой А-участок и связывается своим
антикодоном с расположенным в нем кодоном мРНК. Белковый фактор этого
процесса – EF-Tu.
2. Транспептидация
Перенос полипептида с тРНК, висящей в Р-участке, на аминокислоту тРНК, висящей
в А-участке.
34.
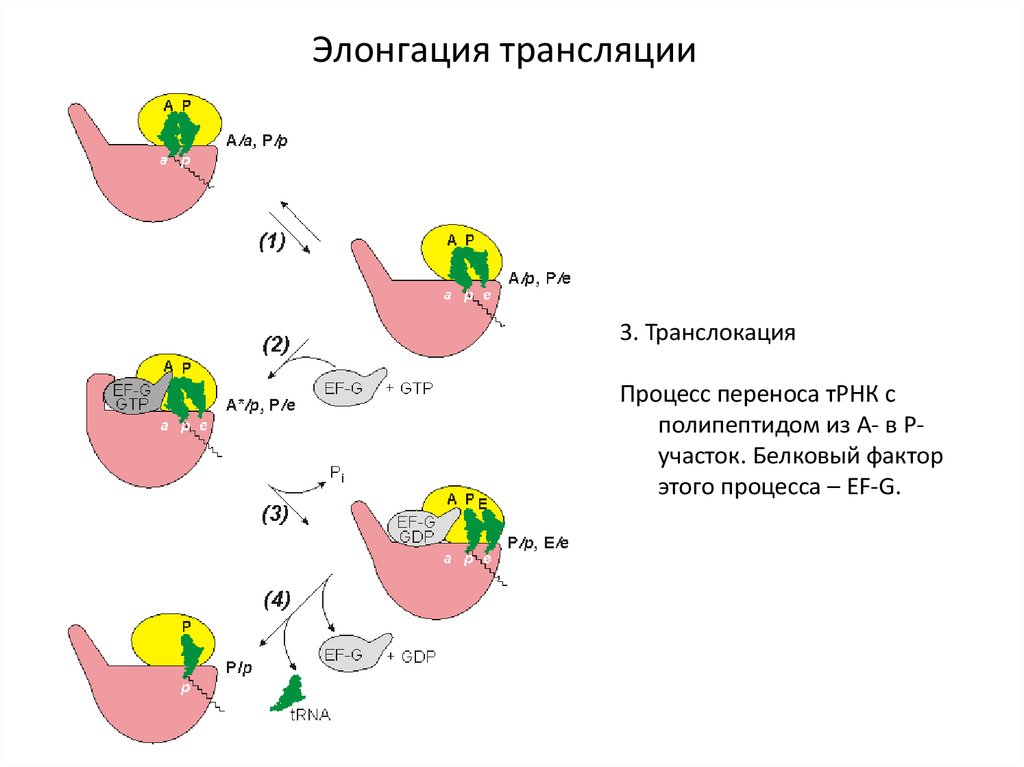
Элонгация трансляции3. Транслокация
Процесс переноса тРНК с
полипептидом из А- в Ручасток. Белковый фактор
этого процесса – EF-G.
35.
Терминация трансляцииТерминация происходит на так называемых
стоп-кодонах мРНК, для которых нет тРНК.
Вместо нее имеется специальный фактор RRF,
структура которого схожа со структурой тРНК,
что и позволяет ему связаться со стоп-кодоном в
А-участке рибосомы.
Такое связывание – сигнал для
остановки трансляции, диссоциации
всех факторов, тРНК и мРНК из
комплекса с рибосомой и диссоциации
самой рибосомы на субъединицы.
После этого с малой субъединицей
связывается фактор инициации IF3, и все
начинается сначала.
36.
Антибиотики, ингибирующие трансляциюСамая большая группа антибиотиков, очень широко использующиеся в клинике,
поскольку бактериальная и эукариотическая системы трансляции структурно и
механистически очень сильно отличаются друг от друга, и поэтому подавляющее
большинство трансляционных антибиотиков никак не воздействуют на трансляцию в
хозяйских клетках.