
























 biology
biologySimilar presentations:



")






Жгутиковые. Простейшие организмы
1.
Зоол.Л.2. ЖгутиковыеМонадный тип организации
Надтип Dinomorpha,
Надтип Euglenozoa: Tип Euglenophyta,
Tип Kinetoplastida: Класс Bodomonadida,
Класс Trypanosomamonadida;
Надтип Polymastigota, Tип Diplomonadida, Tип
Parabasalia: Класс Trichomonadea, Класс
Hypermastiginеa: Tип Chlorophyta: Класс
Euchloromonada
2. Царство Protista, Подцарство Protozoa
• 1. Общая характеристика• К подцарству Protozoa относятся гетеротрофные
эукариоты, среди которых есть одноклеточные
моно и полиэнергидные, многоклеточные и
колониальные организмы. Общее свойство всех
представителей подцарства: их организация не
выходит за пределы клеточного уровня, т.е.
подавляющее большинство дифференцировок,
приводящих к возникновению того или иного плана
организации, происходят внутри клеток. У
многоклеточных простейших никогда не возникает
гетероклеточность, приводящая к образованию
ансамблей клеток, объединенных сходством
выполняемых функций (тканей или тканоидов).
3.
Элементарной единицей организации простейших
служит так называемая энергида. Энергида - часть
клетки, если клетка - это плазма со своим ядром,
органеллами и пограничным слоем, пределом которого
служит клеточная оболочка. Энергида не имеет
собственного пограничного слоя. Это участок плазмы
со своим ядром, органеллами, но без пограничного слоя.
У многоядерной амебы имеется общая плазма, ядра
и часть ядер имеет свою долю пограничного слоя, а
часть нет. Те ядра, которые расположены ближе к
центральной части амебы и не имеют своего
пограничного слоя вместе с органеллами, составляют
энергиду. А само такое многоядерное тело называется
симпластом (если не способно к амебоидному
движению) или плазмодием (если способно к
амебоидному движению или изменению формы тела).
Клетку со многими ядрами называют полиэнергидной,
а с одним - моноэнергидной.
4.
• Размеры простейших• В среднем – меньше 1 мм. Наибольшее
число видов – от 5 до 150 мкм.
• Самые мелкие – Babesia boa – 12,5 мкм,
Leischmania donovani – 14 мкм.
• Наиболее известные крупные –
Paramecium caudatum – 150-250 мкм,
Amoeba proteus – 600 мкм.
• Cамые крупные – инфузория
Spirostomum ambiguum – 3 мм,
фораминиферы ныне живущие – до 25
мм, вымершие – до 1,5 м.
5.
• Форма тела простейших и его окраскачрезвычайно разнообразны и зависят
от условий сред обитания.
• То же относится и к строению тела
простейших. Но оно всегда сложнее
строения отдельной клетки
многоклеточных организмов, т.к.
простейшие – самостоятельные
организмы. Однако, есть примитивные
(паразитические) и более совершенные
(свободно живущие, тем более хищные
и более подвижные) протисты.
6.
Монадный (жгутиковый) тип организацииЖ.т. объединяет организмы, у которых во взрослом состоянии имеется
жгутик как орган передвижения. В то же время, наличие жгутика на
определенных стадиях жизненного цикла и у гамет характерно
практически для всех животных (кроме нематод) и растений, даже
некоторых высших.
Детальное изучение тонкого строения жгутиконосцев (покровных структур
клетки, жгутикового аппарата, митохондрий и хлоропластов и т.д.), их
физиологии и биохимических особенностей отчетливо показало, что это
сборная группа, всех представителей которой фактически объединяет
только один признак — наличие жгутикового аппарата.
Жгутиковые, как таксономическая группа – Mastigophora, выделяются по
наличию целой системы органоидов, отличающих их от саркодовых.
Размеры жгутиконосцев: 46-3000 мкм, в среднем – 50.
Систематика и филогения простейших строится на строении жгутика и его
производных.
Жгутики служат не только для движения, но могут использоваться для
временного или постоянного прикрепления.
7.
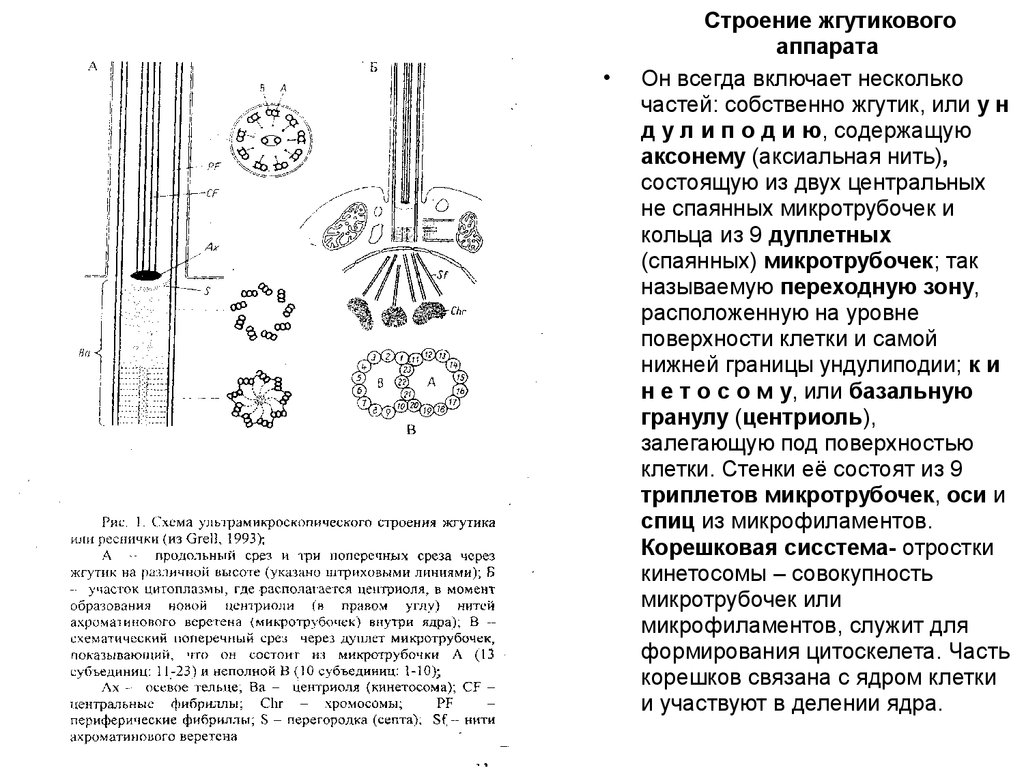
Строение жгутикового
аппарата
Он всегда включает несколько
частей: собственно жгутик, или у н
д у л и п о д и ю, содержащую
аксонему (аксиальная нить),
состоящую из двух центральных
не спаянных микротрубочек и
кольца из 9 дуплетных
(спаянных) микротрубочек; так
называемую переходную зону,
расположенную на уровне
поверхности клетки и самой
нижней границы ундулиподии; к и
н е т о с о м у, или базальную
гранулу (центриоль),
залегающую под поверхностью
клетки. Стенки её состоят из 9
триплетов микротрубочек, оси и
спиц из микрофиламентов.
Корешковая сисстема- отростки
кинетосомы – совокупность
микротрубочек или
микрофиламентов, служит для
формирования цитоскелета. Часть
корешков связана с ядром клетки
и участвуют в делении ядра.
8.
Одни детали строения жгутикового аппарата очень
консервативны и воспроизводятся практически у всех эукариот,
обладающих жгутиками. К их числу относятся аксонема,
проходящая внутри ундулиподии, и кинетосома. Другие же
варьируют в достаточно широких пределах, причем для каждого
таксона высокого ранга (тип, класс) характерен свой набор
признаков.
• Если строение аксонемы универсально для всех жгутиковых и
ресничных эукариот, то внешний вид ундулиподии может сильно
различаться у представителей разных таксонов. Поверхность
может быть гладкой и ровной, а может нести многочисленные
чешуйки или особые палочковидные структуры —
мастигонемы, наличие которых придает жгутику перистый вид.
У некоторых жгутиконосцев параллельно аксонеме тянется
особая волокнистая структура — параксиальный тяж,
присутствие которого, по-видимому, увеличивает упругость
жгутика.
9.
Судя по наличию двух жгутиков у гамет, например, у фораминифер, иподавляющего большинства жгутиконосцев, предками их были
двужгутиковые. Многожгутиковость и одножгутиковость вторичны.
Раньше систематика жгутиковых строилась на числе, форме, длине,
месте прикрепления и характере движения. Сейчас – на тонком
строении отдельных структур.
Выделяют 4 морфотипа жгутиконосцев:
• Изоконты имеют от 2 до 8 жгутиков равной длины, направленных в
одну сторону, с одинаковыми способами биения, К ним относится
большинство подвижных клеток зеленых водорослей.
• Анизоконты имеют 2 жгутика неравной длины, направленные в одну
сторону, отличающиеся по способу биения. Такие жгутики характерны
для бесцветных жгутиконосцев.
• У гетероконтных имеются 2 жгутика неравной длины (один направлен
вперед, второй - назад), различающиеся по способу биения.
Характерны и для подвижных клеток водорослей, и для так называемых
зооспоровых грибов, и для бесцветных жгутиконосцев.
• Стефаноконты имеют венчик жгутиков на переднем конце клетки. Это
характерно для многожгутиковых гамет и зооспор некоторых зеленых
водорослей.
10. Надтип Dinomorpha Панцирные жгутиконосцы
11. Надтип Euglenozoa: Tип Euglenophyta
представлен одиночными и
колониальными ауто-, миксо-и
гетеротрофными организмами. Хотя
внешняя морфология эвгленовых
весьма разнообразна, всем им
присущ ряд общих признаков.
Размеры эвгленовых варьируют в
широких пределах — от 5 до 500 мкм.
Форма тела изменяется от широко
листовидной до вытянуто игольчатой.
Наиболее обычны виды с
веретеновидным или удлиненно
овальным телом. Иногда тело
заключено в «домик».
Количество жгутиков варьирует от
одного до нескольких. Обычно
имеется два жгутика, сильно
различающихся по длине.
Основания жгутиков помещаются на
дне глубокого цилиндрического или
мешковидного впячивания,
называемого жгутиковым
резервуаром или жгутиковым
карманом.
12.
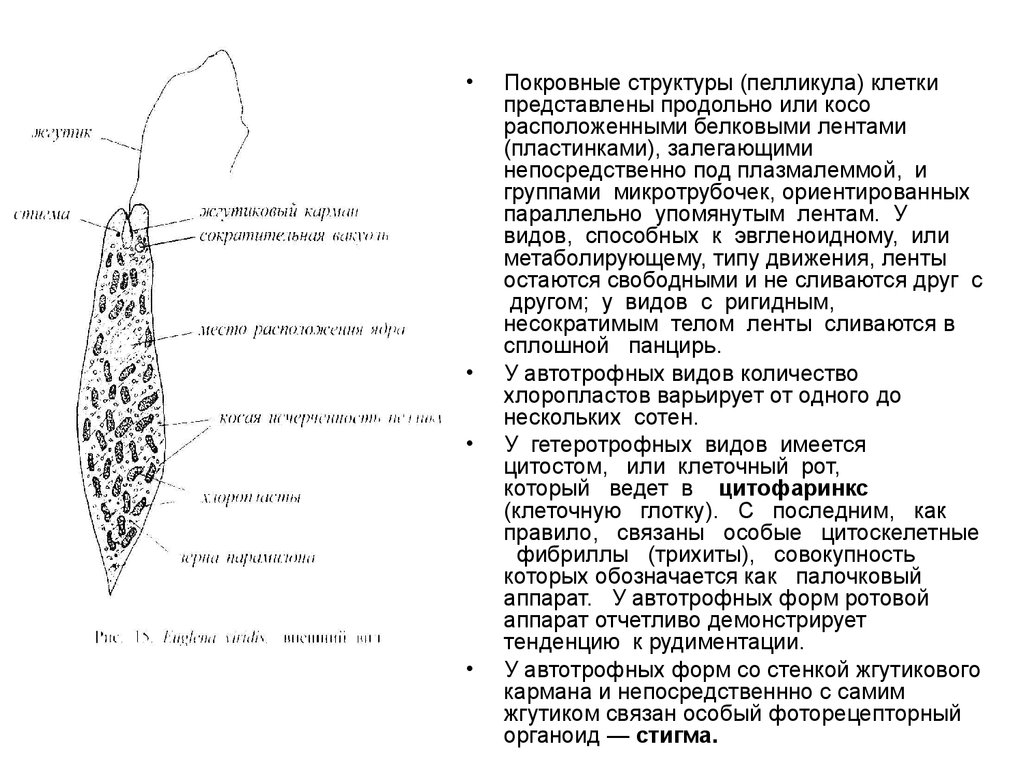
Покровные структуры (пелликула) клетки
представлены продольно или косо
расположенными белковыми лентами
(пластинками), залегающими
непосредственно под плазмалеммой, и
группами микротрубочек, ориентированных
параллельно упомянутым лентам. У
видов, способных к эвгленоидному, или
метаболирующему, типу движения, ленты
остаются свободными и не сливаются друг с
другом; у видов с ригидным,
несократимым телом ленты сливаются в
сплошной панцирь.
У автотрофных видов количество
хлоропластов варьирует от одного до
нескольких сотен.
У гетеротрофных видов имеется
цитостом, или клеточный рот,
который ведет в цитофаринкс
(клеточную глотку). С последним, как
правило, связаны особые цитоскелетные
фибриллы (трихиты), совокупность
которых обозначается как палочковый
аппарат. У автотрофных форм ротовой
аппарат отчетливо демонстрирует
тенденцию к рудиментации.
У автотрофных форм со стенкой жгутикового
кармана и непосредственнно с самим
жгутиком связан особый фоторецепторный
органоид — стигма.
13.
Резервные энергетические вещества у
автотрофных форм представлены особым
полисахаридом — парамилоном, а у
гетеротрофных форм к нему добавляется
гликоген.
Сократительная вакуоль располагается в
переднем конце тела, в непосредственной
близости от жгутикового кармана. В полость
этого кармана при систоле изливается
содержимое вакуоли. У ряда видов
сократительная вакуоль отсутствует.
Ядро одно, округлой или овальной
формы, обычно располагается в
центральной части клетки.
10. Размножаются преимущественно
путем простого бинарного деления.
Для некоторых видов (в частности,
паразитических) характерна
п а л и н т о м и я, то есть серия быстро
следующих друг за другом делений клетки.
Так как период роста вновь образовавшихся
клеток при таком типе их размножения
практически полностью выпадает,
результатом всего этого становится
появление большого числа очень мелких
особей. В жизненных циклах паразитов они
играют роль инвазионных для хозяина
зооспор
14.
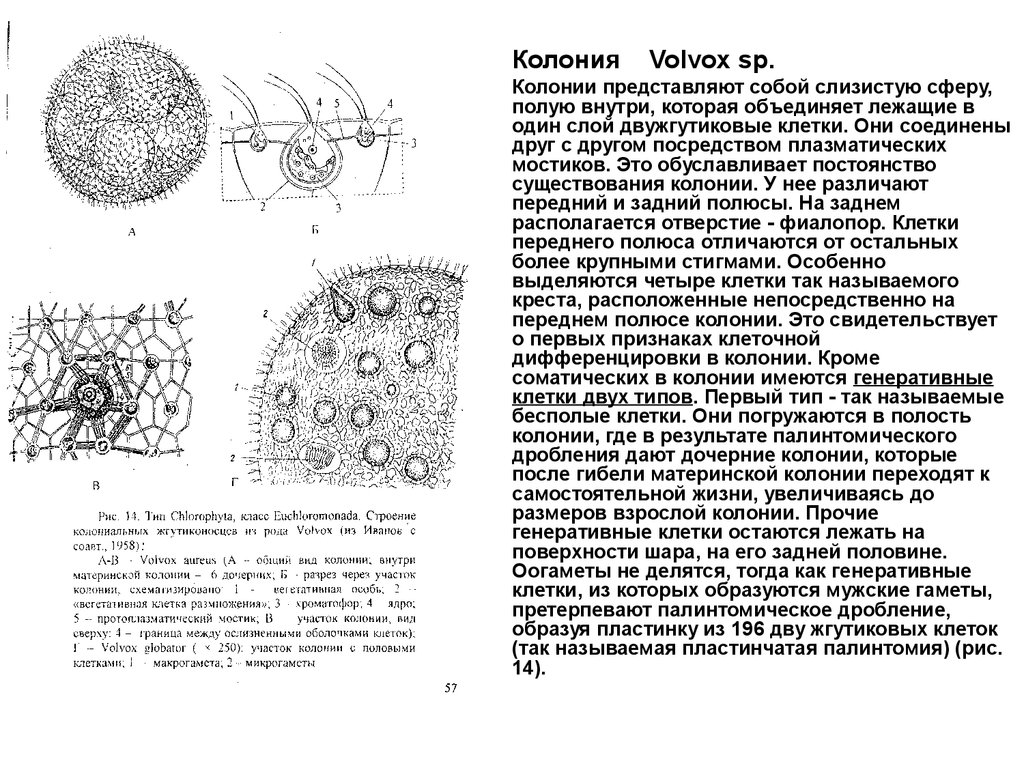
Колония
Колонии представляют собой слизистую сферу,
полую внутри, которая объединяет лежащие в
один слой двужгутиковые клетки. Они соединены
друг с другом посредством плазматических
мостиков. Это обуславливает постоянство
существования колонии. У нее различают
передний и задний полюсы. На заднем
располагается отверстие - фиалопор. Клетки
переднего полюса отличаются от остальных
более крупными стигмами. Особенно
выделяются четыре клетки так называемого
креста, расположенные непосредственно на
переднем полюсе колонии. Это свидетельствует
о первых признаках клеточной
дифференцировки в колонии. Кроме
соматических в колонии имеются генеративные
клетки двух типов. Первый тип - так называемые
бесполые клетки. Они погружаются в полость
колонии, где в результате палинтомического
дробления дают дочерние колонии, которые
после гибели материнской колонии переходят к
самостоятельной жизни, увеличиваясь до
размеров взрослой колонии. Прочие
генеративные клетки остаются лежать на
поверхности шара, на его задней половине.
Оогаметы не делятся, тогда как генеративные
клетки, из которых образуются мужские гаметы,
претерпевают палинтомическое дробление,
образуя пластинку из 196 дву жгутиковых клеток
(так называемая пластинчатая палинтомия) (рис.
14).
Volvox sp.
15.
Колония Volvox sp.
Неповрежденные колонии вольвоксов имеют
правильную сферическую форму (рис. 22, А).
Сколько-нибудь выраженной морфологической
дифференцировки полюсов у таких колоний при
простом микроскопировании обнаружить не
удается. Размеры колоний очень сильно
варьируют в зависимости от возраста. У молодых
и, соответственно, мелких колоний клетки
располагаются компактно и расстояние между
ними очень небольшое, у более старых
экземпляров расстояние между клетками заметно
увеличивается. Рост колонии в подобных случаях
осуществляется не за счет увеличения числа
вегетативных клеток (у видов рода Vоlvох они
вообще утратили способность к делению), а в
результате их расхождения и удаления друг от
друга.
Вегетативные клетки (особи) располагаются
поверхностно, в один слой, так что в центральной
части колонии они всегда отсутствуют. У одних
видов (V. аиreus) все особи, входящие в состав
колонии, соединены с соседними клетками
тонкими ц и т о п л а з м а т и ч е с к и м и м о
с т и к а м и, у других же (V. сиrteri) клетки в
процессе деления полностью обосабливаются
друг от друга.
Клетки одеты клеточной стенкой и заключены в
прозрачный матрикс. Периферический слой этого
матрикса, в который и погружены отдельные
особи, более плотный, а его центральная часть
разжижена.
16.
У различных видов вольвоксов
имеются "разнополые" или
"однополые" колонии. Мужская
гамета, наткнувшись на оогамету,
копулирует с нею. Образовавшаяся
зигота претерпевает редукционное
дробление (зиготическая редукция), а
затем делится митотически.
Образующиеся при палинтомическом
делении клетки образуют
поверхность сферы, но передние
полюса клеток, где в дальнейшем
образуются жгутики, направлены
внутрь (состояние инкурвациии ввернутое внутрь). Когда дробление
завершится, дочерняя колония
выворачивается через фиалопор так,
что теперь передние концы клеток
направлены наружу (явление
экскурвации). После этого колония
переходит к стадии роста (Рис. 15).
Весь комплекс описанных выше
явлений демонстрирует появление
типа жизненного цикла, который
приближается к типичному
жизненному циклу многоклеточных
животных.
17.
• Надтип Polymastigota, Tип Diplomonadida• В состав надтипа входят преимущественно
паразитические жгутиконосцы. В пределах каждого
из типов, объединяемых им, прослеживается
полимеризация (увеличение количества
структурных единиц) жгутикового аппарата.
Структурная единица локомоторной системы
полимастигот (мастигонт) - 2 или 4 кинетосомы со
жгутиками. Для каждого типа характерен свой
специфический набор корневых корешков
кинетосомы. Митохондрии у всех представителей
надтипа отсутствуют.
18.
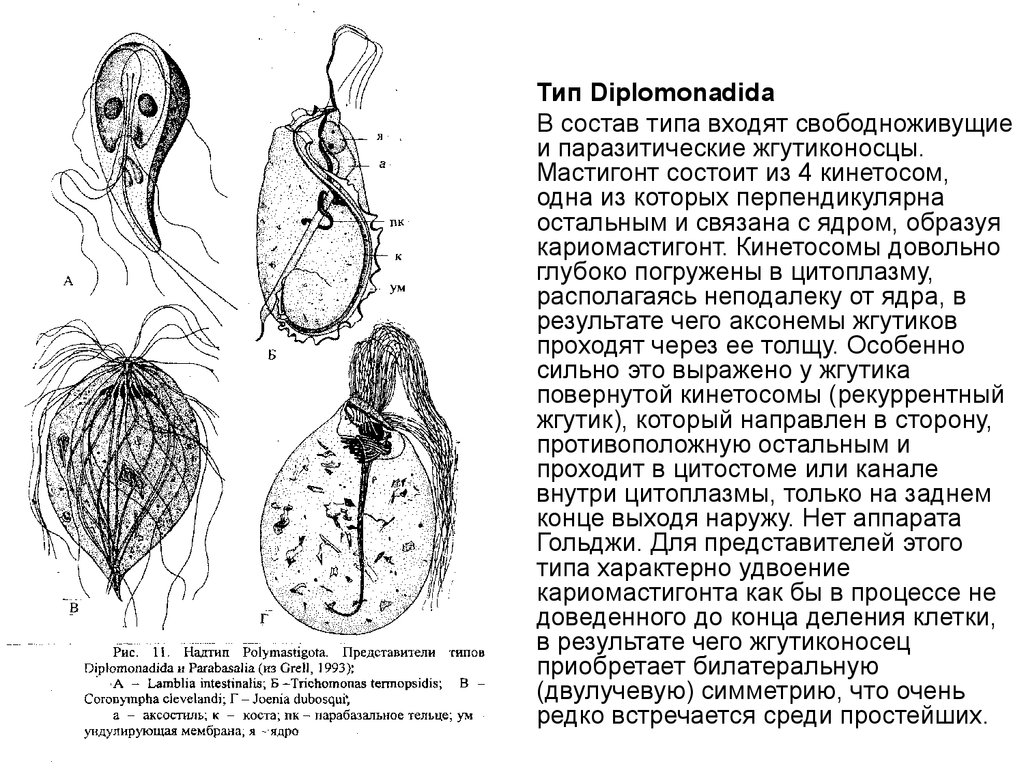
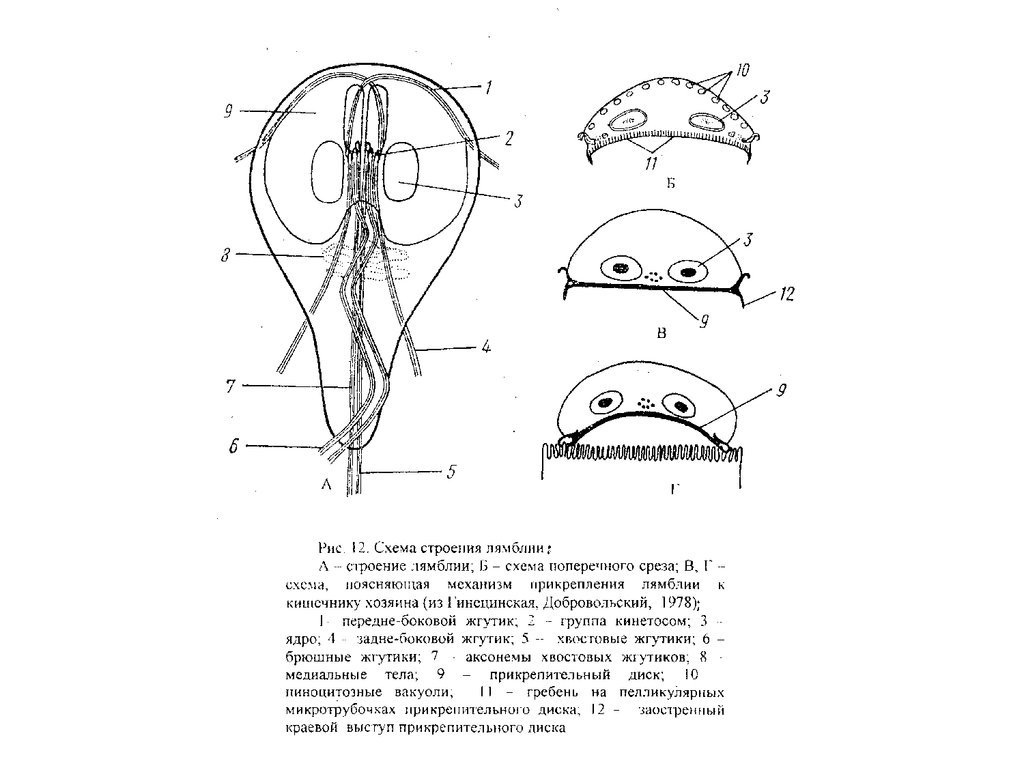
Тип Diplomonadida
В состав типа входят свободноживущие
и паразитические жгутиконосцы.
Мастигонт состоит из 4 кинетосом,
одна из которых перпендикулярна
остальным и связана с ядром, образуя
кариомастигонт. Кинетосомы довольно
глубоко погружены в цитоплазму,
располагаясь неподалеку от ядра, в
результате чего аксонемы жгутиков
проходят через ее толщу. Особенно
сильно это выражено у жгутика
повернутой кинетосомы (рекуррентный
жгутик), который направлен в сторону,
противоположную остальным и
проходит в цитостоме или канале
внутри цитоплазмы, только на заднем
конце выходя наружу. Нет аппарата
Гольджи. Для представителей этого
типа характерно удвоение
кариомастигонта как бы в процессе не
доведенного до конца деления клетки,
в результате чего жгутиконосец
приобретает билатеральную
(двулучевую) симметрию, что очень
редко встречается среди простейших.
19.
20.
21.
• Tип Parabasalia: Класс Trichomonadea,• Класс Hypermastiginеa Тип объединяет исключительно
паразитических жгутиконосцев. Для них характерно наличие
кариомастигонта, образованного 4 кинетосомами и ядром.
Фибриллярные корешки формируют хорошо развитый
цитоскелет. Всегда имеется так называемый парабазальный
аппарат, который представляет из себя аппарат Гольджи. В
клетке имеется особое опорное образование -аксостиль,
который проходит по центральной оси тела.
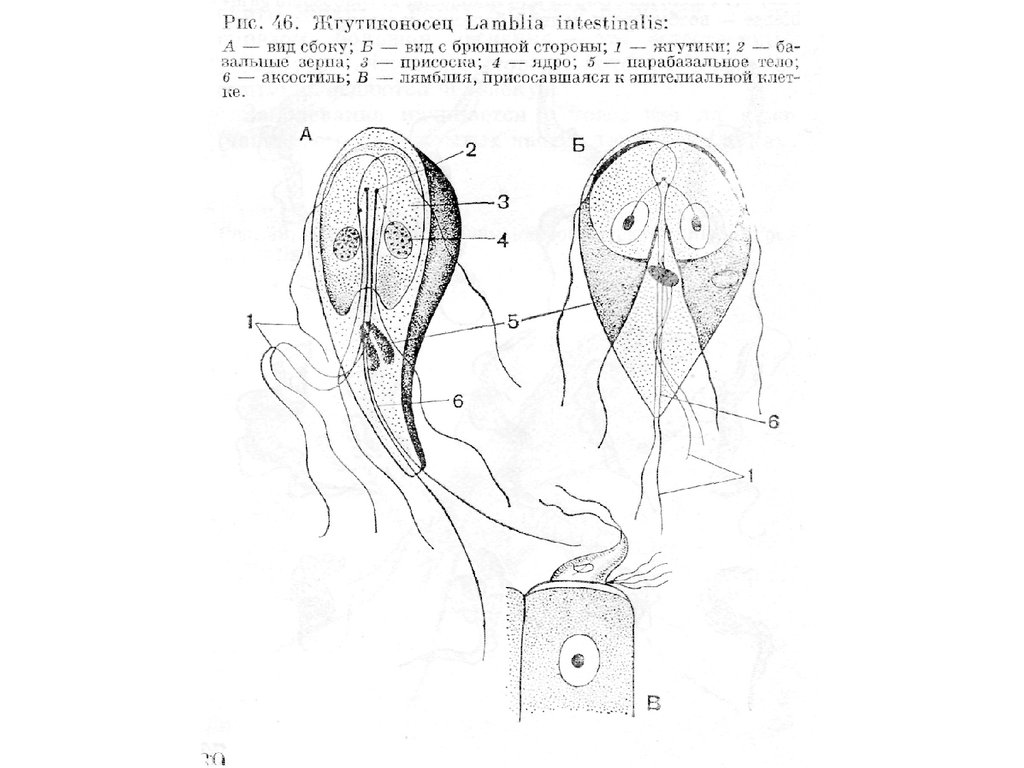
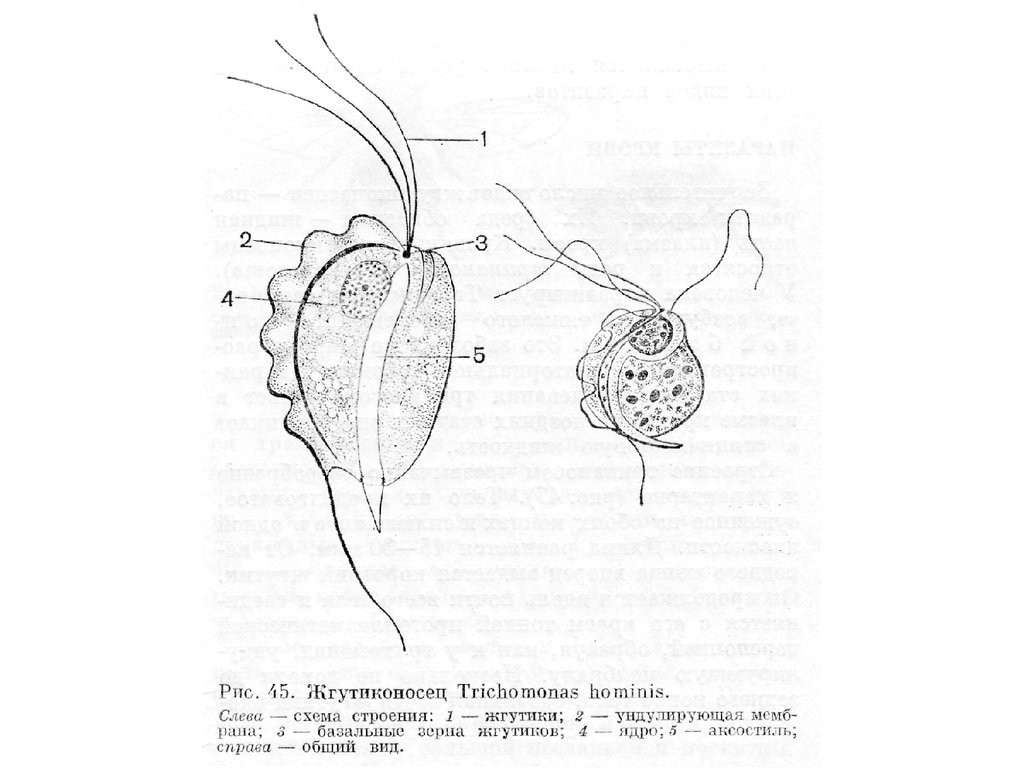
Класс Тrichomonadеа
• Это обширная группа, к которой относятся жгутиконосцы,
обладающие 4-6 жгутиками. Один из них (рекуррентный)
всегда направлен назад и окаймляет идущую вдоль тела
ундулирующую мембрану. Митохондрии отсутствуют.
• Типичными и наиболее известными представителями
класса могут служить жгутиконосцы из рода Trichomonas.
На переднем конце тела трихомонасов имеются четыре
свободных жгутика. Форма клетки овальная, грушевидная
или вытянутая, 5-30 мкм длиной. В передней трети тела
расположено крупное
22.
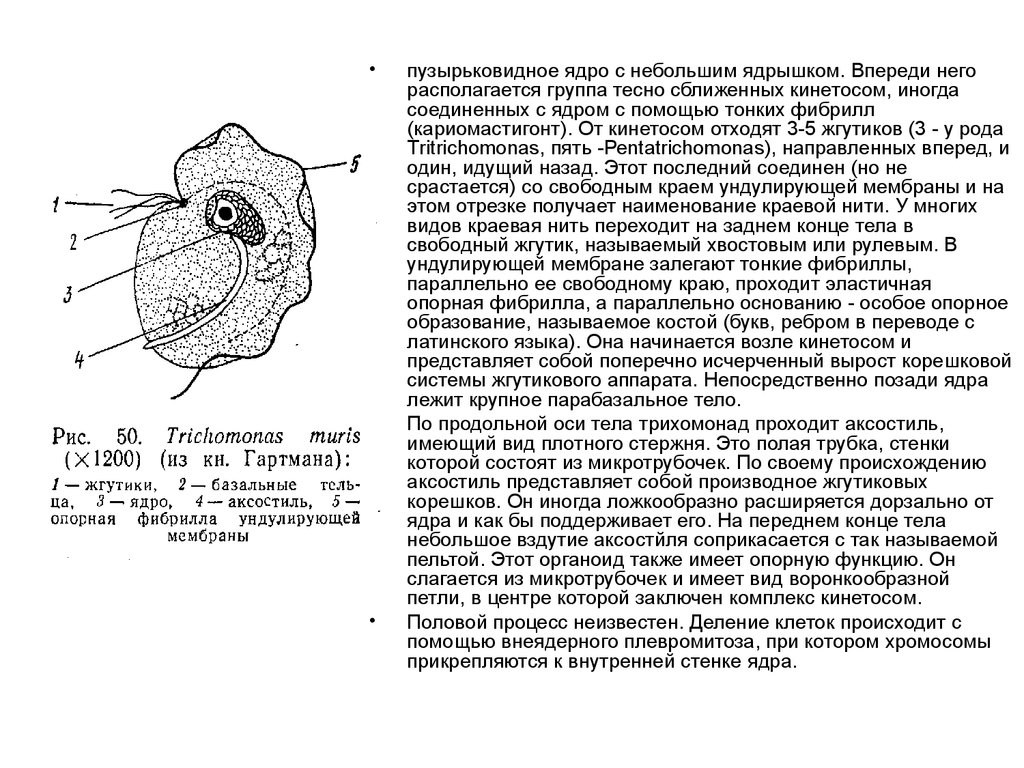
23.
пузырьковидное ядро с небольшим ядрышком. Впереди него
располагается группа тесно сближенных кинетосом, иногда
соединенных с ядром с помощью тонких фибрилл
(кариомастигонт). От кинетосом отходят 3-5 жгутиков (3 - у рода
Тritrichomonas, пять -Реntatrichomonas), направленных вперед, и
один, идущий назад. Этот последний соединен (но не
срастается) со свободным краем ундулирующей мембраны и на
этом отрезке получает наименование краевой нити. У многих
видов краевая нить переходит на заднем конце тела в
свободный жгутик, называемый хвостовым или рулевым. В
ундулирующей мембране залегают тонкие фибриллы,
параллельно ее свободному краю, проходит эластичная
опорная фибрилла, а параллельно основанию - особое опорное
образование, называемое костой (букв, ребром в переводе с
латинского языка). Она начинается возле кинетосом и
представляет собой поперечно исчерченный вырост корешковой
системы жгутикового аппарата. Непосредственно позади ядра
лежит крупное парабазальное тело.
По продольной оси тела трихомонад проходит аксостиль,
имеющий вид плотного стержня. Это полая трубка, стенки
которой состоят из микротрубочек. По своему происхождению
аксостиль представляет собой производное жгутиковых
корешков. Он иногда ложкообразно расширяется дорзально от
ядра и как бы поддерживает его. На переднем конце тела
небольшое вздутие аксостйля соприкасается с так называемой
пельтой. Этот органоид также имеет опорную функцию. Он
слагается из микротрубочек и имеет вид воронкообразной
петли, в центре которой заключен комплекс кинетосом.
Половой процесс неизвестен. Деление клеток происходит с
помощью внеядерного плевромитоза, при котором хромосомы
прикрепляются к внутренней стенке ядра.
24.
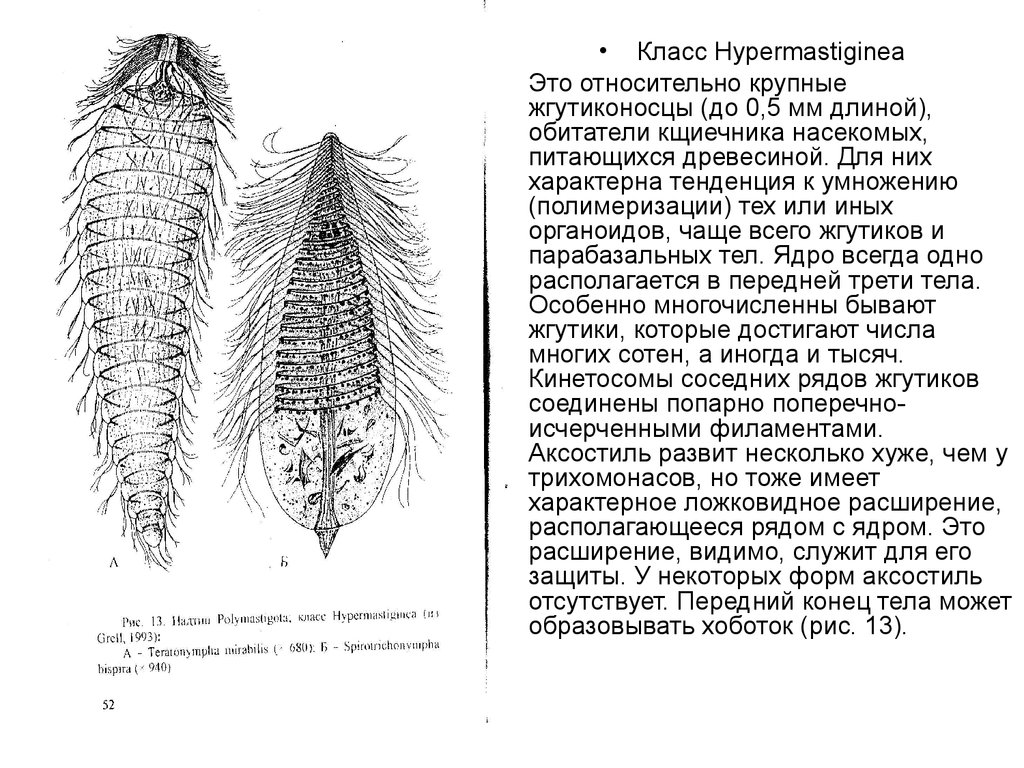
• Класс Нуреrmastiginea
Это относительно крупные
жгутиконосцы (до 0,5 мм длиной),
обитатели кщиечника насекомых,
питающихся древесиной. Для них
характерна тенденция к умножению
(полимеризации) тех или иных
органоидов, чаще всего жгутиков и
парабазальных тел. Ядро всегда одно
располагается в передней трети тела.
Особенно многочисленны бывают
жгутики, которые достигают числа
многих сотен, а иногда и тысяч.
Кинетосомы соседних рядов жгутиков
соединены попарно поперечноисчерченными филаментами.
Аксостиль развит несколько хуже, чем у
трихомонасов, но тоже имеет
характерное ложковидное расширение,
располагающееся рядом с ядром. Это
расширение, видимо, служит для его
защиты. У некоторых форм аксостиль
отсутствует. Передний конец тела может
образовывать хоботок (рис. 13).
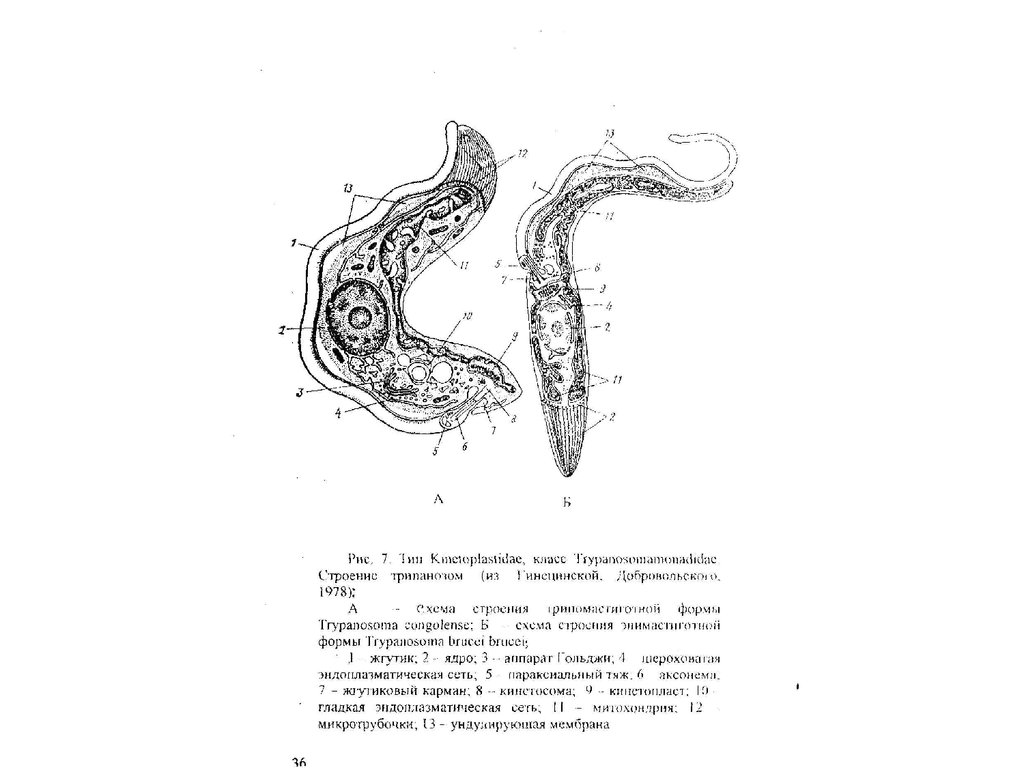
25. Tип Kinetoplastida: Класс Bodomonadida, Класс Trypanosomamonadida
(Тип) Класс Kinetoplastida объединяет в
своем составе облигатно гетеротрофных
свободноживущих и паразитических
жгутиконосцев.
Для всех кипетопластид характерны
следующие признаки.
Тело вытянуто в длину, веретеновидной
формы, иногда образует несколько
изгибов. Редко тело бывает
шаровидным.
Многим кинетопластидам (отряд
Trypanosomatida) присуща способность
изменять морфологические особенности
строения клетки при переходе от одной
стадии развития к другой в процессе
реализации своего жизненного цикла.
Соответственно особи одного и того же
вида на разных этапах жизненною цикла
имеют разное строение и относятся к
разным морфам, или морфотипам
(трипомастиготы, эпимастиготы,
промастиготы, опистомастиготы,
амастиготы и др.). Жизненный цикл
каждого конкретного вида включает
постоянный набор определенных
морфотипов.
26.
В клетке кинетопластид имеется
только одна очень крупная,
разветвленная митохондрия. Ее
отростки пронизывают весь объем
клетки. В непосредственной
близости от кинетосомы жгутика
расположен специализированный
участок митохондрии (иногда он
обособлен небольшой перетяжкой),
в котором сконцентрирована вся
митохондриальная ДНК в виде
компактного тельца —
к и н е т о п л а с т а. Кинетопласт
выявляется ядерными
красителями и хорошо виден в
обычный световой микроскоп.
Размножаются продольным
бинарным делением, реже
почкованием.
Половой процесс неизвестен.
27.
28.
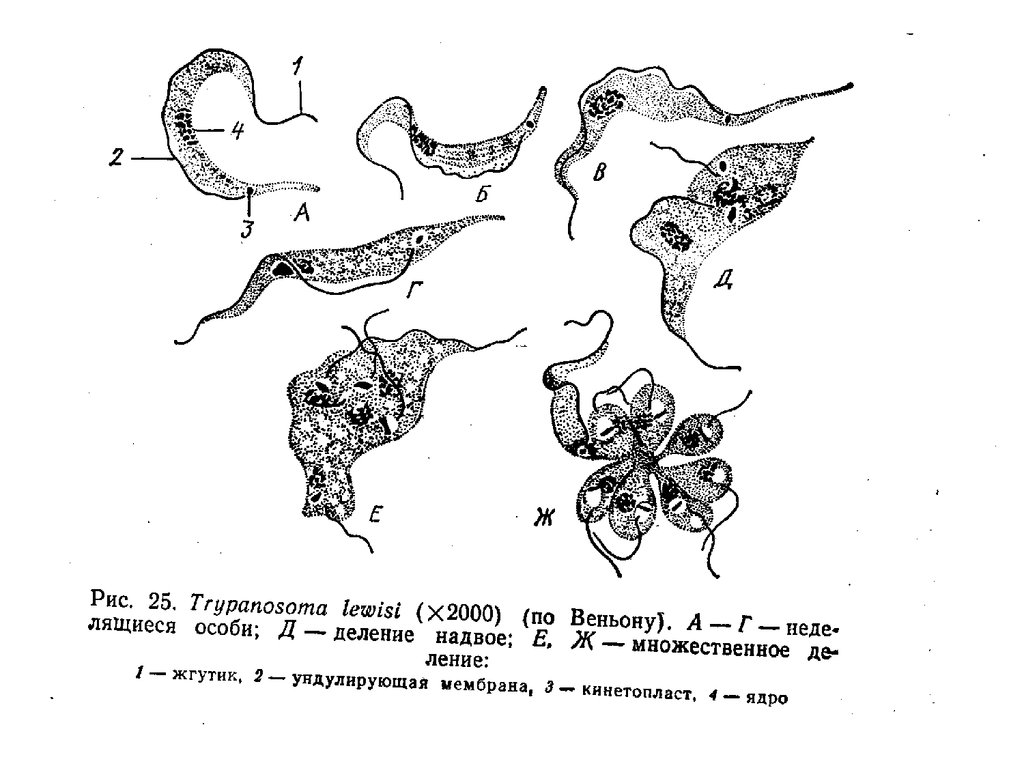
Trypanosoma sp.
Трипаносомы изучаются на окрашенных
мазках крови зараженных животных.
Между эритроцитами отчетливо
выделяются ярко окрашенные,
червеобразно удлиненные и
причудливо изогнутые жгутиконосцы
(рис.).
Тело трипаносом сильно вытянуто в
длину и заострено с обоих концов
(рис.}. В большинстве случаев передний
конец оттянут значительно сильнее, чем
задний. В зависимости от модификации
окраски, использованной при обработке
мазков, цитоплазма трипаносом может
быть светло-голубой или бледнорозовой. В средней части клетки
располагается овальное ядро,
окрашенное в темно-красный или
красно-фиолетовый цвет. В заднем
конце тела отчетливо различимы два
небольших тельца, как правило,
лежащих друг за другом. Одно, более
крупное, занимает самое заднее
положение в клетке. Оно окрашивается
подобно ядру и имеет вид плотной,
слабо преломляющей свет гранулы. Это
и есть к и н е т о п л ас т — плотное
скопление кольцевых молекул
митохондриальной ДНК.
29.
• Немного впереди кинетопласта расположена к и н е т ос о м а (базальное тельце), от которой берет началожгутик. Рассмотреть жгутиковый карман и тем более
открывающийся в него цитостом на тотальных
препаратах невозможно.
• Выйдя из кармана на поверхность клетки, жгутик
поворачивает вперед. Он прочно связан с прозрачным и
тонким цитоплазматическим гребнем—ундулирующ ей
мембраной, которая тянется вдоль всего тела
трипаносомы вплоть до его переднего конца. Жгутик
проходит по вершине мембраны и, достигнув конца тела,
продолжается вперед в виде свободного дистального
участка. Рассмотреть ундулирующую мембрану на всем
ее протяжении удается редко. Обычно хорошо видны
лишь отдельные широкие складки, выступающие за
границы тела. Участки мембраны, оказавшиеся на фоне
тела, увидеть бывает нелегко.
30.
Возбудитель другой очень широко
распространенного и известного широкому кругу
людей заболевания, так называемой сонной
болезни, обитает в Тропической Африке. Это
Тripanosoma gambiense. Она может служит
примером максимально упрощенного
жизненного цикла среди трипаносом.
Хотя сонная болезнь известна очень давно,
природа этого недуга была выяснена лишь в
начале нашего столетия благодаря Д.Даттону,
который в 1902 году в Западной Африке
обнаружил трипаносом, паразитирующих в
крови человека, А.Кастеллани, впервые (в 1903
году) установившему связь между
трипаносомами и сонной болезнью, и, наконец,
Давиду Брюсу, расшифровавшему цикл
развития жгутиконосцев в организме и
доказавшему участие мух це-це (Glossina
palpalis, G. morsitans и др.) в распространении
заболевания (Рис. 10).
Тripanosoma gambiense в переносчике
развивается, проходя две стадии:
трипомастигот, размножающихся в кишечнике
мухи, и эпимастигот, поселяющихся в ее
слюнных железах. Эпимастиготы делятся, а
затем дают начало трипомастиготам,
занимающим "переднюю позицию" (слюнные
железы) в готовности к заражению. Оно
происходит инокулятивно, т.е. в процессе акта
сосания крови, когда в кровь может попасть до
40 тысяч трипаносом, хотя для инфицирования
достаточно 250-350 особей. Первые 9-10 дней
жгутиконосцы обитают в подкожной клетчатке, а
затем постепенно переходят в кровяное русло и
попадают в спинно-мозговую жидкость.
Тripanosoma cruci – болезнь Чагаса в Лат.
Америке. Ререносчики поцелуйные клопы
31.
32.
33.
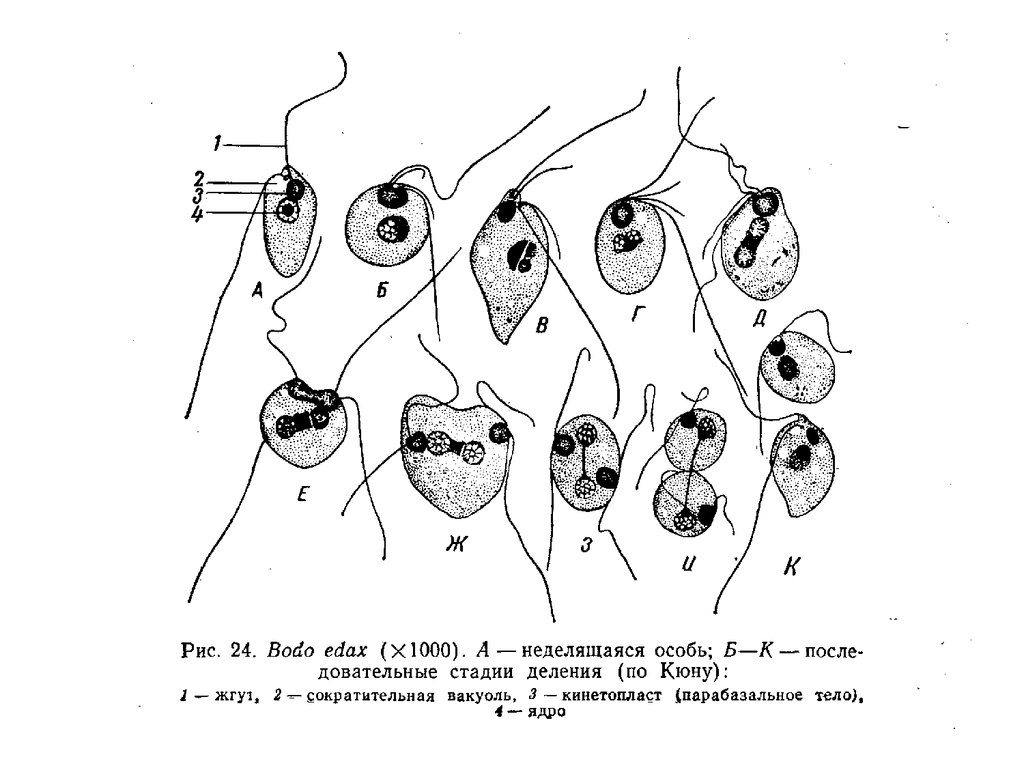
отряд Bodonida
Жгутиков два - начинаются на дне
жгутикового кармана, положение
которого на теле жгутиконосца
варьрирует. У некоторых форм со
жгутиком (или с одним из
жгутиков) связана так называемая
ундулирующая мембрана —
ограниченная поверхностной
мембраной тонкая упругая
складка, к гребню которой и
крепится жгутик.
Покровы клетки представлены
пелликулой особого типа — т у б у
л е м м о й. Непосредственно под
плазмалеммой в один слой
располагаются много
численные продольные
микротрубочки.
Цитостом в большинстве случаев
имеется — в жгутиковый карман
открывается тонкий трубчатый
канал, на свободном конце
которого образуются
пиноцитозные вакуоли.
34.
35.
Трипаносоматиды освоили широкий круг хозяев - от
простейших до позвоночных животных, в том числе
человека. Представители рода Phytomonas обитают в
млечном соке некоторых растений. Их переносчиками служат
растительноядные клопы. Насекомые являются наиболее
обычными хозяевами трипаносоматид, особенно
кровососущие, которые служат переносчиками паразита. Для
человека наибольшее значение представляют
Трипаносоматиды двух родов - Leischmania и Тгураnosoma.
Они служат источниками целого ряда тяжелых заболеваний
домашнего скота и людей.
Лейшмании используют в качестве хозяев широкий
круг позвоночных животных: рыб, рептилий,
млекопитающих, в том числе человека, вызывая у него
тяжелые заболевания лейшманиозы.
Переносчиками для большинства видов лейшманий служат
мелкие кровососущие насекомые - москиты рода Phlebotomus
(рис. 8В). Москиты, питаясь на позвоночном животном, вносят
в его ткани подвижные промастиготные формы лейшманий.
Последние заглатываются лейкоцитами-макрофагами, но не
перевариваются, а превращаются в амастигот, переходя
таким образом к внутриклеточному паразитированию. В
клетках хозяина амастиготы интенсивно делятся. Образуются
плотные скопления паразитов, так называемые
"псевдоцисты". С течением времени клетка разрушается и
паразиты попадают в кровь, заражая новые, еще здоровые
клетки.
Заражение лейшманиями москита происходит при питании
его на больном животном. Вместе с кровью в
пищеварительную систему москита попадают и амастиготы.
Здесь они превращаются в промастигот, которые
расселяются по пищеварительному тракту хозяина и
приступают к размножению. Численность паразитов заметно
увеличивается. Они постепенно перемещаются в передние
отделы пищеварительной системы, а оттуда поступают в
хоботок. С этого момента переносчик способен заражать
новых, еще не зараженных хозяев - позвоночных (рис. 8).
36.
Лейшманиозы человека представлены двумя основными
типами заболеваний - это так называемый кожный
лейшманиоз, при котором паразиты локализуются в коже и
подкожной клетчатке, и висцеральный или общий, при
котором паразитические жгутиконосцы приурочены к
различным внутренним органам и поэтому широко
поражают организм хозяина.
Кожный лейшманиоз или пендинская язва встречается
всюду, где обитают москиты Phlebotomus papatasii,
переносящие возбудителя этой болезни – Leischmania
tropica. Естественным резервуаром болезни служат
суслики, песчанки и другие грызуны, чьи поселения
окружают деревни и поселки. В их норах в трещинах
почвы и скрываются москиты.
Первые внешние проявления заболевания становятся
заметными лишь через 3-8 месяцев (а иногда через 1,5
года) после попадания паразита в организм человека.
Сначала на поверхности кожи, преимущественно лица и
рук, которые чаще всего подвергаются нападению
москитов, появляется небольшой бугорок, приобретающий
буровато-красный цвет. В это время паразит интенсивно
размножается в подкожной клетчатке. На месте бугорка
через 3-6 месяцев образуется сухая корка, прикрывающая
небольшую язву. Заболевание в среднем продолжается
около года, после чего язва начинает рубцеваться. Следы
от язвы часто сохраняются у человека на всю жизнь.
Возбудителем висцерального лейшманиоза, или калаазара, является L. donovani. Инкубационный период
продолжается в среднем 3-5 месяцев. Болезнь имеет
характер неправильной перемежающейся лихорадки.
Паразиты интенсивно размножаются в клетках различных
внутренних органов - печени, селезенки, лимфатических
узлов, стенок кишечника и т.п. В пораженных органах
образуются участки омертвевших тканей, стенки
кишечника изъязвляются. Для этой болезни характерна
также сильная анемия. Особенно опасен кала-азар для
детей, у которых она часто заканчивается смертью.
37.
проникают в кровеносную и лимфатическую системы. Со временем они
поселяются в спинномозговой жидкости, откуда уже попадают в
мозговую ткань, поражая центральную нервную систему. Это
обуславливает и особенности течения сонной болезни.
Заболевание представлено двумя формами - острой и хронической,
отличающимися друг от друга характерами симптомов и скоростью
развития. Инкубационный период длится от 1,5-3 недель до двух лет и
более. Миграция паразитов из подкожной клетчатки в кровь и
лимфатическую систему сопровождается появлением лихорадки и
воспалением лимфатических узлов. Наблюдается потеря аппетита,
головокружения, уменьшается частота пульса. При хронической форме
этот период продолжается от нескольких месяцев до нескольких лет,
при острой - всего несколько недель. Последующее нарушение нервной
системы в конечном итоге приводит к впадению в коматозное
состояние. Продолжительность болезни при хронической форме
составляет несколько лет, при острой - не более 6-9 месяцев. При
отсутствии лечения больной чаще всего погибает.
Возбудитель хронической формы болезни циркулирует среди
населения по берегам крупных рек. Возбудители острой формы
распространены главным образом в природных очагах, приуроченных к
зоне саванн. Резервуарными хозяевами служат антилопы.
Сонная болезнь легче всего поддается лечению на ранних стадиях.
Иммунитет к ней не вырабатывается из-за сложной и изменчивой
системы антигенов, содержащихся в поверхностной пленке
трипаносом. Состав антигенов меняется так быстро, что организм
человека не успевает выработать к ним антитела.