














































 biology
biologySimilar presentations:










Актуальные проблемы структурной эволюции растений
1.
Актуальные проблемы структурнойэволюции растений
Предмет: становление вегетативного тела в ходе
морфологической эволюции растений
2.
Высшие растенияОпределение,
происхождение и
предполагаемые предки
3.
Выход растений на сушу• Могли переносить иссушение
(покровная ткань, кутикула,
паренхиматозное строение)
• Могли осуществлять оксигенный
фотосинтез (устьица и аэренхима)
• “Расплата” за открытые устьица транспирация 99,8% воды необходимость постоянно
всасывать воду
4.
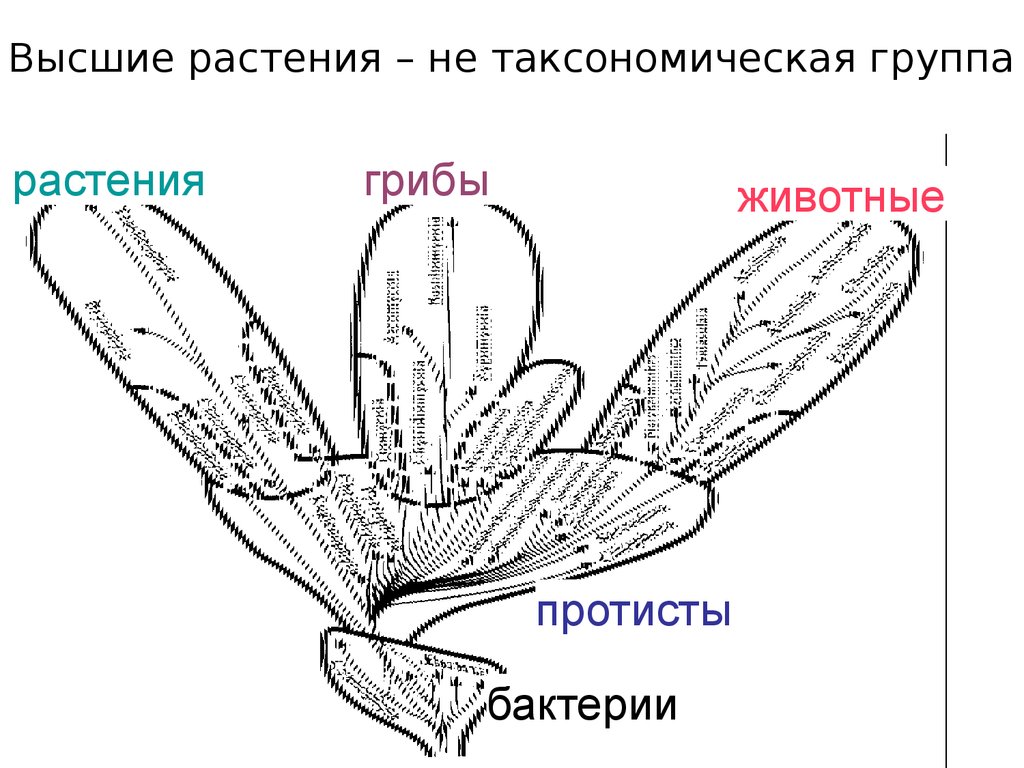
Высшие растения – не таксономическая группарастения
грибы
животные
протисты
бактерии
5.
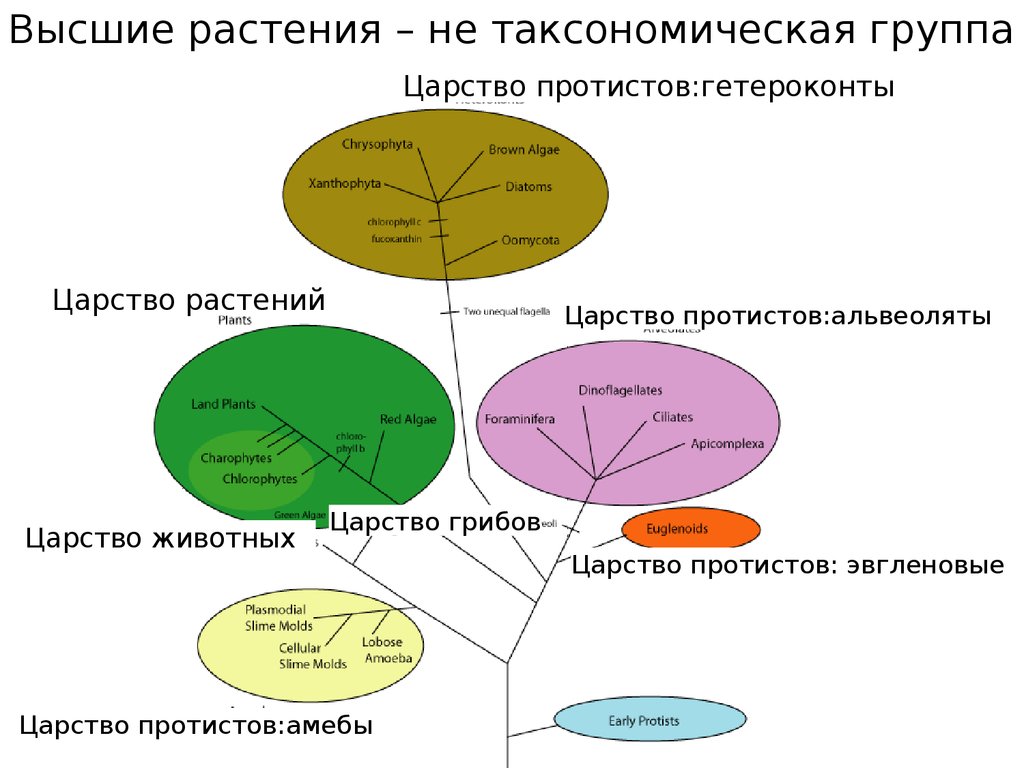
Высшие растения – не таксономическая группаЦарство протистов:гетероконты
Царство растений
Царство животных
Царство протистов:альвеоляты
Царство грибов
Царство протистов:амебы
Царство протистов: эвгленовые
6.
Отд. Chlorophyta, зеленые водоросли Хлорофиллы а, b.Предположительно хлоропласты - результат первичного
эндосимбиоза с цианобактериями. Морфологическое
разнообразие талломов. Все варианты жизненного цикла.
Chlorokybus
Zygnema
Ulva
7.
Вероятные предки высших растений - класс Charophyceae (отд.Charophyta).
Nitella
Charales
Chara
Coleochaete
8.
Харовые водоросли. Признаки, общиес высшими растениями.
Образуют фрагмопласт в процессе цитокинеза.
Клеточная стенка из целлюлозы, гемицеллюлозы, пектиновых
веществ.
Имеют плазмодесмы.
Паренхиматозное строение.
Вещество запаса - крахмал откладывается в пластидах.
9.
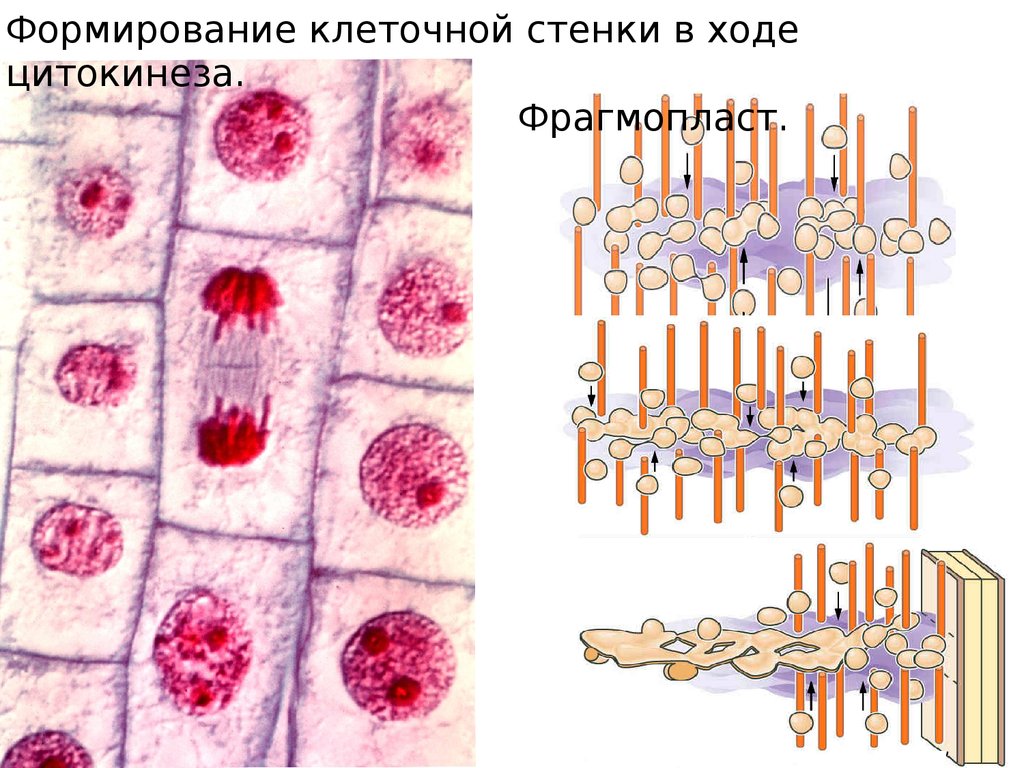
Формирование клеточной стенки в ходецитокинеза.
Фрагмопласт.
10.
Харовые водоросли. Признаки, общиес высшими растениями.
Образуют фрагмопласт в процессе цитокинеза.
Вещество запаса - крахмал откладывается в пластидах.
Клеточная стенка из целлюлозы, гемицеллюлозы, пектиновых
веществ.
Имеют плазмодесмы.
Паренхиматозное строение.
11.
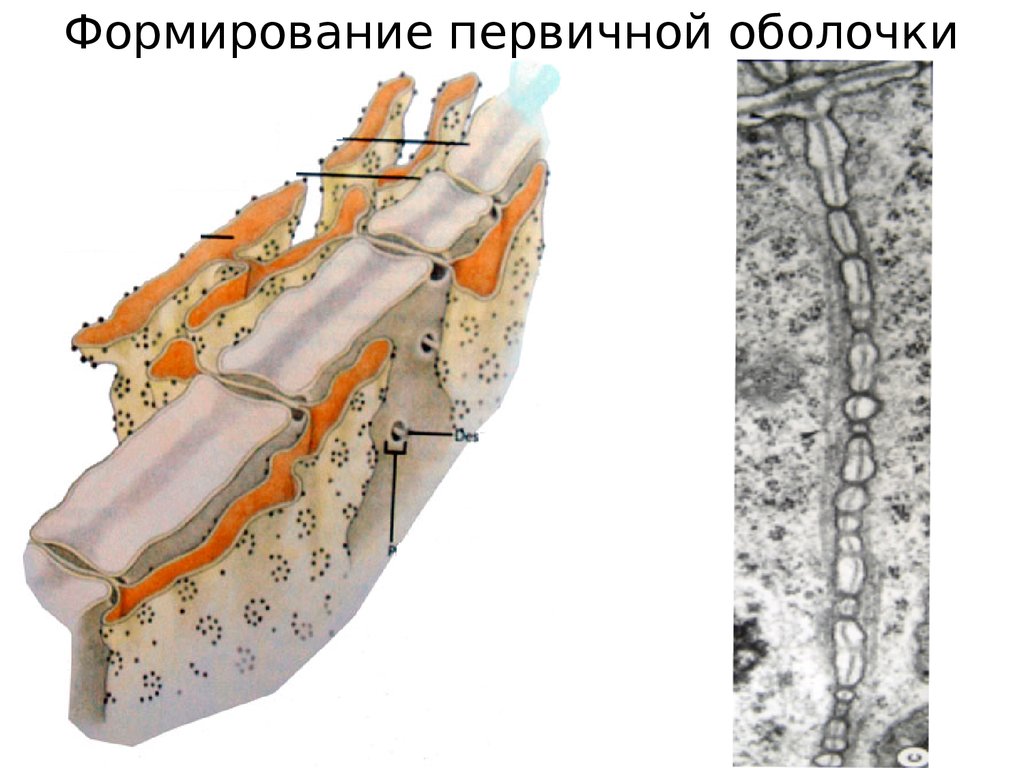
Формирование первичной оболочки12.
Харовые водоросли. Признаки, общиес высшими растениями.
Образуют фрагмопласт в процессе цитокинеза.
Вещество запаса - крахмал откладывается в пластидах.
Клеточная стенка из целлюлозы, гемицеллюлозы, пектиновых
веществ.
Имеют плазмодесмы.
Паренхиматозное строение.
13.
Вероятные предки высших растений - класс Charophyceae (отд.Charophyta).
Nitella
Charales
Chara
Coleochaete
14.
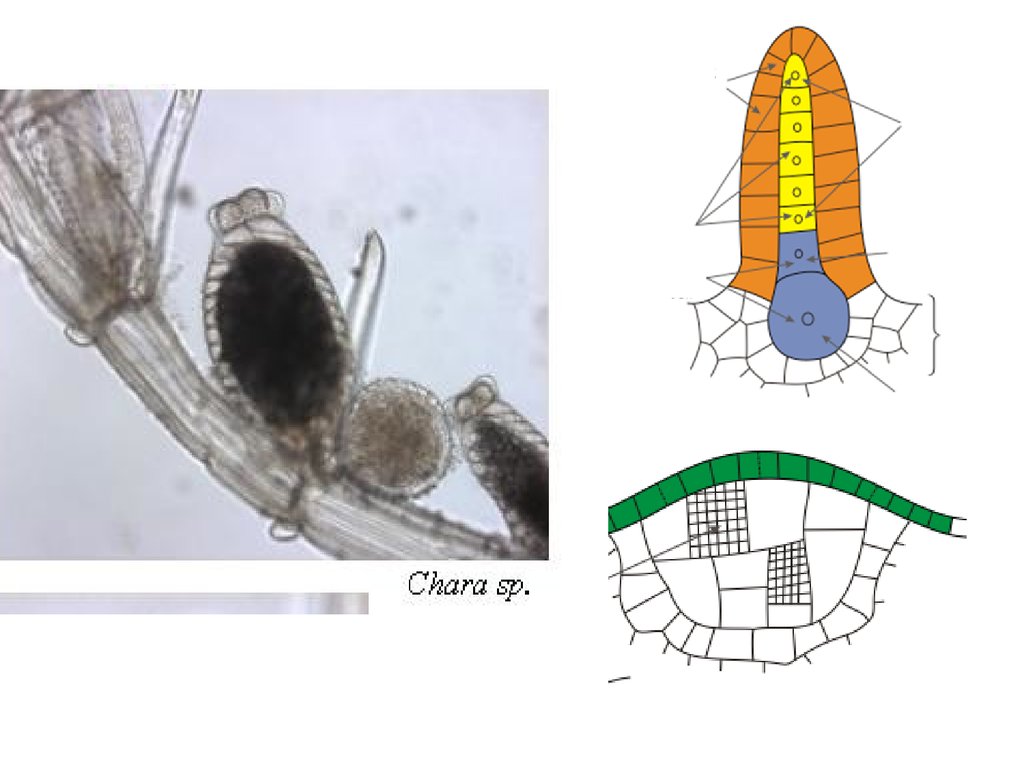
Таллом Chara - гаметофитмногоклеточные гаметангии, оогамный половой процесс, гаплофазный
жизненный цикл с зиготической редукцией.
15.
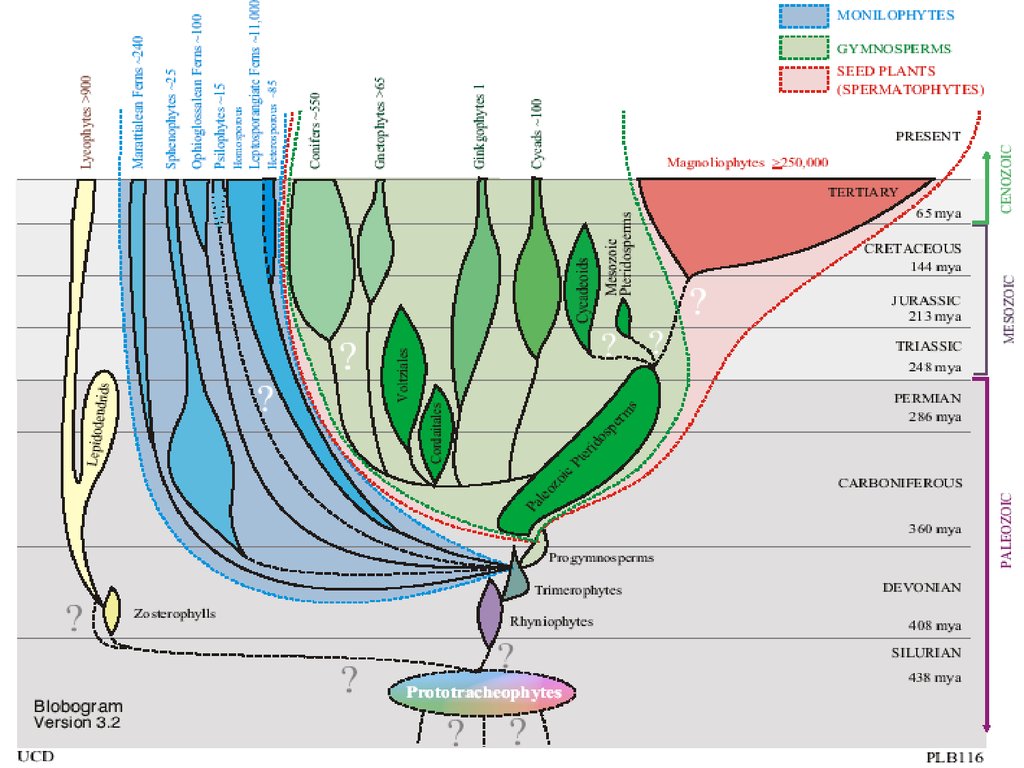
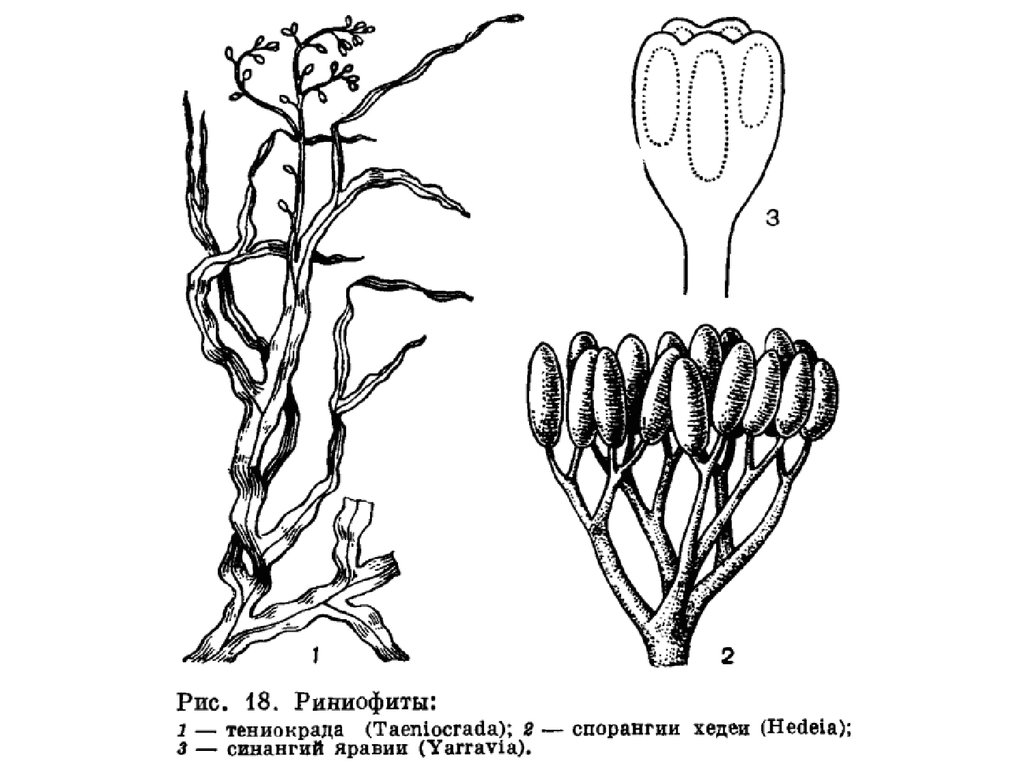
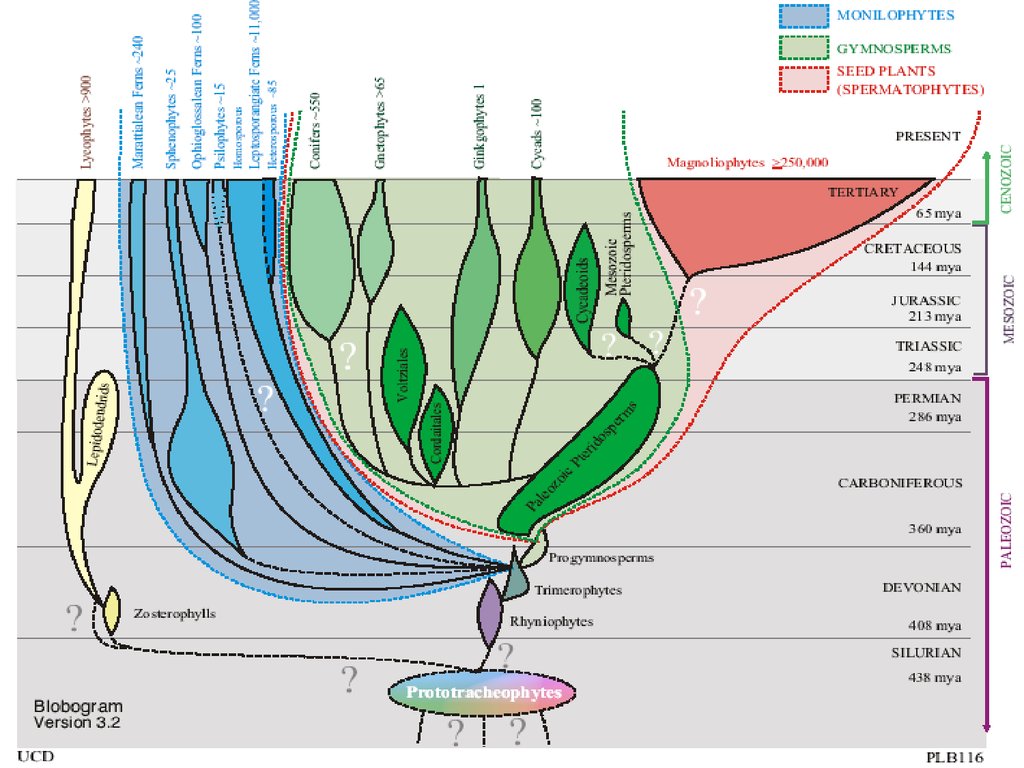
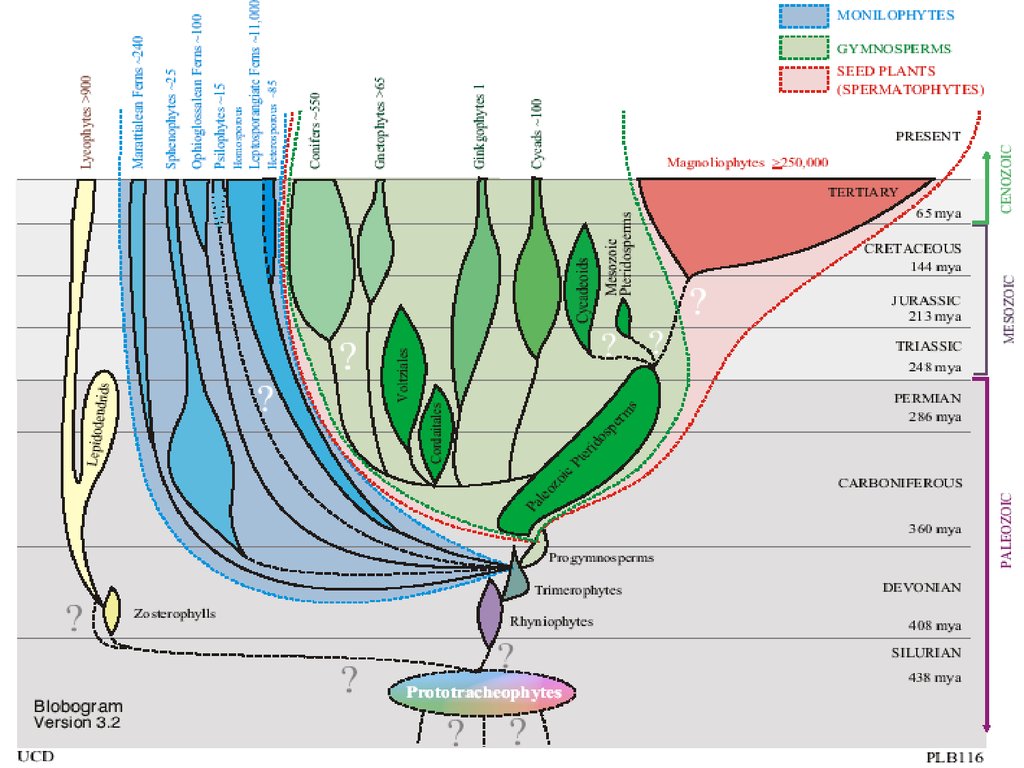
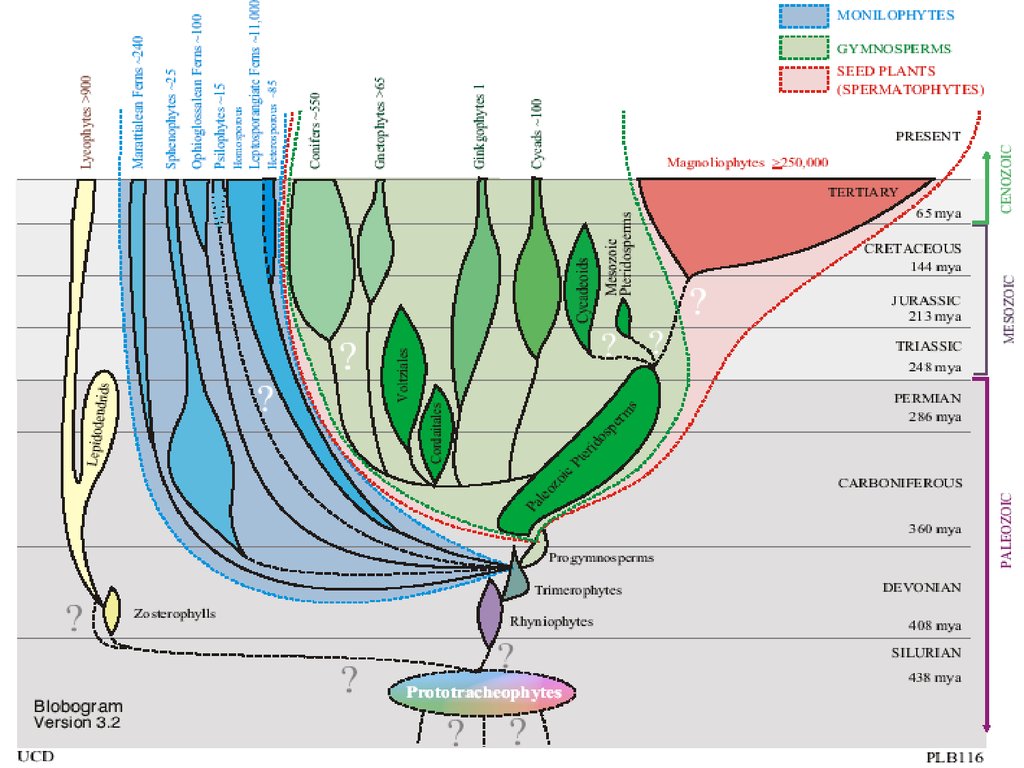
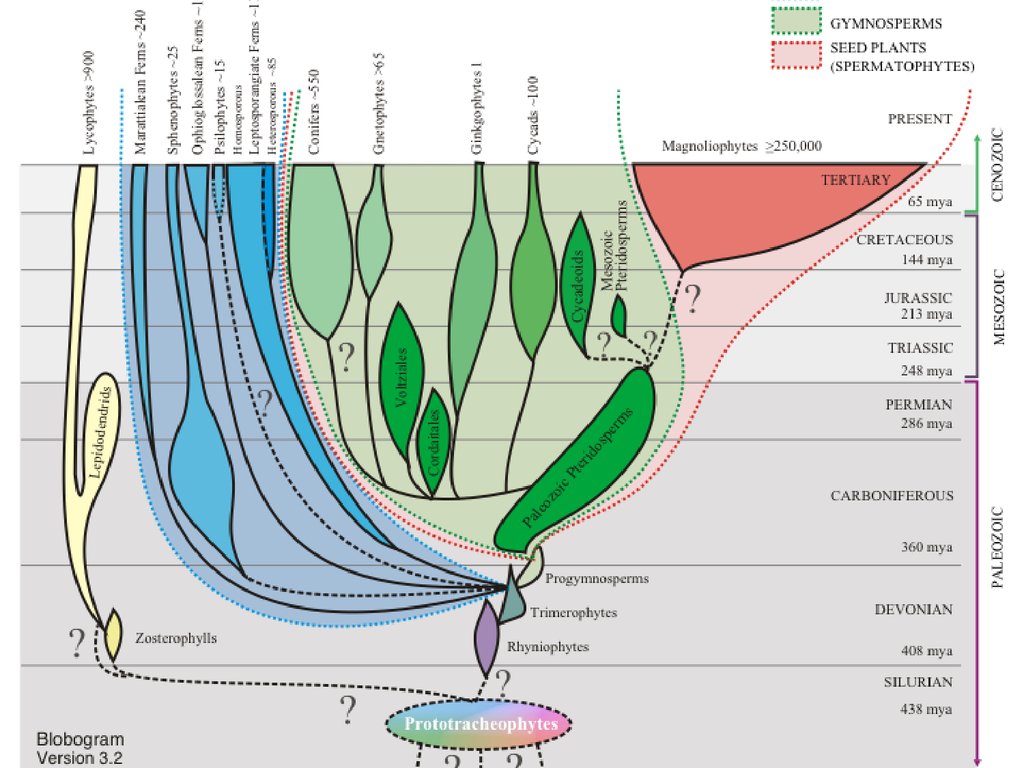
Отделы высших растенийI. Отдел RHYNIOPHYTA (РИНИОФИТЫ, ПСИЛОФИТЫ)
II. Отдел BRYOPHYTA (Marchantiophyta, Anthocerophya, Bryophyta)
– 25 000 видов (МОХООБРАЗНЫЕ)
III. Отдел PSILOTOPHYTA - 12 видов (ПСИЛОТООБРАЗНЫЕ)
IV. Отдел LYCOPODIOPHYTA – 1 100 видов (ПЛАУНООБРАЗНЫЕ)
V. Отдел EQUISETOPHYTA – 30 видов (ХВОЩЕОБРАЗНЫЕ)
VI. Отдел POLYPODIOPHYTA – 12 000 видов
(ПАПОРОТНИКООБРАЗНЫЕ)
VII. Отдел PINOPHYTA - 800 видов (ГОЛОСЕМЕННЫЕ)
VIII. Отдел MAGNOLIOPHYTA – 300 000 видов (ЦВЕТКОВЫЕ,
ПОКРЫТОСЕМЕННЫЕ)
16.
Признаки высшихрастений
17.
Признаки высшихрастений
1. Гаплодиплофазный
жизненный цикл
18.
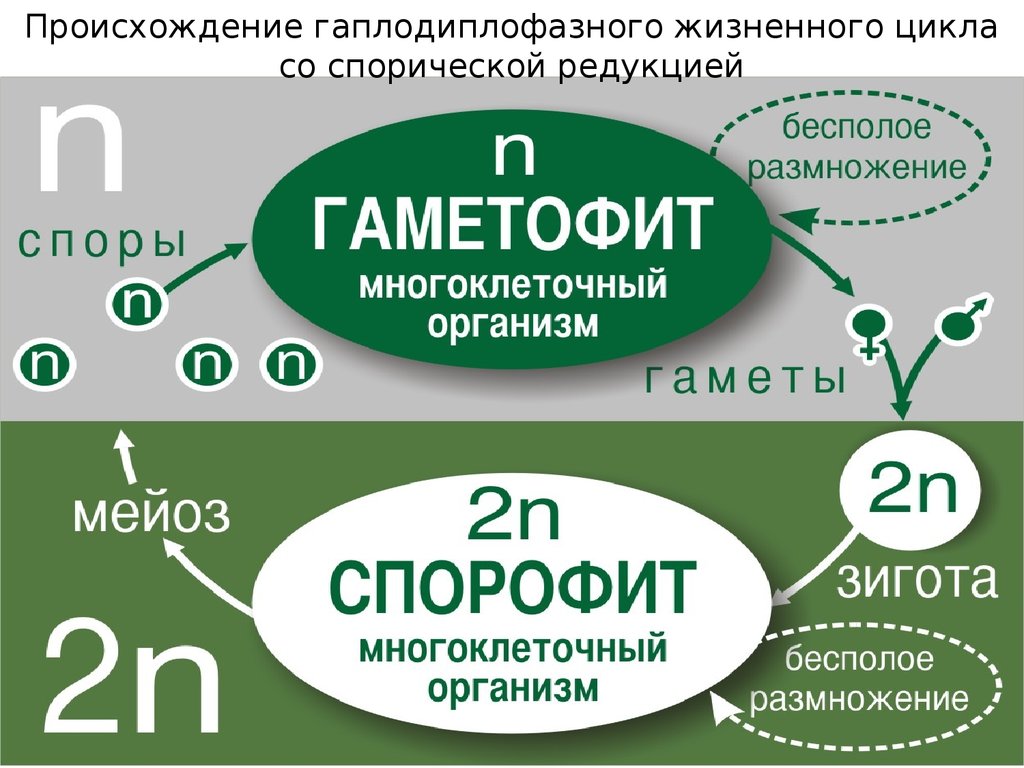
Происхождение гаплодиплофазного жизненного цикласо спорической редукцией
19.
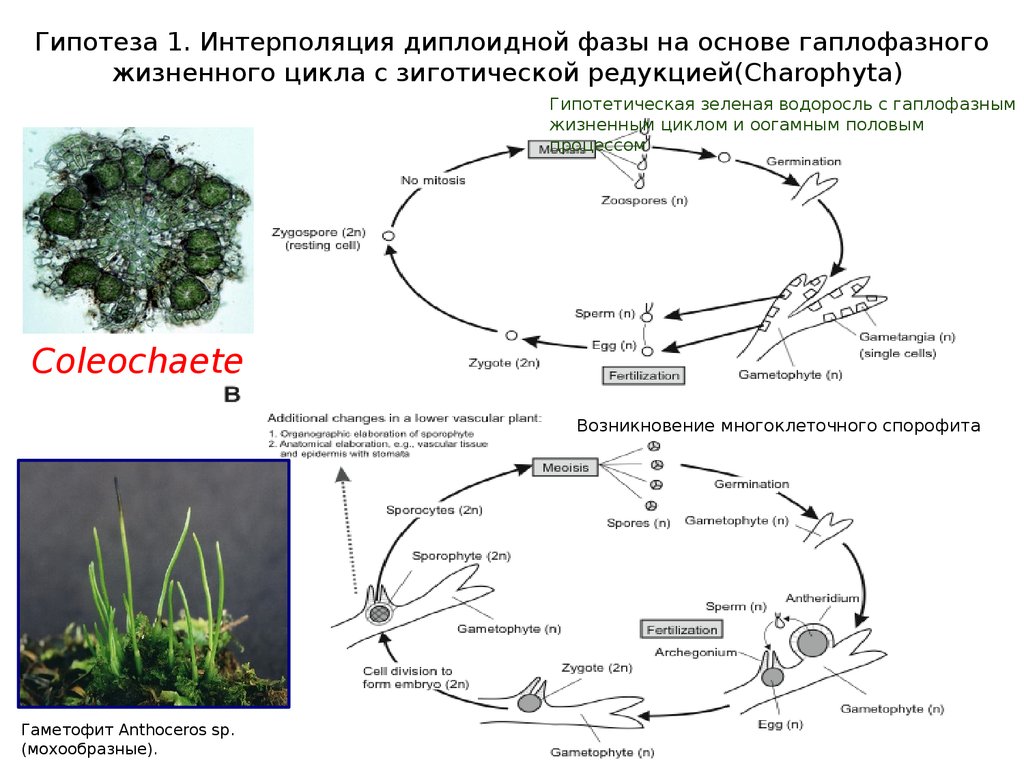
Гипотеза 1. Интерполяция диплоидной фазы на основе гаплофазногожизненного цикла с зиготической редукцией(Charophyta)
Гипотетическая зеленая водоросль с гаплофазным
жизненным циклом и оогамным половым
процессом
Coleochaete
Возникновение многоклеточного спорофита
Гаметофит Anthoceros sp.
(мохообразные).
20.
Гипотеза 1. Интерполяция диплоидной фазы на основе гаплофазного жизненного циклас оогамным половым процессом изиготической релукцией (Chlorophyta, Charophyta)
Coleochaete
Гаметофит Anthoceros sp. (мохообразные).
У первых наземных растений доминировал гаметофит. Усложнение спорофита
происходило постепенно. Высшие растения - монофилетическая группа.
21.
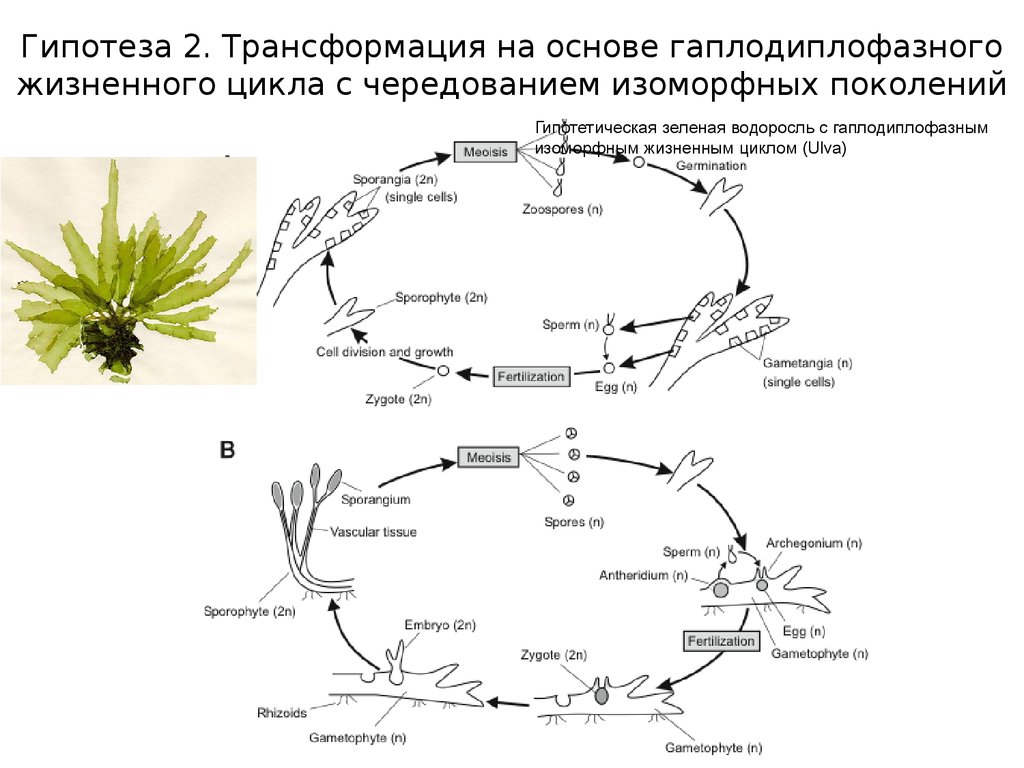
Гипотеза 2. Трансформация на основе гаплодиплофазногожизненного цикла с чередованием изоморфных поколений
Гипотетическая зеленая водоросль с гаплодиплофазным
изоморфным жизненным циклом (Ulva)
22.
Гипотеза 2. Трансформация на основе гаплодиплофазного жизненного цикла счередованием изоморфных поколений (Chlorophyta, Phaeophyta, Rhodophyta)
У первых наземных растений НЕ доминировал гаметофит. Высшие
растения “спорофитной” и “гаметофитной” линии возникли от разных
групп зеленых водорослей
23.
Признаки высшихрастений
2. Строение репродуктивных органов.
Многоклеточные гаметангии.
24.
25.
Признаки высшихрастений
3. Строение репродуктивных органов.
Многоклеточные спорангии.
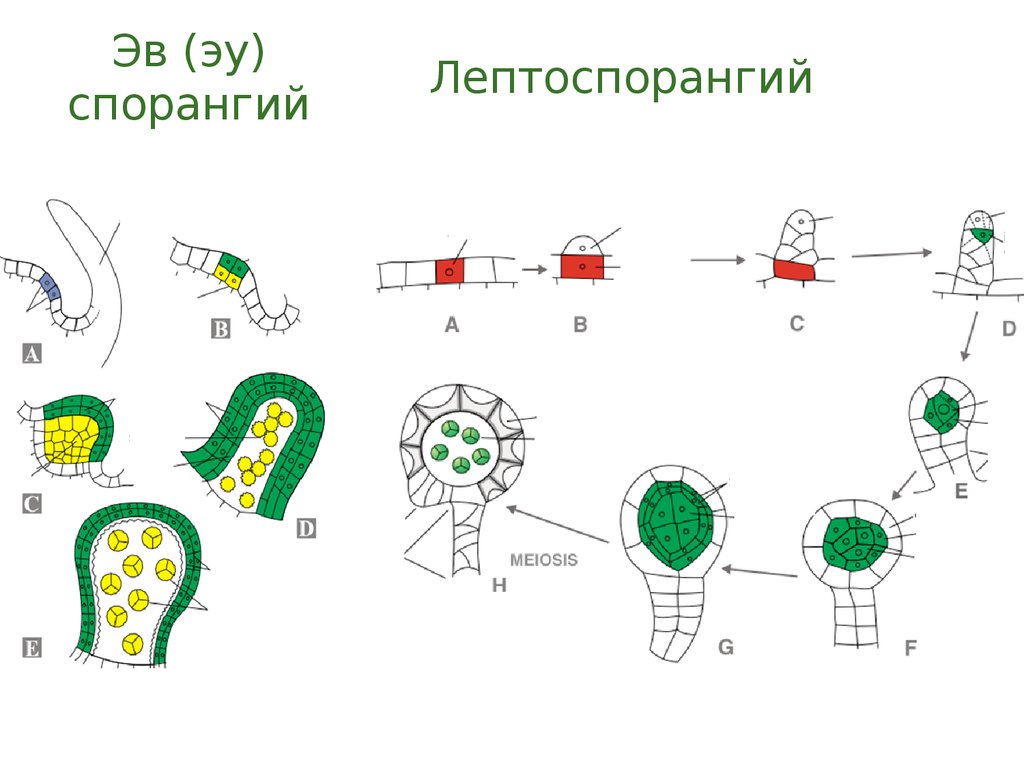
26.
Эв (эу)спорангий
Лептоспорангий
27.
Признаки высших растений4. Фаза зародыща в
онтогенезе.
EMBRYOPHYTA
28.
Признаки высших растений5. Открытый рост. Наличие потенциально бессмертных
образовательных тканей - апикальных меристем.
29.
Признаки высших растений6. Тело разделено на органы. CORMOPHYTA
(THALLOPHYTA)
Наиболее распространенная точка
зрения: два типа вегетативных органов побег и корень. Есть и другие мнения.
Вегетативное тело имеет
метамерное строение.
30.
Признаки высших растений7. Тело дифференцировано на ткани.
Системы тканей.
1.
2.
3.
4.
5.
Образовательные ткани
Проводящие
Покровные
Ассимиляционные (фотосинтезирующие)
Механические
6. Запасающие
7. Всасывающие
8. Выделительные
9. Пограничные
10. Аэренхима, проветривание (система межклетников)
31.
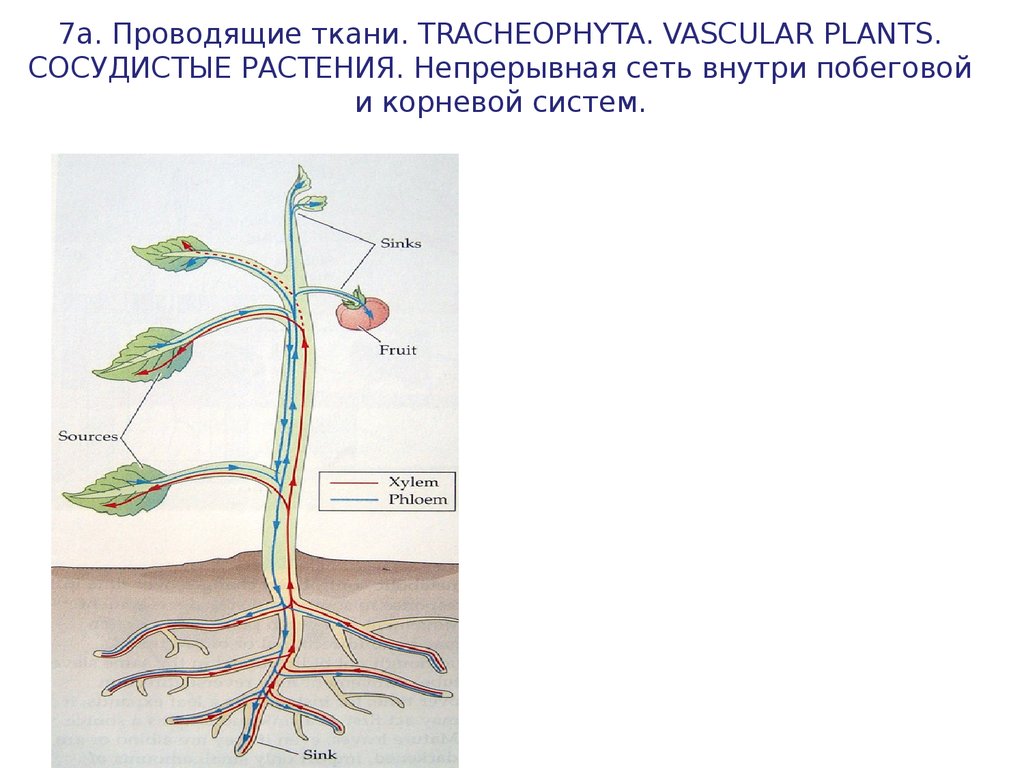
7а. Проводящие ткани. TRACHEOPHYTA. VASCULAR PLANTS.СОСУДИСТЫЕ РАСТЕНИЯ. Непрерывная сеть внутри побеговой
и корневой систем.
32.
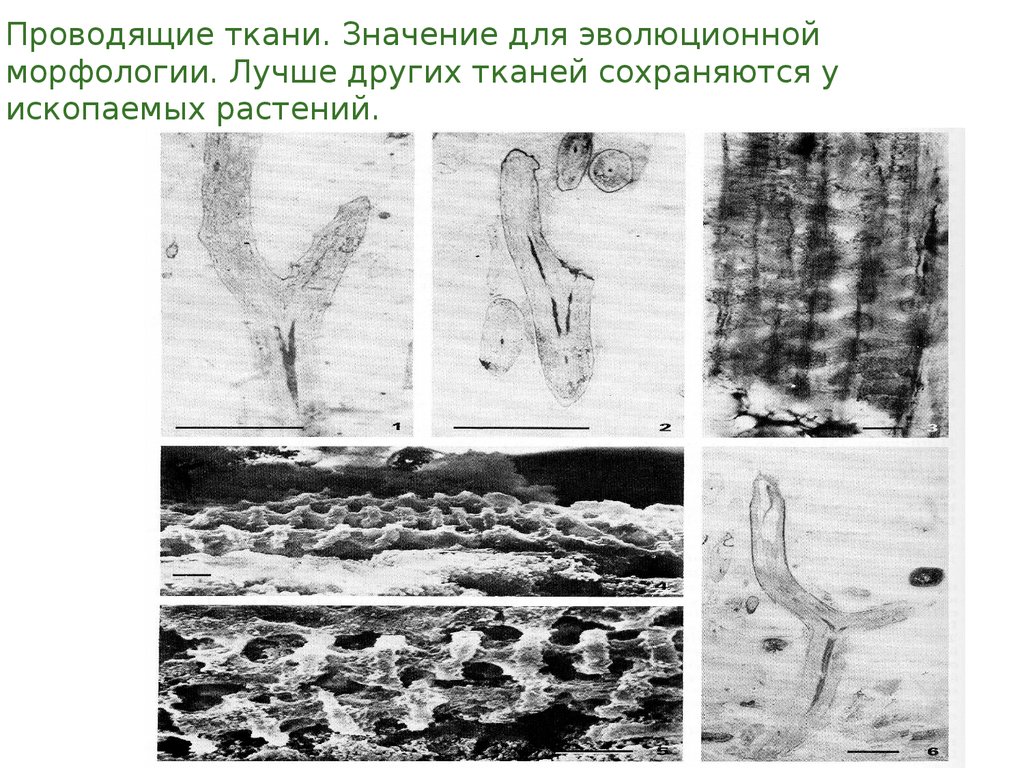
Проводящие ткани. Значение для эволюционнойморфологии. Лучше других тканей сохраняются у
ископаемых растений.
33.
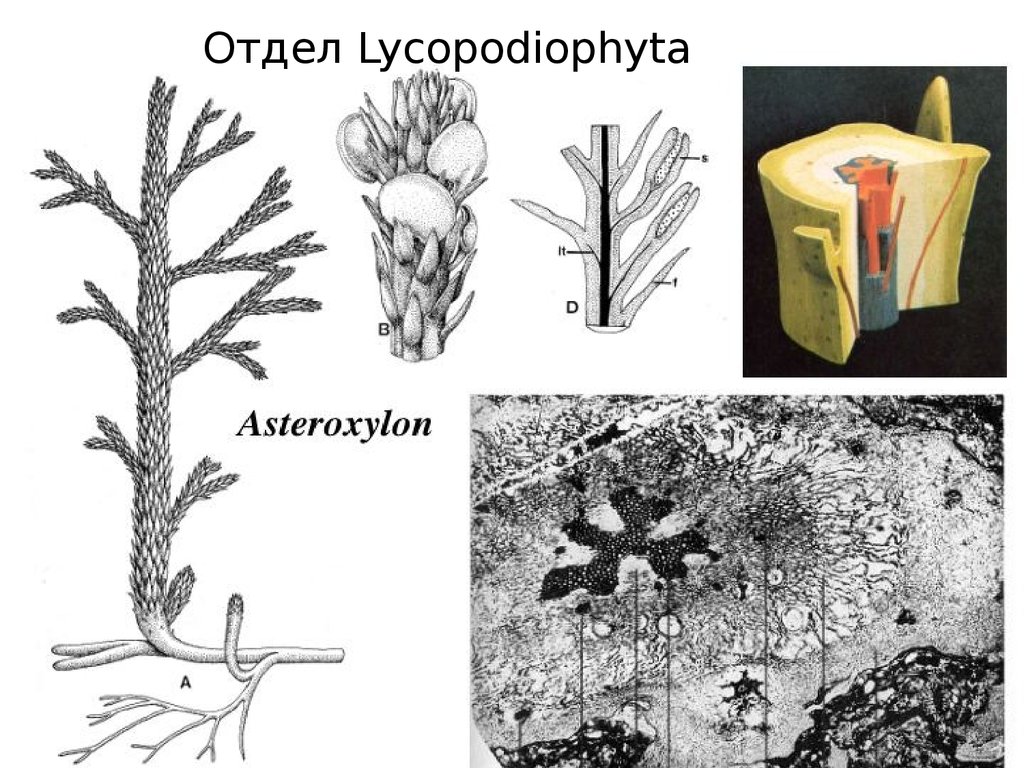
Отдел Lycopodiophyta34.
Эволюция вегетативныхорганов высших растений
35.
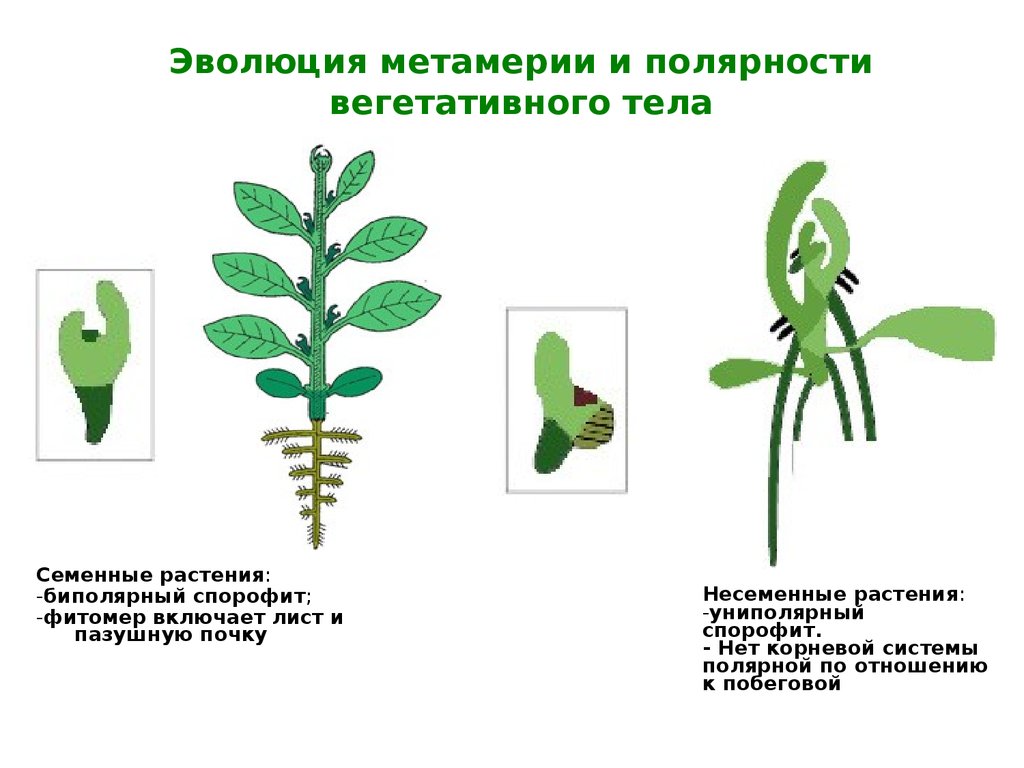
Эволюция метамерии и полярностивегетативного тела
Семенные растения:
-биполярный спорофит;
-фитомер включает лист и
пазушную почку
Несеменные растения:
-униполярный
спорофит.
- Нет корневой системы
полярной по отношению
к побеговой
36.
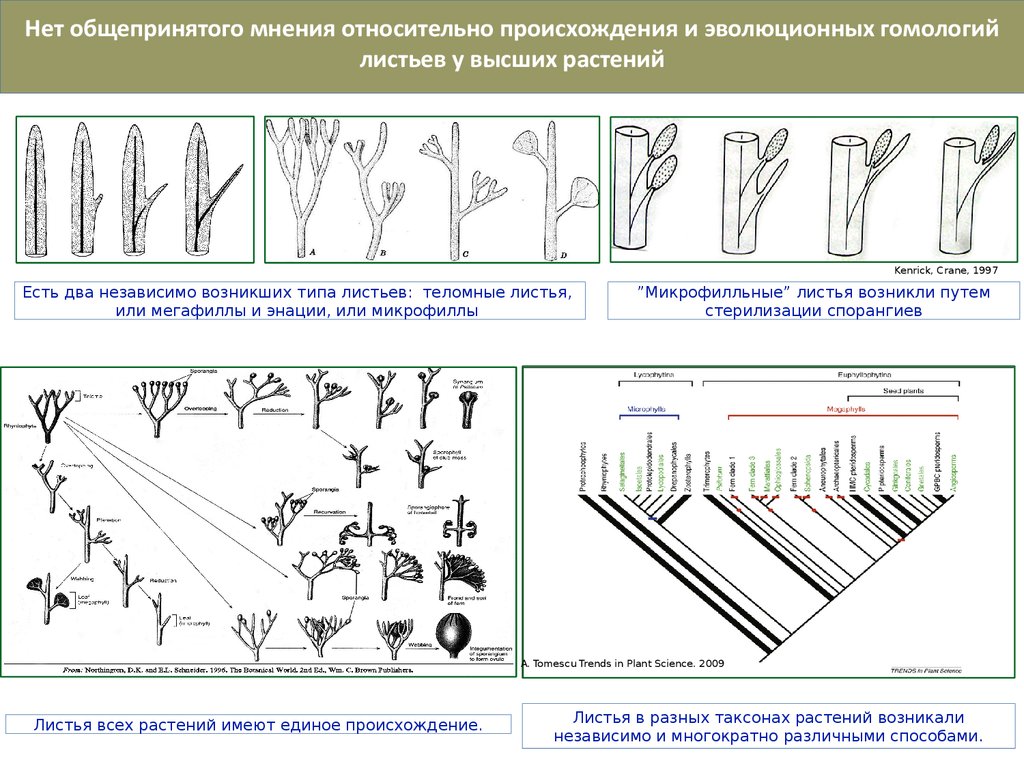
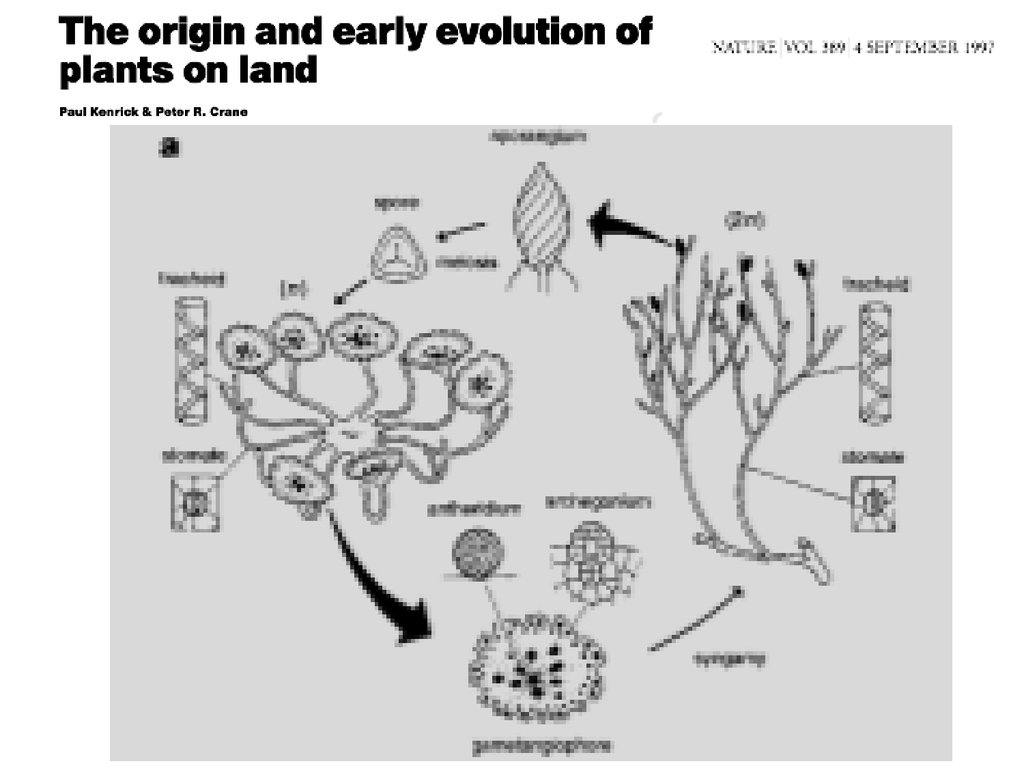
Нет общепринятого мнения относительно происхождения и эволюционных гомологийлистьев у высших растений
Kenrick, Crane, 1997
Есть два независимо возникших типа листьев: теломные листья,
или мегафиллы и энации, или микрофиллы
”Микрофилльные” листья возникли путем
стерилизации спорангиев
A. Tomescu Trends in Plant Science. 2009
Листья всех растений имеют единое происхождение.
Листья в разных таксонах растений возникали
независимо и многократно различными способами.
37.
Способность к ветвлению - общая черта спорофитоввсех высших растений спорофитной линии эволюции.
Роль - вегетативное разрастание, увеличение числа
листьев, то есть интенсивности фотосинтеза,
увеличение числа спорангиев. POLYSPORANGIOPHYTA.
Внешний облик растения определяется
ветвлением и нарастанием. Способ
образования почек = тип ветвления.
Последующий рост новых побегов из
почек - нарастание.
38.
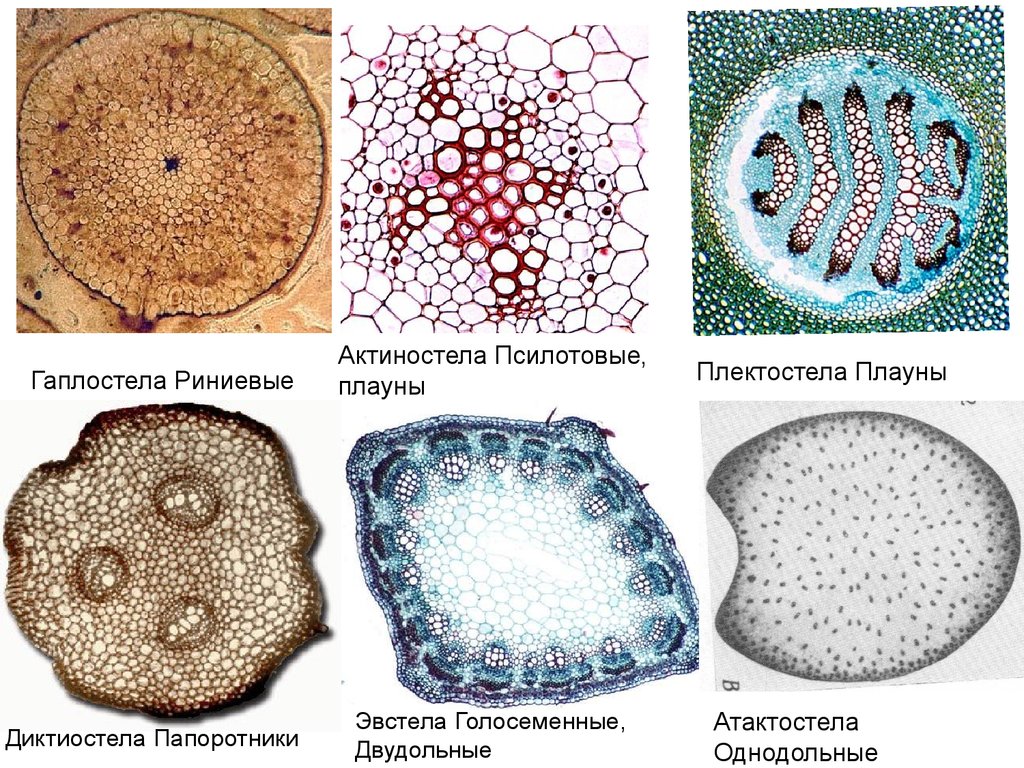
Понятие о стеле.Тип стелы характеризует таксоны на уровне отдела
или класса. Зависит от строения апикальной
меристемы и морфологической природы листа, то
есть отражает общий уровень организации побега.
Сохраняется у ископаемых растений.
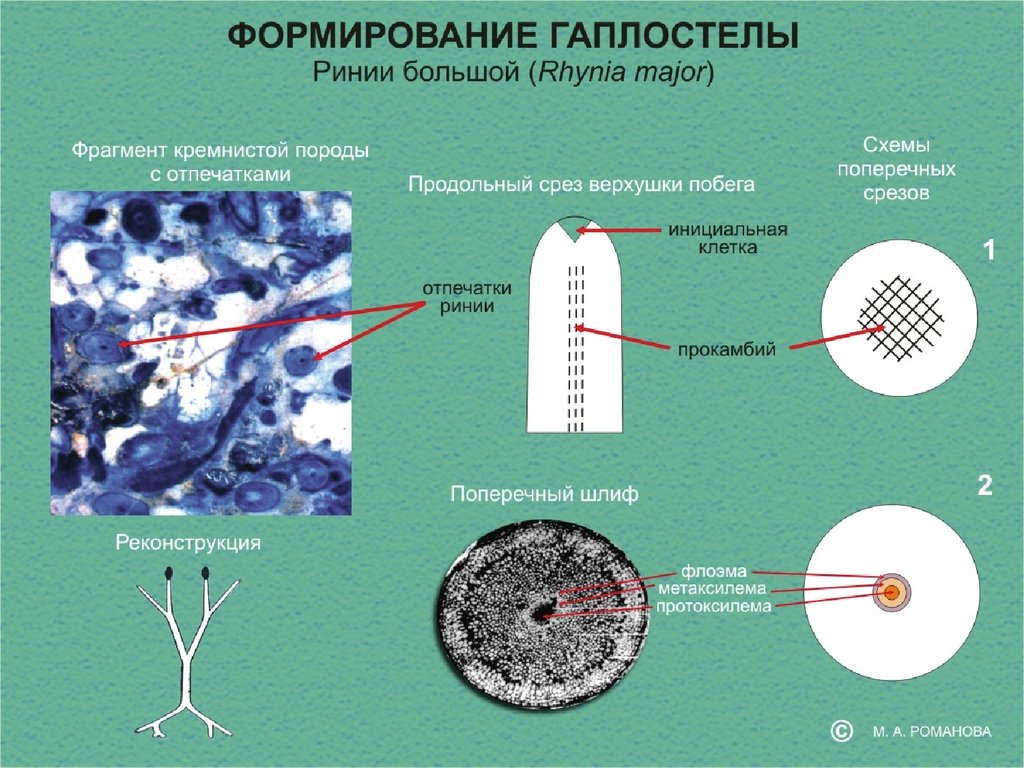
39.
Гаплостела РиниевыеДиктиостела Папоротники
Актиностела Псилотовые,
плауны
Эвстела Голосеменные,
Двудольные
Плектостела Плауны
Атактостела
Однодольные
40.
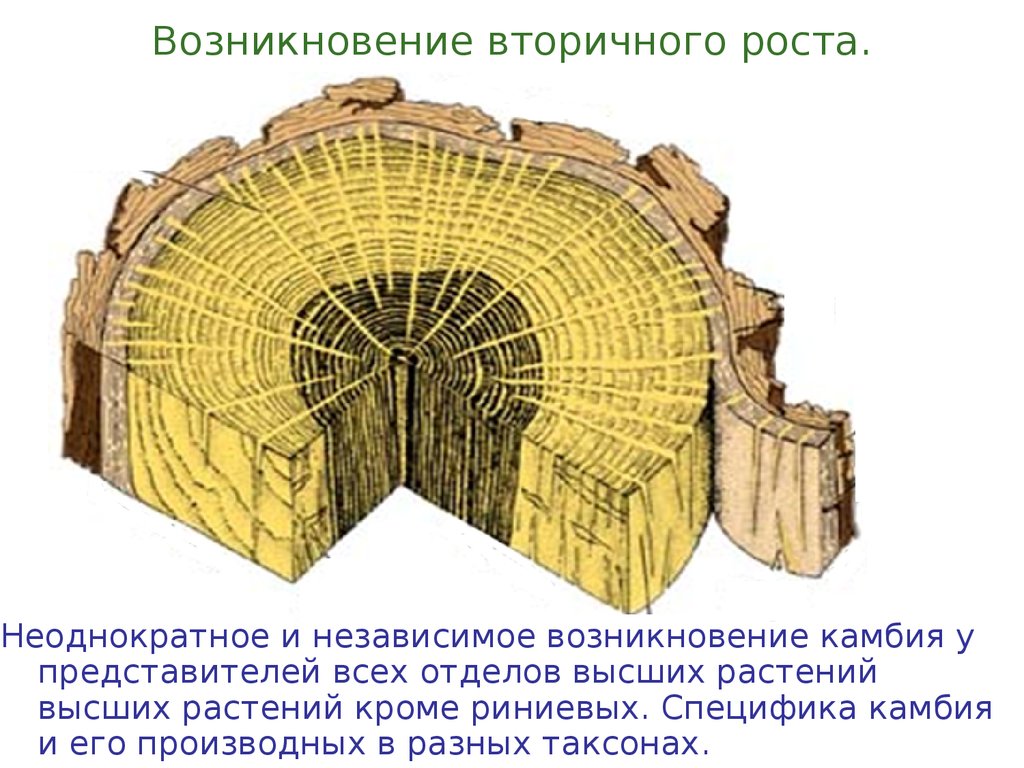
Возникновение вторичного роста.Неоднократное и независимое возникновение камбия у
представителей всех отделов высших растений
высших растений кроме риниевых. Специфика камбия
и его производных в разных таксонах.
41.
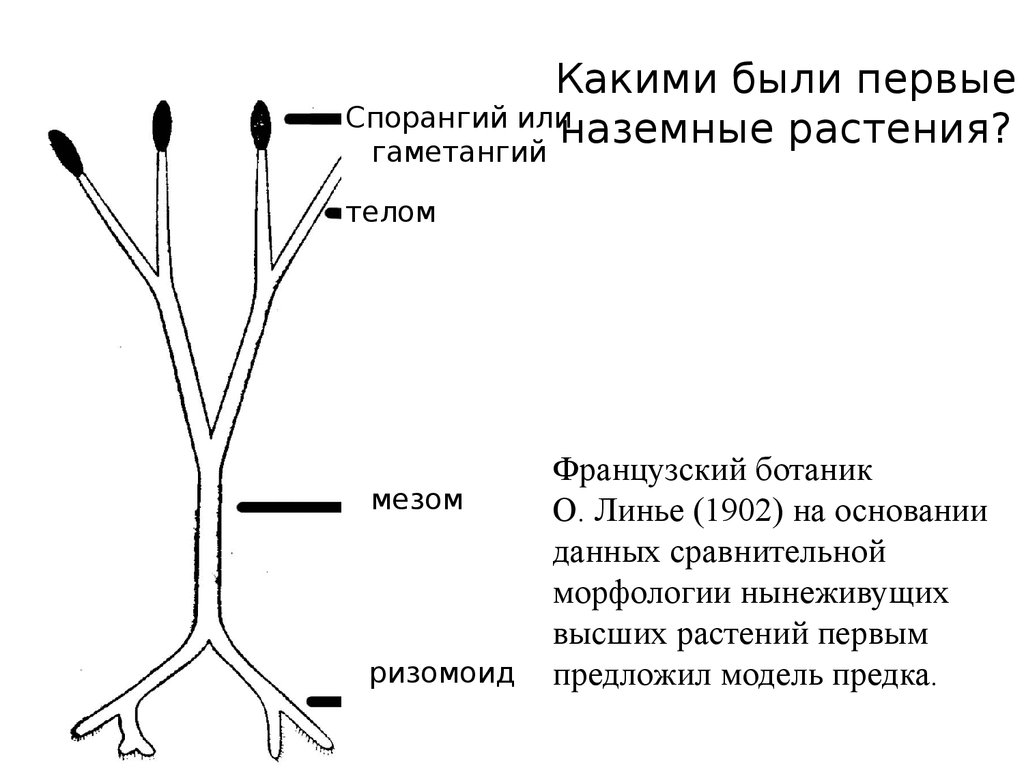
Какими были первыеСпорангий или
наземные растения?
гаметангий
телом
мезом
ризомоид
Французский ботаник
О. Линье (1902) на основании
данных сравнительной
морфологии нынеживущих
высших растений первым
предложил модель предка.
42.
Какая фаза доминировала в жизненномцикле первых наземных растений?
43.
Гаметофит Anthoceros sp. (мохообразные). Спорофит AglaophytonДостоверные остатки 425-375 млн лет.
major (Rhyniophyta) 450Споры и новейшие открытия 475 млн лет.
415 млн лет назад
44.
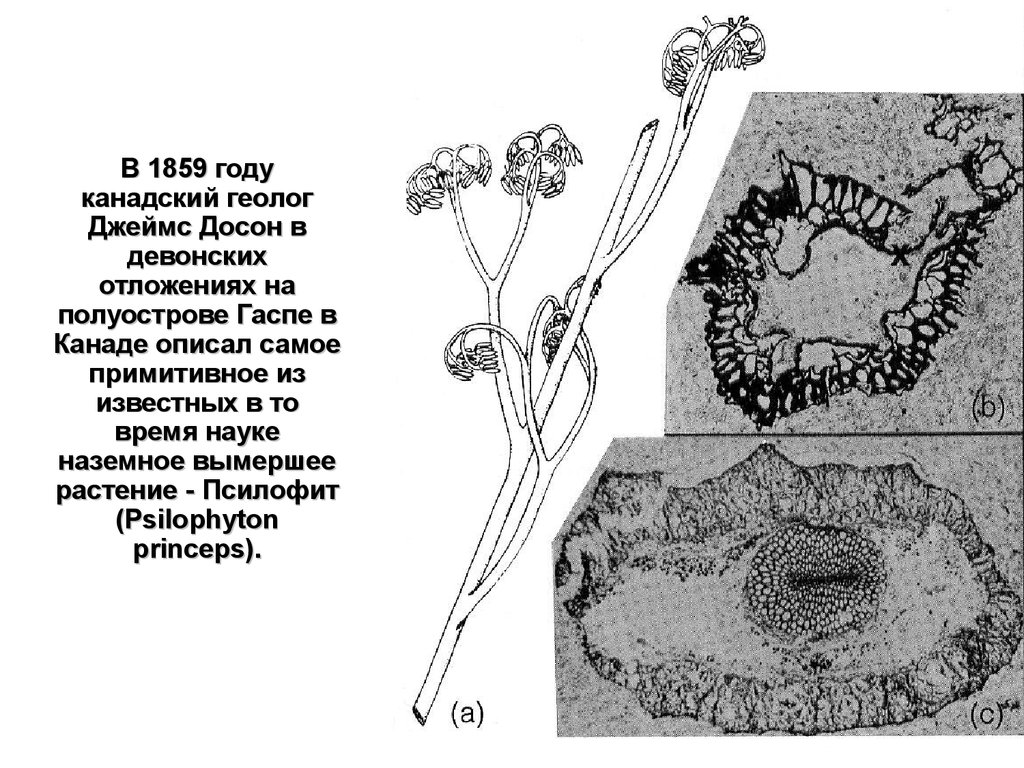
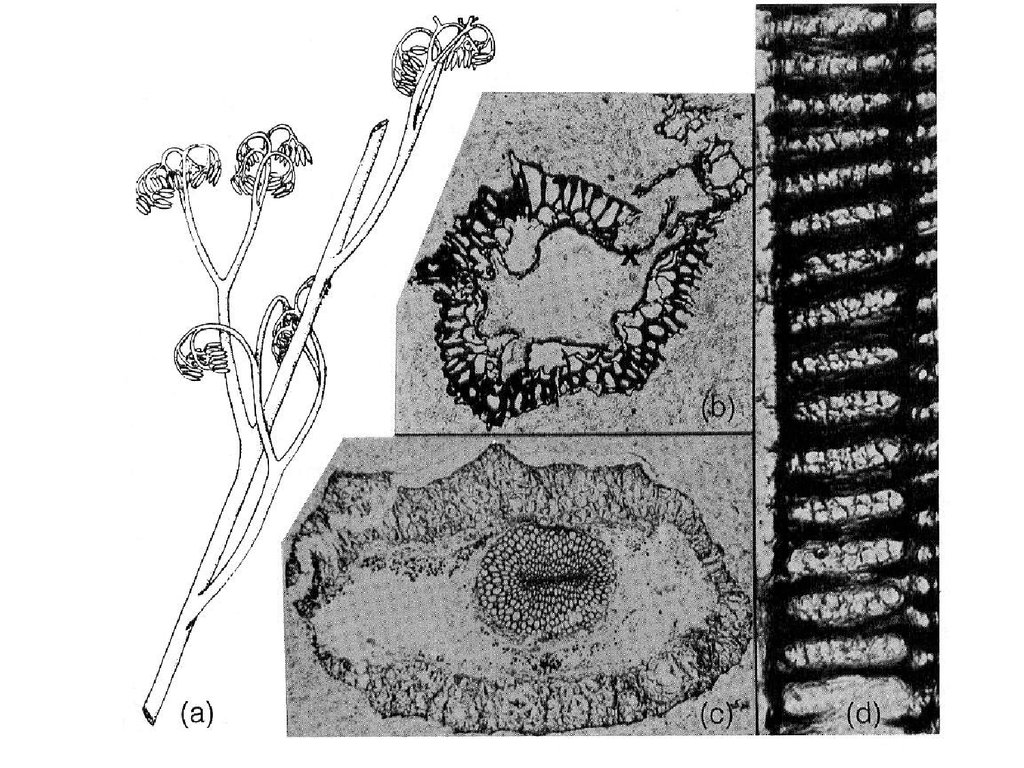
В 1859 годуканадский геолог
Джеймс Досон в
девонских
отложениях на
полуострове Гаспе в
Канаде описал самое
примитивное из
известных в то
время науке
наземное вымершее
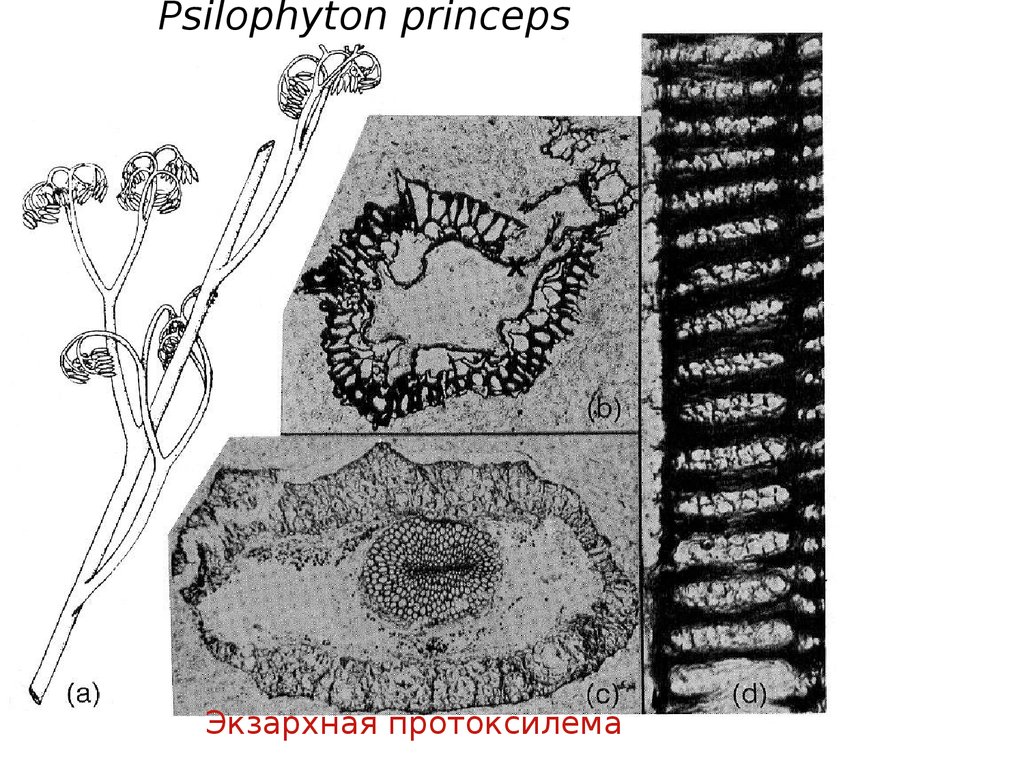
растение - Псилофит
(Psilophyton
princeps).
45.
1917 г46.
47.
RHYNIOPHYTA48.
• Роберт Кидстон и Ланг с 1917 по1921 опубликовали пять работ,получивших мировую известность. Возраст черта - раннедевонский 415 млн. лет назад.
• Ими были описаны: Psilophyton, Rhynia и Horneophyton
49.
ОТДЕЛRhyniophyta
Возникновение в Силуре (430-415
млн лет назад). Вымерли в девоне
(400-380 млн лет назад).
50.
51.
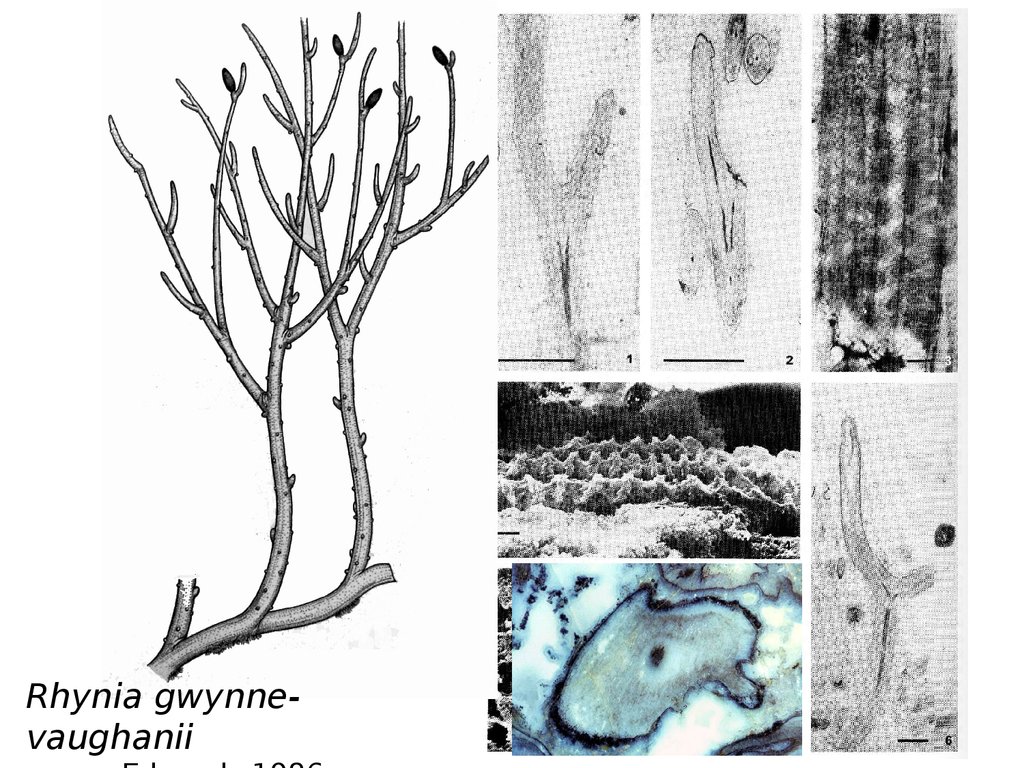
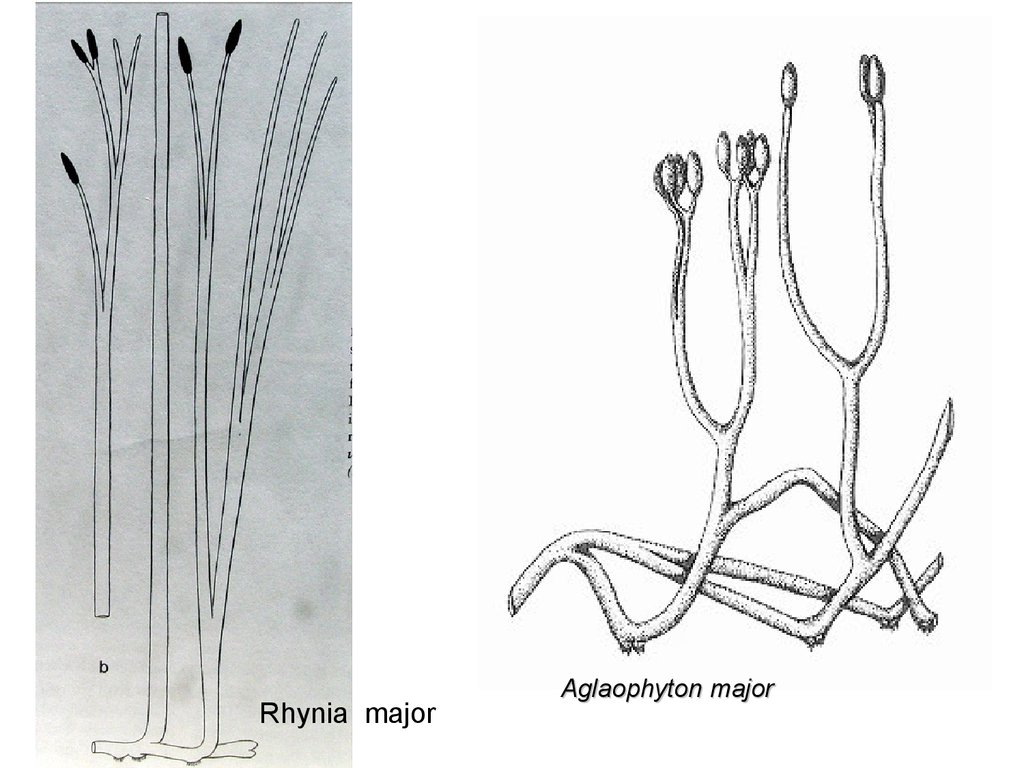
Rhynia gwin-vaughaniiRhynia gwynnevaughanii
52.
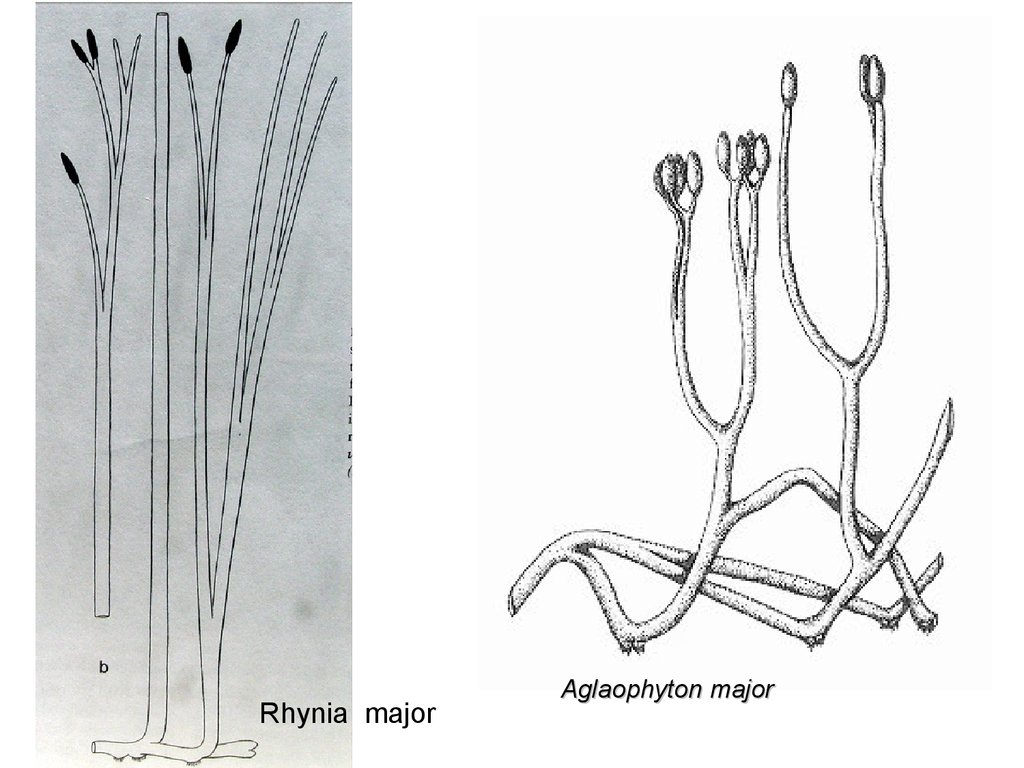
Rhynia majorAglaophyton major
53.
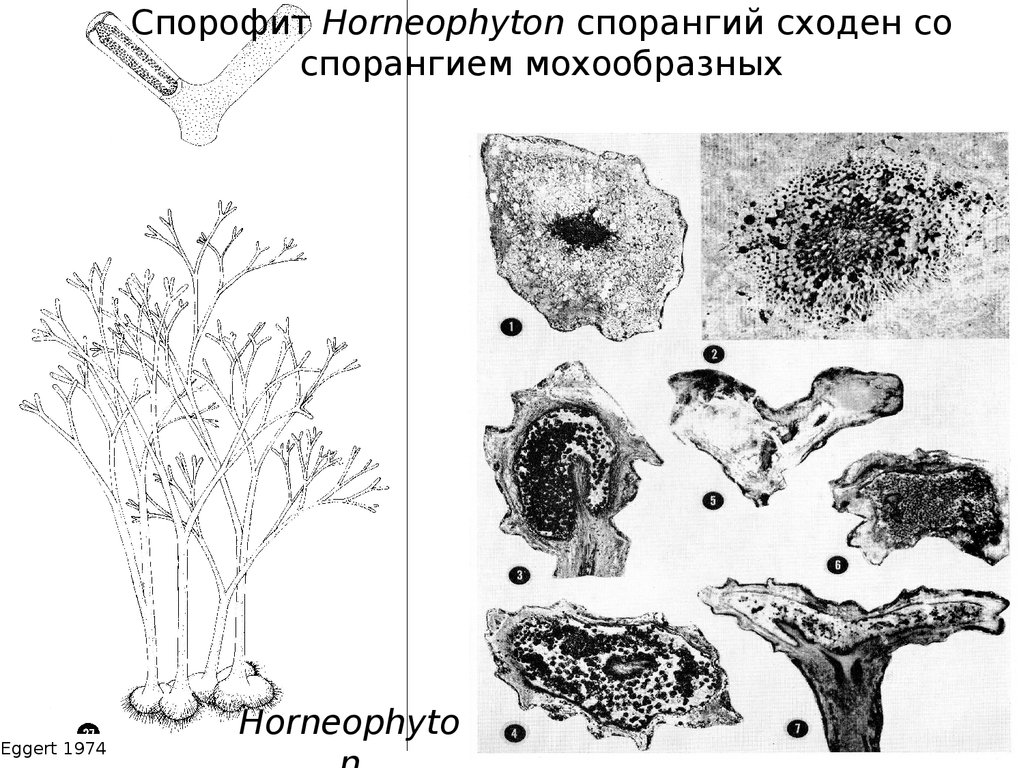
Спорофит Horneophyton спорангий сходен соспорангием мохообразных
Eggert 1974
Horneophyto
54.
Aglaophyton majorСПОРАНГИЙ
55.
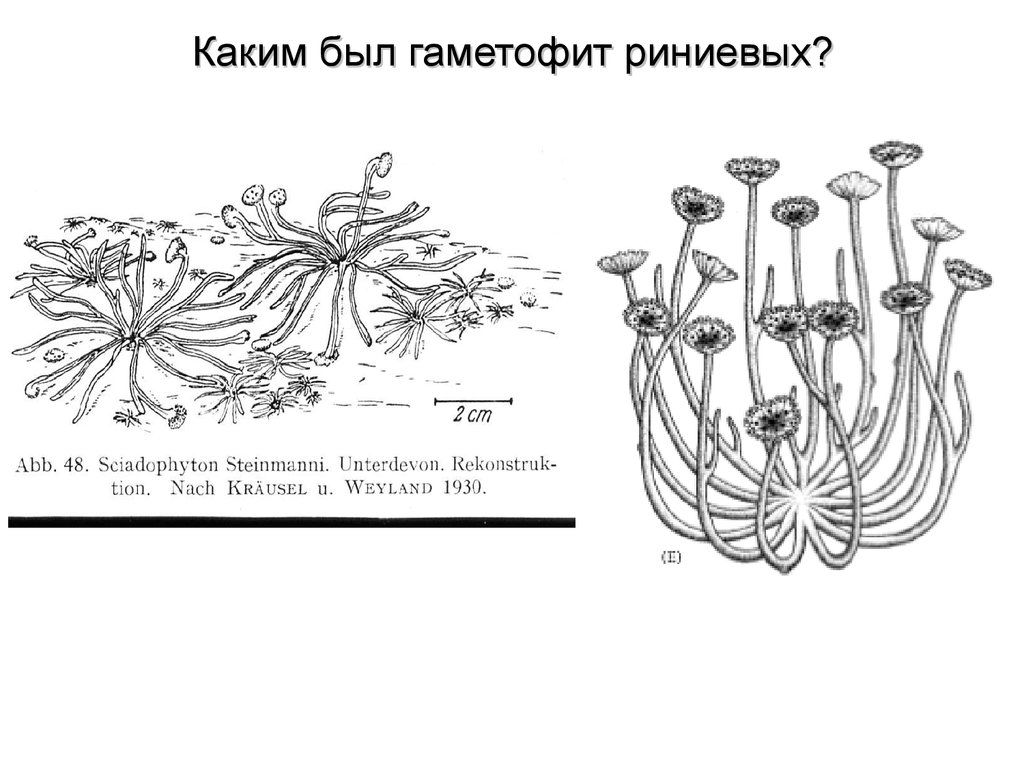
Каким был гаметофит риниевых?56.
57.
Развитие и строение гаметофитаLyonophyton
(D) Multicellular gametophyte (arrow) protruding from broken spore coat.
(E) Polar view of multicellular gametophyte showing arrangement of cells just beneath
apical cell.
(F) Gametophyte showing apical cell (upper arrow) and spore coat remnants (lower
arrow).
(G) Stoma on lower surface of gametangiophore.
(H) Cortical cells with endomycorrhizal hyphae in intercellular spaces (arrows).
(I) Cortical cells of gametangiophore containing arbuscules.
(J) Longitudinal section of several conducting elements.
(K) Longitudinal section of distal end of antheridiophore with two antheridia (arrows).
(L) Longitudinal section of antheridium. (Scale bar, 100 μm.) (M) Section of antheridium
showing sperm packets. (Scale bar, 5 μm.) (N) Longitudinal section of archegonium
showing neck canal (upper arrow) and probable egg chamber (lower arrow). (Scale bar, 40
μm.) (O) Dense cluster of gametophytes showing size of antheridiophores and
archegoniophores. (Scale bar, 6.5 mm.) (P) Spore aggregate. (Scale bar, 1.0 mm.) (Q)
Bulbil (arrow) on vegetative axis. (Scale bar, 1.0 mm.)
Thomas N. Taylor et al. PNAS 2005;102:5892-5897
©2005 by National Academy of Sciences
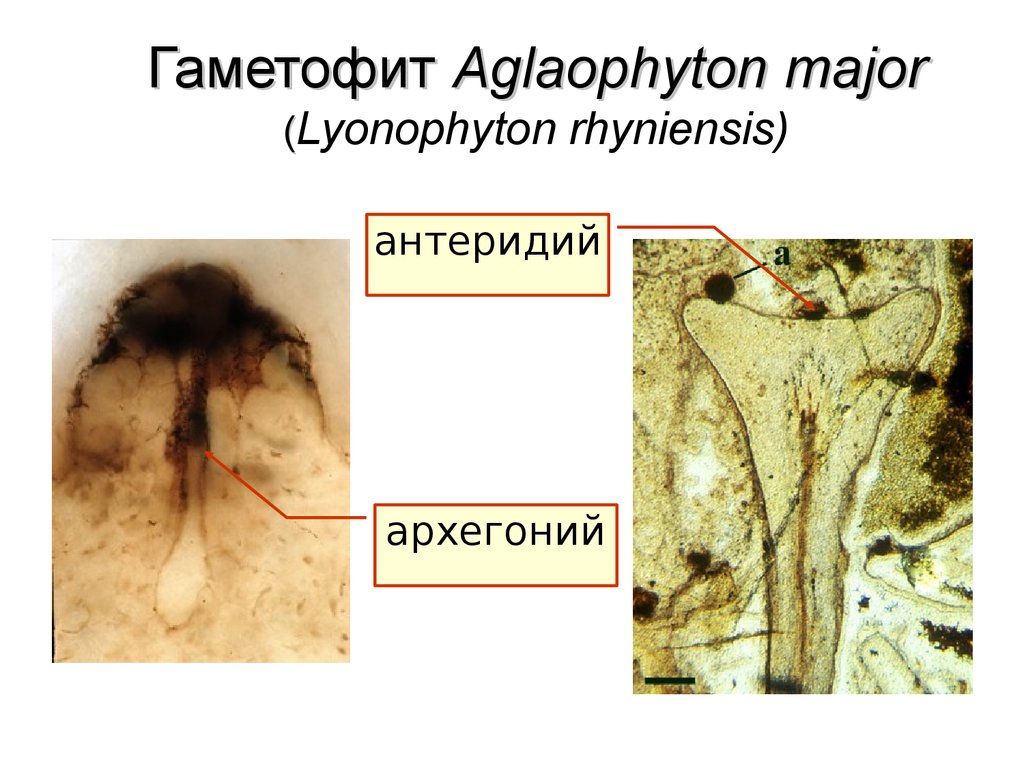
58.
Гаметофит Aglaophyton major(Lyonophyton rhyniensis)
антеридий
архегоний
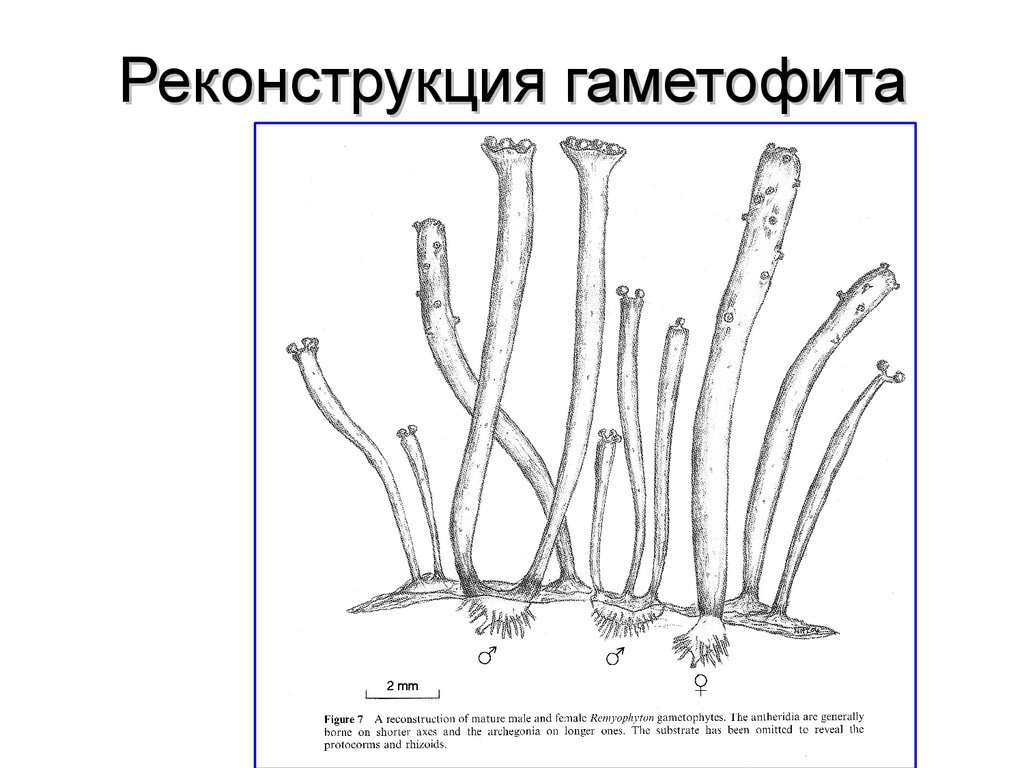
59.
Реконструкция гаметофита60.
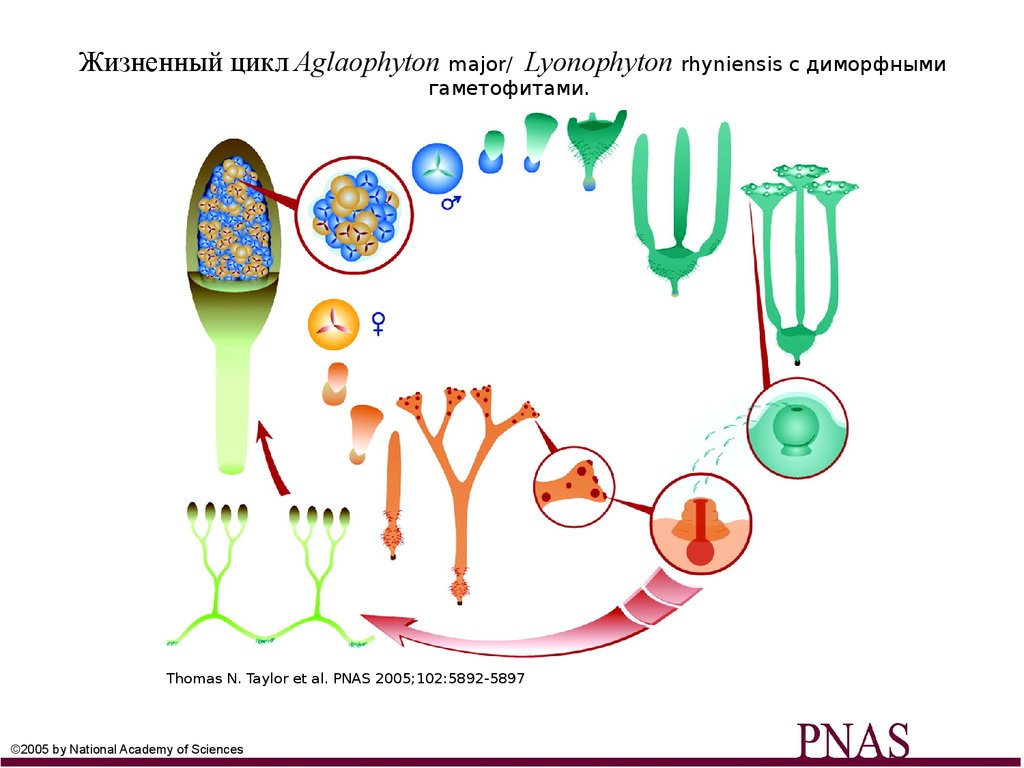
Жизненный цикл Aglaophyton major/ Lyonophyton rhyniensis с диморфнымигаметофитами.
Thomas N. Taylor et al. PNAS 2005;102:5892-5897
©2005 by National Academy of Sciences
61.
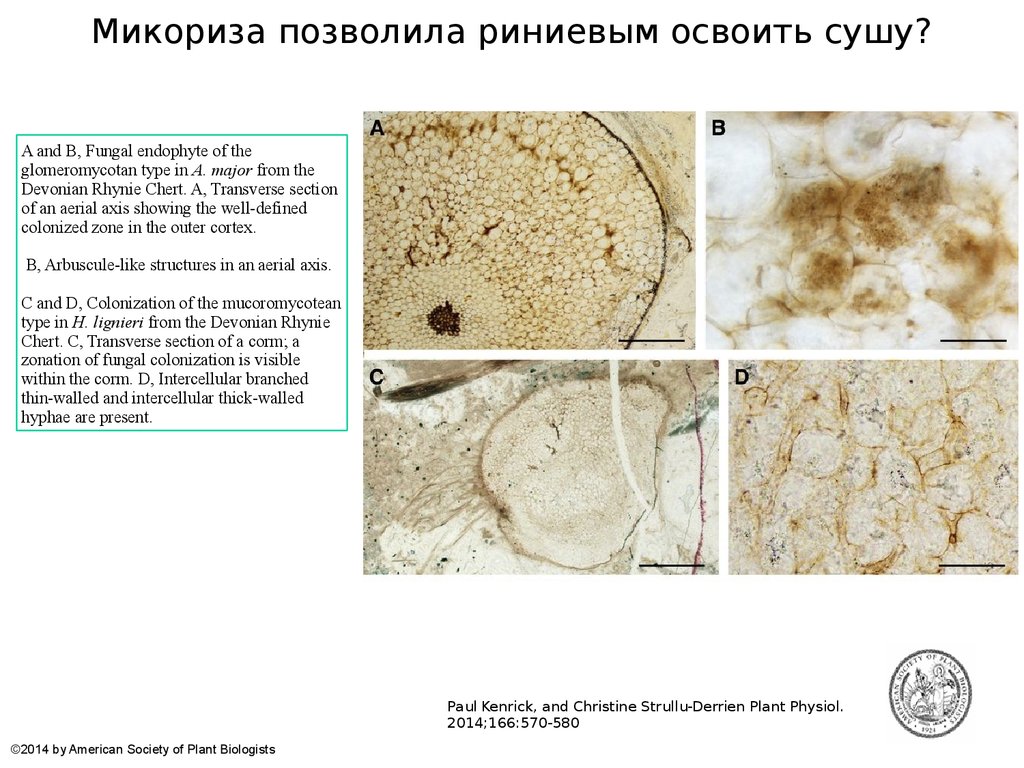
Микориза позволила риниевым освоить сушу?A and B, Fungal endophyte of the
glomeromycotan type in A. major from the
Devonian Rhynie Chert. A, Transverse section
of an aerial axis showing the well-defined
colonized zone in the outer cortex.
B, Arbuscule-like structures in an aerial axis.
C and D, Colonization of the mucoromycotean
type in H. lignieri from the Devonian Rhynie
Chert. C, Transverse section of a corm; a
zonation of fungal colonization is visible
within the corm. D, Intercellular branched
thin-walled and intercellular thick-walled
hyphae are present.
Paul Kenrick, and Christine Strullu-Derrien Plant Physiol.
2014;166:570-580
©2014 by American Society of Plant Biologists
62.
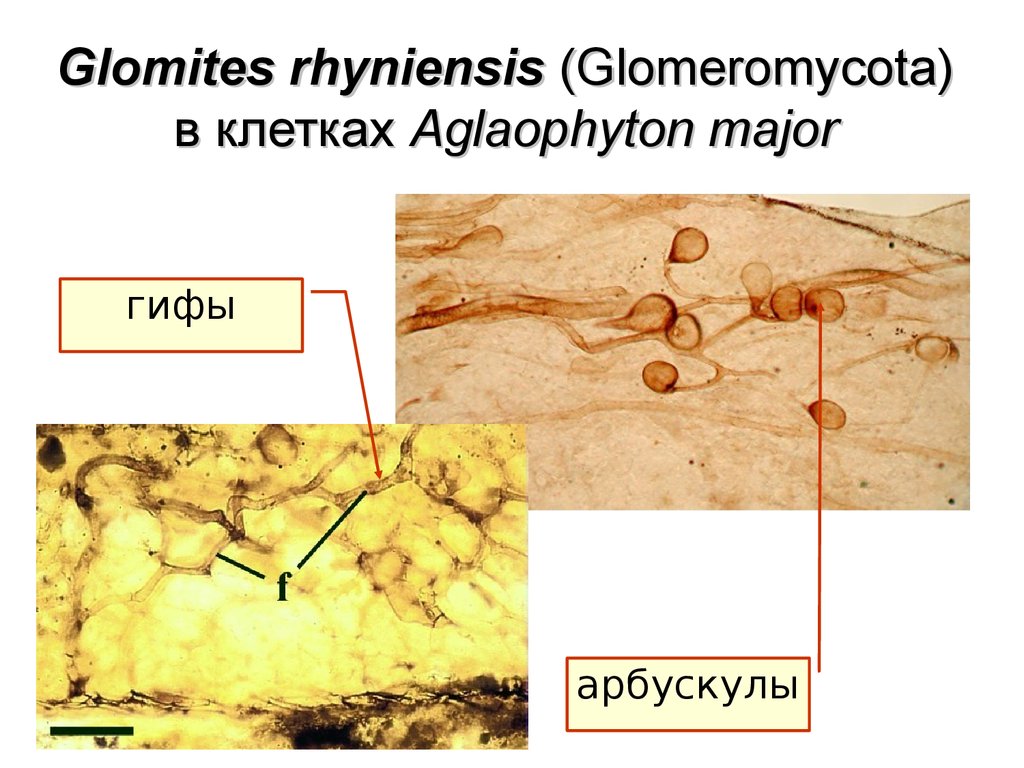
Glomites rhyniensis (Glomeromycota)в клетках Aglaophyton major
гифы
арбускулы
63.
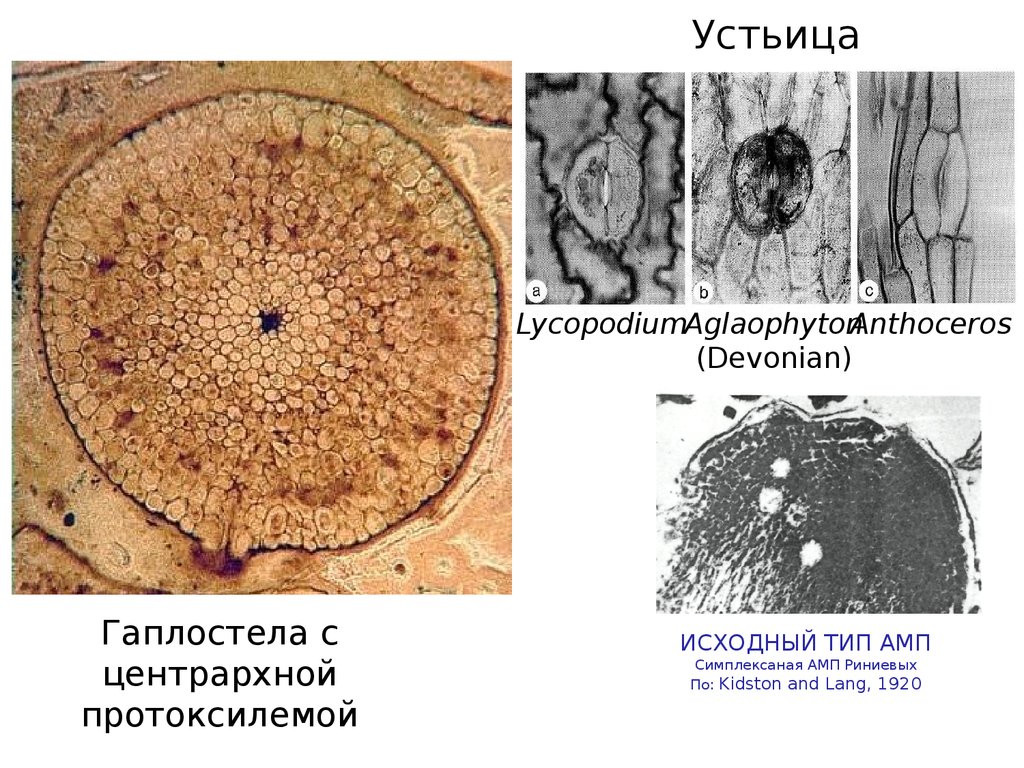
УстьицаLycopodiumAglaophytonAnthoceros
(Devonian)
Гаплостела с
центрархной
протоксилемой
ИСХОДНЫЙ ТИП АМП
Симплексаная АМП Риниевых
По: Kidston and Lang, 1920
64.
Rhynia gwynnevaughanii65.
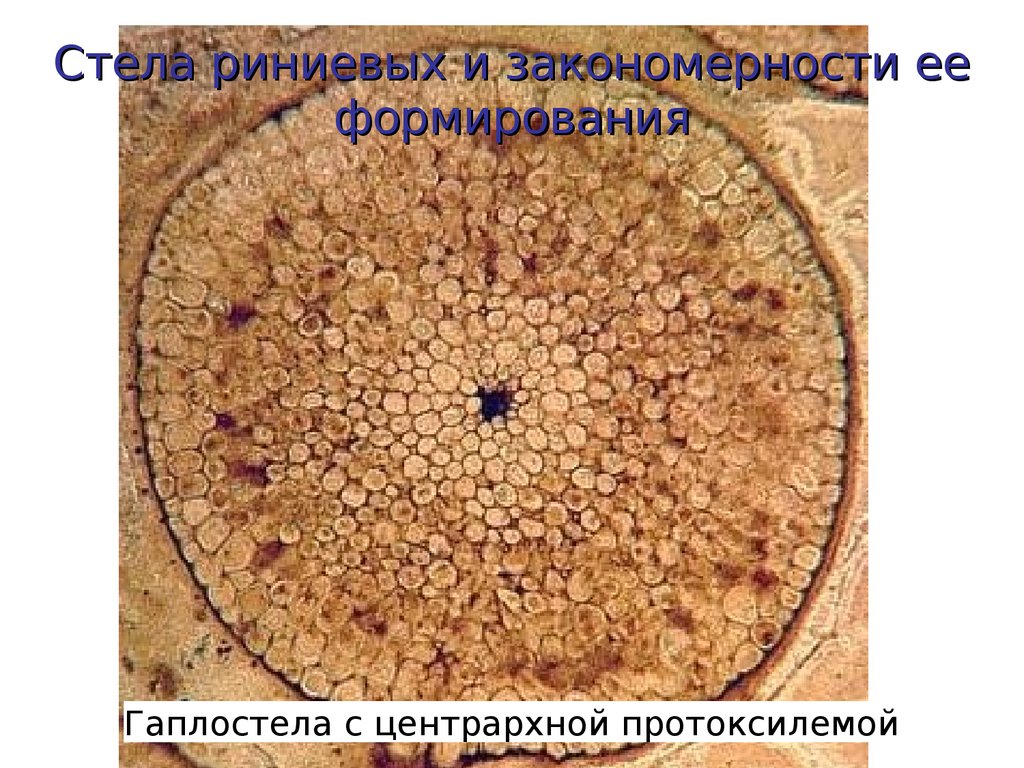
Стела риниевых и закономерности ееформирования
Гаплостела с центрархной протоксилемой
66.
67.
Осознание искусственностиотд.
Rhyniophyta
Концепция Protracheophyta
(«предсосудистых» растений).
Спорофиты с ветвящимися
теломами, но без одревесневшей
водопроводящей ткани (ксилемы)
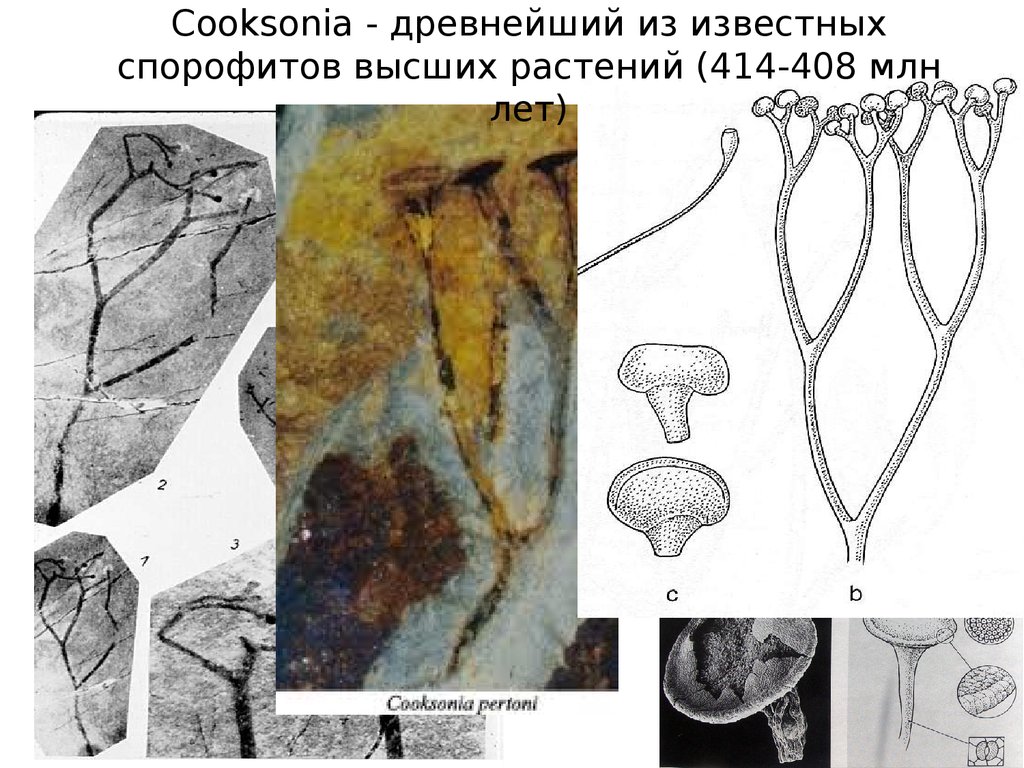
68.
Cooksonia - древнейший из известныхспорофитов высших растений (414-408 млн
лет)
69.
Rhynia majorAglaophyton major
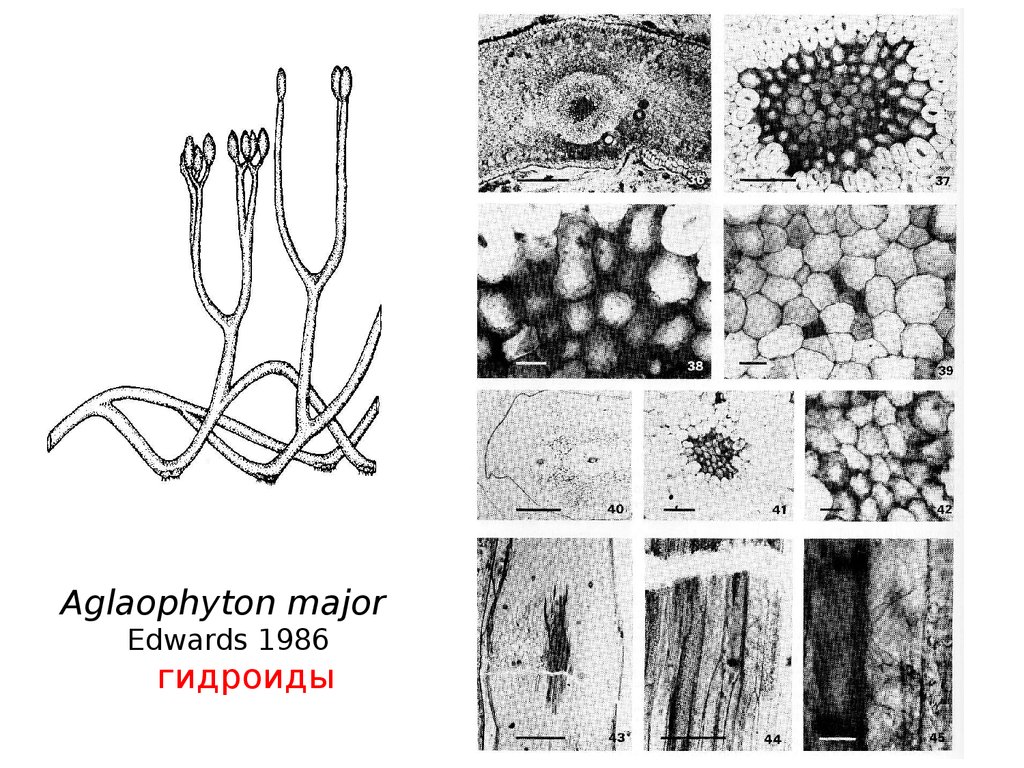
70.
Aglaophyton majorEdwards 1986
гидроиды
71.
72.
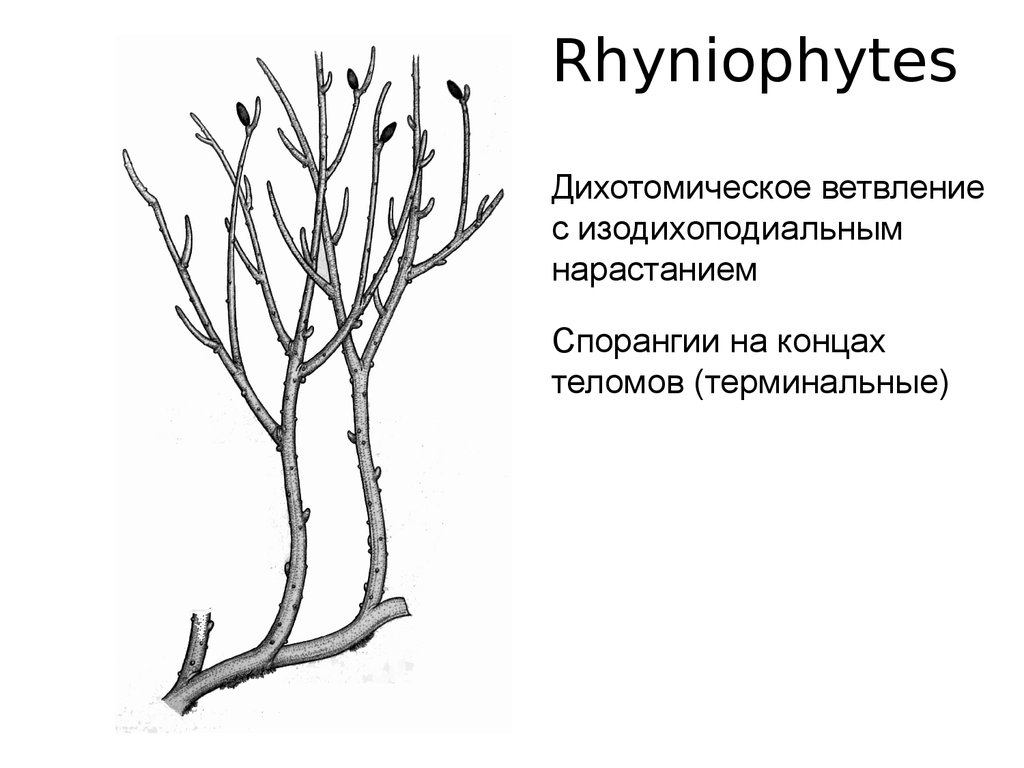
RhyniophytesДихотомическое ветвление
с изодихоподиальным
нарастанием
Спорангии на концах
теломов (терминальные)
73.
SteganothecaUskiella
74.
Celatheca beckii75.
76.
77.
TrimerophytesАнизодихоподиальное
нарастание при
дихотомическом
ветвлении
Хорошо выражена
главная ось
Psilophyton
78.
Psilophyton princeps79.
Psilophyton princepsЭкзархная протоксилема
80.
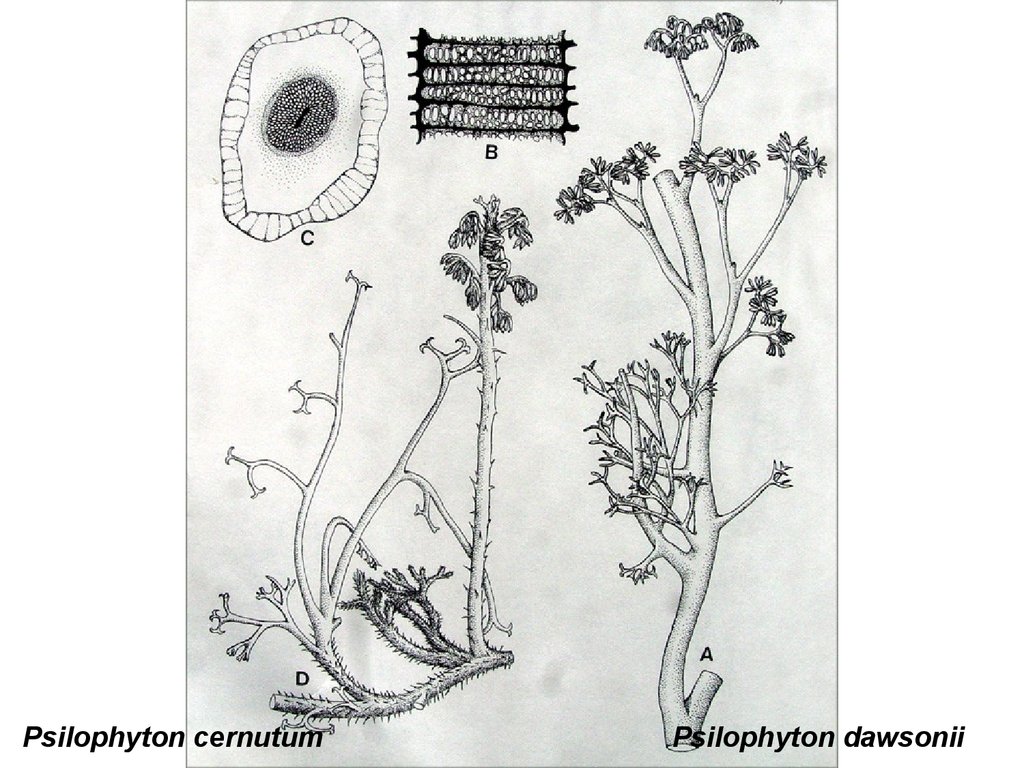
Psilophyton cernutumPsilophyton dawsonii
81.
Hsua robusta82.
83.
Pertica84.
85.
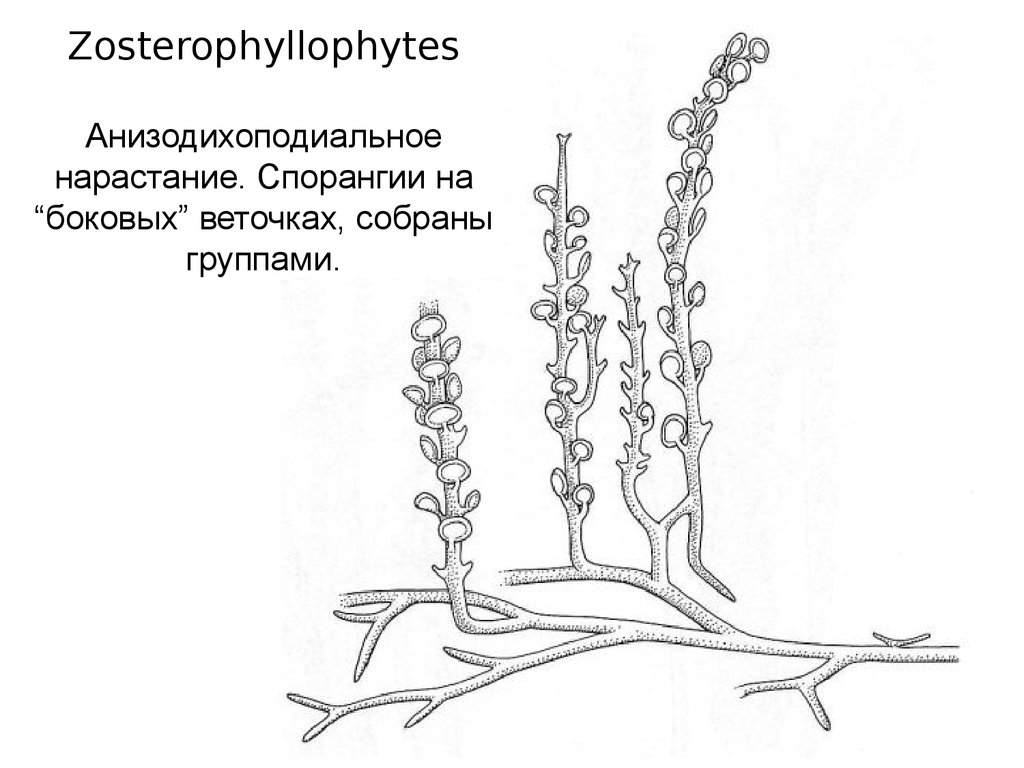
ZosterophyllophytesАнизодихоподиальное
нарастание. Спорангии на
“боковых” веточках, собраны
группами.
86.
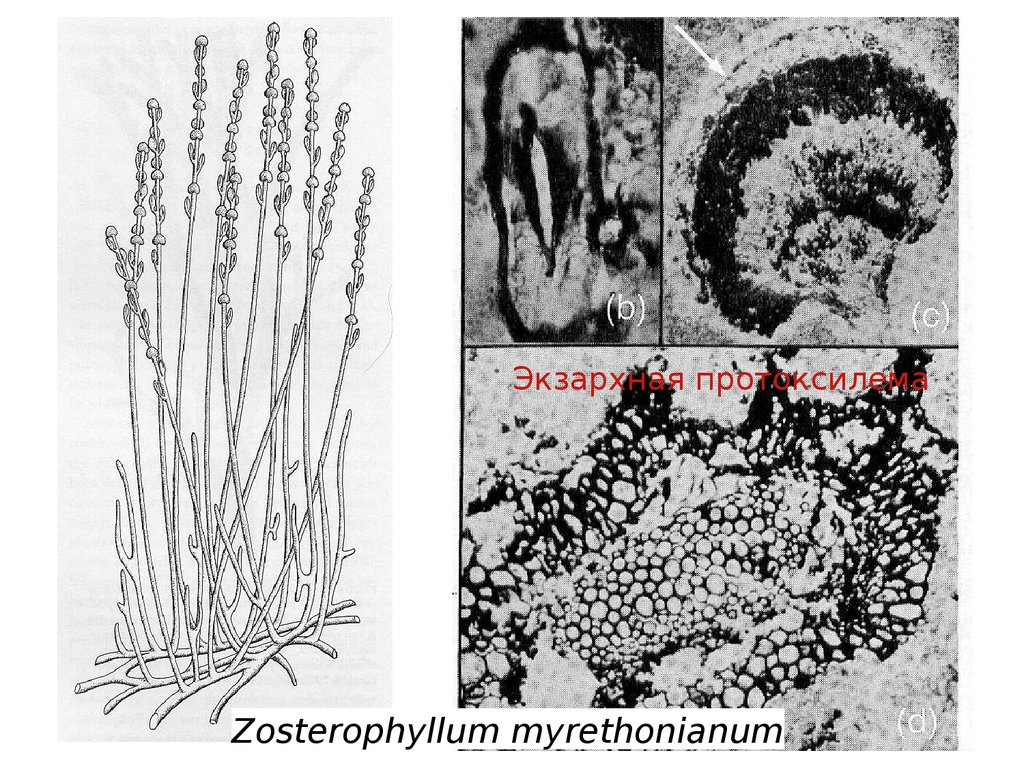
Экзархная протоксилемаZosterophyllum myrethonianum
87.
88.
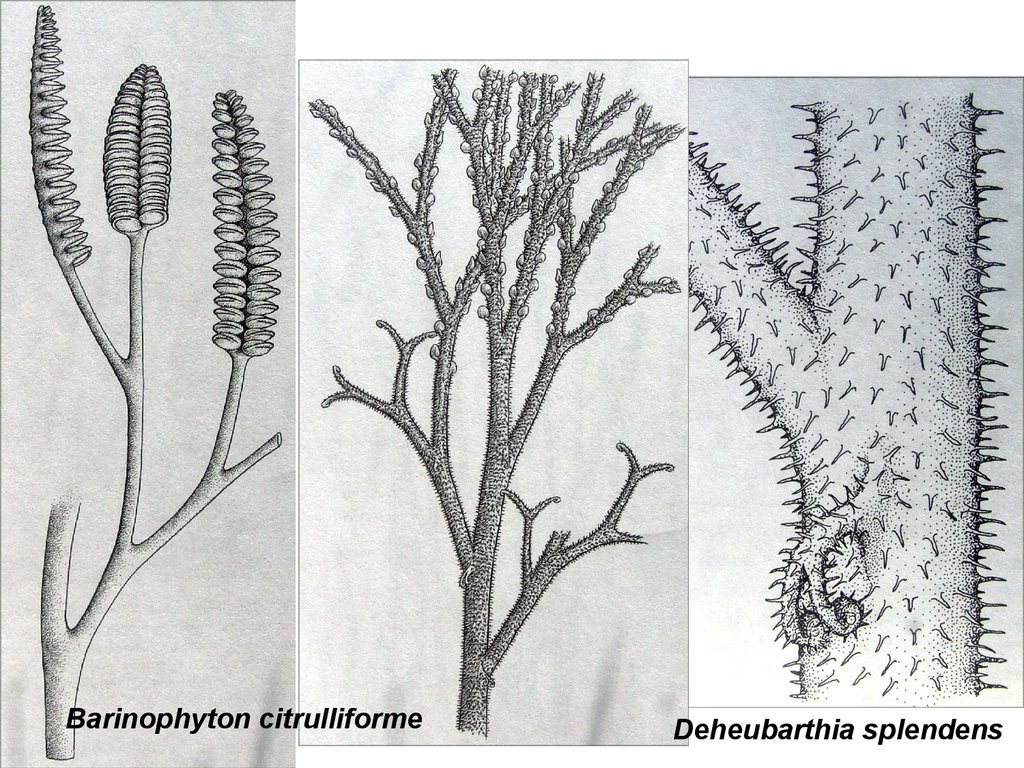
Barinophyton citrulliformeDeheubarthia splendens
89.
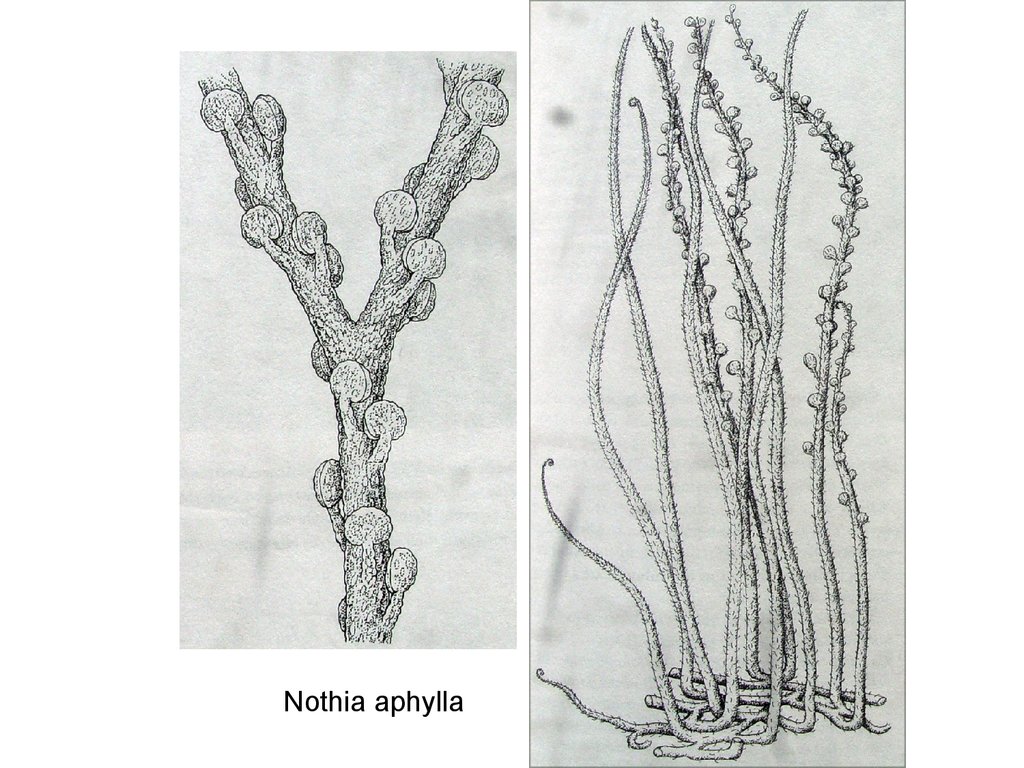
Nothia aphylla90.
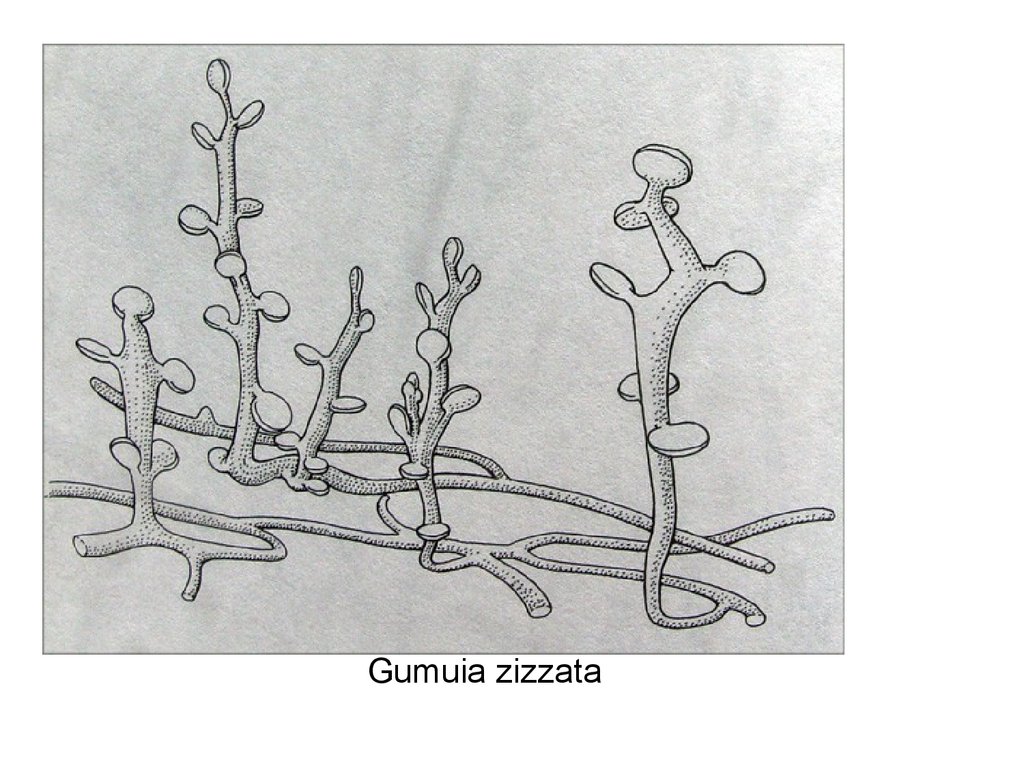
Gumuia zizzata91.
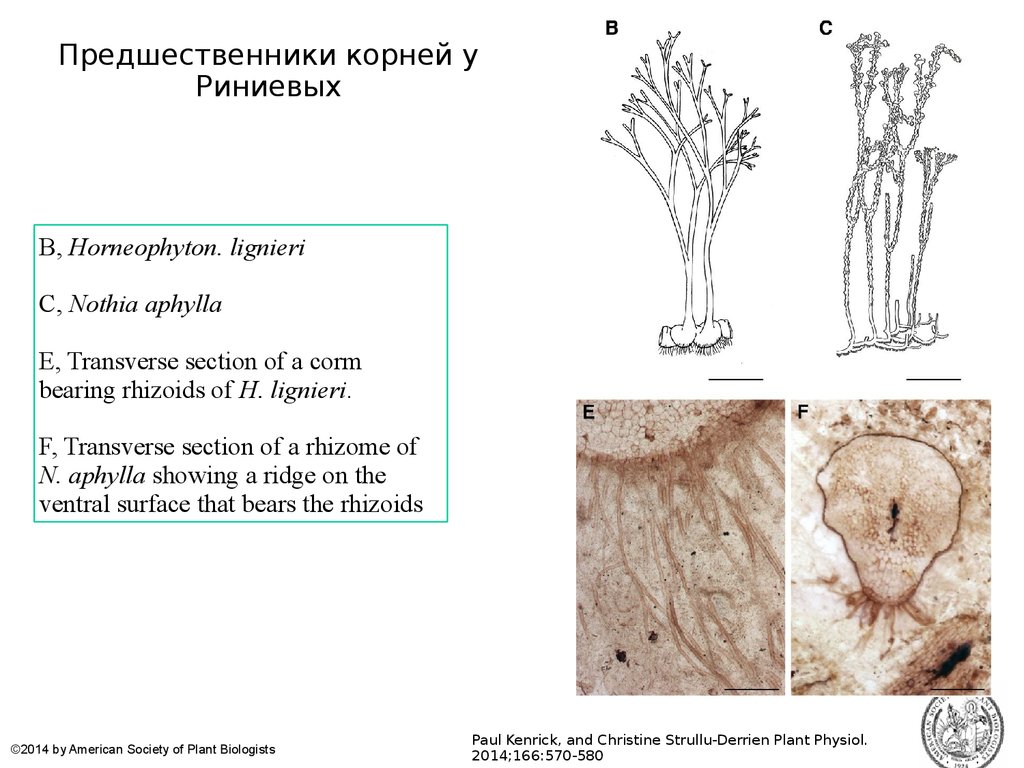
Предшественники корней уРиниевых
B, Horneophyton. lignieri
C, Nothia aphylla
E, Transverse section of a corm
bearing rhizoids of H. lignieri.
F, Transverse section of a rhizome of
N. aphylla showing a ridge on the
ventral surface that bears the rhizoids
©2014 by American Society of Plant Biologists
Paul Kenrick, and Christine Strullu-Derrien Plant Physiol.
2014;166:570-580
92.
Предшественники корней у РиниевыхB, Early rooting system in Zosterophyllum shengfengense The plant shows a tufted habit of numerous aerial axes,
which are supported by a rhizome and shallow rooting system. C, Reconstruction of Z. shengfengense
©2014 by American Society of Plant Biologists
Paul Kenrick, and Christine Strullu-Derrien Plant Physiol.
2014;166:570-580
93.
94.
95.
96.
97.
98.
Ландшафт среднего раннего Девона. Раннедевонские ( 408 - 387 млн летназад) Cooksonia, Zosterophyllum, Aglaophyton. Среднедевонский
Psilophyton (387-374 млн лет назад), плауны Drepanophycus и
Protolepidodendron. На переднем плане - мхи, «вышедшие на сушу» 510-
99.
100.
ОТДЕЛ RHYNIOPHYTAПреобладание спорофита в жизненном цикле
Спорофиты ветвящиеся
Верхушечный рост
Верхушечное (= дихотомическое) ветвление
Отсутствие листьев; фотосинтез осевыми
органами (теломами)
Отсутствие корней (их функцию выполнял
ризомоид с ризоидами)
Есть все основные ткани кроме механической.
Они в равной степени развиты у спорофита и
гаметофита.
101.
Почему вымерлиRHYNIOPHYTA?
Нет листьев - мала
поверхность фотосинтеза
Нет корней - низкая
эффективность
почвенного питания