





. Вьетнам.")













 biology
biologySimilar presentations:










Практический курс морфологии архегониальных растений
1.
2.
К.И. Мейер.Практический курс
морфологии
архегониальных
растений. М, 1982
3.
4. Takakia
(Тимонин, Филин, 2009)Род был описан в 1958 году. Его относили к
печеночникам
(а иногда даже сближали с плауновидными)
до 1990, когда были найдены спорофиты
Takakia: 2 вида
(тихоокеанское побережье Сев.
Америки, Алеутские
о-ва, Борнео, Гималаи)
Takakia
5.
Wollemia nobilis –третий род семейства
Araucariaceae
Новый род, описан из
Австралии в 1994 г.
6. Ticodendron incognitum
http://www.plantsystematics.orgTicodendron
incognitum
7.
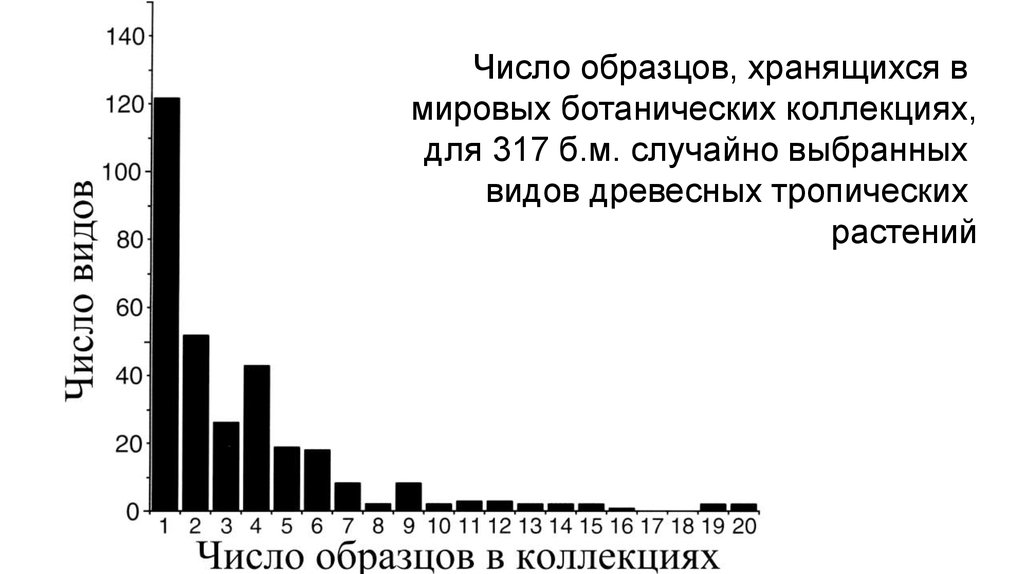
Число образцов, хранящихся вмировых ботанических коллекциях,
для 317 б.м. случайно выбранных
видов древесных тропических
растений
8.
Vaccinium (Ericaceae): около 450 видов, большое разнообразие в горных районах тропиковПервое описание и изображение цветка
Пыльники склеены в трубку
Придатки пыльников диморфные,
перекрещиваются через один
Vaccinium uliginosum - голубика
узкий ареал
Vaccinium
bullatum
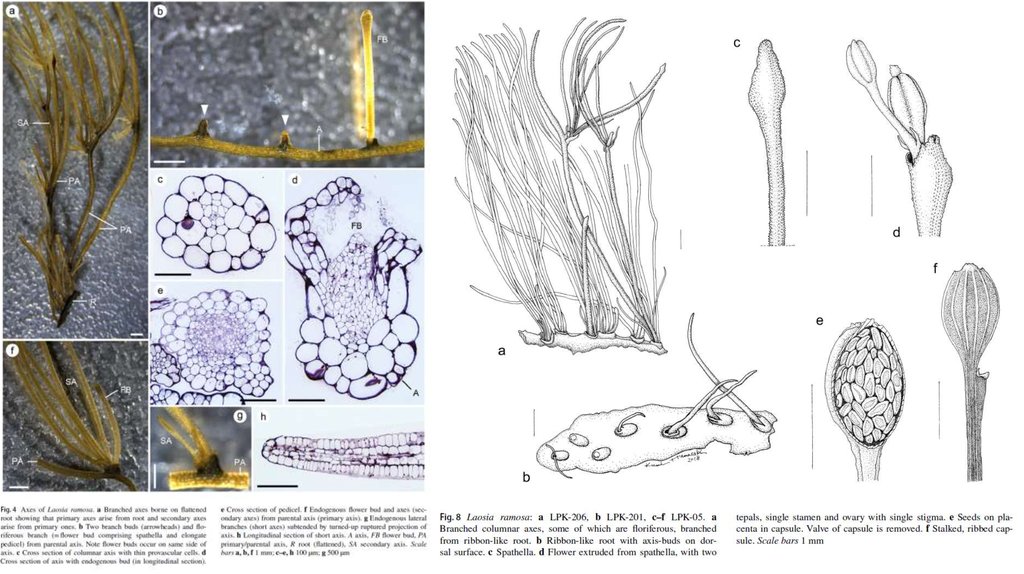
9. Shortia rotata Gaddy & Nuraliev (Diapensiaceae). Вьетнам.
Новые для науки виды, подвиды и разновидности, описанныесотрудниками и аспирантами кафедры высших растений
за 1993-2018 гг. (значки – типовые местонахождения)
сайт кафедры:
msu-botany.ru
Shortia rotata Gaddy & Nuraliev
(Diapensiaceae). Вьетнам.
Biermannia longicheila Aver. & Nuraliev
(Orchidaceae). Вьетнам.
Arthrocnemum franzii Sukhor.
(Amaranthaceae). Кабо-Верде.
Tripleurospermum nathaliae A.Zernov (Asteraceae).
Краснодарский край, Россия.
всего более 80 новых видов за 25
лет
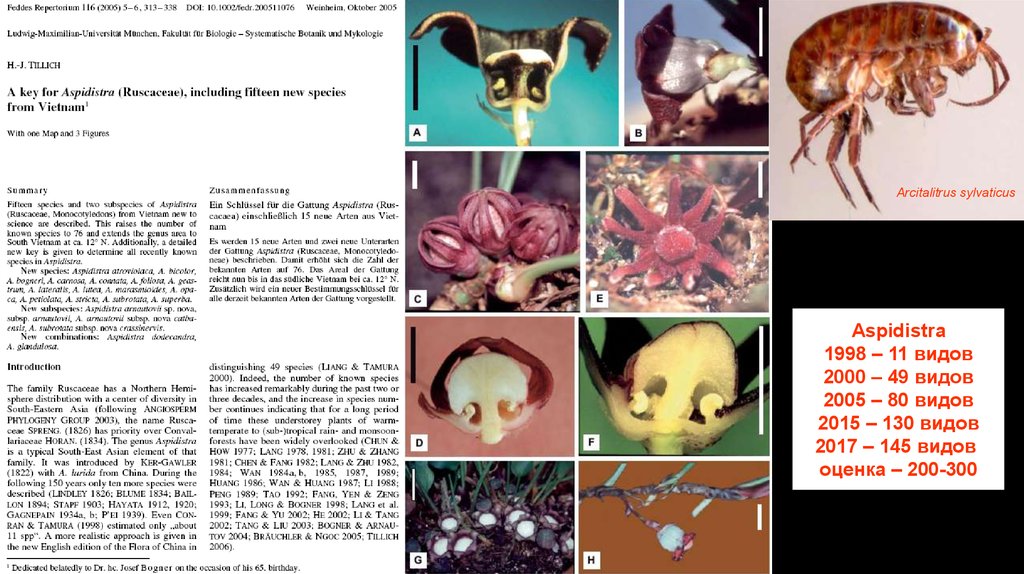
Aspidistra xuansonensis Vislobokov
(Asparagaceae). Вьетнам.
Symphytum × mosquense
S.R.Majorov & D.D.Sokoloff (Boraginaceae).
Москва.
Strobilanthes barbigera J.R.I.Wood, Nuraliev &
Scotland (Acanthaceae). Вьетнам.
Lotus stepposus Kramina
(Fabaceae). Украина.
Corbichonia exellii Sukhor.
(Lophiocarpaceae). Намибия.
Argostemma cordatum
Nuraliev (Rubiaceae).
Вьетнам.
Seychellaria barbata
Nuraliev & Cheek (Triuridaceae).
Мадагаскар.
Thismia mucronata Nuraliev
(Thismiaceae). Вьетнам.
Trithuria cowieana D.D.Sokoloff, Remizowa,
T.D.Macfarl. & Rudall (Hydatellaceae).
Австралия.
10.
Aspidistraelatior
11.
Arcitalitrus sylvaticusAspidistra
1998 – 11 видов
2000 – 49 видов
2005 – 80 видов
2015 – 130 видов
2017 – 145 видов
оценка – 200-300
12.
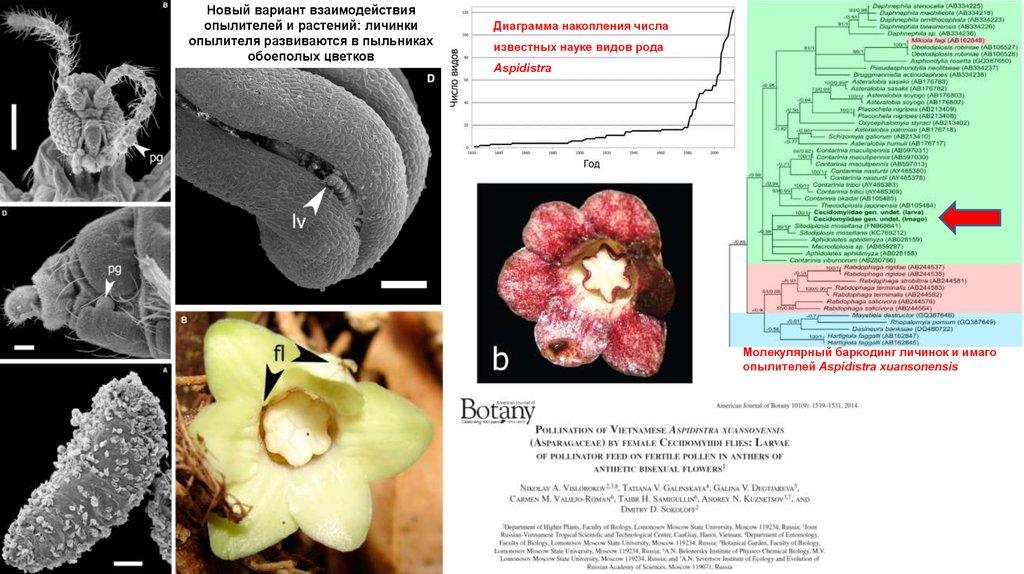
Новый вариант взаимодействияопылителей и растений: личинки
опылителя развиваются в пыльниках
обоеполых цветков
Диаграмма накопления числа
известных науке видов рода
Aspidistra
Молекулярный баркодинг личинок и имаго
опылителей Aspidistra xuansonensis
13.
14.
15.
16.
Ginkgo ♀17.
18. Типы систем живых организмов
• Искусственные – произвольный выбор небольшогочисла признаков как основы системы
• Естественные – анализ большого числа признаков и
выделение самых значимых как результат работы
систематика; попытка отразить в системе «естественно
присущий природе порядок» (который мог пониматься,
например, как план творения).
• Эволюционные – система должна отражать
представления об эволюции
• Кладистические – система должна однозначно
следовать из представлений об эволюции. Два аспекта –
переход от филогенетического дерева к системе и
технические алгоритмы построения деревьев.
19.
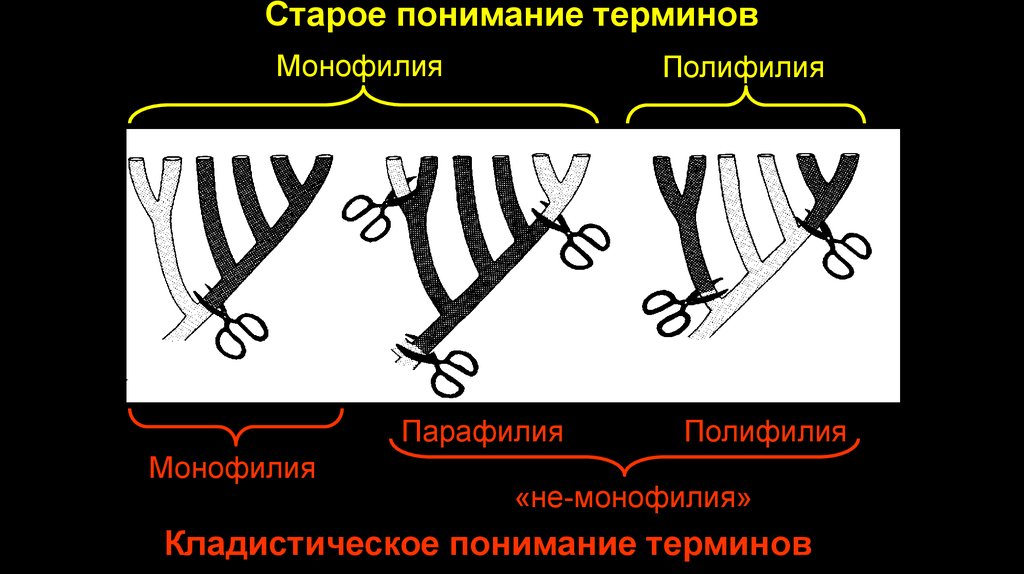
Старое понимание терминовМонофилия
Полифилия
Парафилия
Монофилия
Полифилия
«не-монофилия»
Кладистическое понимание терминов
20.
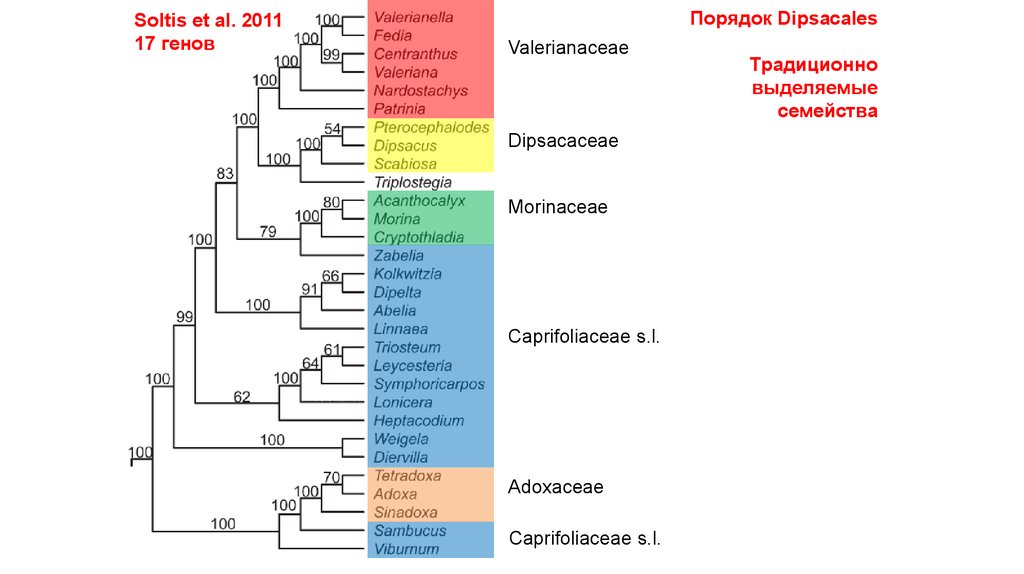
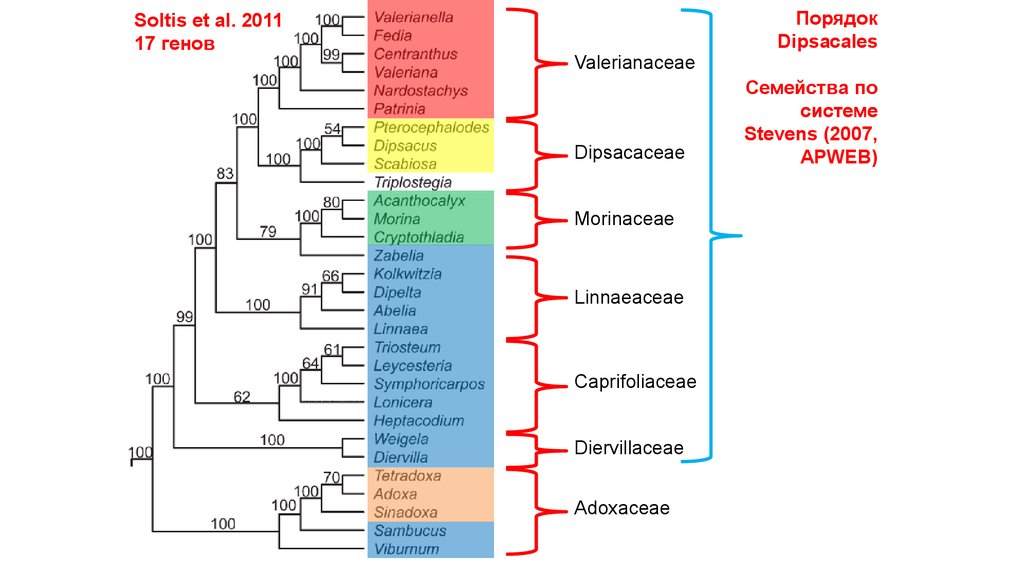
Soltis et al. 201117 генов
Порядок Dipsacales
Valerianaceae
Dipsacaceae
Morinaceae
Caprifoliaceae s.l.
Adoxaceae
Caprifoliaceae s.l.
Традиционно
выделяемые
семейства
21.
ПорядокDipsacales
Soltis et al. 2011
17 генов
Valerianaceae
Dipsacaceae
Morinaceae
Linnaeaceae
Caprifoliaceae
Diervillaceae
Adoxaceae
Семейства по
системе
Stevens (2007,
APWEB)
22. Кладизм, или «филогенетическая систематика»
W. Hennig (1950, 1966) – основатель кладизма
монофилия – парафилия – полифилия
признаки – состояния признаков
апоморфные и плезиоморфные состояния
признаков
синапоморфии и симплезиоморфии
использование внешней группы
принцип максимальной экономии (парсимонии)
23.
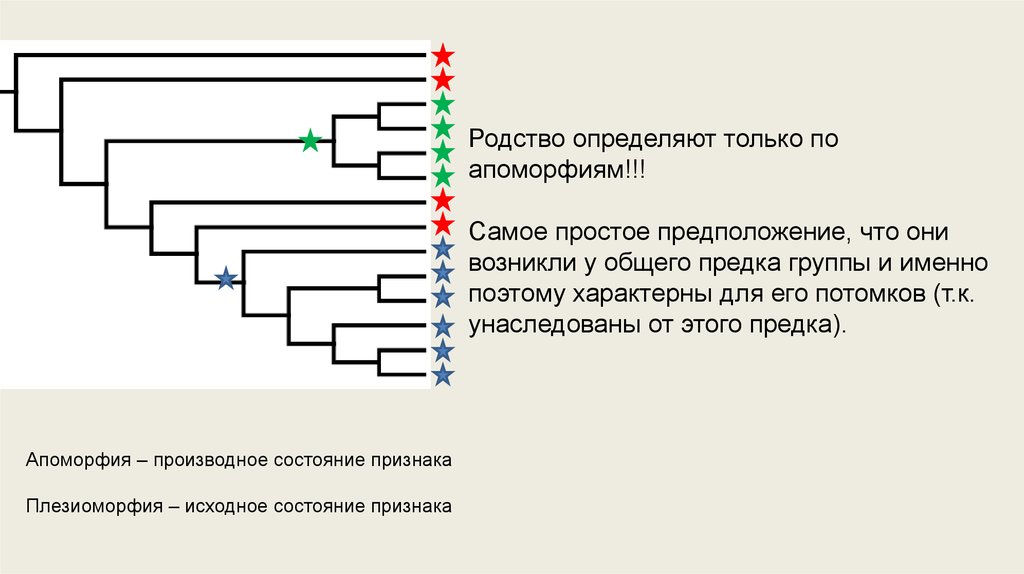
Родство определяют только поапоморфиям!!!
Самое простое предположение, что они
возникли у общего предка группы и именно
поэтому характерны для его потомков (т.к.
унаследованы от этого предка).
Апоморфия – производное состояние признака
Плезиоморфия – исходное состояние признака
24.
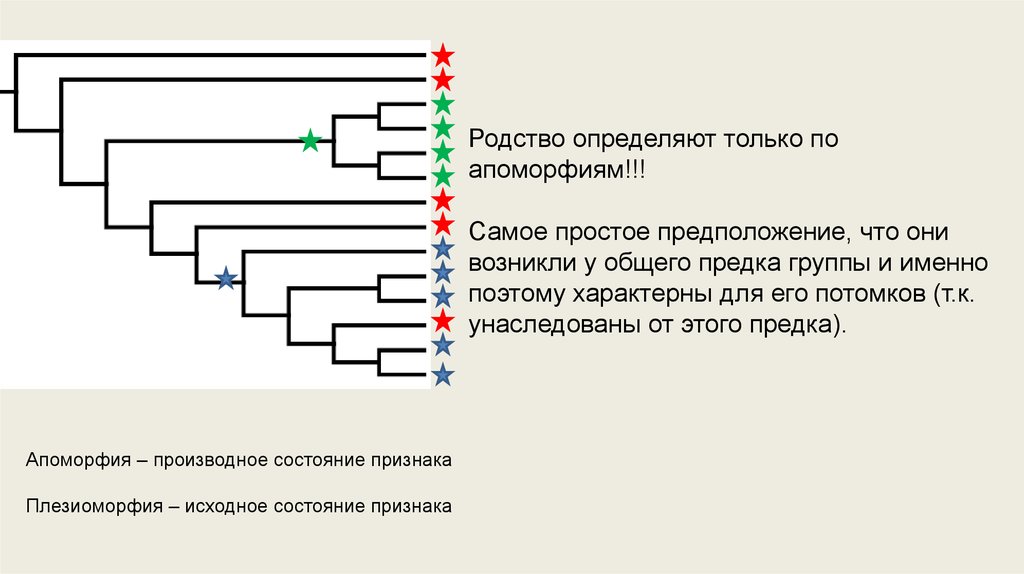
Родство определяют только поапоморфиям!!!
Самое простое предположение, что они
возникли у общего предка группы и именно
поэтому характерны для его потомков (т.к.
унаследованы от этого предка).
Апоморфия – производное состояние признака
Плезиоморфия – исходное состояние признака
25.
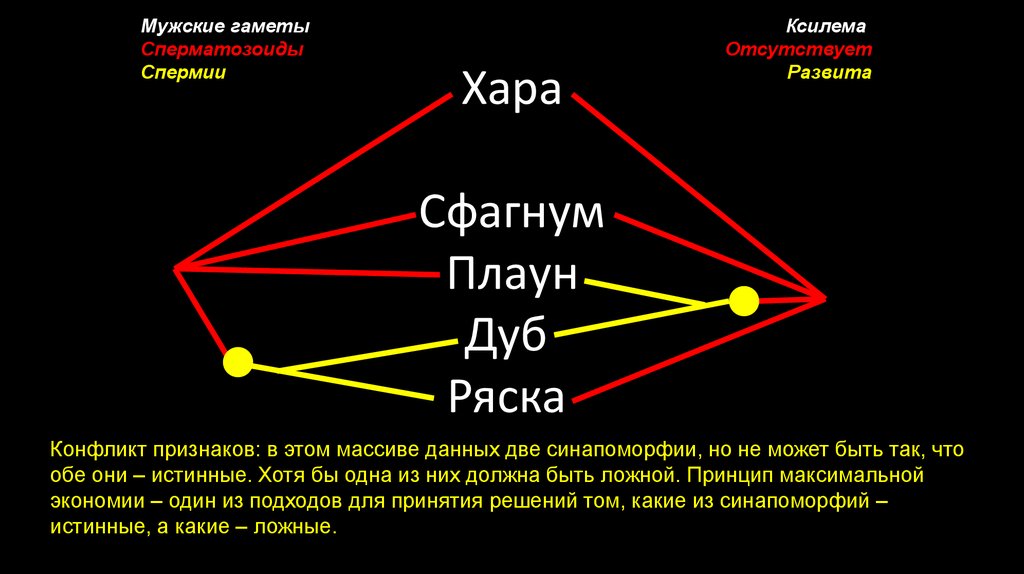
Мужские гаметыСперматозоиды
Спермии
Хара
Ксилема
Отсутствует
Развита
Сфагнум
Плаун
Дуб
Ряска
Конфликт признаков: в этом массиве данных две синапоморфии, но не может быть так, что
обе они – истинные. Хотя бы одна из них должна быть ложной. Принцип максимальной
экономии – один из подходов для принятия решений том, какие из синапоморфий –
истинные, а какие – ложные.
26.
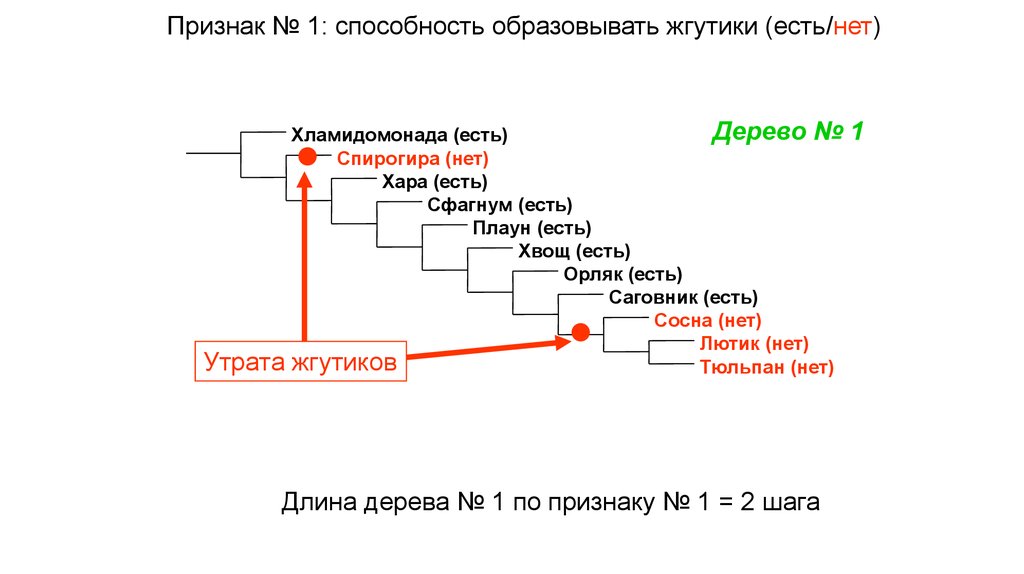
Признак № 1: способность образовывать жгутики (есть/нет)Утрата
Дерево №
Хламидомонада (есть)
Спирогира (нет)
Хара (есть)
Сфагнум (есть)
Плаун (есть)
Хвощ (есть)
Орляк (есть)
Саговник (есть)
Сосна (нет)
Лютик (нет)
жгутиков
Тюльпан (нет)
Длина дерева № 1 по признаку № 1 = 2 шага
1
27.
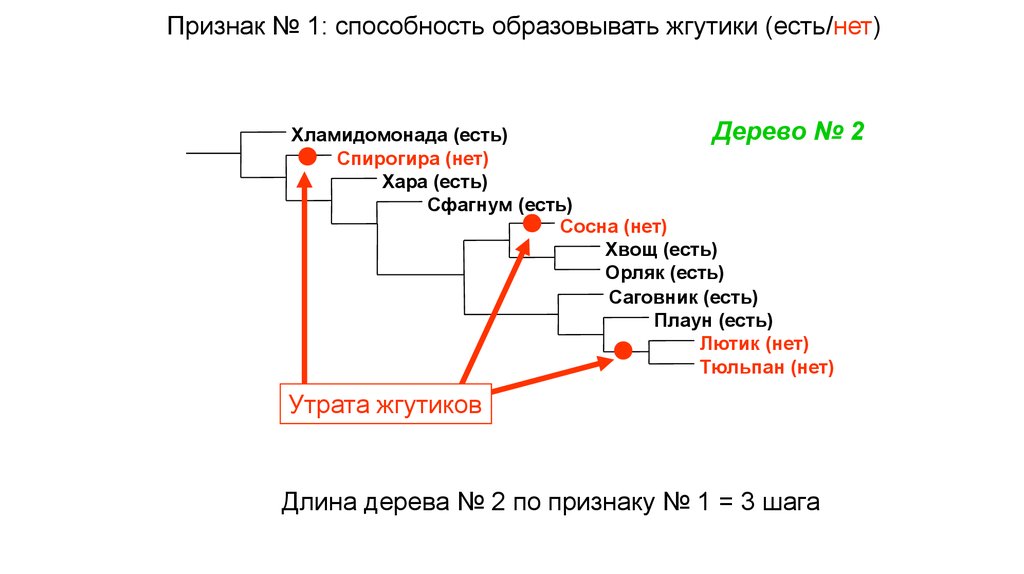
Признак № 1: способность образовывать жгутики (есть/нет)Дерево №
Хламидомонада (есть)
Спирогира (нет)
Хара (есть)
Сфагнум (есть)
Сосна (нет)
Хвощ (есть)
Орляк (есть)
Саговник (есть)
Плаун (есть)
Лютик (нет)
Тюльпан (нет)
Утрата жгутиков
Длина дерева № 2 по признаку № 1 = 3 шага
2
28.
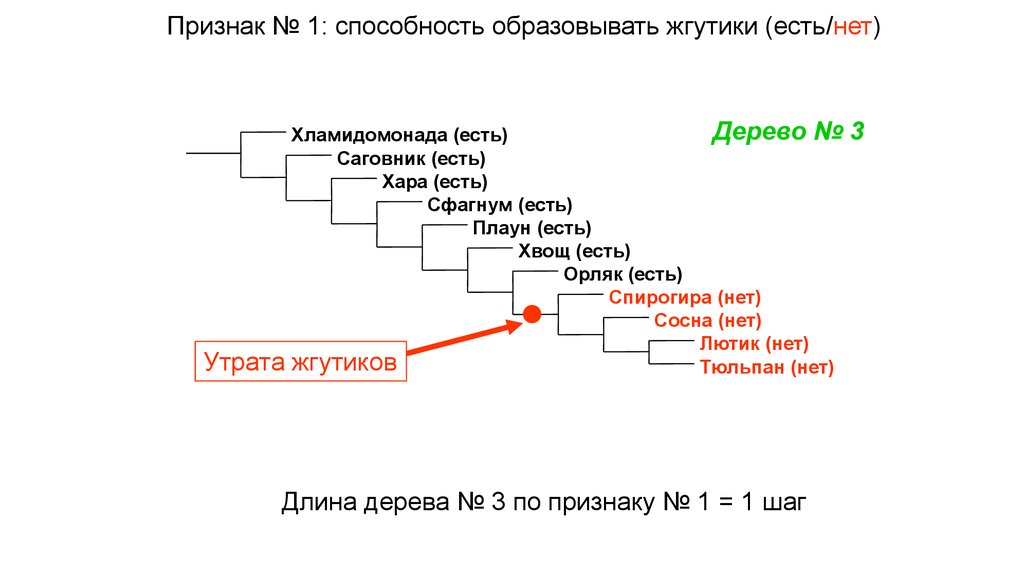
Признак № 1: способность образовывать жгутики (есть/нет)Утрата
Дерево №
Хламидомонада (есть)
Саговник (есть)
Хара (есть)
Сфагнум (есть)
Плаун (есть)
Хвощ (есть)
Орляк (есть)
Спирогира (нет)
Сосна (нет)
Лютик (нет)
жгутиков
Тюльпан (нет)
Длина дерева № 3 по признаку № 1 = 1 шаг
3
29.
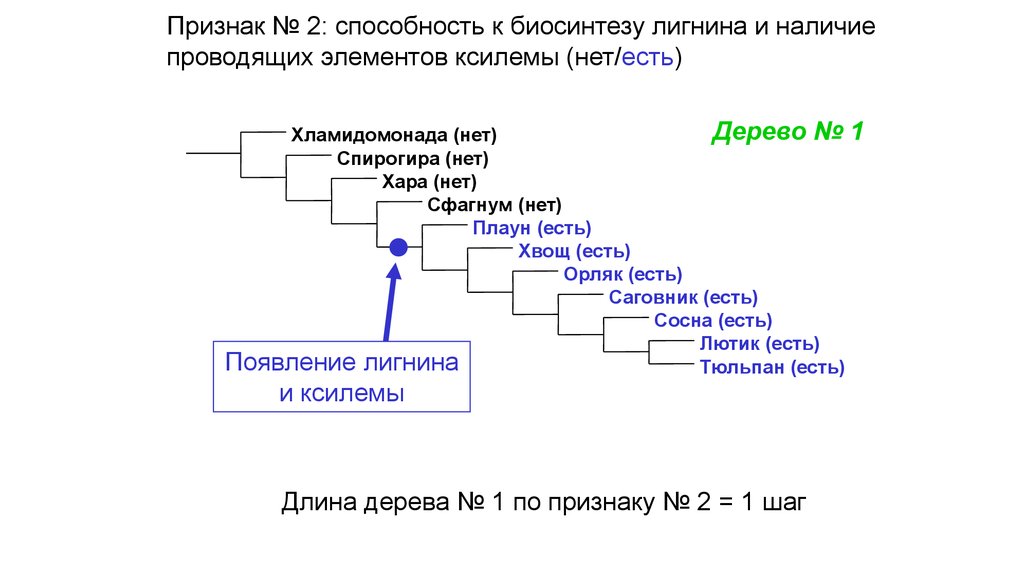
Признак № 2: способность к биосинтезу лигнина и наличиепроводящих элементов ксилемы (нет/есть)
Дерево № 1
Хламидомонада (нет)
Спирогира (нет)
Хара (нет)
Сфагнум (нет)
Плаун (есть)
Хвощ (есть)
Орляк (есть)
Саговник (есть)
Сосна (есть)
Лютик (есть)
Появление лигнина
Тюльпан (есть)
и ксилемы
Длина дерева № 1 по признаку № 2 = 1 шаг
30.
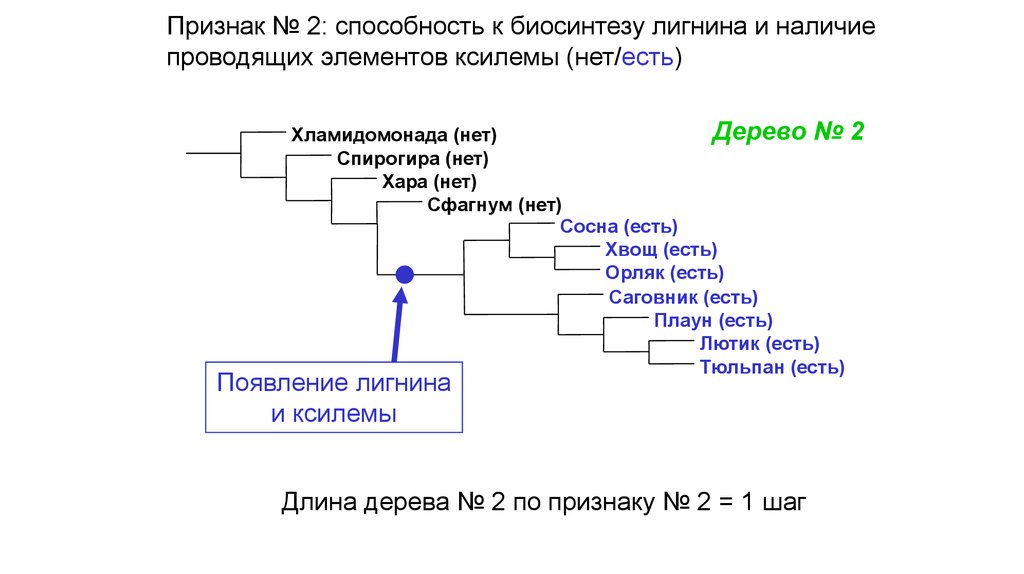
Признак № 2: способность к биосинтезу лигнина и наличиепроводящих элементов ксилемы (нет/есть)
Дерево № 2
Хламидомонада (нет)
Спирогира (нет)
Хара (нет)
Сфагнум (нет)
Сосна (есть)
Хвощ (есть)
Орляк (есть)
Саговник (есть)
Плаун (есть)
Лютик (есть)
Тюльпан (есть)
Появление лигнина
и ксилемы
Длина дерева № 2 по признаку № 2 = 1 шаг
31.
Признак № 2: способность к биосинтезу лигнина и наличиепроводящих элементов ксилемы (нет/есть)
Дерево № 3
Хламидомонада (нет)
Саговник (есть)
Хара (нет)
Сфагнум (нет)
Плаун (есть)
Хвощ (есть)
Орляк (есть)
x Спирогира (нет)
Сосна (есть)
Появление лигнина
Лютик (есть)
Тюльпан (есть)
и ксилемы
Утрата лигнина и
ксилемы
Длина дерева № 3 по признаку № 2 = 3 шага
32.
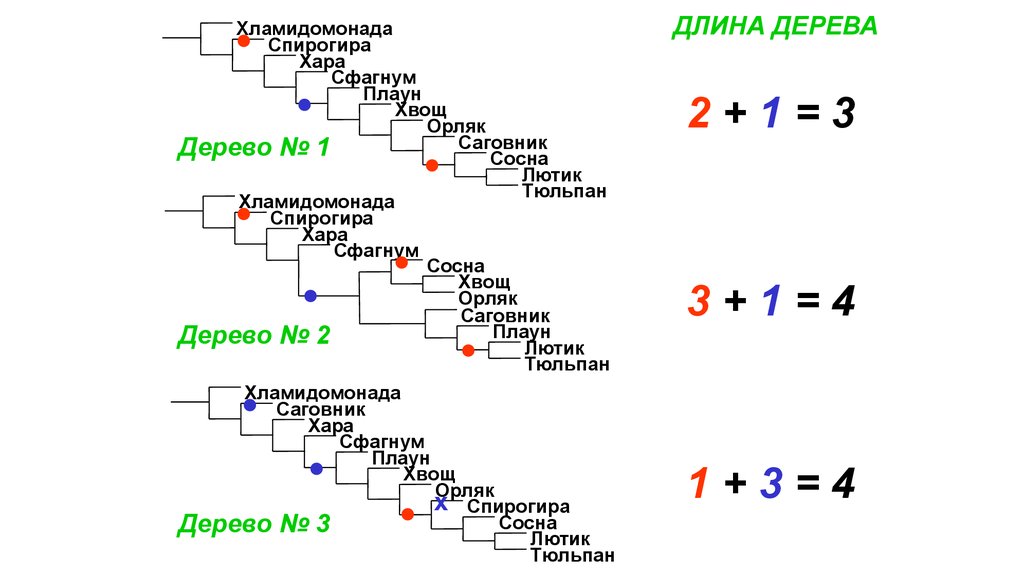
ХламидомонадаСпирогира
Хара
Сфагнум
Плаун
Хвощ
Орляк
Саговник
Дерево № 1
Сосна
Лютик
Тюльпан
Хламидомонада
Спирогира
Хара
Сфагнум
Сосна
Хвощ
Орляк
Саговник
Плаун
Дерево № 2
Лютик
Тюльпан
Хламидомонада
Саговник
Хара
Сфагнум
Плаун
Хвощ
Орляк
x Спирогира
Сосна
Дерево № 3
Лютик
Тюльпан
ДЛИНА ДЕРЕВА
2+1=3
3+1=4
1+3=4
33.
ХламидомонадаСпирогира
Хара
Сфагнум
Плаун
Хвощ
Орляк
Саговник
Сосна
Лютик
Тюльпан
Хламидомонада
Спирогира
Хара
Сфагнум
Плаун
Хвощ
Орляк
Саговник
Сосна
Лютик
Тюльпан
34.
Общий вид первичных данных, используемых в молекулярнофилогенетическом анализеПризнак – позиция нуклеотида
Состояние признака – A, T, G, C, наличие/отсутствие
35.
Примитивныедвудольные
Однодольные
Высшие
двудольные
Филогения цветковых
растений из работы
Angiosperm Phylogeny
Group (2003), данные
в основном
по 3 генам
(18S rDNA, rbcL, atpB)
Ближайший общий предок
примитивных двудольных
– одновременно общий
предок всех
покрытосеменных
36.
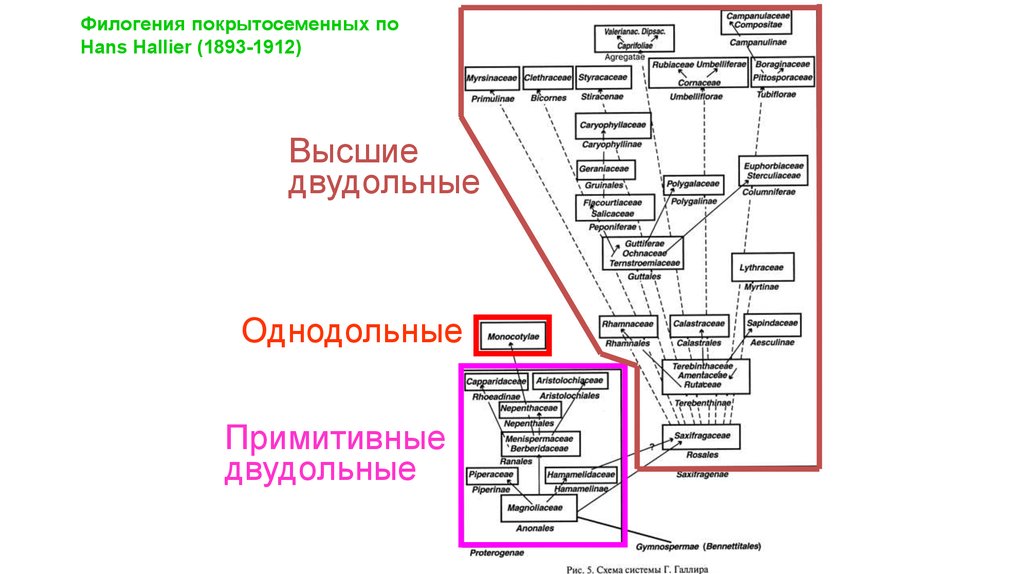
Филогения покрытосеменных поHans Hallier (1893-1912)
Высшие
двудольные
Однодольные
Примитивные
двудольные
37.
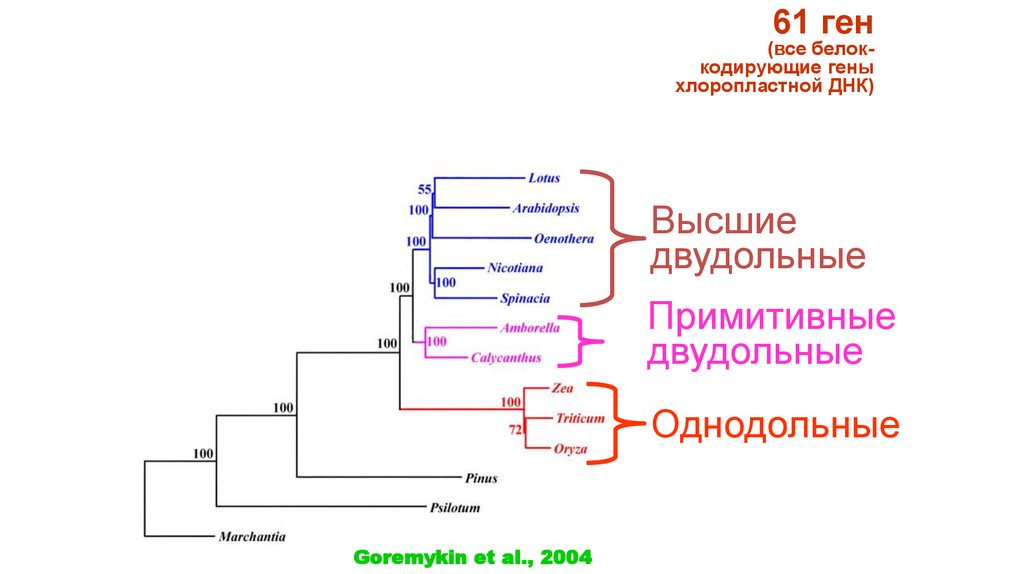
61 ген(все белоккодирующие гены
хлоропластной ДНК)
Высшие
двудольные
Примитивные
двудольные
Однодольные
Goremykin et al., 2004
38.
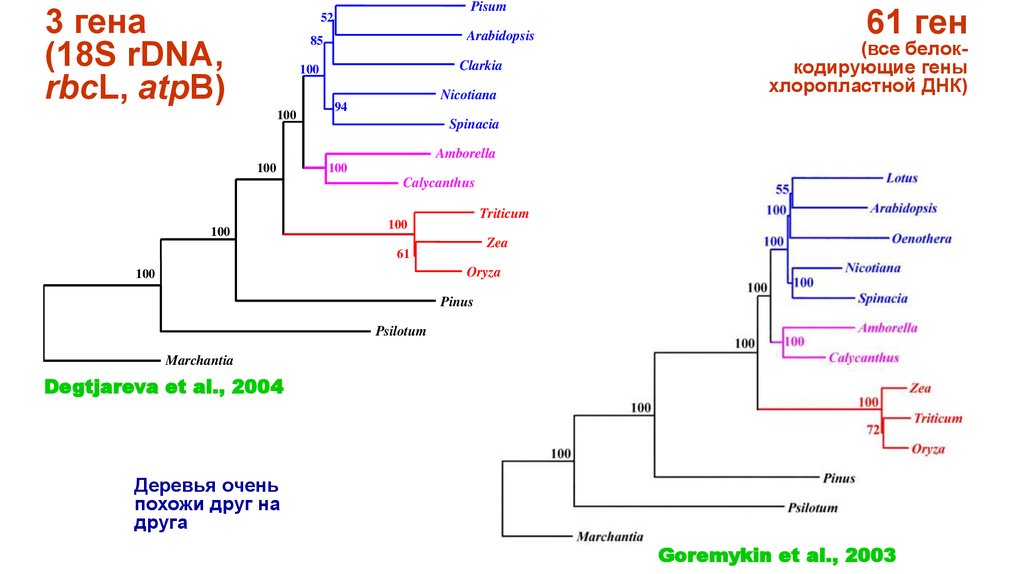
3 гена(18S rDNA,
rbcL, atpB)
Pisum
52
Arabidopsis
85
Clarkia
100
Nicotiana
100
61 ген
(все белоккодирующие гены
хлоропластной ДНК)
94
Spinacia
Amborella
100
100
Calycanthus
100
Triticum
100
Zea
61
Oryza
100
Pinus
Psilotum
Marchantia
Degtjareva et al., 2004
Деревья очень
похожи друг на
друга
Goremykin et al., 2003
39.
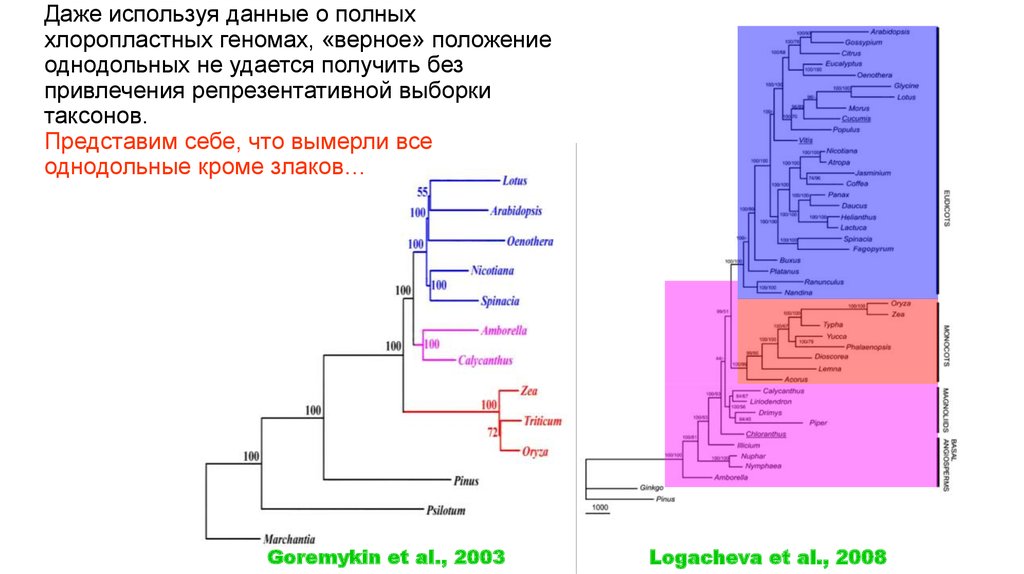
Даже используя данные о полныххлоропластных геномах, «верное» положение
однодольных не удается получить без
привлечения репрезентативной выборки
таксонов.
Представим себе, что вымерли все
однодольные кроме злаков…
Goremykin et al., 2003
Logacheva et al., 2008
40.
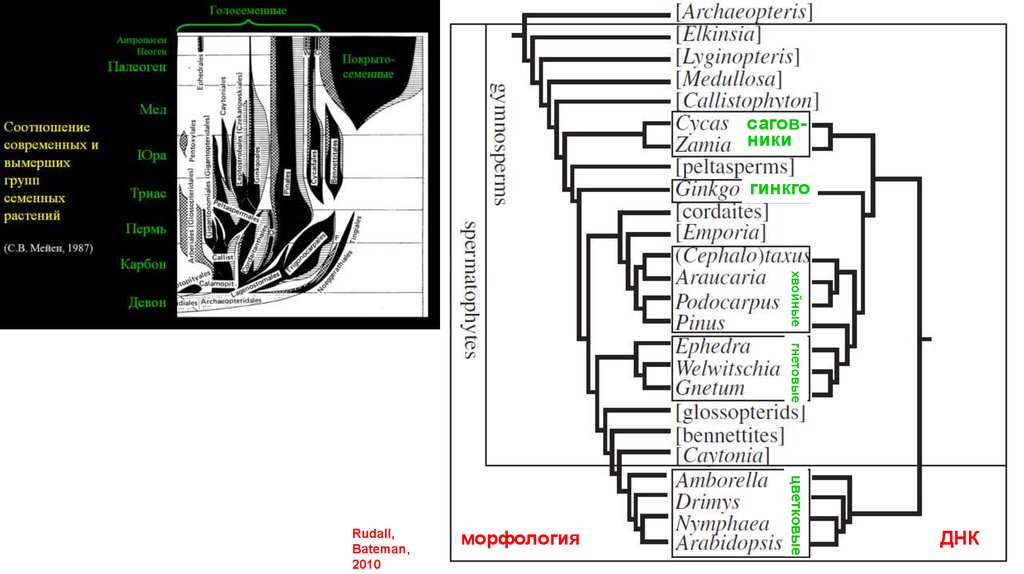
саговникигинкго
хвойные
гнетовые
морфология
цветковые
Rudall,
Bateman,
2010
ДНК
41.
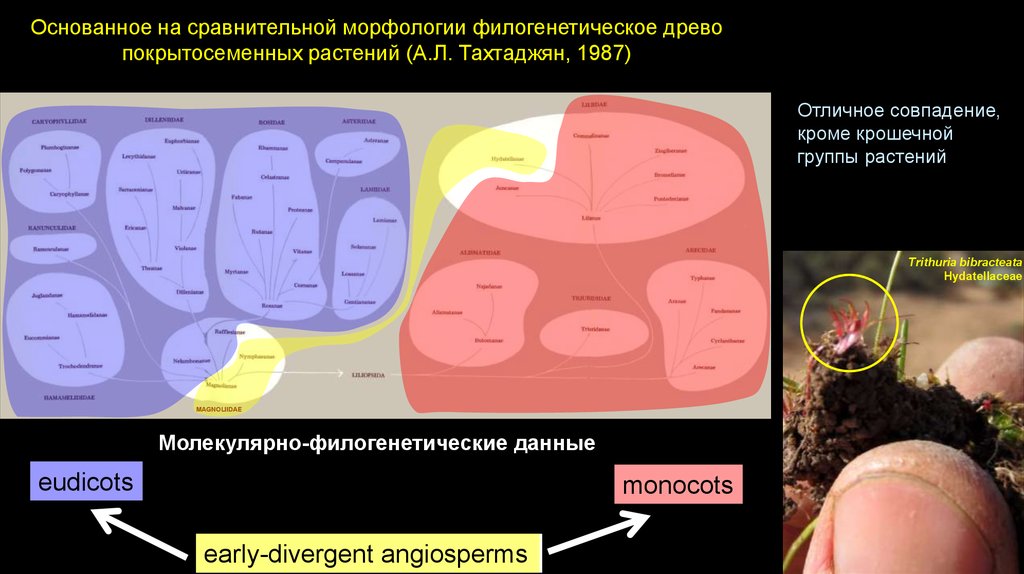
Основанное на сравнительной морфологии филогенетическое древопокрытосеменных растений (А.Л. Тахтаджян, 1987)
Отличное совпадение,
кроме крошечной
группы растений
Trithuria bibracteata
Hydatellaceae
MAGNOLIIDAE
Молекулярно-филогенетические данные
eudicots
eudicots
monocots
monocots
Early-divergent
angiosperms
early-divergent
angiosperms
42.
Основанное на сравнительной морфологии филогенетическое древо покрытосеменных растений(А.Л. Тахтаджян, 1987). Цветом показана выделяемая сейчас группа розид
43.
A ‘sandwich effect’: the degree ofincongruence between morphologybased systems and molecular
phylogenies depends on taxon rank
subkingdom Embryophyta
superdivision Tracheophyta
division Spermatophyta
subdivision Angiospermae
class Dicotyledones
subclass Rosidae
order Fabales
family Leguminosae
subfamily Papilionoideae
group of tribes ‘Galegoid complex’
tribe Loteae
genus Lotus
subgenus Lotus
section Lotus
Lotus corniculatus species group
species Lotus corniculatus
На разных ступенях иерархической
системы степень «революционности»
изменений различна
OK
OK
OK
ОК
almost ОК
not ОК
not ОК
ОК
ОК
not ОК
ОК
almost ОК
not ОК
not ОК
ОК
44.
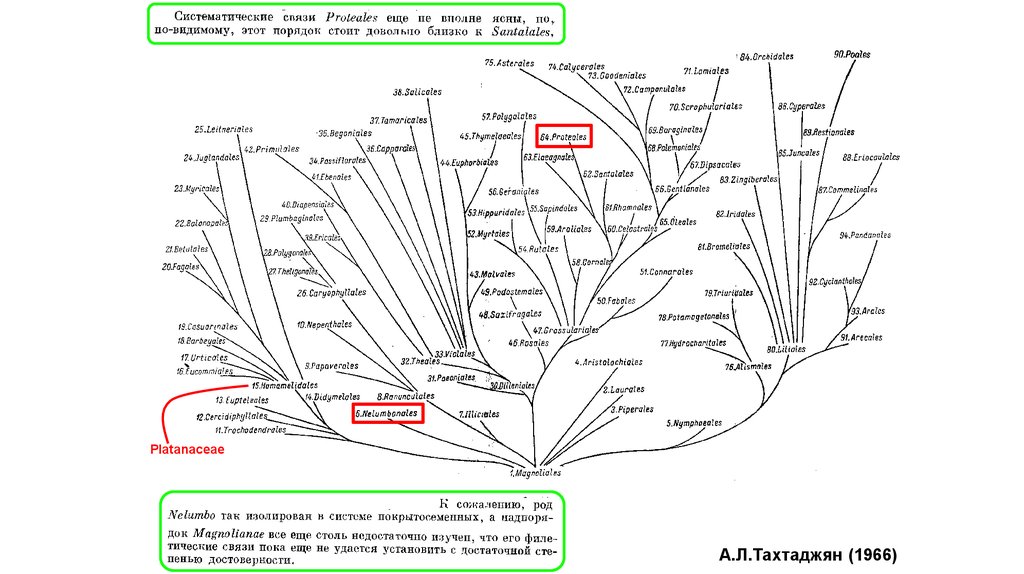
Platanus, Nelumbo, Proteaceae45.
PlatanaceaeА.Л.Тахтаджян (1966)
46.
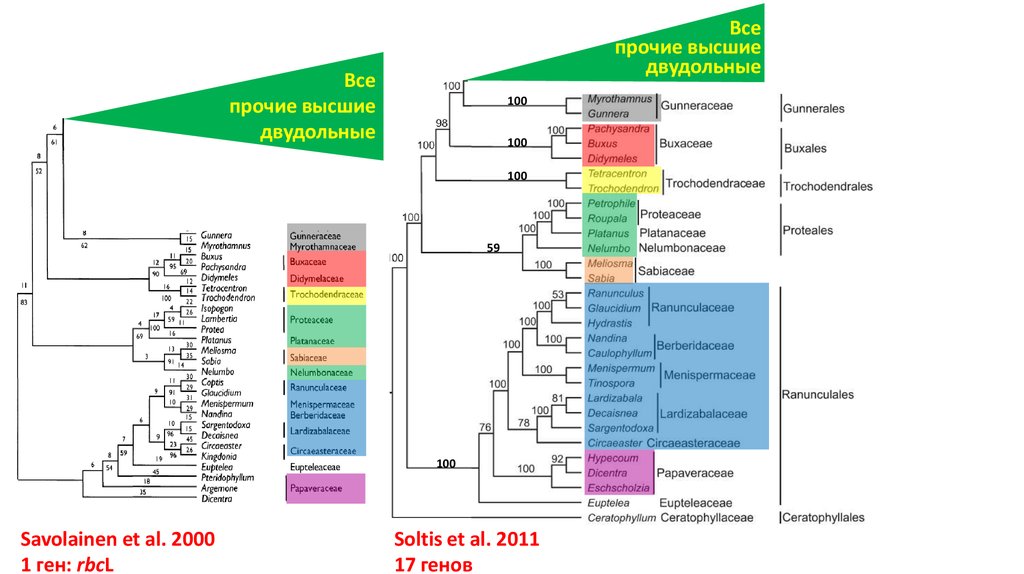
Всепрочие высшие
двудольные
Все
прочие высшие
двудольные
100
100
100
59
100
Savolainen et al. 2000
1 ген: rbcL
Soltis et al. 2011
17 генов
47.
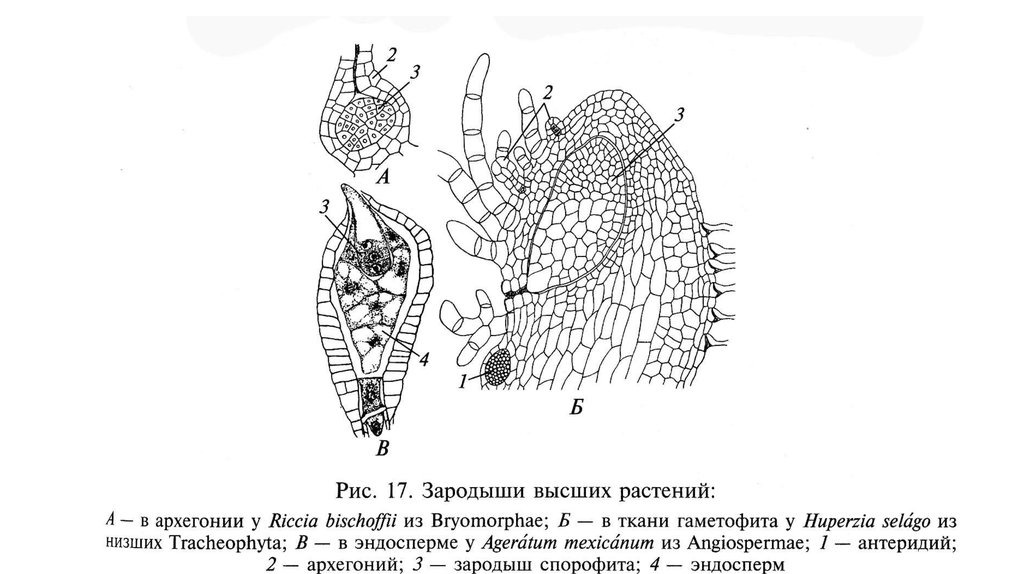
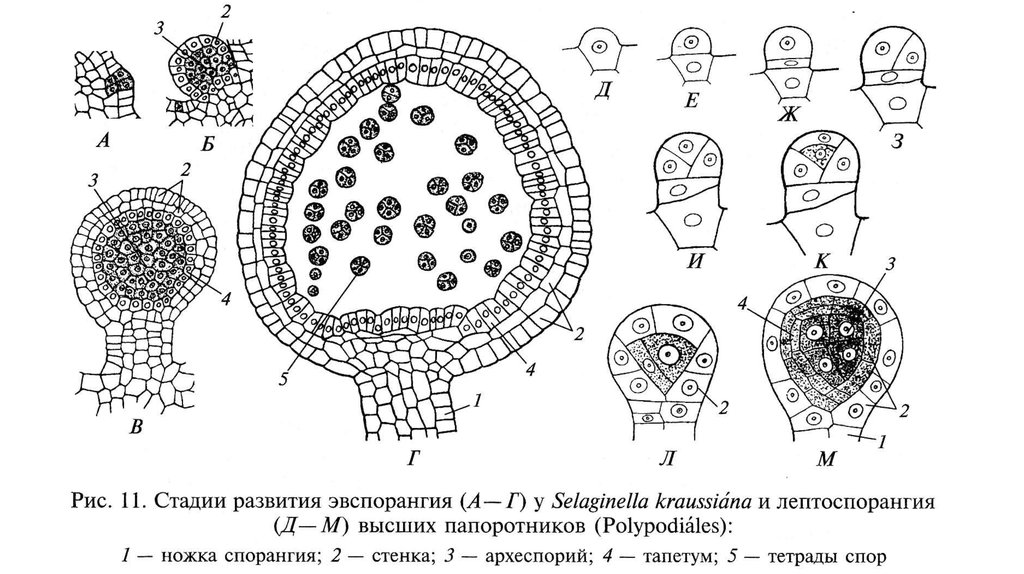
СПОРОФИТЗИГОТА
СПОРАНГИИ
ОПЛОДОТВОРЕНИЕ
ГАМЕТЫ
МЕЙОЗ
СПОРЫ
ГАМЕТАНГИИ
(антеридии и
архегонии)
ГАМЕТОФИТ
48.
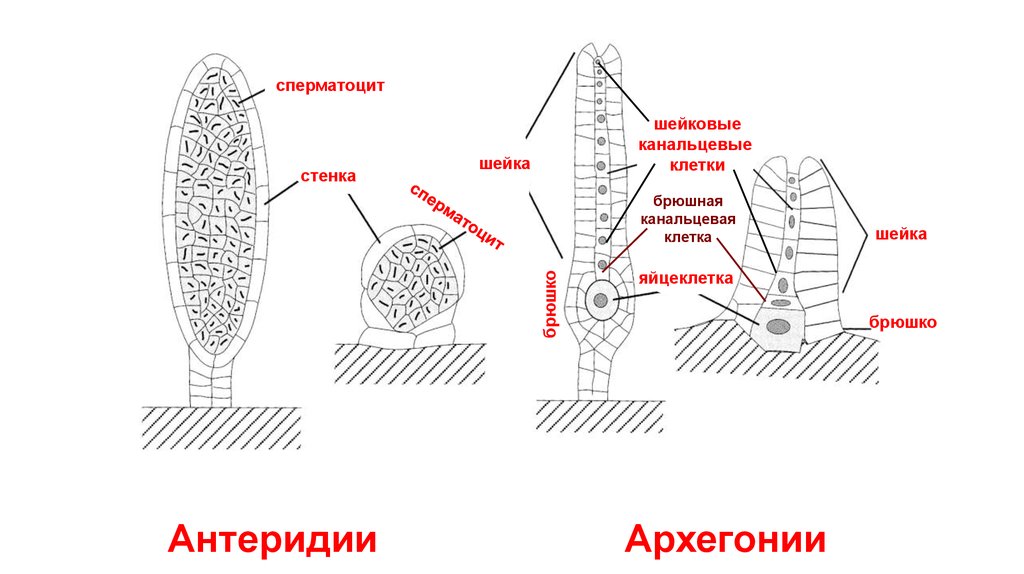
сперматоцитстенка
шейковые
канальцевые
клетки
шейка
брюшко
брюшная
канальцевая
клетка
Антеридии
шейка
яйцеклетка
брюшко
Архегонии
49.
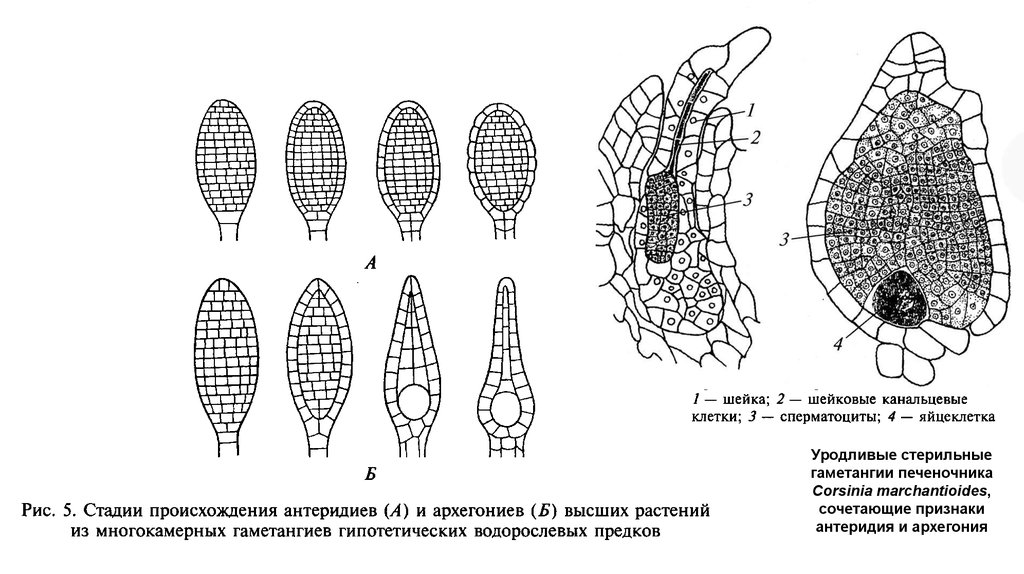
Уродливые стерильныегаметангии печеночника
Corsinia marchantioides,
сочетающие признаки
антеридия и архегония
50.
51.
52.
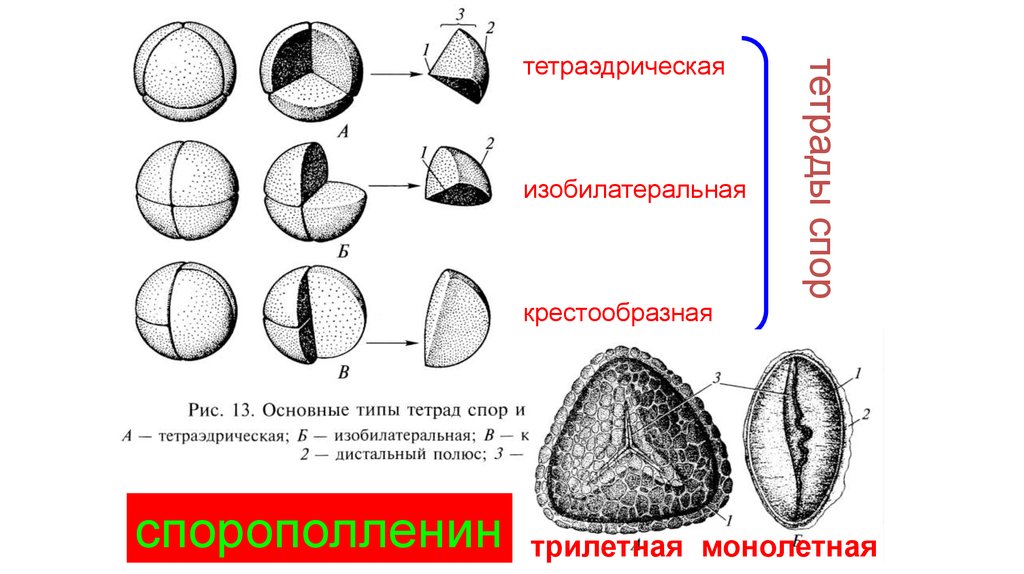
изобилатеральнаякрестообразная
спорополленин
тетрады спор
тетраэдрическая
трилетная монолетная