")
 – сукупність всіх хімічних реакцій і всіх видів перетворень речовин та енергії в організмі.")


 і катаболізму (процеси розпаду речовин)")









")












")

")





















 biology
biologySimilar presentations:




")





Обмін речовин та енергії (метаболізм)
1. ОБМІН РЕЧОВИН ТА ЕНЕРГІЇ (МЕТАБОЛІЗМ)
2. Обмін речовин (метаболізм) – сукупність всіх хімічних реакцій і всіх видів перетворень речовин та енергії в організмі.
3.
Хімічні речовини, що беруть участь в обмініречовин називаються метаболітами.
Поза клітиною майже всі ці перетворення
проходили б дуже повільно і не направлено.
Впорядковані послідовності хімічних реакцій, що
проходять з високою продуктивністю, так звані
метаболічні шляхи, можливі тільки завдяки
присутності в клітині специфічних ферментів.
4.
Метаболізмзабезпечує:
1. Розвиток
2. Життєдіяльність
3. Самовідтворення організмів
4. Зв'язок з оточуючим середовищем
5. Адаптацію до змін зовнішніх умов
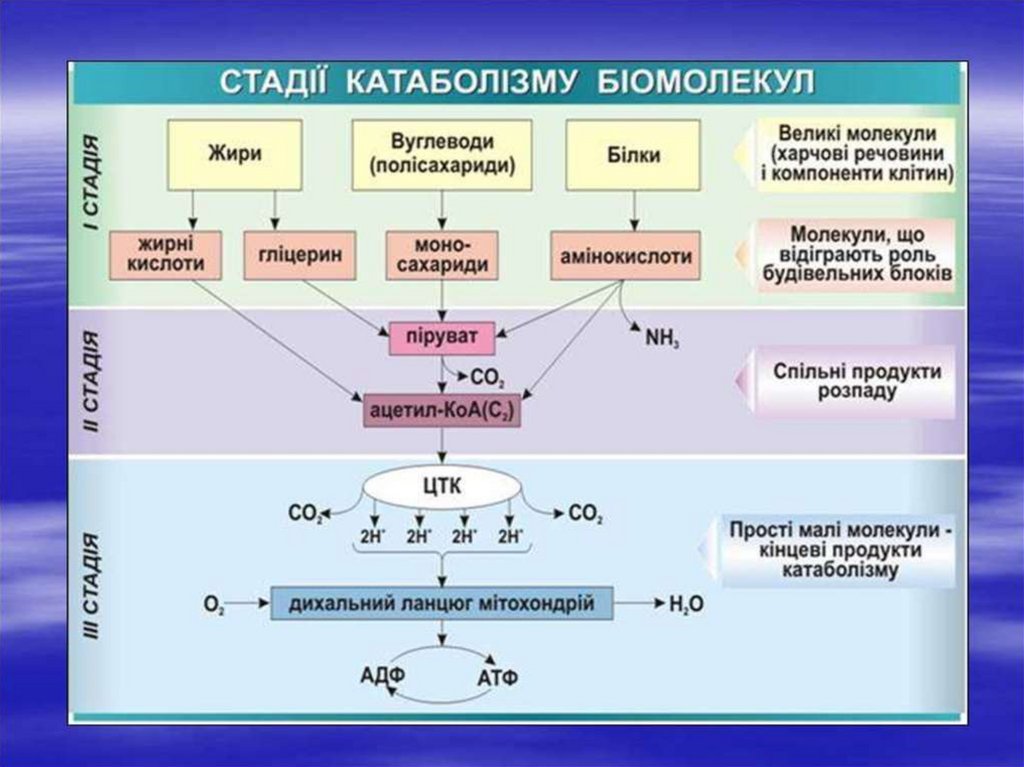
5. Метаболізм складається з анаболізму (процеси синтезу) і катаболізму (процеси розпаду речовин)
6.
7. Основа обміну речовин
Асиміляція (анаболізм):хімічні
перетворення,
що
приводять до використання
органічних речовин, які
надходять із зовнішнього
середовища для синтезу
специфічних білків, НК,
ліпідів, полісахаридів та ін.
Супроводжуються
поглинанням енергії
(ендергонічні)
Дисиміляція (катаболізм):
процеси розпаду органічних
речовин до кінцевих
продуктів – води, СО2,
амоніаку, сечовини, сечової
кислоти.
Супроводжуються
виділенням енергії
(екзергонічні)
8.
Ці процеси взаємопов'язані і направлені набезперервне поновлення живого матеріалу і
забезпечення його необхідною енергією.
Організм тварин і людини – відкрита
система, яка потребує притоку енергії
ззовні.
9.
Живі клітини постійно потребують органічні інеорганічні речовини, а також хімічну енергію, яку вони
отримують переважно з АТФ (АТР).
Всі організми ділять на автотрофні та гетеротрофні.
Автотрофні організми, до яких належать рослини і
більшість
мікроорганізмів,
можуть
синтезувати
органічні молекули з неорганічних попередників (CO2),
наприклад, за рахунок фотосинтезу.
Гетеротрофи, наприклад тварини та гриби, залежать
від одержання органічних речовин з їжею.
10.
Із багаточисельних метаболітів виділяють 3найбільш важливих – піруват (ПВК), ацетил-КоА
і гліцерин.
Вони є ланкою, що зв'язує між собою метаболізм
білків, вуглеводів та ліпідів.
Кінцевими продуктами розпаду органічних
речовин у тварин є диоксид Карбону (CO2), вода
(H2O) та амоніак (NH3). Амоніак перетворюється
в сечовину і в такій формі виводиться з
організму.
11.
12.
Ні вуглеводи, ні жири, ні білки не можутьбезпосередньо бути «паливом» для клітинних
процесів.
Клітини тварин можуть використовувати
тільки
енергію
розриву
хімічних
зв'язків!
Вона може передаватися з однієї частини
клітини в іншу і використовуватися
економно.
13. Макроергічні сполуки
- це сполуки, що містять макроергічні (багаті на енергію)фосфатні зв'язки.
В результаті їх гідролізу виділяється більше 5 ккал/моль
енергії.
Знаком ~ (тильда) позначають макроергічний зв'язок.
До макроергів відносять: АТФ, УТФ, ЦТФ, ГТФ, ТТФ,
14.
Центральне місце серед макроергічнихсполук займає АТФ – молекула, багата
енергією,
оскільки
вона
містить
2
фосфоефірних зв'язки.
В результаті гідролізу АТФ перетворюється в
АДФ і ортофосфат (Р).
При цьому виділяється 7,3 ккал/моль вільної
енергії.
15. Структура АТФ
16. Аденозинтрифосфорна кислота (АТФ)
17.
АТФ – універсальна «енергетичнавалюта» всіх живих організмів.
АТФ мобільна, потрапляє в любу частину
клітини, де вона може підлягати гідролізу з
виділенням вільної енергії.
В залежності від глибини гідролізу АТФ в
якості кінцевого продукту може утворитися
АДФ або АМФ.
18.
В клітині молекула АТФ використовується протягом 1хвилини після її утворення.
У людини кількість АТФ, рівне за величиною масі його
тіла, використовується протягом 24 годин.
Клітина повинна мати надійний механізм поповнення
АТФ і продукції тепла.
Синтез АТФ у клітині може здійснюватися
двома шляхами:
Субстратне фосфорилування, під час якого
відбувається безпосередня передача молекули
активного фосфату на АДФ від більш енерговмісних
сполук.
Окисне
фосфорилування, яке відбувається в
мітохондріях і спряжене з тканинним диханням.
19. Біологічне окиснення може відбуватися кількома шляхами:
- відщеплення відсубстратів атомів
Гідрогену і електронів
(дегідрування):
на внутрішній мембрані
мітохондрій;
Енергія: АТФ + тепло;
Ф-ція: забезпечення
процесів біоокиснення.
- приєднання до
субстрату Оксигену
(оксигенація):
в мікросомах;
тепло;
знезараження
чужорідних сполук,
токсинів. Синтез
органічних речовин.
20. Біологічне окиснення
Це сукупність окисно-відновних процесів убіологічних об'єктах, які відбуваються в
клітинах
організму
в
період
внутрішньоклітинного обміну та мають
важливу роль в забезпеченні організму
енергією і метаболітами.
21. Функції біологічного окиснення:
1.2.
3.
4.
5.
Енергетичне забезпечення процесів
життєдіяльності: підтримання температури
тіла, хімічні синтези, електричні процеси,
механічна робота.
Усунення кінцевих і шкідливих продуктів
обміну речовин.
Знезараження чужорідних сполук.
Синтез ключових метаболітів.
Регуляція обміну речовин шляхом змінення
співвідношення окиснених і відновлених форм
коферментів (НАД ⁄ НАД·Н).
22. Окиснення органічних речовин Оксигеном до СО2 і Н2О називається тканинним диханням або біологічним окисненням.
23.
Тканинне дихання відбувається за участіланцюгу перенесення електронів (ЛПЕ) або
дихального ланцюгу,
який утворений окисно-відновними
ферментами, локалізованими в
ліпідному шарі внутрішньої мембрани
мітохондрій.
24.
25.
26.
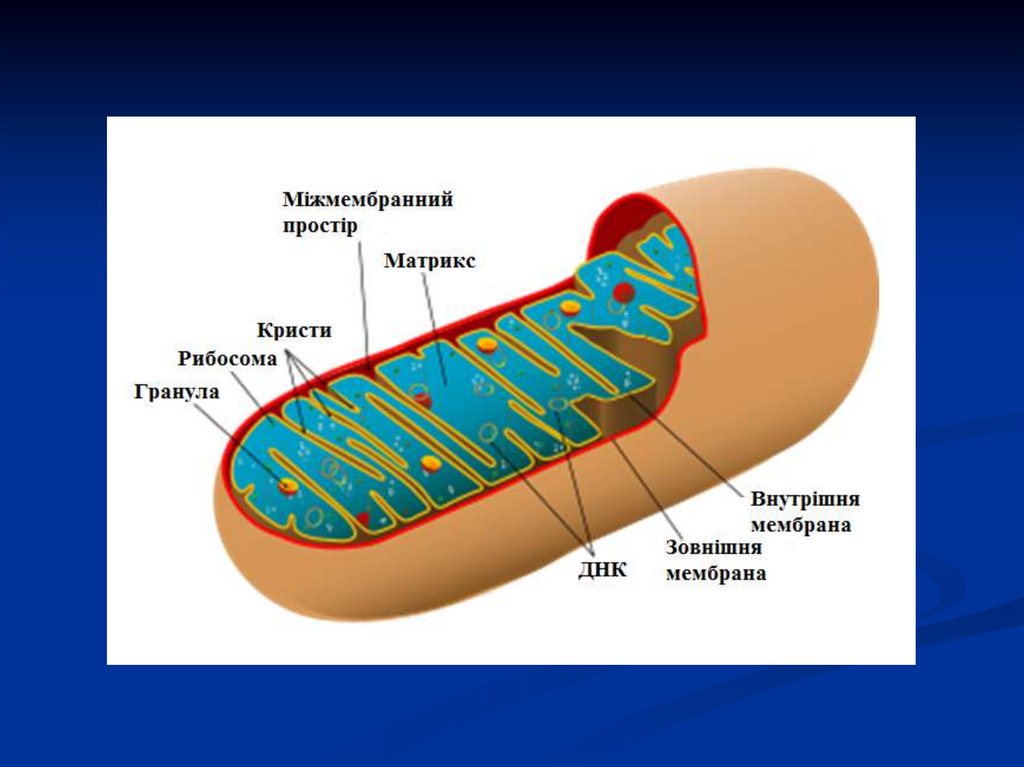
Уперше мітохондрії (МХ) були виявлені у вигляді гранул ум'язових клітинах (1850 р.). Кількість МХ у клітинах мінлива. Їх
особливо багато у клітинах з великою потребою у кисні. Залежно
від того, в яких ділянках клітини відбувається споживання
енергії, МХ здатні переміщуватись туди по цитоплазмі.
МХ мають дві замкнені, ізольовані мембрани: внутрішню
та зовнішню, які розділені водним міжмембранним простором.
Простір між крістами внутрішньої мембрани МХ
заповнений напіврідкою гелеподібною масою – матриксом, що
складається на 50% з білка. Матрикс містить ферменти: циклу
Кребса, β-окиснення жирних кислот (основні постачальники
субстратів
окиснення),
окиснення
амінокислот
та
піруватдегідрогеназний комплекс ферментів.
На відміну від інших внутрішньоклітинних органоїдів МХ
мають свій геном, тому в матриксі також знаходяться ферменти
автономного мітохондріального синтезу ДНК, РНК, білків тощо.
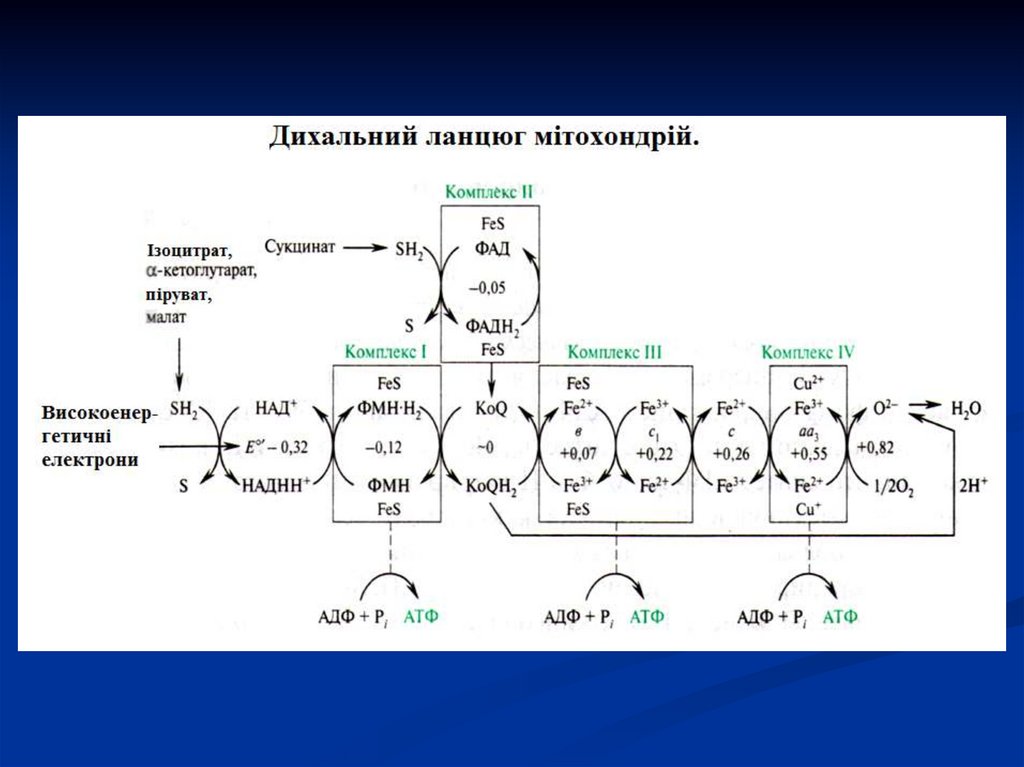
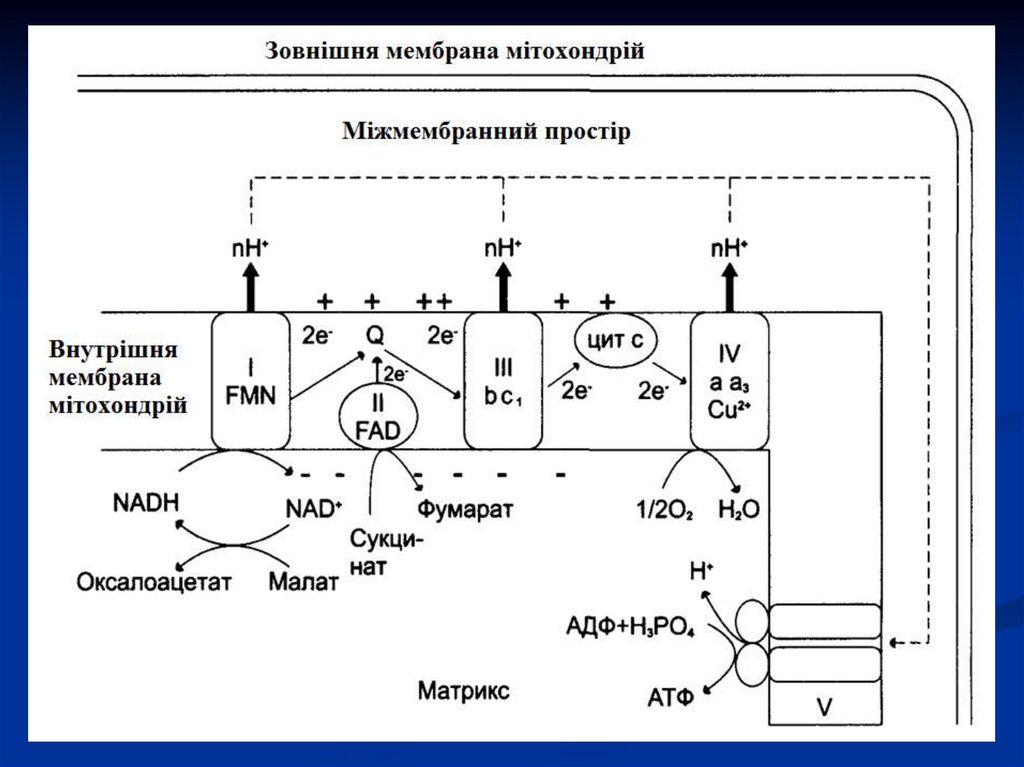
27. Дихальний ланцюг
Сукупність ферментів, що здійснюють транспортелектронів від речовини, що окиснюється, на О2
називається дихальним ланцюгом або ланцюгом
перенесення електронів (ЛПЕ).
Перенесення електронів в дихальному ланцюгу
забезпечується
різницею
потенціалів
між
окремими ланками цього ланцюгу.
Послідовний рух електронів в дихальному ланцюгу
супроводжується
ступінчатим
вивільненням
хімічної
енергії,
яка
акумулюється
в
макроергічних зв'язках АТФ.
28.
В ЛПЕ приймають участь4 групи ферментів класу оксидоредуктаз:
1. піридинзалежні дегідрогенази,
2. флавінзалежні дегідрогенази,
3. убіхінон або коензим Q;
4.
цитохромні системи (в, с, а), представлені
комплексом гемінових ферментів і
цитохромоксидаза (а3).
Всі 4 групи коферментів сприяють перенесенню
протонів і електронів від органічних речовин,
що окиснюються, на Оксиген.
29.
30.
31.
32.
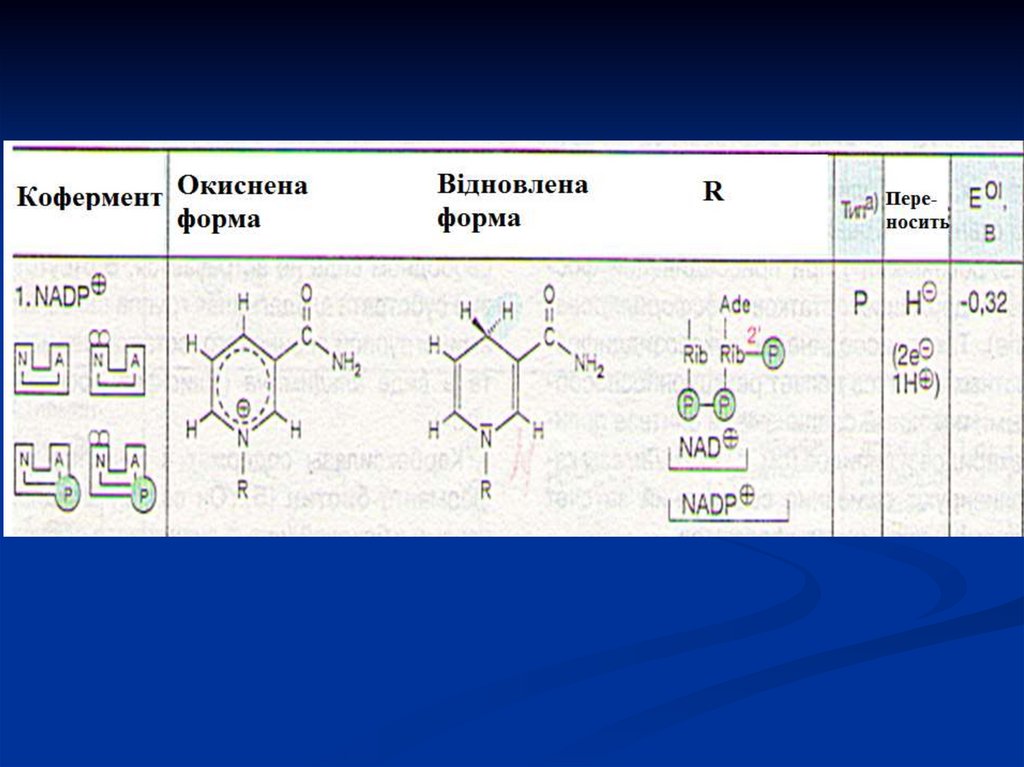
33. Піридинзалежні дегідрогенази (двохкомпонентні)
Специфічний білок (апофермент) +кофермент НАД або НАДФ.
Є універсальними акцепторами Гідрогену
для багатьох субстратів (спиртів, альдегідів
і т.д.).
Відбираючи від субстрату атоми Гідрогену,
вони
відновлюються, а субстрати
окиснюються.
34.
NAD - залежна дегідрогеназа каталізуєреакції окиснення безпосередньо субстрату
(α-кетоглутарат, ізоцитрат, ПВК, малат,
глутамат та ін.).
NAD+ є коферментом (утворюється в організмі
людини і тварин з вітаміну В5) і виконує роль
акцептора Гідрогену:
Н
НАДН2
H
НАД
ФМН
ФМНН2(в дых. цепь)
Субстрат
субстрат другой (восстанавл.)
35.
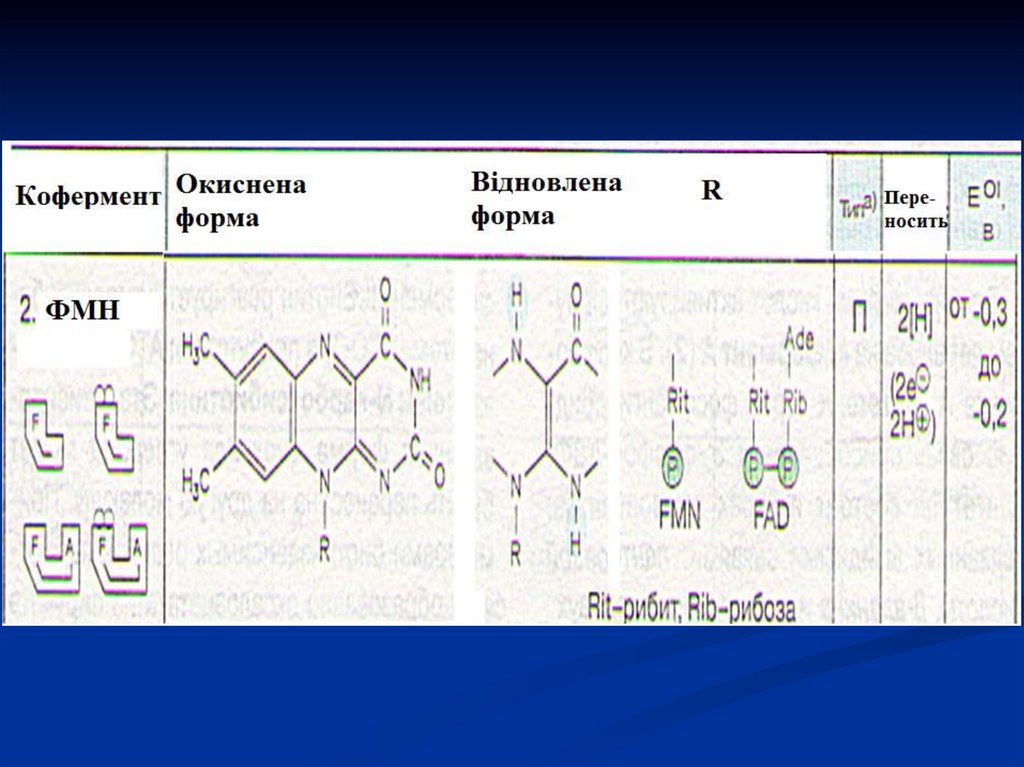
36. Флавінзалежні дегідрогенази (двохкомпонентні)
Специфічний білок (апофермент) +кофермент ФМН або ФАД.
Флавінові коферменти щільно зв'язані з
апоферментами (на відміну від піридинзалежних
дегідрогеназ).
Ці коферменти утворюються в організмі людини з
вітаміну В2.
Окиснюють відновлені субстрати (сукцинат,
ацил-КоА, α-гліцерофосфат).
37.
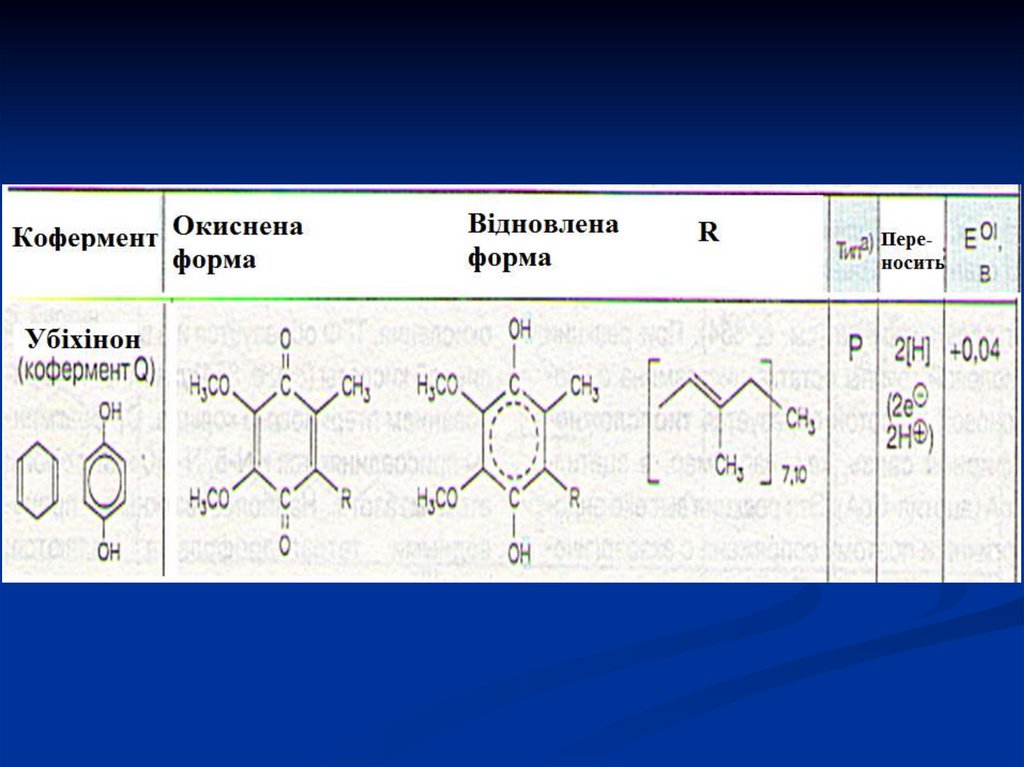
38. УБІХІНОН або коензим Q – проміжний переносник атомів Гідрогену, тобто електронів і протонів в мітохондріальній мембрані,
окиснює відновленуформу флавінових ферментів.
39.
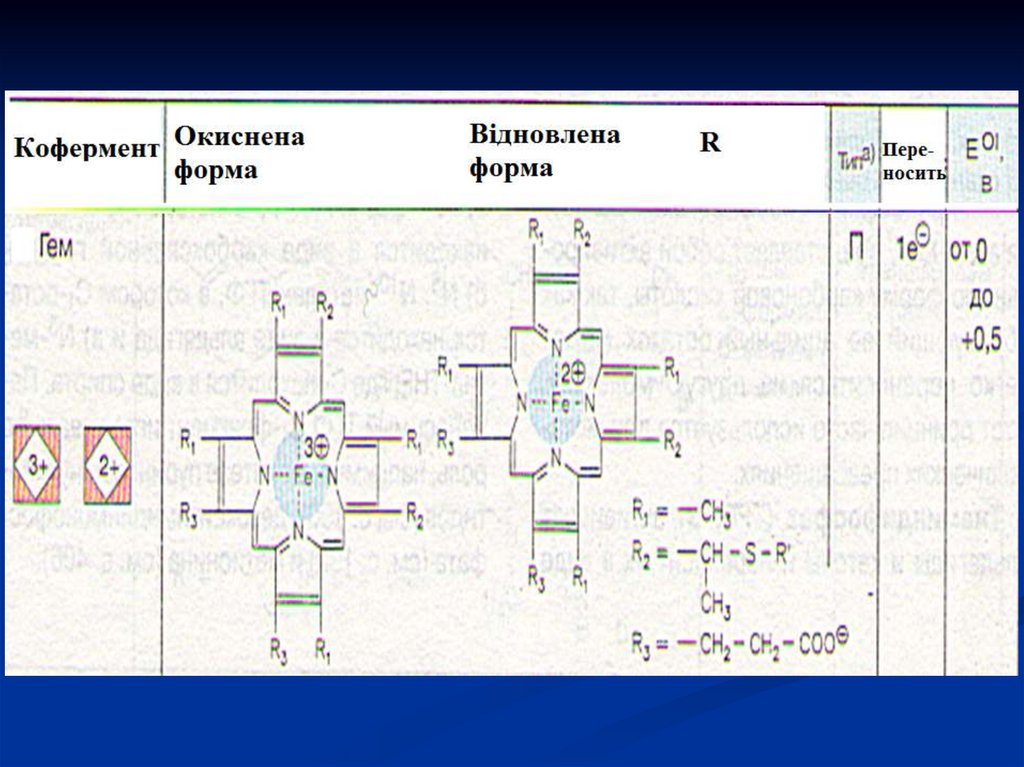
40. Цитохроми
Відносяться до гемвмісних хромопротеїнів. Їхпростетичною групою є гем, у якого атом заліза
має
здатність
легко
приєднувати
(відновлюватися) і віддавати (окиснюватися)
електрони.
41.
42.
В природі існують сімейства цитохромів (в, с, а),які відрізняються між собою величиною
електричного потенціалу, який збільшується в
напрямку в → с → аа3. В такому порядку
електрони в ланцюгу цитохромів послідовно
передаються від в до а3.
Цитохроми називають проміжними
переносниками електронів
(між дегідрогеназами і О2).
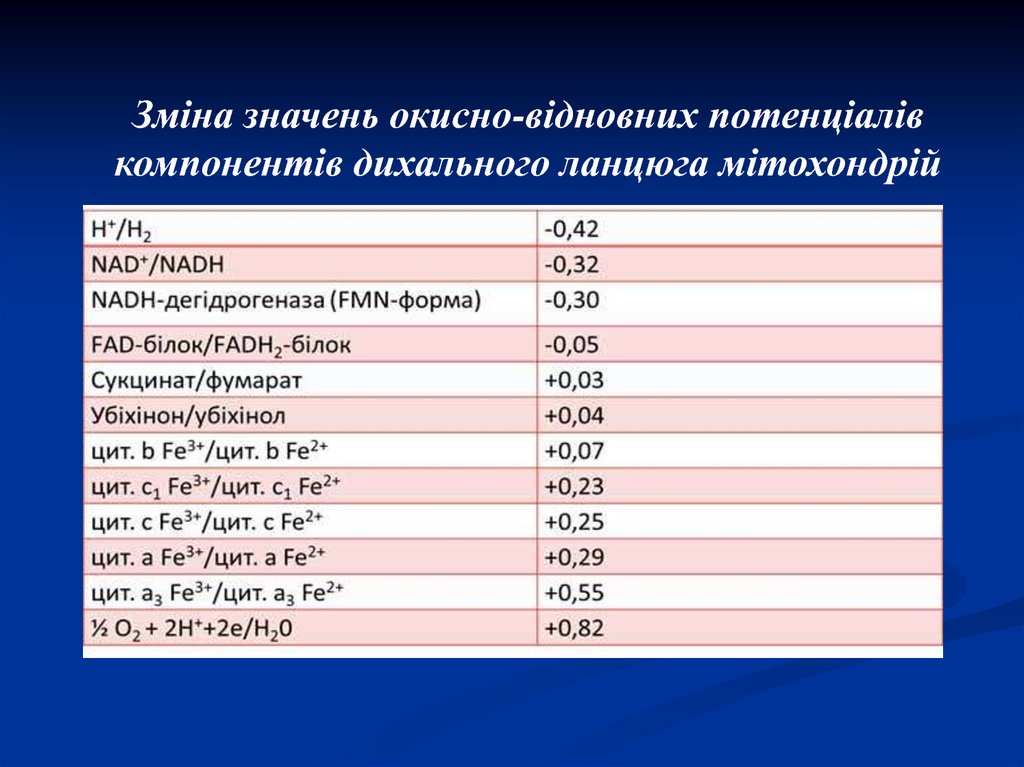
43. Редокс-потенціал
Напрямок потоку електронів приспряженні однієї окисної системи з
іншою визначається їх стандартними
окисно-відновними потенціалами або
редокс-потенціалами (Е0).
44.
Зміна значень окисно-відновних потенціалівкомпонентів дихального ланцюга мітохондрій
45. Ендогенна вода
Убіхінон передає на систему цитохромів тількиелектрони, а протони (Н+) надходять в
середовище, де взаємодіють з відновленим О2,
утворюється вода, що називається ендогенною:
2Н+ + 1⁄2О2 → Н2О.
Утворюється в результаті окиснення вуглеводів,
білків та ліпідів (100г ліпідів при окисненні дають
організму 107 г води).
Верблюд в пустелі використовує жирові
відкладення в горбах для одержання ендогенної
води.
46. Інгібітори дихального ланцюгу
Речовини, що блокують певні етапи цього процесу.Для НАД-залежної дегідрогенази:
- ротенон – високотоксична сполука, що міститься в
деяких водоростях та є отрутою для риб;
- амітал – лікарський препарат з групи барбітуратів.
Для убіхінону – антиміцин А, токсичний антибіотик.
Для цитохромоксидази – ціанід, СО, Н2S. Ціанід є
найбільш токсичним для людини, він приєднується
до Fe3+ цитохромоксидази і блокує перенесення
електронів до кисню.
47. Окисне фосфорилування
Синтез АТФ з АДФ відбувається врезультаті окисного фосфорилування, яке
сполучене з процесом біологічного
окиснення (тканинного дихання).
Цей процес був відкритий Енгельгардтом.
48.
49.
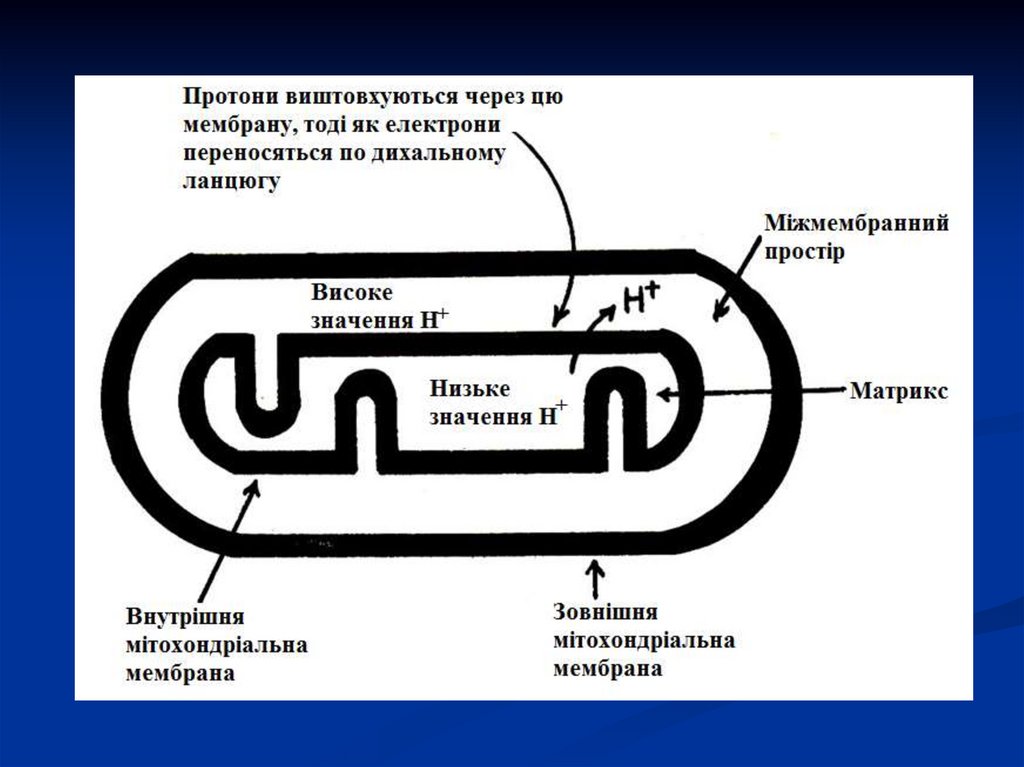
створюється протонний градієнт, при якомуконцентрація протонів у міжмембранному
просторі більше, а рН менше, ніж у матриксі.
з'являється різниця потенціалів по обидві
сторони мембрани: негативний заряд на
внутрішній стороні і позитивний – на зовнішній.
Разом
вони
утворюють
електрохімічний
потенціал - джерело енергії для синтезу АТФ.
50.
В дихальному ланцюгу можна виділити3 ділянки, в яких перенесення електронів
супроводжується зниженням вільної енергії.
Ці ділянки називають пунктами сполучення
дихання і фосфорилування.
Кількість енергії, що тут виділяється,
приблизно така, яка є необхідною для синтезу
АТФ з АДФ і фосфату:
АДФ + Н3РО4 ↔ АТФ
51.
Енергія електрохімічного потенціалувикористовується для синтезу АТФ, якщо
протони повертаються в матрикс через іонні
канали АТФ-синтази.
АТФ-синтаза (Н-АТФаза) – інтегральний білок
внутрішньої мембрани мітохондрій. Він
розташований безпосередньо поблизу
дихального ланцюга.
Підвищення концентрації протонів у
міжмембранному просторі активує АТФ-азу.
52. АТФ-синтаза
Складається з 2-х білкових комплексів.Гідрофобний
комплекс
занурений в мембрану. Він є
основою, яка фіксує АТФ-азу в
мембрані.
Складається
з
декількох субодиниць, що
утворюють канал, по якому
протони
переносяться
в
матрикс.
Другий комплекс виступає в
мітохондріальний матрикс. Він
складається з 9 субодиниць, в
яких і відбувається синтез
АТФ.
53.
Для синтезу 1 моля АТФ з АДФ необхідно7,5 ккал енергії, що є рівносильним різниці редокспотенціалів 0,15-0,22 V.
Така енергія вивільнюється при переході
електронів:
- від НАД до ФМН (1 комплекс),
- від цитохрому в до цитохрому с (3 комплекс),
- від цитохромоксидази аа3 до О2 (4 комплекс).
54. Мітохондріальний ланцюг перенесення електронів
55.
В НАД-залежних реакціях при окисненні 1 молясубстрату утворюється 3 АТФ,
у ФАД-залежних реакціях – 2 АТФ,
у випадку окиснення аскорбінової кислоти–1АТФ
Загальна кількість АТФ в організмі 30-50 г, але
кожна молекула АТФ в клітині може «жити»
менше хвилини.
Збільшення
концентрації
АДФ
негайно
приводить
до
прискорення
дихання
і
фосфорилування.
56. Роз'єднання дихання і фосфорилування
Роз'єднання дихання іфосфорилування
Деякі хімічні речовини (протонофори) можуть
переносити протони або інші іони (іонофори) з
міжмембранного простору через мембрану в
матрикс, минаючи протонні канали АТФсинтази.
В результаті зникає електрохімічний потенціал і
припиняється синтез АТФ.
Це явище називається роз'єднанням дихання і
фосфорилування.
57.
В результаті роз'єднання кількість АТФзменшується, а АДФ збільшується. Швидкість
окиснення НАДН і ФАДН2 підвищується,
збільшується
і
кількість
кисню,
що
поглинається, але енергія виділяється тільки у
вигляді теплоти.
Приклади роз'єднуючих агентів:
дикумарол (антикоагулянт);
білірубін (метаболіт гему);
тироксин (гормон щитоподібної залози).
Всі вони проявляють свою дію тільки за умови
високої концентрації.
58. МІКРОСОМАЛЬНЕ ОКИСНЕННЯ
Це ще один тип біологічного окиснення, якийвідбувається в мікросомах (переважно в
печінці і надниркових залозах) і пов'язаний з
безпосереднім включенням Оксигену в
сполуку, яка окиснюється.
В мікросомальному окисненні, на відміну від
мітохондріального, Оксиген використовується
не з енергетичною, а з «пластичною» метою.
59.
До ферментів мікросомального окисненнявідтносяться МОНООКСИГЕНАЗИ і
ДИОКСИГЕНАЗИ.
Монооксигенази (гідроксилази) при окисненні
речовини включають 1 атом Оксигену.
Диоксигенази каталізують реакції окиснення з
включенням в молекулу 2 атомів молекули
кисню.
60. ФУНКЦІЇ МІКРОСОМАЛЬНОГО ОКИСНЕННЯ:
Окиснення речовин циклічної структури, які неможуть окиснюватися шляхом дегідрування
(триптофан, гомогентизинова кислота та ін.);
Синтез стеринів (холестерин, жовчні кислоти,
гормони кіркової речовини надниркових залоз,
статеві гормони);
Метаболізм лікарських препаратів, а також
знезараження токсичних речовин.
61. Утворення пероксиду водню.
При тканинному диханні можливеутворення пероксиду водню (Н2О2), який є
сильним окисником сульфгідрильних груп
ферментів. Він токсичний для клітин.
В процесі еволюції утворився ряд
ферментів, які його розщеплюють, в тому
числі КАТАЛАЗА, що міститься в
еритроцитах, в печінці та інших тканинах.
62.
Каталаза:каталаза
2H 2O 2
2H 2O + O 2
Пероксидаза:
OH
O
пероксидаза
+ H 2O
+ H2O2
OH
O