




 мышц")


 тип дыхания")
 тип дыхания")


































")














































 biology
biologySimilar presentations:










Физиология дыхания
1. Физиология дыхания
Общие принципы2. Дыхание
Многоэтапный процесс переноса О2из атмосферного воздуха к клеткам
организма
и
СО2
от
клеток
в
окружающую среду. Он складывается
из:
- обмена между атмосферным воздухом и альвеолами —
легочная вентиляция;.
- обмена между альвеолами и кровью — легочная диффузия;
- переноса между легочными и системными капиллярами (к
тканям и обратно) — транспорт кровью;
- обмена между системными капиллярами и клетками —
тканевая диффузия.
Первые две стадии - внешнее дыхание, последние две —
3. Система дыхания
Система дыхания- комплекс структур,
которые
принимают
участие в газообмене,
и
механизмы
их
регуляции.
Есть
два
вида
дыхания: внешнее и
внутреннее.
4. Внешнее дыхание - это обмен газов между внешней средой и альвеолами
Внешнее дыхание – это совокупность процессов,происходящих
в
легких
и
обеспечивающих
нормальное содержание в крови О2 и СО2
В легких происходит 3 основных процесса:
– вентиляция
– диффузия
– перфузия
Чередование дыхательных движений (вдоха и
выдоха)
называется
дыхательным
циклом
(соотношение 1:1,2).
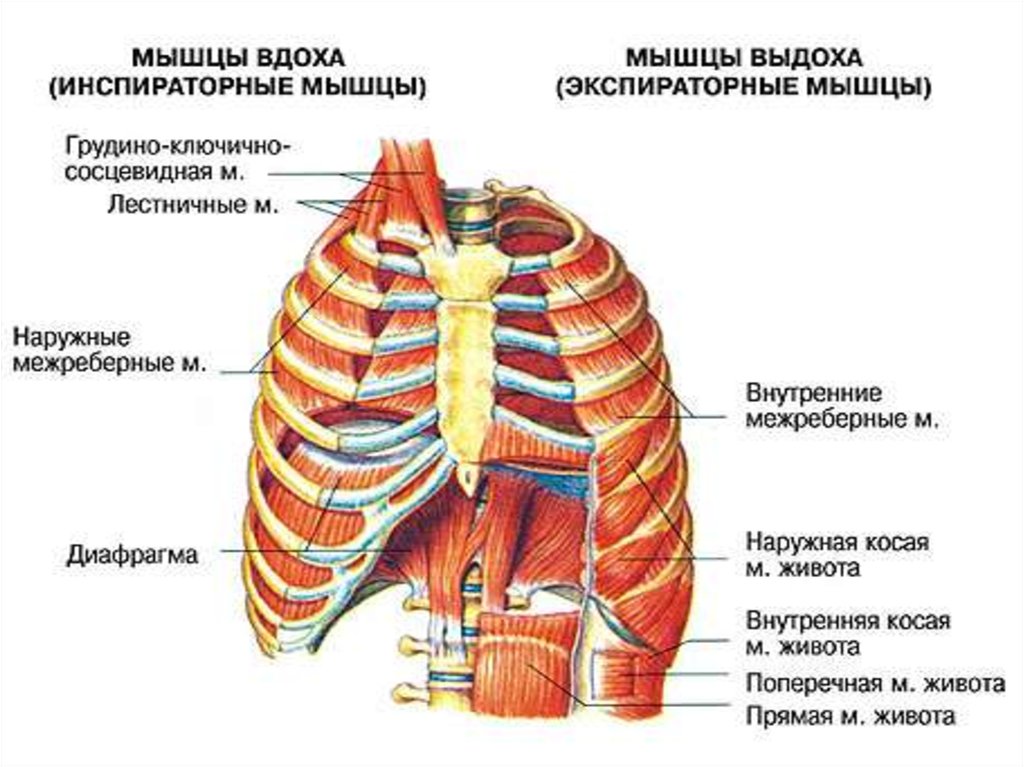
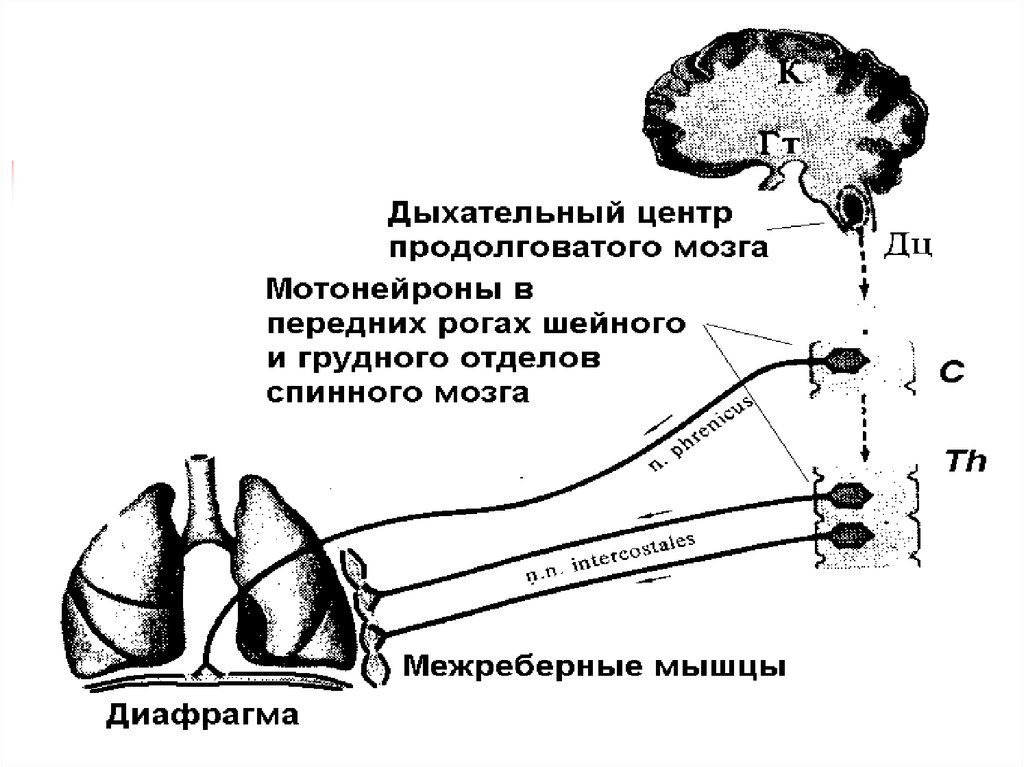
5. Основные и вспомогательные дыхательные мышцы
К основным относят диафрагму и межреберныемышцы, обеспечивающие вентиляцию легких в
физиологических условиях.
К вспомогательным относятся мышцы шеи,
часть мышц верхнего плечевого пояса, мышцы,
брюшного пресса, принимающие участие в
форсированном
вдохе
или
выдохе
в
обстоятельствах,
затрудняющих
вентиляцию
легких.
6. Вдох начинается с сокращения дыхательных (респираторных) мышц
Мышцы,сокращение
которых
приводит
к
увеличению объема грудной полости инспираторные
Мышцы,
сокращение
которых
приводит
к
уменьшению объема грудной полости экспираторные.
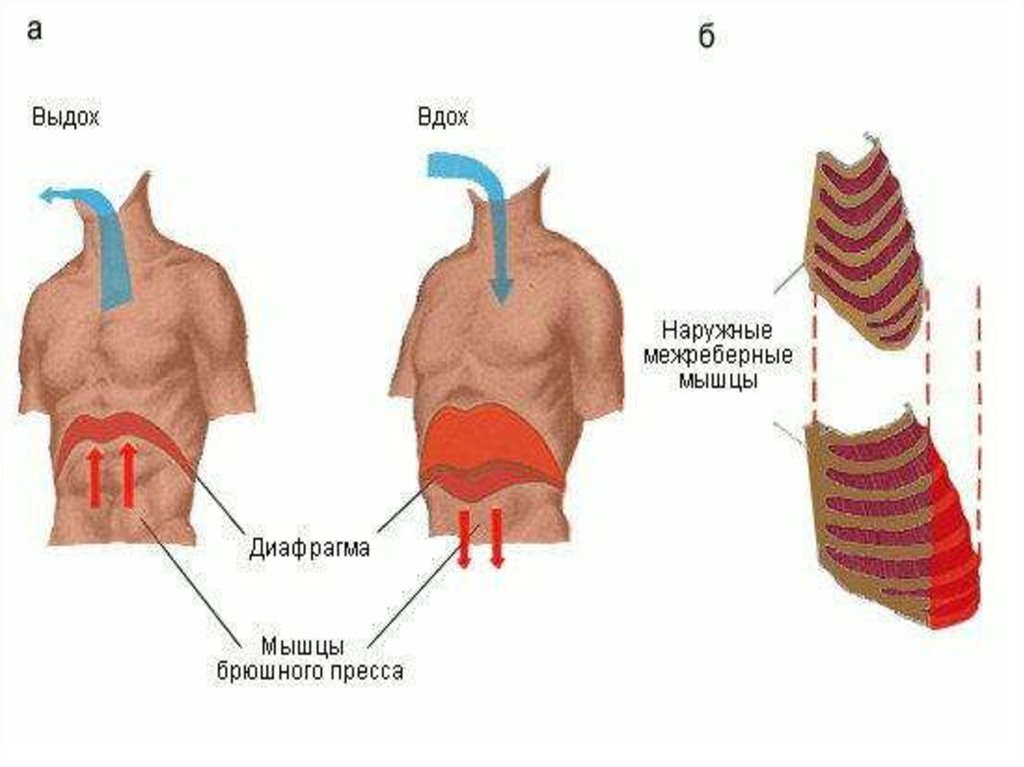
Основная инспираторная мышца - диафрагма. При ее
сокращении купол диафрагмы уплощается, внутренние
органы оттесняются вниз, увеличение объема грудной
полости происходит вертикальном направлении.
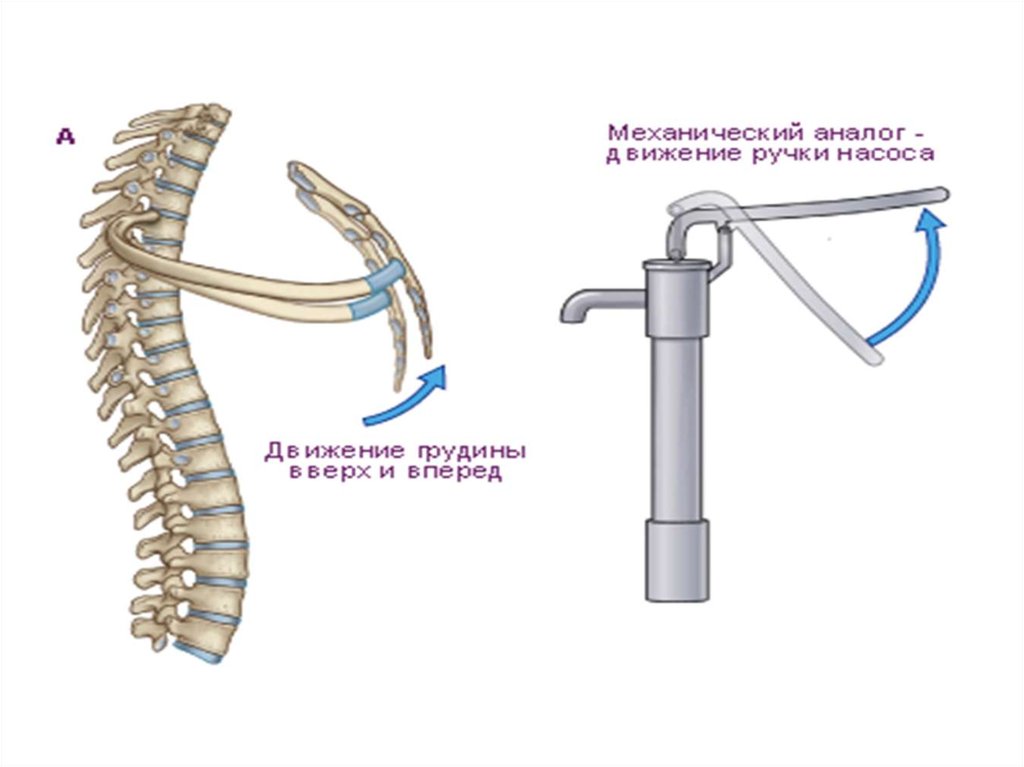
Сокращение наружных межреберных и межхрящевых
мышц приводит к увеличению объема грудной полости
в сагиттальном и фронтальном направлениях.
7.
8.
9.
10.
В процессе работы дыхательные мышцыпреодолевают
сопротивление,
2/3
которого
приходится на эластическое сопротивление тканей
легких, которое создается за счет поверхностно—
активных веществ — сурфактантов, тонким слоем
выстилающих изнутри альвеолы.
Сурфактанты
состоят из липопротеинов. Они
стабилизируют сферическую форму альвеол,
препятствуя их перерастяжению на вдохе и
спадению на выдохе.
11. Для нормальной легочной вентиляции необходимо структурное обеспечение
работа дыхательных мышц для измененияразмеров грудной клетки
эластичность легочной ткани, которая позволяет
ей следовать за изменениями размеров грудной
клетки
транспульмональное
давление,
которое
поддерживает легкие в расправленном состоянии
легочный
сурфактант,
препятствующий
спадению альвеол
12.
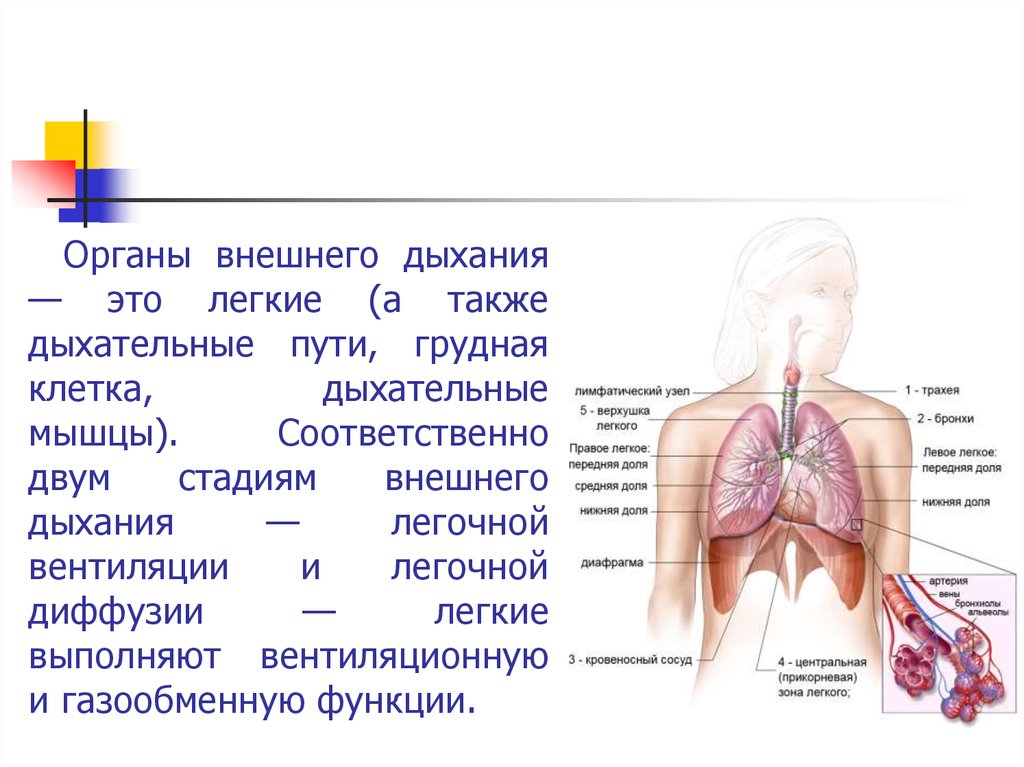
Органы внешнего дыхания— это легкие (а также
дыхательные пути, грудная
клетка,
дыхательные
мышцы).
Соответственно
двум
стадиям
внешнего
дыхания
—
легочной
вентиляции
и
легочной
диффузии
—
легкие
выполняют вентиляционную
и газообменную функции.
13. Типы дыхания Грудной (реберный) тип дыхания
Чаще встречается у женщин, дыхательныедвижения осуществляются за счет сокращения
межреберных мышц, а грудная клетка расширяется
и слегка приподнимается во время вдоха,
суживается и несколько опускается при выдохе
14. Брюшной (диафрагмальныйм) тип дыхания
Чаще встречающемся у мужчин, дыхательныедвижения
осуществляются
преимущественно
диафрагмой, которая во время вдоха сокращается
и опускается, это увеличивает отрицательное
давление в грудной полости, легкие заполняются
воздухом. Внутрибрюшное давление повышается и
брюшная стенка выпячивается. Во время выдоха
диафрагма расслабляется, поднимается, брюшная
стенка возвращается в исходное положение.
15. Смешанный тип
В акте дыхания участвуют межреберные мышцы идиафрагма
16. Ацинус У взрослого 150 000 ацинусов суммарная площадь 80 м2
Легочныйацинус
—
это
функциональная
единица
легких,
вентилируемая
конечной
бронхиолой,
от
которой отходят дыхательные
бронхиолы,
образующие
альвеолярные
каналы
или альвеолярные ходы. В
конце каждого альвеолярного
канала находятся альвеолы
17.
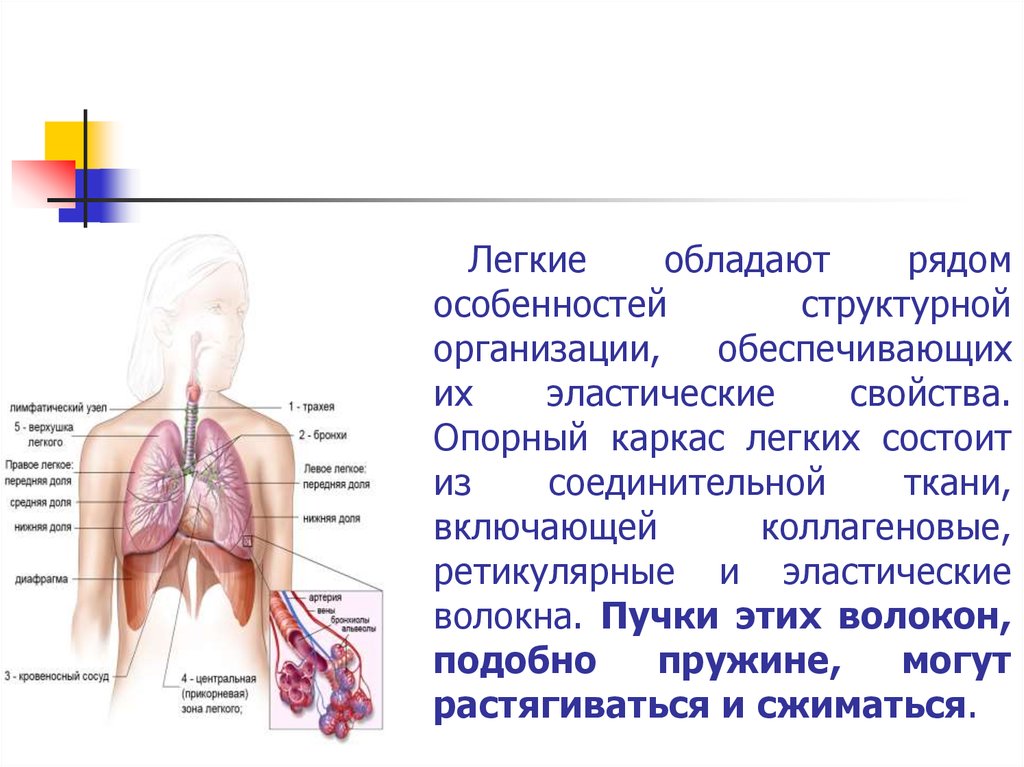
Легкиеобладают
рядом
особенностей
структурной
организации,
обеспечивающих
их
эластические
свойства.
Опорный каркас легких состоит
из
соединительной
ткани,
включающей
коллагеновые,
ретикулярные и эластические
волокна. Пучки этих волокон,
подобно
пружине,
могут
растягиваться и сжиматься.
18.
Таким образом, легкие содержат структуры,которые, с одной стороны, эластичны и могут
растягиваться, а с другой - обладают выраженной
способностью к ретракции. Во время вдоха легкие
подвергаются растяжению под действием сил
сокращения дыхательной мускулатуры (размер
грудной клетки увеличивается). Когда эти силы
прекращают действовать, легкие благодаря своим
упругим
свойствам
возвращаются
в
первоначальное состояние
19. Легочный сурфактант
Пленка жидкости, выстилающая внутреннююповерхность
альвеол,
содержит
вещество,
понижающее поверхностное натяжение. Это
вещество называется сурфактант, который
обеспечивает
во-первых,
повышение
растяжимости легких и уменьшении работы,
совершаемой
во
время
вдоха,
во-вторых,
обеспечивает стабильности альвеол препятствуя
их слипанию.
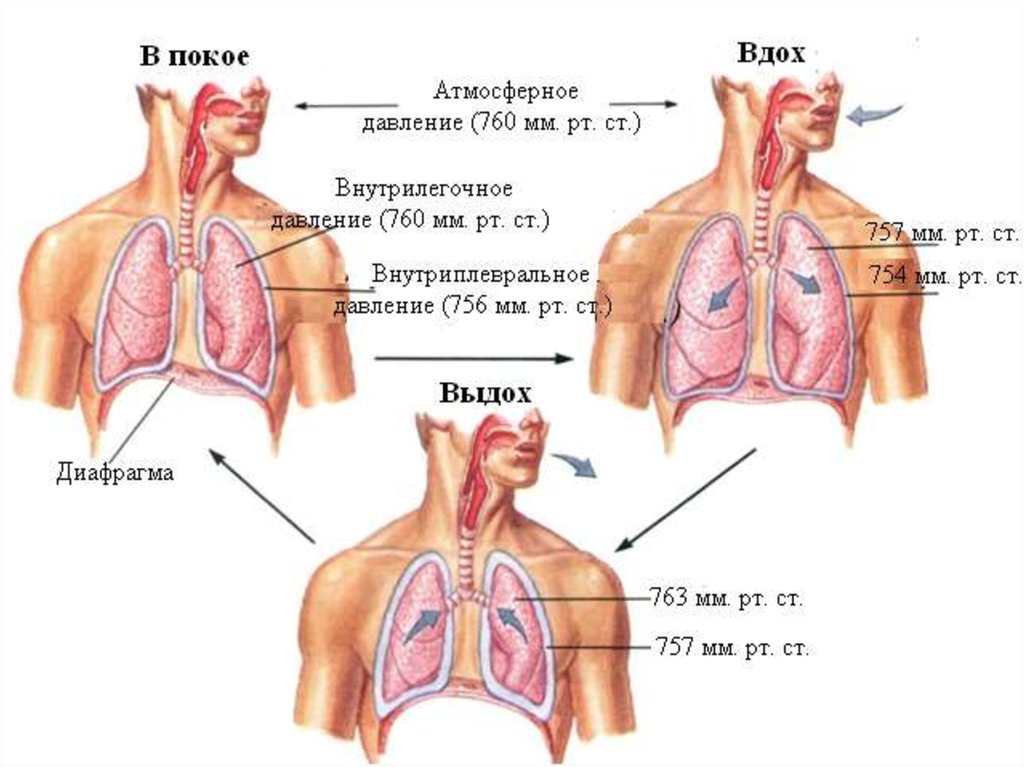
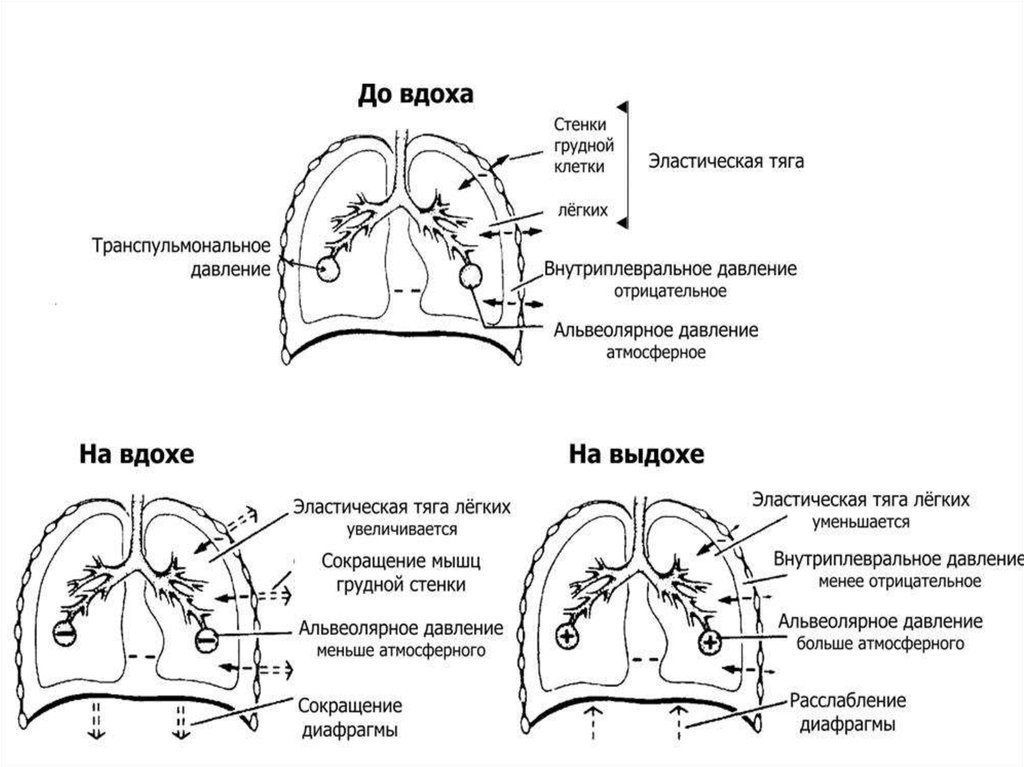
20. Транспульмональное давление
Растянутыелегкие
постоянно
стремятся
уменьшить свой размер за счет способности
эластических волокон к ретракции.
Эта сила
эластической тяги легких постоянно «оттягивает»
легкие от грудной клетки, поэтому давление в
плевральной полости всегда немного ниже, чем
давление в альвеолах. У человека в состоянии
покоя в конце выдоха внутриплевральное
давление примерно на 3-4 мм рт. столба (5см.
водного столба) ниже атмосферного.
21. Плевральное давление
Внутриплевральное давление ниже давления вальвеолах на величину эластической тяги легких:
Р плевр. = Р альв. - Р эласт. тяги легких
Т.о., между внутренней поверхностью альвеол и
плевральной полостью существует разность
давлений,
которая
всегда
в
пользу
альвеолярного пространства
22. Транспульмональное давление
Разницу между давлением в альвеолах идавлением в плевральной полости называют
транспульмональным давлением.
Р транспульм. = Ральв. - Р плевр.
Транспульмональное давление это тот
градиент давлений, который поддерживает
легкие
в
расправленном
состоянии
(давление
«изнутри»
выше
давления
«снаружи»).
23.
24. Движение диафрагмы и грудной клетки
Легкие покрыты серозной оболочкой - плеврой,состоящей из висцерального и париетального листков.
Париетальный листок соединен с грудной клеткой, а
висцеральный - с тканью легких. С увеличением
объема грудной клетки, в результате сокращения
инспираторных мышц, париетальный листок следует за
грудной клеткой. Появляются адгезивные силы между
листками плевры, висцеральный листок следует за
париетальным, а вслед за ними и легкие. Возрастает
отрицательного давления в плевральной полости и
увеличивается объем легких, что сопровождается
снижением в них давления, оно становится ниже
атмосферного и воздух начинает поступать в легкие происходит вдох.
25. Механизм вдоха
Сигнализация о дыхательной потребности организмареализуется возбуждением ДЦ. Далее возбуждение через
спинномозговые центры (диафрагмальное ядро и ядра
межреберных мышц) поступает к дыхательной мускулатуре
(диафрагме и межреберным мышцам), в результате
происходит сокращение дыхательной мускулатуры и
увеличение объема грудной клетки в вертикальном,
фронтальном и сагиттальном направлениях. Париетальный
листок плевры, следуя за грудной клеткой и диафрагмой и
удаляется от висцеральной. Объем плевральной полости
увеличивается
26.
Иннервация органов дыхания27. Механизм вдоха
Внутриплевральноедавление
снижается,
а
транспульмональное становится преобладающим.
С увеличением объема легких, транспульмональное
давление снижается. Нарушается соотношение между
атмосферным
и
транспульмональным
давлением.
Преобладающим становится атмосферное давление. В
результате возникшего градиента давлений происходит
заполнение легких воздухом. Обратная афферентация о
количестве
поступающего
в
легкие
воздуха
осуществляется
на
основе
легочно-вагусной
афферентации.
28. Механизм вдоха
Обратная афферентация возникает в механорецепторахальвеол и по блуждающим нервам направляется в ДЦ.
Легочно-вагусная афферентация прекращает вдох, как
только в легкие поступило необходимое количество
воздуха.
В
зависимости от условий пребывания организма –
особенностей газовой среды, активности мышечной
работы,
эмоционального
состояния
–
количество
вдыхаемого воздуха будет различным, но всегда легочная
афферентация будет прекращать вдох в тот самый момент,
когда в легкие поступит необходимое количество воздуха.
29. Механизм выдоха
Выдох наступает в результате подавления активностиинспираторного отдела.
Нисходящая импульсация к дыхательной мускулатуре
уменьшается.
Мышцы расслабляются.
Ребра, грудина и диафрагма занимают первоначальное
положение (в силу тяжести и давления брюшных органов
на диафрагму).
Объем грудной клетки уменьшается.
Париетальный листок плевры приближается к
висцеральному.
30. Механизм выдоха
Объем плевральной полости уменьшается.Внутриплевральное
давление
увеличивается
и
становится больше транспульмонального.
Объем легких в результате уменьшается.
Транспульмональное
давление
увеличивается
и
становится больше атмосферного.
В силу возникшего градиента давлений воздух покидает
легкие, т.е. осуществляется выдох. Выдох при спокойном
дыхании осуществляется пассивно т.к. грудная клетка и
легкие после вдоха стремятся занять положение, из
которого они были выведены сокращением дыхательных
мышц.
31.
32. Пневмоторакс
Сообщение плевральной полости с внешнейсредой в результате нарушения герметичности
грудной клетки называется пневмоторакс. При
пневмотораксе выравниваются внутриплевральное
и атмосферное давления, вызывающее спадение
легкого, что делает невозможной его вентиляцию
при дыхательных движениях грудной клетки и
диафрагмы.
33. Пневмоторакс
34. Легочные объемы
дыхательный объем: объем воздуха,вдыхаемый (или выдыхаемый) при одном
вдохе (выдохе). В норме при спокойном
дыхании — до 500 мл;
резервный объем вдоха: объем воздуха,
который можно дополнительно вдохнуть
после спокойного вдоха. В норме — 2000 3000 мл;
резервный
объем
выдоха:
объем
воздуха, который можно дополнительно
выдохнуть после спокойного выдоха. В норме
— около 1000-1500 мл;
остаточный
объем
легких:
объем
воздуха, который остается в легких после
максимального выдоха. В норме — около
1000-1500 мл.
35. Легочные емкости
жизненная емкость легких:объем воздуха, который можно
выдохнуть после максимального
вдоха;
сумма
дыхательного
объема, резервного объема вдоха
и резервного объема выдоха. В
норме — 3000-4500 мл;
общая емкость легких: объем
воздуха, содержащийся в легких
на высоте максимального вдоха;
сумма жизненной емкости легких
и остаточного объема легких. В
норме — 4000—6000 мл;
36. Легочные емкости
функциональная остаточнаяемкость:
объем
воздуха,
содержащийся в легких после
спокойного
выдоха;
сумма
резервного объема выдоха и
остаточного объема легких. В
норме — 2000—3000 мл;
емкость вдоха: объем воздуха,
который можно вдохнуть после
спокойного
выдоха;
сумма
дыхательного
объема
и
резервного объема вдоха. В норме
— 2500-3500 мл.
37. Спирограмма
По вертикальной оси: объем легких, мл. ДО — дыхательныйобъем; Евд — емкость вдоха; ЖЕЛ — жизненная емкость легких;
ОЕЛ — общая емкость легких; ООЛ — остаточный объем легких;
РОвд — резервный объем вдоха; РОвыд — резервный объем
выдоха; ФОЕ — функциональная остаточная емкость
38.
Приведенныеколичественные
значения
варьируют и зависят от пола, возраста, роста и
других факторов.
Остаточный объем легких (а следовательно, и те
легочные емкости, в которые он входит ─
функциональную остаточную емкость и общую
емкость легких) нельзя измерить при спирометрии.
Эти показатели определяются другими методами.
39. Мертвое пространство
В легких можно выделить два отдела:альвеолярное пространство — все отделы
легких, в которых идет газообмен (альвеолы и
альвеолярные ходы);
мертвое пространство — все дыхательные
пути, в которых не идет газообмен (верхние
дыхательные пути, трахея, бронхи и бронхиолы
вплоть до терминальных бронхиол).
40. Мертвое пространство
Объем мертвого пространства составляет около150 мл. Таким образом, при каждом вдохе 150 мл
поступающего воздуха не участвуют в газообмене.
41. Физиологическое значение мертвого пространства
Соотношение между объемом мертвого пространстваи дыхательным объемом — это один из факторов,
определяющих эффективность дыхания.
Если объем мертвого пространства = 150 мл, а
дыхательный объем = 500 мл, то до альвеол доходит (и
участвует в газообмене) две трети дыхательного
объема; если же дыхательный объем = 350 мл, то до
альвеол дойдет всего половина. Таким образом,
при частоте дыхания 12 раз в минуту
альвеолярная вентиляция равна 12 х (500 150), или 4200 мл/мин
42. Диагностическое значение мертвого пространства
Некоторыеальвеолы
вентилируются,
но
газообмен в них не идет (они не снабжаются
кровью); объем мертвого пространства возрастает.
В связи с этим выделяют анатомическое
мертвое пространство (дыхательные пути) и
функциональное
мертвое
пространство
(анатомическое мертвое пространство + альвеолы,
в которых идет вентиляция, но не газообмен).
43. Показатели вентиляции
Основных таких показателей три.Минутный объем дыхания (МОД) - объем воздуха,
проходящий через легкие за минуту. В покое МОД
составляет 6—8 л/мин.
Максимальная вентиляция легких (МВЛ) - объем
воздуха, который мог бы пройти через легкие за минуту
при максимально интенсивном (частом и глубоком)
дыхании.
Альвеолярная вентиляция (АВ) - объем воздуха,
проходящий через альвеолярное пространство за минуту.
44. Характеристика показателей
МВЛотражает
максимальные
резервы
дыхательной системы, которые никогда не
используются: МВЛ может достигать 180 л/мин, а
МОД даже при самой интенсивной нагрузке не
превышает 100 л/мин. Чтобы рассчитать МВЛ,
человека просят дышать максимально интенсивно в
течение 10 с и полученный объем воздуха
перерассчитывают на 1 мин.
45. Характеристика показателей
МОД всегда больше АВ, так как часть МОДидет на вентиляцию мертвого пространства.
Для газообмена важнее величина АВ, но
работа дыхательных мышц зависит от МОД.
46.
Динамическиепоказатели
связаны
со
статическими - с дыхательным объемом и объемом
мертвого пространства, через частоту дыхания:
МОД=ДО х ЧД, (I)
АВ=(ДО-ОМП) х ЧД, (2)
где ДО - дыхательный объем; АВ – альвеолярная
вентиляция; ОМП - объем мертвого пространства;
ЧД - частота дыхания.
47. Соотношение вентиляции и перфузии легких
Для нормального процесса обмена газов влегочных альвеолах необходимо, чтобы их
вентиляция
воздухом
находилась
в
определенном соотношении с перфузией их
капилляров кровью.
Минутному
объему
дыхания
должен
соответствовать минутный объем крови
48. Соотношение вентиляции и перфузии легких
Вентиляционно—перфузионный коэффициенту здорового человека составляет 0,8—0,9.
Если альвеолярная вентиляция 6 л/мин, то
минутный объем крови может составить около 7
л/мин.
В отдельных областях легких соотношение
между вентиляцией и перфузией может быть
неравномерным - это либо патология, либо
предпатология
49. Диффузия газов через аэрогематический барьер
В организме газообмен О2 иСО2,
а
так
же
других
газообразов
происходит
с
помощью
диффузии
через
альвеолокапиллярную мембрану
легких в два этапа. На первомдиффузионный перенос газов
происходит
по
концентрационному
градиенту
через
аэрогематический
барьер
(1мкм).
На
втором
этапе
происходит связывание газов в
крови легочных капилляров.
50. Закон Фика
Диффузия газов осуществляется в соответствии сградиентом парциальных давлений газов и
описывается законом Фика:
51. Закон Фика
Согласно этому закону газообмен (M/t) в легкихпрямо
пропорционален
градиенту
(ΔР)
концентрации 02 и С02 по обе стороны от
альвеолярной мембраны, площади ее поверхности
(S), коэффициентам (k) растворимости 02 и С02 в
биологических средах альвеолярной мембраны и
обратно пропорционален толщине альвеолярной
мембраны (L), а также молекулярной массе газов
(М).
52.
С02 примерно в 20 раз более растворим вальвеолярной мембране, чем 02.
Поэтому,
несмотря на различие в
градиентах парциальных давлений 02 и С02
по обе стороны от альвеолярной мембраны,
диффузия этих газов совершается за очень
короткий
отрезок
времени
движения
эритроцитов
крови
через
легочные
капилляры.
53. Состав атмосферного, альвеолярного и выдыхаемого воздуха (%, парциальное давление в мм рт.ст.)
ГазАтмосферный, Альвеолярный, Выдыхаемый,
%, мм. рт. ст.
%, мм. рт. ст.
%, мм. рт. ст.
О2
20,85 (160)
13,5 (104)
15,5 (120)
СО2
0,03 (0,2)
5,3 (40)
3,7 (27)
N2
78,62 (596)
74,9 (569)
74,6 (566)
Н2О
0,5 (3.8)
6,3 (47)
6,,2 (47)
Общий
100 (760)
100 (760)
100 (760)
54. Диффузия О2 из альвеол в капиллярную кровь легких
Ро2 в альвеолярной газовойсмеси - 104 мм рт. ст., а Ро2
в венозной крови, входящей
в легочный капилляр через
его артериальный конец 40 мм рт. ст. Т.о. разница в
парциальном давлении,
составляет 104 - 40, или 64
мм рт. ст. – это причина
диффузии
55. Диффузия О2 из периферических капилляров в тканевую жидкость
Po2 в интерстиции 40 мм рт. ст., вклетках ткани - 23
мм рт. ст.
В артериальной крови Ро2 составляет 95 мм рт. ст. В
окружающей тканевые клетки интерстициальной
жидкости Ро2 всего 40 мм рт. ст. По градиенту
давления О2 диффундирует из капиллярной крови в
интерстиций
и ткани. Поэтому Ро2 в крови,
покидающей капилляры и поступающей в системные
вены, составляет около 40 мм рт. ст.
56.
О2 в клетках превращается в СО2, в результате вклетке повышается Рсо2,которая диффундирует из
клеток в тканевые капилляры и переносится кровью
в легкие и выводится. В отличие от О2 скорость
диффузии СО2 в 20 раз выше, поэтому необходим
меньший градиент давления - 5 мм рт. ст.
57. Транспорт двуокиси углерода в растворенном виде
Оъем СО2, растворенный в жидкой части крови,равен примерно 2,7 мл/дл (2,7 об%). Для
выделения
в
легких
транспортируется
в
растворенном виде только 0,3 мл СО2 в 100 мл
крови. Это составляет около 7% всего объема,
транспортируемого
кровью
в
нормальных
условиях.
58. Транспорт двуокиси углерода в виде иона бикарбоната
Карбоангидразаэритроцитов катализирует реакцию
между растворенным в крови
СО2 и водой, образуя Н2СО3,
которая диссоциирует на
ионы
водорода
и
бикарбоната (H+ и НСОз-).
59. Транспорт двуокиси углерода в виде иона бикарбоната
Ионы H+ присоединяются вэритроцитах к гемоглобину.
Ионы НСОз- диффундируют из
эритроцитов в плазму, откуда в
эритроцит возвращаются ионы
Сl-.
Это
обеспечивается
переносчиком ионов НСОз- и
Сl- в мембране эритроцитов.
Около 70% СО2 из тканей в
легкие выводится по этому
механизму.
60. Транспорт двуокиси углерода в связи с гемоглобином
Часть углекислого газа соединяется вэритроцитах с дезоксигемоглобином через
аминогруппы,
образуя
карбаминовые
соединения. Реакция протекает следующим
образом:
HbNH2+ CO2 HbNHCOOH HbNHCOO- + H+
61. Дыхательный центр
Дыхательный центр состоит из 3 групп нейронов,расположенных в стволе мозга по обе стороны
продолговатого мозга и моста:
- дорсальная
группа дыхательных нейронов в
дорсальной части продолговатого мозга, вызывает
вдох;
- вентральная группа дыхательных нейронов
продолговатого мозга, вызывает выдох;
- пневмотаксический центр, расположен дорсально в
верхней части моста, контролирует скорость и
глубину дыхания.
62. Дыхательный центр
63. Дорсальная группа дыхательных нейронов
Большинство этих нейронов расположено в ядреодиночного тракта. Ядро одиночного тракта
является сенсорным ядром для блуждающего и
языкоглоточного нервов, которые передают в
дыхательный центр сенсорные сигналы от:
- периферических хеморецепторов;
- барорецепторов;
- разного типа рецепторов легких
64. Дорсальная группа дыхательных нейронов
Базовый ритм дыхания генерируется в основномдорсальной группой дыхательных нейронов. Даже
после перерезки всех входящих в продолговатый
мозг периферических нервов и ствола мозга ниже
и выше продолговатого мозга эта группа
нейронов
продолжает
генерировать
повторяющиеся залпы потенциалов действия
инспираторных нейронов.
65. Дорсальная группа дыхательных нейронов
Большая часть аксонов нейронов дорсальнойгруппы направляется в спинной мозг к
диафрагмальным
ядрам,
расположенным
в
шейном
отделе.
Часть
аксонов
отдают
коллатерали в вентральное ядро
66. Вентральная группа дыхательных нейронов
Вентральное дыхательноеядро
расположено
от
каудального
края
ядра
лицевого нерва до первого
шейного сегмента спинного
мозга. В нем выделяют три
скопления
дыхательных
нейронов.
67. Вентральная группа дыхательных нейронов
Рядом с ядром лицевого нерванаходятся скопления экспираторных нейронов (комплекс
Бетцингера).
2. Каудальнее
расположено
параамбигуальное
ядро,
в
которое входит обоюдное ядро,
в нем находятся мотонейроны
мышц гортани и глотки. Здесь
же
расположено
скопление
инспираторных нейронов
1.
68. Вентральная группа дыхательных нейронов
Частьвентрального
ядра,
расположенная латеральнее и
каудальнее обоюдного ядра,
называется ретроамбигуальным
ядром, здесь располагаются
экспираторные нейроны. Таким
образом,
вентральное
ядро
имеет сложную структуру и
включает как инспираторные,
так и экспираторные нейроны.
69. Вентральная группа дыхательных нейронов
Нейронывентральной
группы
посылают
нисходящие волокна к спинномозговым мотонейронам межреберных мышц и мышц живота. В
вентральной
группе
нейронов
находятся
эфферентные
преганглионарные
волокна
блуждающего нерва, которые обеспечивают
изменение
просвета
дыхательных
путей.
Максимальная активность нейронов блуждающего
нерва соответствует концу выдоха, минимальная
концу вдоха.
70. Пневмотаксический центр
В экспериментах с перерезкой мозга нижеваролиева моста установлно, что в регуляции
дыхания принимают участие и дыхательные
нейроны, расположенные в районе моста.
После перерезки и отделения продолговатого
мозга от моста периодические дыхательные
движения сохраняются, но ритм становится
другим:
длительный
выдох
периодически
прерывается короткими вздохами
71. Пневмотаксический центр
Для ритмичного дыхания и равномерной сменыдыхательных
фаз
необходима
целостность
нейронов,
расположенных
в
медиальном
парабрахиальном ядре и ядре Келликера-Фузе и их
связь с дыхательными нейронами продолговатого
мозга. Этот отдел моста получил название
пневмотаксического центра.
72. Механорецепторы
Дыхательный центр получает информацию отмеханорецепторов - от чувствительных нервных
окончаний, расположенных в гладких мышцах, в
подслизистом слое и эпителии воздухоносных
путей. Выделяют три основные вида рецепторов:
1.
рецепторы растяжения легких
2.
ирритантные рецепторы
3.
J - рецепторы - юкстакапиллярные рецепторы
легких
73. Рецепторы растяжения легких
находятся в гладких мышцах воздухоносныхпутей. В каждом легком около 100 таких
рецепторов, они связаны с дыхательным
центром толстыми миелинизированными
волокнами
блуждающего
нерва
с
высокой (40м/сек) скоростью проведения
74. Рецепторы растяжения легких
Возбуждениерецепторов
возникает
или
усиливается при увеличении объема легких.
Рецепторы
обладают
разными
порогами
возбуждения. Часть рецепторов обладает низким
порогом – они возбуждены в течение выдоха и
даже при спадении легких, во время вдоха частота
импульсов резко возрастает. Высокопороговые
рецепторы возбуждаются только при увеличении
объема легких сверх функциональной остаточной
емкости
75. Рецепторы растяжения легких
Способностьрецепторов
растяжения
возбуждаться при увеличении объема легких лежит
в основе рефлекторной саморегуляции дыхания, в
соответствии
с
которой
длительность
фаз
дыхательного цикла и дыхательный объем
определяются
частотой
импульсов
от
механорецепторов
76. Рецепторы растяжения легких
Возбуждениерецепторов
растяжения
легких вызывает рефлекторное торможение
вдоха и переход к выдоху. Этот рефлекс
называется
инспираторно-тормозящим
рефлексом Геринга-Брейера. У взрослого
человека
рефлексы
Геринга-Брейера
включаются, когда дыхательный объем
превышает 1 литр
77. Ирритантные рецепторы
Быстроадаптирующиеся
рецепторы
воздухоносных путей, слизистой оболочки
трахеи и бронхов. Особенно много их у корней
легких. Ирритантные рецепторы обладают
одновременно свойствами и механо- и
хеморецепторов. Пороги возбуждения этих
рецепторов
выше,
чем
у
рецепторов
растяжения, они возбуждаются при резких
изменениях объема легких.
78. Ирритантные рецепторы возбуждаются
подвоздействием
пылевых
частиц,
накапливающейся в воздухоносных путях слизи, при
действии паров едких веществ (аммиака, табачного
дыма), некоторых биологически активных веществ
(гистамина). Это быстро адаптирующиеся рецепторы
при раздражении которых в трахее - возникает
кашель,
першение,
жжение.
Возбуждение
ирритантных рецепторов бронхов провоцирует
частое и поверхностное дыхание, а импульсы от них
вызывают рефлекторное сужение бронхов бронхоконстрикцию.
79. J-рецепторы - юкстакапиллярные рецепторы легких
Этирецепторы
расположены
вблизи
от
капилляров малого круга кровообращения в
интерстициальной ткани альвеол. Раздражителем
для них является повышение АД в малом круге,
увеличение объема интерстициальной жидкости в
легких, эти рецепторы чувствительны к некоторым
биологически активным веществам (никотину,
простагландинам, гистамину)
80. J-рецепторы - юкстакапиллярные рецепторы легких
Импульсы от этих рецепторов поступаютк
дыхательному
центру
по
медленным
немиелинизированным волокнам блуждающего
нерва. Раздражение этих рецепторов приводит к
частому,
поверхностному
дыханию
и
бронхоконстрикции.
Сильное
и
устойчивое
возбуждение J-рецепторов наблюдается при застое
крови в малом круге, отеке легких, закупорке
мелких сосудов и других повреждениях легочной
ткани.
81. Химическая регуляция активности дыхательного центра
Изменения газового состава внутренней средыорганизма оказывают влияние на дыхательный
центр
опосредованно,
через
специальные
хеморецепторы, расположенные в структурах
продолговатого
мозга
«центральные
хеморецепторы» и в сосудистых рефлексогенных
зонах («периферические хеморецепторы»).
82. Химическая регуляция активности дыхательного центра
Хемочувствительнаязона
расположена билатерально и
лежит
под
вентральной
поверхностью
продолговатого мозга на глубине 0,2 мм.
Зона высоко чувствительна к
изменениям Рсо2, изменениям
концентрации Н+.
Возбуждает другие части
дыхательного центра.
83. Химическая регуляция активности дыхательного центра
Центральныехеморецептивные нейроны возбуждаются
только при действии на них
ионов
водорода.
Мультиполярные
нейроны
увеличивают метаболическую
и электрическую активности
при
гиперкапнии
и
при
локальном
повышении
концентрации ионов водорода
во внеклеточной жидкости,
омывающей эти нейроны.
84. Химическая регуляция активности дыхательного центра
На границе между кровью ивнеклеточной
жидкостью
высока активность фермента
карбоангидразы.
Эндотелий
сосудов,
граничащий с внеклеточной
жидкостью
в
области
хеморецептивных полей не
проницаем для ионов Н+ и
НСО3- но хорошо проницаем
для СО2
85. Химическая регуляция активности дыхательного центра
Повышениеконцентрации
СО2
в
крови
сопровождаетсяи диффузией в зону с высокой
карбоангидразной активностью
2) СО2
соединяется с Н2О под влиянием
карбоангидразы,
затем
диссоциирует
с
освобождением Н+
3) Накопление во внеклеточной жидкости ионов
водорода
приводит к повышению активности
мультиполярных нейронов.
1)
86. Периферическая система хеморецепторов в регуляции дыхательной активности
Существуетеще
один
механизм регуляции дыхания
— периферическая система
хеморецепторов,
которые
воспринимают
химические
раздражения и расположены в
нескольких
зонах
за
пределами мозга
87. Периферическая система хеморецепторов в регуляции дыхательной активности
Артериальные хеморецепторырасположены в дуге аорты и
каротидном
синусе,
и
представлены каротидными и
аортальными телами. Здесь же
локализованы и барорецепторы,
принимающие
участие
в
регуляции
артериального
давления.
88. Периферическая система хеморецепторов в регуляции дыхательной активности
В регуляции дыхания болееважна синокаротидная зона,
хотя
двустороннее удаление
каротидных тел у человека не
вызывает заметных изменений
дыхания в состоянии покоя.
Каротидные тела расположены
в месте деления общей сонной
артерии на внутреннюю и
наружную
89. Периферическая система хеморецепторов в регуляции дыхательной активности
В ткани каротидного теларазличают два типа клеток. I
тип - главные клетки, в них
содержатся гранулы, которые
исчезают при острой гипоксии.
С этими клетками контактируют
окончания афферентной ветви
языкоглоточного нерва (нерв
Геринга, синусный нерв).
90. Периферическая система хеморецепторов в регуляции дыхательной активности
Клетки I типа основные ввосприятии
хемочувствительности - разрушение этих клеток
прекращает
хеморецептивную
активность каротидного тела.
Мелкие клетки II
типа
гомологичны глиальным клеткам
и напоминают Шванновские.
Своими
отростками
они
оплетают главные клетки
91. Периферическая система хеморецепторов в регуляции дыхательной активности
Стимуляторами хеморецепторов каротидного теласлужат:
1) снижение напряжения кислорода
2) увеличение напряжения СО2
3) увеличение концентрации водородных ионов в
составе омывающей их артериальной крови
Главным
стимулятором активности каротидного
центра является гипоксия, которая сопровождается
более выраженным увеличением частоты импульсов
синусного нерва, чем сильная гиперкапния.
92. Механизм восприятия снижения напряжения кислорода в крови
Оценка уровня кислорода осуществляетсяспециальными
рецепторами,
которые
расположены на мембране клеток I типа
каротидного
синуса,
взаимодействие
которых
с
кислородом
приводит
к
активации калиевых каналов, выход К+ из
цитоплазмы
поддерживает
потенциал
клетки на уровне мембранного потенциала
покоя
93. Механизм восприятия снижения напряжения кислорода в крови
Снижениенапряжения
О2
в
крови
освобождает кислородный сенсор, калиевые
каналы
закрываются,
мембранный
потенциал
уменьшается,
достигает
критического уровня деполяризации, в
клетках I типа возникает потенциал
действия.
Возникновение
ПД
сопровождается открыванием Са 2+ каналов
и выделением дофамина.
94. Механизм восприятия снижения напряжения кислорода в крови
Артериальныехеморецепторы
возбуждаются и при повышении напряжения
СО2
в
артериальной
крови.
Гиперкапническая стимуляция артериальных
хеморецепторов происходит за счет прямого
влияния ионов Н+ при снижении рН крови.
Воздействие водородных ионов в клетках
каротидного тела обусловлено сдвигом
метаболизма
95. Механизм восприятия снижения напряжения кислорода в крови
Гипоксия и гиперкапния приводят кизменению метаболических процессов в
клетках, а стимуляторами каротидных
хеморецепторов
служат
метаболиты.
Однако реакция на снижение напряжения
кислорода наступает значительно быстрее
96. Механизм восприятия снижения напряжения кислорода в крови
Возникшее возбуждение по афферентнымволокнам синусного нерва проводится в
дорзальную
группу
дыхательных
нейронов
продолговатого
мозга,
возбуждение
которых
повышает
инспираторную активность в диапазоне
напряжения кислорода от 80 до 20 мм рт.ст.
97. Механизм восприятия снижения напряжения кислорода в крови
Хеморецепторыкаротидного
синуса
находятся
под
нервным
контролем:
повышение
активности
симпатической
нервной системы и выделение НА повышает
их чувствительность, а парасимпатические
импульсы и АХ – снижают.
98. Аортальные тела
Аортальные тела по строению подобныкаротидным телам, и выполняют функции
кислородных сенсоров. Хеморецепторы этой
зоны принимают незначительное участие в
регуляции дыхания, их основная роль
проявляется в регуляции деятельности
сердца и тонуса сосудов.
99. Физиологический смысл дублирования
Центральные хеморецепторыв условиях
гипоксии могут потерять чувствительность, что
приведет к снижению активности дыхательных
нейронов. В этих условиях дыхательный центр
получает основную возбуждающую стимуляцию от
периферических хеморецепторов, для которых
основным стимулом является дефицит кислорода.
Т.о., артериальные хеморецепторы - «аварийный»
механизм стимуляции дыхательного центра в
условиях снижения снабжения мозга кислородом