





















 biology
biologySimilar presentations:







")

")
Эволюционно-генетические отношения при симбиозе. (Тема 3)
1. Тема 3. Эволюционно-генетические отношения при симбиозе
2. 1. Коэволюция и коадаптация симбионтов.
Коэволюция – параллельная, взаимосвязаннаясовместная эволюция неродственных, но экологически
тесно сопряженных видов организмов, такое изменение
популяционной структуры одного из видов, которое
происходит под действием изменения в популяциях
партнера и оказывает обратное влияние на него.
Коадаптация – взаимное приспособление разных форм
живого, обитающих совместно.
Самец малахитовой нектарницы Nectarinia
famosa, сосущий нектар из растения Babiana
ringens. Видно, что, дотягиваясь до цветка,
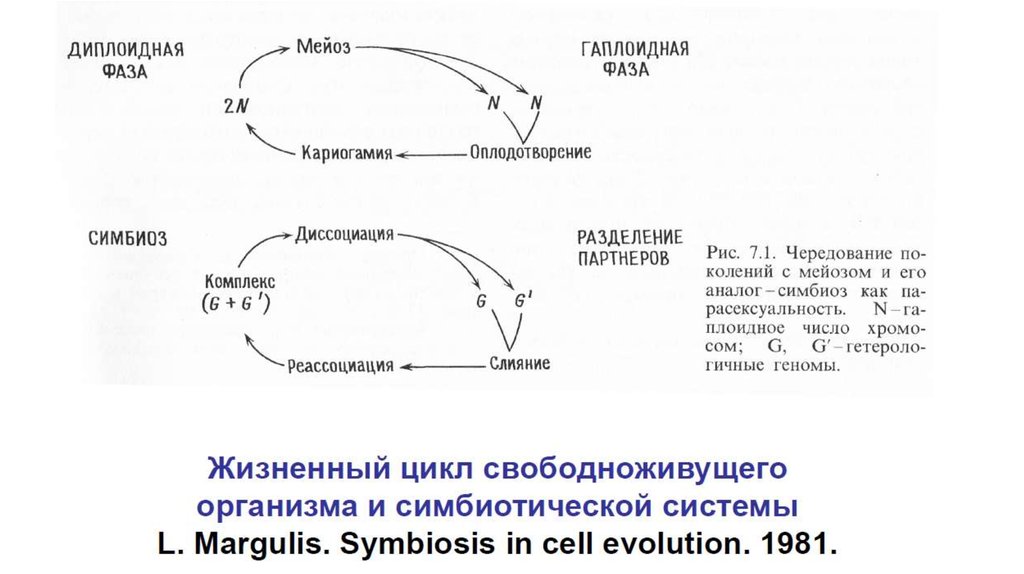
птица касается грудью тычинок и пестиков.
Вырост цветка, на котором сидит птица, повидимому, образовался в ходе коэволюции с
опылителями. Его единственная функция —
служить насестом для птиц, потребляющих
нектар и осуществляющих при этом
опыление. Фото с сайта
academic.sun.ac.za/botzoo
3.
Виноград Margravia evenia ,произрастающий в кубинских лесах, имеет
тарелочную форму листа, что помогает
летучим мышам легко находить цветки в
темноте с помощью эхолокации.
Heliconia и гранатовый колибри —
единственный опылитель цветков
Heliconia.
4.
Развитие многих видов бабочек-голубянок проходит в гнездах муравьев, которые выращивают гусениц, принимаяих по запаху за своих личинок. Голубянка алькон (Maculinea alcon) заражает гнезда нескольких видов муравьев
из рода Myrmica. В популяциях одного из этих видов (M. rubra) вторжения голубянки стимулируют локальные
эволюционные процессы: отбор поощряет изменение запаха муравьиных личинок, позволяя муравьям не путать
с ними паразитов. Бабочки, в свою очередь, эволюционируют вслед за муравьями, подстраиваясь под новый
преобладающий личиночный запах. Происходит эволюционная «гонка вооружений», идущая в разных
направлениях и вызывающая мозаичное распределение преобладающих запахов в популяциях как муравьев, так
и голубянок.
Самец голубянки алькон (Maculinea alcon)
на цветке эрики четырехмерной (Erica
tetralix). Фото с сайта news.bbc.co.uk
A — Молодая гусеница голубянки Maculinea alcon, на бутоне
горечавки Gentiana pneumonanthe; видны также яйца бабочки. B —
муравей Myrmica rubra (рабочая особь) несет гусеницу голубянки
в свое гнездо. Фото из статьи в Science
5. Сущность эволюционно-генетической теории симбиоза
• У симбионтов имеются генывирулентности (AVR) и гены
резистентности (R).
• Природные популяции
симбионтов полиморфны по
«генам симбиоза» (2 аллеля у
патогена AVR-avr, 2 аллеля у
хозяина R-r).
• Симбиотическая система имеет
собственный фенотип,
обусловленный
взаимодействием аллелей генов
обоих партнеров (Inf+, Inf-).
• Наследование фенотипа
симбиосистемы и
свободноживущего организма
осуществляется сходным
образом.
6. Генетический контроль взаимодействия при симбиозе
http://www.arriam.spb.ru/rus/.bin/noc/Lection1.pdf7. Гипотеза «Ген-на-ген»: каждому гену вирулентности паразита соответствует ген устойчивости хозяина.
• Сопряженные колебания частот аллелей в популяцияхрастений и патогенов.
8. Механизмы поддержания полиморфизма по генам устойчивости к паразитам
• Гипотеза Холдейна (1949): хозяева-носители редких биохимическихфенотипов имеют преимущество в борьбе с паразитами.
9. Эволюционная роль паразитизма
• Гипотеза Красной Королевы - Red Queen’s Hypothesis (Van Valen, 1974): Хозяева ипаразиты находятся в постоянном эволюционном соревновании — кто быстрее
изобретет средства нападения и защиты, у кого они окажутся лучше и эффективнее.
Стоит одной стороне замедлить движение в этом направлении, как преимущество
получает вторая сторона, и первая проигрывает: популяция теряет численность и
вымирает.
• Согласно гипотезе Красной Королевы, чем быстрее эволюционирует паразит, тем
быстрее эволюционирует хозяин;
«…здесь, знаешь ли, приходится бежать
со всех ног, чтобы только остаться на том
же месте! Если же хочешь попасть в
другое место, тогда нужно бежать по
меньшей мере вдвое быстрее!»
(Льюис Кэрролл
«Алиса в Зазеркалье»)
10. Закономерности коэволюции паразитов и хозяев
• При длительной совместной эволюции паразита и хозяина происходит снижение степениантагонизма между ними, снижается вредоносность паразита и реактивность хозяина так, что
он конечном итоге может перестать воспринимать паразита как чужеродный объект. В
последнем случае паразитизм может трансформироваться в комменсализм, и далее в
мутуализм.
• В процессе коэволюции возможно замещение некоторых функций хозяина симбионтом.
• Впоследствии такие сообщества становятся все более интегрированными, так что партнеры не
могут существовать по отдельности.
• Далеко не всегда антагонизм партнеров снижается в ходе коэволюции. Паразиты – некротрофы
эволюционируют по пути совершенствования механизмов патогенного воздействия на хозяина.
• В процессе коэволюции возрастает степень специфичности паразита в выборе хозяина. Наличие
большого круга хозяев – признак эволюционной молодости таксона паразита.
• Паразит повторяет эволюционный путь хозяина.
• Коэволюция ведет к возникновению новых свойств, стимулирует эволюцию разных форм
иммунитета.
11.
Эволюция гомеостаза в системе "хозяин-паразит" (хозяин комнатная муха Muscadomestica (I), паразит - оса Nasonia vitripennis (II) (по Ю.Одуму): а - недавно
объединенные популяции, б - популяции взятые из колоний, в которых оба вида
существовали два года.
12. 2. Половой процесс и симбиоз Разнообразие способов размножения организмов
13. Половое размножение и паразитизм
Гипотеза У. Хамильтона (Hamilton, Axelrod, Tanese, 1990. Sexual reproduction
as an adaptation to resist parasites): потомки полового размножения более
устойчивы к паразитам.
14. Доказательства гипотезы Хамильтона в наблюдениях за природными популяциями
Работа Кертисом Лайвлис соавторами была выполнена на
рыбках пецилиопсисах (Poeciliopsis).
Эти рыбки, как правило, заражаются
трематодами, что вызывает
появление на коже у инфицированных
рыб белых пятен.
В разных прудах обитали
пецилиопсисы, размножающиеся
бесполо (гибридные триплоиды) и
половым путем. Наиболее
зараженными были именно бесполые
гибриды, а те, кто размножался
половым путем, вместе с редким
бесполым клоном, оказались менее
подвержены инфекции.
15. Доказательства гипотезы Хамильтона в экспериментах с Caenorhabditis elegans
Caenorhabditis elegans ибактерия Serratia marcescens
Изменение доли скрещивающихся особей в ряду
поколений нематод. Контроль показывает
природную популяцию, которую не подвергали
действию паразитов; эволюция — природную
популяцию поместили в условия стабильной
инфекции; коэволюция — нематод подвергли
действию паразита, постоянно увеличивающего
свою вирулентность.
16. Симбиоз и половой процесс как формы генетической интеграции организмов
• при половых и парасексуальныхпроцессах рекомбинация открывает
возможность для повышения
адаптивного потенциала у потомков
взаимодействующих (родительских)
особей.
• При симбиозе расширение адаптивных
возможностей происходит непосредственно у
взаимодействующих организмов благодаря
функциональной интеграции их генов.
17.
18. Симбиоз и парасексуальные процессы прокариот
19.
20. Перенос генетической информации
Вертикальный• Процесс передачи генов от
родителей к потомству.
Горизонтальный (латеральный)
• процесс передачи генов между одновременно
существующими взрослыми организмами (от одного
генома к другому), часто - между разными видами,
штаммами, формами. Как правило, для этого
необходим посредник (вектор), роль которого могут
выполнять разные генетические конструкции вирусы, мобильные элементы.
21. Распространение и значение горизонтального переноса генов
• Горизонтальный перенос генов чрезвычайно широкораспространен в природе, особенно среди прокариот, от
10 до 80% (по разным оценкам) генов которых
приобретены таким образом.
• Существование горизонтального переноса генов недавно
доказано и для высших организмов – растений,
животных, включая высших приматов.
• Горизонтальный перенос позволяет приобретать новые
свойства гораздо быстрее, чем мутационный процесс и
служит фактором быстрой прогрессивной эволюции.
• Это относительно редкое событие – частота переноса
составляет 3 гена за 1 млн. лет.
• Имеет большое практическое значение в генетической
инженерии.
22. Модель экологической дифференциации двух популяций морской бактерии Vibrio cyclitrophicus. Тонкие серые и черные стрелки символизируют внутри- и ме
Модель экологическойдифференциации двух
популяций морской
бактерии Vibrio
cyclitrophicus. Тонкие серые
и черные стрелки
символизируют внутри- и
межпопуляционный ГГО.
Толстые цветные стрелки
показывают появление (в
результате мутации или ГГО)
адаптивных аллелей
(генетических вариантов),
облегчающих жизнь в двух
разных местообитаниях
(зеленом и красном).
Источник: B. Jesse Shapiro, Jonathan Friedman, Otto X. Cordero, Sarah P. Preheim, Sonia C. Timberlake, Gitta Szabó, Martin F. Polz,
Eric J. Alm. Population Genomics of Early Events in the Ecological Differentiation of Bacteria // Science. 2012. V. 336. Pp. 48–51.
23. Горизонтальный обмен у прокариот заменяет половой процесс
• генетический обмен междублизкородственными микробами
(представителями одной и той же
популяции) идет очень активно,
обеспечивая интенсивную
рекомбинацию и, фактически,
выполняя ту же функцию, что и
половой процесс у высших организмов.
• отбор идет в основном на уровне
отдельных фрагментов ДНК, а не целых
геномов.
• у бактерий, приспосабливающихся к
разным экологическим нишам,
формируется частичная
репродуктивная изоляция — по мере
накопления нуклеотидных различий
снижается вероятность гомологичной
рекомбинации.