





























 psychology
psychologySimilar presentations:

. Эндокринология")








Популяционные эффекты стресса
1.
Популяционные эффекты стресса.2.
Генетико-эволюционный рядЦарство
(по
И.А. Шилову, 1988)
Свойства системы
Вид
Структурированность
Интегрированность (целостность)
Способность к адаптивному ответу
на воздействия
Популяция
Организм
Положение
популяций в
иерархии биосистем
Биоценоз
Экосистема
Функционально-энергетический ряд
3.
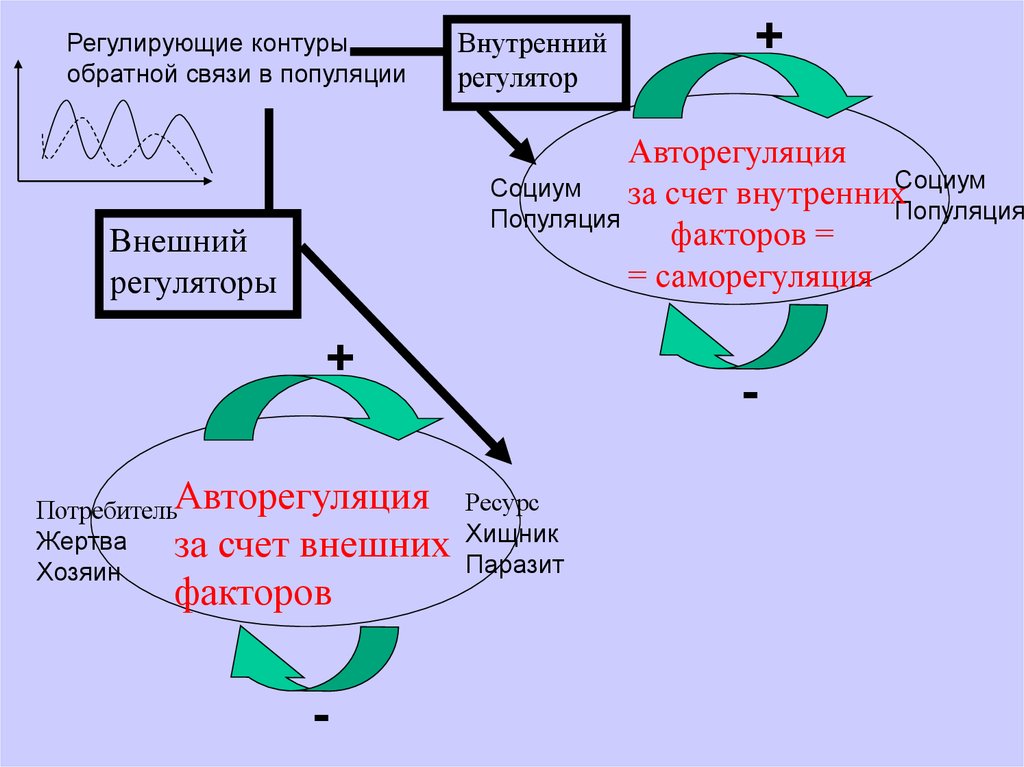
Регулирующие контурыобратной связи в популяции
Внутренний
регулятор
+
Авторегуляция
Социум
за счет внутреннихСоциум
Популяция
Популяция
факторов =
= саморегуляция
Внешний
регуляторы
+
ПотребительАвторегуляция Ресурс
Жертва
за счет внешних Хищник
Паразит
Хозяин
факторов
-
-
4.
Концепция саморегуляциипопуляционной плотности
Wynne-Edwards V.C. 1962 Animal dispersion in
relation to social behaviour Oliver and Boyd,
Edinburg. 653 pp
И.А.Шилов 1977.Эколого-физиологические
основы популяционных отношений у
животных М. МГУ. 260с
5.
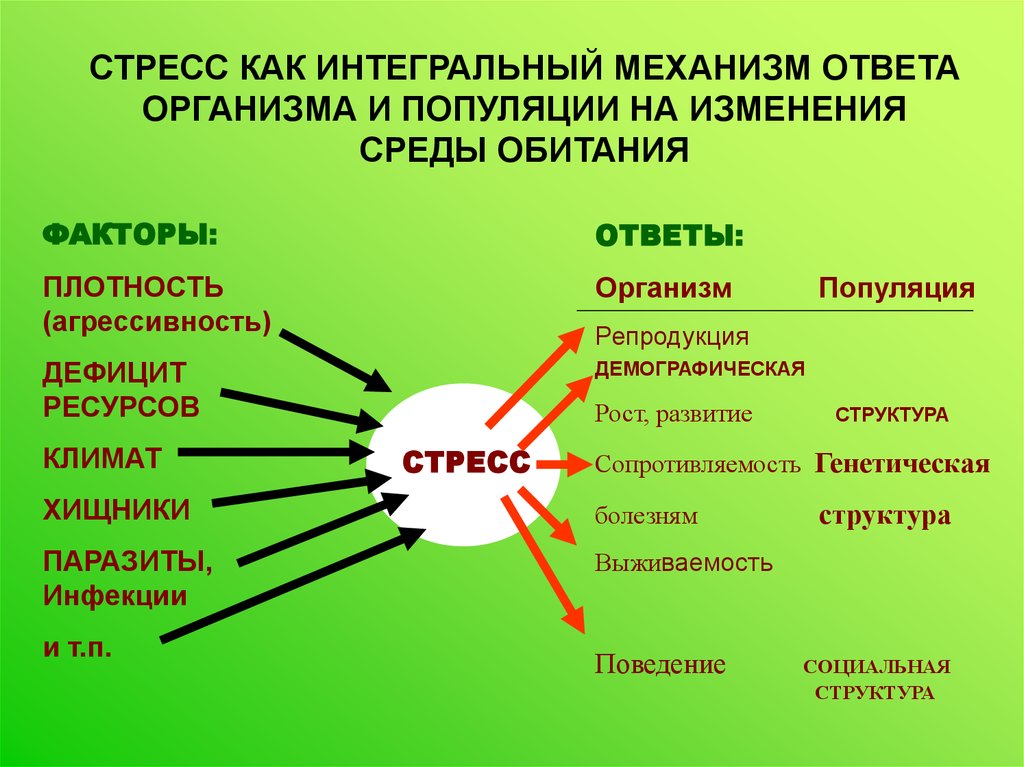
СТРЕСС КАК ИНТЕГРАЛЬНЫЙ МЕХАНИЗМ ОТВЕТАОРГАНИЗМА И ПОПУЛЯЦИИ НА ИЗМЕНЕНИЯ
СРЕДЫ ОБИТАНИЯ
ФАКТОРЫ:
ОТВЕТЫ:
ПЛОТНОСТЬ
(агрессивность)
Организм
ДЕФИЦИТ
РЕСУРСОВ
ДЕМОГРАФИЧЕСКАЯ
КЛИМАТ
Репродукция
Рост, развитие
СТРЕСС
СТРУКТУРА
Сопротивляемость Генетическая
ХИЩНИКИ
болезням
ПАРАЗИТЫ,
Инфекции
Выживаемость
и т.п.
Популяция
Поведение
структура
СОЦИАЛЬНАЯ
СТРУКТУРА
6.
Роль стресса в регуляции численностимлекопитающих
7.
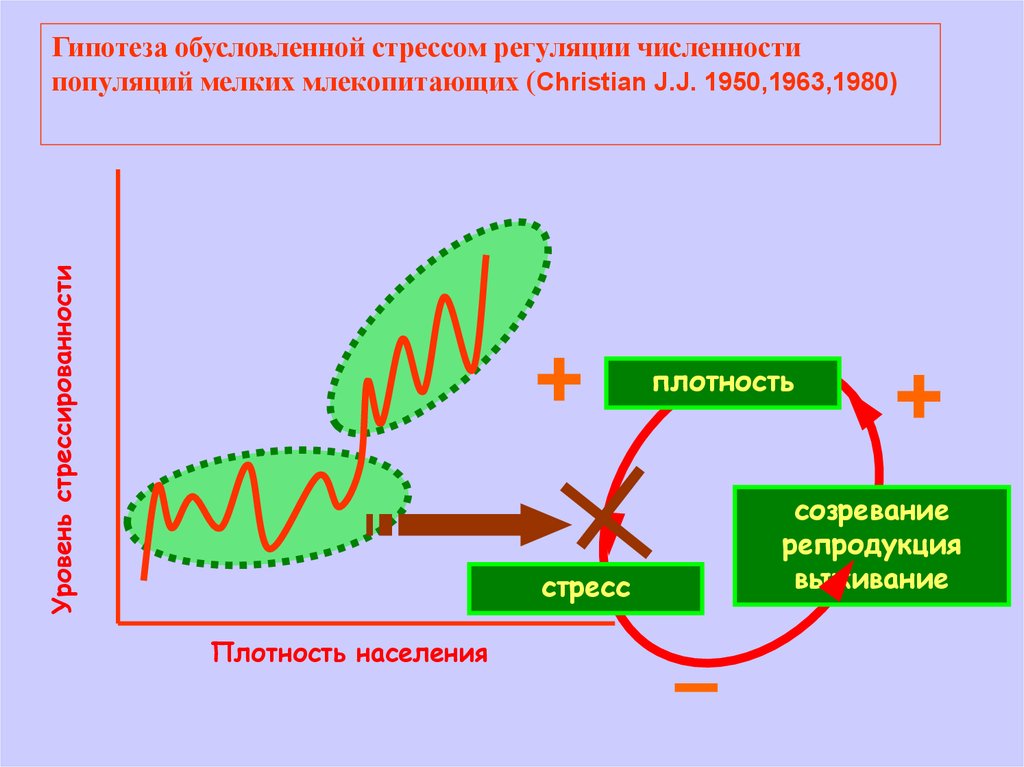
Уровень стрессированностиГипотеза обусловленной стрессом регуляции численности
популяций мелких млекопитающих (Christian J.J. 1950,1963,1980)
плотность
стресс
Плотность населения
созревание
репродукция
выживание
8.
Relationships between population density and stress in redvoles inhabited optimum and non-optimum area
Animals per 100 trap nights
Population dynamics
18
Altai (optimum)
16
Novosibirsk (non-optimum)
14
Corticosterone vs
Density r=0.64
12
*
10
8
Corticosterone vs
Density r=0.01
6
4
2
0
81
82
83
84
85
86
87
Years
С ЛЮБЕЗНОГО РАЗРЕШЕНИЯ
ПРОФ. М.П. МОШКИНА
88
89
90
91
92
9.
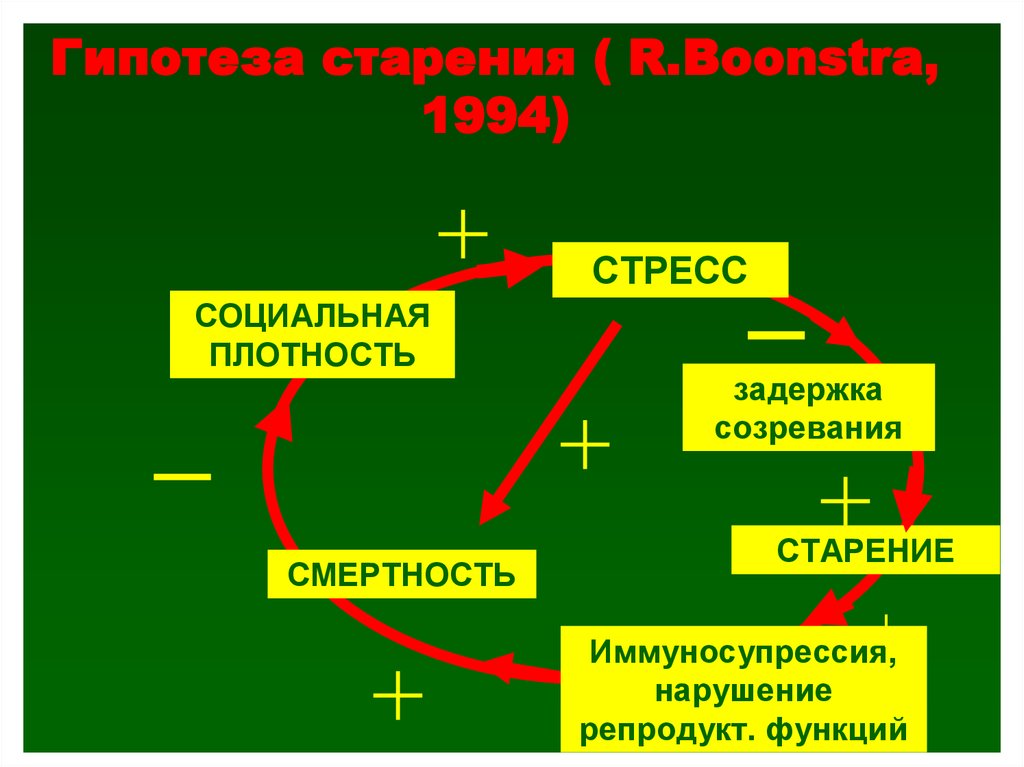
Гипотеза старения ( R.Boonstra,1994)
СТРЕСС
СОЦИАЛЬНАЯ
ПЛОТНОСТЬ
СМЕРТНОСТЬ
задержка
созревания
СТАРЕНИЕ
Иммуносупрессия,
нарушение
репродукт. функций
10.
Maternal effect hypothesis(Ward, 1984;
Lee, McDonald, 1985, Boonstra et al., 1998)
ПЛОТНОСТЬ
нарушение поведения,
репродукт. функций,
иммуносупрессия у
потомков
Стресс матери пренатальный
стресс
нарушение
развития
плода
11.
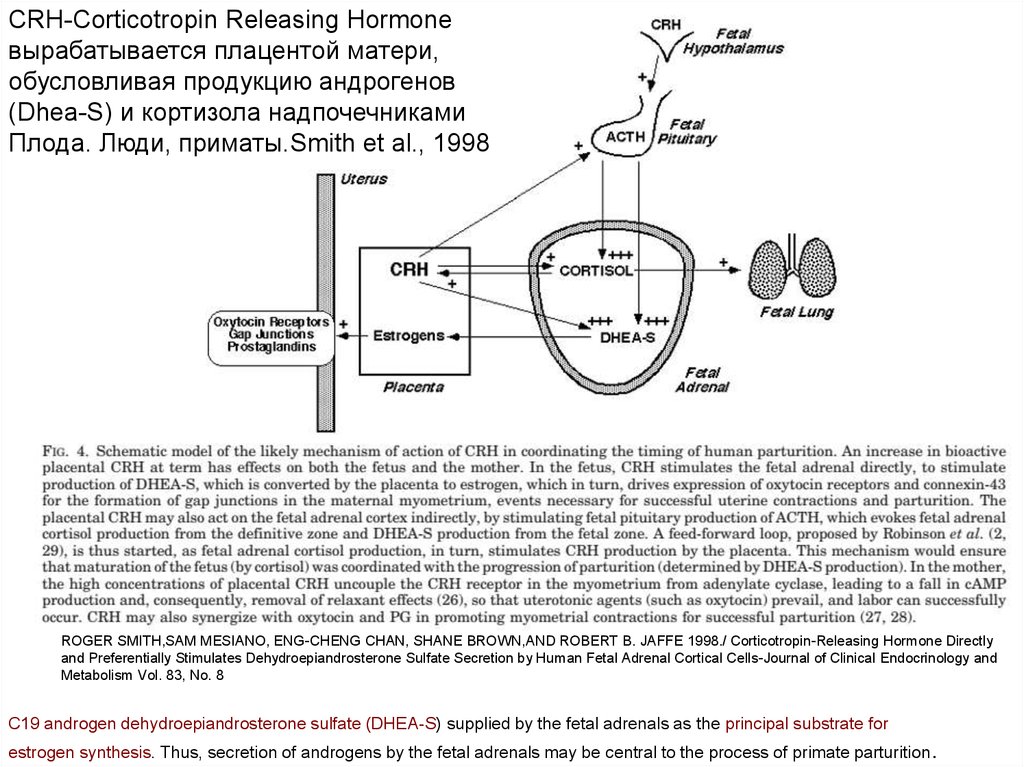
CRH-Corticotropin Releasing Hormoneвырабатывается плацентой матери,
обусловливая продукцию андрогенов
(Dhea-S) и кортизола надпочечниками
Плода. Люди, приматы.Smith et al., 1998
ROGER SMITH,SAM MESIANO, ENG-CHENG CHAN, SHANE BROWN,AND ROBERT B. JAFFE 1998./ Corticotropin-Releasing Hormone Directly
and Preferentially Stimulates Dehydroepiandrosterone Sulfate Secretion by Human Fetal Adrenal Cortical Cells-Journal of Clinical Endocrinology and
Metabolism Vol. 83, No. 8
C19 androgen dehydroepiandrosterone sulfate (DHEA-S) supplied by the fetal adrenals as the principal substrate for
estrogen synthesis. Thus, secretion of androgens by the fetal adrenals may be central to the process of primate parturition .
12.
Гипотеза зависящего от плотностиотбора геннетически обусловленных
типов поведения (агрессивных =
=территориальных особей). Мелкие
Млекопитающие.
(D.Chity, 1960, 67) была
сформулирована вне связи с
теорией стресса
13.
Однако, отбор настресс-устойчивость
(чувствительность) к стрессорам
реально существует в природе,
и вектор отбора может меняться по ходу
цикла численности популяции
14.
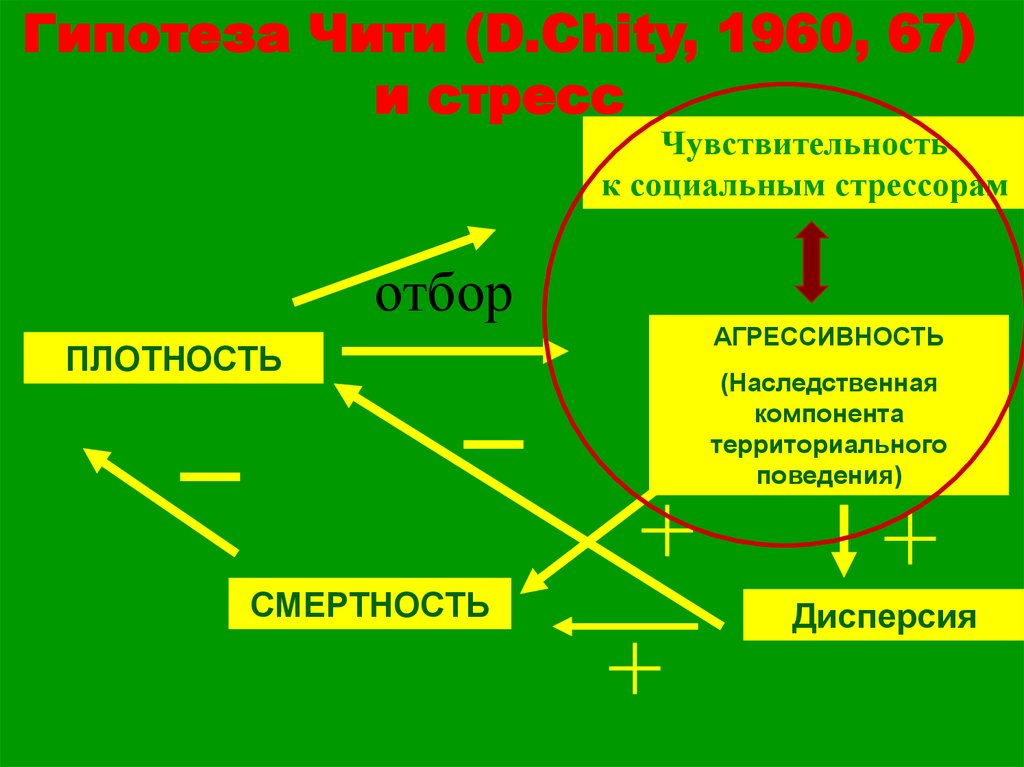
Гипотеза Чити (D.Chity, 1960, 67)и стресс
Чувствительность
к социальным стрессорам
отбор
ПЛОТНОСТЬ
СМЕРТНОСТЬ
АГРЕССИВНОСТЬ
(Наследственная
компонента
территориального
поведения)
Дисперсия
15.
Способность переносить воздействия стрессороввысокореактивный
низкореактивный
генотип
генотип
Эмоциональные
стрессоры
-
+
Sapolsky, 1992
Физические
стрессоры
+
-
Мошкин, 1992
-
+
+
-
Преимушество при высокой
плотности
Преимущество при низкой
плотности
16.
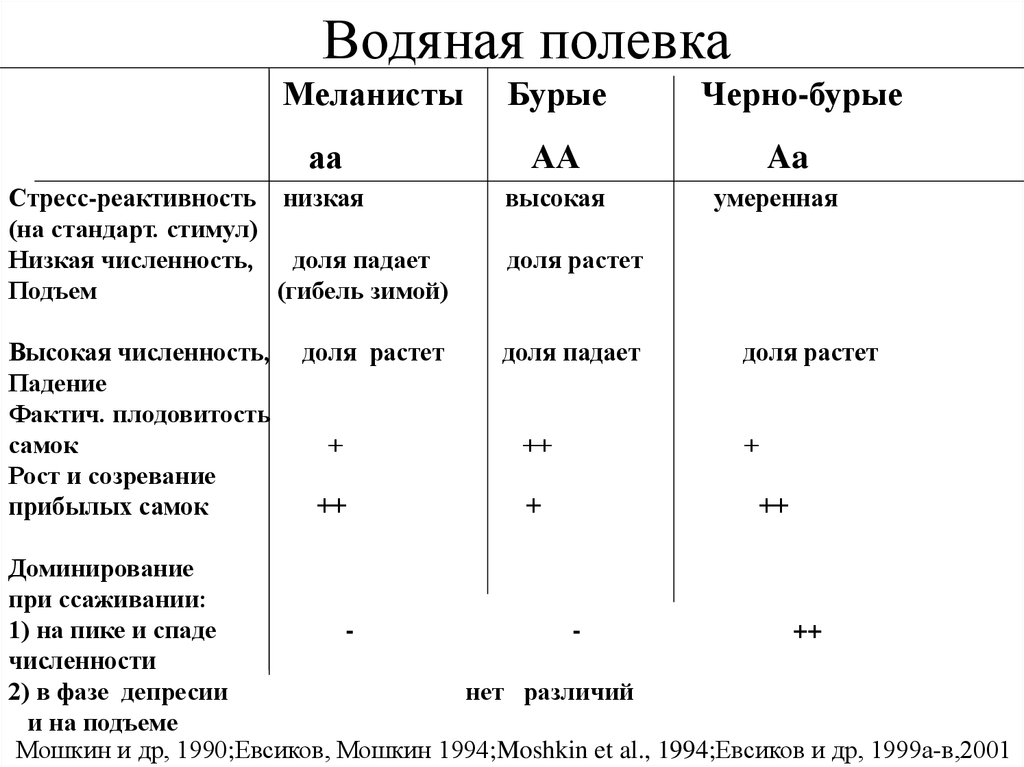
Водяная полевкаМеланисты
аа
Бурые
АА
Стресс-реактивность низкая
(на стандарт. стимул)
Низкая численность,
доля падает
Подъем
(гибель зимой)
высокая
Высокая численность,
Падение
Фактич. плодовитость
самок
Рост и созревание
прибылых самок
доля падает
доля растет
Черно-бурые
Aa
умеренная
доля растет
+
++
++
+
доля растет
+
++
Доминирование
при ссаживании:
1) на пике и спаде
++
численности
2) в фазе депресии
нет различий
и на подъеме
Мошкин и др, 1990;Евсиков, Мошкин 1994;Moshkin et al., 1994;Евсиков и др, 1999а-в,2001
17.
Таким образом, генетическидетерминированные особенности
нейроэндокринной реакции на стресс могут
вовлекаться в микроэволюционные
преобразования популяций животных в т.ч. в
результате направленного отбора на
большую или меньшую стресс-реактивность
18.
Примеры участия стресса врегуляции популяционной
плотности в природе
1. Мелкие хищные сумчатые Австралии.
Классическая саморегуляция
по Кристиану
19.
Antechinus subtropicus (Dasiuridae,Marsupialia)
Bradley et al., 1980,
McDonald et al., 1981,1989
ПОСЛЕ ГОНА
=>
<=
=>
Отсутствие связывающего андрогены протеина =>
Высокая концентрация тестостерона +Низкая конц. CBG
агрессия
Усиление адренокортикальной активности => Активный кортизол
=>
=>Глюконеогенез => Истощение организма => Смерть
20.
Водяная полевкаРоль доступности пищевых
ресурсов
21.
Водяная полевка (Arvicola terrestris)Multiannual dynamics of density and intermale aggression in
water vole population (from Evsikov V.I. et al., 1999)
1200
20
18
Population Density
Wounds
16
14
800
12
600
10
8
400
6
4
200
2
0
0
80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98
Year
Wounds
Animals per sq. km
1000
22.
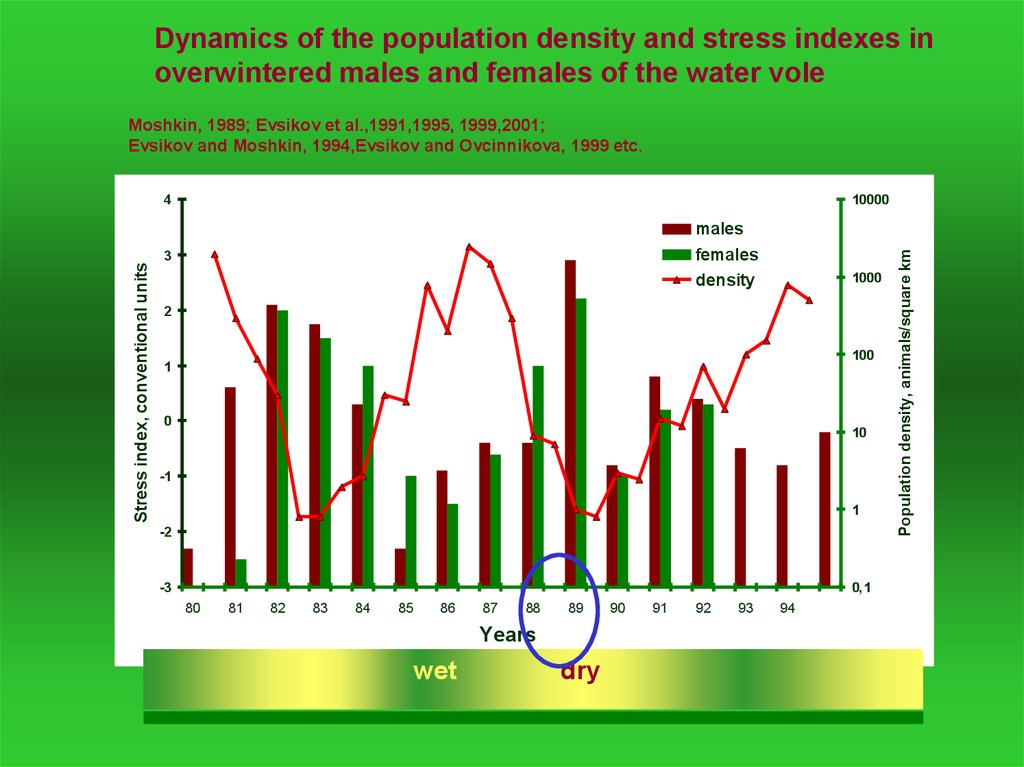
Dynamics of the population density and stress indexes inoverwintered males and females of the water vole
Moshkin, 1989; Evsikov et al.,1991,1995, 1999,2001;
Evsikov and Moshkin, 1994,Evsikov and Ovcinnikova, 1999 etc.
10000
males
females
3
1000
density
2
100
1
0
10
-1
1
-2
-3
0,1
80
81
82
83
84
85
86
87
88
89
Years
wet
dry
90
91
92
93
94
Population density, animals/square km
Stress index, conventional units
4
23.
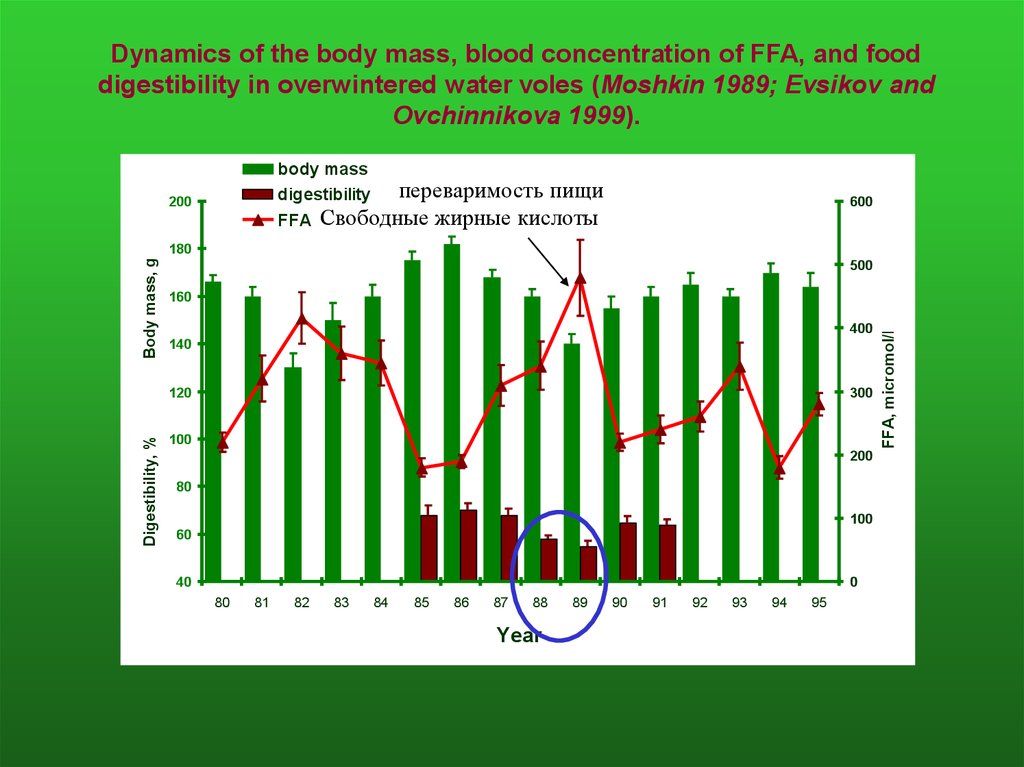
Dynamics of the body mass, blood concentration of FFA, and fooddigestibility in overwintered water voles (Moshkin 1989; Evsikov and
Ovchinnikova 1999).
body mass
переваримость пищи
digestibility
FFA Свободные жирные кислоты
200
600
500
160
400
140
Digestibility, %
120
300
100
200
80
100
60
40
0
80
81
82
83
84
85
86
87
88
Year
89
90
91
92
93
94
95
FFA, micromol/l
Body mass, g
180
24.
Но и роль социума важна.Обеспеченность пищей и самками
25.
**8,0
7,0
200
*
190
180
Масса тела самцов, г
Количество ран, шт.
6,0
Все самцы
5,0
4,0
3,0
2,0
170
160
1,2
1
3
1
150
140
130
120
110
1,0
100
0,0
Обилие ресурсов
Дефицит корма
Обилие ресурсов
Дефицит самок
3
500
2,3
400
2
2
2
300
200
Дефицит корма
Дефицит самок
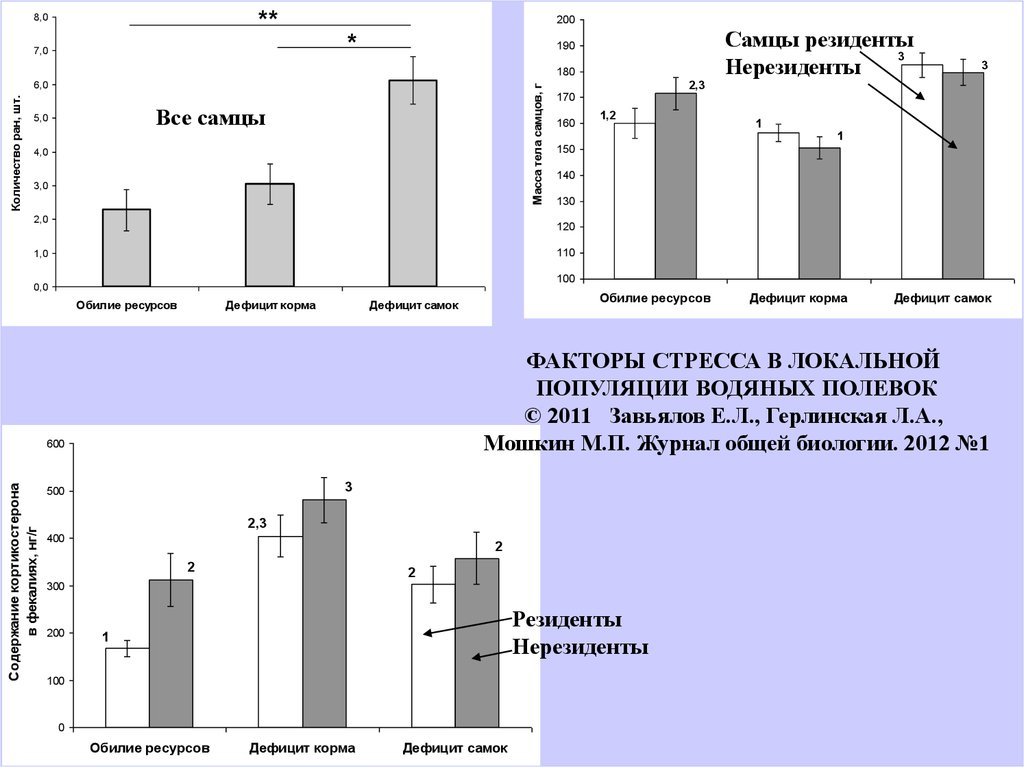
ФАКТОРЫ СТРЕССА В ЛОКАЛЬНОЙ
ПОПУЛЯЦИИ ВОДЯНЫХ ПОЛЕВОК
© 2011 Завьялов Е.Л., Герлинская Л.А.,
Мошкин М.П. Журнал общей биологии. 2012 №1
600
Содержание кортикостерона
в фекалиях, нг/г
2,3
Самцы резиденты
3
Нерезиденты
Резиденты
Нерезиденты
1
100
0
Обилие ресурсов
Дефицит корма
Дефицит самок
26.
Пример с большими песчанкамиРазные факторы стресса и
их относительная роль
27.
БОЛЬШАЯ ПЕСЧАНКА КАК СОЦИАЛЬНЫЙ ВИД•Семейные группы. Самец
заботится о потомстве
•Отсроченное выселение
молодых
•Кооперация:
Запасание корма
Защита от хищников
Охрана территории
годы
28.
Большая песчанкаFECAL CORTICOSTERONE CONCENTRATIONS IN ADULT MALES
IN THE BEGINNING OF SPRING, 1999-2008
VS. NUMBER OF FEMALES IN GROUP
1,90
годы
Spring 2008, recovering density
0
2,45
100
150
14
y = 0,17x - 2,76
12
10
R2 = 0,72
8
6
4
2
0
40
50
60
70
80
90
CORTICOSTERONE, ng/g
100
CO RTICO STERO NE ng/g
плотность
2,15
80
2,05
70
1,95
60
1,85
50
стресс
1,75
40
1,65
30
1,55
20
1,45
1,35
19
15
2
8
11
15
22
8
5
2
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008
YEARS
10
0
100
1,75
1,70
Y = 1.24+0.32*X
r=0.74; p=0.014
1,65
1,60
1,55
1,50
1,45
90
2,25
BURROWS OCCUPIED (%)
CORTICOSTERONE (Lg, ng/g)
2,35
50
16
1,80
1,40
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
# FEMALES IN GROUP
FECAL CORTICOSTERONE CONCENTRATIONS IN ADULT MALES
IN THE BEGINNING OF SPRING 1999-2008
VS. DENSITY OF FAMILY GROUPS
2,0
±Std. Err.
Mean
CORTICOSTERONE, Lg (Ng/g)
TESTOSTERONE, ng/g
TESTOSTERONE, ng/g
Spring 1999, high density
14
12
10
8
6
4
2
0
CORTICOSTERONE, Lg (Ng/g)
1,85
1,9
1,8
Y = 1.58+0.2*X
r=0.77; p=0.009
1,7
1,6
1,5
1,4
0,0
0,2
0,4
0,6
0,8
1,0
1,2
DENSITY OF GERBIL GROUPS (n/1ha)
1,4
1,6
1,8
29.
Взаимодействие факторов среды(ресурсы) и социальной природы
Взрослые самцы весной
годы
1999,2000, 2004 гг – высокая плотность
Cреднее за весну: R2=0.17; F=3.9; d.f.=3, 57; P=0.012
Кортикостерон = - 0,04 Ресурсы*** + 0,16 Число
самок* - 0,15 Дист. до ближ. соседа*
Начало весны: R2=0.17; F=4.3; d.f.=2, 41; P=0.020
Кортикостерон = + 0,20 Число самок* - 0,21 Дист. до
ближ. соседа*
30.
Stress in adult males in spring:Impact of environmental (resources) and
social factors
2001, 2002, 2003 – low density
Whole spring: R2=0.04; F=0.72; d.f.=2, 31; P=0.50
Corticosterone = N. S.
Beginning of spring: R2=0.11; F=3.4; d.f.=1, 28;
P=0.076
Corticosterone = -0.05Resources*
31.
В год высокой численности активность HPA системы веснойотрицательно коррелировала с обилием пищи летом
Body
mass
Ephemeral
herbage
(March-May)
-0.11
0.44* -0.01
Perennial
shrubs
(April- May)
0.12
Succulents
(JuneSeptember)
Haloxylon
trees(SummerFall-Winter)
-0.33*
0.07
0.20
Gerbil N
-0.11
0.05
КОРТИКОСТЕРОН ВЕСНОЙ,
Lg (ng/g)
Cortico
sterone
Food
plants
2,4
1999
High density
2,2
2,0
1,8
1,6
1,4
-0,1
0,1
0,3
0,5
0,7
ОБИЛИЕ ПИЩИ ЛЕТОМ
(летние солянки)
0.03
-0.06
-0.02
0,9
32. Стресс как регулятор темпа роста и созревания ==
== механизм модификациионтогенетических программ
33.
Среди грызунов широкораспространена филопатрия
оседание молодых близ
границ или в пределах
материнского (родительского
участка обитания)
34.
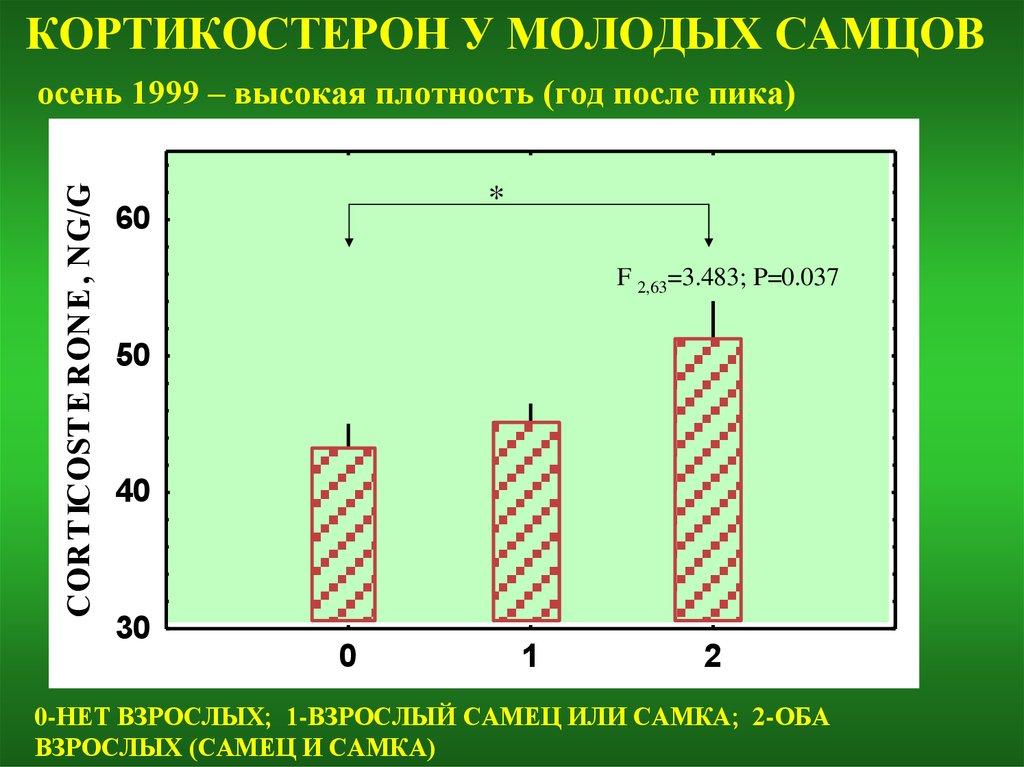
КОРТИКОСТЕРОН У МОЛОДЫХ САМЦОВCORT ICOST E RONE , NG/G
осень 1999 – высокая плотность (год после пика)
*
60
F 2,63=3.483; P=0.037
50
40
30
0
1
2
0-НЕТ ВЗРОСЛЫХ; 1-ВЗРОСЛЫЙ САМЕЦ ИЛИ САМКА; 2-ОБА
ВЗРОСЛЫХ (САМЕЦ И САМКА)
35.
БРЮШНАЯ ЖЕЛЕЗА МОЛОДЫХ САМЦОВосень 1999
MID-VENTRAL GLAND, mm
2
140
F 2,65 =7.66; P=0.001
**
120
*
100
80
60
0
1
2
0-НЕТ ВЗРОСЛЫХ; 1-ВЗРОСЛЫЙ САМЕЦ ИЛИ САМКА; 2-ОБА
ВЗРОСЛЫХ (САМЕЦ И САМКА)
36.
МАССА ТЕЛА МОЛОДЫХ САМЦОВОСЕНЬ 1999
1
8
0
F2,65=4.457; P=0.015
*
1
7
0
BODYMASS,g
1
6
0
1
5
0
1
4
0
1
3
0
0
1
2
0-НЕТ ВЗРОСЛЫХ; 1-ВЗРОСЛЫЙ САМЕЦ ИЛИ САМКА; 2-ОБА ВЗРОСЛЫХ (САМЕЦ И САМКА)
37.
ОТ
Н
О
С
И
Т
Е
Л
Ь
Н
О
Е
О
Б
И
Л
И
Е
В
А
Р
А
Н
О
В
В
1998-2002
Стресс и обилие хищников
(Большая песчанка и серый варан
в Кызылкумах)
4
2
1
0
1998
1999
2000
2001
2002
В
Е
Р
О
Я
Т
Н
О
С
Т
ЬП
О
С
Е
Щ
Е
Н
И
ЯН
О
Р
Ы
-К
О
Л
О
Н
И
И
С
Е
Р
Ы
М
В
А
Р
А
Н
О
М
ВЗ
А
В
И
С
И
М
О
С
Т
ИО
Т
Р
А
С
С
Т
О
Я
Н
И
ЯД
ОБ
Л
И
Ж
А
Й
Ш
Е
ЙН
О
Р
Ы
-К
О
Л
О
Н
И
И
0
,9
S
P
E
A
R
M
A
N
R
=
0
,5
4
;t=
2
,6
3
;P
<
0
,0
5
N
=
1
9
0
,7
0
,5
%ДНЕЙСОСЛЕДАМИВАРАНА
(бальнаяоценка)
ОБИЛИЕ
3
0
,3
0
,1
-0
,1
0
4
0
8
0
1
2
0
1
6
0
2
0
0
2
4
0
2
8
0 >
3
0
0
Р
А
С
С
Т
О
Я
Н
И
ЕД
ОБ
Л
И
Ж
А
Й
Ш
Е
ЙК
О
Л
О
Н
И
И
,m
1c
a
s
e
2c
a
s
e
s
38.
Sp
e
a
rm
a
nR
=
-0
.6
1
,t=
-2
.6
5
,P
<
0
.0
5
N
=
1
4
1
0.8
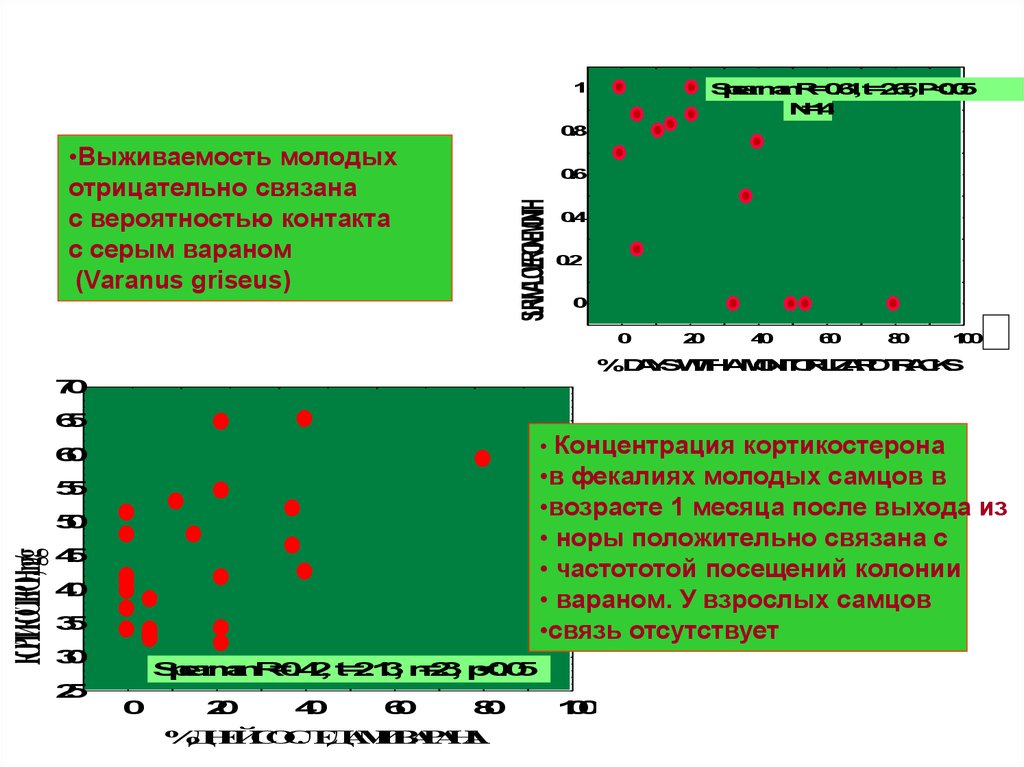
•Выживаемость молодых
отрицательно связана
с вероятностью контакта
с серым вараном
(Varanus griseus)
SURVIVALOVERONEMONTH
0.6
0.4
0.2
0
0
20
40
60
80
100
%D
A
Y
SW
IT
HAM
O
N
IT
O
RL
IZ
A
R
DT
R
A
C
K
S
7
0
6
5
• Концентрация кортикостерона
6
0
•в фекалиях молодых самцов в
•возрасте 1 месяца после выхода из
• норы положительно связана с
• частототой посещений колонии
• вараном. У взрослых самцов
•связь отсутствует
5
5
КОРТИКОСТЕРОН,ng/g
5
0
4
5
4
0
3
5
3
0
2
5
S
p
e
a
rm
a
nR
=
0
.4
2
, t=
2
.1
3
,n
=
2
3
,p
<
0
.0
5
0
2
0
4
0
6
0
8
0
%
Д
Н
Е
Й
С
О
С
Л
Е
Д
А
М
И
В
А
Р
А
Н
А
1
0
0
39.
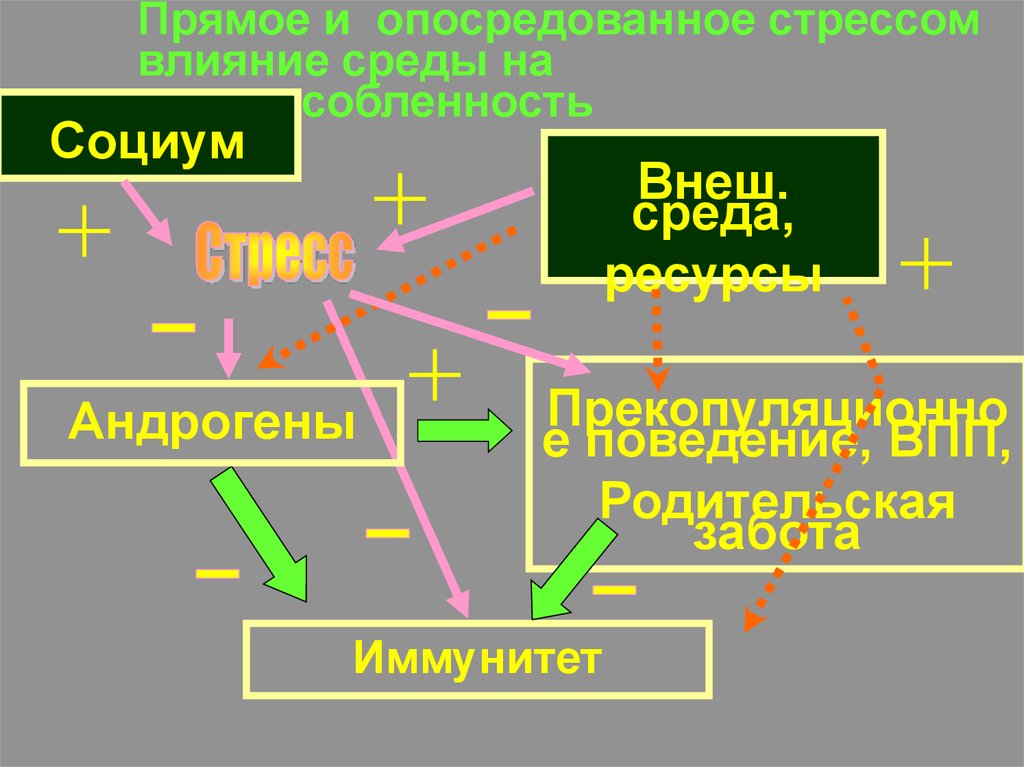
Прямое и опосредованное стрессомвлияние среды на
приспособленность
Социум
Внеш.
среда,
ресурсы
Андрогены
Прекопуляционно
е поведение, ВПП,
Родительская
забота
Иммунитет
40.
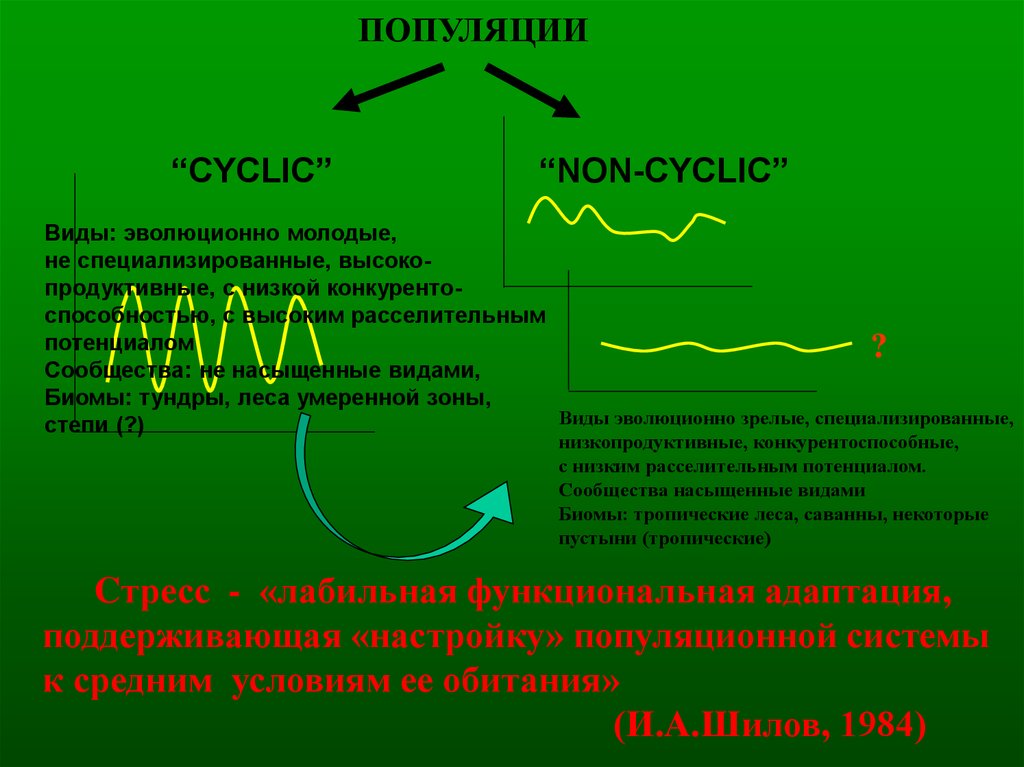
ПОПУЛЯЦИИ“CYCLIC”
“NON-CYCLIC”
Виды: эволюционно молодые,
не специализированные, высокопродуктивные, с низкой конкурентоспособностью, с высоким расселительным
потенциалом
?
Сообщества: не насыщенные видами,
Биомы: тундры, леса умеренной зоны,
Виды эволюционно зрелые, специализированные,
степи (?)
низкопродуктивные, конкурентоспособные,
с низким расселительным потенциалом.
Сообщества насыщенные видами
Биомы: тропические леса, саванны, некоторые
пустыни (тропические)
Cтресс - «лабильная функциональная адаптация,
поддерживающая «настройку» популяционной системы
к средним условиям ее обитания»
(И.А.Шилов, 1984)
41.
Перспективные направления исследованийстресса в природе
1. Факторы стресса и его роль в динамике
популяций разных видов, представителей разных
жизненных форм.
2. Роль стресса в регуляции поведения животных.
(нейроэндокринные механизмы на внутри и
межиндивидуальном уровнях, стресс в раннем
онтогенезе и его последствия,
3. Стресс как фактор эволюции. Роль стресса в
эволюции семейно-группового образа жизни
4. Положительная связь стрессированности с
репродукцией и выживаемостью в природе.
42.
Для чтения к экзаменуШилов И.А. 1977.Эколого-физиологические
основы популяционных отношений у
животных М. МГУ. 260с
Шилов И.А. 1984. Стресс как экологическое явление //
Зоологический журнал Т. 63, вып.6, с.805-810.
Роговин К.А., Мошкин М.П. 2007. Авторегуляция
численности в популяциях млекопитающих и
стресс (Штрихи к давно написанной картине)
//Журнал общей биологии Т.68. N 4. С. 244-267.
Новиков Е.А. Мошкин М.П. 2009. Роль стресса в
модификации онтогенетических программ //Успехи
соврем. Биол. Т129, №3,с. 1-12
43.
Селье Г. На уровне целого организма.— М.: Наука, 1972.
Селье Ганс. Стресс без дистресса
. — М.: Прогресс, 1982.
Кокс Т. Стресс. — М.: Медицина, 1981.
44.
THE END45.
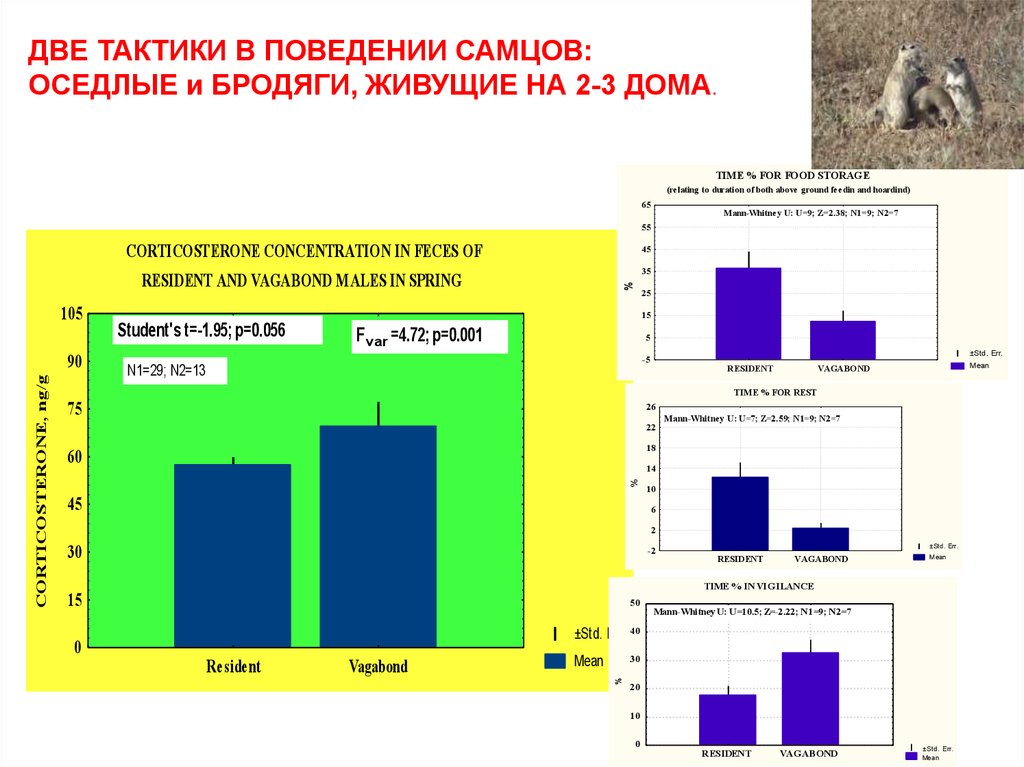
ДВЕ ТАКТИКИ В ПОВЕДЕНИИ САМЦОВ:ОСЕДЛЫЕ и БРОДЯГИ, ЖИВУЩИЕ НА 2-3 ДОМА.
TIME % FOR FOOD STORAGE
(relating to duration of both above ground feedin and hoardind)
65
Mann-Whitney U: U=9; Z=2.38; N1=9; N2=7
55
105
RESIDENT AND VAGABOND MALES IN SPRING
35
%
45
25
15
Student's t=-1.95; p=0.056
Fvar =4.72; p=0.001
5
±Std. Err.
-5
N1=29; N2=13
RESIDENT
Mean
VAGABOND
TIME % FOR REST
75
26
22
Mann-Whitney U: U=7; Z=2.59; N1=9; N2=7
18
60
%
14
45
10
6
2
30
-2
0
±Std. Err.
RESIDENT
VAGABOND
Mean
TIME % IN VIGILANCE
15
50
Resident
Vagabond
±Std. Err.
40
Mean
30
%
CORTICOSTERONE, ng/g
90
CORTICOSTERONE CONCENTRATION IN FECES OF
Mann-Whitney U: U=10.5; Z=-2.22; N1=9; N2=7
20
10
0
RESIDENT
VAGABOND
±S td. E rr.
Mean