












 biology
biologySimilar presentations:

. Лекция 2")
")







Топогенез белков. Посттрансляционная модификация
1.
ТОПОГЕНЕЗ БЕЛКОВПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ
СПЕЦГЛАВЫ БИОХИМИИ. ЛЕКЦИЯ 5.
2.
Топогенез белков- фолдинг (сворачивание) белка
- посттрансляционная модификация белка
- транспорт белков внутри клетки
- транспорт белков во внешнюю среду
2
3.
Посттрансляционная модификацияПолипептидная
цепь
–
первичный
продукт
биосинтеза
–
часто
подвергается химическим превращениям, изменяющим ее ковалентную
структуру. Такие превращения могут происходить как ко-трансляционно
(в процессе трансляции), так и посттрансляционно (после окончания
биосинтеза – трансляции).
Химические
видоизменяют
модификации
его
пострансляционной
завершают
функциональные
модификации
созревание
свойства.
катализируются
белка
Обычно
или
реакции
специфическими
ферментами и являются нематричными процессами (не кодируются).
- модификации обогащают возможности изменения свойств белков;
- модификации могут быть обратимыми, что регулирует проявление
активности белков в ответ на изменяющиеся условия и стимулы.
3
4.
1. Йодирование остатков тирозинаРеакция характерна для тиреоглобулина – высокомолекулярного белка,
содержащегося в щитовидной железе и являющегося источником
тироксина – гормона, влияющего на обменные процессы в тканях.
- йодирование остатков Tyr-5, 1291. 2555, 2748
- йод атакует ОН-группу тирозина по радикальному механизму, отщепля
HI и оставляя неспаренный электрон на кислороде тирозина
- электрон перемещается по ароматической системе, локализуясь в параположении и стимулируя разрыв связи С-СН2
- ароматический фрагмент с неспаренным электроном переносится
внутримолекулярно на кислород пространственно сближенного остатка
дийодтирозина
- высвобождение тироксина под действием аспартильной протеазы
4
5.
2. Образование остатков -карбоксиглутаминовой кислотыРеакция
происходит
в
некоторых
животных
белках,
функционирование которых требует образования комплекса с
ионами кальция.
Реакция
состоит
в
прямом
включении
СО2
в
гамма-
метиленовую группу остатка глутаминовой кислоты. Протекает
только в присутствии витамина К.
Реакция свойственна протромбину и другим белкам системы
свертывания
крови,
в
которых
карбоксилированию
повергаются несколько (обычно 10) остатков глутамата в
аминоконцевом участке полипептидной цепи.
Модифицированный
кальция,
клеточных
глутамат
одновременно
взаимодействует
связанными
с
с
ионами
фосфолипидами
мембран. Это способствует концентрированию
белков системы свертывания крови на поверхности клеток,
что облегчает их взаимодействие друг с другом.
5
6.
2. Образование остатков -карбоксиглутаминовой кислотыСходную роль играют остатки гамма-карбоксиглутаминовой
кислоты в белке костной ткани – остеокальцине, который
взаимодействует с ионами кальция на поверхности частиц
гидроксилапатита – минерального компонента кости.
Аналогично модифицированы остатки глутаминовой кислоты в
овокальцине и атерокальцине, некоторых белках хрящей,
почечных канальцев, а также в некоторых белках рибосом,
которые содержат и бета-карбоксиаспарагиновую кислоту.
Дефицит
витамина
(декумарина)
приводит
к
или
блокирует
торможению
взаимодействий.
6
К
присутствие
его
ингибиторов
гамма-карбоксилирование,
свертывания
крови
и
что
других
7.
3. Гликозилирование белковГликопротеины – белки, содержащие ковалентно связанный углеводный
компонент. По структурной роли углеводного компонента гликопротеины
можно разделить на 2 класса: протеогликаны и гликопротеины.
Протеогликаны
имеют
полипептидную
цепь,
плотно
унизанную
олигосахаридными цепочками, молекула в целом выступает подобно
разветвленному полисахариду, ее полипептидный скелет постоянно
скрыт.
В гликопротеинах полипептидная цепь формирует пространственную
структуру,
свойственную
белкам,
присоединены к белковой глобуле.
7
а
углеводные
компоненты
8.
3. Гликозилирование белковВ N-гликопротеинах углеводный компонент присоединяется N-
гликозидной связью к амидному азоту остатка аспарагина в
составе
полипептидной
цепи.
Первым
звеном
углеводного
компонента всегда оказывается N-ацетилглюкозамин.
N-гликозидная связь легко расщепляется в кислой среде.
В гликопротеинах к остатку N-ацетилглюкозамина присоединен 14-связью еще один остаток N-ацетилглюкозамина, далее следуют
несколько остатков маннозы, могут встречаться остатки Nацетилглюкозамина, галактозы, нейраминовой кислоты, фукозы и
некоторые другие моносахаридные звенья.
Антибиотик туникамицин блокирует N-гликозилирование белков.
Гликопептидаза F избирательно отщепляет олигосахариды от Nгликопротеинов.
Модификации подвергаются секреторные белки, попадающие в
ЭПР.
8
N-ацетилглюкозамин
9.
3. Гликозилирование белковВ О-гликопротеинах углеводный компонент присоединяется О-
гликозидной связью к оксигруппе остатка серина или треонина
в составе полипептидной цепи. Первым звеном углеводного
компонента всегда оказывается N-ацетилгалактозамин.
О-гликозидная связь легко расщепляется в щелочной среде.
Модификации подвергаются муцины (вещества, определяющие
группу крови).
В иммуноглобулинах встречаются одновременно оба типа
гликозилирования.
9
N-ацетилглюкозамин
10.
4. Фосфорилирование белковБелки фосфорилируют протеинкиназы, которые
катализируют перенос концевого фосфата АТФ на
гидроксильные группы остатков серина и треонина.
Фосфорилирование
тирозина
наблюдается
значительно реже.
Фосфоэфирные
гидролизоваться
связи
фосфопротеинов
ферментами
могут
-
фосфопротеинфосфатазами.
Фосфорилирование белков – обратимый процесс,
регулирующий активность белков. происходит в
области контакта субъединиц или доменов.
10
11.
5. Метилирование белков11
12.
6. Присоединение изопренильных групп12
13.
7. Модификация N- и С-конца13
14.
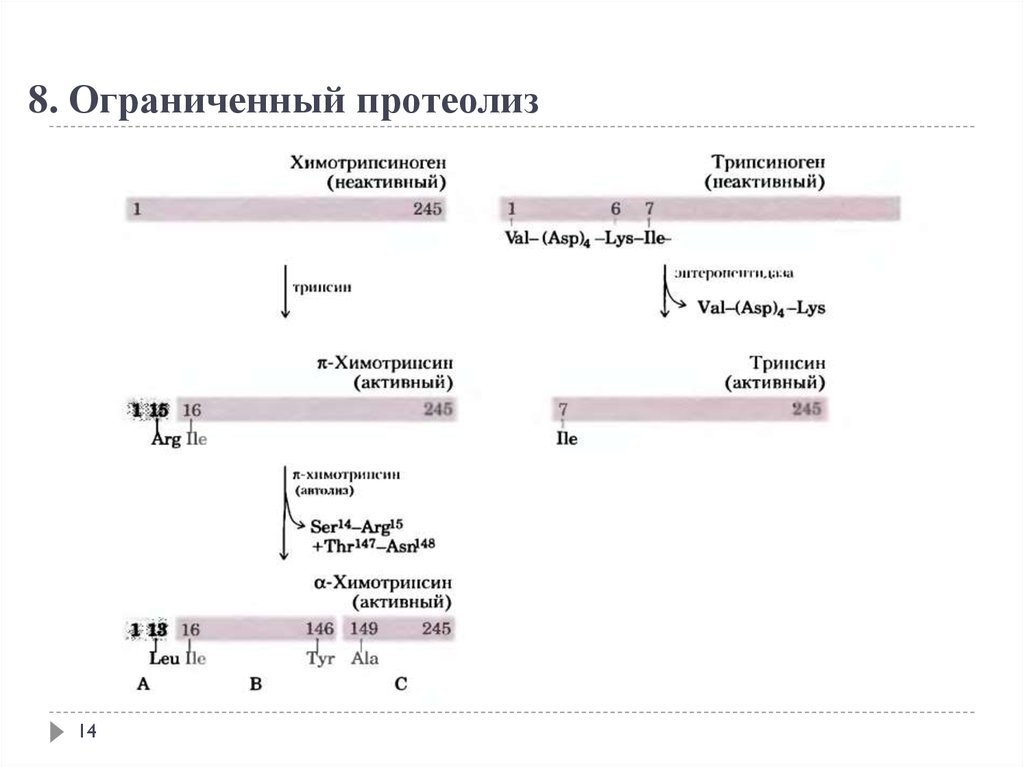
8. Ограниченный протеолиз14