
 – перегородки, разделяющие гифу на компартменты или клетки после деления")

 и гомобазидиомицетов")



")
")
")





")
 септы с тельцами Воронина у Aspergillus nidulans (А,В) и Neurospora crassa (C)")
 и сумки (9)")





 окружена группой везикул или трубочек (структурно идентичных ЭР)")


")
")


 и модифицированное с током")






 и Schizosaccharomyces pombe (Б)")

 Aspergillus oryzae (Б) (септинов в том числе)")
 лежит под клеточной мембраной, неизвестные белки (черные")






 вокруг поры.")
")




 biology
biologySimilar presentations:


. Лекция 8 -9")


")




Септы у грибов
1. Септы у грибов
2. СЕПТЫ (от лат. septum, saeptum, перегородка) – перегородки, разделяющие гифу на компартменты или клетки после деления
(одноклеточные грибы)3. Функции септ
• структурная поддержка гифы,• разделение мицелия на компартменты (с разными
физиологическими и биохимическими
процессами, например, при формировании спор),
• контролируемое распределение органелл (аско- и
базидиомицеты),
• первая линия обороны в случае повреждения
клеток (образование у несептированных грибов и
закупоривание пор у септированных грибов),
• конечный этап цитокинеза (материнская клетка
делится на две).
4. Быстрое закупоривание септальной поры у аскомицетов (включая дейтеромицеты с аскомицетным аффинитетом) и гомобазидиомицетов
Markham, 19945. Органеллы, которые закрывают септальную пору.
1.-2. Тельца Воронина1.Электронно-плотные тельца
Воронина округлой формы, одно
закрыло пору, второе расположено
рядом, и два в поврежденном
компартменте Aspergillus.
2. Тельце Воронина гексагональной
формы в поре Neurospora crassa.
3. Три вытянутых кристаллических
тела, лежащих бок о бок и
закрывающих пору Discina venosa.
6.
7. Химический состав септ аско- и базидиомицетов
• Грибная септа содержит хитин, который отсутствует уживотных и растений. Септальная пластина помимо
хитина у аскомицетов состоит из β-глюканов, но αглюкан - компонентом латеральных стенок - не
обнаружен. Старая септа может быть покрыта
аморфным слоем белка. Подобно аскомицетам,
химический состав септы у базидиомицетов
отличается от латеральных клеточных стенок.
Латеральные клеточные стенки состоят из хитина,
β-1,3/β-1,6-глюкана, но не α-1,3-глюкана. Более того,
долипоровое вздутие/утолщение содержит больше β1,6-глюкана, чем септальная пластинка.
8. Типы септ у разных таксономических групп грибов (Lutzoni et al., 2004)
микропоровая септамежду ризоидальной клеткой
и зооспоранигием
Плазмодесмы (=десмопоры)— микроскопические
цитоплазматические мостики, соединяющие
соседние клетки
9. Типы септ у вегетативных клеток разных таксономических групп грибов (Lutzoni et al., 2004)
пор.Kickxellalesоднопоровая септа
мицелий
пор.Mucorales
микропоровая септа
гаметангий
Септы большинства зигомицетовых грибов – сплошная поперечная перегородка,
которая формируется для изоляции старых или поврежденных районов мицелия или
при формировании репродуктивных структур
10. Типы септ у вегетативных клеток разных таксономических групп грибов (Lutzoni et al., 2004)
микропоровая септаоднопоровая септа с
тельцами Воронина
диаметр поры 50-500 нм
11. Однопоровая септа с поровыми включениями
12. Долипоровая септа с парентосомой
13. Септы с плазмодесмами между ризоидом и спорангиумом у Chytridium confervae из отдела Chytridiomycota
14. Септы с плазмодесмами у дрожжей
Fig. 9. Saccharomycopsissynnaedendra. Plasrnodesmata in
septum of a hypha.
Fig. 10. S. synnaedendra (Pichia
crossotarsi). Multiple scars
(arrowed) on neck of a yeast cell.
Fig. 11. Arthroascus javanensis.
Two separating cells with a single
connection in the center
of the septum.
Fig. 12. Saccharomycopsis
lipolytica. Mother cell and bud with
a central connection in the
cross wall.
15. Септа Geotrichum candidum с микропорами
16. Септы с одной порой с вильчатым утолщением и внутрипоровым включением (пор Kickzellales класс Zygomycetes отдела Zygomycota)
рис. 7-8 Mycoëmiliascoparia
рис. 9 Ramicandelaber
brevisporus
17. Однопоровые (=простые) септы с тельцами Воронина у Aspergillus nidulans (А,В) и Neurospora crassa (C)
18. Септы в генеративных клетках Sordaria humana : аскогенные гифы (5-8) и сумки (9)
19. Ультраструктура септы и септальной поры у Basidiomycota
20. Pucciniomycetes представлены ржавчинными грибами, которые имеют септы как у мицелиальных Ascomycota, но без телец Воронина (к
септам бывают приуроченымикротельца), трехламеллярная септа с простой порой, окруженной везикулами,
электронноплотными глобулами или микротельцами
или мультиперфорированная септа с простыми порами у Kriegeria eriophori
Microbotryomycetes
21. Pucciniomycetes
22. Ustilaginomycetes или головневые грибы. Имеют септы близкие по структуре к септам ржавчинных грибов, но могут иметь
незначительныйвздутый ободок вокруг
поры. Септальные поры
могут ассоциировать с
мембранными
шапочками или
мембранными
поясками/ободками
23. Редко у Tilletiales и Entorrhizomycetales – долипоровые септы без шапочек и парентосом, иногда могут иметь окружение из мембран
ЭПР24. Долипора - бочонкообразное вздутие вокруг поры (долипора) окружена группой везикул или трубочек (структурно идентичных ЭР)
класс Tremellomycetesпор.Tremellales
Trichosporon sporotrichoides - долипору окружает везикулярнотрубчатая шапочка, из мембран ЭПР
25.
Tremella globospora26. Долипора без парентосомы, окружена мембранами ЭПР
• Cryptococcus (Filobasidiella neoformans)из пор.Tremellales
27. Auricularia auricula-judae Lu, H. and D. J. McLaughlin (1991)
Auriculariaceae, Auriculariales,Incertae sedis, Agaricomycetes,
Basidiomycota, Fungi
Auricularia auricula-judae
Lu, H. and D. J. McLaughlin
(1991)
28. Долипора с перфорированной парентосомой и маленькими перфорациями как у Schizophyllum commune. Диаметр SPC (septal pore cap)
Schizophyllum commune от 450 до600 нм с перфорациями 100 нм.
29. Долипора с перфорированной парентосомой и крупными перфорациями у Rhizoctonia solani Диаметр SPC Rhizoctonia solani от 1600 до
2000 нм,имеющий 3-5 перфораций с диаметром 800 нм.
30. У некоторых агарикоидных грибов еще наблюдается наружная шапочка, например, у Agaricus bisporus, Coprinus cinereus, Agrocybe
praecox, которая напоминаетцистерны ЭР, связанные с перфорированными
парентосомами. Присутствие наружной шапочки вокруг
перфорированной SPC может зависеть от стадии
развития гифы.
31. Диаграмма продольного среза септального аппарата показывающая нормальное положение (точковая линия) и модифицированное с током
цитоплазмы (твердая линия)(Bracker, Butler, 1964)
Септальное
вздутие состоит
из хитина и
глюкана (альфа и
бета)
Долипоровый канал
от 70-500 нм
32.
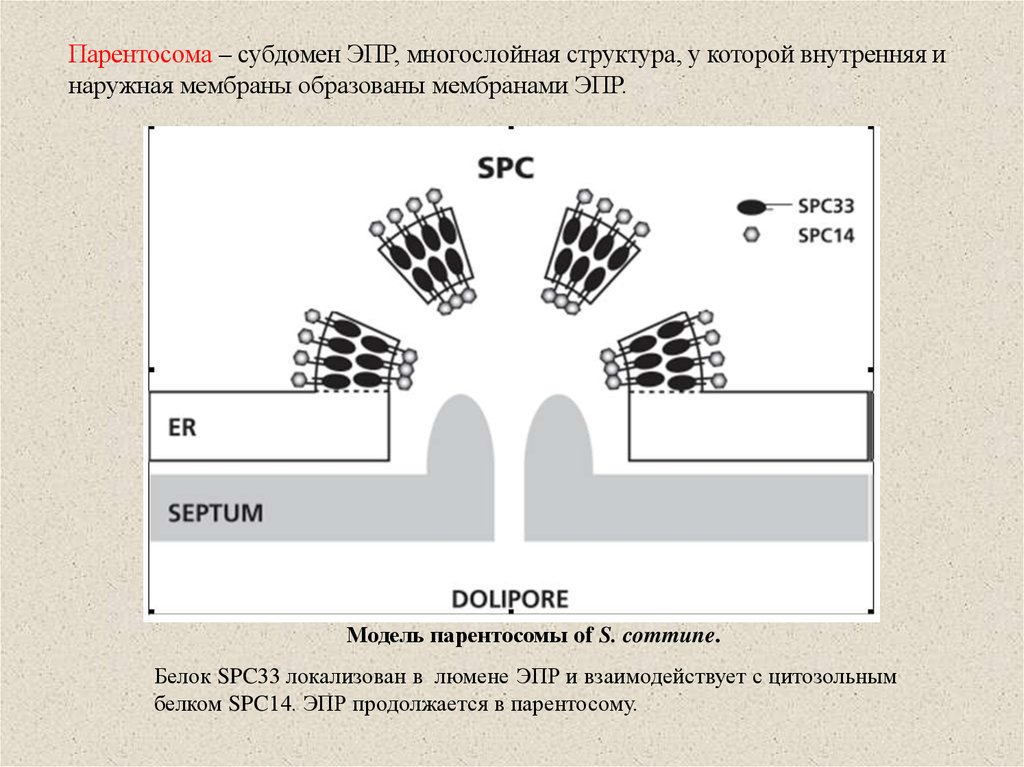
Парентосома – субдомен ЭПР, многослойная структура, у которой внутренняя инаружная мембраны образованы мембранами ЭПР.
Модель парентосомы of S. commune.
Белок SPC33 локализован в люмене ЭПР и взаимодействует с цитозольным
белком SPC14. ЭПР продолжается в парентосому.
33. Реорганизация долипоровой септы
• Начинается с лизиса долипорового утолщения.Долипоровое вздутие прогрессивно исчезает,
пока не исчезнет совсем. Реорганизация септы
как было показано ассоциировано со
вторжением везикул, мультивезикулярных тел
и вакуолей в район септы, которые, повидимому, содержат литические и
синтетические ферменты и компоненты
матрикса. Процесс лизиса направлен от
септальной поры к стенке гифы и
продолжается до исчезновения септы.
34. Частичный лизис долипорового вздутия у шампиньона двуспорового
35. Септирование и цитокинез у грибов
Септирование или формирование септы ицитокинез (деление цитоплазмы после
митотического деления ядра) у грибов имеют
ряд особенностей:
• Во-первых, образование септы не всегда
приурочено к делению ядра и это приводит к
образованию многоядерных клеток или даже к
существованию асептированного мицелия
(зигомицеты).
• Во-вторых, у грибов можно встретить два типа
цитокинеза (полный и неполный).
36.
• Грибы проявляют одну из двух моделей роста:дрожжеподобную или мицелиальную. Дрожжеподобный
рост, примером которого может быть рост дрожжей
Saccharomyces cerevisiae, характеризуется полным
цитокинезом – процессом, который заключается в разделении
дочерней клетки и материнской за счет частичной деградации
хитинсодержащей первичной (см. далее) септы.
Мицелиальные грибы обладают поляризованным ростом за
счет деления апикальной клетки и формируют гифу, которая
у большинства видов грибов (аскомицеты и базидиомицеты)
разделена на клетки септами. В септировании мицелиальных
грибов и цитокинезе дрожжей участвуют гомологичные
белки. Однако септа мицелиальных грибов не разделяет
полностью цитоплазму смежных компартментов из-за
наличия септальной поры (неполный цитокинез наблюдается
у большинства грибов). В местах септирования не
происходит гидролиза хитина, что приводит к формированию
многоклеточного мицелия.
37.
Цитокинез и формирование септы - завершающие этапыклеточного цикла и поэтому нуждаются в тщательном
генетическом контроле, также как и другие процессы,
такие как рост дочерней клетки/апикальный рост,
удвоение ДНК и митоз. Концепция деления клетки
одна из фундаментальных проблем всех клеток,
которая находит разные решения при развитии
бактерий, грибов, растений и животных. Клеточное
деление у грибов происходит в три этапа:
• Выбор места деления
• Сборка белкового комплекса
• Формирование сократительного акто-миозинового
кольца еще до окончания митоза и отложение хитина и
других компонентов септы
38.
• Формирование септы у мицелиальныхгрибов имеет место в прорастающих
спорах, в апикальных компартментах
гифы и в местах бокового ветвления,
при формировании пряжек и спор
(базидиоспор в т.ч.).
39. Выбор сайта деления у Saccharomyces cerevisiae (А) и Schizosaccharomyces pombe (Б)
голодание40. Образцы септирования у мицелия
41. Сборка белкового комплекса у S.cerevisiae (А) Aspergillus oryzae (Б) (септинов в том числе)
42. Образование септы у почкующихся дрожжей. Кольцо из септинов (желтым) лежит под клеточной мембраной, неизвестные белки (черные
стрелки) связывают акто-миозиновоекольцо (голубое) с мембраной.
Доставка трансмембранной хитин-синтазы Chs2 (розовым) в
мембранных везикулах (не показано) по соседству с мембраной и
сократительным кольцом.
43.
Формирование септы у Saccharomycescerevisiae
Формирование септы происходит в три этапа.
А. Первый этап происходит очень рано, когда
хитиновое кольцо (зеленым) появляется в
основании почки «почечеый рубец» (СsIII –
хитинсинтетаза III- черным, Rho1p и Fks1p –
компоненты β 1-3 глюкан синтазы). На этой
стадии дочерняя почка растет апикально.
В - дочерняя клетка меняет рост на
изодиаметрический, а Rho1p и Fks1p –
компоненты не выявляются.
С- Второй этап происходит в процессе
цитокинеза: когда сокращение актомиозинового
кольца приводит к инвагинации плазмалеммы.
Мембрана впячивается и синтезируется хитин
(реакция катализируется хитинсинтетазой II
коричневым цветом, CsII), который формирует
диск – первичную септу (зеленым), и которая
является первой структурой разделяющей
клетки.
D – первичная септа полная и происходит
образование вторичной септы (SS), которая
откладывается по обе стороны от первичной
септы (желтым).
Е – Третий этап: хитиназа лизирует первичную
септу, дочерняя клетка отделяется от
материнской. Лиловым цветом показана
хитинсинтетаза I, CsI, которая выполняет
репаративную функцию, если процесс лизиса
заходит слишком далеко.
44. Цитокинез у Schizosaccharomyces pombe
Сdс ("celldivision
cycle«)
proteins фосфатазы
45. Механизм сборки сократительного кольца у делящихся дрожжей
46. Митозы и септирование после прорастания конидии у A.nidulans
47.
Изображение схематичной модели сборки идинамики септальной пластинки у
Aspergillus nidulans.
Сегменты гифы ориентированы
параллельно друг другу так, что кончик
находится справа:
а). В ответ на сигнал митотического
веретена SepA локализуется в месте септы
как кортикальное пятно (зеленое). Хотя это
не показано, но возможно, маленькие
частицы актина и/или септина AspB могут
быть колокализованы с SepA.
b). Септальная полоска, которая состоит из
ансамбля ко-локализованного актина
(красное кольцо), AspB (желтое кольцо) и
SepA (зеленое кольцо). Дочерние ядра
завершают митоз.
с). AspB септиновое кольцо расщепляется
на два кольца, которые располагаются сбоку
от актинового и SepA колец. Было
предположено, что расщепление AspB
кольца может быть триггером сокращения
актинового кольца.
d). Актин и SepA кольца сокращаются и
откладывается материал септы (синий)
е). В процессе формирования септы, актин,
SepA и базальное AspB кольца исчезают, в
то время как апикальное AspB сохраняется.
48.
• Формирование септы на структурном уровнепроисходит следующим образом. Вслед за
инвагинацией плазмалеммы с небольшим
количеством материала стенки происходит
инициация синтеза септы. Появляются
микровезикулы предположительно с
предшественниками КС (хитосомы).
Микровезикулы (производные аппарата Гольджи)
и промежуточные филаменты обнаруживаются
поблизости от образующейся септы.
• Скорость формирования септы:
от 2 мин.(Alternaria solani), 4 мин. (N. crassa), 10
мин. (Rhizoctonia) до 20 мин. (Rhizopus nigricans).
49. Формирование долипоровой септы и парентосомы (septal pore cap, SPC) вокруг поры.
• Долипора формируется в процессе синтеза септы убазидиомицетов. После того, как септа приобретет свой
окончательный размер, мембрана септы отходит от порового
отверстия и вытягивается, формируя долипору. Утолщение затем
расширяется. Мало, что известно о формировании SPCs.
Основываясь на данных электронной микроскопии, была
предложена следующая модель синтеза этих структур. В этой
модели футляр из ЭР выстраивается параллельно септе и
аггрегирует в несимметричную по размеру структуру. Эта
структура затем развивается в два SPCs, которые остаются
закрепленными в процессе их роста. После разделения они
занимают положение у долипоровой септы. Положение SPCs
вдоль септе предположительно закрепляется на финальных
стадиях процесса формирования септы.
50. Ранние стадии развития долипоровой септы Pisolithus tinctorius (5 мин)
51.
52. Формирование септального аппарата у Auricularia auricula-judae
• Септа формируется за 4 минуты.:• На ранних этапах появляются тонкие филаменты с внешней
стороны септальной поры, которые связаны в районе отверстия
поры с промежуточными структурами. Филаменты 7,6-9,4 нм в
диаметре, обычно связывают ЦПМ окаймляя перегородку септы.
ЭПР продолжается в шапочку поры. Наблюдается центральная
пластина в септальной поре. Рибосомы отсутствуют между
пластиной и шапочкой. В инициацию септы и ее развитие
вовлечены микровезикулы, очевидно продукт аппарата Гольджи и
третий тип филаментов 10,6-13,2 нм в диаметре. Септальная
структура подтверждает отношение Auricularia к Tremella и
функциональный анализ предполагает отсутствие транспорта
крупных органелл между клетками, в отличие от
гомобазидиомицетов
53.
54.
55.
Стратегия цитокинеза у растений, дрожжей и клеток животныхМеханизм цитокинеза через сократительное
кольцо предка амеб, грибов и животных.