")









 НС, симпатического (Б) и парасимпатического (В) отделов вегетативной НС")







 узлы")
 узлы")

")



")
")

")





















 biology
biologySimilar presentations:










Частная гистология (учение о тканевом строении, развитии и жизнедеятельности отдельных органов и систем)
1. ЧАСТНАЯ ГИСТОЛОГИЯ (учение о тканевом строении, развитии и жизнедеятельности отдельных органов и систем)
2. СПИСОК ЛИТЕРАТУРЫ
• 1. Быков В.Л. Частная гистология человека (краткий обзорный курс).1999 г.
• 2. Данилов Р.К., Клишов А.А., Боровая Т.Г. Гистология. 2003 г.
• 3. Кузнецов С.Л., Мушкамбаров Н.Н. Гистология, цитология и
эмбриология. 2016 г.
• 4. Кузнецов С.Л., Мушкамбаров Н.Н., Горячкина В.Л. Атлас по
гистологии, эмбриологии и цитологии. 2010 г.
• 5. Гистология, цитология и эмбриология. , под ред. Афанасьева Ю.И. и
др., 1999 г., 2005 г., 2013 г.
• 6. Виноградова М.С. Органогенез.
3. ОРГАНЫ НЕРВНОЙ СИСТЕМЫ
4. Структура и функции нервной системы
Нервная система осуществляет объединение частей организма вединое целое (интеграцию), обеспечивает регуляцию разнообразных
процессов, координацию функции различных органов и тканей и
взаимодействие организма с внешней средой. Нервная система
воспринимает многообразную информацию, поступающую из внешней
среды и из внутренних органов, перерабатывает ее и генерирует
сигналы, обеспечивающие ответные реакции, адекватные
действующим раздражителям.
Функции нервной системы:
1.
2.
3.
4.
Обеспечение взаимодействия организма с внешней средой
Регуляция разнообразных жизненных процессов
Интеграция частей организма в единое целое
Координация работы органов
5. Развитие нервной системы
4 неделя развитияСтенка трубки подразделяется на
3 слоя:
1. внутренний слой – эпендима –
образует впоследствии
эпендимную глию
2. средний слой – плащевой –
даёт начало коре г.м., серому
веществу с.м., т.е. скоплению тел
нейронов
3. наружный слой – краевая вуаль
– превращается в белое
вещество, состоящее из
миелиновых нервных волокон.
Нервный гребень
(ганглиозная пластинка)
6.
7. Структура и функции нервной системы
- соматическая (анимальная) нервная система регулирует преимущественно функциипроизвольного движения;
-автономная (вегетативная) нервная система регулирует деятельность внутренних
органов и желез. Влияя на активность обмена веществ в различных органах и тканях в
соответствии с меняющимися условиями их функционирования и внешней среды, она
осуществляет адаптационно-трофическую функцию. Подразделяется на:
взаимодействующие друг с другом симпатический и парасимпатический отделы,
которые различаются локализацией центров в мозге и периферических узлов, а
также характером влияния на внутренние органы. В нее входят звенья,
расположенные в центральной и периферической нервных системах.
метасимпатический отдел (МНС)- комплекс микроганглионарных образований
(интрамуральных ганглиев) и соединяющих их нервов, а также отдельные
нейроны и их отростки, расположенные в стенках внутренних органов, которые
обладают сократительной активностью. Основными эффекторными аппаратами
стенок полых висцеральных органов, которые регулируются МНС, являются:
гладкая мышца, секреторный, всасывающий и экскреторный эпителий,
капиллярная сеть, местные эндокринные и иммунные образования.
Характеризуется высокой степенью относительной независимости от центральной
нервной системы. Не имеет ядерной структуры.
8. Основные понятия
Скопления нервных клеток (точнее, их тел):вне ЦНС называются нервными узлами (или ганглиями),
в головном и спинном мозгу – серым веществом (А), которое
представлено:
ядрами и (только в головном мозгу) корой.
Совокупность нервных волокон :
вне ЦНС называется нервным стволом (нервом),
а в головном и спинном мозгу – белым веществом (Б), в котором
миелиновые волокна образуют различные проводящие пути.
В нерве могут одновременно присутствовать :
и афферентные волокна (содержащие дендриты чувствительных
нейронов), и эфферентные;
а в каждом конкретном проводящем пути –либо только афферентные,
либо только эфферентные волокна.
А
Б
9. Основные понятия
Нервные центры — скопление нервных клеток в центральной и периферическойнервных системах, между которыми осуществляется синаптическая передача. Обладают
сложной структурой, богатством и разнообразием внутренних и внешних связей и
специализированы на выполнении определенных функций.
По характеру морфофункциональной организации различают:
нервные центры ядерного типа (нейроны располагаются без видимой
упорядоченности. Это вегетативные ганглии, ядра спинного и головного мозга);
нервные центры экранного типа (нейроны, выполняющие однотипные функции,
собраны в виде отдельных слоев, сходных с экранами, на которых проецируются нервные
импульсы (кора мозжечка, кора полушарий большого мозга, сетчатка глаза). Внутри слоев и
между ними имеются многочисленные ассоциативные связи.
В нервных центрах происходят процессы конвергенции и дивергенции нервного
возбуждения, функционируют механизмы обратной связи.
Конвергенция — схождение различных путей проведения нервных импульсов к
меньшему числу нервных клеток. На нейронах могут иметься окончания клеток разных типов,
что обеспечивает конвергенцию влияний из различных источников.
Дивергенция — образование связей одного нейрона с большим числом других, оказывая
влияние на их деятельность, обеспечивая перераспределение импульсов с иррадиацией
(распространением) возбуждения.
Механизмы обратной связи обеспечивают нейронам самостоятельное регулирование
величины поступающих к ним сигналов благодаря связям их аксонных коллатералей со
вставочными клетками. Последние оказывают влияние (обычно тормозное) как на нейроны, так
и на терминали конвергирующих к ним волокон.
10. В основе деятельности нервной системы лежат рефлекторные дуги, следовательно, общая организация НС системы лучше всего
В основе деятельности нервной системы лежат рефлекторные дуги, следовательно,общая организация НС системы лучше всего выявляется при составлении типичной
для неё рефлекторной дуги - цепочки из чувствительного, ассоциативного (одного,
нескольких или вообще без такового) и эффекторного нейронов, связанных друг с
другом синапсами, которая функционирует при типичном ответе организма на
определённое внешнее или внутреннее раздражение.
Рефлекторные дуги являются «морфологическим субстратом» нервной системы
В любой рефлекторной реакции участвует не одна цепочка нейронов, а некая
совокупность “параллельных” цепочек
11. Рефлекторные дуги соматической (А) НС, симпатического (Б) и парасимпатического (В) отделов вегетативной НС
11
1
1
2
1
А:
1-чувствительный нейрон
2-двигательный нейрон
2
1
1
1
2
1
3
1
Б, В:
2-преганглионарный нейрон
3-постганглионарный нейрон
D1-постганглионарный нейрон
(или клетка Догеля 1 типа)
D2- клетка Догеля 2 типа
D3- клетка Догеля 3 типа
12.
Принцип построения трехнейронных спинальных рефлекторных дуг13. Органы периферической нервной системы
• Нервы (нервные стволы)• Нервные узлы (нервные ганглии)
• Нервные окончания
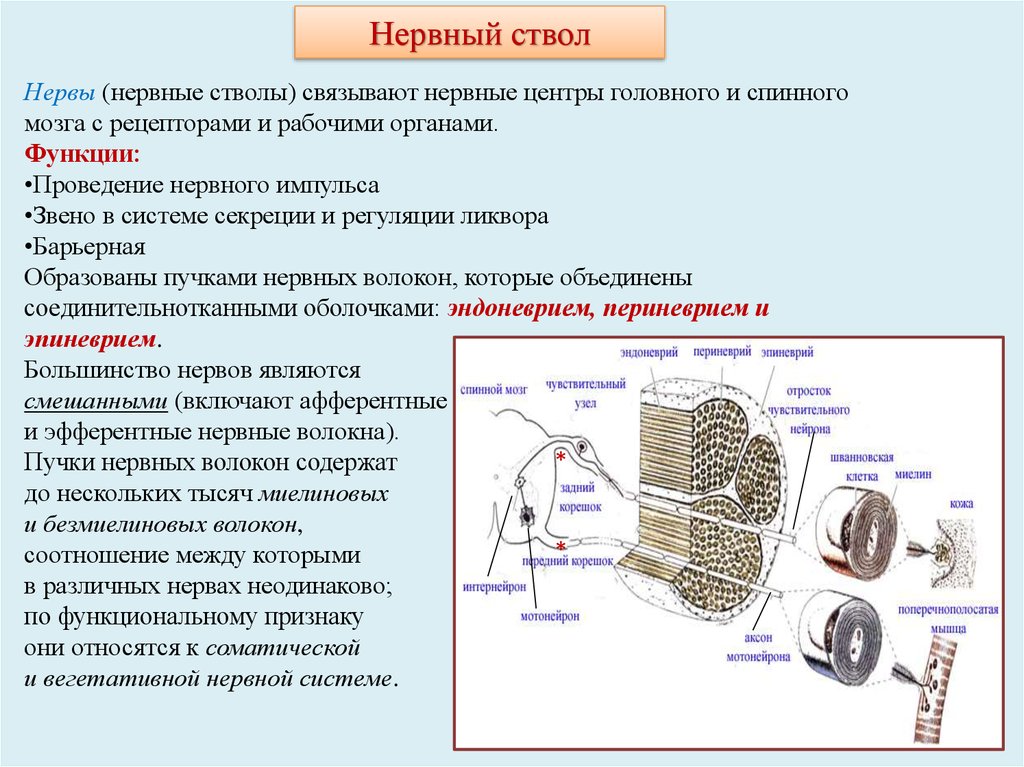
14.
Нервный стволНервы (нервные стволы) связывают нервные центры головного и спинного
мозга с рецепторами и рабочими органами.
Функции:
•Проведение нервного импульса
•Звено в системе секреции и регуляции ликвора
•Барьерная
Образованы пучками нервных волокон, которые объединены
соединительнотканными оболочками: эндоневрием, периневрием и
эпиневрием.
Большинство нервов являются
смешанными (включают афферентные
и эфферентные нервные волокна).
*
Пучки нервных волокон содержат
до нескольких тысяч миелиновых
и безмиелиновых волокон,
*
соотношение между которыми
в различных нервах неодинаково;
по функциональному признаку
они относятся к соматической
и вегетативной нервной системе.
15. Нервный ствол
Эндоневрий - тонкие прослойки рыхлой волокнистойсоединительной ткани с мелкими кровеносными
сосудами, окружающие отдельные нервные волокна и
связывающие их в единый пучок.
Периневрий - оболочка, покрывающая каждый пучок
нервных волокон снаружи и отдающая перегородки
вглубь пучка. Он имеет пластинчатое строение и
образован концентрическими пластами уплощенных
фибробластоподобных клеток. Между слоями клеток
в
пространствах,
заполненных
жидкостью,
располагаются компоненты базальной мембраны и
продольно ориентированные коллагеновые волокна.
Эпиневрий - наружная оболочка нерва, связывающая
пучки нервных волокон. Он состоит из плотной
волокнистой соединительной ткани, содержащей
жировые клетки, кровеносные и лимфатические
сосуды
16.
Общая характеристика нервного ствола17.
Ганглии (нервные узлы, скопление клеток за пределами ЦНС)Спинномозговой ганглий
Вегетативный ганглий
Строма:
•Капсула – покрывает ганглий снаружи, образована РСТ
•Прослойки РСТ внутри органа
Паренхима:
Нервные клетки и нервные волокна
Нервные клетки:
• располагаются диффузно
• располагаются группами
• все нейроны мультиполярные
• все нейроны униполярны
• все нейроны в основном двигательные
• все нейроны чувствительные
• нейромедиаторы: в симпатической НС –
• нейромедиаторы: АТФ, субстанция Р,
норадреналин, в парасимпатической НС кальцитонин-ген-родственный пептид
ацетилхолин
Нервные волокна:
•присутствуют в виде дендритов и аксонов нейронов, покрытых оболочками
В симпатических ганглиях имеются МИФ-клетки (малые интенсивно флюоресцирующие
клетки) – мелкие нейроны, содержащие серотонин, регулирующие проведение импульсов с
преганглионарных волокон на нейроны ганглия, от которых отходят постганглионарные
волокна.
В парасимпатических ганглиях нейроны делятся на три типа:
Клетки Догеля I типа –постганглионарные эфферентные нейроны
Клетки Догеля II типа – чувствительные нейроны местных рефлекторных дуг, образуют
синапсы с клетками 1 типа
Клетки Догеля III типа – ассоциативные нейроны, связывающие соседние ганглии
18. Общая характеристика спинномозговых ганглиев
19. Чувствительный ганглий спинномозгового нерва
1 - задний корешок;2 - чувствительный ганглий спинномозгового нерва:
2.1 - соединительнотканная капсула,
2.2 - тела псевдоуниполярных чувствительных
нейронов,
2.3 - нервные волокна;
3 - передний корешок;
4 - спинномозговой нерв
Спинномозговой (спинальный) узел (ганглий)
имеет веретеновидную форму, капсулу из ПВСТ,
периферические скопления тел псевдоуниполярных
нейронов, в центре - их отростки, между ними тонкие прослойки
эндоневрия с сосудами.
Переключения нервного импульса с одного нейрона
на другой в пределах спинномозговых узлов не
происходит, они не являются нервными центрами.
Псевдоуниполярные нейроны:
Крупные,
промежуточные
и
мелкие,
различаются
видами
проводимых
импульсов
(проприоцептивная,
тактильная
и
болевая+температурная соответственно).
В цитоплазме - многочисленные митохондрии,
цистерны грЭПС, элементы ПК, лизосомы.
Оболочки: слой клеток олигодендроглии
(мантийными
глиоцитами,
или
клеткамисателлитами), базальная мембрана и тонкая
соединительнотканная оболочка.
Отросток
разделяется
Т-образно
на
афферентную
(дендритную)
и
эфферентную
(аксональную) ветви, покрытые миелиновыми
оболочками. Афферентная ветвь заканчивается на
периферии рецепторами, эфферентная в составе
заднего корешка вступает в спинной мозг.
Нейромедиаторы: ацетилхолин, глутаминовая
кислота, вещество Р, соматостатин, холецистокинин,
гастрин, вазоинтестинальный пептид.
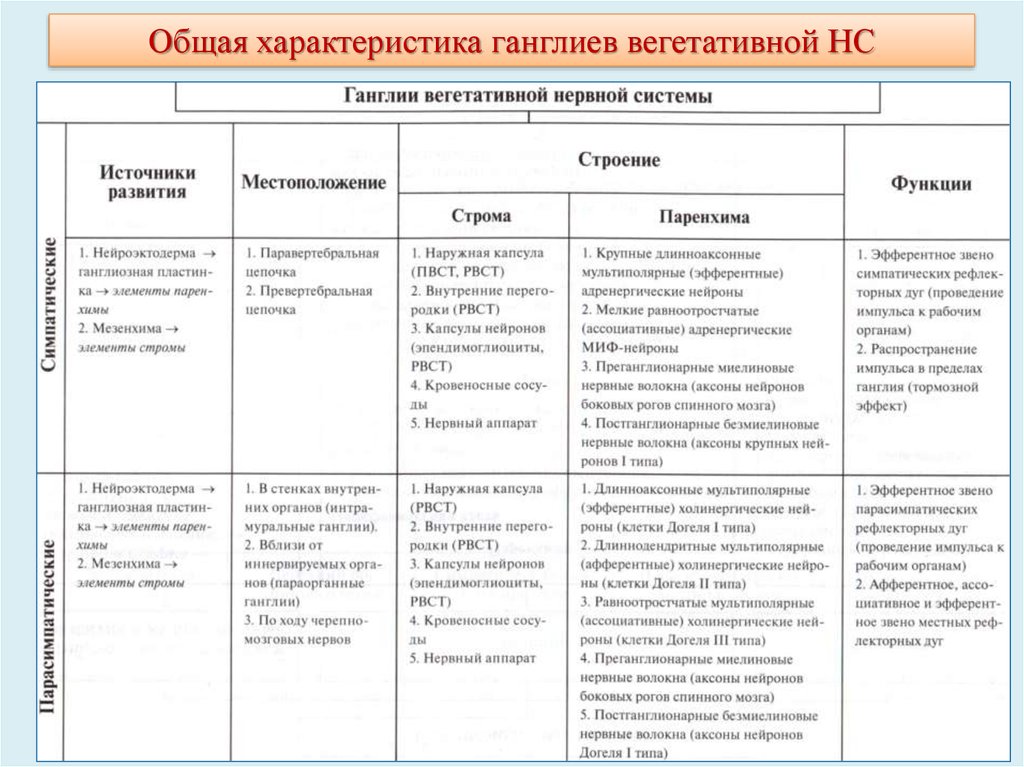
20. Автономные (вегетативные) узлы
Расположение: вдоль позвоночника (паравертебральные ганглии), или впереди него(превертебральные ганглии), а также в стенке органов — сердца, бронхов,
пищеварительного тракта, мочевого пузыря и других (интрамуральные ганглии) или
вблизи их поверхности. Могут иметь вид мелких (от нескольких клеток до нескольких
десятков клеток) скоплений нейронов, расположенных по ходу некоторых нервов или
лежащих интрамурально (микроганглии). К вегетативным узлам подходят
преганглионарные волокна (миелиновые), содержащие отростки клеток, тела которых
лежат в центральной нервной системе. Эти волокна сильно ветвятся и образуют
многочисленные синаптические окончания на клетках вегетативных узлов. Благодаря
этому осуществляется конвергенция большого числа терминалей преганглионарных
волокон на каждый нейрон ганглия. В связи с наличием синаптической передачи
вегетативные узлы относят к нервным центрам ядерного типа.
Разделяются (по функциональному признаку и локализации ): симпатические и
парасимпатические.
Общий план строения симпатических и парасимпатических нервных узлов сходен.
Вегетативный узел покрыт соединительнотканной капсулой и содержит диффузно или
группами расположенные тела мультиполярных нейронов, их отростки в виде
безмиелиновых или, реже, миелиновых волокон и эндоневрий. Тела нейронов имеют
неправильную форму, содержат эксцентрично расположенное ядро, окружены (обычно не
полностью) оболочками из глиальных клеток-сателлитов (мантийных глиоцитов). Часто
встречаются многоядерные и полиплоидные нейроны.
21. Автономные (вегетативные) узлы
Симпатические нервные узлы(пара- и превертебральные) получают преганглионарные
волокна от клеток, расположенных в вегетативных ядрах
грудных и поясничных сегментов спинного мозга.
Нейромедиаторы: преганглионарных волокон ацетилхолин, постганглионарных - норадреналин (за
исключением потовых желез и некоторых кровеносных
сосудов, имеющих холинергическую симпатическую
иннервацию), а также энкефалины, вещество Р,
соматостатин, холецистокинин.
узел солнечного сплетения
Парасимпатические нервные узлы (интрамуральные,
лежащие вблизи органов или узлы головы) получают
преганглионарные волокна от клеток, расположенных в
вегетативных ядрах продолговатого и среднего мозга, а
также крестцового отдела спинного мозга. Эти волокна
покидают центральную нервную систему в составе 3, 7,
9, 10 пар черепномозговых нервов и передних корешков
крестцовых сегментов спинного мозга.
Нейромедиаторы: пре- и постганглионарных волокон –
ацетилхолин, а также серотонин, АТФ, возможно,
некоторые пептиды.
интрамуральный ганглий в стенке мочевого
22. Интрамуральные ганглии
Автономные (вегетативные) узлыИнтрамуральные ганглии
Интрамуральный ганглий выявляется как скопление нервных клеток в толще органа.
Нейроны имеют крупные размеры, базофильную цитоплазму Вокруг нейронов, как обычно,
находятся глиальные клетки-сателлиты и соединительнотканные элементы и светлые ядра.
Интрамуральные узлы выполняют двойную функцию:
1. Их нейроны участвуют в образовании как центральных, так и периферических
рефлекторных дуг.
2. Ввиду соответствующей принадлежности эффекторных нейронов, периферические
рефлекторные дуги в большинстве своём вызывают парасимпатические эффекты.
Интрамуральные узлы и связанные с ними проводящие пути в виду их высокой
автономии, сложности организации и особенностей медиаторного обмена некоторыми
авторами выделяются в самостоятельный метасимпатический отдел вегетативной нервной
системы. В частности, общее число нейронов в интрамуральных узлах кишки выше, чем в
спинном мозге, а по сложности их взаимодействия в регуляции перистальтики и секреции их
сравнивают с миникомпьютером.
По своей функции нейроны интрамуральных ганглиев неоднородны: среди них
встречаются клетки всех трёх функциональных типов :
эффекторные (клетки 1-го типа)
чувствительные (клетки 2-го типа)
ассоциативные (клетки 3-го типа).
23. Интрамуральный ганглий (мышечная оболочка тонкой кишки)
Клетки Догеля в сплетении АуэрбахаЭффекторные нейроны (клетки 1-го типа,
или длинноаксонные клетки), воспринимают
сигналы от преганглионарных волокон – с
помощью холинергических синапсов и
передают эти сигналы к эффекторным, или
рабочим, структурам (например, к гладким
миоцитам) того органа, где расположен ганглий.
Чувствительные нейроны (клетки 2-го типа,
или равноотростчатые клетки),
мультиполярные, имеют несколько дендритов.
а) дендриты образуют в органе рецепторные
окончания;
б) поступающие от них сигналы передаются
клеткой сразу на эффекторный нейрон (клетку
1-го типа) того же узла.
Ассоциативные нейроны (клетки 3-го типа),
получают сигналы от чувствительных нейронов
узла и передают их эффекторным нейронам
соседних интрамуральных узлов.
24.
Общая характеристика ганглиев вегетативной НС25. Органы центральной нервной системы
• Спинной мозг• Мозжечок
• Кора больших полушарий головного
мозга
26. Спинной мозг
27. Спинной мозг
Располагается в позвоночном канале и имеет вид округлого тяжа, расширенного в шейном ипоясничном отделах и пронизанного центральным каналом. Он состоит из 31 сегмента в виде
двух симметричных половин, разделенных спереди срединной щелью, сзади срединной
бороздой, и характеризуется сегментарным строением; с каждым сегментом связана пара
передних (вентральных) и пара задних (дорсальных) корешков. В спинном мозге различают
серое вещество, расположенное в его центральной части, и белое вещество, лежащее по
периферии. Центральный канал заполнен спинномозговой жидкостью.
Покрыт тремя оболочками - мягкой, паутинной и твёрдой. Мягкая и паутинная оболочки
образованы РВСТ и покрыты одним слоем плоских эпителиоподобных клеток (мягкая –
снаружи, паутинная - с обеих сторон). Между ними - подпаутинное (субарахноидальное)
пространство, заполненное соединительнотканными трабекулами. Твёрдая мозговая
оболочка образована ПВСТ.
1 - серое вещество:
1.1 - передний (вентральный) рог,
1.2 - задний (дорсальный) рог,
1.3 - боковой (латеральный) рог;
2 - передняя и задняя серые спайки:
2.1 - центральный канал;
3 - передняя срединная щель;
4 - задняя срединная борозда;
5 - белое вещество (тракты):
5.1 - дорсальный канатик,
5.2 - латеральный канатик,
5.3 - вентральный канатик;
6 - мягкая оболочка спинного мозга
28. Спинной мозг (серое вещество)
Серое вещество: парные передние (вентральные), задние(дорсальные) и боковые (латеральные) рога (непрерывные
столбы, идущие вдоль спинного мозга), связанные спайкой
(комиссурой). В сером веществе находятся тела, дендриты
и (частично) аксоны нейронов (изодентритических: везде;
Идиодендритических: передние и задние рога, ядро Кларка;
Промежуточных:передние задние рога, собственное ядро з.р.),
глиальные клетки, формирующие глиальные мембраны на
поверхности сосудов. Между телами нейронов находится
нейропиль — сеть, образованная нервными волокнами и
отростками глиальных клеток (астроцитов).
Цитоархитектоника спинного мозга
Мультиполярные нейроны располагаются в виде скоплений (ядер), в которых происходит переключение
нервных импульсов с клетки на клетку (нервный центр ядерного типа). По топографии аксонов
нейроны СМ подразделяются на:
•корешковые нейроны, аксоны которых образуют передние корешки;
•внутренние нейроны, отростки которых заканчиваются в пределах серого вещества спинного мозга;
•пучковые нейроны, отростки которых образуют пучки волокон в белом веществе спинного мозга в
составе проводящих путей.
Наиболее важные ядра:
•задние рога содержат диффузные нейроны, собственное ядро и грудное ядро (ядро Кларка);
•в промежуточной зоне и в боковых рогах соответственно - медиальное промежуточное и латеральное
промежуточное ядра;
•в передних рогах - 5 соматомоторных ядер с самыми крупными (в спинном мозгу) нейронами (альфамотонейроны,35—70 мкм), мелкие гамма-мотонейроны (15—35 мкм), интерстициальное ядро Кахаля.
29. Спинной мозг (серое вещество)
Пластины Рекседа - это разнородные анатомические структуры сероговещества спинного мозга, выделенные на основе морфологии составляющих их
нейронов, иначе - колонки нейронов, которые на поперечных срезах спинного
мозга воспринимаются как ядра.
•Пластина I представляет собой самый поверхностный слой дорсального рога,
ее еще называют краевым слоем. Он содержит большие плоские "маргиальные
клетки" и нейроны промежуточного размера.
•Пластину II называют "желатинозной" из-за ее желатиноподобного вида на
свежем срезе спинного мозга. В ее состав входят мелкие плотно расположенные
клетки.
• Пластина III содержит крупные рыхло располагающиеся клетки.
• Пластина IV, самая толстая из расположенных в заднем роге, скомпанована из больших нейронов с
дендритами, распространяющимися в другие пластины. Вместе пластины III и IV образуют собственное
ядро (nucleus proprius).
• Пластина V состоит из мелких нейронов. Весь дорсальный рог сформирован пластинами I-V.
• Пластина VI локализуется в самом основании дорсального рога и прослеживается только в зонах
утолщения спинного мозга (шейный и поясничный отделы).
• Пластина VII занимает неправильной формы область в центре серого вещества спинного мозга.
Пластины промежуточной зоны – VI- VII.
• Пластина VIII охватывает внутреннюю половину переднего рога в области шейного и поясничного его
утолщений.
• Пластина IX соответствует расположению группы двигательных нейронов в вентральном роге. Таким
образом, вентральный рог сформирован пластинами VIII – IX.
• Пластина Х окружает центральный канал.
30. Спинной мозг. Основные структурные характеристики серого вещества
31. Спинной мозг (белое вещество)
Белое вещество - это совокупность нервных миелиновых волокон.Волокна идут, в основном, вдоль длинной оси спинного мозга
и образуют различные проводящие пути - восходящие
и нисходящие, связывающие различные сегменты спинного
мозга, либо спинной мозг с головным.
Рогами серого вещества и глиальными перегородками
белое вещество разбивается на 3 пары канатиков:
•задние канатики (6), отделённые друг от друга
срединной перегородкой (9) и содержащие преимущественно восходящие пути;
•боковые канатики (7), в которых идут и восходящие, и нисходящие пути;
•передние канатики (8), между которыми находится глубокая срединная
вырезка (10); в этих канатиках – преимущественно нисходящие пути.
•В срединную вырезку заходит мягкая мозговая оболочка; на срезе она выглядит
как чёрная (при импрегнации серебром) и произвольно изгибающаяся полоска.
•б) Посередине спинного мозга находится центральный канал (11), выстланный
эпендимоцитами
32. Спинной мозг
33. Мозжечок. Развитие
В эмбриональном периоде сначала формируется древняя часть мозжечка –червь, а затем – его полушария.
На 4-5-м месяце внутриутробного развития разрастаются поверхностные
отделы мозжечка, образуются борозды и извилины. В первый год жизни наиболее интенсивный рост мозжечка (в 4 раза) , когда ребенок учится сидеть и
ходить. К 3 годам размеры мозжечка приближаются к его размерам у взрослого
(150 г). Кроме того, быстрое развитие мозжечка происходит и в период полового
созревания.
Серое и белое вещество мозжечка развивается неодинаково. До 7 лет
количество серого вещества увеличивается приблизительно в 2 раза, а белого –
почти в 5 раз.
Из ядер мозжечка раньше других формируется зубчатое ядро. До первых лет
жизни детей, ядерные образования выражены лучше, чем нервные волокна.
Клеточное строение коры мозжечка у новорожденного значительно отличается от
взрослого. Ее клетки во всех слоях отличаются по форме, размерам и количеству
отростков. У новорожденного еще не полностью сформированы клетки
Пуркинье, в них не развито тигроидное вещество, ядро почти полностью
занимает клетку, ядрышко имеет неправильную форму, дендриты клеток
слаборазвиты. Клеточные слои значительно тоньше. Полное формирование
клеточных структур мозжечка осуществляется к 7-8 годам.
34. Мозжечок
• Расположение: Мозжечок располагается надпродолговатым мозгом и варолиевым мостом .
• Функции: являясь частью комплексного
регуляторного механизма с обратной связью, мозжечок
выполняет роль координационного центра,
обеспечивающего равновесие тела и поддержание
мышечного тонуса. Как отмечает Петер Дуус (P. Duus,
1995, мозжечок обеспечивает возможность выполнения
дискретных и точных движений, отслеживая и
координируя сенсорную информацию на входе и
моделируя моторные сигналы на выходе.
• Серое вещество формирует кору мозжечка и
находящиеся в его глубине парные ядра (подкорковые):
зубчатые ядра (1) (расположены в полушариях) –
регуляция работы мышц конечностей;
• ядра шатра (3) (в центральной части червя) –
регуляция работы вестибулярного аппарата;
шаровидные (4) и пробковидные ядра (2) (между
зубчатыми и ядрами шатра) – регуляция работы мышц
туловища.
1 – борозды
2 – извилины
3 – кора
4 – белое вещество
35. Мозжечок. Кора
Кора мозжечка является нервнымцентром экранного типа и характеризуется
высокой упорядоченностью расположения
нейронов, нервных волокон и глиальных
клеток.
В ней различают три слоя (снаружи
внутрь):
• молекулярный слой, содержащий
сравнительно небольшое количество мелких
клеток;
• ганглионарный слой, образованный одним
рядом тел крупных грушевидных клеток
(клеток Пуркинье);
• зернистый слой, с большим количеством
плотно лежащих клеток.
• Помимо нейронов в каждом слое
присутствуют глиоциты и сосуды.
1.
2.
3.
4.
1
2
3
молекулярный слой
ганглионарный слой
зернистый слой
белое вещество
36. Кора мозжечка
Молекулярный слой по ширине - самыйбольшой, а по концентрации нейронов – самый
бедный. Содержит тела корзинчатых и звездчатых
клеток (коротко- и длинноаксонных), дендриты
которых образуют синапсы с аксонами клеток-зерен.
Корзинчатые
клетки
располагаются
во
внутренней части молекулярного слоя. Это
мультиполярные нейроны неправильной формы и
небольших размеров. Их короткие дендриты образуют
связи с параллельными волокнами в наружной части
молекулярного слоя, а длинный аксон идет
параллельно поверхности извилины, отдавая через
определенные интервалы коллатерали, которые
спускаются к телам клеток Пуркинье (способны
охватывать до 240 клеток) и, разветвляясь,
охватывают их наподобие корзинок, образуя
тормозные аксо-соматические синапсы.
Звездчатые клетки — мелкие нейроны, тела
которых лежат выше тел корзинчатых клеток. У
короткоаксонных звездчатых клеток дендриты
образуют связи с параллельными волокнами, а
разветвления аксона формируют тормозные синапсы
на дендритах клеток Пуркинье. У длинноаксонных
звездчатых клеток аксон может участвовать в
образовании корзинки вокруг тела клетки Пуркинье.
37. Кора мозжечка
Ганглионарный слой содержит лежащие в один ряд тела клетокПуркинье (грушевидных нейронов), оплетенные коллатералями аксонов
корзинчатых клеток ("корзинками").
Клетки Пуркинье (грушевидные нейроны) — крупные клетки с телом
грушевидной формы, содержащим хорошо развитые органеллы. Их
количество у человека составляет 30 млн, размер – 35-60 мкм. От него в
молекулярный слой отходят 2—3 первичные (стволовые) дендрита,
интенсивно ветвящиеся в плоскости, перпендикулярной направлению
извилины, с образованием конечных (терминальных) дендритов,
достигающих поверхности молекулярного слоя. На дендритах находятся
60—100 тыс. шипиков — контактных зон возбуждающих синапсов,
образуемых параллельными волокнами (аксонами клеток-зерен) и
возбуждающих синапсов, образуемых лазящими волокнами.
Аксон клетки Пуркинье отходит от основания ее тела, одевается
миелиновой оболочкой, пронизывает зернистый слой и проникает в белое
вещество, являясь единственным эфферентным путем его коры,
заканчивается на нейронах ядер мозжечка. Аксоны небольшой части клеток
направляется в вестибулярные ядра продолговатого мозга и моста. По ходу
аксон отдает коллатерали, которые возвращаются в область расположения
тел клеток Пуркинье и образуют тормозные синапсы на телах соседних
клеток Пуркинье и клеток Гольджи.
Количество клеток Пуркинье заметно снижается при старении : на 2040% к 70—90 годам (по сравнению с их числом у 40—50 летних), что,
вероятно, служит одной из причин нарушения функции мозжечка у
пожилых людей.
Клетки Пуркинье
38. Кора мозжечка
Зернистый слой содержит близко расположенные тела клеток-зерен, больших клеток-зерен (клетокГольджи), веретеновидных горизонтальных нейронов, а также клубочки мозжечка — особые округлые
сложные синаптические контактные зоны между моховидными волокнами, дендритами клеток-зерен и аксонами
больших клеток-зерен.
Клетки-зерна — мелкие (4 мкм) и наиболее многочисленные нейроны коры мозжечка со слабо развитыми
органеллами и короткими дендритами, имеющими вид "птичьей лапки", на которых в клубочках мозжечка
розетки моховидных волокон образуют многочисленные синаптические контакты. Аксоны клеток-зерен
направляются в молекулярный слой, где Т-образно делятся на две ветви, идущие параллельно длине извилины
(параллельные волокна), образуя возбуждающие синапсы на дендритах клеток Пуркинье, корзинчатых,
звездчатых клеток и больших клеток-зерен. Через дендритное дерево каждой клетки Пуркинье проходит до 200300 тыс. параллельных волокон, образуя на каждой клетке 60-100 тыс. синапсов (не все волокна образуют
синапсы). Аксон каждой клетки-зерна образует связи с дендритами 250-500 клеток Пуркинье.
Большие клетки-зерна (клетки Гольджи)
крупнее клеток-зерен, содержат хорошо развитые
органеллы. Их аксоны в пределах клубочков мозжечка
образуют синапсы на дендритах клеток-зерен, а
длинные дендриты поднимаются в молекулярный слой,
где ветвятся и образуют связи с параллельными
волокнами. Большие клетки-зерна оказывают
угнетающее влияние на активность клеток-зерен.
Веретеновидные горизонтальные клетки имеют
небольшое вытянутое тело, от которого в обе стороны
отходят длинные горизонтальные дендриты,
заканчивающиеся в ганглионарном и зернистом слоях.
Аксоны этих клеток дают коллатерали в зернистый
слой и уходят в белое вещество.
Клубочек мозжечка – скопления
терминальных ветвлений отростков различных
нейронов мозжечка и моховидных волокон.
Окружен глиальной капсулой. Вокруг клубочка
расположены клетки-зерна.
39. Мозжечок Белое вещество. Миелоархитектоника
Афферентные волокна коры мозжечка включают моховидные (мшистые) илазящие.
Моховидные (мшистые) волокна мозжечка проходят в составе спинно- и
мостомозжечковых путей и, разветвляясь, заканчиваются расширениями (розетками)
в особых контактных зонах — клубочках мозжечка, образуя синаптические контакты
с дендритами клеток-зерен, на которых оканчиваются также и аксоны больших
клеток-зерен. Клубочки мозжечка снаружи не полностью окружены плоскими
отростками астроцитов.
Лазящие (лиановидные) волокна мозжечка идут в составе оливомозжечковых
путей и проникают в кору из белого вещества, проходя через зернистый слой до
ганглионарного и заканчиваются возбуждающими синапсами на телах и дендритах
клеток Пуркинье. Коллатеральные ветки лазящих волокон образуют синапсы на
других нейронах всех типов, включая клетки-зерна, клетки Гольджи, звездчатые и
корзинчатые клетки. С каждой клеткой Пуркинье обычно контактирует одно лазящее
волокно.
Эфферентные волокна коры мозжечка представлены аксонами клеток
Пуркинье, которые в виде миелиновых волокон направляются в белое вещество и
достигают глубоких ядер мозжечка и вестибулярного ядра, образуя на их нейронах
тормозные синапсы (клетки Пуркинье являются тормозными нейронами).
40. Мозжечок. Межнейрональные связи в коре мозжечка
Межнейронные связи в коре мозжечка обеспечиваютпереработку поступающей в нее разнообразной
сенсорной информации. Возбуждающие импульсы
поступают в кору мозжечка по лазящим и моховидным
волокнам. В первом случае возбуждение передается на
дендриты клеток Пуркинье непосредственно, во втором
— через клубочки мозжечка — на дендриты клетокзерен и далее по их аксонам (параллельным волокнам).
Последние образуют возбуждающие синапсы также на
дендритах корзинчатых и звездчатых клеток и больших
клеток-зерен. Аксоны корзинчатых клеток образуют
тормозные синапсы на телах клеток Пуркинье, а аксоны
звездчатых клеток на их дендритах. Аксоны больших
зернистых клеток в клубочках мозжечка образуют
тормозные синапсы на дендритах клеток-зерен.
Сформированные в коре мозжечка тормозные сигналы
передаются с клеток Пуркинье на ядра мозжечка и
вестибулярные ядра, а через них в конечном итоге
контролируют активность нисходящих двигательных
путей.
Клетки:
1- клетки Пуркинье
2- звездчатые клетки
3-корзинчатые клетки
4- зерновидные клетки
5- звездчатые клетки Гольджи
Волокна:
6- моховидные волокна
7- лазящие волокна
«+» – активирующее
(возбуждающее) влияние
«-» - тормозное влияние
41. Мозжечок. Глия
Глиальные элементы коры мозжечка обеспечивают функции нейронов,располагаются во всех ее слоях и весьма разнообразны; они включают:
• олигодендроциты (участвуют в образовании миелиновых оболочек
нервных волокон),
• Астроциты (своими уплощенными на концах отростками образуют
периваскулярные пограничные мембраны - компонент гематоэнцефалического барьера, и оболочки вокруг клубочков мозжечка.
• В ганглионарном слое между грушевидными нейронами лежат особые
астроциты с темными ядрами - клетки Бергмана. Отростки этих
клеток направляются к поверхности коры и образуют глиальные
волокна молекулярного слоя мозжечка (волокна Бергмана),
формируют поверхностную пограничную глиальную мембрану,
окружают и поддерживают дендриты клеток Пуркинье.
• Микроглия в большом количестве содержится в молекулярном и
ганглионарном слоях
42. Головной мозг
43. Головной мозг
Кора больших полушарий мозга - сложно организованный нервный центр экранного типа,обеспечивает регуляцию разнообразных функций организма и сложные формы поведения.
Кора образована слоем серого вещества толщиной 3—5 мм на поверхности извилин (30 %) и в
глубине борозд (70 %) общей площадью 1500—2500 см2 при объеме около 300 см3. Серое вещество
содержит нервные клетки (около 10—15 млрд), нервные волокна и клетки нейроглии (более 100 млрд).
На основании различий плотности расположения и строения клеток (цитоархитектоники), хода
волокон (миелоархитектоники) и функциональных особенностей различных участков коры в ней
выделяют 52 нерезко разграниченные поля.
А, Б. Расположение клеток
(цитоархитектоника).
В. Расположение миелиновых
волокон (миелоархитектоника).
1. Молекулярный слой.
2. Наружный зернистый слой.
3. Пирамидный слой.
4. Внутренний зернистый слой.
5. Ганглионарный слой.
6. Слой полиморфных клеток.
7. Наружная полоска Байарже.
8 Внутренняя полоска Байарже.
44. Головной мозг Кора больших полушарий
Среди нейронов коры мозга различают ассоциативные – связывающие участки коры внутриодного полушария, комиссуральные – их аксоны идут в другое полушарие, и проекционные –
их аксоны идут в нижележащие отделы мозга.
Нейроны коры — мультиполярные, различных размеров и форм, включают более 60 видов,
среди которых выделены два основных типа — пирамидные и непирамидные. Пирамидные
клетки — специфический для коры полушарий тип нейронов; составляет 50—90 % всех
нейроцитов коры. От апикального полюса их конусовидного (на срезах — треугольного) тела,
который обращен к поверхности коры, отходит длинный (апикальный) покрытый шипиками
дендрит, направляющийся в молекулярный слой коры, где он ветвится. От базальной и
латеральных частей тела вглубь коры и в стороны от тела нейрона расходятся 5—16 более
коротких боковых (латеральных) дендритов, которые, ветвясь, распространяются в пределах того
же слоя, где находится тело клетки. От середины базальной поверхности тела отходит длинный и
тонкий аксон. Он тянется в белое вещество, на расстоянии 60—90 мкм начинает давать
коллатерали. Размеры пирамидных нейронов варьируются от 10 до 140 мкм; различают
гигантские, крупные, средние и малые пирамидные клетки.
Непирамидные клетки располагаются практически во всех слоя коры, воспринимая
поступающие афферентные сигналы, их аксоны распространяются в пределах самой коры,
передавая импульсы на пирамидные нейроны. Эти клетки весьма разнообразны и
преимущественно являются разновидностями звездчатых клеток. Они включают шипиковые,
звездчатые, корзинчатые, аксоаксональные клетки, клетки—"канделябры", клетки с двойным
букетом дендритов, горизонтальные клетки Кахаля, клетки Мартинотти и другие. Основная
функция непирамидных клеток —интеграция нейронных цепей внутри коры.
45. Головной мозг Цитоархитектоника коры больших полушарий
Специфическую организацию нейронов называют цитоархитектоникой. Так, всенсорных зонах коры пирамидный и ганглиозный слои выражены слабо, а зернистые слои
- хорошо. Такой тип коры называется гранулярным. В моторных зонах, напротив,
зернистые слои развиты плохо (II и IV), а пирамидные хорошо (III, V и VI ). Это
агранулярный тип коры.
I. Молекулярный слой располагается под мягкой мозговой оболочкой; содержит
небольшое число мелких нейронов — горизонтальных клеток Кахаля с длинными
ветвящимися дендритами, отходящими в горизонтальной плоскости от веретеновидного
тела. Их аксоны участвуют в образовании тангенциального сплетения волокон этого слоя. В
молекулярном слое имеются многочисленные дендриты, и аксоны клеток более глубоко
расположенных слоев, образующих межнейронные связи.
II. Наружный зернистый слой образован многочисленными мелкими пирамидными и
звездчатыми клетками, дендриты которых ветвятся и поднимаются в молекулярный слой, а
аксоны либо уходят в белое вещество, либо образуют дуги и также направляются в
молекулярный слой.
III. Пирамидный слой значительно варьирует по ширине и максимально выражен в
ассоциативных и сенсомоторных областях коры. В нем преобладают пирамидные клетки,
размеры которых увеличиваются вглубь слоя от мелких до крупных. Апикальные дендриты
пирамидных клеток направляются в молекулярный слой, а латеральные образуют синапсы с
клетками данного слоя. Аксоны этих оканчиваются в пределах серого вещества или
направляются в белое. Помимо пирамидных клеток, слой содержит разнообразные
непирамидные нейроны. Слой выполняет преимущественно ассоциативные функции,
связывая клетки как в пределах данного полушария, так и с противоположным полушарием.
46. Головной мозг Цитоархитектоника коры больших полушарий
IV. Внутренний зернистый слой широкий в зрительной и слуховой областях коры, а всенсомоторной области практически отсутствует. Он образован мелкими пирамидными и
звездчатыми клетками. В этом слое заканчивается основная часть таламических
(шипиковых) афферентных волокон. Аксоны клеток этого слоя образуют связи с клетками
выше- и нижележащих слое коры.
V. Ганглионарный слой образован крупными, а в области моторной коры
(прецентральной извилины) — гигантскими пирамидными клетками (Беца). Апикальные
дендриты пирамидных клеток достигают I слоя, образуя там верхушечные букеты,
латеральные дендриты распространяются в пределах того же слоя. Аксоны гигантских и
крупных пирамидных клеток проецируются на ядра головного и спинного мозга, наиболее
длинные из них в составе пирамидных путей достигают каудальных сегментов спинного
мозга. В V слое сосредоточено большинство корковых проекционных эфферентов.
VI. Слой полиморфных клеток образован разнообразными по форме нейронами
(веретеновидными, звездчатыми, клетками Мартинотти). Наружные участки слоя
содержат более крупные клетки, внутренние — более мелкие и редко расположенные.
Аксоны этих клеток уходят в белое вещество в составе эфферентных путей, а дендриты
проникают до молекулярного слоя. Аксоны мелких клеток Мартинотти поднимаются к
поверхности коры и ветвятся в молекулярном слое.
47. Головной мозг Миелоархитектоника коры больших полушарий
Нервные волокна коры полушарий большого мозга включаюттри группы:
• афферентные;
• ассоциативные и комиссуральные;
• эфферентные волокна.
Афферентные волокна в виде пучков в составе радиальных
лучей (рл) приходят в кору от ниже расположенных отделов
головного мозга, в частности, от зрительных бугров и коленчатых
тел. Большая часть этих волокон заканчивается на уровне IV слоя.
Ассоциативные и комиссуральные волокна —
внутрикорковые волокна, которые соединяют между собой
различные области коры в том же или в другом полушариях,
соответственно. Эти волокна образуют пучки, которые проходят
параллельно поверхности коры в I слое (тангенциальные волокна,
тв), во II слое (полоска Бехтерева, пб), в IV слое (наружная
полоска Байярже, нпб) и в V слое (внутренняя полоска Байярже,
впб). Последние две системы являются сплетениями,
образованными конечными отделами афферентных волокон.
Эфферентные волокна связывают кору с подкорковыми
образованиями. Эти волокна идут в нисходящем направлении в
составе радиальных лучей (рл) (например, пирамидные пути).
тв
I
пб
II
III
IV
нпб
V
впб
IV
рл
48. Модуль коры головного мозга
Модуль - это совокупность нейроцитов всех 6-ти слоев,расположенных на одном перпендикулярном пространстве и
тесно взаимосвязанных между собой и подкорковыми
образованьями. В пространстве модуль можно представить
как цилиндр, пронизывающий все 6 слоев коры,
ориентированный своей длинной осью перпендикулярно к
поверхности коры и имеющий диаметр около 300 мкм. В
коре БПШ человека насчитывается около 3 млн. модулей. В
каждом модуле содержится до 2 тысяч нейроцитов. Каждый
модуль включает афферентные пути, систему локальных
связей и эфферентные пути.
К афферентным путям относятся кортико-кортикальные и
таламо-кортикальные волокна.
Модуль организован вокруг кортико-кортикальных волокон,
представляющих собой аксоны пирамидных клеток либо
этого же полушария, либо противоположного. Кортикокортикальные волокна образуют окончания во всех слоях
коры данного модуля.
В модуль входят также таламо-кортикальные волокна,
оканчивающиеся в IV слое коры на шипиковых звездчатых
нейронах и базальных дендритах пирамидных нейронов.
Эфферентные пути формируются аксонами крупных и
гигантских пирамидных нейронов, а также аксонами
веретеновидных и некоторых других клеток VI слоя коры.
49.
Модуль коры головного мозга(продолжение)
Система локальных связей формируется вставочными нейронами колонки, которые включают
более десятка типов клеток. Часть из них обладает тормозной функцией и регулирует
преимущественно активность пирамидных клеток.
Из тормозных нейронов колонки наибольшее значение имеют:
аксо-аксональные клетки, тела которых лежат во II и III слоях, а аксоны идут горизонтально,
отдавая многочисленные терминальные веточки, которые образуют тормозные синапсы на
начальных сегментах аксонов пирамидных клеток II и III слоев;
клетки-"канделябры" встречаются во всех внутренних слоях коры. Их аксонные коллатерали
идут горизонтально и дают несколько восходящих и нисходящих веточек, которые образуют
спиральные ветвления вокруг апикальных дендритов пирамидных клеток;
корзинчатые клетки, которые располагаются во II слое, на границе III и IV, а также IV и V
слоев. Их аксоны проходят горизонтально на расстояние до 2—3 мм и, оплетая тела крупных и
средних пирамидных клеток, влияют на 20—30 соседних колонок. Колонковые корзинчатые
клетки обеспечивают торможение пирамидных клеток по вертикали внутри данной колонки;
клетки с двойным букетом дендритов, отходящих вертикально от полюсов тела,
расположенного во II—III слоях. Их аксон дает коллатерали, образующие контакты с дендритами
как пирамидных клеток, так и непирамидных (в том числе тормозных) нейронов. Первый тип
контактов опосредует угнетение пирамидных клеток, а второй — их активацию путем снятия
торможения;
клетки с аксонным пучком (кисточкой) — звездчатые нейроны II слоя, аксоны которых
ветвятся в I слое, образуя связи с дистальными сегментами апикальных дендритов пирамидных
клеток и с горизонтальными ветвями кортико-кортикальных волокон.
50. Глия головного мозга
Головной мозг содержит все виды макроглии (астроцитарную, эпендимную и олигодендроглию), а такжемикроглию.
Астроцитарная глия обеспечивает микроокружение нейронов, выполняет опорную и трофическую
функции в сером и белом веществе, участвует в метаболизме нейромедиаторов. Астроциты уплощенными
пластинчатыми концевыми участками своих отростков образуют три вида пограничных глиальных мембран:
периваскулярные, поверхностную и субэпендимальную.
Периваскулярные пограничные мембраны окружают капилляры головного мозга и входят в состав гематоэнцефалического барьера, отделяющего нейроны центральной нервной системы от крови и тканей внутренней
среды. Гемато-энцефалический барьер препятствует проникновению в центральную нервную систему
переносимых кровью токсических веществ, нейромедиаторов, гормонов, антибиотиков (что затрудняет лечение
инфекционных поражений мозга и его оболочек), поддерживает электролитный баланс мозга, обеспечивает
избирательный транспорт ряда веществ (глюкозы, аминокислот) из крови в мозг.
Гемато-энцефалический барьер включает в себя следующие компоненты:
эндотелий кровеносных капилляров (с непрерывной выстилкой)главный компонент гемато-энцефалического
барьера. Его клетки связаны мощными плотными соединениями, образование которых индуцируется контактом с
астроцитами. Эндотелий препятствует переносу одних веществ, содержит специфические транспортные системы
для других и метаболически изменяет третьи, превращая их в соединения, неспособные проникнуть в мозг;
базальную мембрану капилляров;
периваскулярную пограничную глиальную мембрану из отростков астроцитов.
Поверхностная пограничная глиальная мембрана (краевая глия) мозга, расположена под мягкой мозговой
оболочкой, образует наружную границу головного и спинного мозга, отделяя ткани центральной нервной
системы от мозговых оболочек.
Субэпендимальная (перивентрикулярная) пограничная глиальная мембрана располагается под слоем
эпендимы и входит в состав нейро-ликворного барьера, который отделяет нейроны от спинномозговой жидкости,
называемой также ликвором. Этот барьер представлен эпендимной глией, ее базальной мембраной (присутствует
не везде) и отростки астроцитов.
Эпендимная глия образует выстилку желудочков головного мозга и входит в состав гематоликворного
барьера (между кровью и спинномозговой жидкости).
Олигодендроглия встречается в сером и белом веществе; она обеспечивает барьерную функцию, участвует в
формировании миелиновых оболочек нервных волокон, регулирует метаболизм нейронов, захватывает
нейромедиаторы.
Микроглия — специализированные макрофаги центральной нервной системы, обладающие значительной
подвижностью. Активируется при воспалительных и дегенеративных заболеваниях. Выполняет в центральной
нервной системе роль антиген-представляющих дендритных клеток.
51.
1Гематонейральныйбарьер часто называют гематоэнцефалическим
52.
Оболочки головного и спинного мозгаГоловной и спинной мозг покрыты тремя соединительнотканными оболочками: внутренней мягкой,
средней паутинной и внешней твердой. Мозговые оболочки выполняют защитную, в т.ч.
амортизирующую, функцию, обеспечивают выработку и всасывание ликвора.
Мягкая мозговая оболочка прилежит к ткани мозга и отграничена от нее краевой глиальной
мембраной. В РВСТ оболочки имеются большое количество кровеносных сосудов, питающих мозг,
нервных волокон, концевые аппараты и одиночные нервные клетки. Мягкая мозговая оболочка окружает
сосуды, проникающие в мозг, образуя вокруг них периваскулярную глиальную мембрану. В желудочках
мозга мягкая мозговая оболочка совместно с эпендимой принимает участие в образовании сосудистых
сплетений, вырабатывающих ликвор.
Паутинная оболочка представлена тонким слоем РВСТ. Между ней и мягкой мозговой оболочкой
лежит сеть перекладин из тонких пучков коллагеновых и тонких эластических волокон, связывающая
оболочки между собой. Между мягкой мозговой оболочкой, повторяющей рельеф ткани мозга, и
паутинной располагается подпаутинное (субарахноидальное) пространство, пронизанное тонкими
коллагеновыми и эластическими волокнами. Субарахноидальное пространство сообщается с
желудочками мозга и содержит цереброспинальную жидкость. В этом пространстве проходят крупные
кровеносные сосуды, ветви которых питают мозг.
Ворсинки паутинной оболочки (наиболее крупные называются пахионовы грануляции) служат
участками, через которые вещества из ликвора возвращаются в кровь. Они представляют собой
бессосудистые выросты паутинной оболочки с сетью щелевидных пространств, выпячивающиеся в
просвет синусов твердой мозговой оболочки.
Твердая мозговая оболочка образована ПВСТ, содержащей много эластических волокон. В
полости черепа она плотно сращена с надкостницей, в спинномозговом канале отграничена от периоста
позвонков эпидуральным пространством, заполненным слоем РВСТ, что обеспечивает ей некоторую
подвижность. Между твердой мозговой и паутинной оболочками располагается субдуральное
пространство. В субдуральном пространстве содержится небольшое количество жидкости.
Оболочки со стороны субдурального и субарахноидального пространства покрыты слоем плоских
клеток глиальной природы.