












































 biology
biologySimilar presentations:










Фотосинтез
1.
Глава 10Фотосинтез
PowerPoint® Lecture Presentations for
Biology
Eighth Edition
Neil Campbell and Jane Reece
Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
2.
Ознакомление• Фотосинтез-процесс превращения
солнечной энергии в химическую
• Фотосинтез – кормит всю живую планеты,
прямым или косвенным способом
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
3.
• Автотрофы поддерживать себя без потребленияорганических веществ полученных от других
организмов
• Автотрофы являются производителями биосферы,
которые получяют органические молекулы из CO2 и
других неорганических молекул
• Почти все растения являются фотоавтотрофами,
используя энергию солнечного света для получения
органических молекул из Н2О и СО2
Copyright © 2008 Pearson Education, Inc., publing as Pearson Benjamin Cummings
4.
Fig. 10-15.
• Фотосинтез происходит в растениях,водорослях, некоторых других простейших и
некоторых прокариот
• Эти организмы питают не только себя, но и
большую часть живого мира
BioFlix: Photosynthesis
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
6.
Fig. 10-2(a)
Растения
(c) Одноклеточные
протисты
10 µm
(e) Серобактерий
(b) Многоклеточные
водоросли
(d) Цианобактерии
40 µm
1.5 µm
7.
• Гетеротрофы получают органическиевещества из других организмов
• Гетеротрофы являются потребителями
биосферы
• Почти все гетеротрофы, включая человека,
зависят от фотоавтотрофов которые в свою
очередь являются источником питания и О2
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
8.
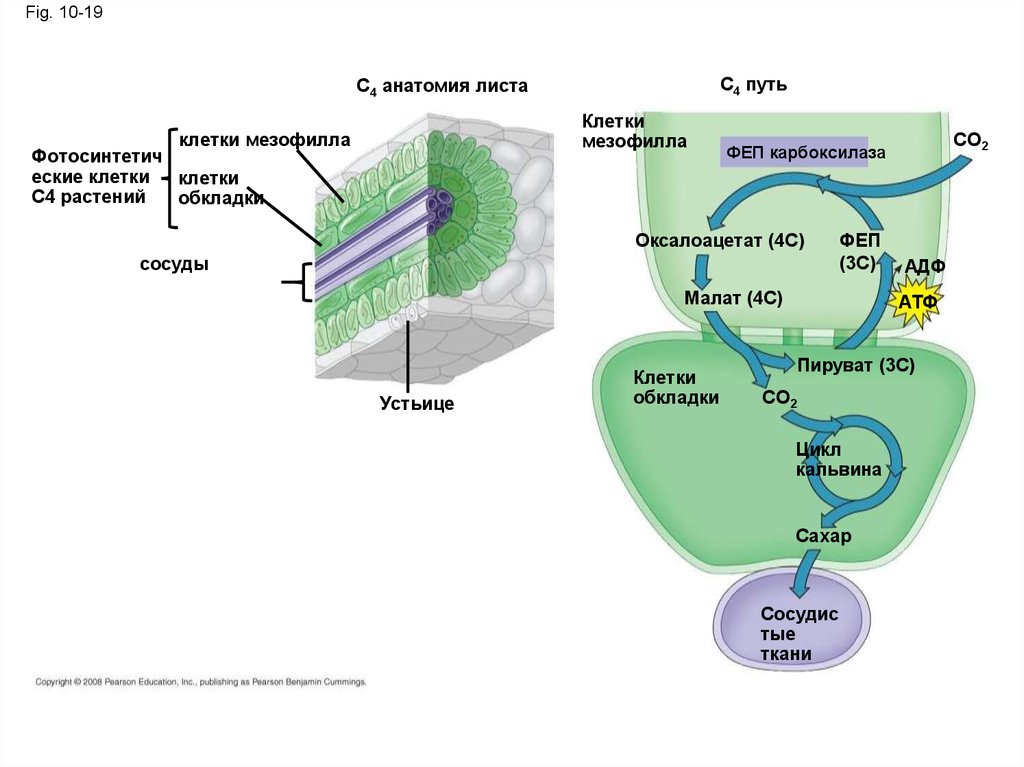
Концепция 10.1: Фотосинтез преобразуетсветовую энергию в химическую энергию пищи
• Хлоропласты, структурно похож на и,
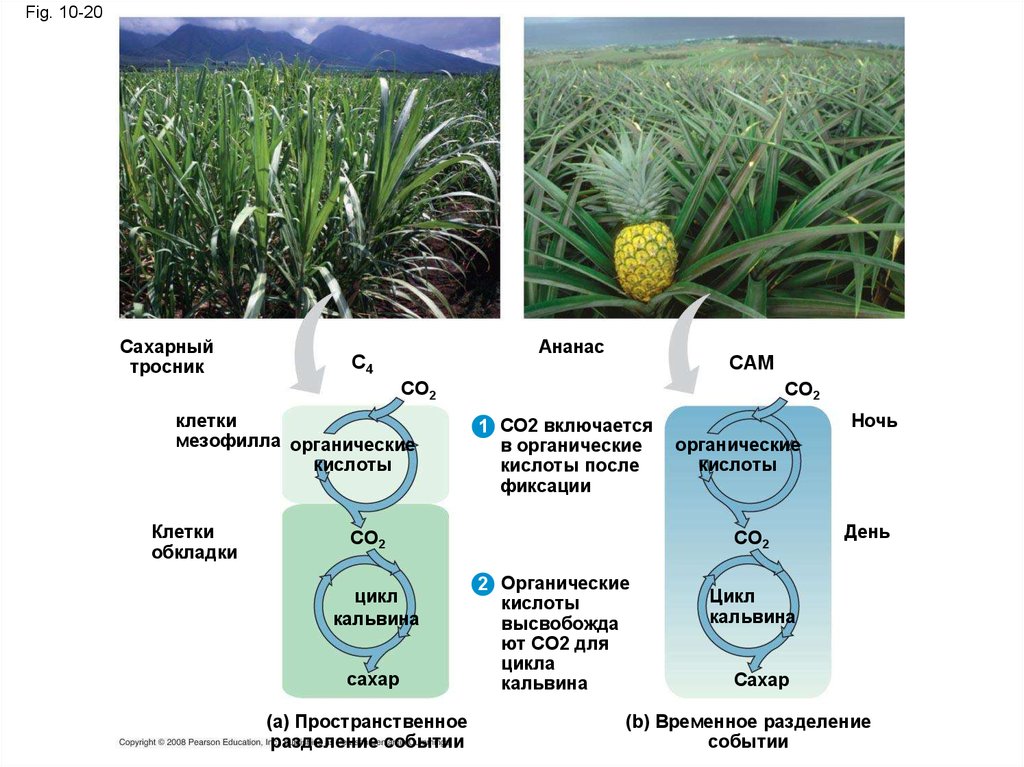
вероятно, произошли от
фотосинтезирующих бактерий
• Структурная организация этих клеток
позволяет химические реакций фотосинтеза
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
9.
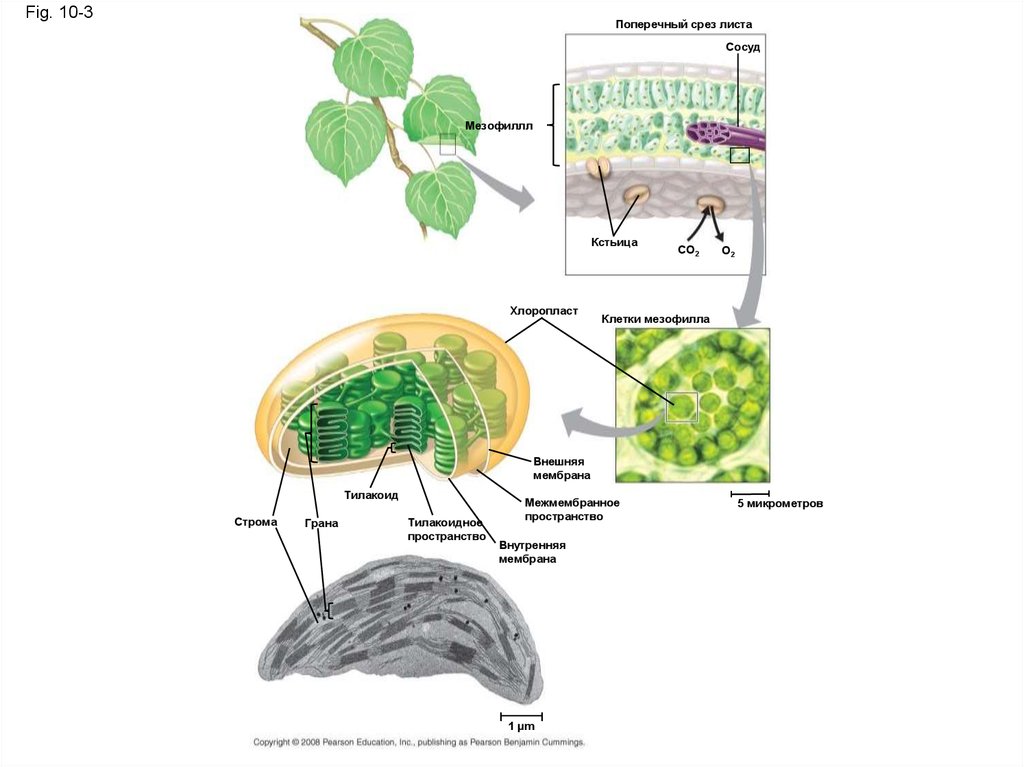
Хлоропласты - сайт фотосинтеза растении• Листья - основное место для фотосинтеза
• Их зеленый цвет исходит из хлорофилла,
зеленого пигмента в хлоропластах
• Световая энергия поглощаемая
хлорофиллом позволяет синтез
органических молекул в хлоропластах
• СО2 поступает и О2 выходит с листьев
через микроскопические поры на листьях
называемых устьицами
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
10.
• Хлоропласты находятся в основном в клеткахмезофилла, во внутренней ткани листа
• Типичная клетка мезофилла имеет 30-40 хлоропластов
• Хлорофилл находятся на мембранах тилакоидов
(связанные мешочки в хлоропластах); тилакоиды
может быть сложены в колонки называемые гранами
• Хлоропласты содержат строму, плотную жидкость
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
11.
Fig. 10-3Поперечный срез листа
Сосуд
Мезофиллл
Кстьица
Хлоропласт
CO2
O2
Клетки мезофилла
Внешняя
мембрана
Тилакоид
Строма
Грана
Тилакоидное
пространство
Межмембранное
пространство
Внутренняя
мембрана
1 µm
5 микрометров
12.
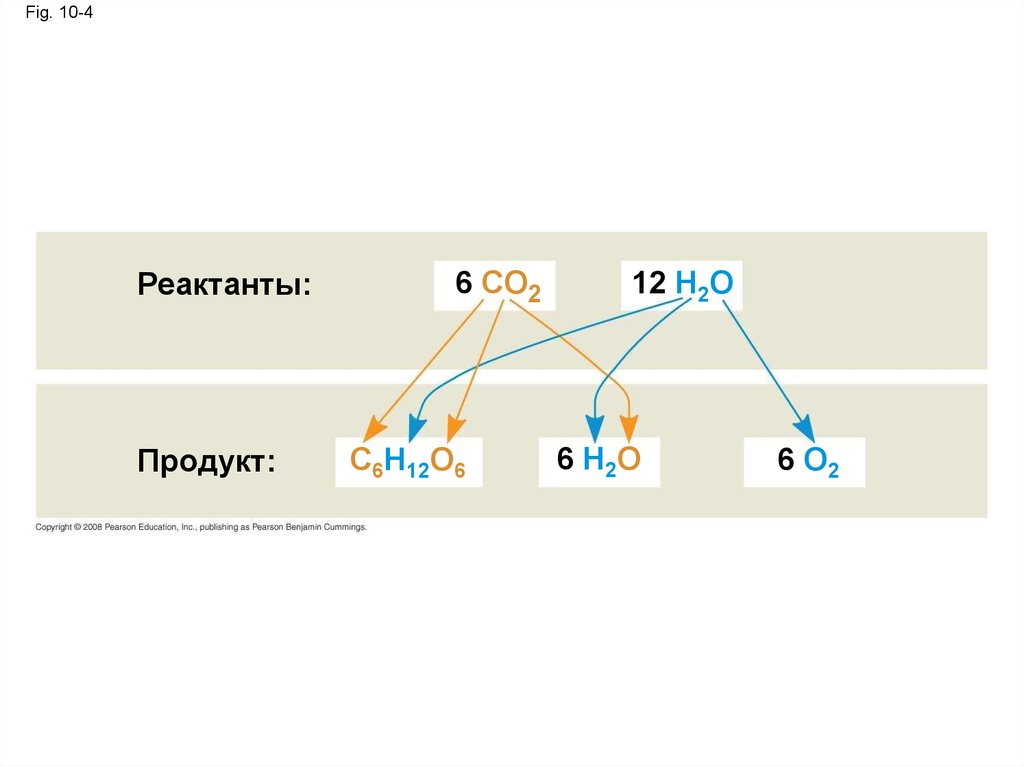
• Фотосинтез можно резюмироватьследующим уравнением:
6 CO2 + 12 H2O + энергия света → C6H12O6 + 6 O2 + 6 H2O
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
13.
Расщепление воды• Хлоропласты расщипляют H2O на водород
и кислород, и включают электроны
водорода в молекулы сахара
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
14.
Fig. 10-4Реактанты:
Продукт:
6 CO2
C6H12O6
12 H2O
6 H2O
6 O2
15.
Фотосинтез-окислительно-восстановительнаяреакция
• Фотосинтез является окислительно
восстановительной реакцией где Н2О
окисляется и СО2 восстанавливается
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
16.
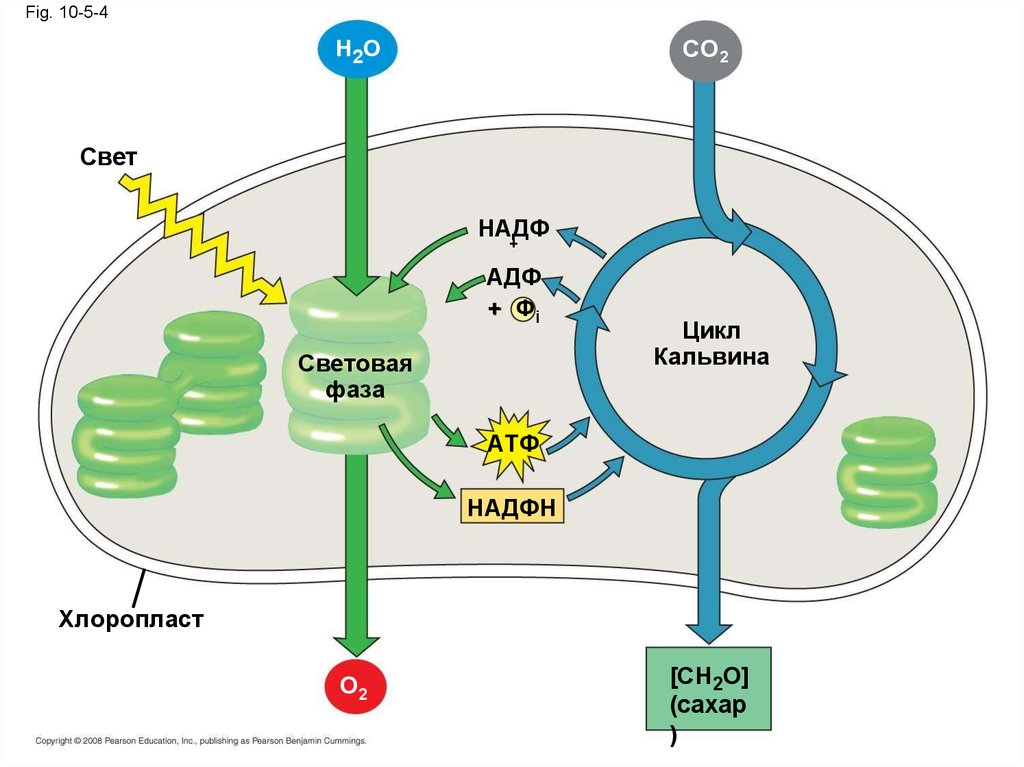
Две фазы фотосинтеза• Фотосинтез состоит из световой и темновой
фазы
• Световая фаза (в тилакоидах):
– Расщипление H2O
– Высвобождение O2
– Восстановление НАДФ+ в НАДФН
– Ситез AТФ из AДФ путем
фотофосфорилирования
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
17.
• Цикл Кальвина (в строме) образует сахар изCO2, используя AТФ и НАДФН
Цикл Кальвина начинает ся с фиксацией СО2
которая затем вкючается в состав
орнанических молекул
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
18.
Fig. 10-5-4CO2
H2O
Свет
НАДФ
+
AДФ
+ Фi
Световая
фаза
Цикл
Кальвина
AТФ
НАДФН
Хлоропласт
O2
[CH2O]
(сахар
)
19.
Концепция 10.2: Световая фаза превращяет солнечнуюэнергию в химическую энергию ATФ и НАДФН
• Хлоропласты являются солнечными
батареями растении
• Тилакоиды превращяют энергию света в
химическую энергию ATФ и НАДФН
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
20.
Природа света• Свет является формой электромагнитной энергии,
которая также называется электромагнитное излучение
• Как и другие электромагнитной энергии, свет проходит
по ритмичным волнам
• Длина волны является расстояние между гребнями
волн
• Длина волны определяет тип электромагнитной
энергии
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
21.
• Электромагнитный спектр это - весь спектрэлектромагнитной энергии, или излучения
• Видимый свет состоит из длин волн (в том
числе те, которые зажигают фотосинтез),
которые производят цвета, которые мы
можем видеть
• Свет ведет себя так, как будто оно состоит
из дискретных частиц, называемых
фотонами
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
22.
Fig. 10-610–5 нм 10–3нм
1 нм
Гамма
X-лучи
лучи
103 нм
У
Ф
106 нм
Инфрак
расные
1м
(109 нм)
Микров
олны
103 м
Радио
лучи
Видимый свет
380
450
500
Короткие волны
Юольше
энергии
550
600
650
750
nm
Длинные волны
Меньше
энергии
700
23.
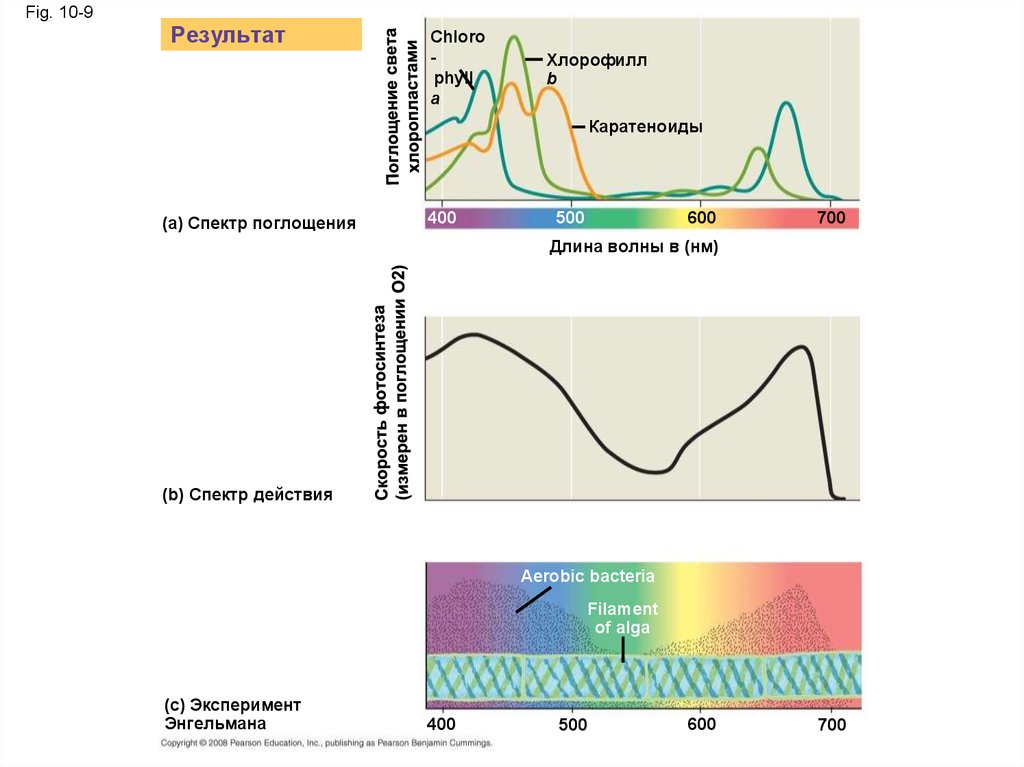
Фотосинтетические пигменты:световыерецепторы
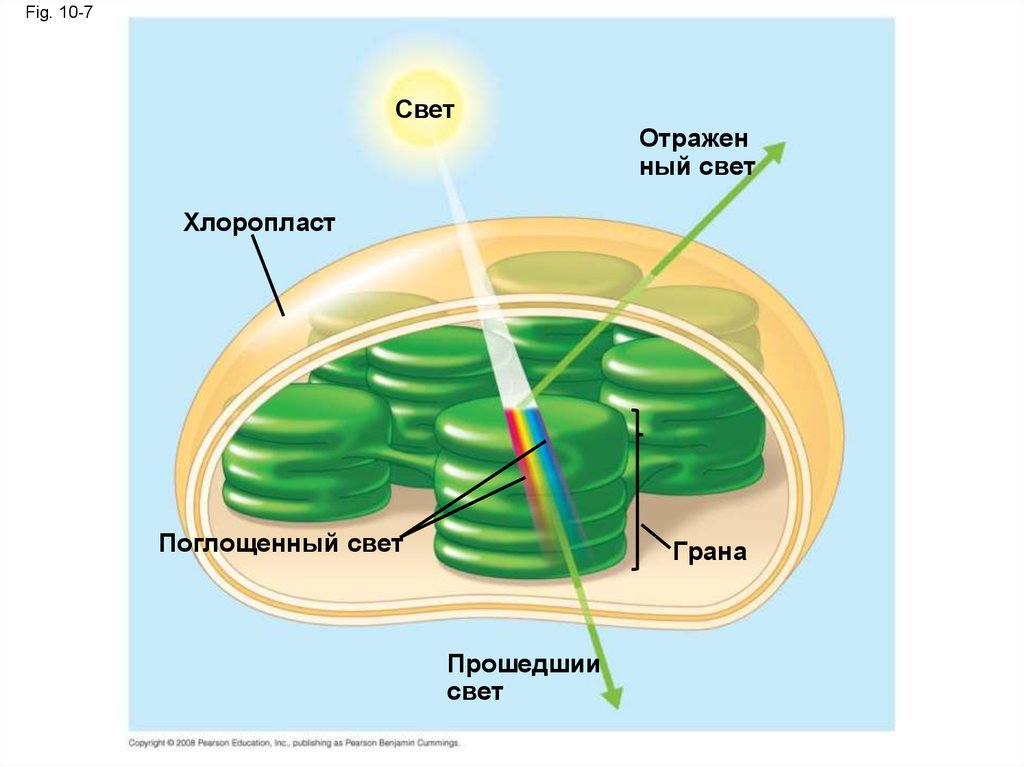
• Пигменты - молекулы, которые поглощают
видимый свет
• Различные пигменты поглощают различные
длины волн
• Длины волн, которые не поглощаются
отражаются или передаются
• Листья являются зелеными, потому что
хлорофилл отражает и передает зеленый
свет
Animation: Light and Pigments
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
24.
Fig. 10-7Свет
Отражен
ный свет
Хлоропласт
Поглощенный свет
Грана
Прошедшии
свет
25.
Fig. 10-9Результат
Chloro
phyll
a
Хлорофилл
b
Каратеноиды
(a) Спектр поглощения
400
500
600
700
Длина волны в (нм)
(b) Спектр действия
Aerobic bacteria
Filament
of alga
(c) Эксперимент
Энгельмана
400
500
600
700
26.
• Хлорофилл является основным пигментомфотосинтеза
• Вспомогательные пигменты, такие как
хлорофилл b, расшириют спектр,
используемый для фотосинтеза
• Вспомогательные пигменты - каротиноиды,
поглощают излишек света, которые могут
повредить хлорофилл
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
27.
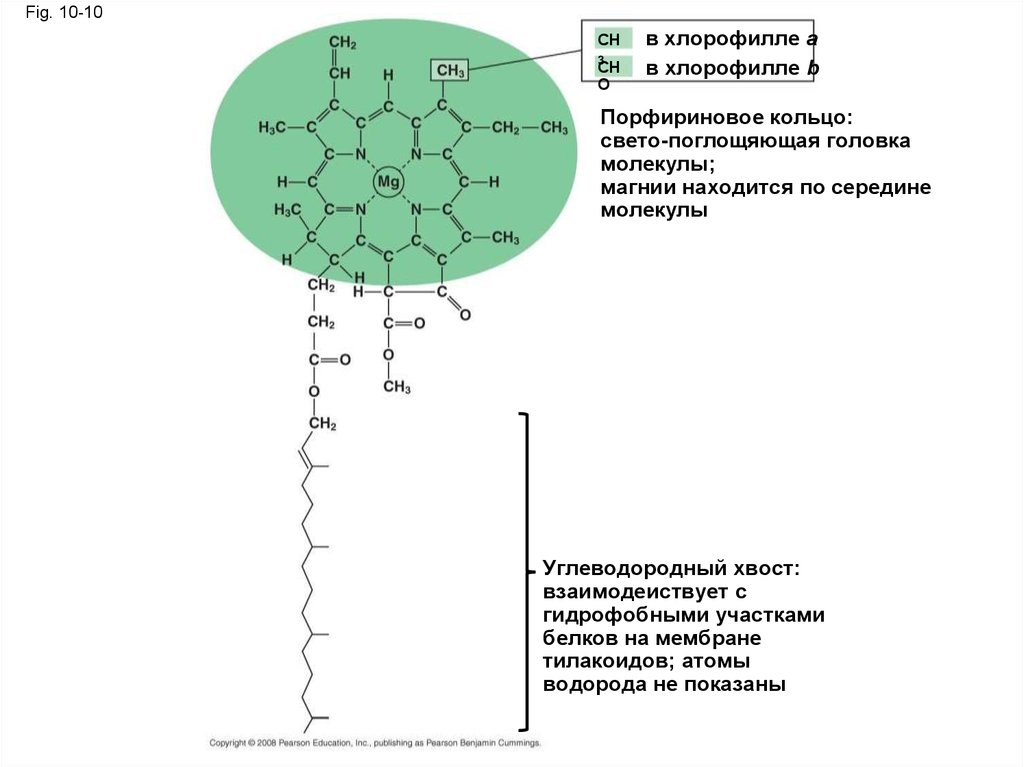
Fig. 10-10CH
3
CH
O
в хлорофилле a
в хлорофилле b
Порфириновое кольцо:
свето-поглощяющая головка
молекулы;
магнии находится по середине
молекулы
Углеводородный хвост:
взаимодеиствует с
гидрофобными участками
белков на мембране
тилакоидов; атомы
водорода не показаны
28.
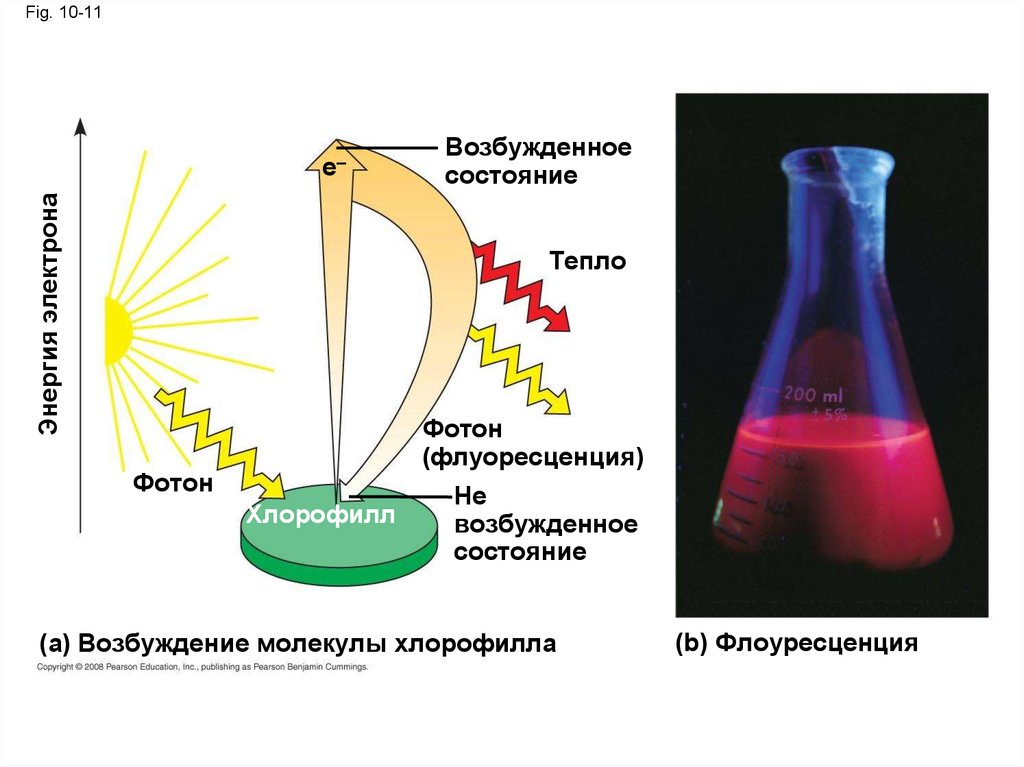
Возбуждение хлорофилла энергией света• Когда пигмент поглощает свет, он идет из не
возбужденного состояния в возбужденное
состояние, которое является неустойчивым
• Когда возбужденные электроны попадают
обратно в не возбужденное состояние,
фотоны выделяется, послесвечение
называется флуоресценции
• Светящийся хлорофилл высвечивается,
испуская свет и тепло
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
29.
Fig. 10-11Энергия электрона
e–
Возбужденное
состояние
Тепло
Фотон
(флуоресценция)
Фотон
Хлорофилл
Не
возбужденное
состояние
(a) Возбуждение молекулы хлорофилла
(b) Флоуресценция
30.
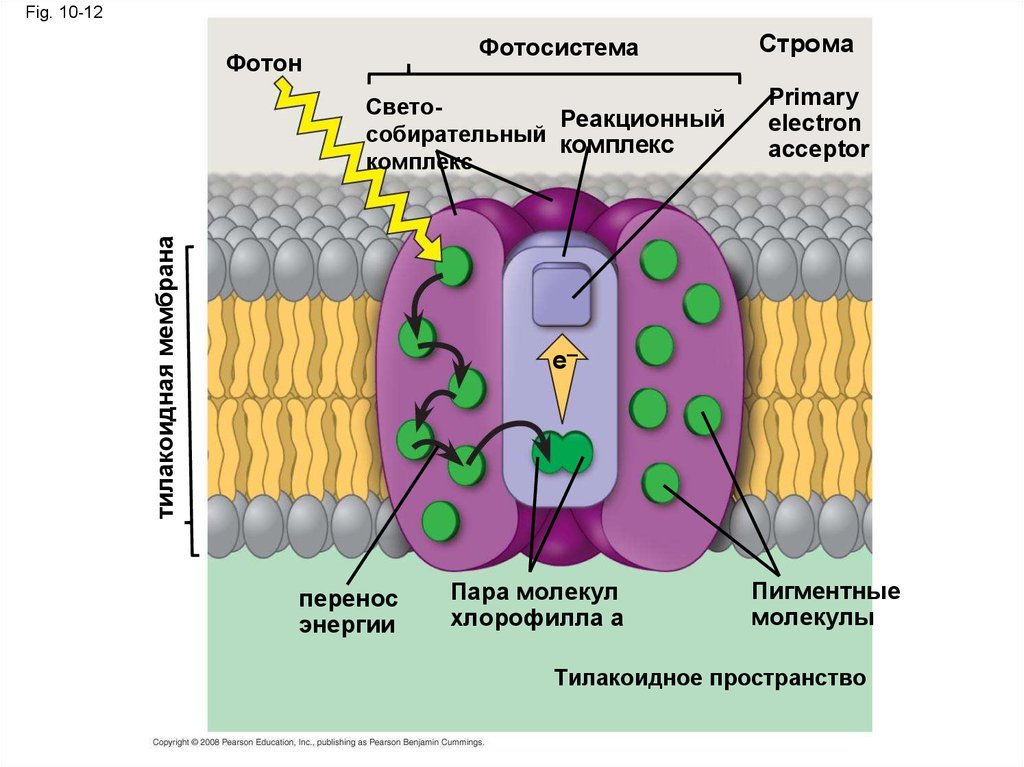
Фотосистема: Комплекс центра реакции связанныйсо свето-собирательным комплексом
• Фотосистема состоит из реакционного
центра (вид белкового комплекса)
окруженная свето-собирательным
комплексом
• Свето-собирательный комплекс (пигменты
прикрепленные к белкам) наводят энергию
фотонов на реакцонный центр
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
31.
• Первичный акцептор электронов вреакционном центре принимает
возбужденный электрон от хлорофилла а
• Перенос электрона от хлорофилла а на
первичный акцептор электронов является
первым шагом световой реакций
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
32.
Fig. 10-12Фотосистема
Фотон
тилакоидная мембрана
СветоРеакционный
собирательный комплекс
комплекс
Строма
Primary
electron
acceptor
e–
перенос
энергии
Пара молекул
хлорофилла а
Пигментные
молекулы
Тилакоидное пространство
33.
Есть два типа фотосистем в тилакоидноймембраны
Фотосистема II функционирует (PS II) первой
(цифры отражают порядок открытия
фотосистем) и лучше поглощает длину волны
680 нм
Реакционный центр хлорофилла ФС II
называется Р680
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
34.
• Фотосистемы I (ФС I), лучше поглощаетдлину волны 700 нм
• Реакционный центр хлорофилла ФС I
называется P700
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
35.
Linear Electron Flow• Существует 2 пути движение электронов в
световой фазе: циклическая и
нециклическая
• Нециклический путь, основной путь,
который проходит через обе фотосистемы и
в резултате образует АТФ и НАДФН
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
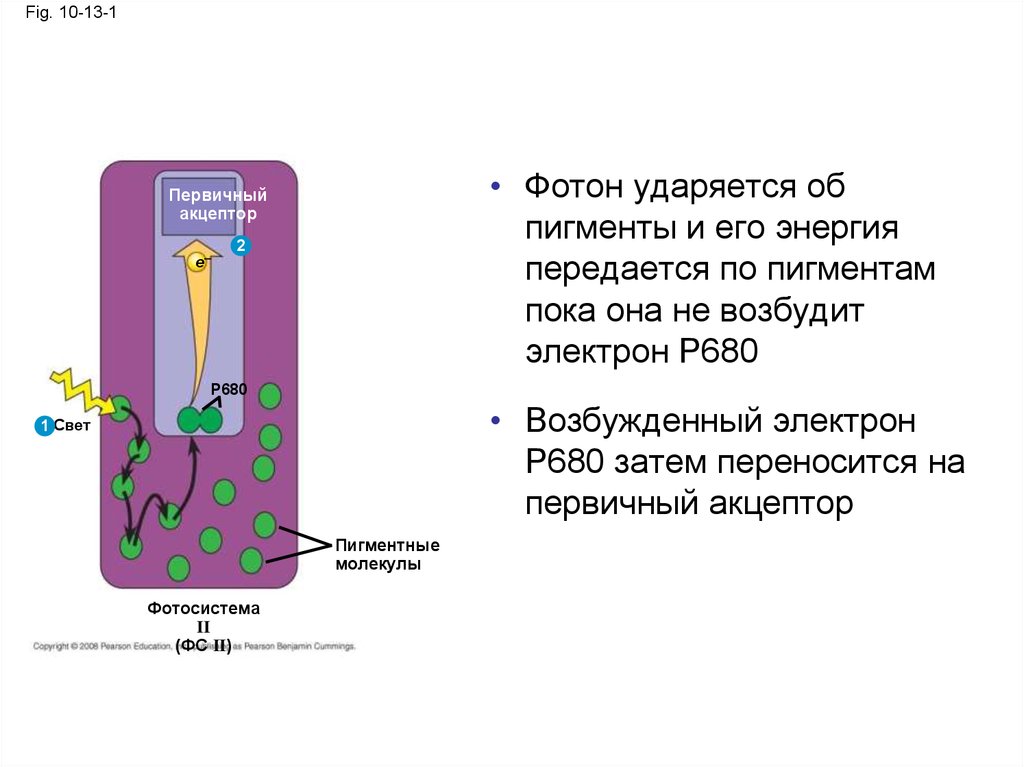
36.
Fig. 10-13-1• Фотон ударяется об
пигменты и его энергия
передается по пигментам
пока она не возбудит
электрон P680
Первичный
акцептор
e–
2
P680
• Возбужденный электрон
Р680 затем переносится на
первичный акцептор
1 Свет
Пигментные
молекулы
Фотосистема
II
(ФС II)
37.
Fig. 10-13-2• P680+ (P680 которая
потеряла электрон)
преврящается в очень
сильный окислитель
Первичный
акцептор
2 H+
+
1/ O
2
2
e–
H2O
2
3
e–
e–
P680
1 Свет
Пигментные
молекулы
Фотосистема
II
(ФС II)
• H2O расщипляется
ферменом, и электрон
волорода переносится на
P680+, который затем
восстанавлявается до P680
обратно.
• O2 высвобождается как
конечный продукт данной
реакции
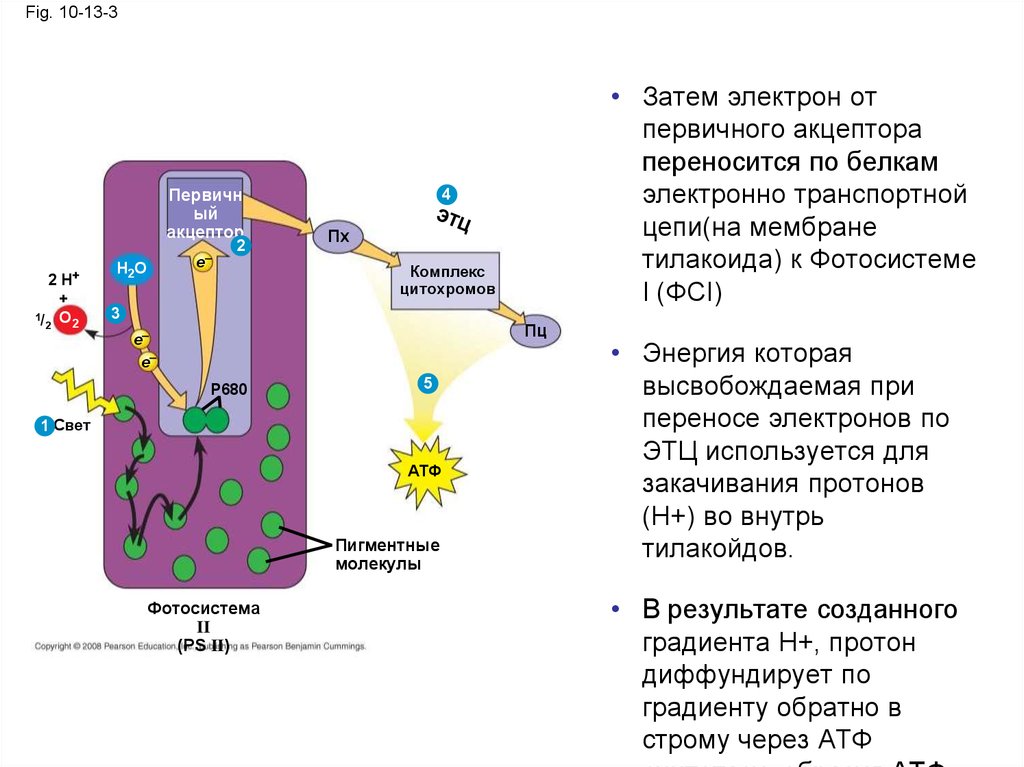
38.
• Затем электрон от первичного акцепторапереносится по белкам электронно
транспортной цепи(на мембране тилакоида)
к Фотосистеме I (ФСI)
• Энергия которая высвобождаемая при
переносе электронов по ЭТЦ используется
для закачивания протонов (Н+) во внутрь
тилакойдов.
• В результате созданного градиента Н+,
протон диффундирует по градиенту обратно
в строму через АТФ синтетазу, образуя АТФ
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
39.
Fig. 10-13-3Первичн
ый
акцептор
1/
2
H+
2
+
O2
e–
H2O
2
• Затем электрон от
первичного акцептора
переносится по белкам
электронно транспортной
цепи(на мембране
тилакоида) к Фотосистеме
I (ФСI)
4
Пх
Комплекс
цитохромов
3
Пц
e–
e–
P680
5
1 Свет
АТФ
Пигментные
молекулы
Фотосистема
II
(PS II)
• Энергия которая
высвобождаемая при
переносе электронов по
ЭТЦ используется для
закачивания протонов
(Н+) во внутрь
тилакойдов.
• В результате созданного
градиента Н+, протон
диффундирует по
градиенту обратно в
строму через АТФ
40.
• Свет попавшии на ФС I так же возбуждаетэлектрон P700 и переносит его на
первичный акцептор
• Образовавшаяся дыра от электрона в P700+
заполняется электроном идущей от ФС II.
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
41.
Fig. 10-13-4первичн
ый
акцептор
1/
2
H+
2
+
O2
e–
H2O
2
Первичный
акцептор
4
e–
Пх
Комплекс
цитохрома
3
Пц
e–
e–
P700
P680
5
Свет
1 свет
6
ATФ
Пигментные
молекулы
Фотосистема
II
(ФС II)
ФотосистемаI
(ФС I)
42.
• Электро от первичного акцептора Р700переносится по ЭТЦ к белковому комплексу,
ферродоксину (Фд)
• Этот жлектрон затем используется для
восстановления НАДФ+ до НАДФН
• Электрон в составе НАДФН затем
используется для реакции в цикле Кальвина
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
43.
Fig. 10-13-54
Primary
acceptor
2
H+
+
1/ O
2
2
e–
H2O
2
Первичн
ый
акцептор
e–
Pq
Cytochrome
complex
7
Фд
e–
e–
8
НАДФ+
редуктаза
3
Pc
e–
e–
P700
5
P680
Свет
1 Light
6
ATP
Pigment
molecules
Photosystem
II
(PS II)
Photosystem I
(PS I)
НАДФ++
+ H+
НАДФН
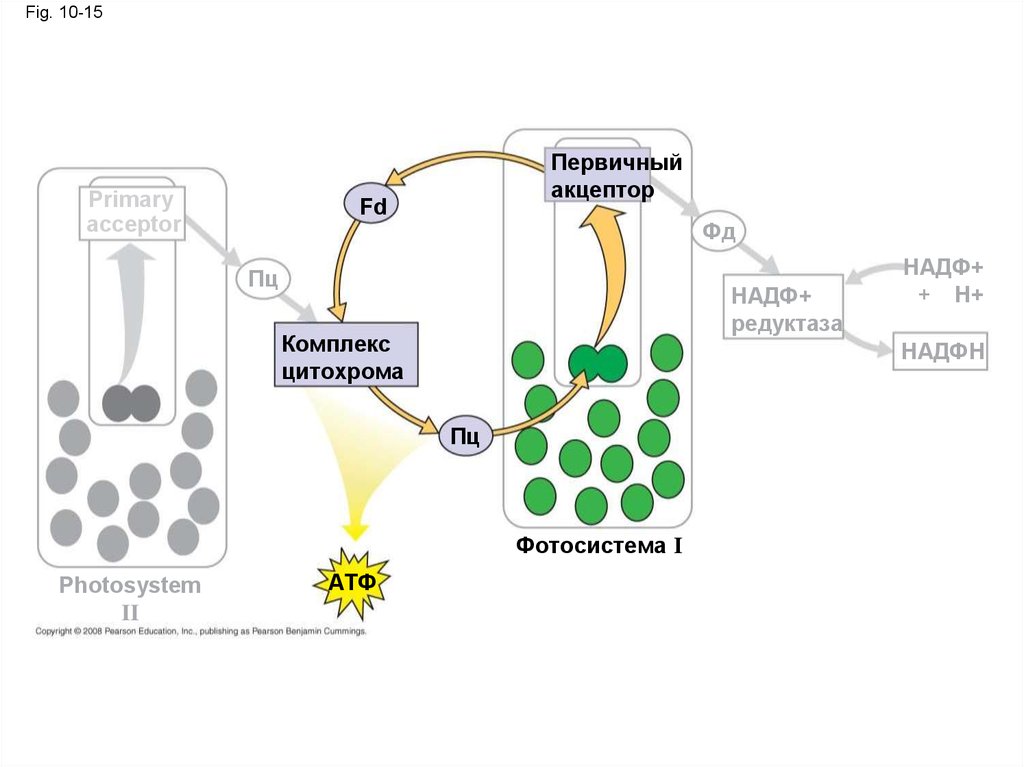
44.
Cyclic Electron Flow• Циклическая фотосистема только
использует ФС I и образует только АТФ
• Этим самым нужда в АТФ может
заполниться для дальнейшего
использования в цикле Кальвина
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
45.
Fig. 10-15Primary
acceptor
Первичный
акцептор
Fd
Фд
Пц
НАДФ+
редуктаза
Комплекс
цитохрома
НАДФН
Пц
Фотосистема I
Photosystem
II
АТФ
НАДФ+
+ Н+
46.
• Некоторые организмы как серные бактерийимеют только ФС I
• Есть гипотеза что циклический путь был
развит первее у организмов в процессе
эволюции
• Циклический путь мог уберечь организмы от
вреда наносимого от света
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
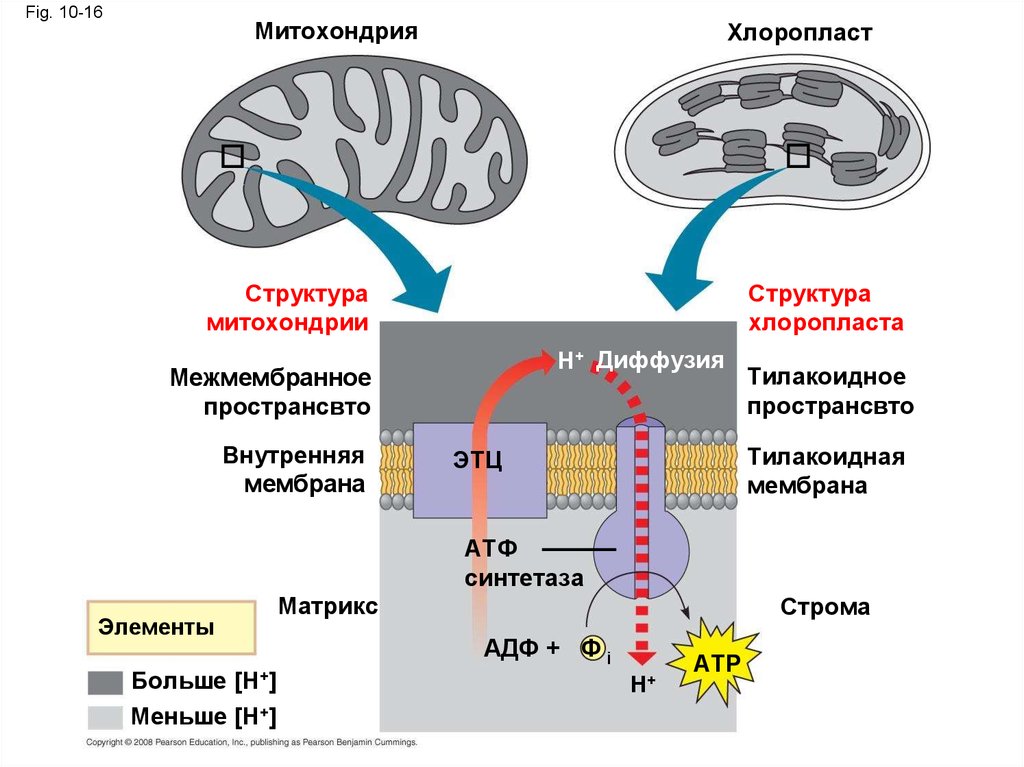
47.
Fig. 10-16Митохондрия
Хлоропласт
Структура
митохондрии
Структура
хлоропласта
H+ Диффузия
Межмембранное
пространсвто
Внутренняя
мембрана
Тилакоидное
пространсвто
Тилакоидная
мембрана
ЭТЦ
АТФ
синтетаза
Элементы
Больше [H+]
Меньше [H+]
Матрикс
Строма
AДФ + Ф i
H+
ATP
48.
• АТФ и НАДФН образуются на сторонестромы которые затем используются в
цикле Кальвина
• В кратце световая фаза синтезирует АТФ
увеличивая потенциальную энергию
электрона в процессе переноса от Н2О к
НАДФН
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
49.
Fig. 10-17Строма
(низкая
концентрация Н+
Комплекс
цитохрома
Фотосистема II
4 H+
Свет
Фотосистема I
Свет
Фд
НАДФ
редуктаза
H2O
Тилакоидное
пространсвто
(высокая концентрация
Н+)
1
e–
1/
O2
+2 H+
4 H+
к циклу
Кальвина
Тилакоидная
мембрана
строма
(lнизкая концентрация Н+)
Пц
2
2
НАДФ+ +
H+
НАДФН
Пх
e–
3
АТФ
синтетаза
AДФ
+
Фi
АТФ
H
+
50.
Концепция 10.3: Цикл Кальвина используетАТФ и НАДФН чтобы синтезировать сахар
• Цикл Кальвина, как в цикле лимонной
кислоты, восстанавливает его исходный
материал после как молекулы входят и
выходят из цикла
• Цикл строит сахар из небольших молекул с
помощью АТФ и уменьшения мощности
электронов осуществляется с помощью
НАДФН
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
51.
• Углерод входит в цикл в виде CO2 и выходит в видесахара глицеральдегид-3-фосфата (Г3Ф)
• Для синтеза 1 молекулы Г3Ф, цикл должен пройти 3
раза, и фиксировать 3 молекулы CO2
• Цикл Кальвина состоит из трех фаз:
– Фиксация углерода (катализируется РУБИСКО(
рибулоза-бисфосфат карбоксилаза)
– Восстановления
– Регенерация акцептора CO2 (РиБФ)
Copyright © 2008 Pearson Education, Inc., ishing as Pearson Benjamin Cummings
52.
Fig. 10-18-3Вход 3
CO2
(Заходит по
одной)
Фаза 1: Фиксация СО2
Рубиско
3 P
Промежуточный
продукт
P
6
P
3-Фосфоглицерат
3P
P
Рибулоза-бисфосфат
(РиБФ)
6
АТФ
6
AДФ
Цикл
Кальвина
3
AДФ
3
AТФ
6 P
P
1,3-Бисфосфоглицерат
6 НАДФН
Фаза 3:
Регенерация
акцептора СО2
6 НАДФ
6 Pi
P
5
Г3Ф
6
P
Глицеральдегид-3-фосфат
(Г3Ф)
1
Выход
P
Г3Ф
(сахар)
Глюкоза и другие
органические
продукты
Фаза 2:
Восстановления
53.
Concept 10.4: Альтернативные способыфиксации СО2 в сухих и жарких климатах
• Обезвоживание растений иногда ставит растения
перед выбором осуществления фотосинтеза. ( так как в
фотосинтезе используется вода)
• В жарких климатических условиях устьица растений
закрываются для сохранения больше воды и
фотосинтез также уменьшается
• Закрывание устьиц прекращает вход СО2 в растении и
прриводит к накоплению О2 внутри.
• Этот процесс благоприятно действует на процесс
фотодыхания
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
54.
Фотодыхание• Во многих растениях(C3 plants), фиксация
СО2 через Рубиско образует трех
углеродные соединения
• В процессе фотодыхания,Рубиско включает
O2 вместо CO2 в цикл Кальвина
• Фотодыхание употребляет O2 органические
соединения и образует CO2 без
образования АТФ и сахара
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
55.
• Фотодыхание может быть эволюционнымреликтом, потому что Rubisco впервые
появились в то время, когда атмосфера
была гораздо меньше O2 и больше СО2,
• Фотодыхание ограничивает пагубные
продукты световых реакций, которые строят
в отсутствие цикла Кальвина
• У многих растений, фотодыхание является
проблемой, потому что в жаркий, сухой день
может истощить целых 50% от углерода,
фиксируемого циклом Кальвина
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
56.
C4 Растения• C4-растени минимизировать эффект фотодыхания
путем включения CO2 в четырех углеродных
соединений в клетках мезофилла
• Этот шаг требует фермент ФЕП-карбоксилазы
• ФЕП карбоксилазы имеет высокое сродство к СО2, чем
RUBISCO ; он может фиксировать СО2 даже при
низких концентрации CO2
• Эти четырех углеродное соединение экспортируется в
клетки обкладки, где они высвобождают CO2, который
затем используется в цикле Кальвина
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
57.
Fig. 10-19C4 путь
C4 анатомия листа
Клетки
мезофилла
клетки мезофилла
Фотосинтетич
еские клетки
клетки
С4 растений
обкладки
Оксалоацетат (4C)
сосуды
ФЕП
(3C)
Малат (4C)
Устьице
Клетки
обкладки
CO2
ФЕП карбоксилаза
AДФ
ATФ
Пируват (3C)
CO2
Цикл
кальвина
Сахар
Сосудис
тые
ткани
58.
CAM Растения• Некоторые растения, в том числе суккуленты,
используют CAM путь метаболизма, чтобы
фиксировать углерода (СО2)
• CAM растения открывают свои устьица ночью и
включают СО2 в органические кислоты
• Устьица закрываются в течение дня, и CO2
высвобождается из органических кислот и
используются в цикле Кальвина
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
59.
Fig. 10-20Сахарный
тросник
Ананас
C4
CAM
CO2
клетки
мезофилла органические
кислоты
Клетки
обкладки
CO2
1 СО2 включается
в органические
кислоты после
фиксации
CO2
цикл
кальвина
сахар
(a) Пространственное
разделение событии
Ночь
органические
кислоты
CO2
2 Органические
кислоты
высвобожда
ют СО2 для
цикла
кальвина
День
Цикл
кальвина
Сахар
(b) Временное разделение
событии