





























 biology
biologySimilar presentations:
")









Эволюция филогенетических групп
1. ЭВОЛЮЦИЯ ФИЛОГЕНЕТИЧЕСКИХ ГРУПП
2. ТЕНДЕНЦИИ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
• "Закон постепенного образования всего сущего" (Рулье,1852).
• "Закон увеличения разнообразия организмов" (Рулье,
1852). В кембрии было 50-100 семейств, сейчас — 900.
2250 семейств вымерло.
• Тенденция увеличения размеров организмов в процессе
эволюции. Клетки прокариот в 10—106 раз мельче клеток
эукариот. Амеба — 1 клетка, мелкая нематода — 560
клеток (по Шенбергу), коловратка — 103, человек — 1014
клеток.
• Тенденция увеличения разнообразия органических
веществ, входящих в состав организмов.
3. Правила эволюции
• 1. Необратимость эволюции (Л.Долло, 1893).• 2. Правило прогрессирующей специализации (Ш.Депре,
1876)
теломорфоз,
гиперморфоз,
катаморфоз,
гипоморфоз
• 3. Правило происхождения от неспециализированных
предков (Э.Коп, 1896).
• 4. Принцип гетеробатмии (А.Л.Тахтаджян, 1959)
• 5. Правило адаптивной радиации (Г.Ф.Осборн, 1902).
• 6. Правило чередования главного направления
эволюции (Шмальгаузен, 1939). Арогенная эволюция
чередуется с аллогенной.
• 7.
Правило
усиления
интеграции
биосистем
(Шмальгаузен, 1961).
• 8. Закон неравномерности эволюции (Ч. Дарвин ).
4. СКОРОСТЬ ЭВОЛЮЦИИ
Скорость эволюции —количество эволюционных изменений заединицу времени. Чаще всего скорость эволюции измеряется
числом таксонов (видов, родов, семейств), которые появлялись за
единицу времени, например, за 1 миллион лет.
• Горотелия — средний, обычный темп
эволюционного процесса (хищные млекопитающие
и брюхоногие моллюски, амфибии в карбоне,
рептилии — в перми).
• Тахителия — быстрый темп эволюции (слоны: в
течение 40 млн. лет кайнозойской эры образовали
6 семейств, 35 родов, млекопитающие – за 30 млн.
лет кайнозоя возникло 100 семейств).
• Брадителия — медленная эволюция (характерна
для «живых ископаемых»—(мамонтовое дерево,
щитень, гаттерия, неопилина, гинкго, наутилус).
5. НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
• Аллогенез – в пределах однойадаптивной зоны (идиоадаптация),
а- алломорфозы – изменение без
сужения адаптивной зоны,
б- специализации – сужение
адаптивной зоны.
• Арогенез – с расширением
адаптивной зоны – выход в новую
адаптивную зону.
6.
7. ФОРМЫ ФИЛОГЕНЕЗА
Эволюция филогенетических группФОРМЫ ФИЛОГЕНЕЗА
1. Филетическая эволюция или ортогенез.
2. Дивергенция.
3. Конвергенция.
4. Параллелизм,
а- синхронный,
б- асинхронный
8.
Дивергенция9.
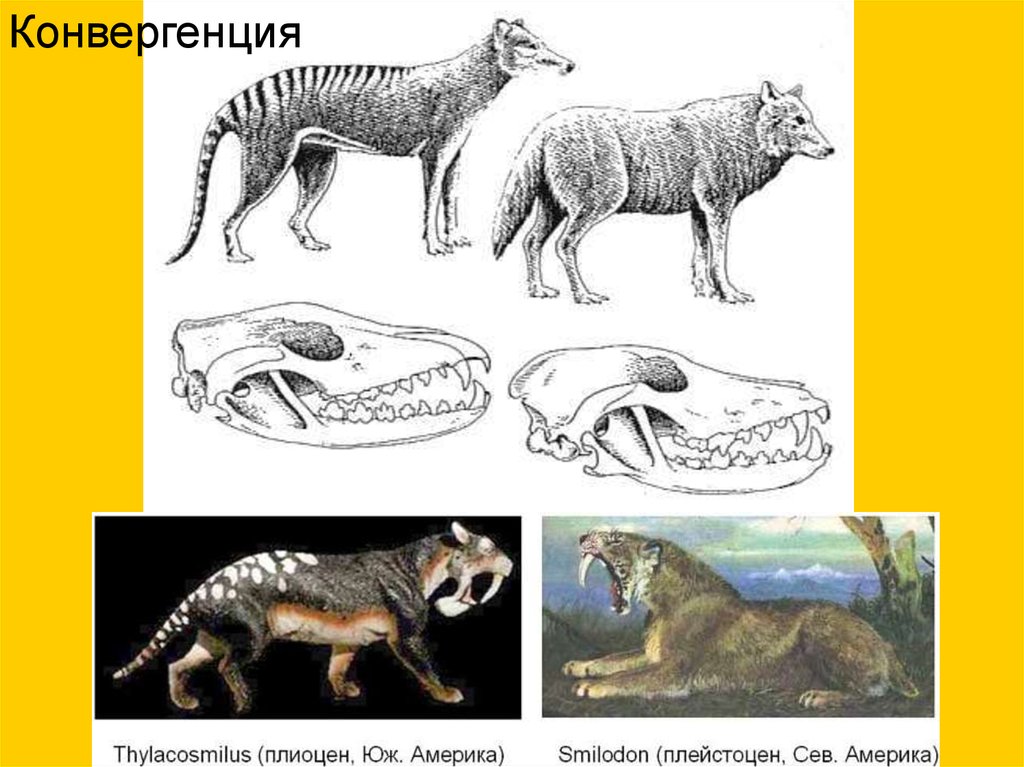
Конвергенция10.
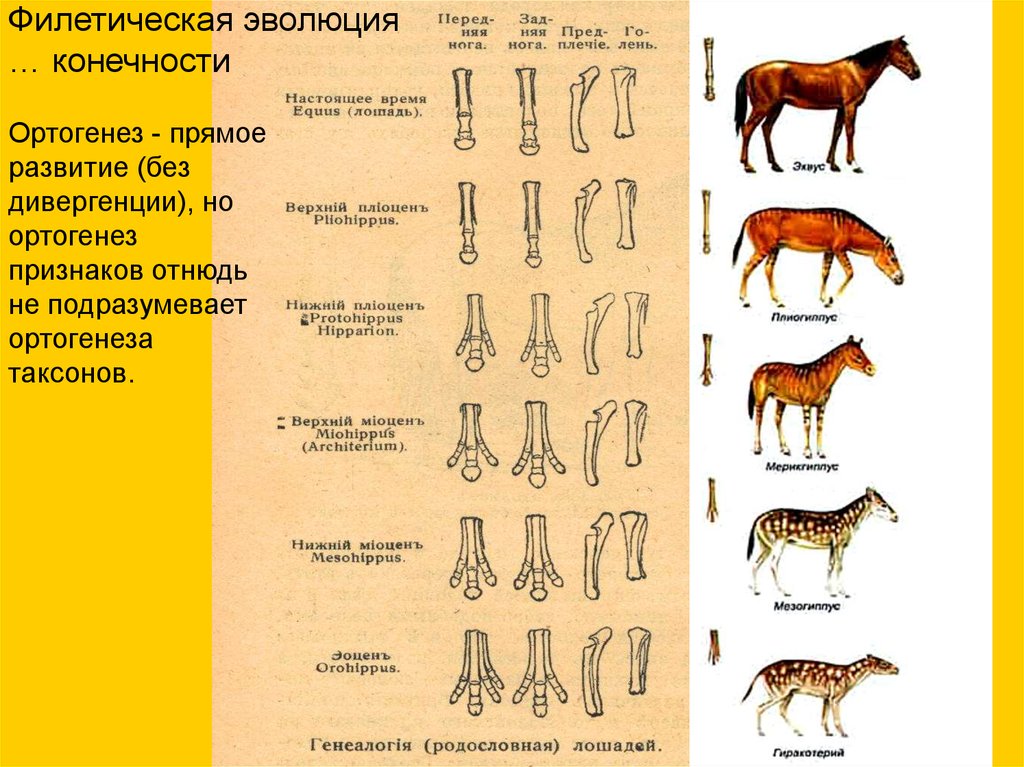
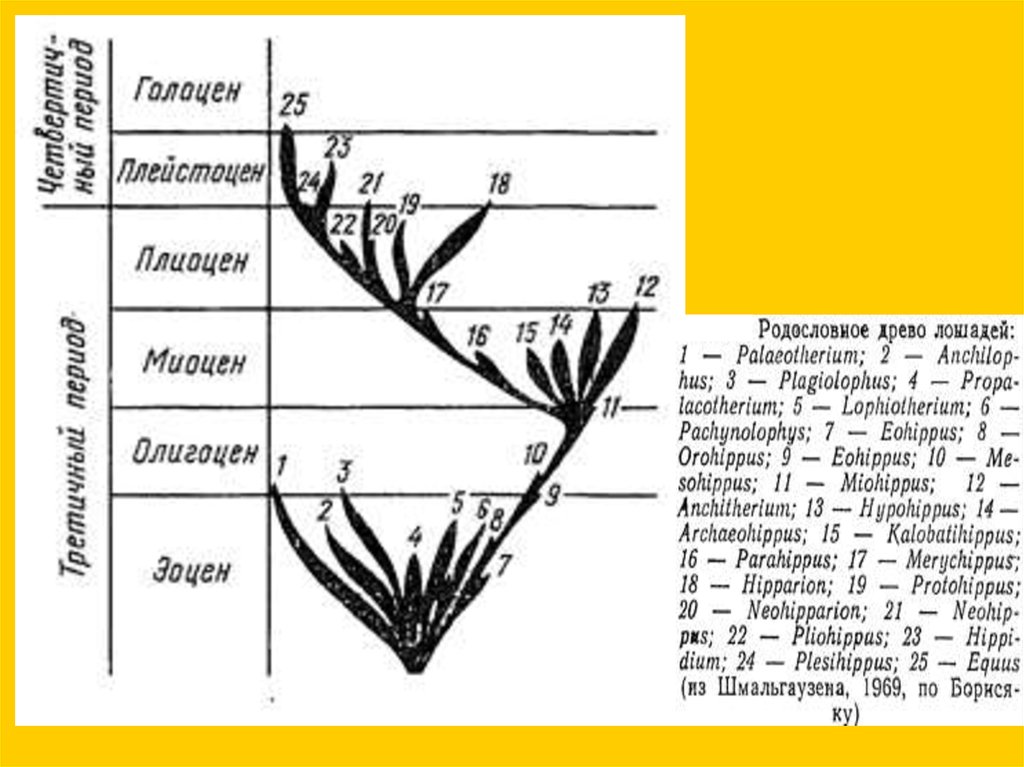
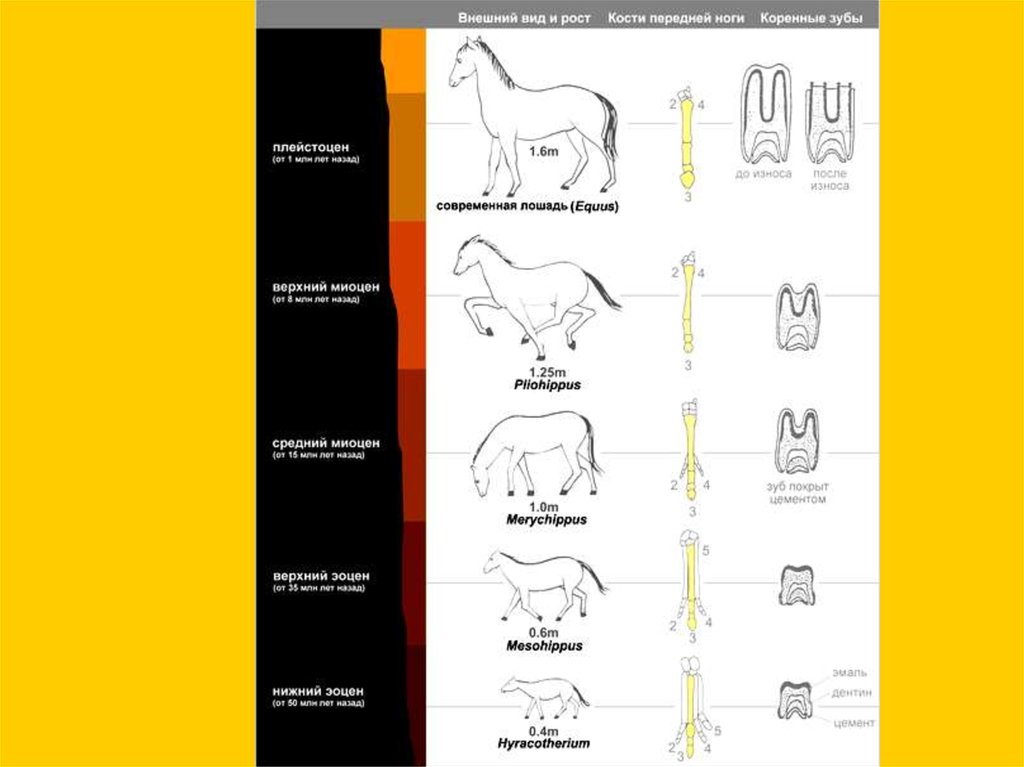
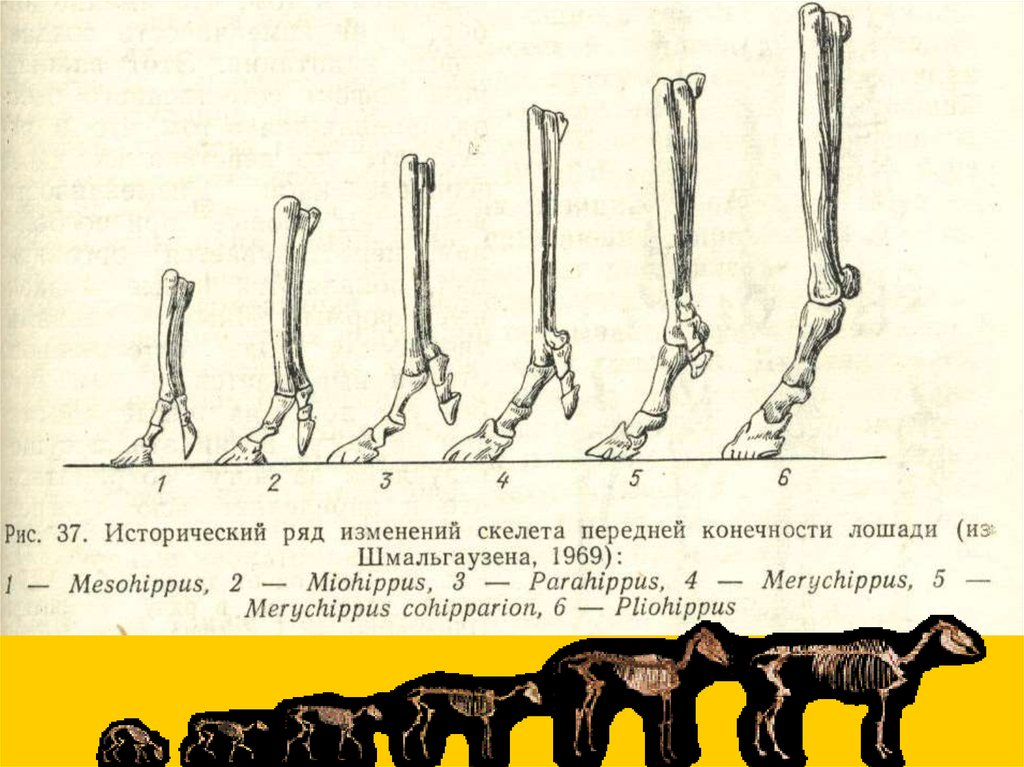
Филетическая эволюция… конечности
Ортогенез - прямое
развитие (без
дивергенции), но
ортогенез
признаков отнюдь
не подразумевает
ортогенеза
таксонов.
11.
12.
13.
14.
Синхронный параллелизмредукция боковых
пальцев почти без
компенсации инадаптивная
эволюция
прогрессивное
(компенсаторное)
развитие 3-го пальца
и редукция
остальных
15.
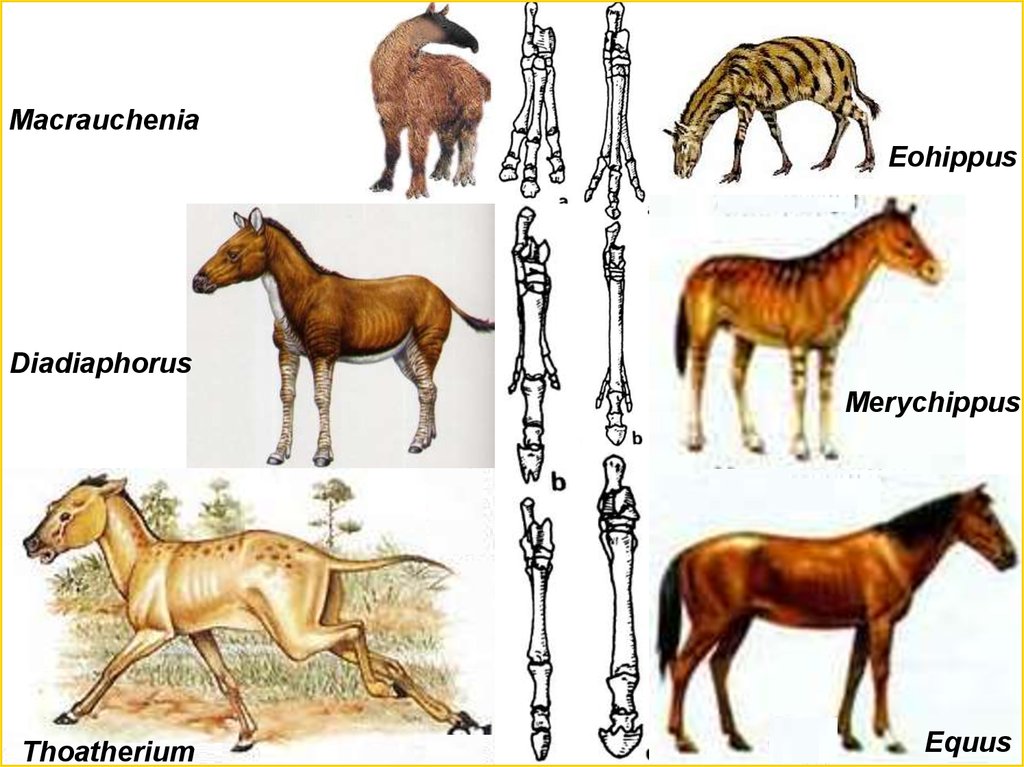
MacraucheniaEohippus
Diadiaphorus
Merychippus
Thoatherium
Equus
16.
MacraucheniaEohippus
17.
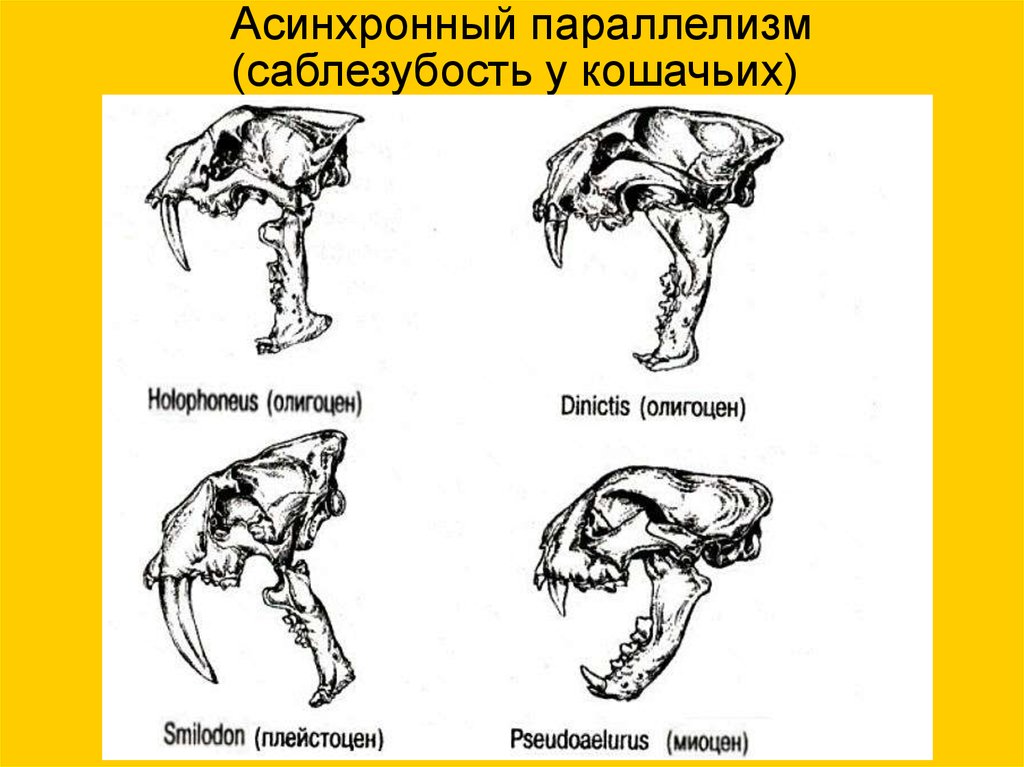
Асинхронный параллелизм(саблезубость у кошачьих)
18.
Асинхронный параллелизм(саблезубость у кошачьих)
19.
Асинхронный параллелизм(саблезубость у кошачьих)
Holophoneus, олигоцен
Smilodon, плейстоцен
Dinictis, олигоцен
Pseudoaelurus миоцен
20.
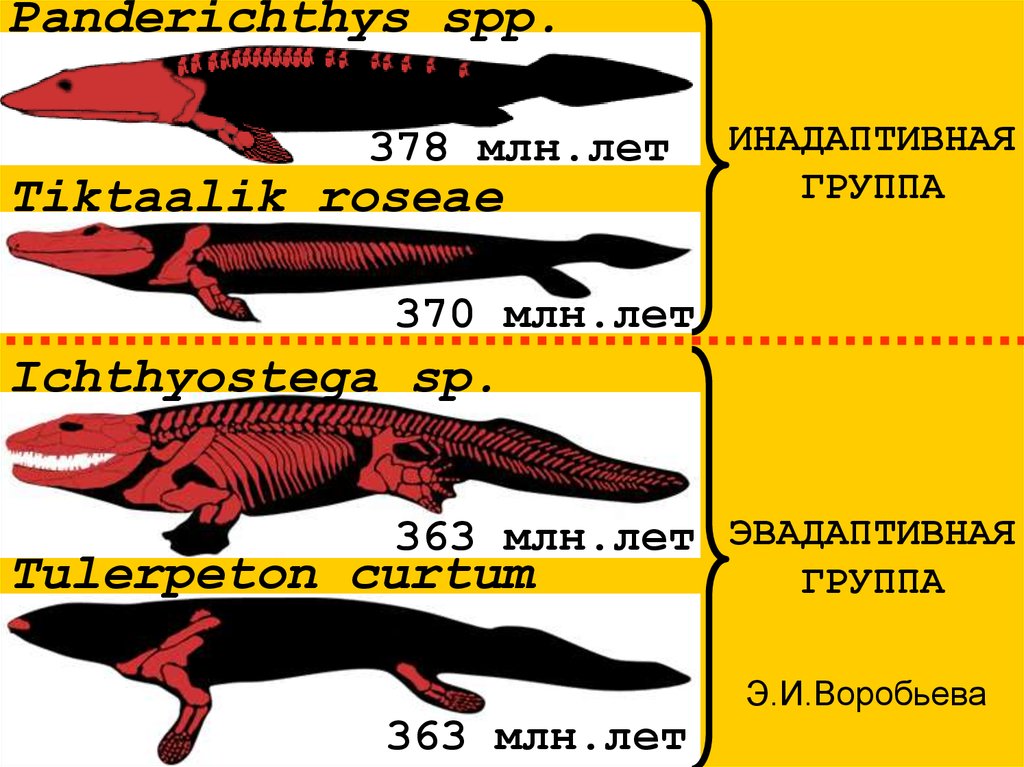
Panderichthys spp.378 млн.лет
Tiktaalik roseae
ИНАДАПТИВНАЯ
ГРУППА
370 млн.лет
Ichthyostega sp.
363 млн.лет ЭВАДАПТИВНАЯ
Tulerpeton curtum
363 млн.лет
ГРУППА
Э.И.Воробьева
21.
22.
Поздняя юрамел
23.
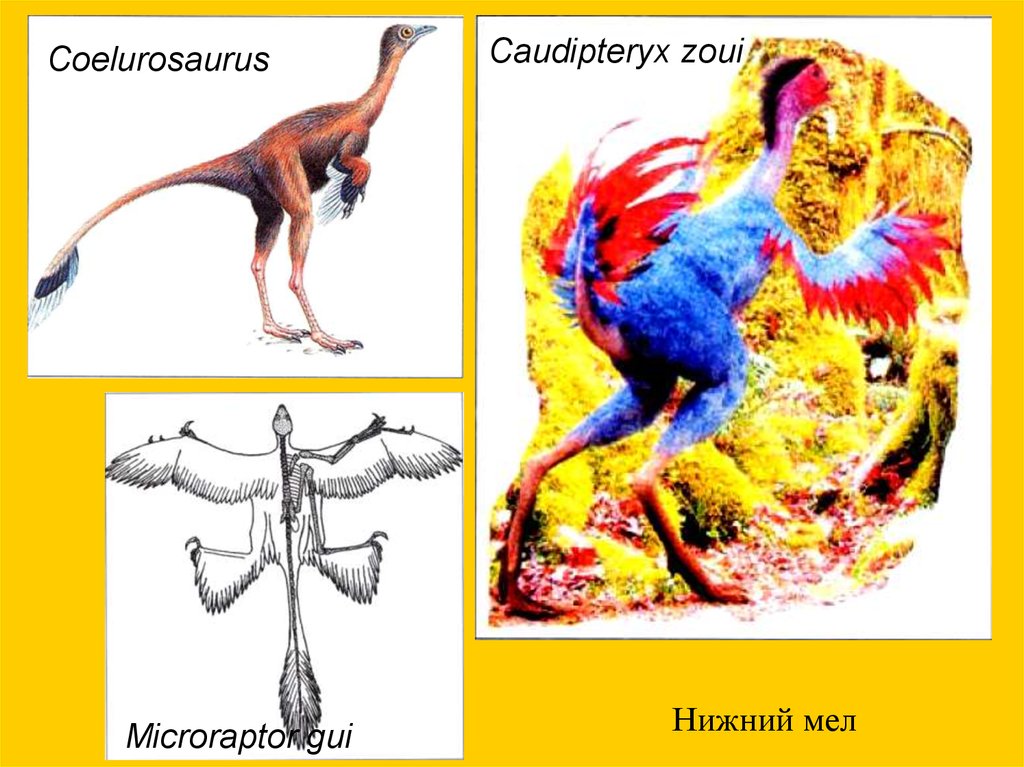
CoelurosaurusMicroraptor gui
Caudipteryx zoui
Нижний мел
24.
поздний триасранний мел
25.
Параллельное появление перьев и полета26.
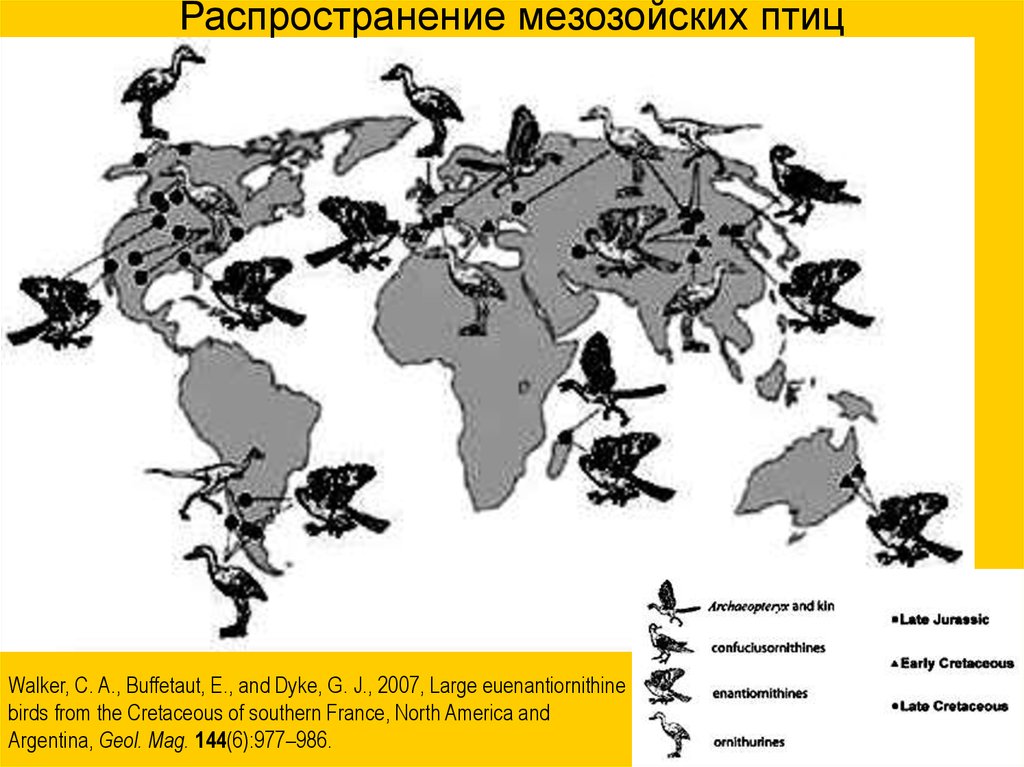
Распространение мезозойских птицWalker, C. A., Buffetaut, E., and Dyke, G. J., 2007, Large euenantiornithine
birds from the Cretaceous of southern France, North America and
Argentina, Geol. Mag. 144(6):977–986.
27.
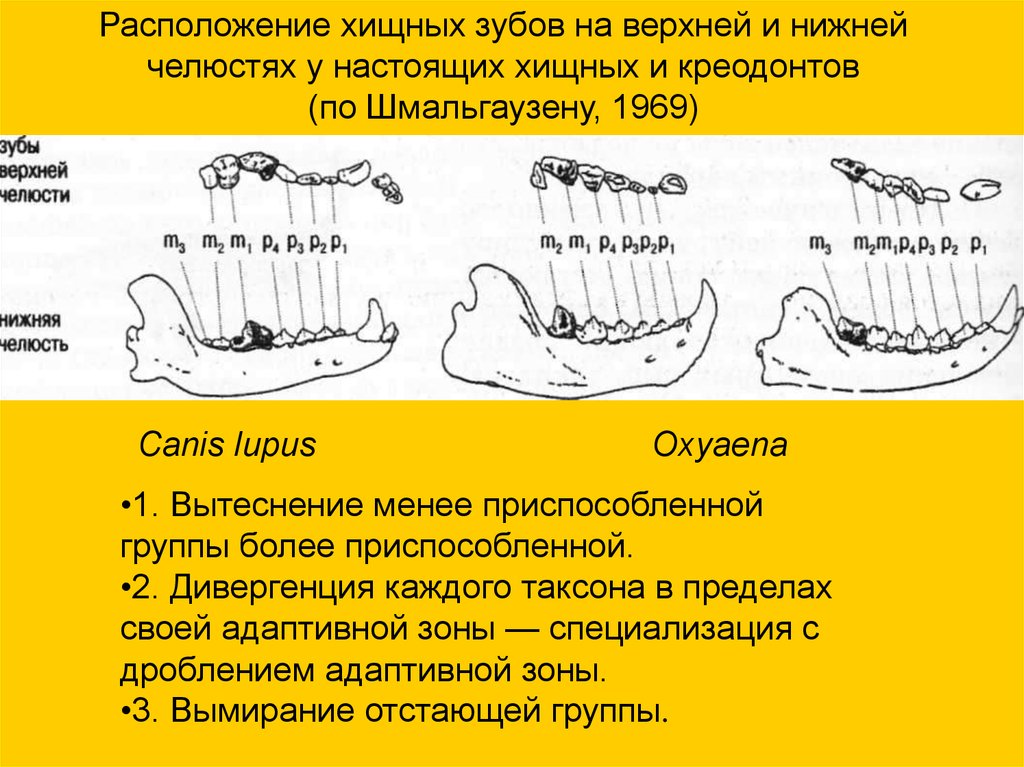
Расположение хищных зубов на верхней и нижнейчелюстях у настоящих хищных и креодонтов
(по Шмальгаузену, 1969)
Canis lupus
Oxyaena
•1. Вытеснение менее приспособленной
группы более приспособленной.
•2. Дивергенция каждого таксона в пределах
своей адаптивной зоны — специализация с
дроблением адаптивной зоны.
•3. Вымирание отстающей группы.
28.
Пример возникновения похожих наборов "жизненных форм" наразобщенных континентах
Сумчатый саблезубый тигр Thylacosmilus
из плиоцена Южной Америки
При параллельном развитии сходство объясняется частью
общностью происхождения, а частью — приспособлением к
сходной среде (Шмальгаузен,1969)
29.
Представители планцентарных (слева) и сумчатых (справа) млекопитающих30.
Ambystoma macrodactylum Pseudacris maculataSpea bombifrons
Bufo boreas
Bufo hemiophrys
Rana sylvatica
Rana luteiventris
Rana pipiens
Salamandrella keyserlingii
Pelobates fuscus
Bufo bufo
Bufo viridis
Rana temporaria
Rana arvalis
Rana amurensis
Rana ridibunda
31. Параллелизмы и их эволюционное значение Л.П.Татаринов,1987
• 1. Генотипические (в основе лежат гомологическиемутации).
• 2. Эпигенетические
а) биохимические (независимое появление сходных метаболитов),
б) гистологические (независимое появление сходных тканей),
в) морфологические (основаны на полях, градиентах, индукции),
г) физиологические (сходные физиологические функции),
д) этологические (независимое сходство в поведении).
• Сходство при параллелизме объясняется общностью
происхождения и проявлением в эволюции латентных
потенций, заложенных в механизмах морфогенеза, генотипе,
метаболизме.
• Различия - начальным расхождением признаков. Латентные
потенции, лежащие в основе параллелизмов, приводятся в
действие давлением естественного отбора.
32. Причины параллелизмов
• Принципиальное сходство в строенииосновных типов сообществ, или биоценозов;
сходство наборов экологических ниш в разных
сообществах.
• «Случайные и ненаправленные» мутации
вовсе не такие уж «случайные». («Метод
случайного поиска» оптимизируется
природой).
• Строение организма допускает ограниченный
набор возможных преобразований, что во
многом предопределяет пути его возможных
эволюционных изменений.
33. Daniel M. Weinreich, Nigel F. Delaney, Mark A. DePristo, Daniel L. Hartl. Darwinian Evolution Can Follow Only Very Few
Адаптация бактерий к цефотаксиму«дикий тип» гена бета-лактамазы пять мутаций (вместе)
увеличивают устойчивость в 100 000 раз;
• существует 5! = (1х2х3х4х5)=120 различных вариантов пути
от «дикого типа» (– – – – –) к устойчивому (+ + + + +);
• из 120 путей - 102 не могут реализоваться (снижение
приспособленности);
18 путей сильно различаются по вероятности реализации;
• в 99% случаев эволюция «выберет» один из 10, а в 50%
случаев — один из двух наиболее вероятных путей;
• молекулярной эволюции свойственна высокая
повторяемость: разные организмы должны независимо
друг от друга двигаться по одним и тем же
«разрешенным» эволюционным траекториям.
Daniel M. Weinreich, Nigel F. Delaney, Mark A. DePristo, Daniel L.
Hartl. Darwinian Evolution Can Follow Only Very Few Mutational
Paths to Fitter Proteins // Science. 2006. V. 312. P.111-114.
34.
35.
• Система, состоящая из взаимосвязанных элементов (блоков),может существовать в различных устойчивых состояниях,
переходя из одного в другое, но число этих состояний ограничено,
а их характер определяется свойствами элементов и структурой
их взаимодействий.
• Дискретный и упорядоченный характер изменчивости
накладывает жесткие ограничения на эволюционные возможности
организмов.
• Сходная изменчивость предопределяет сходство эволюционных
изменений у разных групп, а ограниченное число возможных
вариаций ограничивает и число возможных путей эволюционного
развития.
• При анализе причин канализации филогенеза выделяются три
основные группы факторов, определяющих направление
эволюции таксона: 1) тип онтогенеза, 2) морфофункциональная
организация особи, 3) условия внешней среды.
36. Эволюционный прогресс и регресс Признаки биологического прогресса:
1- увеличение численности,
2- увеличение ареала,
3- расширение экологической ниши,
4- увеличение числа дочерних таксонов.
37. Причины эволюционного прогресса
Общая тенденция - усложнение организации.В открытых системах с постоянным поступлением вещества
и энергии извне, закономерно происходит самоорганизация образование «порядка из хаоса», т.е. прогресс (И.Пригожин).
Основа прогресса - блочный принцип организации.
Новые, сложные системы складываются из готовых
функциональных блоков уже имеющихся, простых
систем.
- на молекулярном уровне (новые гены/белки могут
собираться путем дублирования и перекомбинации кусков
старых генов/белков);
- на организменном (через коадаптацию и симбиоз новый, сложный организм может сложиться из сообщества
нескольких простых организмов).
38. … а также «преадаптации». Только часть регуляторных связей важна для адаптации в конкретный момент. Все остальные - слабые,
«непроявленные» составляют «банк регуляторных преадаптаций», длявозникновения новых «проявленных» регуляторных
связей.
Принцип «регуляторного (адаптивного) компромисса»
(А.П.Расницын) - конфликт между необходимостью
поддерживать целостность и нормальное
функционирование сложного организма (старой системы
и необходимостью адекватно реагировать на изменения
внешних условий.
Требуется дальнейшее усложнение организма...
Итог - самоускорение (автокаталитичность)
прогресса.
39. - прогресс тормозится необходимостью сохранения целостности и нормального функционирования старой, сложившейся системы
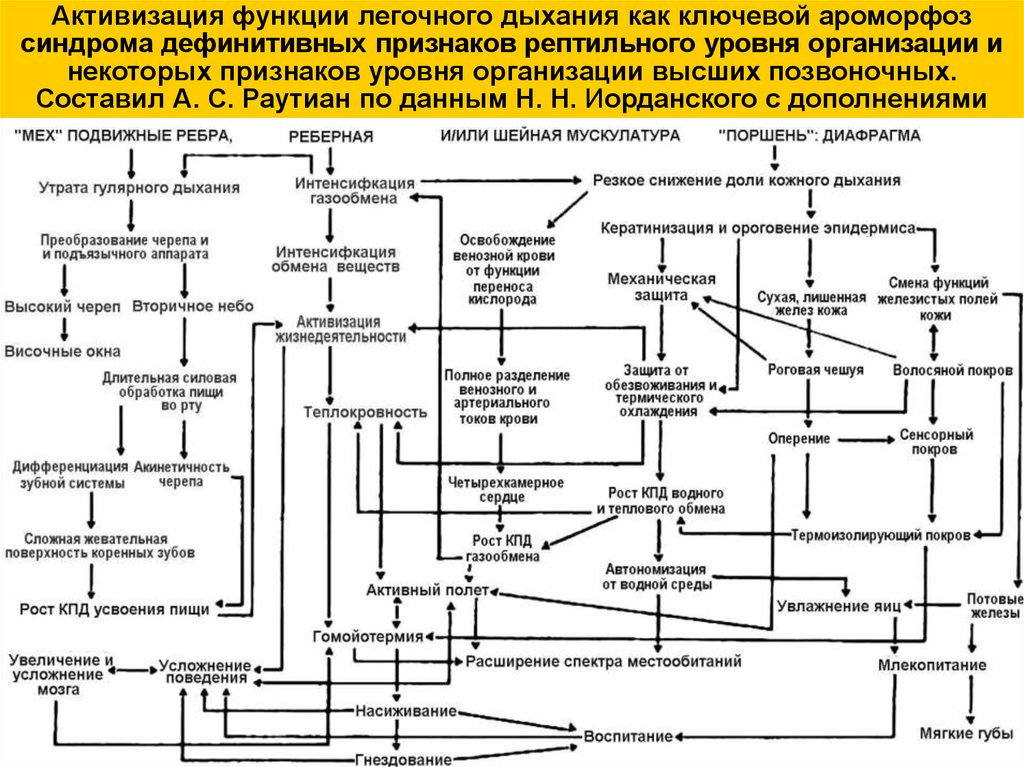
(А.П.Расницын).Принцип «ключевого ароморфоза» (Н.Н.Иорданский)
- часто одно ключевое новообразование открывает
путь для возникновения целого комплекса новшеств.
Ключевые адаптации открывают путь для целого
каскада достаточно строго детерминированных
направленных преобразований (от активизации
легочного дыхания к комплексу признаков,
соответствующих рептильному уровню организации).
40.
Активизация функции легочного дыхания как ключевой ароморфозсиндрома дефинитивных признаков рептильного уровня организации и
некоторых признаков уровня организации высших позвоночных.
Составил А. С. Раутиан по данным Н. Н. Иорданского с дополнениями
41.
Наличие оперения как ключевой ароморфоз синдрома признаковуровня организации птиц. Составил А. С. Раутиан
42.
Устойчивость может быть повышена путем усложнения системы• Биохимия: (ранние этапы) появлялись и совершенствовались
новые ферментые системы, новые метаболические пути. Нарастала
роль регуляторных систем (рецепторов, различных регуляторов
метаболизма и всех прочих клеточных процессов в зависимости от
внешних и внутренних факторов); у эукариот прогресс
внутриклеточной регуляторики, вероятно, вышел на первый план,
а усложнение биохимических систем приостановилось.
• Морфология: с появлением многоклеточности, через дальнейшее
усложнение регуляторных систем, начался ее быстрый прогресс
(усложнялась форма организма, ткани и органы).
• Поведение: усложняется с появлением и развитием нервной
системы (регуляторика высшего уровня).
• Культура: с появлением человека прогресс сконцентрировался в
этой области.
«Элементарное усложнение» - появление новой регуляторной связи
Процесс усложнения автокаталитичен и идет с ускорением.
43.
• Брадителия1) высокий темп эволюции,
2) средний темп эволюции,
3) низкий темп эволюции.
• Эвадаптивная эволюция
1) быстрый слабо скоординированный процесс,
2) медленный скоординированный процесс,
3) стазис.
• Биологический прогресс
1) усложнение организации,
2) рост приспособленности,
3) рост изменчивости.