




")
")



.")
 как пример, иллюстрирующий одновременно два крайних типа формообразования внутри вида:")































 biology
biologySimilar presentations:










Адаптация, как результат эволюции
1. АДАПТАЦИЯ КАК РЕЗУЛЬТАТ ЭВОЛЮЦИИ
2. Появление определенного генотипа на эволюционной арене и размножение его в определенных условиях – элементарное адаптационное
явление3.
Адаптации – возникновение и развитиеконкретных морфофизиологических свойств,
значение которых для организма однозначно
связано с теми или иными конкретными
условиями среды (Тимофеев-Ресовский, 1977).
Адаптация — приспособление организма к
внешним условиям в процессе эволюции,
включая морфофизиологическую и
поведенческую составляющие.
Адаптации – приспособления живых систем к
изменениям среды, переход живой системы из
одного устойчивого состояния в другое (Озернюк,
2003).
4.
Классификация адаптаций по уровнюорганизации биологических систем
(Шкорбатов, 1971):
• онтогенетических систем - организмов,
• филогенетических систем - популяций
и видов,
• ценотических систем - биоценозов.
5. С.С.Шварц - процесс видообразования, как процесс смены уровня адаптаций: развитие популяций в своеобразной среде развитие
С.С.Шварц - процесс видообразования, какпроцесс смены уровня адаптаций:
развитие популяций в своеобразной среде
развитие морфофизиологически
необратимых адаптаций
развитие тканевых адаптаций
нескрещиваемость (возникновение
репродуктивной изоляции)
видообразование.
“Виды не потому виды, что они не скрещиваются, а
они потому не скрещиваюся, что они виды”
(Шварц,1980).
6. Классификация адаптаций (по Н.В.Тимофееву-Ресовскому)
Принцип классификацииПо происхождению
Группа адаптаций
Преадаптивные, комбинативные
(комплементация, эпистаз, полимерия),
постадаптивные
По принадлежности к разным
аспектам среды
Генотипические, популяционные,
По эволюционной значимости
Частные, общие
По морфофизиологическому
содержанию
биоценотические, абиотические
Упрощающие строение,
усложняющие строение
7. Преадаптации (по Cuenot, 1914)
Преадаптации – бесполезные или полуполезныеособенности, способные превращаться в явные
приспособления при изменении образа жизни.
•преадаптации к физическим факторам среды,
•к новой пище,
•к воздействию повреждающих факторов среды.
8.
• Чтобы иметь возможность успешно проникнуть в новуюнишу или адаптивную зону, вид должен быть
преадаптирован к ней; организм называют
преадаптированным, если он способен переходить в
новый биотоп; структуру называют преадаптированной,
если она может взять на себя новую функцию без
ущерба для первоначальной (Майр, 1974).
• ...наличие определенных преадаптаций необходимое
условие для успешной быстрой эволюции популяций в
условиях сильного загрязнения (Porter, 1972).
• Преадаптации, безусловно, являются одним из
важнейших условий, позволяющих сделать животным
первый шаг к освоению новой среды (Шварц, 1980).
9. Комбинативные адаптации
взаимодействие новых мутаций друг с другом и сгенотипом.
• комплементация – усиление выраженности,
• эпистаз – подавление выраженности,
• полимерия – градуированное проявление.
10. they are considered the same species: Theridion grallator
Популяционный полиморфизми его роль в процессах адаптациогенеза
they are considered the same species:
Theridion grallator
11. Термин “полиморфизм” используется только для обозначения прерывистой генетической изменчивости (Майр, 1974).
Полиморфизм - сосуществование в более или менеединамически равновесном состоянии двух или более
морфофизиологически различимых форм в пределах
популяции или группы популяций (ТимофеевРесовский, 1977).
Термин “полиморфизм” используется только для
обозначения прерывистой генетической
изменчивости (Майр, 1974).
Существование двух (или более) генетически
различных форм в популяции в состоянии
длительного равновесия (Яблоков, Юсуфов, 2004).
Генетический полиморфизм природных популяций—
универсальное явление, составляющее основу
генетической пластичности вида (Гершензон, 1983).
12. Феногеография барсука (Meles meles) как пример, иллюстрирующий одновременно два крайних типа формообразования внутри вида:
образование количественных градиентовопределенных признаков внутри всего ареала вида (цифрами обозначены проценты встреч
особей с присутствующим первым предкоренным зубом) и наличие резких различий внутри
ареала вида (по характеру окраски головы)
1 — встречаемость темной полосы на голове, охватывающей все ухо; 2 — встречаемость
узкой темной полосы, проходящей выше уха; з — распространение общей темно-бурой
окраски головы (по Геншеру, 1968, из Тимофеева-Ресовского, Яблокова, 1973)
13. Гетерозиготный полиморфизм в лабораторной популяции Drosophila melanogaster по мутации ebony, устанавливающийся в результате
Гетерозиготный полиморфизм …в сущности, положительноотбирается лишь одна форма — гетерозигота
Гетерозиготный полиморфизм в лабораторной популяции Drosophila
melanogaster по мутации ebony, устанавливающийся в результате
некоторого повышения жизнеспособности гетерозигот по сравнению с
жизнеспособностью обеих гомозигот (из Тимофеева-Ресовского,
Свирежева, 1965)
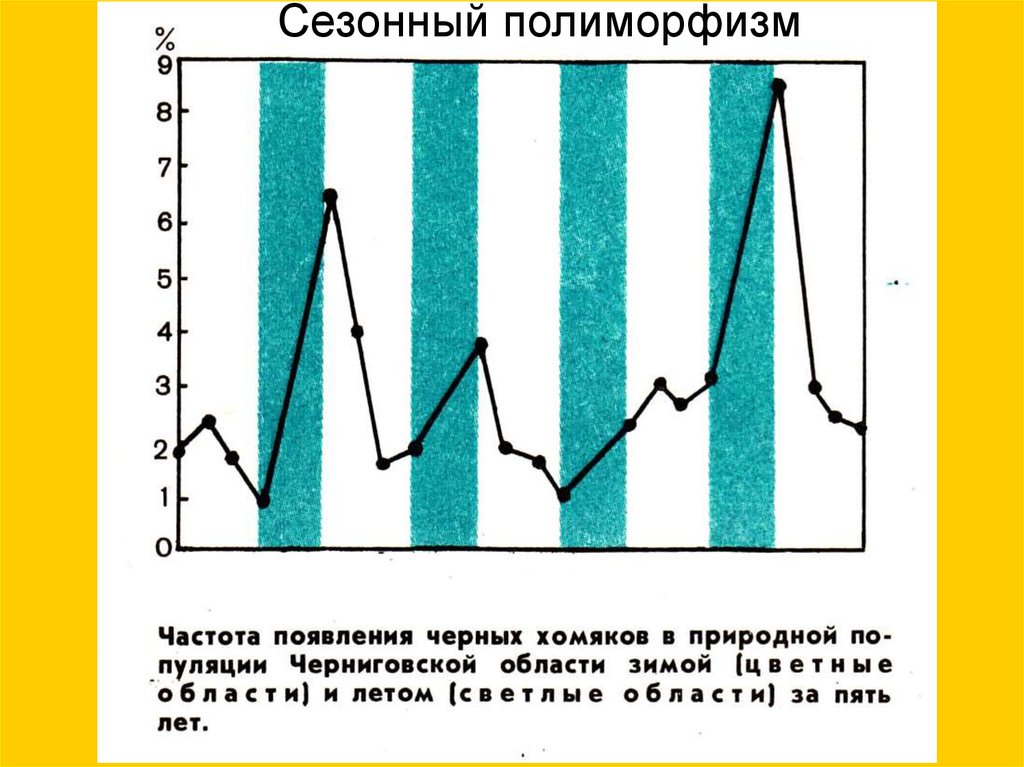
14.
Сезонный полиморфизм15.
Сезонный полиморфизм16.
«Индустриальный меланизм»17.
18.
19.
Кора березы, покрытая лишайникомКора березы, покрытая копотью
20.
Различная смертность особей березовой пяденицы(Bision betularia), выпущенных в разных лесистых
местностях (Кеттлуэлл, 1961)
Ч и с л о в ы п у ще н н ы х
особей
Т и п ме с тн о с ти
меланисты светлые
Равное
С сероватым
фоном
Равное
Загрязненная
копотью
473
496 С сероватым
фоном
447
137 Загрязненная
копотью -
Число особей,
съеденных
п ти ц а ми
В то р и ч н о
выловленные
особи (живые) %
меланисты светлые меланисты светлые
164
26
—
—
15
43
—
—
—
—
6,3
12,5
—
—
27,5
13,0
21. С.М.Гершензон Микроэволюция, полиморфизм и доминантные мутации // Природа. 1985. № 4. Гершензон С. М. «Мобилизационный резерв»
внутривидовой изменчивости // Журн. общ. биол.1941. Т. 2, № 1.
• Для появления гомозигот необходимо, чтобы
скрещивание всегда происходило между особями,
гетерозиготными по данной рецессивной мутации.
• Нужно, чтобы у таких гомозиготных рецессивов не
было никаких дефектов, мешающих нормальной
работе организма.
• Необходимо, чтобы образовавшиеся мутантные
гомозиготы обладали какими-либо преимуществами
перед немутантными.
НИ ОДНО ИЗ ЭТИХ УСЛОВИЙ НЕ СОБЛЮДАЕТСЯ.
22.
Характеристика мутантных генов,присутствующих в природных популяциях
дрозофил (Гершензон, 1985)
23.
• Скорее всего, рецессивные мутации участвуют в эволюциитогда, когда адаптивную ценность приобретает редукция
некоторых структур и физиологических процессов, ставших
лишними, как бывает, скажем, при переходе к паразитизму,
подземному, пещерному, малоподвижному или сидячему
образу жизни и т. п., что большей частью сопровождается
увеличением частоты инбридинга.
• В целом же эволюционное значение рецессивных мутаций,
по-видимому, невелико главным образом из-за
конститутивной слабости гомозигот.
• Большинство доминантных мутаций, встречаемых в
природных популяциях дрозофил, вызывают столь
небольшие фенотипические отклонения от основной массы
особей, что на первый взгляд представляются
“нейтральными”.
24.
• Первый этап: многократное возникновениедоминантных, полудоминантных и кодоминантных
мутаций в разных точках популяций, занимающих
ареал вида и их закрепление.
• Второй этап: формирование адаптивного
сбалансированного генетического полиморфизма.
• Третий этап: интенсивное вытеснение
мутантными - немутантных особей, и появление
нового экотипа.
• При изоляции экотипа от прочих популяций вида,
мы будем иметь дело с постепенным
образованием новой разновидности, которая,
изменяясь далее, может стать новым видом.
25.
Генетический полиморфизм природныхпопуляций — универсальное явление,
составляющее основу генетической
пластичности вида (Гершензон, 1983).
26.
27.
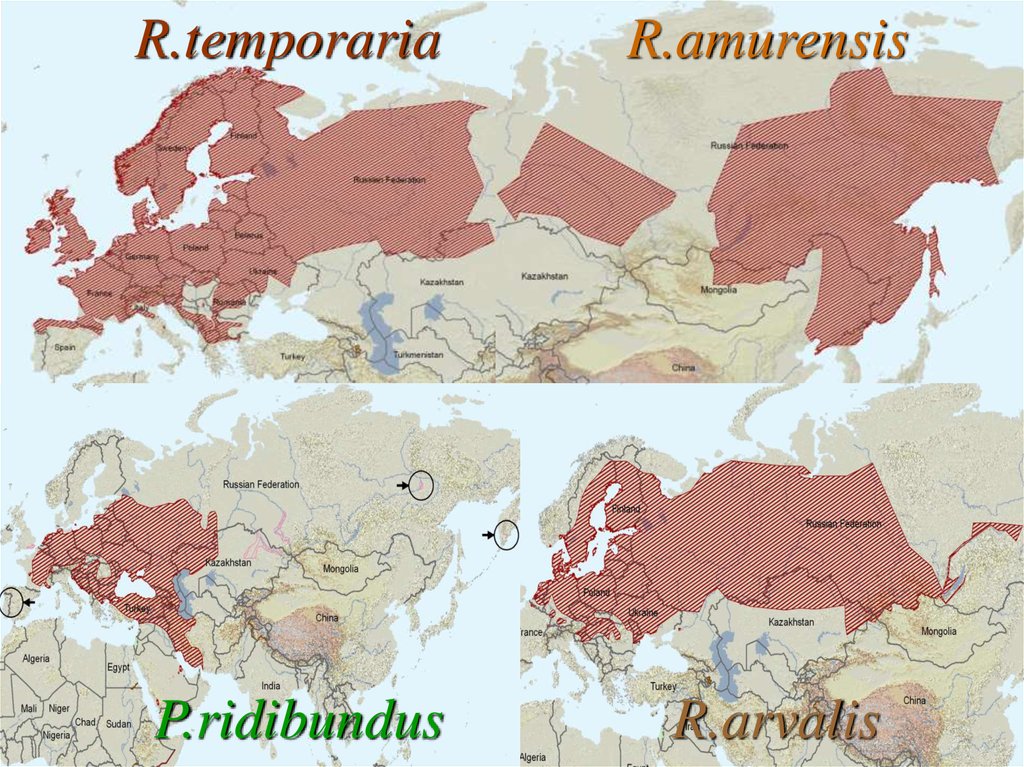
R.temporariaR.amurensis
P.ridibundus
R.arvalis
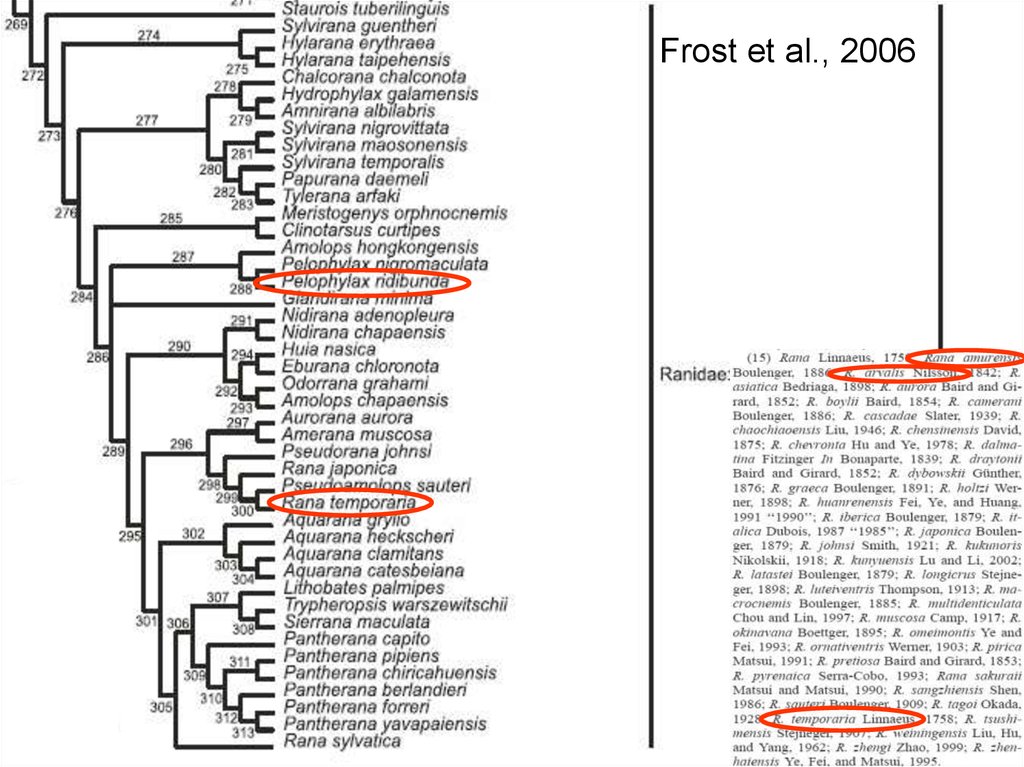
28.
Frost et al., 200629.
Ranaarvalis
Rana
temporaria
Pelophylax
ridibundus
Rana
amurensis
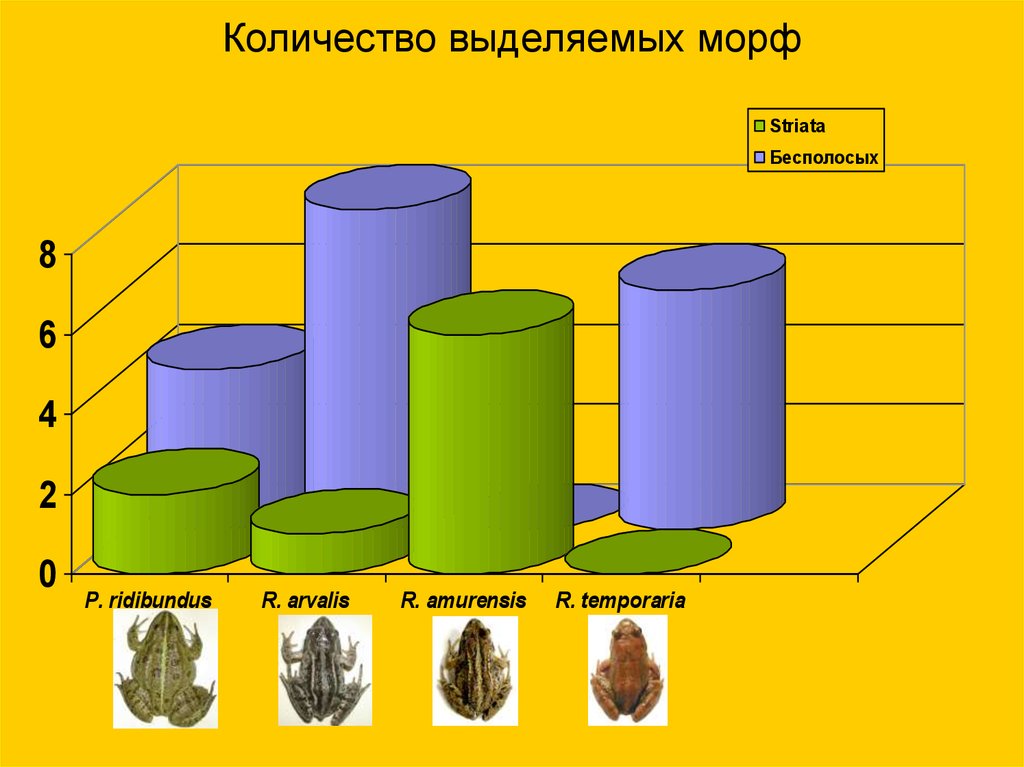
30.
Количество выделяемых морфStriata
Бесполосых
8
6
4
2
0
P. ridibundus
R. arvalis
R. amurensis
R. temporaria
31.
Встречаемость морфы striata уR.arvalis
45
40
35
30
25
20
15
10
5
0
II
Зона
III
IV
K
Се голе тки
Взрослые
32.
Возрастные изменения во встречаемостиморфы striata у R.arvalis
%
40
30
Ad
20
Juv
10
II
IV
III
Зоны
K
33. Встречаемость морфы striata у P.ridibundus
10080
60
40
% 20
0
II
Зона
III
IV
Сеголетки
Взрослые
34.
Возрастные изменения во встречаемостиморфы striata у P.ridibundus
%
100
50
0
II
III
Зона
IV
Juv
Ad
35. Средний возраст половозрелых R.arvalis
Возраст(годы)Средний возраст половозрелых R.arvalis
F
(1
,1
2
2
)=
4
.3
4
;p
<
0
.0
3
9
3
3
,2
3
,1
3
,0
2
,9
2
,8
2
,7
2
,6
2
,5
2
,4
Á
å
ñ
ïîë
îñ
û
å
S
tria
ta
Ìîð
ô
à
36.
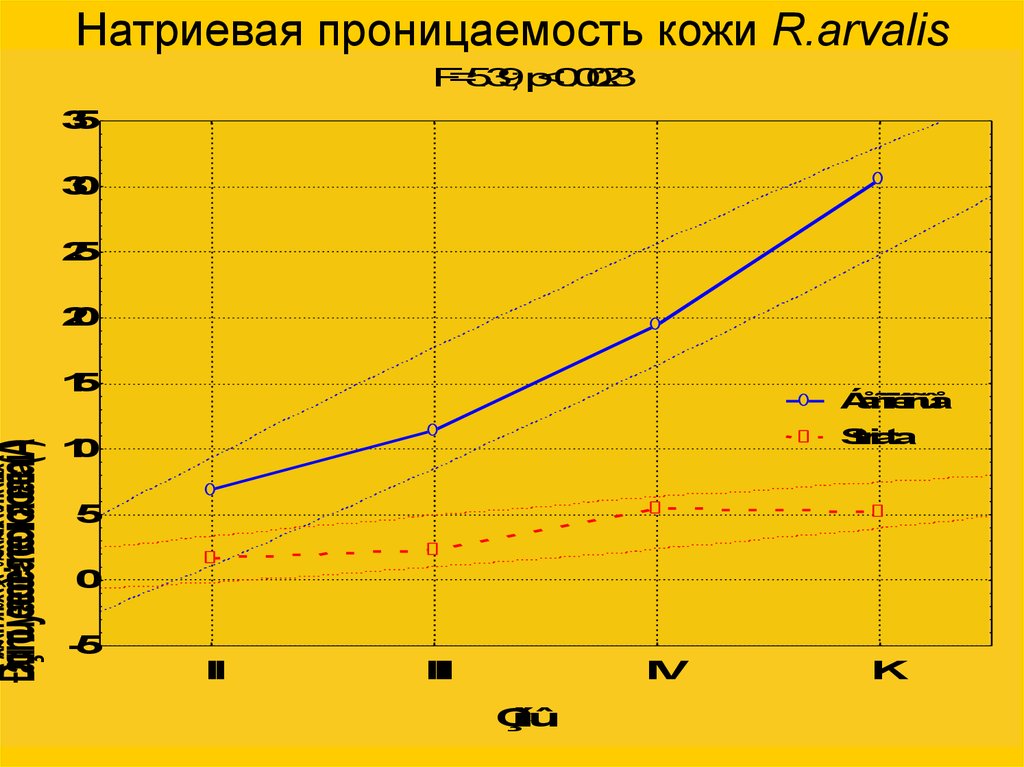
Ðàçíîñòüýëåêòðè÷åñêèõïîòåíöèàëîâ(ìÂ)Натриевая проницаемость кожи R.arvalis
F
=
5
.3
9
;p
<
0
.0
0
2
3
3
5
3
0
2
5
2
0
1
5
Á
å
ñ
ïîë
îñ
û
å
S
tria
ta
1
0
5
0
-5
II
III
IV
Ç
îíû
K
37. Уровень -активности у R.arvalis c ВУРСа
Уровеньбетаактивности(Бк/г)Уровень -активности у R.arvalis c
ВУРСа
1
8
0
0
1
6
0
0
1
4
0
0
1
2
0
0
1
0
0
0
8
0
0
6
0
0
4
0
0
2
0
0
0
Б
е
с
п
о
л
о
с
ы
е
S
tria
ta
М
о
р
ф
а
38.
Порогвозбудимости(мВ)ПОРОГ ВОЗБУДИМОСТИ НЕРВНОЙ
ТКАНИ (R.arvalis)
F
(1
,5
6
)=
6
.7
7
;p
<
0
.0
1
1
8
0
,4
8
0
,4
7
0
,4
6
0
,4
5
0
,4
4
0
,4
3
0
,4
2
0
,4
1
0
,4
0
0
,3
9
0
,3
8
Б
е
с
п
о
л
о
с
ы
е
S
tria
ta
М
о
р
ф
а
39.
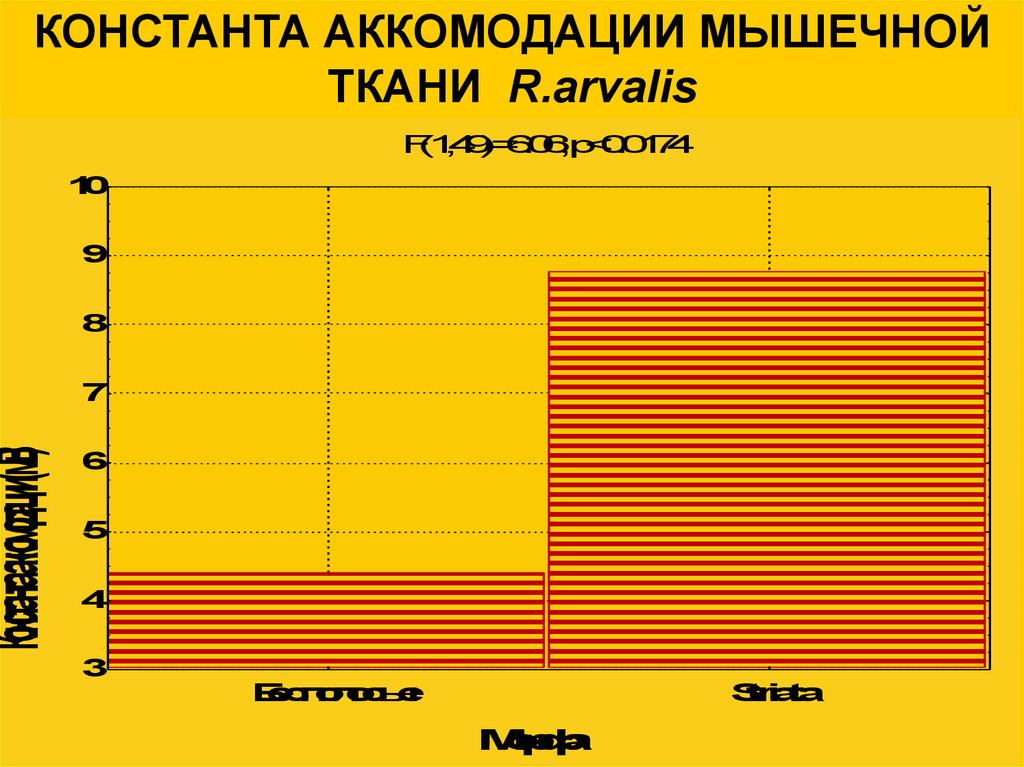
Константааккомодации(мВ)КОНСТАНТА АККОМОДАЦИИ МЫШЕЧНОЙ
ТКАНИ R.arvalis
F
(1
,4
9
)=
6
.0
6
;p
<
0
.0
1
7
4
1
0
9
8
7
6
5
4
3
Б
е
с
п
о
л
о
с
ы
е
S
tria
ta
М
о
р
ф
а
40.
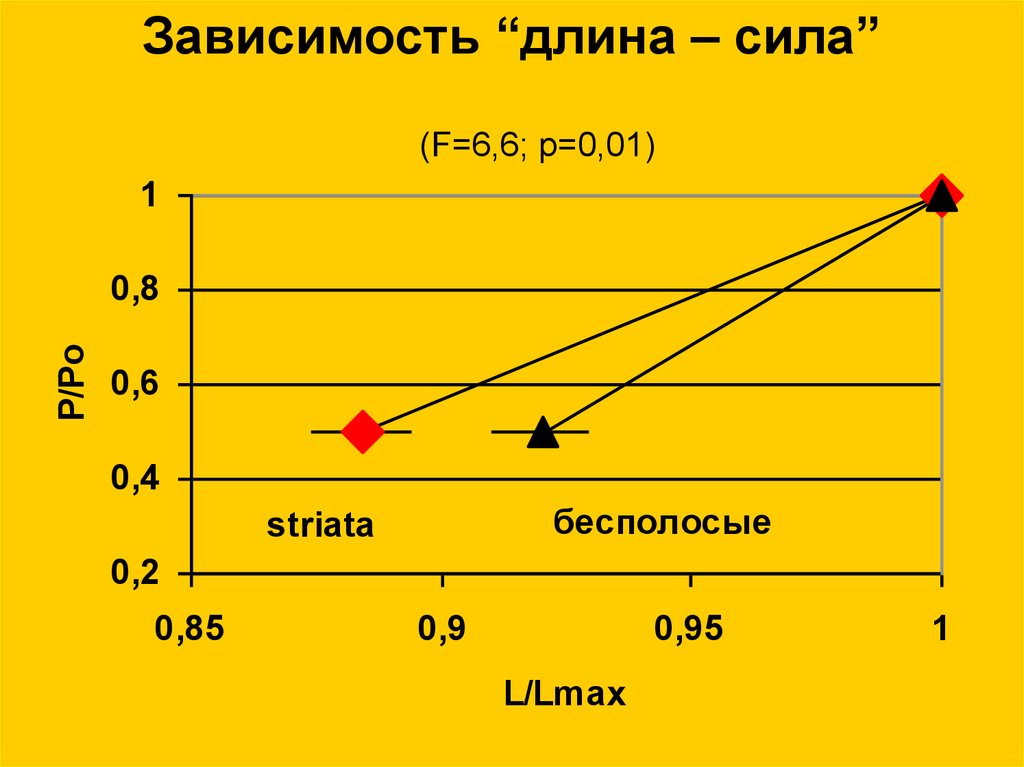
Зависимость “длина – сила”(F=6,6; p=0,01)
1
P/Po
0,8
0,6
0,4
бесполосые
striata
0,2
0,85
0,9
0,95
L/Lmax
1
41.
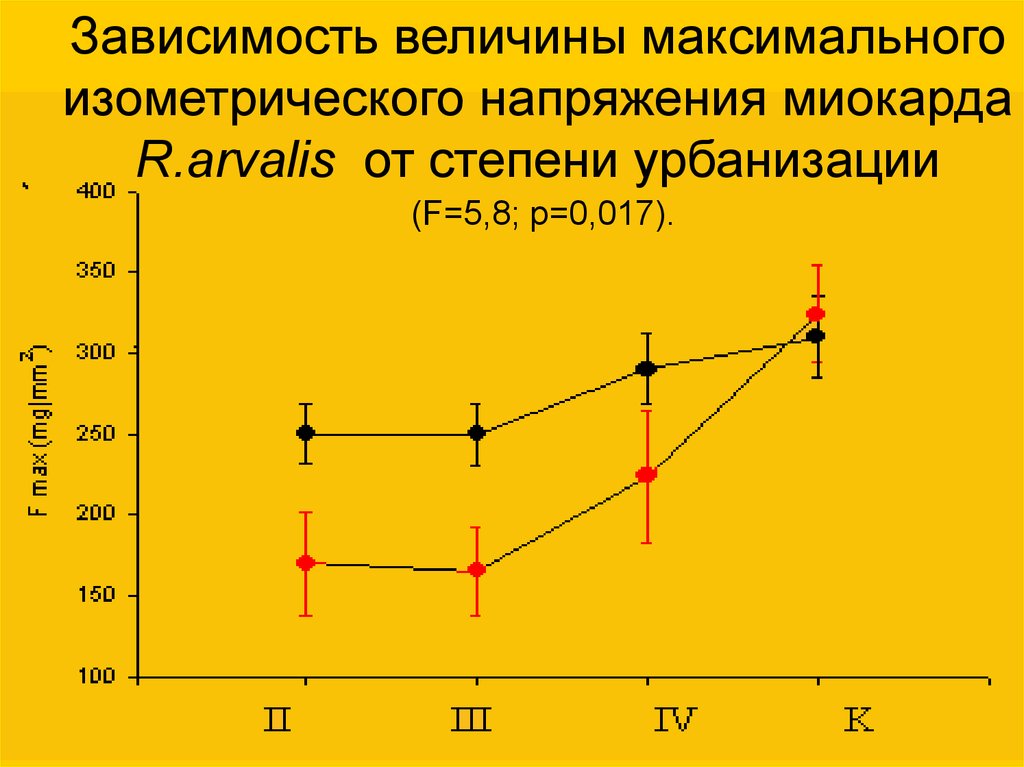
Зависимость величины максимальногоизометрического напряжения миокарда
R.arvalis от степени урбанизации
(F=5,8; p=0,017).
42.
Доля общих нейтрофиловP.ridibundus
(F=5.1, p=0.03)
28
R.arvalis
17
16
16
(F=7.995, p=0.005)
15
14
15
23
R.temporaria
13
14
12
13
Áåñïîëîñûå
Striata
8
Îáùèåíåéòðîôèëû(%)
Îáùèåíåéòðîôèëû(%)
13
12
11
10
III
IV
Çîíà
K
Striata
9
8
II
Áåñïîëîñûå
Îáùèåíåéòðîôèëû(â%)
11
18
10
9
8
7
6
II
III
IV
Çîíà
K
II
III
IV
Çîíà
K
43. Связь между количеством эритроцитов и индексом печени у R.temporaria
44. Связь между долей эритроидных предшественников и индексом печени у P.ridibundus
45.
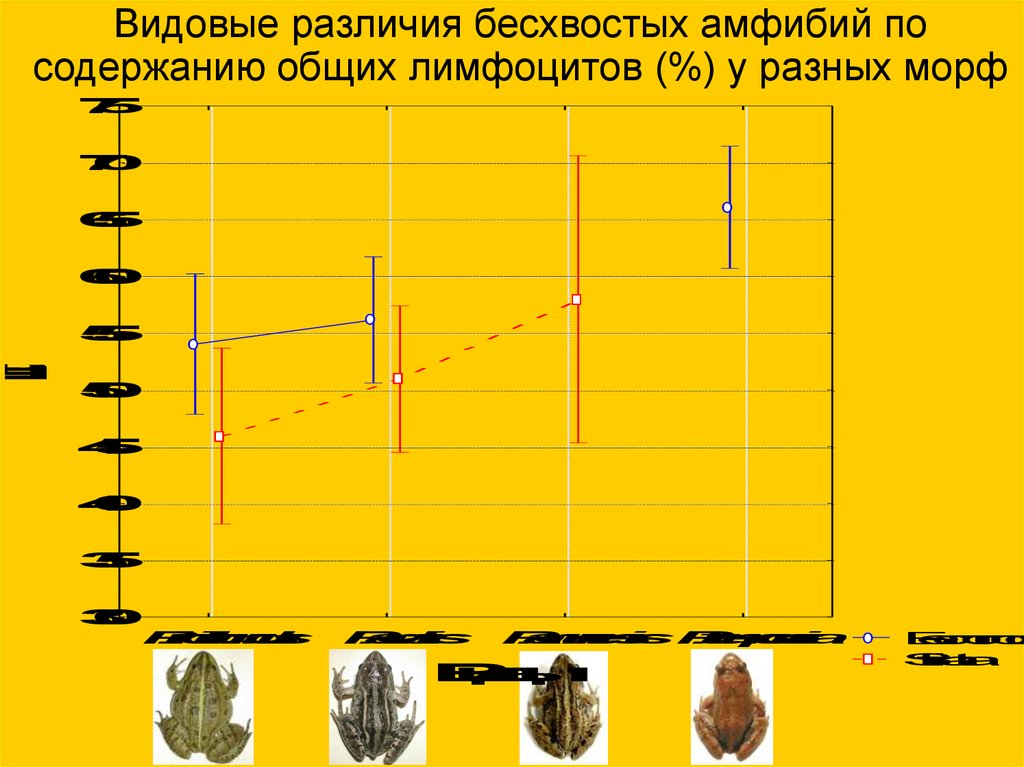
Видовые различия бесхвостых амфибий посодержанию общих лимфоцитов (%) у разных морф
7
5
7
0
6
5
6
0
LFt
5
5
5
0
4
5
4
0
3
5
3
0
P
.
r
i
d
i
b
u
n
d
u
s
R
.
a
r
v
a
l
i
s
R
.
a
m
u
r
e
n
s
i
sR
.
t
e
m
p
o
r
a
r
i
a
В
и
д
ы
Б
е
с
п
о
л
о
с
ы
S
t
r
i
a
t
a
46.
EFВидовые различия бесхвостых амфибий по
содержанию эозинофилов (%) у разных морф
2
5
2
0
1
5
1
0
5
0
5
P
.
r
i
d
i
b
u
n
d
u
s
R
.
a
r
v
a
l
i
s
R
.
a
m
u
r
e
n
s
i
sR
.
t
e
m
p
o
r
a
r
i
a
В
и
д
ы
Б
е
с
п
о
л
о
с
ы
е
S
t
r
i
a
t
a
47.
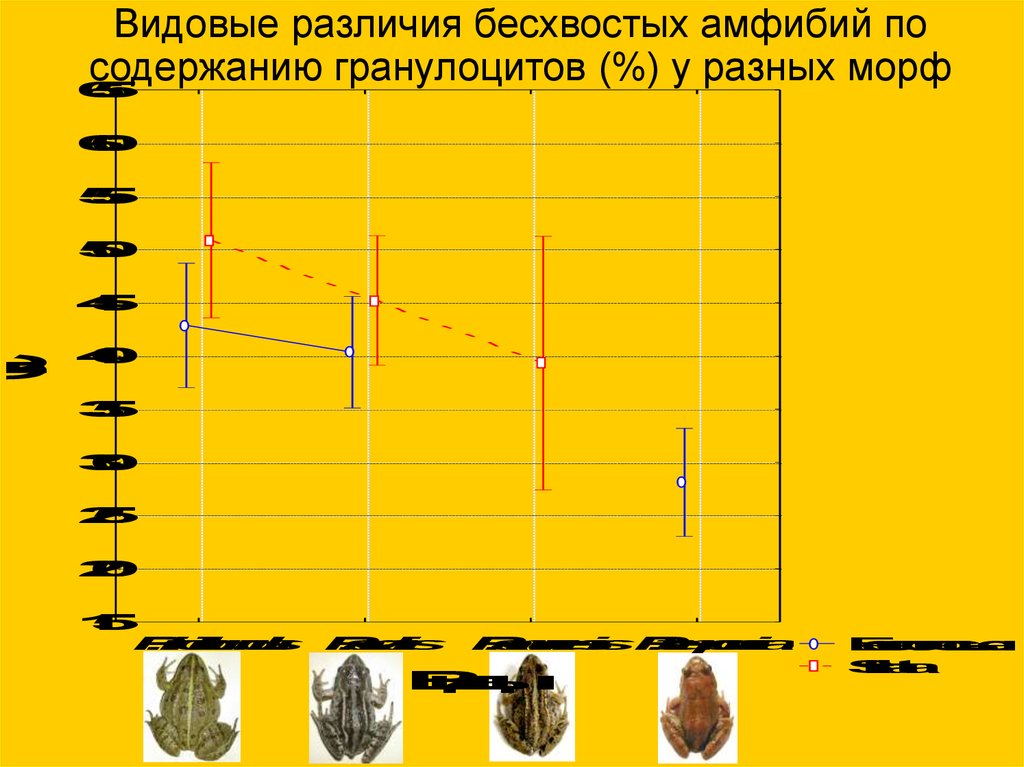
GrCВидовые различия бесхвостых амфибий по
содержанию гранулоцитов (%) у разных морф
6
5
6
0
5
5
5
0
4
5
4
0
3
5
3
0
2
5
2
0
1
5
P
.r
i
d
i
b
u
n
d
u
s
R
.
a
r
v
a
l
i
s
R
.
a
m
u
r
e
n
s
i
sR
.
t
e
m
p
o
r
a
r
i
a
В
и
д
ы
Б
е
c
п
о
л
о
с
ы
е
S
t
r
i
a
t
a
48.
ECВидовые различия бесхвостых амфибий по
содержанию эритроцитов (%) у разных морф
6
,5
E
5
6
E
5
5
,5
E
5
5
E
5
4
,5
E
5
4
E
5
3
,5
E
5
3
E
5
2
,5
E
5
2
E
5
1
,5
E
5
P
.r
id
ib
u
n
d
u
s
R
.a
r
v
a
lis
R
.t
e
m
p
o
r
a
r
ia R
.a
m
u
r
e
n
s
is
В
и
д
ы
Б
е
c
п
о
л
о
с
ы
е
S
t
r
ia
t
a
49.
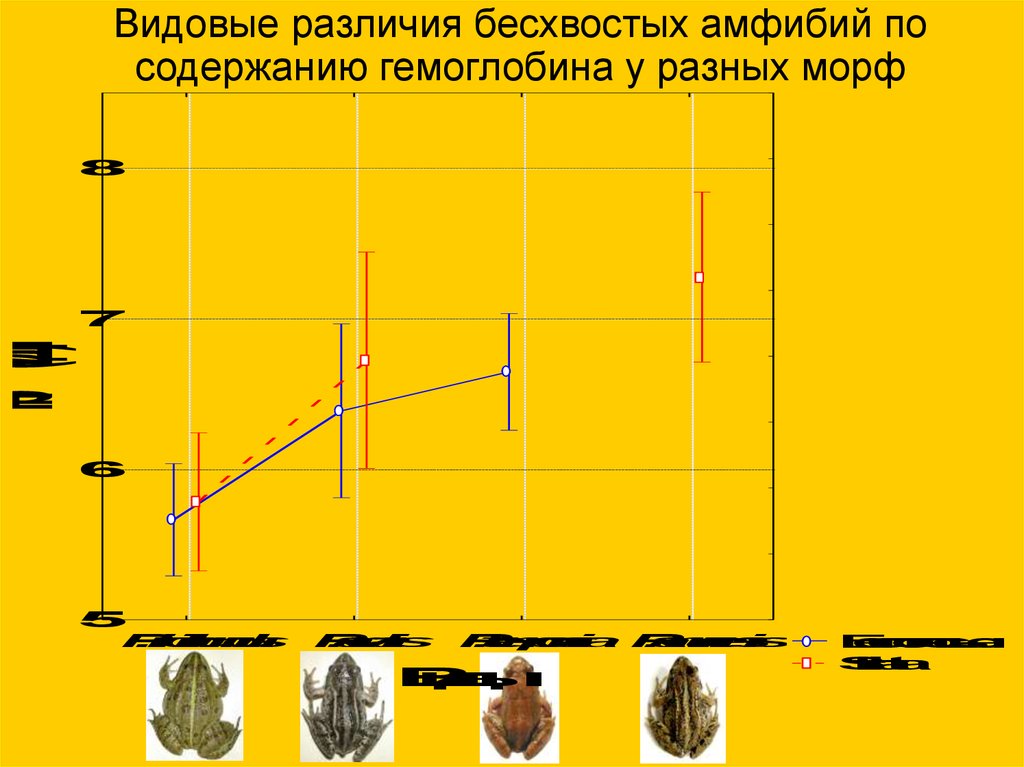
Видовые различия бесхвостых амфибий посодержанию гемоглобина у разных морф
8
Hb (г/дл)
7
6
5
P
.r
i
d
i
b
u
n
d
u
s
R
.
a
r
v
a
l
i
s
R
.
t
e
m
p
o
r
a
r
i
a R
.
a
m
u
r
e
n
s
i
s
В
и
д
ы
Б
е
c
п
о
л
о
с
ы
е
S
t
r
i
a
t
a
50.
LinkageDistanceДистанцированность бесхвостых амфибий по
изученным параметрам без учета морфы
9
0
8
0
7
0
6
0
5
0
4
0
3
0
2
0
R
.
a
m
u
r
e
n
s
i
s
R
.
t
e
m
p
o
r
a
r
i
a R
.
a
r
v
a
l
i
sP
.
r
i
d
i
b
u
n
d
u
s
51.
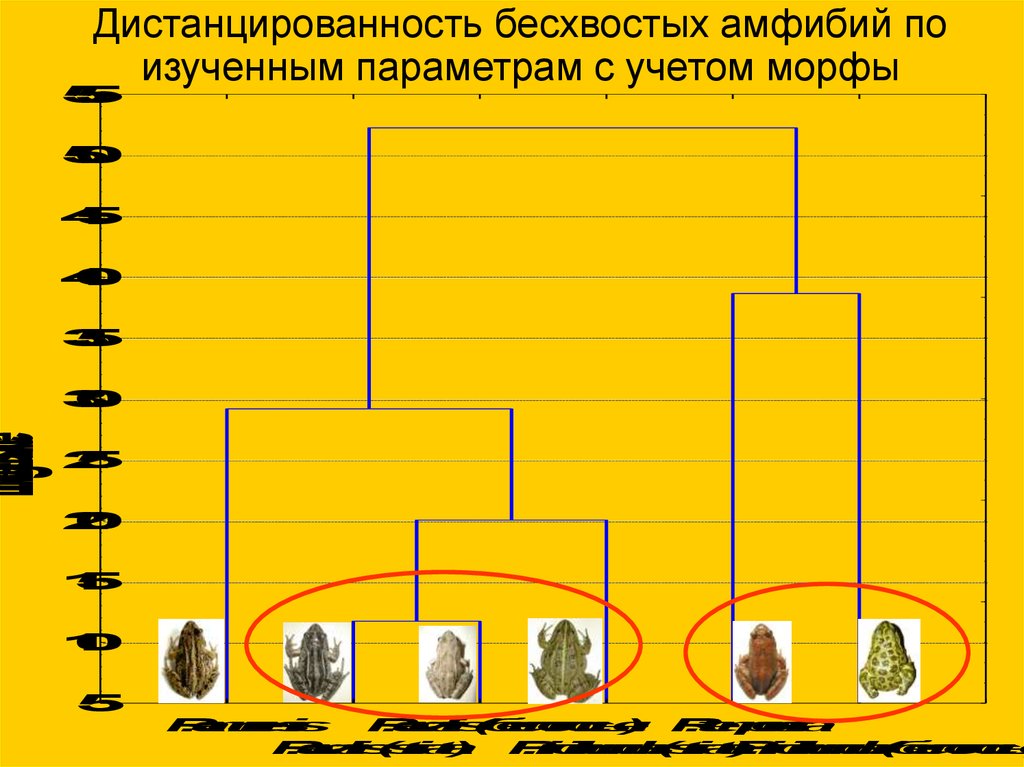
LinkageDistanceДистанцированность бесхвостых амфибий по
изученным параметрам с учетом морфы
5
5
5
0
4
5
4
0
3
5
3
0
2
5
2
0
1
5
1
0
5
R
.a
m
u
r
e
n
s
i
s
R
.a
r
v
a
l
i
s
(
б
е
с
п
о
л
о
с
ы
е
) R
.t
e
m
p
o
r
a
r
i
a
R
.
a
r
v
a
l
i
s
(
s
t
r
i
a
t
a
)
P
.r
i
d
i
b
u
n
d
u
s
(
s
t
r
i
a
t
a
P
)
.r
i
d
i
b
u
n
d
u
s
(
б
е
с
п
о
л
о
с
ы
е
52.
• Адаптивный потенциал во многом определяетсянаследственно обусловленными особенностями
физиологии, спецификой структуры полиморфизма
видов и популяций.
• Индивидуальная аккомодация и пределы ее
изменчивости являются преадаптивной основой
популяционной устойчивости к условиям
дестабилизированной среды.
• В условиях современных экосистем, доминантные
мутации могут способствовать быстрому
адаптивному успеху их носителей.
• Внутривидовое разнообразие, выражающееся в
популяционном полиморфизме - условие повышения
гомеостатичности видов - устойчивости в
меняющихся условиях среды.
53. Широкое распространение генетических механизмов, которые обусловливают и поддерживают полиморфизм, непосредственно определяется
отбором исамо по себе представляет компонент
адаптированности.
Таким образом, правильно, видимо, говорить
об “адаптивном полиморфизме”
(Э.Майр, 1974).
54. полиморфизм популяций - это универсальная стратегия, обеспечивающая сохранение целостности вида на основе постоянного
взаимодействия наследственнойизменчивости, случайного дрейфа генов
и естественного отбора в нормально
флуктуирующей среде.
55.
• Адаптация:1) нейтральное изменение генотипа,
2) приспособления живых систем к изменениям
среды,
3) процесс эволюции.
• Эпистаз:
1) отсутствие взаимодействия мутаций,
2) градуированное проявление мутаций,
3) подавление выраженности мутаций.
• Преадаптация:
1) исходно бесполезные особенности,
2) существование дискретных форм,
3) использование ранее существовавшего
признака в других целях.