




























 biology
biologySimilar presentations:








. Физиология нервов и нервных волокон. Физиология синапсов")

Законы возбудимости. Физиология нервного волокна, нерва, синапса
1. ЗАКОНЫ ВОЗБУДИМОСТИ. ФИЗИОЛОГИЯ НЕРВНОГО ВОЛОКНА, НЕРВА, СИНАПСА.
2. Зависимость возникновения возбуждения от длительности и силы раздражения
3.
Кривая «сила-длительность» описывает соотношение междудлительностью различных применяемых стимулов и их
амплитудой, необходимой для активирования нервных или
мышечных волокон при данной продолжительности стимулов. Эта
зависимость представляет собой гиперболу и имеет
обратнопропорциональный характер. Из этого следует, что ток
ниже некоторой минимальной силы или напряжения не
вызывает возбуждение, как бы длительно он не действовал, и
чем короче импульсы тока, тем меньшую раздражающую
способность они имеют.
Причиной такой зависимости является мембранная емкость.
Очень «короткие» токи просто не успевают разрядить эту емкость
до критического уровня деполяризации. Минимальная величина
тока, способная вызвать возбуждение при неограниченно
длительном его действии, называется «реобазой». Время, в течении
которого должен действовать ток удвоенной реобазы, чтобы
вызвать возбуждение, носит название "хронаксия". Оба эти порога
имеют более высокие значения для мышечных волокон, чем для нервов.
4.
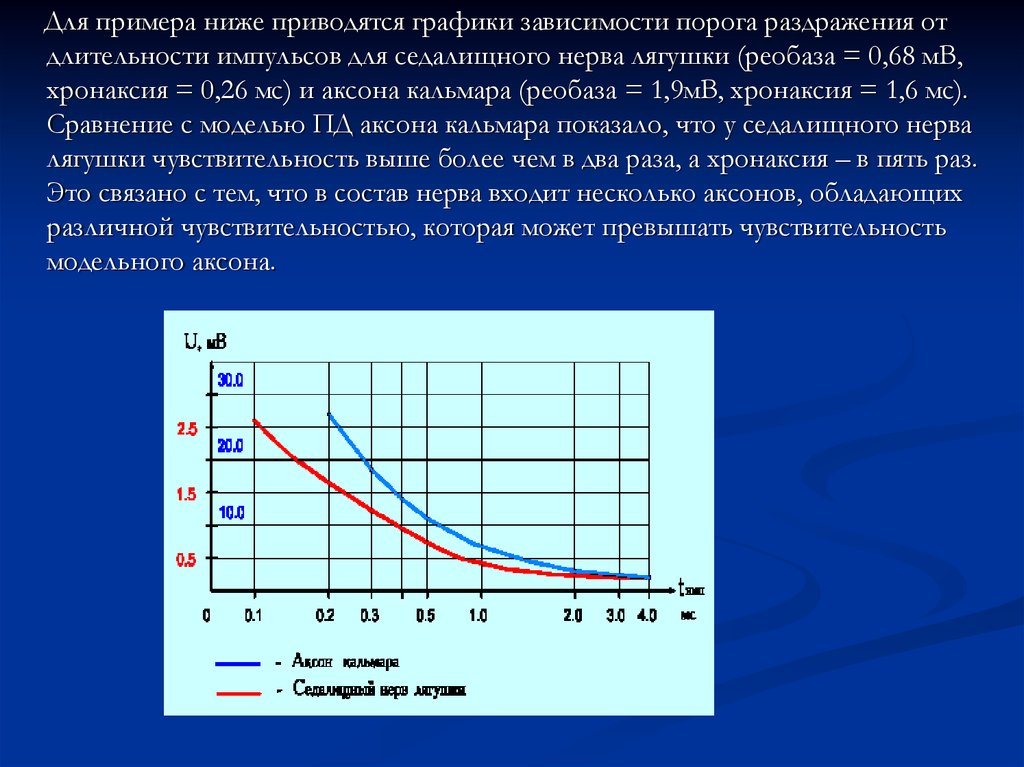
Для примера ниже приводятся графики зависимости порога раздражения отдлительности импульсов для седалищного нерва лягушки (реобаза = 0,68 мВ,
хронаксия = 0,26 мс) и аксона кальмара (реобаза = 1,9мВ, хронаксия = 1,6 мс).
Сравнение с моделью ПД аксона кальмара показало, что у седалищного нерва
лягушки чувствительность выше более чем в два раза, а хронаксия – в пять раз.
Это связано с тем, что в состав нерва входит несколько аксонов, обладающих
различной чувствительностью, которая может превышать чувствительность
модельного аксона.
5. ЗАКОН «ВСЕ ИЛИ НИЧЕГО»
Если ткань подчиняется закону, то наподпороговый раздражитель ответной реакции
нет. На пороговый или надпороговый
раздражитель возникает ответ сразу максимальной
амплитуды. Дальнейшая сила раздражения
амплитуды не изменяет.
Если ткань не подчиняется закону – ответ
градуальный, т.е. амплитуда ответа зависит от силы
действующего раздражителя. Если раздражитель
подпороговый, возникает локальный ответ, или
местный потенциал. Этот процесс не переходит в
потенциал действия.
6. ВОЗБУДИМОСТЬ И ВОЗБУЖДЕНИЕ ПРИ ДЕЙСТВИИ ПОСТОЯННОГО ТОКА НА ТКАНЬ
Мышцы и нервы – электролиты и проводят электрический ток.При накладывании на мышцу или нерв двух электродов,
соединенных с источником постоянного тока, происходит
перемещение ионов. К аноду устремляются анионы, а к катоду –
катионы.
При действии на нерв или мышцу постоянным током Е.Пфлюгер
выяснил следующие закономерности (что было подтверждено
Дюбуа – Реймоном). Постоянный ток раздражает ткань только в
момент размыкания и замыкания электрической цепи. Раздражение
ткани происходит не на всем участке, а лишь под электродами. При
замыкании тока средней силы возбуждение возникает на катоде, а
при размыкании – на аноде. Эти закономерности получили
название «полярный закон раздражения».
ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ НА КАТОДЕ НАЗЫВАЕТСЯ
КАТЭЛЕКТРОТОН, А НА АНОДЕ –АНЭЛЕКТРОТОН.
7. ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ В ПРОЦЕССЕ ВОЗБУЖДЕНИЯ
Длительность потенциала действия в мышечном волокнесоставляет около 5 мс (в нервном - 0,5 ÷ 2 мс). Следует
тут же отметить, что время абсолютной рефрактерной
фазы (АРФ), т.е. фазы, когда нервная структура совсем
невосприимчива к раздражению - независимо от ее
интенсивности - и относительной рефрактерной фазы
(ОРФ) – фазы, когда возбудимость возможна при
повышенной силе раздражающего стимула - для
мышечных волокон составляют порядка 2,5 мс и 12 мс
(соответственно) и для нервных волокон – обе фазы
около 1 мс. Далее следуют супернормальный период
(или фаза экзальтации, т.е. фаза повышенной
возбудимости) и субнормальный период (или фаза
пониженной возбудимости).
8. ОПТИМУМ И ПЕССИМУМ РИТМА И СИЛЫ РАЗДРАЖЕНИЯ.
Если каждый последующий импульс возбуждениявозникает во время фазы экзальтации, созданной
предшествующим импульсом, то мышца функционирует
в наиболее оптимальном ритме, а этот ритм раздражений
называется ОПТИМУМ. Например, максимальный ритм
для двигательного нерва лягушки составляет 500, а
оптимальный 100-150 импульсов.
При очень частых раздражениях, превышающих
оптимальных ритм, сокращения мышцы уменьшаются и
даже совсем прекращаются – это ПЕССИМУМ ритма
раздражения. Пессимум возникает в том случае, когда
частота раздражений превышает меру лабильности.
9.
Последующее раздражения приходится на фазуабсолютной рефрактерности. Хотя это раздражение и не
вызывает ответной реакции, оно все же не остается без
последствий: углубляется фаза абсолютной
рефрактерности и затрудняется действие следующих за
ним раздражений. Поэтому каждое новое раздражение,
посылаемое в очень частом ритме, снижает лабильность
ткани и вызывает не возбуждение, а качественно новое
состояние – ТОРМОЖЕНИЕ.
Эти положения, разработанные Н.Е.Введенским,
получили дальнейшее развитие и уточнение в результате
описания механизма синаптической передачи. В нервно –
мышечном препарате наименьшая возбудимость и
лабильность выявлены у синапса. Поэтому синапс и
лимитирует частоту раздражений, передаваемых с нерва
на мышцу.
10.
Редкие или оптимальные по частоте импульсыпроводятся синапсом без изменений. При этом каждый
импульс попадает в фазу убывания постсинаптического
потенциала, вызванного предыдущим импульсом. При
частом раздражении постсинаптические потенциалы
суммируются, что приводит к стойкой деполяризации
постсинаптической мембраны и развитию блока
проведения – развитию пессимума. Это явление получило
название ПЕССИМАЛЛЬНОЕ ТОРМОЖЕНИЕ.
Явление оптимума и пессимума - общебиологические
свойства живой ткани, образующиеся в ответ на любые
физические или химические воздействия. Умеренные
раздражения оказывают оптимальное (возбуждающее)
действие, а раздражения большей силы, частоты и
длительности вызывают тормозящее действие на реакции
различных органов как при непосредственном, так и при
рефлекторном воздействии на них.
11. Функциональная подвижность - лабильность
Свойство лабильности открыл Н.Е.Введенский (1892), изучаядействие ритмических раздражений различной частоты на нервномышечный препарат.
В каждой ткани одиночный импульс возбуждения продолжается
определенное время. Поэтому определение лабильности
следующее: СКОРОСТЬ ПЕРЕХОДА ВОЗБУДИМОЙ ТКАНИ ИЗ
СОСТОЯНИЯ ПОКОЯ В СОСТОЯНИЕ ВОЗБУЖДЕНИЯ И НАОБОРОТ
БЕЗ ТРАНСФОРМАЦИИ РИТМА ДЕЙСТВУЮЩЕГО РАЗДРАЖИТЕЛЯ.
Измеряется лабильность в ПД/с. Мера лабильности – это
максимальное число импульсов возбуждения, которые возникают за
одну секунду в ответ на такое же максимальное число раздражений.
В ПРОЦЕССЕ РОСТА И РАЗВИТИЯ ЛАБИЛЬНОСТЬ УВЕЛИЧИВАЕТСЯ,
ПРИ СТАРЕНИИ УМЕНЬШАЕТСЯ.
ИЗМЕНЕНИЕ ЛАБИЛЬНОСТИ В СТОРОНУ ПОВЫШЕНИЯ ИЛИ
ПОНИЖЕНИЯ ПО СРАВНЕНИЮ С ИСХОДНЫМ УРОВНЕМ В СВЯЗИ С
ДЕЯТЕЛЬНОСТЬЮ ТКАНИ НАЗЫВАЮТ УСВОЕНИЕ РИТМА. Лучше
усваивается частый ритм при невысокой исходной лабильности.
12.
Например: при стимуляции нервного волокна, с частотой400 импульсов в секунду, по нервному волокну будет
проводиться каждый импульс. При увеличении
стимуляции до 700 импульсов в секунду, будет
проводиться каждый второй импульс. При еще
большей частоте (800 импульсов), будет проводиться
каждый третий импульс. Однако при повышенной
частоте стимуляции лабильность нервной ткани может
повыситься и при частоте 700 импульсов вначале
будет проводиться каждый второй, а затем каждый
импульс. Однако, повышение лабильности не
безгранично, и через некоторое время может снизиться
проводимость возбудимого образования. Наибольшей
лабильностью отличаются аксоны, способные
воспроизводить до 500-1000 импульсов в 1 сек.; менее
лабильны синапсы, например двигательное нервное
окончание может передать на скелетную мышцу не более
100-150 возбуждений в 1 сек
13. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ ПО НЕРВНЫМ ВОЛОКНАМ
Потенциал действия или нервный импульс можетвозникать в любой точке возбудимой мембраны нервного
или мышечного волокна и способен распространяться
вдоль ее поверхности. При этом роль потенциала
действия заключается в передаче информации по
нервным волокнам от тела нейрона к нервному
окончанию. Когда потенциалы действия достигают
терминалей аксона, то информация передается на другие
нейроны благодаря выделению из нервных окончаний
молекул медиаторов. В мышечных клетках потенциалы
действия распространяются по сарколемме и активируют
механизм сокращения мышц. Проведение нервного
импульса от тела нейрона к окончанию аксона
различается в немиелинизированных и
миелинизированных нервных волокнах.
14.
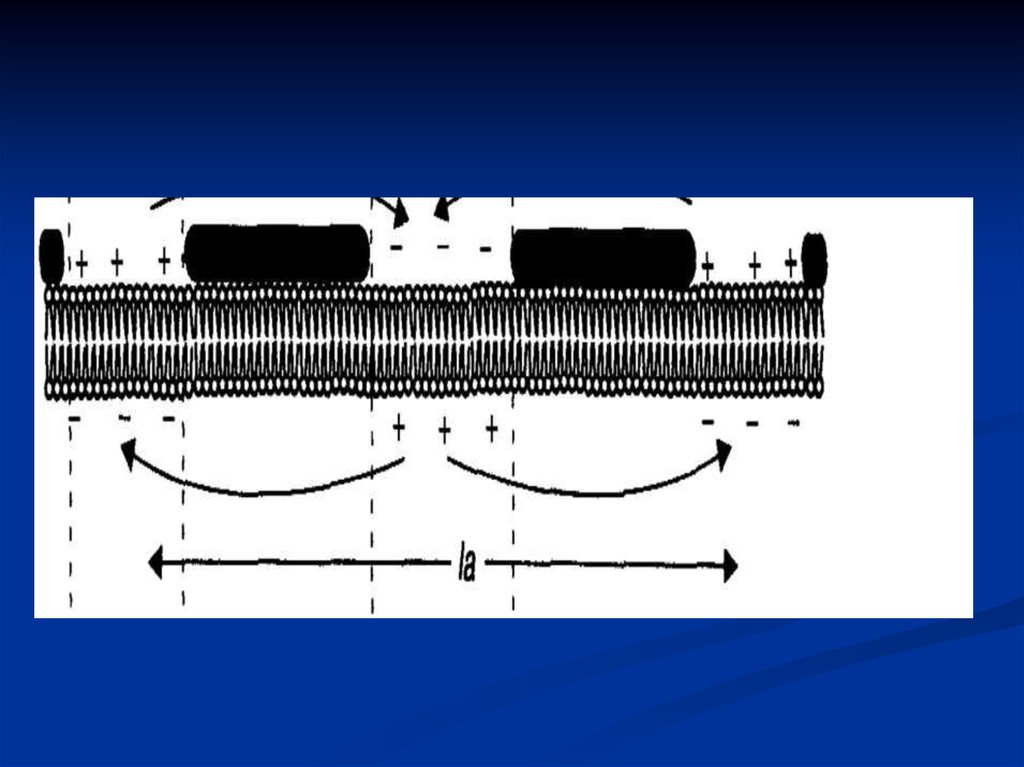
15. НЕМИЕЛИНИЗИРОВАННОЕ ВОЛОКНО
Проведение потенциала действия понемиелинизированному нервному волокну происходит
путем активации потенциалзависимых натриевых ионных
каналов участка мембраны волокна, прилегающего к тому
месту, где возник потенциал действия. При этом между
возбужденным и невозбужденным участками мембраны
нервного волокна возникают локальные электрические
токи, которые вызывают деполяризацию мембраны
невозбужденного участка до критического уровня, и
только после этого в мембране нервного волокна
мгновенно открываются потенциалзависимые натриевые
каналы. ПД генерируется в соседнем участке нервного
волокна.
16. МИЕЛИНИЗИРОВАННОЕ ВОЛОКНО
В миелинизированном нервном волокне потенциалыдействия генерируются только в области перехвата
Ранвье, т. е. той части мембраны, которая не покрыта
шванновскими клетками. С одной стороны, это
обусловлено тем, что цитоплазма шванновской клетки
содержит липид - сфингомиелин, который уменьшает поток
ионов через мембрану нервного волокна примерно в 5000
раз и снижает ее емкость в 50 раз. С другой стороны, в
области перехвата Ранвье в мембране нервного волокна
имеется наибольшее число потенциалзависимьтх
натриевых ионных каналов, а сама мембрана обладает
нормальным уровнем возбудимости. При нанесении
точечного раздражения на миелинизированное нервное
волокно потенциал действия генерируется в зоне
перехвата Ранвье и возникают электрические токи,
которые текут вдоль силовых линий от плюса к минусу
потенциала на мембране.
17.
Одновременно в аксоплазме нервного волокна возникаютпродольные токи, которые направлены от места генерации
потенциала действия в обе стороны. Продольные токи вызывают
открывание потенциалзависимьтх натриевых ионных каналов, а
следовательно, движение ионов натрия через мембрану и
генерацию ПД в соседних перехватах Ранвье, минуя часть нервного
волокна, покрытого шванновскими клетками. Поскольку ПД
передаются от одного перехвата Ранвье к другому, то механизм
проведения потенциалов действия в миелинизированных нервных
волокнах получил название сальтаторного. В миелинизированных
нервных волокнах реполяризация мембраны после потенциала
действия происходит с высокой скоростью. В результате нервные
волокна имеют высокую функциональную лабильность и
способны проводить значительное число потенциалов действия в
единицу времени. Скорость распространения ПД в зависимости от
диаметра и типа миелинизированных нервных волокон высока. В
толстых миелинизированных нервных волокнах скорость
проведения возбуждения пропорциональна диаметру волокна, а
проводимость нервного волокна имеет обратную зависимость от
его диаметра.
18. ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНОМУ ВОЛОКНУ
Проведение возбуждения по нервному волокну возможно приусловии его структурной целостности и физиологической
непрерывности (ЗАКОН ФИЗИОЛОГИЧЕСКОЙ
НЕПРЕРЫВНОСТИ). Физиологическая непрерывность нерва
может быть нарушена, например, при сдавлении нерва без его
структурного повреждения, что препятствует проведению
потенциалов действия. При проведении возбуждения по нервному
волокну потенциал действия не распространяется с одного волокна
на другое, например рядом расположенное (ЗАКОН
ИЗОЛИРОВАННОГО ПРОВЕДЕНИЯ). Несмотря на то что
нервные волокна проводят потенциалы действия либо в
направлении тела нейрона (центростремительно), либо от тела
нервной клетки к окончаниям аксона (центробежно), отдельное
нервное волокно обладает двусторонней проводимостью (ЗАКОН
ДВУСТОРОННЕГО ПРОВЕДЕНИЯ). При искусственном
электрическом раздражении в любой точке по ходу нервного
волокна может возникать потенциал действия и распространяться
как центростремительно, так и центробежно.
19. ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА НЕРВНЫХ ВОЛОКОН
ВОЗБУДИМОСТЬ (у миелиновых выше, чем у безмиелиновых)ЛАБИЛЬНОСТЬ (у миелиновых выше, чем у безмиелиновых)
ИЗОЛИРОВАННОЕ ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ
ДВУСТОРОННЕЕ ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ
СКОРОСТЬ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ (зависит от толщины
нервного волокна и наличия или отсутствия миелиновой
оболочки). Классифицируют все волокна по классам А, В и С.
А – толстые миелиновые, В – тонкие миелиновые, С – тонкие
безмиелиновые.
ОБМЕН ВЕЩЕСТВ. Интенсивность обмена веществ в нерве
очень мала. Во время возбуждения обмен веществ в нерве
усиливается возрастает потребление кислорода, глюкозы и
выделение двуокиси углерода.
УТОМЛЕНИЕ. Нерв практически неутомляем. Это доказано
Н.Е.Введенским.
20. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ ЧЕРЕЗ СИНАПС
Передача возбуждения с отростка одной нервной клетки наотросток или тело другой нервной клетки возможна двумя
способами: электрическим и химическим. Электрический способ
передачи возбуждения осуществляется с помощью щелевых
контактов (НЕКСУСОВ) в том случае, если мембраны клеток
отделены щелью размером примерно 2 нм. Щелевые контакты
распространены, например, в миокарде и гладких мышцах
пищеварительного тракта и представляют собой межклеточные
тубулярные белковые гидрофильные каналы, состоящие их шести
молекул интегральных БЕЛКОВ-КОННЕКСОНОВ. Передача
возбуждения электрическим способом осуществляется аналогично
его проведению по нервным волокнам с помощью местных токов,
возникающих между деполяризованным участком мембраны
нервного волокна и поляризованным участком мембраны нервной
клетки. Локальные токи деполяризуют мембрану нейрона до
критического уровня, после чего возникает спонтанный процесс
регенеративной деполяризации.
21.
Электрическая передача возбуждения осуществляется с высокойскоростью, близкой к скорости проведения возбуждения по
нервным волокнам.
Основным способом передачи возбуждения между нейронами, а
также между нервными и мышечными клетками является
химический, который реализуется с помощью синапсов. СИНАПС
— это специализированное по структуре и функции место контакта
мембран между двумя возбудимыми клетками, которое обеспечивает
передачу импульсов от одной возбудимой клетки к другой с
помощью молекул химических веществ, или медиаторов. В нервной
системе проведение возбуждения осуществляется между отростками
нейронов и между отростками и телами нервных клеток. Так,
проведение возбуждения с аксона одного нейрона на аксон другого
происходит при участии аксо-аксональных синапсов. Передача
импульсов от аксона нейрона на на дендриты другой нервной
клетки осуществляется аксо-дендритными синапсами, а от одних
дендритов на другие — с помощью дендро-дендритных синапсов .
22.
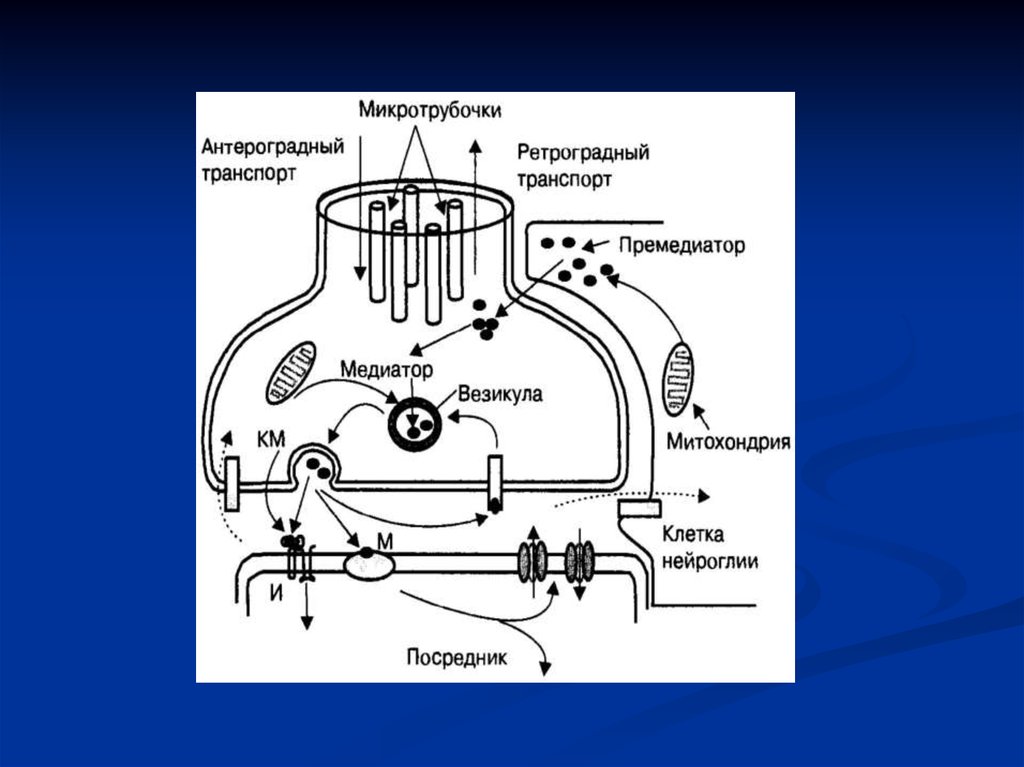
23. Функция пресинаптического окончания нейронов
В нервных терминалях локализовано два типа везикул — мелкие икрупные. Мелкие везикулы терминалей нейронов центральной и
периферической нервной системы содержат разные группы
медиаторов: амины (ацетилхолин, дофамин, норадреналин,
серотонин и гистамин), аминокислоты (глютамат, глицин, гаммааминомасляная кислота) и пурины (АТФ, аденозин). Крупные или
пептидные везикулы нервных терминалей центральной и
периферической нервной систем содержат нейроактивные пептиды
(субстанция Р, вазопрессин, окситоцин, кортиколиберин,
тиреолиберин, соматолиберин, гонадолиберин, энкефалины,
ангиотензин 2 и др.). В отличие от мелких везикул пептидные
синтезируются в соме нейронов, наполняются молекулами
пептидов и транспортируются с помощью аксотока в нервное
окончание. Функции медиаторов мелких и крупных везикул
взаимосвязаны между собой в процессе синаптической
передачи: первые обеспечивают химическую передачу возбуждения
через синапс, а вторые — модулируют, т. е. усиливают или
уменьшают этот процесс.
24.
25. ВЕЗИКУЛЯРНЫЙ МЕХАНИЗМ ЭКЗОЦИТОЗА МЕДИАТОРА ИЗ НЕРВНОЙ ТЕРМИНАЛИ
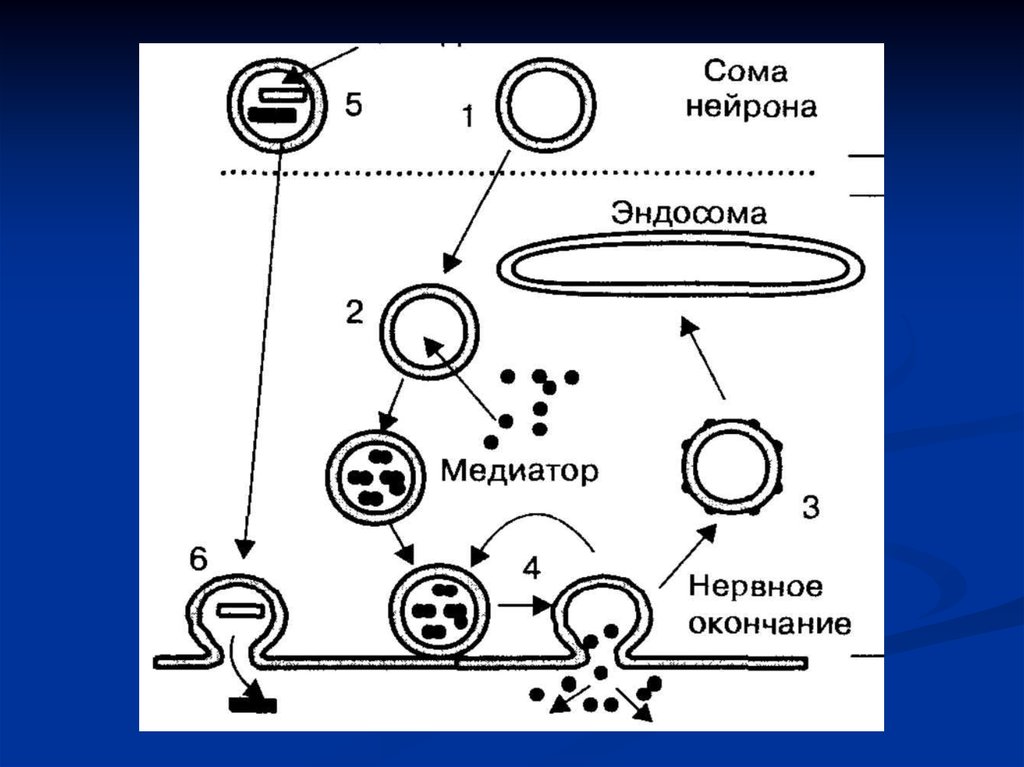
Везикулы (1) формируются в теле нейрона,транспортируются в нервное окончание, где
заполняются медиатором (2). При экзоцитозе, с
помощью механизма полного слияния и с
последующим образованием покрытых везикул
(З), запасы медиатора пополняются через стадию
эндосом. При неполном слиянии и повторном
заполнении медиатором везикулы сразу могут
участвовать в экзоцитозе (4). Крупные везикулы
заполняются пептидами в теле нейрона (5), их
экзоцитоз происходит в других зонах нервного
окончания (6).
26. ПРЕСИНАПТИЧЕСКИЙ МЕХАНИЗМ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
Пресинаптический механизм проведения возбужденияосуществляет трансформацию энергии нервных импульсов в
кванты вьделяющегося медиатора. В фазу деполяризации
потенциала действия в мембране нервной терминали открываются
потенциалзависимые кальциевые ионные каналы, и ионы кальция
по концентрационному градиенту поступают из внеклеточной
среды внутрь нервного окончания. Ионы Са вблизи внутренней
поверхности пресинаптической мембраны образуют микродомен и
связываются со специфическим везикулярным белком кальциевым
сенсором — синаптотагмином. Последовательное взаимодействие
специфических белков экзоцитоза, локализованых на мембране
везикул (синапсин, синаптотагмин, синаптобревин) и на
пресинаптической мембране (синтаксин, синапсоассоциированный
белок) связывает везикулу с активной зоной пресинаптической
мембраны, формируя секретосому. При этом мембрана везикулы
полностью сливается с пресинаптической мембраной, и медиатор
освобождается в синаптическую щель через пору.
27.
В синапсах нервной системы экзоцитоз медиаторов можетпроходить через постоянную или временную пору. В первом
случае происходит полное слияние мембран везикулы и
пресинаптического окончания и высвобождение всего количества
молекул медиатора из везикул. Во втором случае не происходит
полного слияния мембран везикулы и пресинаптической терминали. При образовании временной поры освобождается только часть
медиатора в синаптическую щель, а затем везикула отделяется от
активной зоны и возвращается в цитоплазму нервной терминали.
Этот механизм позволяет одной и той же везикуле без пополнения
запаса медиатора неоднократно участвовать в экзоцитозе.
После высвобождения молекулы нейротрансмиттера
диффундируют от мест экзоцитоза к постсинаптической мембране.
Во время диффузии часть медиатора может подвергаться
ферментному разрушению в синаптической щели или
захватываться обратно внутрь нервного окончания.
Постсинаптической мембраны могут достигать все или только
часть молекул нейротрансмттеров, что обусловливает
соответственно большее или меньшее количество передаваемых
через аксосоматический синапс импульсов.
28. ПОСТСИНАПТИЧЕСКИЙ МЕХАНИЗМ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
Постсинаптический механизм проведения возбуждения ваксосоматическом синапсе нервной системы осуществляет передачу
нервного импульса на постсинаптический нейрон (быстрые реакции).
Передача нервного импульса от одного нейрона к другому
осуществляется в течение короткого отрезка времени (мс). Этот
процесс на этапе постсинаптического механизма передачи
возбуждения осуществляется при участии ионотропных рецепторов
постсинаптической мембраны. Ионотропные рецепторы представляют
собой часть молекулярной структуры хемозависимых ионных
каналов (натриевых, калиевых, кальциевых) постсинаптической
мембраны нейронов. Количество ионотропных рецепторов на
постсинаптической мембране нейронов является регулируемой
величиной: оно повышается в ответ на длительное воздействие на
мембрану нейрона малой концентрации молекул медиаторов
(явление сенситизации). Напротив, число рецепторов на
постсинаптической мембране понижается в ответ на длительное
воздействие на постсинаптическую мембрану медиаторов в
высокой концентрации (явление десенситизации). Вещества, которые
связываются с рецепторами хемозависимых ионных каналов
постсинаптической мембраны, называются лигандами. Лиганды
подразделяются на агонисты и антагонисты.
29.
Результатом взаимодействия медиаторов с ионотропнымирецепторами постсинаптической мембраны является ее
кратковременная деполяризация или гиперполяризация
постсинаптической мембраны. ДЕПОЛЯРИЗАЦИЯ
ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЫ В ОБЛАСТИ
ОДНОГО СИНАПСА НАЗЫВАЕТСЯ ВОЗБУЖДАЮЩИМ
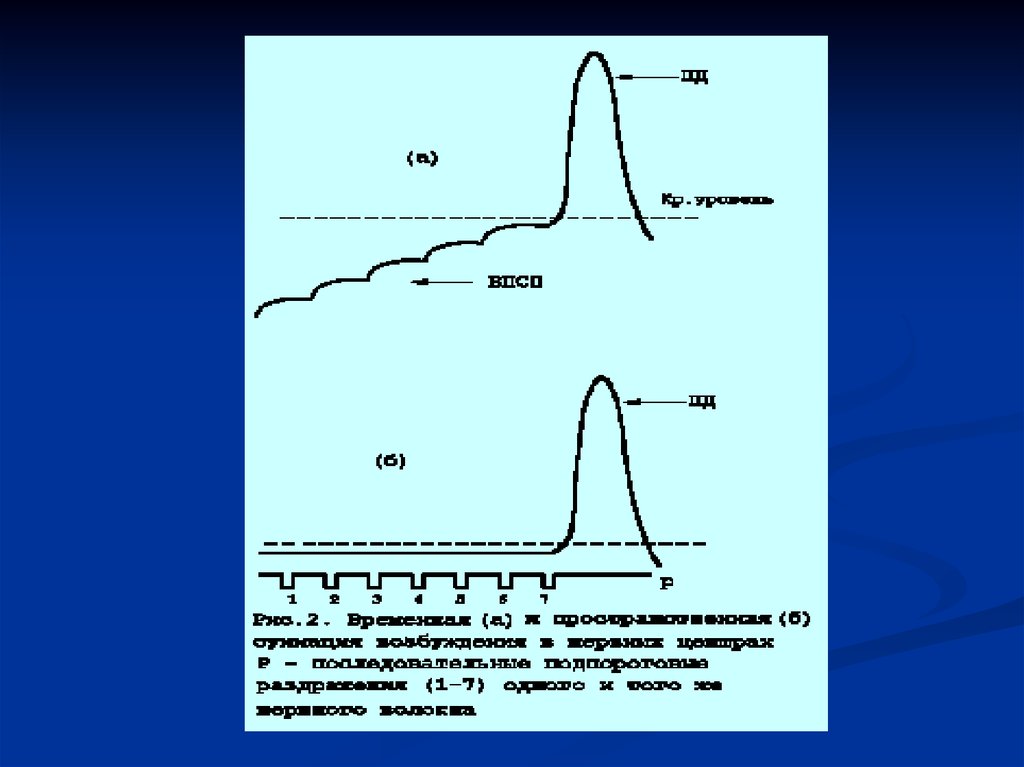
ПОСТСИНАПТИЧЕСКИМ ПОТЕНЦИАЛОМ (ВПСП).
Химические синапсы, в которых при передаче возбуждения
возникают ВПСП, называются возбуждающими. По аналогии
медиатор, вызывающий возникновение ВПСП, называется
возбуждающим медиатором. Возникновение ВПСП обусловлено
открытием преимущественно натриевых ионных каналов. ПРИ
ВПСП ПРОИСХОДИТ ДЕПОЛЯРИЗАЦИЯ
ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЫ И УМЕНЬШЕНИЕ
ПОРОГА ЕЕ ВОЗБУДИМОСТИ. Амплитуда ВПСП прямо
зависит от количества молекул медиатора взаимодействующих с
рецептора ми постсинаптической мембраны, а также
последовательностью выделения квантов медиатора из
пресинаптического окончания.
30.
Результатом взаимодействия медиаторов с ионотропнымирецепторами хемозависимых ионных каналов постсинаптической
мембраны может быть ее кратковременная гиперполяризация. В
результате понижается возбудимость постсинаптического нейрона
и в нем не происходит генерация потенциала действия.
ГИПЕРПОЛЯРИЗАЦИЯ ПОСТСИНАПТИЧЕСКОЙ
МЕМБРАНЫ В ОБЛАСТИ ОДНОГО СИНАПСА
НАЗЫВАЕТСЯ ТОРМОЗНЫМ ПОСТСИНАПТИЧЕСКИМ
ПОТЕНЦИАЛОМ (ТПСП), такие химические синапсы называются
тормозными. По аналогии медиаторы, вызывающие ТПСП,
называются тормозными медиаторами. Взаимодействие тормозного
медиатора с рецепторами постсинаптической мембраны открывает
хлорные и калиевые ионные каналы, что резко повышает ионную
проводимость постсинаптической мембраны для ионов хлора или
калия. В соответствии с электрохимическими градиентами ионов
калия и хлора по обе стороны от постсинаптической мембраны,
ТПСП ПРЕДСТАВЛЯЕТ СОБОЙ СДВИГ ПОТЕНЦИАЛА НА
ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ В СТОРОНУ
УВЕЛИЧЕНИЯ ЕГО ВЕЛИЧИНЫ. В результате происходит
гиперполяризация постсинаптической мембраны и увеличение порога
ее возбудимости.
31.
32. ЯВЛЕНИЕ ПАРАБИОЗА
В классическом труде «Возбуждение, торможение инаркоз» (1901) Н.Е. Введенский изложил свои
представления о парабиозе (от греческих слов para –
около, bios – жизнь, т.е. состояние на грани жизни). Эта
теория объясняет каким образом возбуждение переходит
в торможение. Последнее состояние может закончиться
прекращением жизнедеятельности ткани.
ОПЫТ ВВЕДЕНСКОГО
На среднюю часть нерва нервно-мышечного препарата
воздействуют эфиром, хлороформом, холодом, теплом,
сильным электрическим током. Под влиянием этих
раздражителей данный участок изменяется, или
альтерируется. На этот участок, а также выше и ниже,
накладывали электроды индукционной катушки.
Электрическую активность нерва измеряли с помощью
33.
1.2.
3.
гальванометра. До действия альтерирующего вещества
высота тетанического сокращения мышцы зависела от
силы или частоты раздражения: чем больше сила или
частота, тем выше сокращение мышцы.
СТАДИИ ПАРАБИОТИЧЕСКОГО ПРОЦЕССА
Уравнительная, или трансформирующая. Через
некоторое время мышца на разные по силе и частоте
раздражения начинала отвечать одинаковыми
сокращениями.
Парадоксальная стадия. Во время нее на сильные и
частые раздражения – слабая ответная реакция, или
вообще ее отсутствие, а на редкие и слабые –
неожиданно сильная.
Стадия торможения. При воздействии на нерв
раздражителем любой силы и частоты мышца не
сокращается.
34.
Стадия торможения заканчиваетсясостоянием, при котором отсутствуют
видимые проявления жизни – возбудимость
и проводимость. Это состояние называется
парабиоз.