






















 biology
biologySimilar presentations:










Закономерности передачи генетической информации. Генетический код
1.
Закономерности передачи генетическойинформации. Генетический код.
2.
История изучение наследования признаковВ начале XIX века Дж. Госс (John Goss), экспериментируя с горохом, показал, что
при скрещивании растений с зеленовато-голубыми горошинами и с желтоватобелыми в первом поколении получались жёлто-белые. Он впервые установил
наличие признаков, названных Г. Менделем «рецессивными».
Огюстен Сажрэ, французский растениевод, проводил
эксперименты по гибридизации тыквенных, главным
образом дынь. Сажрэ впервые в истории гибридизации
стал изучать отдельные признаки скрещивающихся
растений (мякоть, кожура и т. д.). Он установил, что при
гибридизации родительские признаки распределяются
между потомками без всякого смешения между собой.
Таким образом, Сажрэ пришёл к установлению
решающего свойства наследственности: в своей статье
«Соображения об образовании гибридов, вариант и
разновидностей» (1825 г.) он указывал на наличие
наследственности «константной» вместо «слитной»
наследственности.
3.
Таким образом, к середине XIXвека
было
открыто
явление
доминантности,
единообразие
гибридов в первом поколении (все
гибриды первого поколения похожи
друг на друга), расщепление и
комбинаторику признаков во втором
поколении. Тем не менее, Мендель,
высоко
оценивая
работы
предшественников, указывал, что
всеобщего закона образования и
развития гибридов ими не было
найдено, и их опыты не обладают
достаточной достоверностью для
определения
численных
соотношений.
4.
В 1865 году монах Грегор Мендель обнародовал на заседании местного обществаестествоиспытателей результаты исследований о передаче по наследству признаков
при скрещивании гороха (работа «Опыты над растительными гибридами» была
опубликована в трудах общества в 1866 г.). Мендель показал, что наследственные
задатки не смешиваются, а передаются от родителей к потомкам в виде дискретных
единиц. Сформулированные им закономерности наследования позже получили
название законов Менделя. При жизни Менделя работы были малоизвестны и
воспринимались критически.
Грегор Мендель
Единственная сохранившееся страничка с расчётами
из записной книжки Г. Менделя
5.
Карл Вильгельм фон НегелиНегели был разносторонне и широко
образованный
исследователь;
его
заслуги велики — и не только в ботанике,
но вообще в биологии. Он открыл и
описал целый ряд водорослей, изучил их
развитие
и
строение,
разработал
систематику этих растений; далее, он
открыл сперматозоиды у наземных и
водяных папоротников, работал по
физиологии грибов и бактерий. Ещё
важнее его исследования по анатомии в
физиологии клетки и органов.
Директор Ботанического института в
Мюнхене
6.
В начале XX века работы Менделя вновь привлекли внимание в связи сисследованиями Карла Корренса, Эриха фон Чермака и Гуго Де Фриза по
гибридизации растений, в которых были подтверждены основные выводы о
независимом наследовании признаков и о численных соотношениях при
«расщеплении» признаков в потомстве.
Исследователи, заложившие основы
современной генетики
Грегор Мендель
Карл Корренс
Гуго де Фриз
Эрих Чермак
7.
Основу исследований Г. Менделя составляли скрещивания и изучения гибридов садовогогороха. Г. Мендель начал исследования располагая 22 чистыми линиями садового гороха,
которые имели хорошо выраженные альтернативные различия между собой по семи парам
признаков.
Признаки растений исходных
линий
Фенотипическое выражение
признака
Форма семян
Круглые - шероховатые
Окраска семядолей
Жёлтые - зелёные
Окраска кожуры семян
Серая - белая
Форма бобов
Выполненные - морщинистые
Окраска незрелых бобов
Зелёная – жёлтая
Расположение цветков
Пазушные – верхушечные
Высота растений
Высокие – карликовые
8.
Основные подходы, позволившие Г. Менделюоткрыть законы наследования признаков
1. В отличие от своих предшественников,
изучавших наследование сразу нескольких
признаков, Г. Мендель в скрещивании
растений одной линии с растениями другой
линии
изучал
поведение
признаков
раздельно.
2. Учитывая строение цветка садового
гороха, Г. Мендель при скрещивании
растений исходных родительских линий
исключал самоопыление.
3. В поисках объяснения своих
результатов он использовал математику.
Анализируя полученные данные, Г. Мендель
формулировал рабочие гипотезы, проверяя
их
правильность
в
последующих
экспериментах.
9.
Аллели генов. Доминантность и рецессивностьАллели (аллеломорфы) — различные формы одного и того же гена,
расположенные в одинаковых участках (локусах) гомологичных хромосом;
определяют варианты проявления одного и того же признака. В диплоидном
организме может быть два одинаковых аллеля одного гена, в этом случае организм
называется гомозиготным, или два разных, что приводит к гетерозиготному
организму.
Доминантность (доминирование) — форма взаимоотношений между
аллелями одного гена, при которой один из них (доминантный) подавляет
(маскирует) проявление другого (рецессивного) и таким образом определяет
проявление признака как у доминантных гомозигот, так и у гетерозигот.
Гомозиготный организм по
доминантным генам
Фенотип
Гетерозиготный организм
Фенотип
Гомозиготный организм по
рецессивным генам
Фенотип
10.
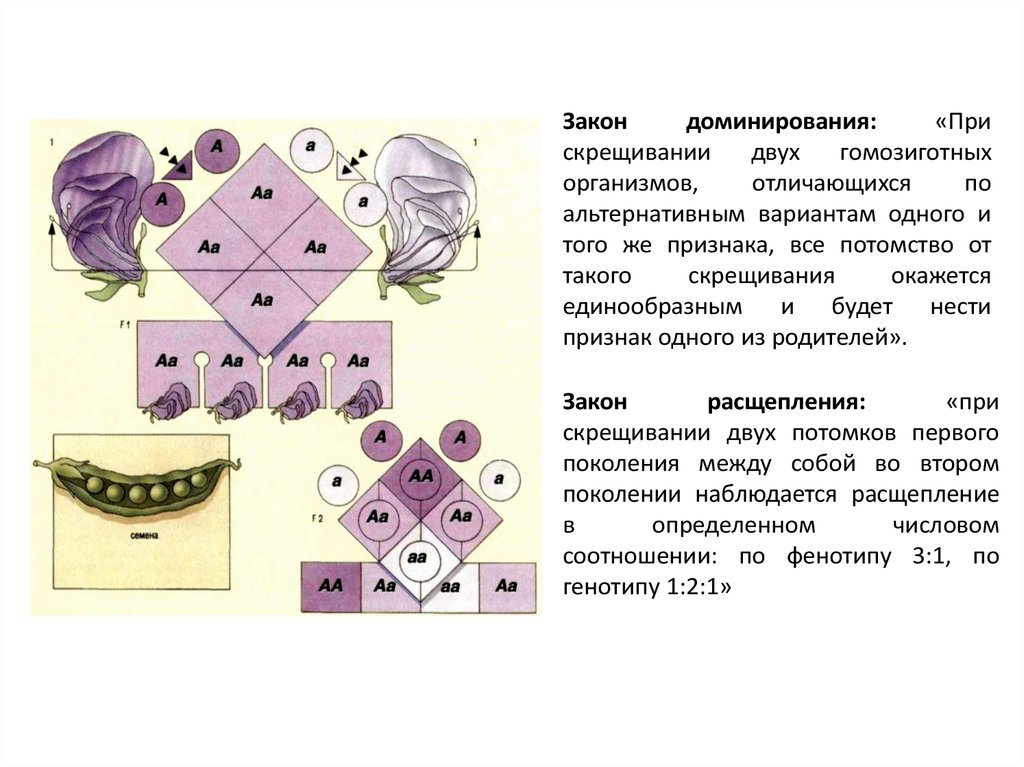
Закондоминирования:
«При
скрещивании
двух
гомозиготных
организмов,
отличающихся
по
альтернативным вариантам одного и
того же признака, все потомство от
такого
скрещивания
окажется
единообразным и будет нести
признак одного из родителей».
Закон
расщепления:
«при
скрещивании двух потомков первого
поколения между собой во втором
поколении наблюдается расщепление
в
определенном
числовом
соотношении: по фенотипу 3:1, по
генотипу 1:2:1»
11.
Неполноедоминирование.
При
неполном
доминировании
гетерозиготы
имеют
признаки,
промежуточные между признаками
рецессивной
и
доминантной
гомозигот.
Кодоминирование.
При
кодоминировании, в отличие от
неполного
доминирования,
у
гетерозигот признаки проявляются
одновременно (смешанно).
12.
Явления кодоминирования и неполного доминирования признаков слегкавидоизменяет первый закон Менделя: «Гибриды первого поколения от
скрещивания чистых линий особей с противоположными признаками всегда
одинаковы по этому признаку: проявляют доминирующий признак, если
признаки находятся в отношении доминирования, или смешанный
(промежуточный) признак, если они находятся в отношении
кодоминирования (неполного доминирования)».
13.
Закон независимого комбинирования: «при скрещивании двух гомозиготныхособей, отличающихся друг от друга по двум и более парам альтернативных
признаков, гены и соответствующие им признаки наследуются независимо друг от
друга и комбинируются во всех возможных сочетаниях».
14.
Независимое комбинирование признаков (третий закон Менделя) осуществляется приусловии, что гены, определяющие эти признаки, находятся в разных парах гомологичных
хромосом. Следовательно, у каждого организма число генов, способных независимо
комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов
значительно превышает количество хромосом. Это дало основание предположить, что в каждой
хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме,
образуют группу сцепления и наследуются вместе (закон сцепленного наследования Т.
Моргана).
Группы сцепления на карте 12
хромосомы свиньи. Красный
цвет – картированные гены,
черный – микросателлитные
последовательности.
15.
Способ наследования сцепленных генов отличается от наследования генов,локализованных в разных парах гомологичных хромосом. Так, если при независимом
комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и
ab) в равных количествах, то при сцепленном наследовании (в отсутствие
кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab)
тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме
родителя.
16.
Генетика полаXY
XO
X1X2…XY
X1X2…XO
XY1Y2…Y
Гаметы
Механизмы определения пола у разных
животных
Определение пола у человека: женщины - гомогаметны,
мужчины - гетерогаметны
Число групп сцепления соответствует гаплоидному числу хромосом, поскольку
группу сцепления составляют две гомологичные хромосомы, в которых локализованы
одинаковые гены. (У особей гетерогаметного пола, например, у самцов
млекопитающих, групп сцепления на самом деле на одну больше, так как X- и Yхромосомы содержат разные гены и представляют собой две разные группы
сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24.
17.
Наследование признаков сцепленных с поломУ человека носителем гена гемофилии являются женщины, но болеют только
мужчины потому что они имеют только одну Х-хромосому и, следовательно, при
получении дефектного гена, не имеют нормальной аллели. У кошек ген В кодирует
жёлтую окраску, а ген b – чёрную, сочетание Bb – черепаховую окраску. При
скрещивании жёлтой кошки и черного кота все кошки будут иметь черепаховую
окраску, а коты чёрную, т.к. ген В не может быть сцеплен с Y-хромосомой.
18.
КроссинговерКроссинго́ вер (другое название в биологии перекрёст) — явление обмена
участками гомологичных хромосом во время конъюгации при мейозе. Помимо
мейотического описан также митотический кроссинговер.
Поскольку кроссинговер вносит возмущения
в картину сцепленного наследования, его
удалось использовать для картирования «групп
сцепления»
(хромосом).
Возможность
картирования была основана на предположении
о том, что, чем чаще наблюдается кроссинговер
между двумя генами, тем дальше друг от друга
расположены эти гены в группе сцепления и тем
чаще будут наблюдаться отклонения от
сцепленного наследования. Первые карты
хромосом были построены в 1913 г. для
классического экспериментального объекта
плодовой мушки Drosophila melanogaster
Альфредом
Стёртевантом,
учеником
и
сотрудником Томаса Ханта Моргана.
Группа сцепления
19.
Линейный порядок и мозаичность генов.Современная концепция гена
Гены на хромосомах располагаются в линейном порядке, причём, чем больше
расстояние между генными локусами, тем большей является частота кроссинговера
между ними. У эукариот сегменты ДНК, соответствующие индивидуальным генам,
разделены не кодирующими последовательностями – интронами, что определяет
«мозаичность» генов. Во многих генах обнаружено по нескольку интронов.
1. Ген является крайней единицей структуры – его нельзя разделить на
субъединицы рекомбинацией или разрывом хромосом.
2. Ген является крайней единицей функции – кодирует только одну функцию,
которую нельзя разделить на субфункции.
Ген глобина
ДНК
мРНК
Построение генетической карты для генов A,
B, C, частоты рекомбинаций между которыми
составляют A – B = 6 %, B – C = 14 %, A – C = 8 %
Мозаичность гена
20.
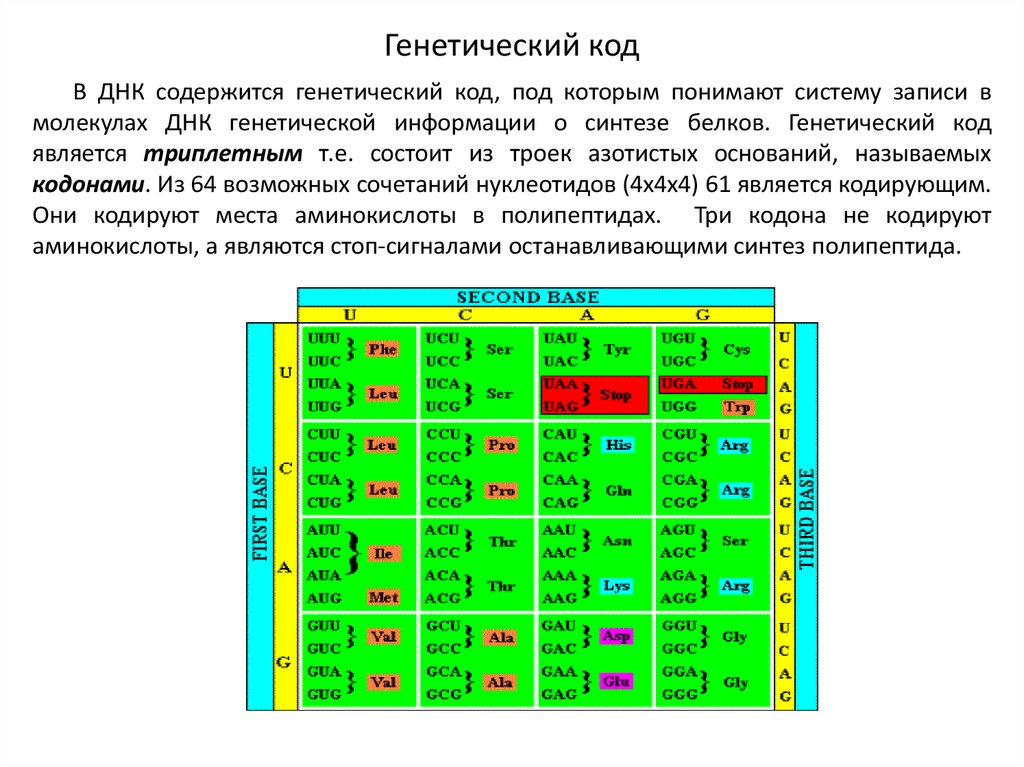
Генетический кодВ ДНК содержится генетический код, под которым понимают систему записи в
молекулах ДНК генетической информации о синтезе белков. Генетический код
является триплетным т.е. состоит из троек азотистых оснований, называемых
кодонами. Из 64 возможных сочетаний нуклеотидов (4х4х4) 61 является кодирующим.
Они кодируют места аминокислоты в полипептидах. Три кодона не кодируют
аминокислоты, а являются стоп-сигналами останавливающими синтез полипептида.
21.
Свойства генетического кода1. Триплетность. Значащей единицей является сочетание трёх нуклеотидов.
2. Непрерывность — между триплетами нет знаков препинания, то есть
информация считывается непрерывно
3. Однозначность (специфичность) — определённый кодон соответствует только
одной аминокислоте
4. Вырожденность (избыточность) — одной и той же аминокислоте может
соответствовать несколько кодонов
5. Универсальность — генетический код работает одинаково в организмах разного
уровня сложности — от вирусов до человека
6. Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене
класса кодируемой аминокислоты, называют консервативными; мутации замен
нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют
радикальными
7. Знаки препинания — триплеты выполняют функцию знаков препинания
22.
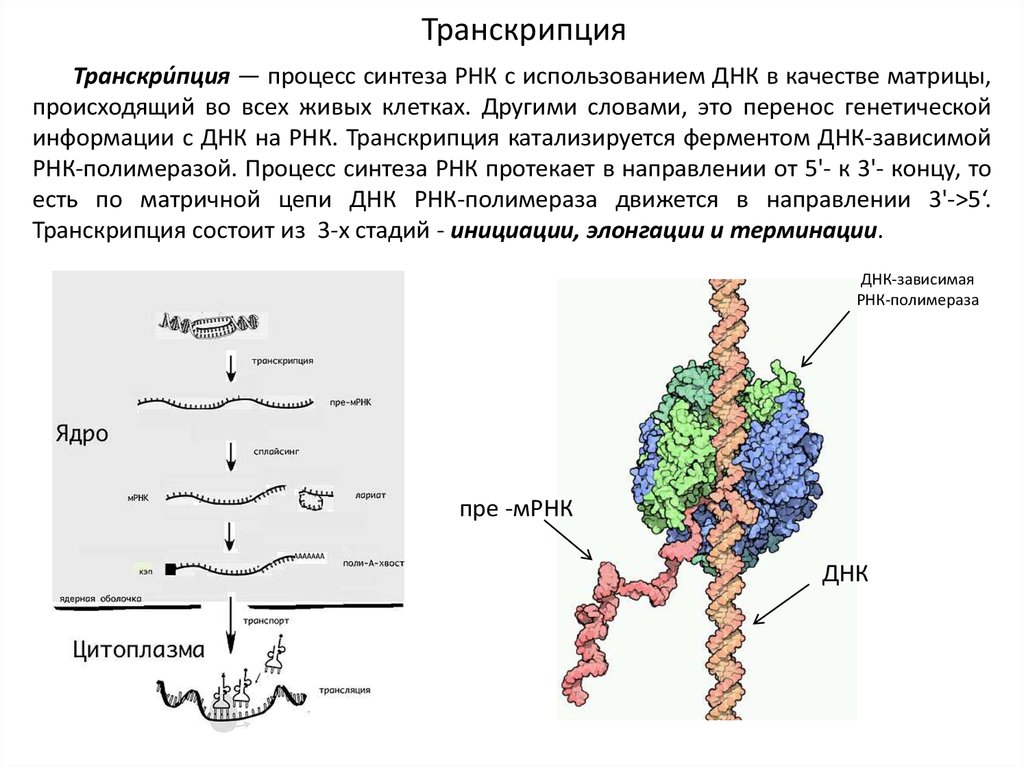
ТранскрипцияТранскри́пция — процесс синтеза РНК с использованием ДНК в качестве матрицы,
происходящий во всех живых клетках. Другими словами, это перенос генетической
информации с ДНК на РНК. Транскрипция катализируется ферментом ДНК-зависимой
РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5'- к 3'- концу, то
есть по матричной цепи ДНК РНК-полимераза движется в направлении 3'->5‘.
Транскрипция состоит из 3-х стадий - инициации, элонгации и терминации.
ДНК-зависимая
РНК-полимераза
пре -мРНК
ДНК
23.
Синтез мРНКСинтез молекул мРНК происходит в ядре клетки. В качестве матрицы для
копирования цепи используется лишь одна нить ДНК. Копирование мРНК может
начинаться с любого участка ДНК и осуществляется ферментом РНК – полимеразой.
5’
3’
ГГГГГГГГГГГГГ ГГГГГГГГГ
3’
5’
ЦЦЦЦЦЦЦЦЦЦЦЦЦЦЦЦЦ
5’
3’
ГГГГГГГГГГГГГ ГГГГГГГГГ
ЦЦЦЦЦЦЦЦЦЦЦЦЦЦЦЦЦ
3’
5’
ЦЦЦЦЦЦЦЦЦЦ
Транскрипт мРНК
ГГГГГГГГГГГГГ
Транскрипт мРНК
1
2
1. При движении РНК-полимеразы слева на право матрицей для синтеза мРНК
служит верхняя цепь.
2. При движении РНК-полимеразы справа на лево матрицей для синтеза служит
нижняя цепь.
24.
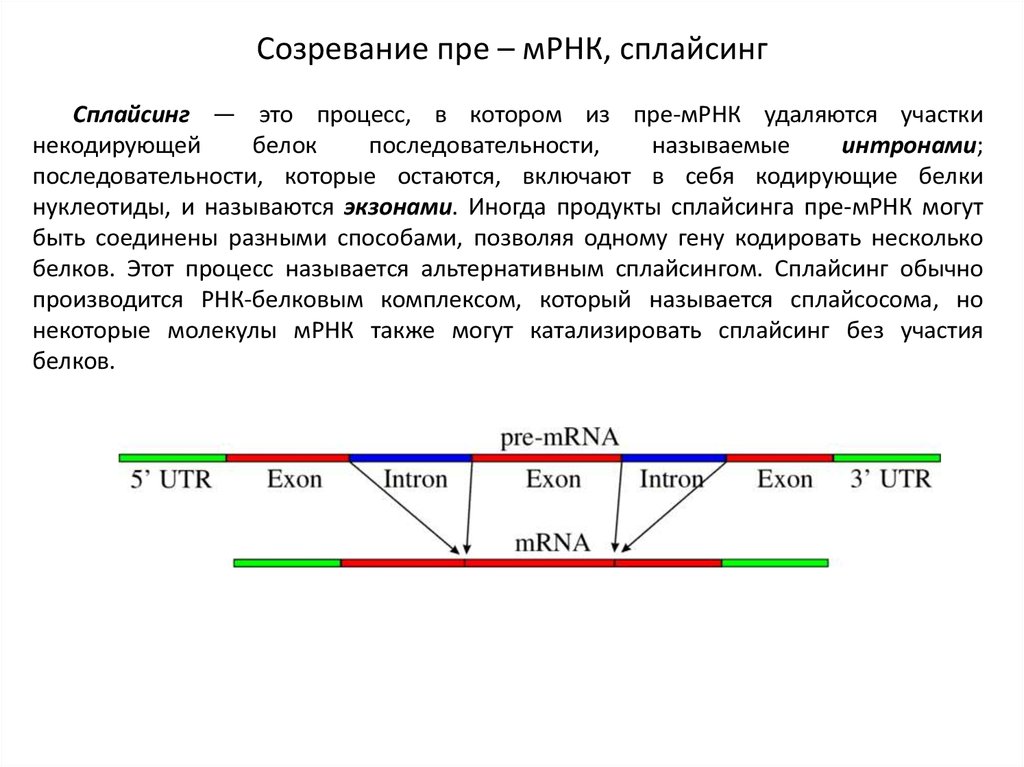
Созревание пре – мРНК, сплайсингСплайсинг — это процесс, в котором из пре-мРНК удаляются участки
некодирующей
белок
последовательности,
называемые
интронами;
последовательности, которые остаются, включают в себя кодирующие белки
нуклеотиды, и называются экзонами. Иногда продукты сплайсинга пре-мРНК могут
быть соединены разными способами, позволяя одному гену кодировать несколько
белков. Этот процесс называется альтернативным сплайсингом. Сплайсинг обычно
производится РНК-белковым комплексом, который называется сплайсосома, но
некоторые молекулы мРНК также могут катализировать сплайсинг без участия
белков.
25.
Строение зрелой мРНКЗрелая мРНК состоит из нескольких участков, различающихся по функциям: «5'
кэп», 5' нетранслируемая область, кодирующая (транслируемая) область, 3'
нетранслируемая область и 3' полиадениновый «хвост».
5' кэп (от англ. cap — шапочка) — это модифицированный гуанидиновый нуклеотид,
который добавляется на 5' (передний) конец незрелой мРНК. Эта модификация очень важна
для узнавания мРНК при инициации трансляции, а также для защиты от 5’нуклеаз —
ферментов, разрушающих цепи нуклеиновых кислот с незащищённым 5'-концом.
Нетранслируемые области — участки РНК, расположенные до старт-кодона и после стопкодона, которые не кодируют белок. Нетранслируемые области имеют несколько функций в
жизненном цикле мРНК, включая регуляцию стабильности мРНК, локализации мРНК и
эффективности трансляции.
Полиадениновый хвост. У высших эукариот поли-А-хвост добавляется к
транскрибированной РНК, которая содержит специфическую последовательность, AAUAAA. По
прошествии некоторого времени, определяемого длиной полиаденинового участка на 3' конце
молекулы, мРНК разрушается на составляющие её нуклеотиды с участием РНКаз.
26.
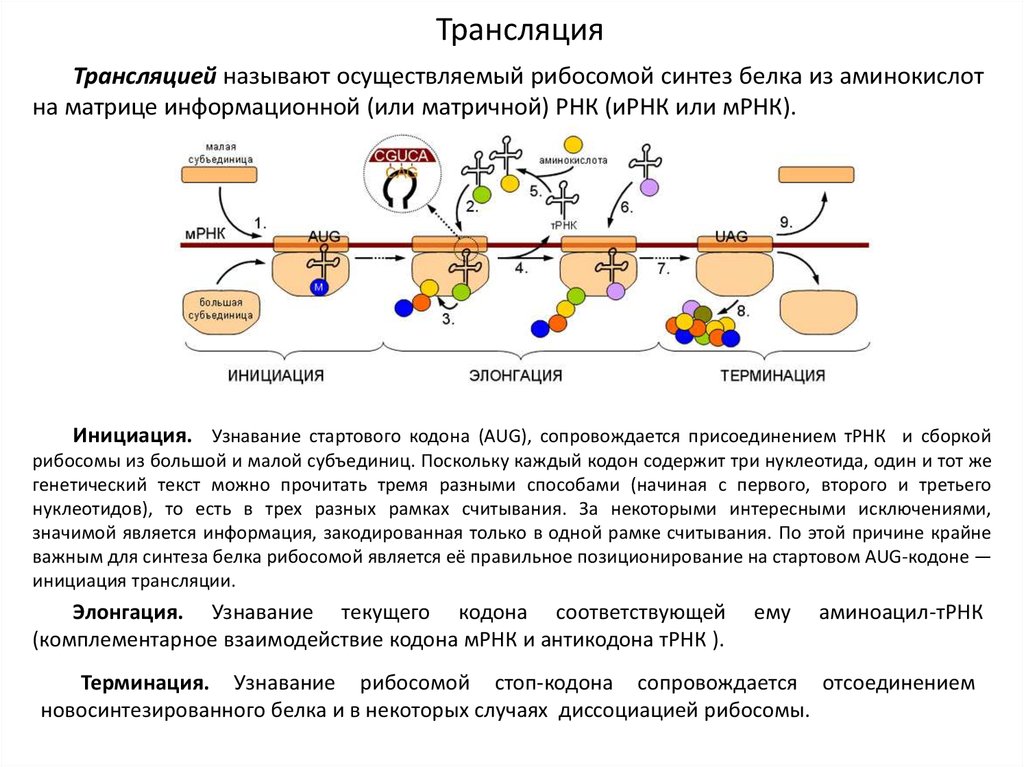
ТрансляцияТрансляцией называют осуществляемый рибосомой синтез белка из аминокислот
на матрице информационной (или матричной) РНК (иРНК или мРНК).
Инициация. Узнавание стартового кодона (AUG), сопровождается присоединением тРНК и сборкой
рибосомы из большой и малой субъединиц. Поскольку каждый кодон содержит три нуклеотида, один и тот же
генетический текст можно прочитать тремя разными способами (начиная с первого, второго и третьего
нуклеотидов), то есть в трех разных рамках считывания. За некоторыми интересными исключениями,
значимой является информация, закодированная только в одной рамке считывания. По этой причине крайне
важным для синтеза белка рибосомой является её правильное позиционирование на стартовом AUG-кодоне —
инициация трансляции.
Элонгация. Узнавание текущего кодона соответствующей
(комплементарное взаимодействие кодона мРНК и антикодона тРНК ).
ему
аминоацил-тРНК
Терминация. Узнавание рибосомой стоп-кодона сопровождается отсоединением
новосинтезированного белка и в некоторых случаях диссоциацией рибосомы.
27.
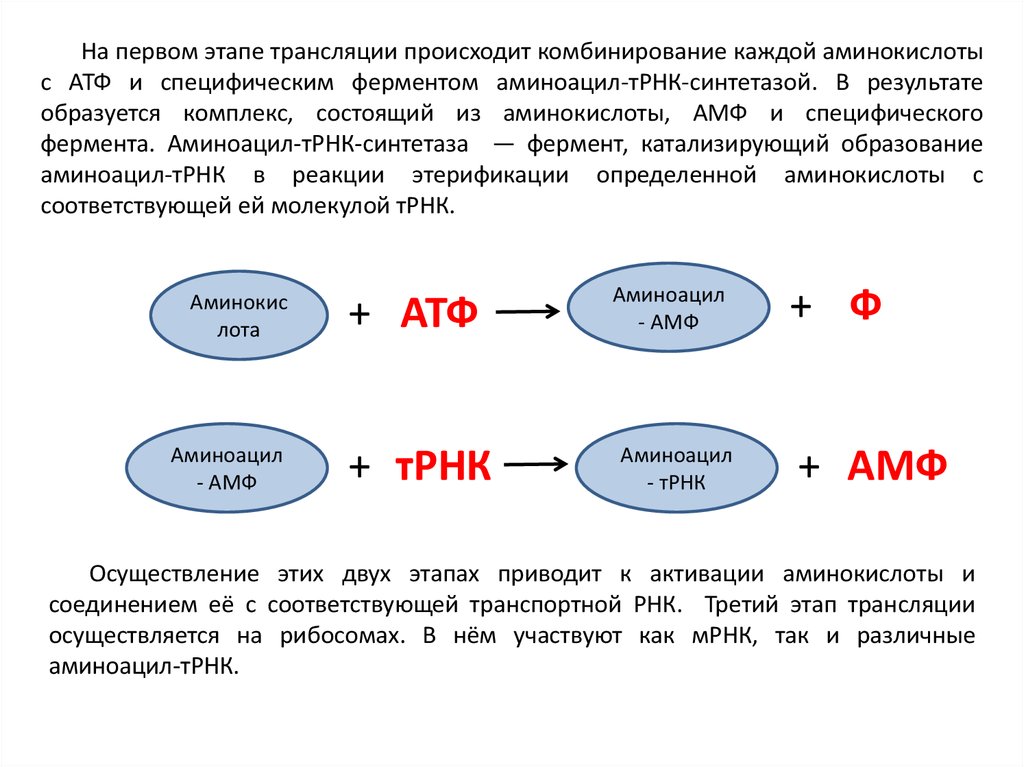
На первом этапе трансляции происходит комбинирование каждой аминокислотыс АТФ и специфическим ферментом аминоацил-тРНК-синтетазой. В результате
образуется комплекс, состоящий из аминокислоты, АМФ и специфического
фермента. Аминоацил-тРНК-синтетаза — фермент, катализирующий образование
аминоацил-тРНК в реакции этерификации определенной аминокислоты с
соответствующей ей молекулой тРНК.
Аминокис
лота
Аминоацил
- АМФ
+ АТФ
+ тРНК
Аминоацил
- АМФ
Аминоацил
- тРНК
+ Ф
+ АМФ
Осуществление этих двух этапах приводит к активации аминокислоты и
соединением её с соответствующей транспортной РНК. Третий этап трансляции
осуществляется на рибосомах. В нём участвуют как мРНК, так и различные
аминоацил-тРНК.
28.
Транспортная РНКФункцией тРНК является транспортировка аминокислот к месту синтеза белка. тРНК также
принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь —
будучи в комплексе с аминокислотой — к кодону мРНК и обеспечивая необходимую для
образования новой пептидной связи конформацию комплекса. Для каждой аминокислоты
существует своя тРНК.
тРНК является одноцепочечной РНК, однако в функциональной форме имеет конформацию
«листа клевера» или «кловерлиф» (англ. cloverleaf). Аминокислота ковалентно присоединяется к
3'-концу молекулы с помощью специфичного для каждого типа тРНК фермента аминоацил-тРНКсинтетазы. На участке «C» находится антикодон, соответствующий аминокислоте.
29.
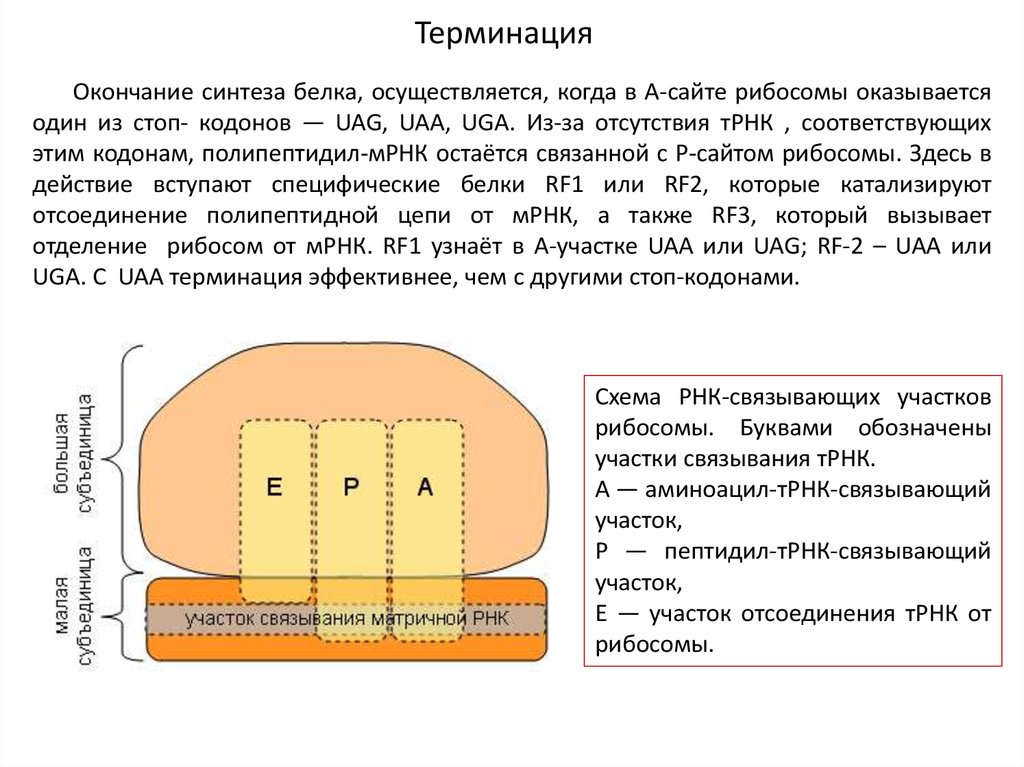
ТерминацияОкончание синтеза белка, осуществляется, когда в А-сайте рибосомы оказывается
один из стоп- кодонов — UAG, UAA, UGA. Из-за отсутствия тРНК , соответствующих
этим кодонам, полипептидил-мРНК остаётся связанной с Р-сайтом рибосомы. Здесь в
действие вступают специфические белки RF1 или RF2, которые катализируют
отсоединение полипептидной цепи от мРНК, а также RF3, который вызывает
отделение рибосом от мРНК. RF1 узнаёт в А-участке UAA или UAG; RF-2 – UAA или
UGA. С UAA терминация эффективнее, чем с другими стоп-кодонами.
Схема РНК-связывающих участков
рибосомы. Буквами обозначены
участки связывания тРНК.
А — аминоацил-тРНК-связывающий
участок,
Р — пептидил-тРНК-связывающий
участок,
Е — участок отсоединения тРНК от
рибосомы.
30.
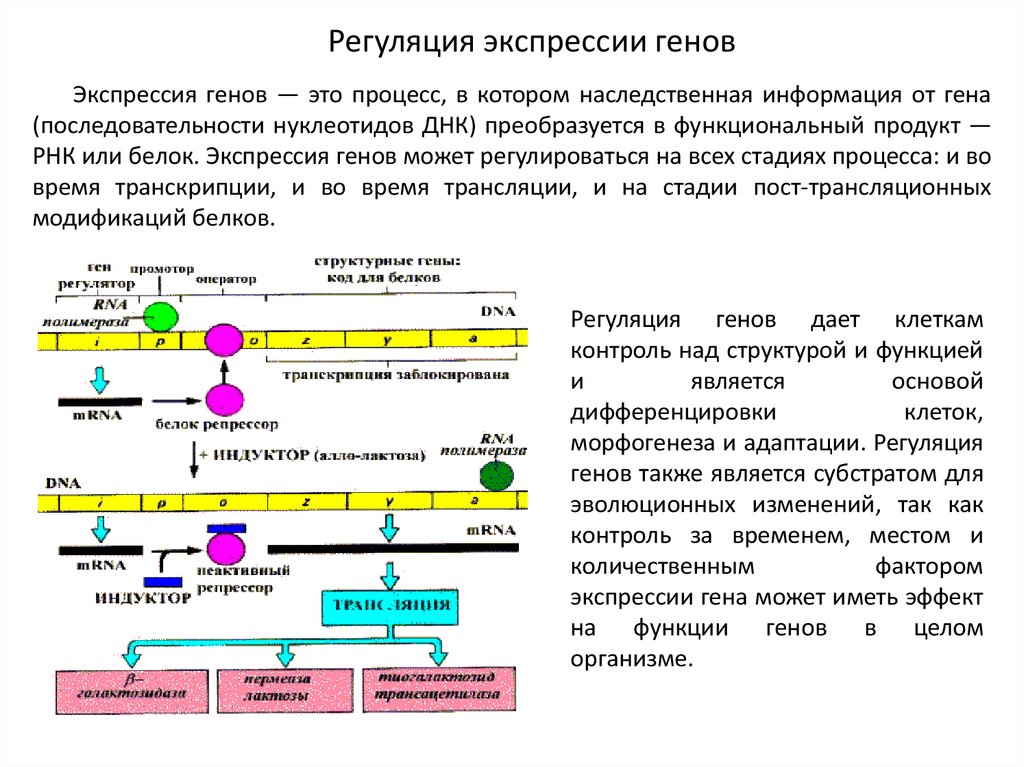
Регуляция экспрессии геновЭкспрессия генов — это процесс, в котором наследственная информация от гена
(последовательности нуклеотидов ДНК) преобразуется в функциональный продукт —
РНК или белок. Экспрессия генов может регулироваться на всех стадиях процесса: и во
время транскрипции, и во время трансляции, и на стадии пост-трансляционных
модификаций белков.
Регуляция генов дает клеткам
контроль над структурой и функцией
и
является
основой
дифференцировки
клеток,
морфогенеза и адаптации. Регуляция
генов также является субстратом для
эволюционных изменений, так как
контроль за временем, местом и
количественным
фактором
экспрессии гена может иметь эффект
на функции генов в целом
организме.
31.
Общие выводы1. В 1865 году, проводя опыты по скрещиванию растений гороха, Г. Мендель показал, что
наследственные задатки не смешиваются, а передаются от родителей к потомкам в виде
дискретных единиц. Сформулированные им закономерности наследования позже получили
название законов Менделя.
2. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе
(закон сцепленного наследования Т. Моргана). Число групп сцепления соответствует
гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные
хромосомы, в которых локализованы одинаковые гены.
3. Кроссинго́ вер— явление обмена участками гомологичных хромосом во время конъюгации
при мейозе.
4. Ген является крайней единицей структуры – его нельзя разделить на субъединицы
рекомбинацией или разрывом хромосом. Ген является крайней единицей функции – кодирует
только одну функцию, которую нельзя разделить на субфункции.
5. Генетический код является триплетным т.е. состоит из троек азотистых оснований,
называемых кодонами.
Свойства генетического кода: неперекрываемость, линейность,
вырожденность.
6. Транскрипция-процесс переписывания генетической информации с ДНК на РНК.
7. Трансляция – перевод генетической информации с последовательности нуклеотидов в
последовательность аминокислот.