



























 biology
biologySimilar presentations:






")



Основні проблеми макроеволюції
1.
Макроеволюція2.
Морськілілії
3.
Основні проблеми макроеволюціїЗакономірності
еволюції груп (таксонів)
Типогенез (формування
таксонів вищіх рангів)
Адаптаціогенез
Закономірності
еволюції онтогенезу
Прогрес і еволюція
Еволюція біогеоценозів
Великі вимирання
Спрямованість
еволюційного розвитку
та паралельна еволюція
Еволюція механізмів
еволюції
Темпи еволюційних
перетворень
Систематика і еволюція
4.
Закономірності еволюції груп•Правило незворотності еволюції (закон Долло)
•Правило походження нових груп від
неспеціалізованих предків(закон Копа)
•Правило прогресуючої спеціалізації (закон
Депере)
•Правило збільшення росту у філогенетичних
гілках (закон Копа-Депере)
•Правило споріднених відхилень (закон Кренке)
•Закон росту середньої тривалості існування
родів у процесі еволюції
5.
Правилонезворотності
еволюції
Форма тіла
швидкоплаваючих тварин
набуває
конвергентної
схожості,
проте
повернення до
предкових
форм у
вторинноводн
их істот не
відбувається
6.
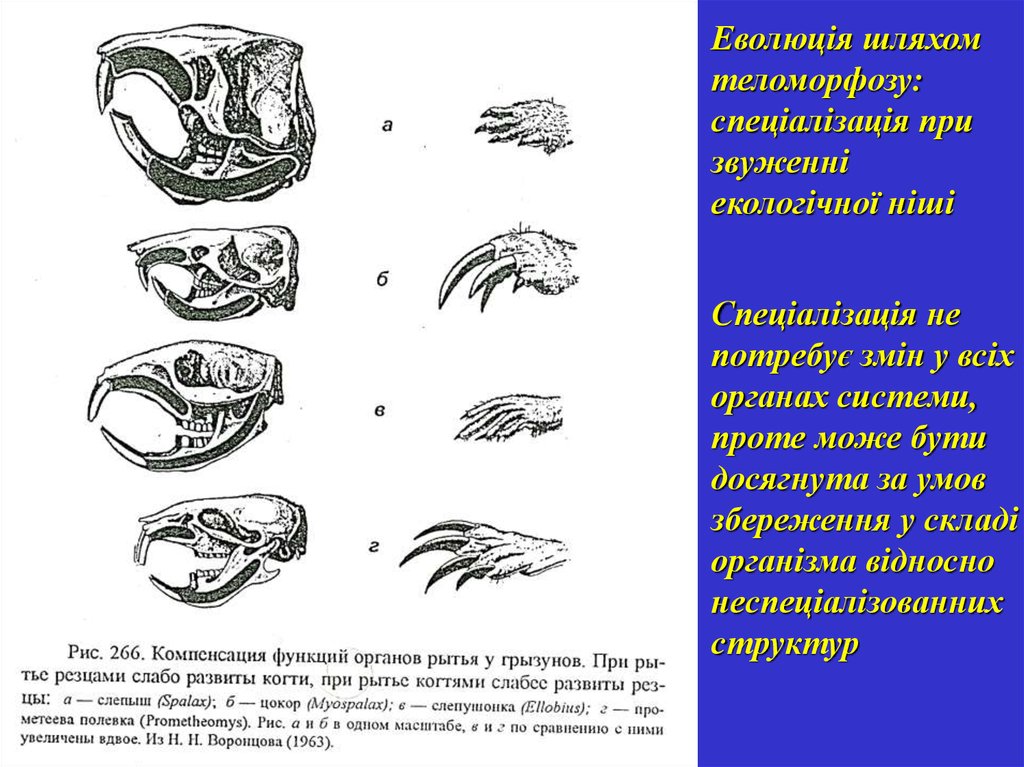
Еволюція шляхомтеломорфозу:
спеціалізація при
звуженні
екологічної ніші
Спеціалізація не
потребує змін у всіх
органах системи,
проте може бути
досягнута за умов
збереження у складі
організма відносно
неспеціалізованних
структур
7.
Еволюція шляхом гіперморфозу: Закон Копа-ДепереПредком могутнього плейстоценового південного слона був
невеличкий ссавець мерітерій.
8.
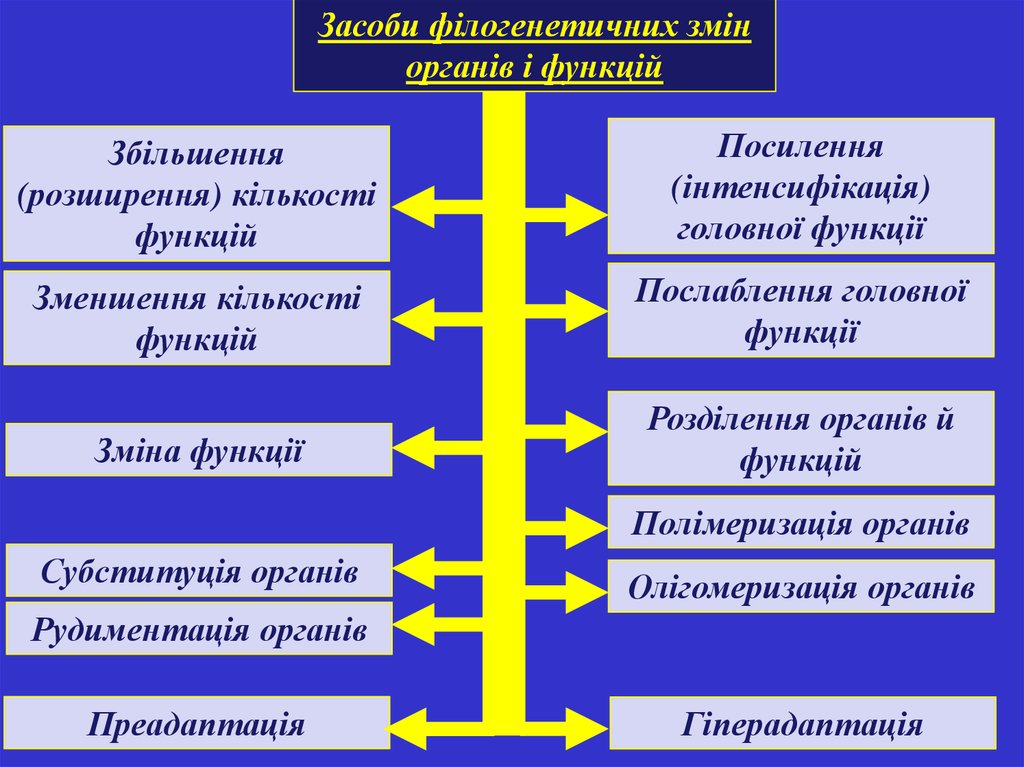
Засоби філогенетичних змінорганів і функцій
Збільшення
(розширення) кількості
функцій
Посилення
(інтенсифікація)
головної функції
Зменшення кількості
функцій
Послаблення головної
функції
Зміна функції
Розділення органів й
функцій
Полімеризація органів
Субституція органів
Олігомеризація органів
Рудиментація органів
Преадаптація
Гіперадаптація
9.
Гетеробатмія (А.Л. Тахтаджян, 1954) – це неоднаковийеволюційний рівень розвитку різних органів, що виникає
внаслідок неоднакового темпу еволюції різних частин
організму.
Компенсація функції. Недостатній рівень виконання тієї чи
іншої функції внаслідок недостатньої швидкості еволюційних
перетворень відповідних органів (або послаблення функції
внаслідок редукції якогось органа) може бути компенсований
прискореним темпом еволюції інших органів чи структур
організму.
10.
Форми еволюційних змінонтогенезу
Перебудови
кореляційних
взаємозв’язків
Автономізація
онтогенеза
Ембріонізація
онтогенеза
Неотенізація
Філембріогенези
Анаболія
Архалаксис
Девіація
11.
Біогенетичнийзакон та закон
зародкової
подібності
12.
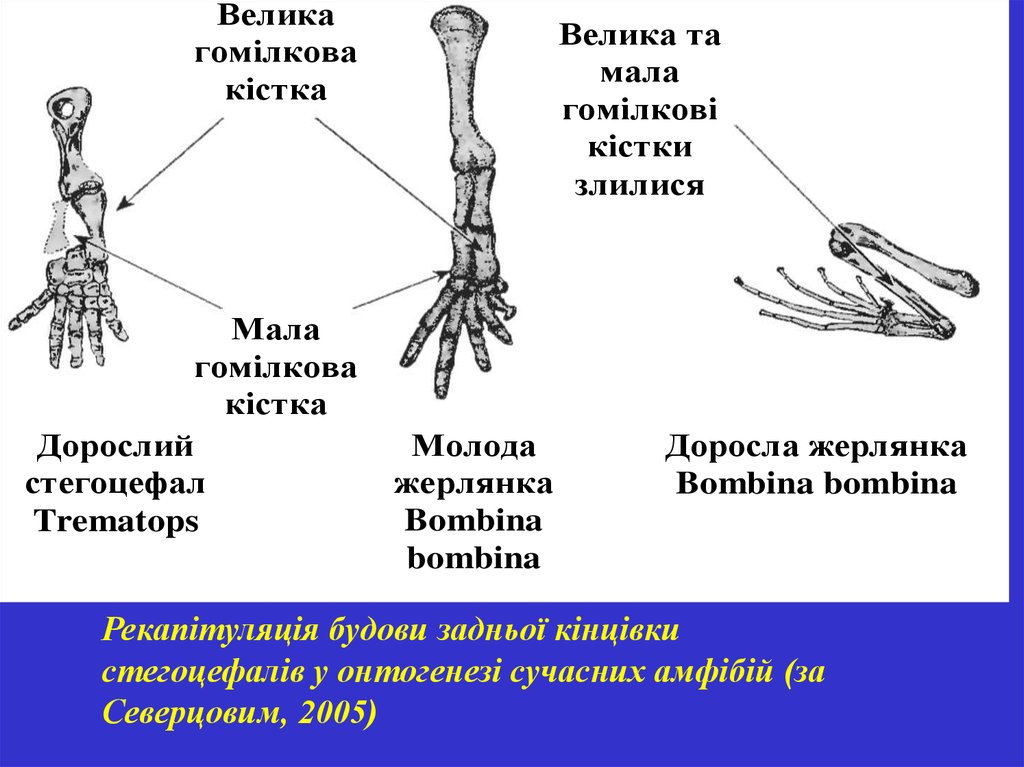
Великагомілкова
кістка
Мала
гомілкова
кістка
Дорослий
стегоцефал
Trematops
Велика та
мала
гомілкові
кістки
злилися
Молода
жерлянка
Bombina
bombina
Доросла жерлянка
Bombina bombina
Рекапітуляція будови задньої кінцівки
стегоцефалів у онтогенезі сучасних амфібій (за
Северцовим, 2005)
13.
Листкова жабаПаразитичні оси
14.
анаболіяN
анаболія H
G
F
G1 G2 G3
анаболія
H1 H 2
G
4
F1 F2
H3
E E1 E
F3
G5
2
F
4
G6
E3
G7
E4
E
5
F5
F6
D D1
F7
D2
D3
E6
C
анаболія
девіація
C1
C2
ембріонізація
B
A
B1
ембріонізація
D4
С3
D4
І
E6
E7
ембріоадаптація
архалаксис
Z
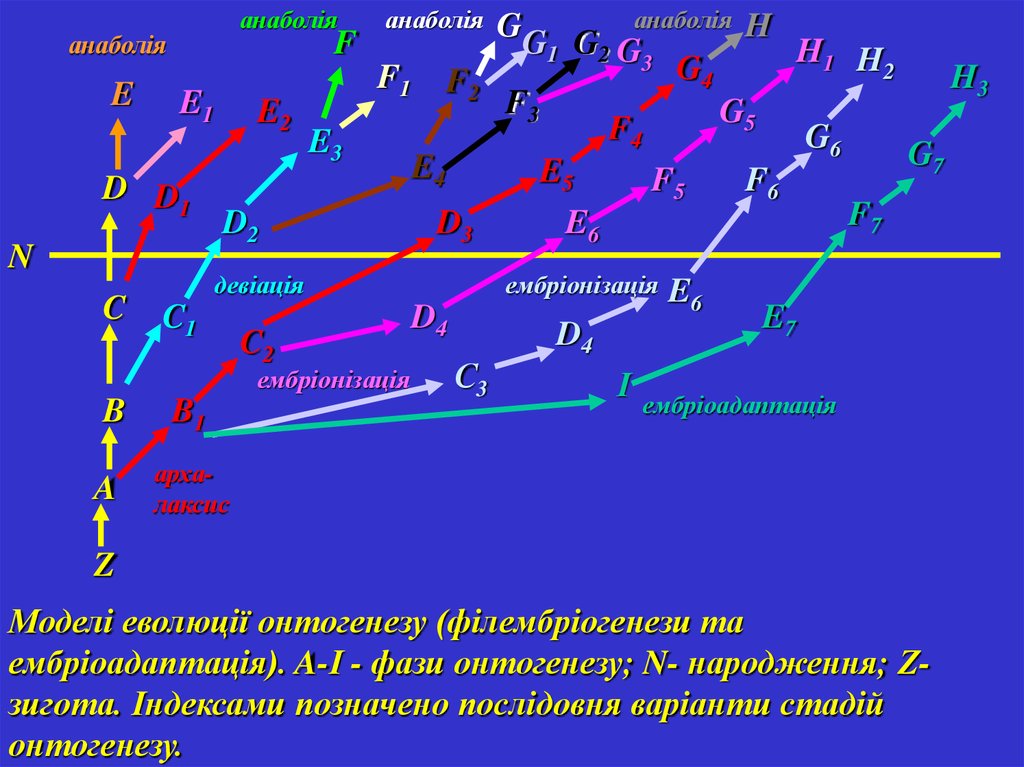
Моделі еволюції онтогенезу (філембріогенези та
ембріоадаптація). A-І - фази онтогенезу; N- народження; Zзигота. Індексами позначено послідовня варіанти стадій
онтогенезу.
15.
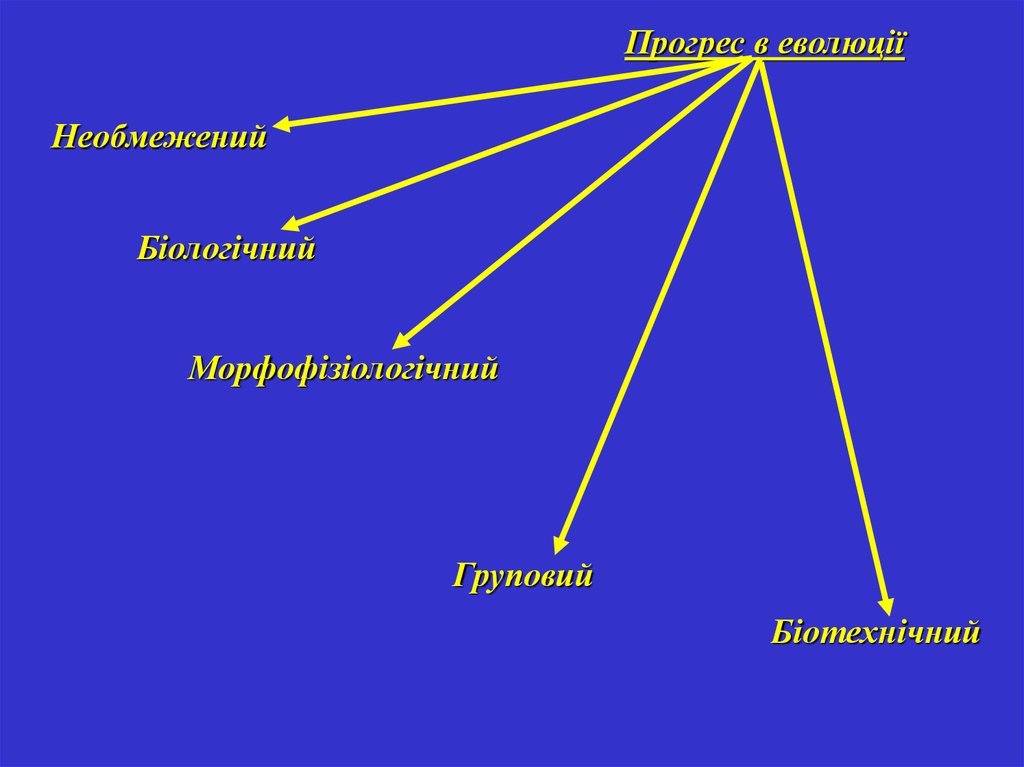
Прогрес в еволюціїНеобмежений
Біологічний
Морфофізіологічний
Груповий
Біотехнічний
16.
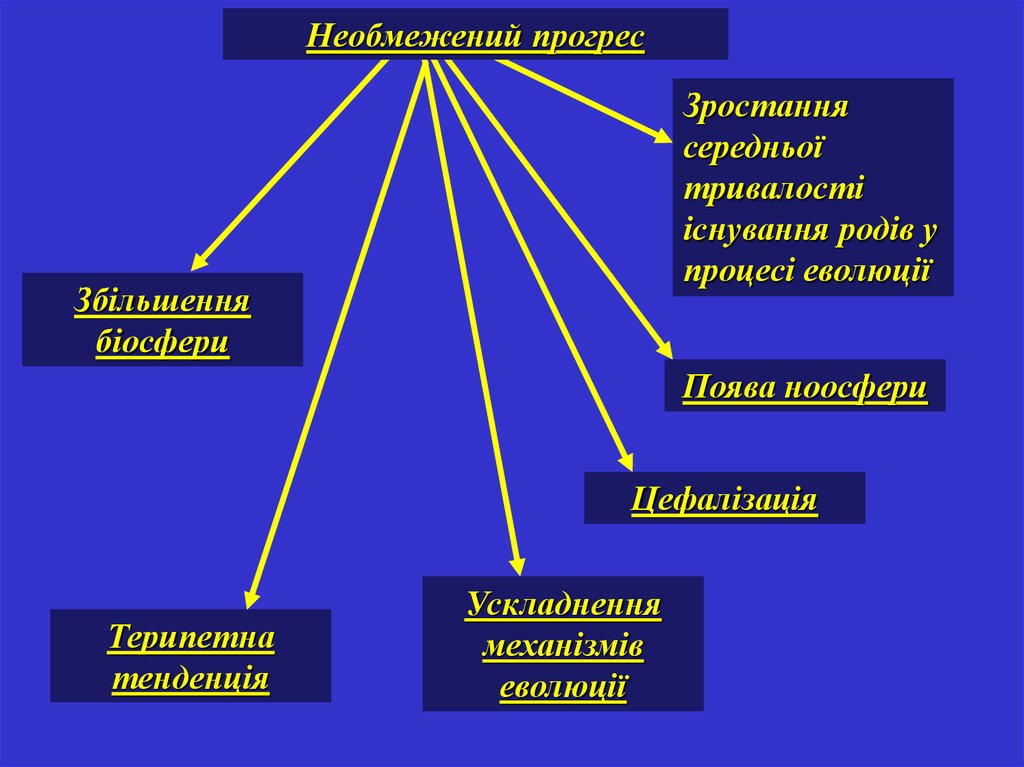
Необмежений прогресЗростання
середньої
тривалості
існування родів у
процесі еволюції
Збільшення
біосфери
Поява ноосфери
Цефалізація
Терипетна
тенденція
Ускладнення
механізмів
еволюції
17.
Зростаннясвітимості
Сонця +30%
4,0млрд.р. до
сучасності
Зростання
об’єму світового
океану (4,00,5млрд.р.)
Террестризація
Зростання
кількості
атмосферного
кисню (0,70,3млрд.р.)
Зростання кількості енергії, доступної екосистемам
Збільшення біомаси біосфери
Ускладнення екосистем
Ускладнення біосфери
Терипетна
тенденція
Ускладнення оточуючого середовища
Ускладнення організмів
Зростання стійкості біол.систем,
середньої тривалості існування родів
Цефалізація
Поява ноосфери
18.
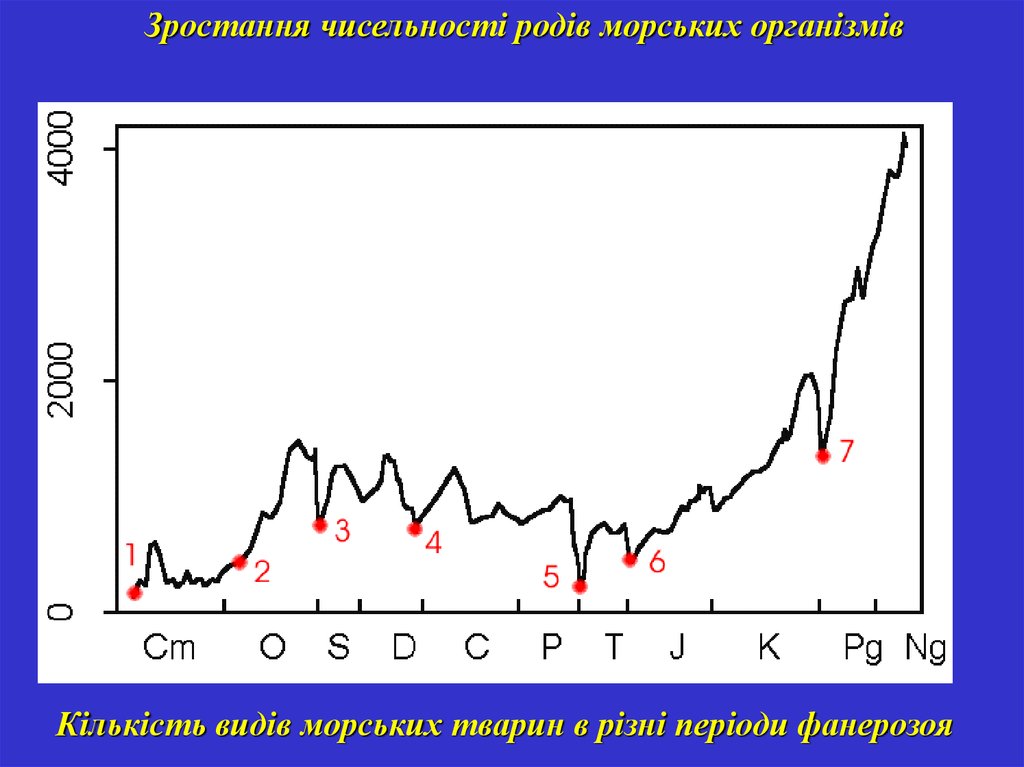
Зростання чисельності родів морських організмівКількість видів морських тварин в різні періоди фанерозоя
19.
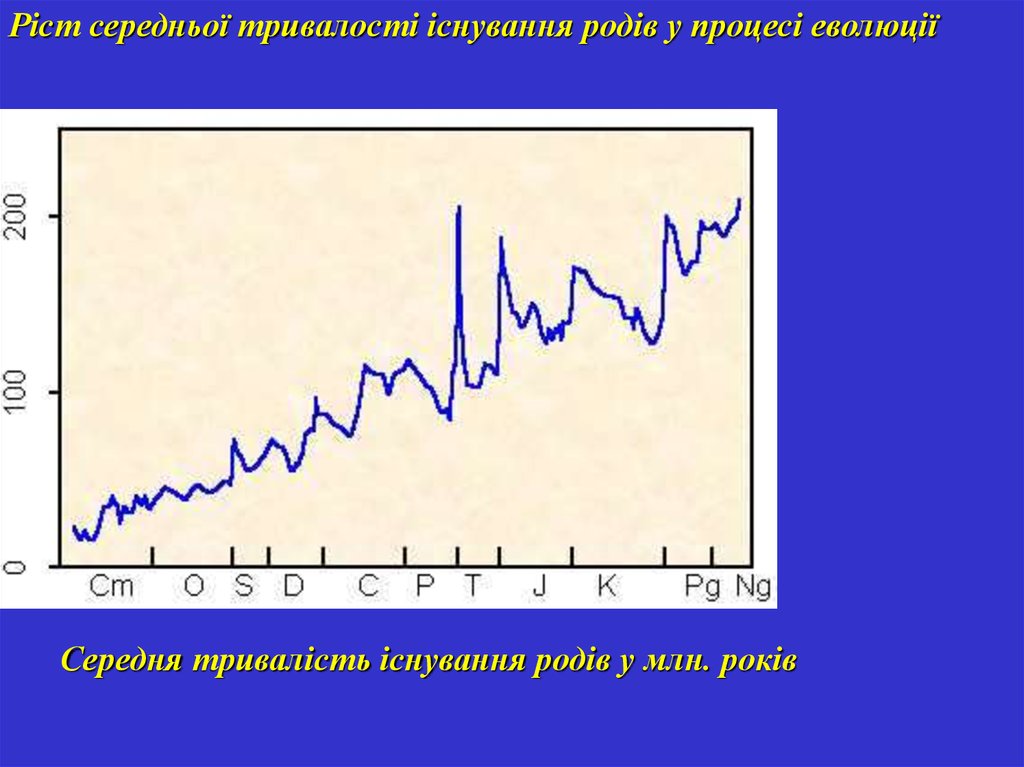
Ріст середньої тривалості існування родів у процесі еволюціїСередня тривалість існування родів у млн. років
20.
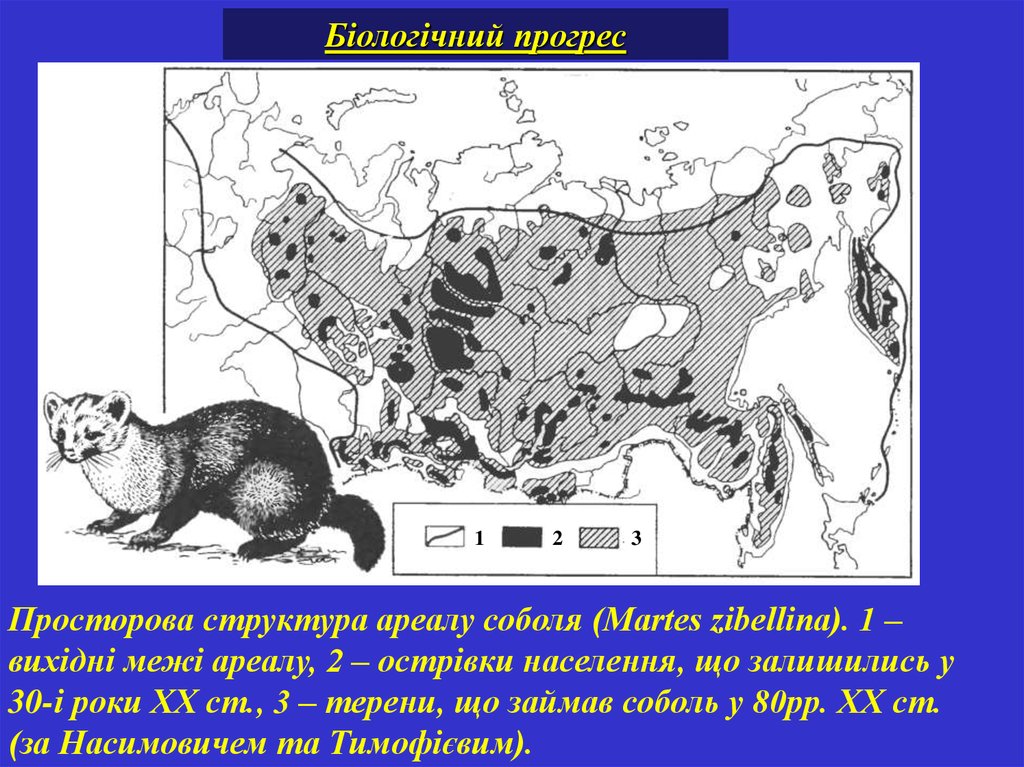
Біологічний прогрес1
2
3
Просторова структура ареалу соболя (Martes zibellina). 1 –
вихідні межі ареалу, 2 – острівки населення, що залишились у
30-і роки ХХ ст., 3 – терени, що займав соболь у 80рр. ХХ ст.
(за Насимовичем та Тимофієвим).
21.
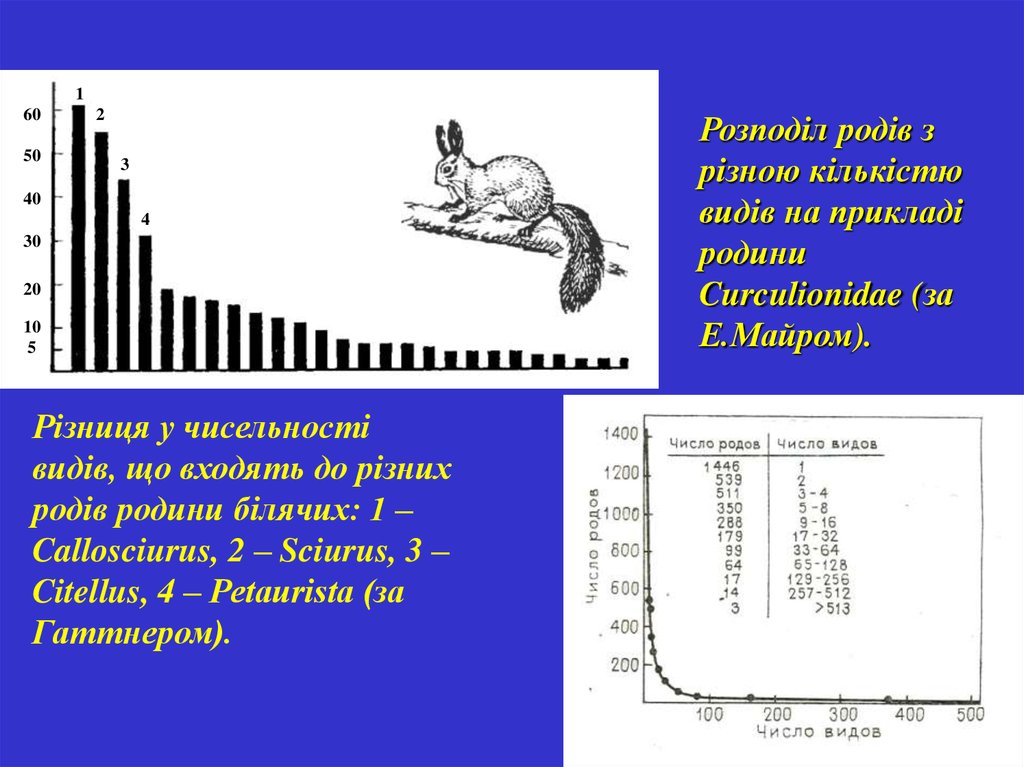
160
50
2
3
40
4
30
20
10
5
Різниця у чисельності
видів, що входять до різних
родів родини білячих: 1 –
Callosciurus, 2 – Sciurus, 3 –
Citellus, 4 – Petaurista (за
Гаттнером).
Розподіл родів з
різною кількістю
видів на прикладі
родини
Curculionidae (за
Е.Майром).
22.
Зростання ступеня різноманіття в межахпевного таксону при захопленні їм нової
адаптивної зони отримало назву
адаптивної радіації.
23.
Морфофізіологічний прогресАроморфоз - еволюційні перебудови організмів, що ведуть
до морфофізіологічного прогресу.
Катаморфоз - регресивні зміни організмів.
Епектоморфози та ідіоадаптації - адаптації, які не
призводять до суттєвих змін рівня організації. При цьому
епектоморфози значно збільшують адаптивні
можливості даної групи.
Форми еволюційного процеса, в результаті яких
утворюються аро-, ката-, епектоморфози та
ідіоадаптації отримали назву аро-, ката-, епекто- та
алогенезів відповідно.
24.
Співвідношення між основнимиформами еволюційного процеса
25.
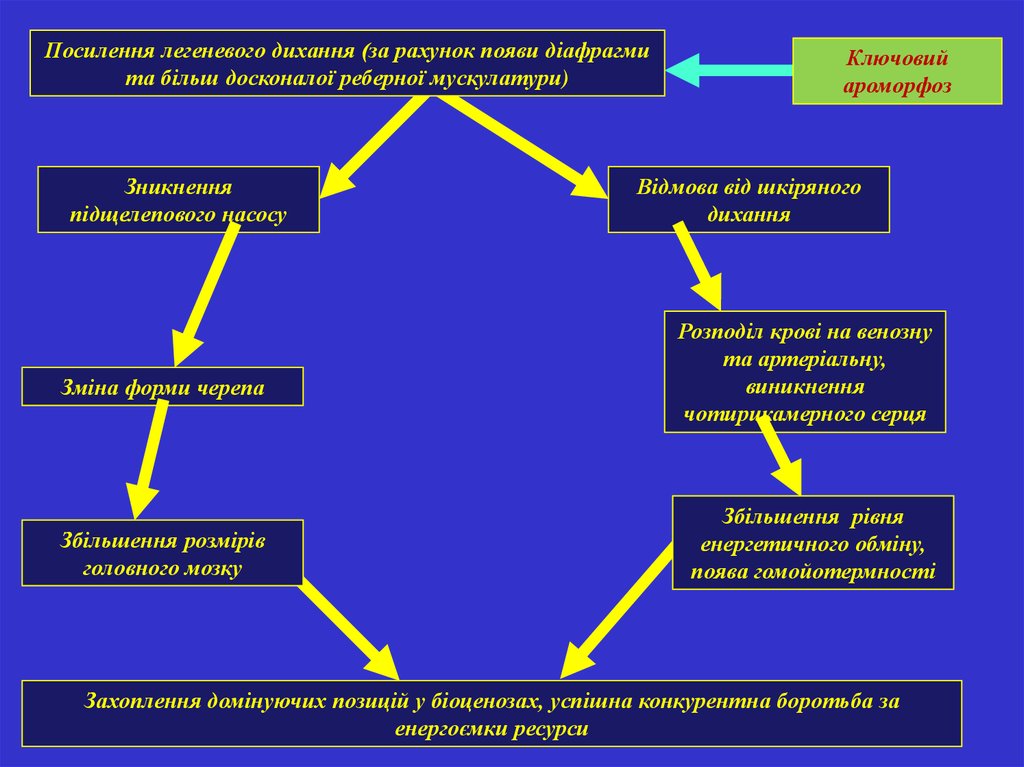
Посилення легеневого дихання (за рахунок появи діафрагмита більш досконалої реберної мускулатури)
Зникнення
підщелепового насосу
Зміна форми черепа
Збільшення розмірів
головного мозку
Ключовий
ароморфоз
Відмова від шкіряного
дихання
Розподіл крові на венозну
та артеріальну,
виникнення
чотирикамерного серця
Збільшення рівня
енергетичного обміну,
поява гомойотермності
Захоплення домінуючих позицій у біоценозах, успішна конкурентна боротьба за
енергоємки ресурси
26.
Паралельнаеволюція. Ссавці.
Процес набуття
ознак маммального
рівня організації
йшов паралельно в
кількох споріднених
групах
теріоморфних
рептилій і отримав
назву маммалізації
27.
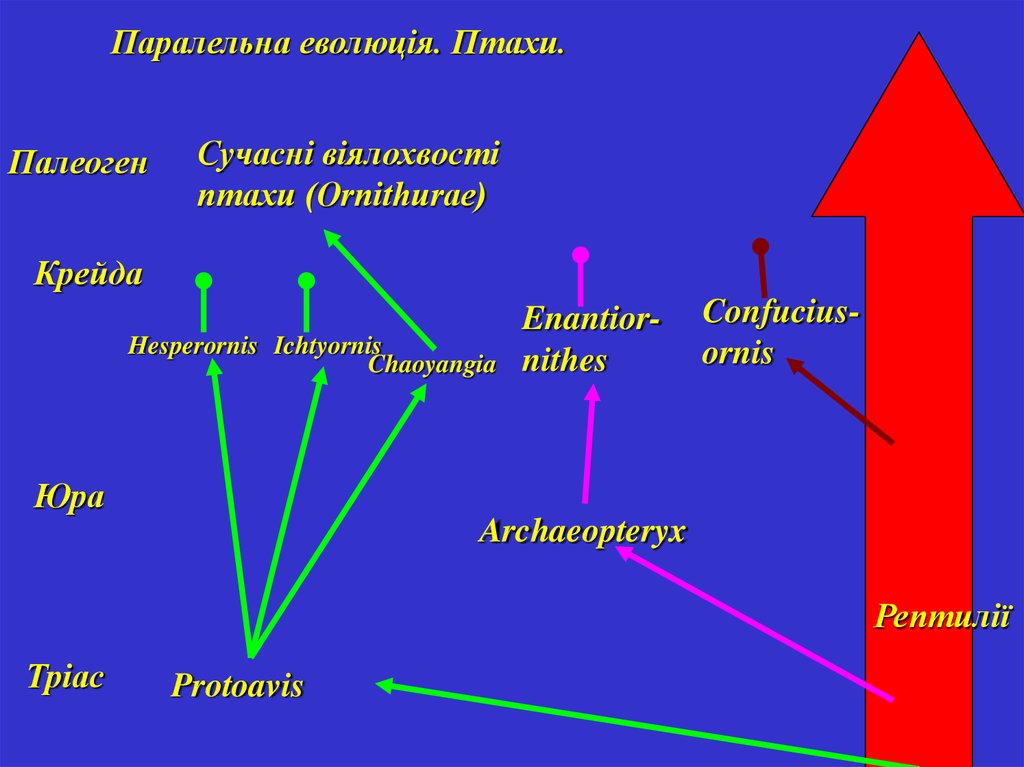
Паралельна еволюція. Птахи.Палеоген
Сучасні віялохвості
птахи (Ornithurae)
Крейда
Hesperornis Ichtyornis
Chaoyangia
Enantiornithes
Confuciusornis
Юра
Archaeopteryx
Рептилії
Тріас
Protoavis
28.
Деяки опірені динозаврипротоптахиАвімім
Конфуціусорніс, рахонавіс, енанціорніс
Криптоволанс
29.
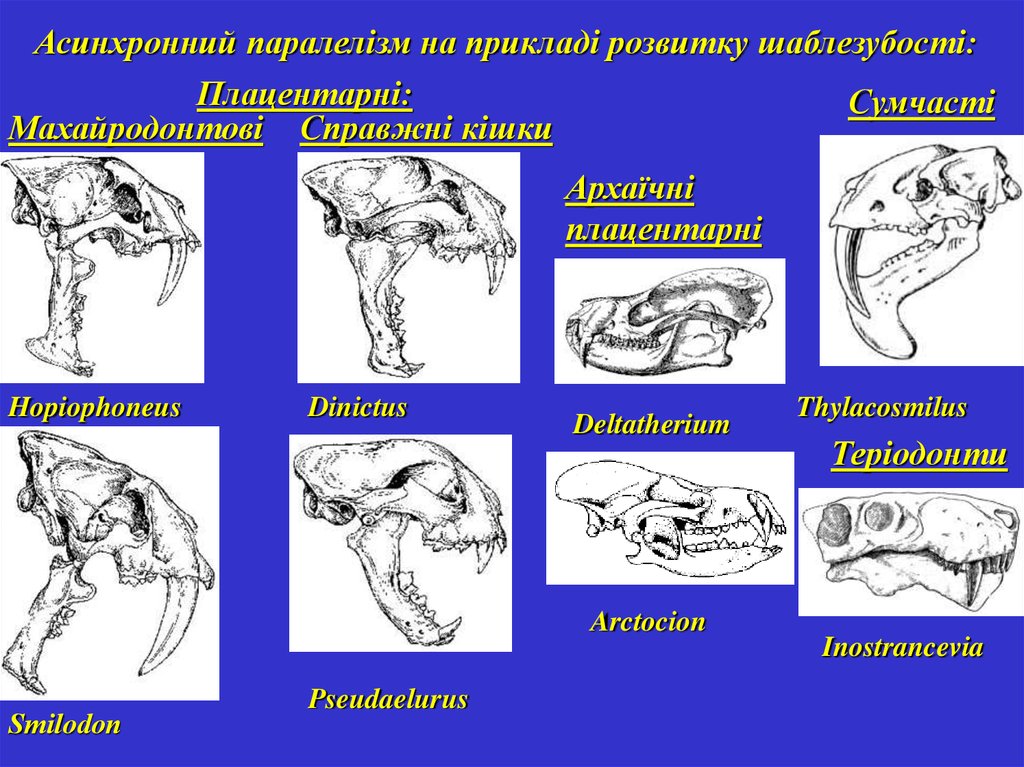
Асинхронний паралелізм на прикладі розвитку шаблезубості:Плацентарні:
Сумчасті
Махайродонтові Справжні кішки
Архаїчні
плацентарні
Hopiophoneus
Dinictus
Deltatherium
Arctocion
Smilоdon
Pseudaelurus
Thylacosmilus
Теріодонти
Inostrancevia
30.
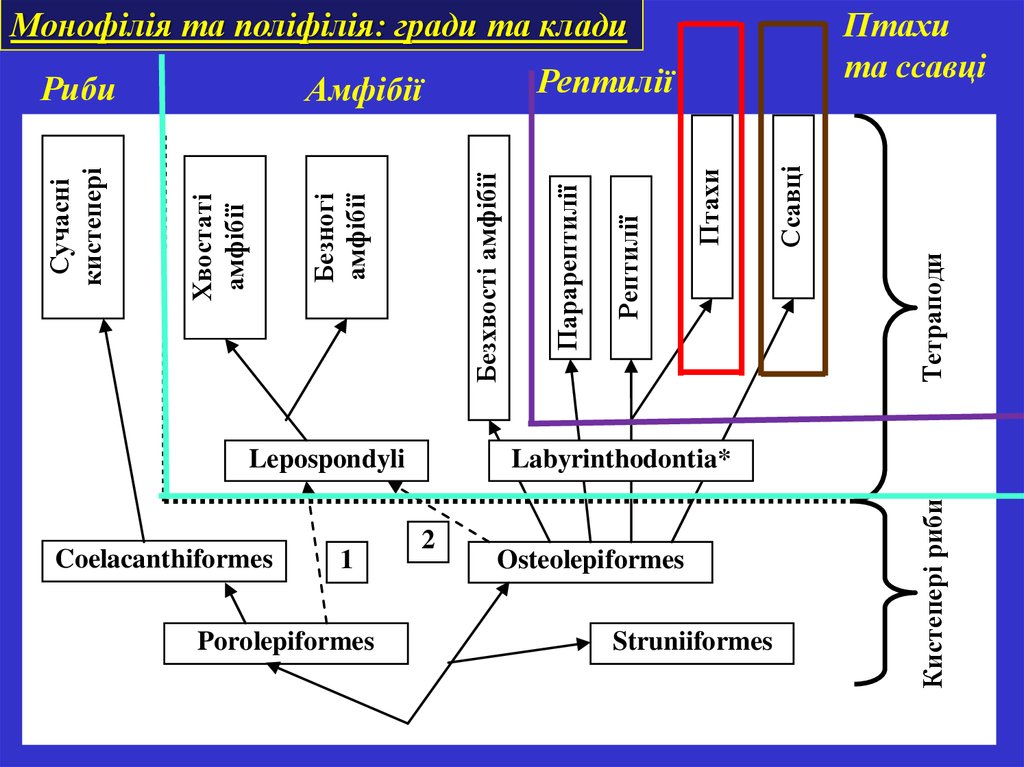
Монофілія та поліфілія: гради та клади1
Porolepiformes
Тетраподи
Ссавці
Птахи
Labyrinthodontia*
2
Osteolepiformes
Struniiformes
Кистепері риби
Coelacanthiformes
Рептилії
Безхвості амфібії
Безногі
амфібії
Lepospondyli
Парарептилії
Рептилії
Амфібії
Хвостаті
амфібії
Сучасні
кистепері
Риби
Птахи
та ссавці
31.
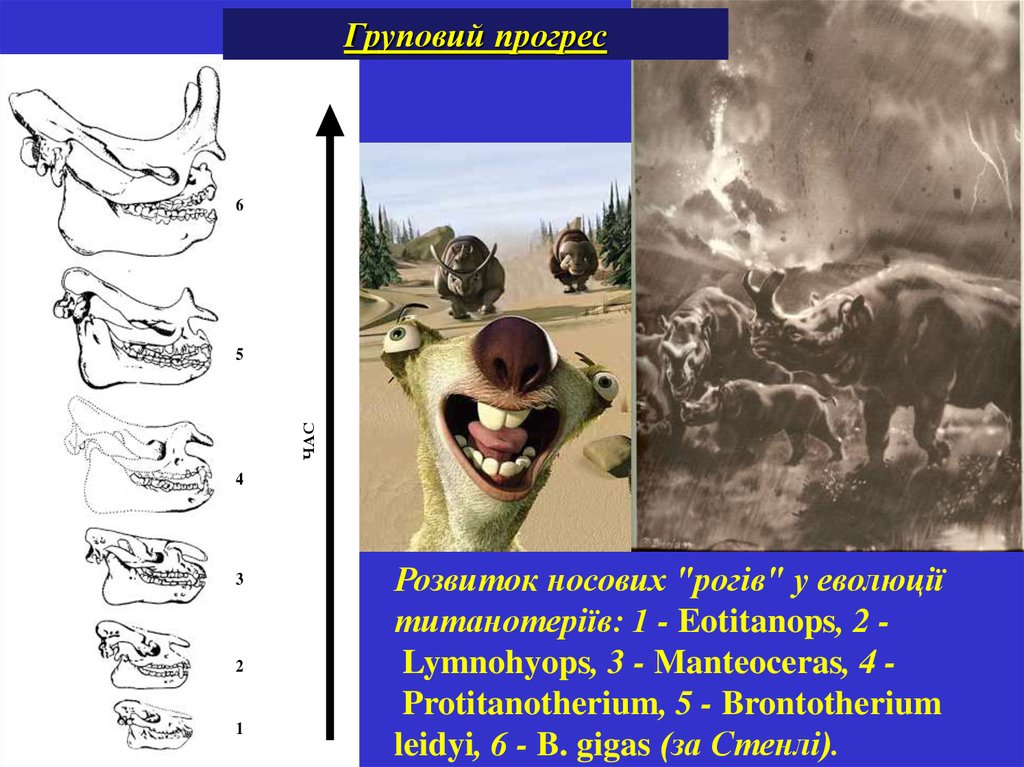
Груповий прогрес6
ЧАС
5
4
3
2
1
Розвиток носових "рогів" у еволюції
титанотеріїв: 1 - Eotitanops, 2 Lymnohyops, 3 - Manteoceras, 4 Protitanotherium, 5 - Brontotherium
leidyi, 6 - B. gigas (за Стенлі).
32.
Вік родів3-0
5-3
11-5
18-11
20-18
23-20
28-23
34-28
59-34
79-59
95-79
110-95
134-110
185-134
293-185
476-293
0%
20%
40%
60%
80%
100%
%
% вимерлих родів
% родів, що вижили
Найбільш стійкими до глобальної екологічної кризи на
границі крейди та палеогену виявились старі роди.
33.
Філогенетичний циклКількість видів
Середній вік видів
Об’єм вільного простору в
адаптивній зоні таксону
Середній об’єм видової
ніші (неспеціалізованість)
Швидкість появи видів
Швидкість вимирання
видів
Ріст - Розквіт - Занепад - Релікт
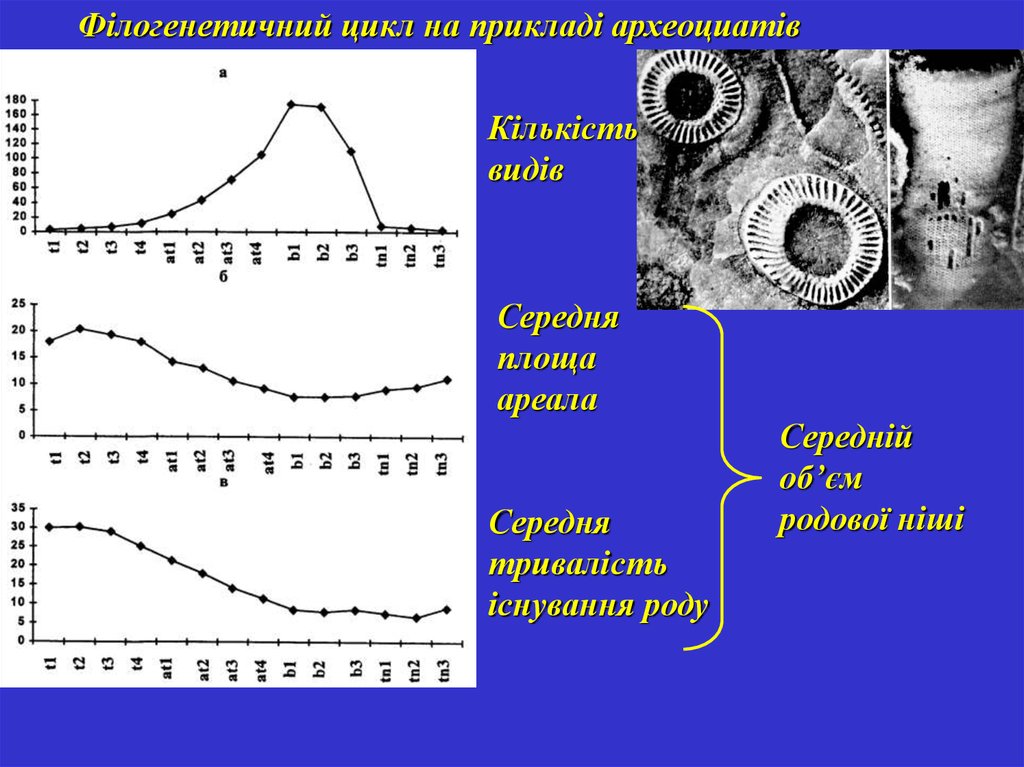
34.
Філогенетичний цикл на прикладі археоциатівКількість
видів
Середня
площа
ареала
Середня
тривалість
існування роду
Середній
об’єм
родової ніші
35.
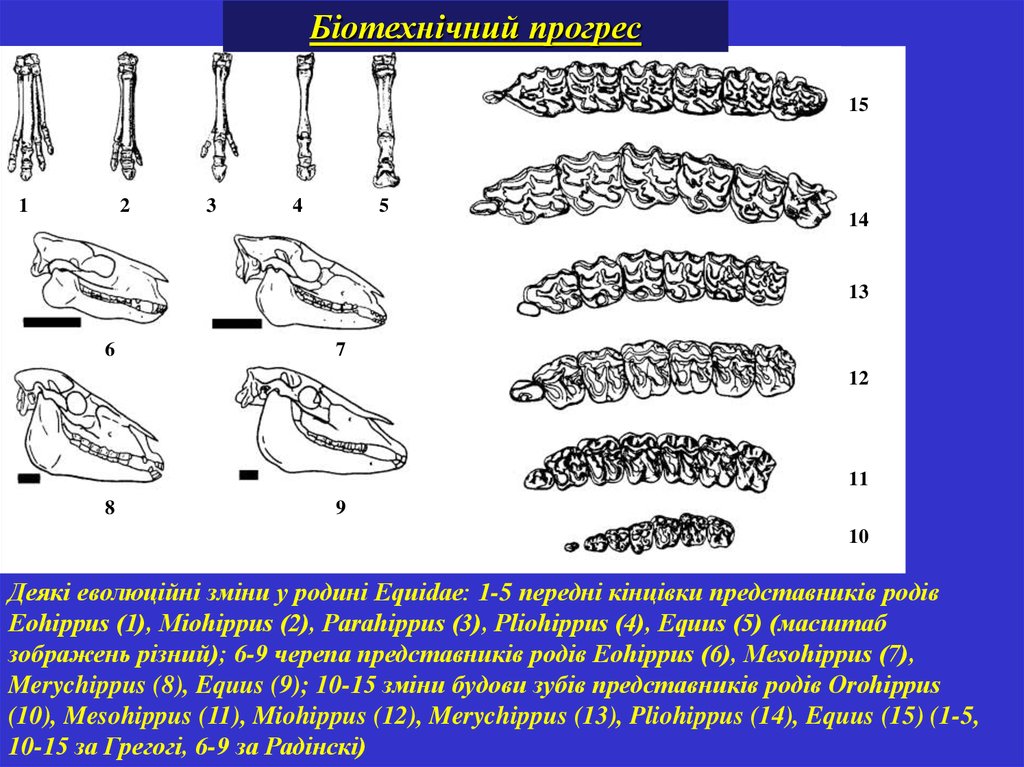
Біотехнічний прогрес15
1
2
3
4
5
14
13
6
7
12
11
8
9
10
Деякі еволюційні зміни у родині Equidae: 1-5 передні кінцівки представників родів
Eohippus (1), Miohippus (2), Parahippus (3), Pliohippus (4), Equus (5) (масштаб
зображень різний); 6-9 черепа представників родів Eohippus (6), Mesohippus (7),
Merychippus (8), Equus (9); 10-15 зміни будови зубів представників родів Orohippus
(10), Mesohippus (11), Miohippus (12), Merychippus (13), Pliohippus (14), Equus (15) (1-5,
10-15 за Грегогі, 6-9 за Радінскі)
36.
Адаптивний компроміс...а у пінгвіна спеціалізована
для плавання.
Форма тіла тіла
альбатроса
оптимальна для
польоту...
На відміну від
спеціалізованих
видів, генералісти,
такі як кайри,
мають проміжний
фенотип, компромісний для пересування у двох різних
типах середовища
37.
38.
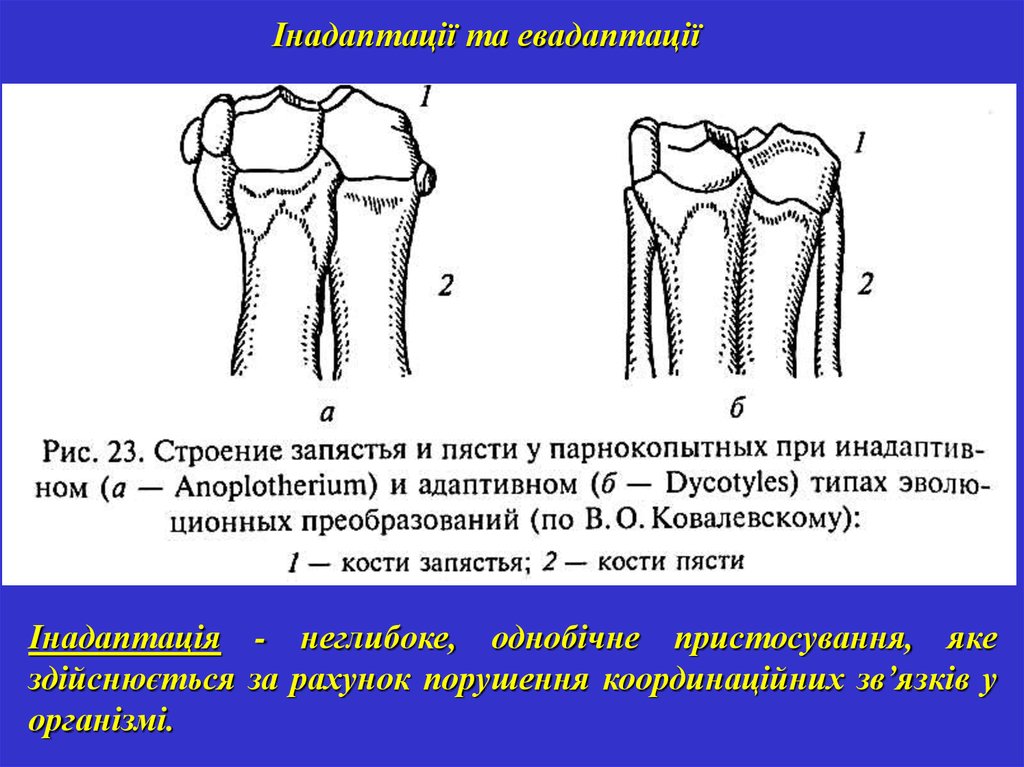
Інадаптації та евадаптаціїІнадаптація - неглибоке, однобічне пристосування, яке
здійснюється за рахунок порушення координаційних зв’язків у
організмі.
39.
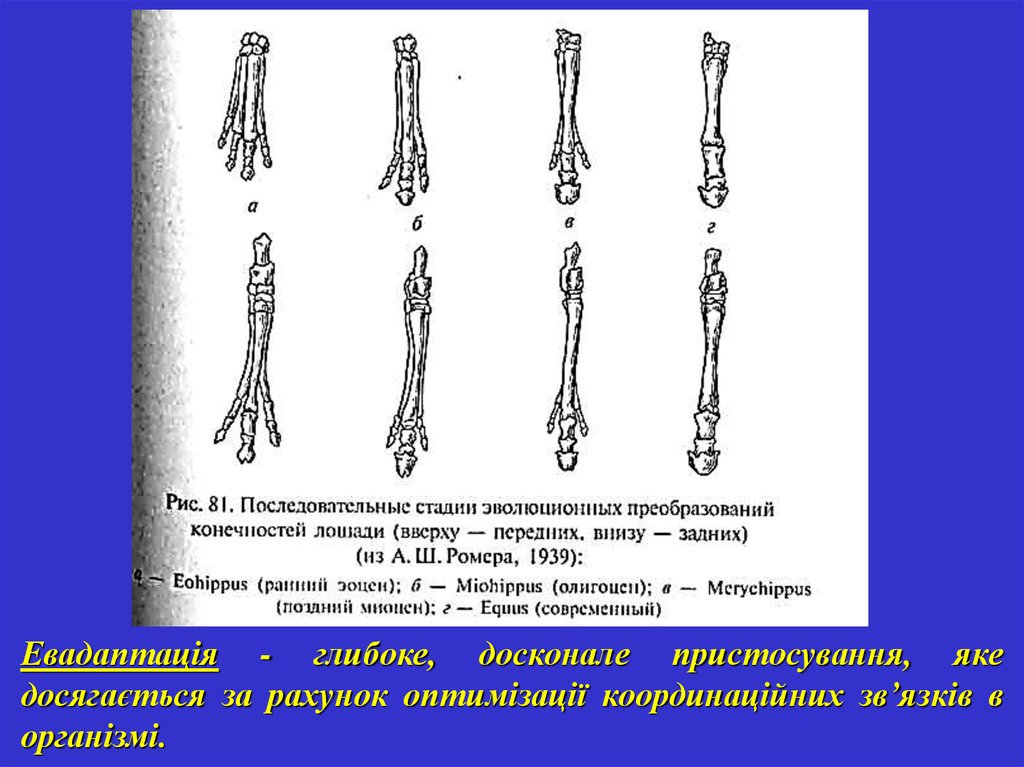
Евадаптація - глибоке, досконале пристосування, якедосягається за рахунок оптимізації координаційних зв’язків в
організмі.
40.
Група креодонтівдосягла швидкого
успіху, проте згодом
поступилася
справжнім хижим.
Оксієна
41.
Темпи еволюційних перетворень1 Дарвін : зміна розмірів органа на 0,001 його величини
за 1000 років
1 Сімпсон : зміна розмірів органа на 1 стандартне
(квадратичне) відхилення за 1 млн. років
1 Холдейн : зміна розмірів органа на 1 стандартне
(квадратичне) відхилення за 1 покоління
Час напіввимирання таксона : час за який вимирає 1/2
частина видів таксона
Вік напівсучасної фауни (фауни, в якій 1/2 видів
сучасна, а друга половина вимерла).
42.
Вік напівсучасної фауни родів деяких таксонівСсавці - 4 млн р.
Птахі - 10 млн. р.
Амфібії - 10 млн. р.
Рептилії - 20 млн. р.
Риби - 30-50 млн. р.
Комахи - 40 млн. р.
Фораменіфери - 230 млн. р.
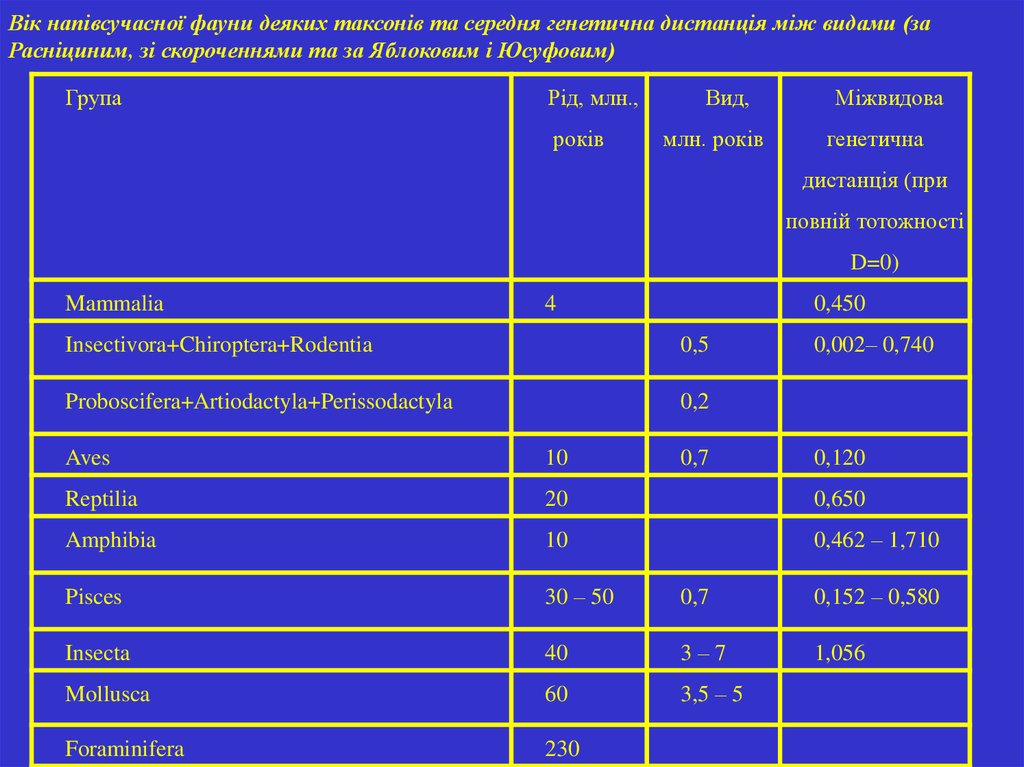
43.
Вік напівсучасної фауни деяких таксонів та середня генетична дистанція між видами (заРасніциним, зі скороченнями та за Яблоковим і Юсуфовим)
Група
Рід, млн.,
років
Вид,
млн. років
Міжвидова
генетична
дистанція (при
повній тотожності
D=0)
Mammalia
4
0,450
Insectivora+Chiroptera+Rodentia
0,5
Proboscifera+Artiodactyla+Perissodactyla
0,2
0,7
0,002– 0,740
Aves
10
0,120
Reptilia
20
0,650
Amphibia
10
0,462 – 1,710
Pisces
30 – 50
0,7
0,152 – 0,580
Insecta
40
3–7
1,056
Mollusca
60
3,5 – 5
Foraminifera
230
44.
Живі викопні роди, які існуютьна Землі десяткисотні міліонів років
45.
46.
Типи живих викопнихВиди – ізоляти (мешканці
рефугіумів)
Види – мешканці
реліктових біогеоценозів
Види, що займають
підпорядковане
положення у
біогеоценозах
Гаттерія
Деревовидна
папороть
Fannia
canicularis
47.
Популяції видуDelamarentulus tristani
Silv. з групи Protura
мешкають у КостаРіці та у Західній
Африці. Ізоляція
популяцій цього виду
відбулася близько
80млн. років тому.
Еволюційному стазису сприяє стабільність умов,
стабільність екологічної ніші, яка більша у видів, що
займають підпорядковане положення у біоценозах.
Види, що займають домінуючі позиції у біоценозах (віоленти)
навпаки виявляються найбільш чутливими до перебудови
екосистем завдяки прунінгу – випадіння останнях стадій
сукцесії.
48.
Фактори, що сприяють швидкій еволюції груп:Статеве розмноження
Можливість дії статевого добору
Складність будови
Великі розміри
Довгий онтогенез
Мала кількість нащадків
Турбота про нащадків
Великий діапазон внутрішньовидової мінливості ознак
Мінімальний діапазон внутрішньовидової генетичної мінливості
Складність міжгенних взаємодій
Панівні позиції в біогеоценозі
Різноманіття умов середовища
Максимально комфортні абіотичні умови
Кондиціонування зовнішнього середовища
Великий ареал
Складність екосистем