






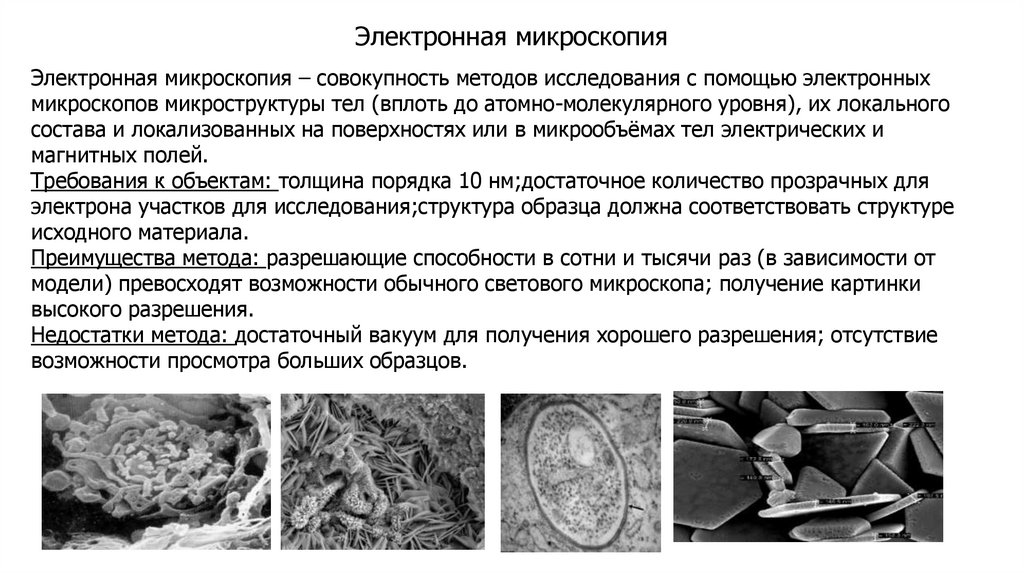
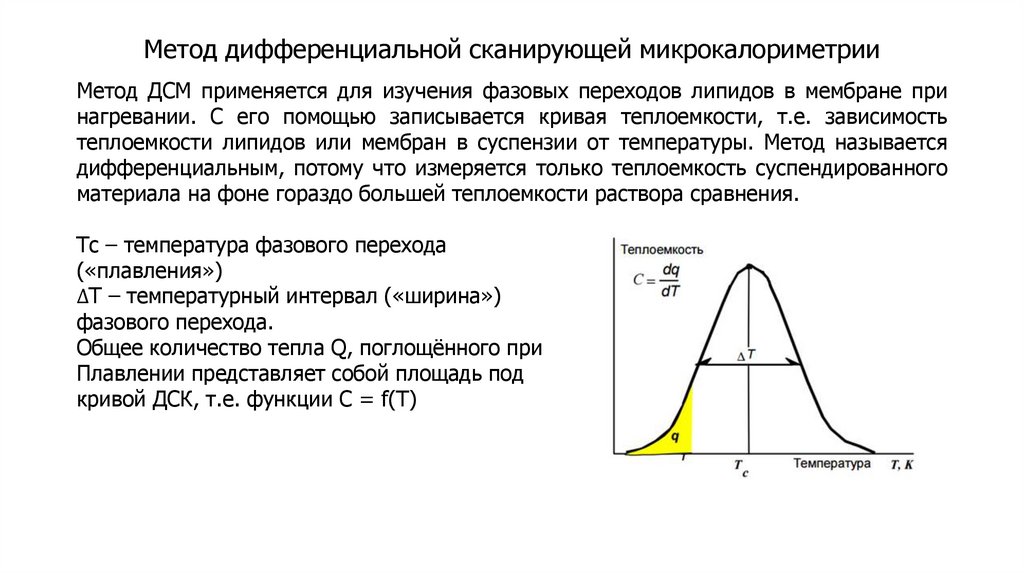




 biology
biologySimilar presentations:










Структурно-функциональная организация биологических мембран. Состав, строение и методы изучения биологических мембран
1.
Лекция №1Структурно-функциональная организация
биологических мембран.
Состав, строение и методы изучения
биологических мембран.
2.
Термин «клеточная мембрана» (в переводе с лат. «membrana» означает - кожица,плёнка) был введен в конце 19 века для обозначения клеточной границы, служащей:
- барьером между содержимым клетки и окружающей средой;
- полупроницаемой перегородкой, через которую могут переходить вода и
некоторые вещества.
Однако,
этим функционал клеточных мембран не исчерпывается, поскольку
биологические мембраны составляют основу структурно-функциональной организации
жизни на клеточном уровне.
3.
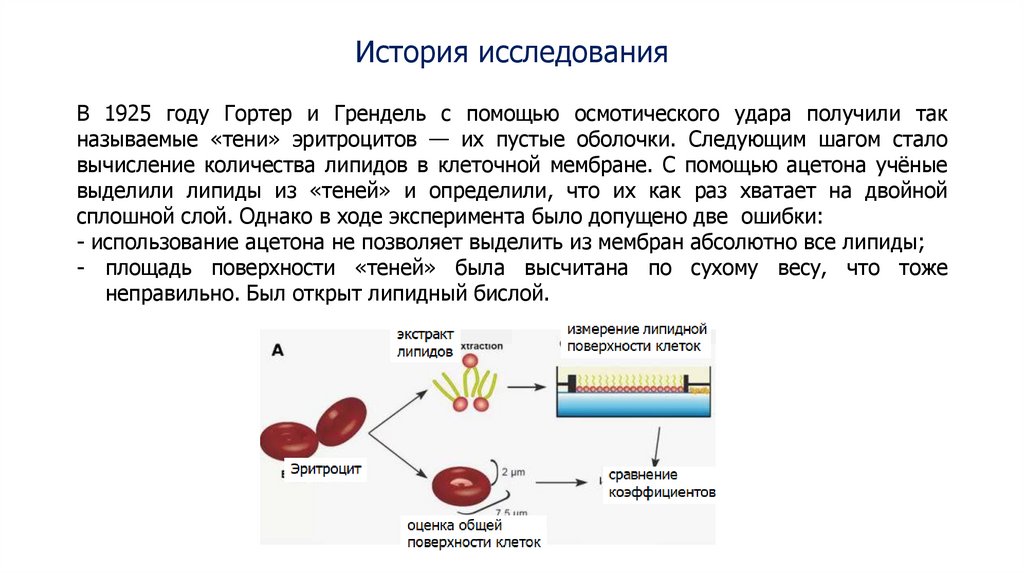
История исследованияВ 1925 году Гортер и Грендель с помощью осмотического удара получили так
называемые «тени» эритроцитов — их пустые оболочки. Следующим шагом стало
вычисление количества липидов в клеточной мембране. С помощью ацетона учёные
выделили липиды из «теней» и определили, что их как раз хватает на двойной
сплошной слой. Однако в ходе эксперимента было допущено две ошибки:
- использование ацетона не позволяет выделить из мембран абсолютно все липиды;
- площадь поверхности «теней» была высчитана по сухому весу, что тоже
неправильно. Был открыт липидный бислой.
4.
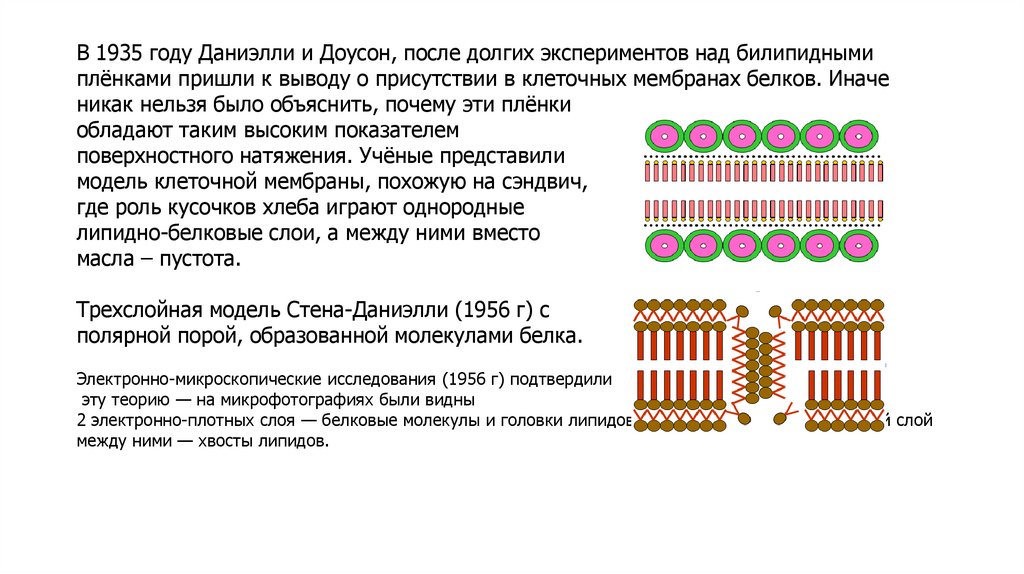
В 1935 году Даниэлли и Доусон, после долгих экспериментов над билипиднымиплёнками пришли к выводу о присутствии в клеточных мембранах белков. Иначе
никак нельзя было объяснить, почему эти плёнки
обладают таким высоким показателем
поверхностного натяжения. Учёные представили
модель клеточной мембраны, похожую на сэндвич,
где роль кусочков хлеба играют однородные
липидно-белковые слои, а между ними вместо
масла – пустота.
Трехслойная модель Стена-Даниэлли (1956 г) с
полярной порой, образованной молекулами белка.
Электронно-микроскопические исследования (1956 г) подтвердили
эту теорию — на микрофотографиях были видны
2 электронно-плотных слоя — белковые молекулы и головки липидов и один электронно-прозрачный слой
между ними — хвосты липидов.
5.
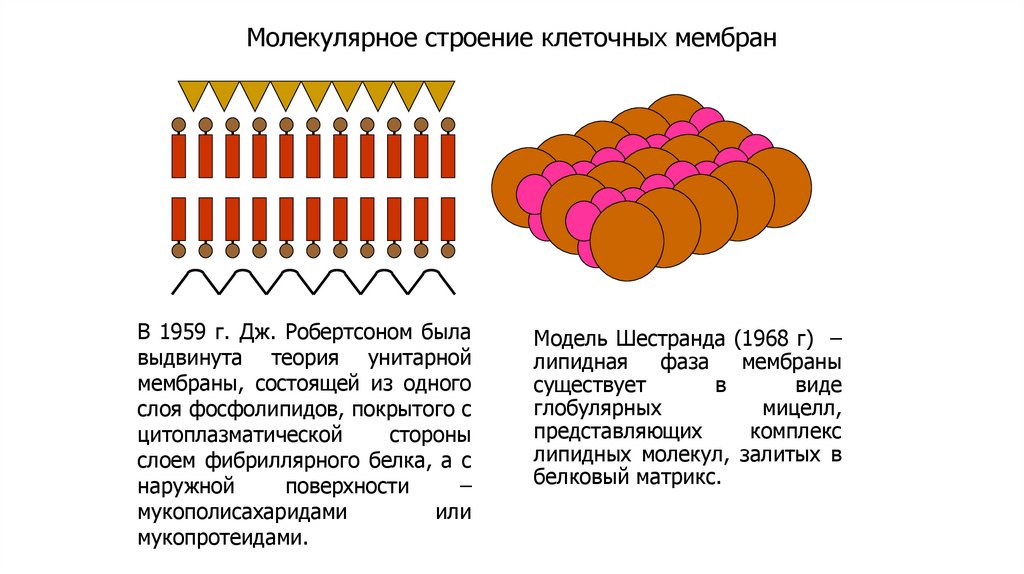
Молекулярное строение клеточных мембранВ 1959 г. Дж. Робертсоном была
выдвинута теория унитарной
мембраны, состоящей из одного
слоя фосфолипидов, покрытого с
цитоплазматической
стороны
слоем фибриллярного белка, а с
наружной
поверхности
–
мукополисахаридами
или
мукопротеидами.
Модель Шестранда (1968 г) –
липидная
фаза
мембраны
существует
в
виде
глобулярных
мицелл,
представляющих
комплекс
липидных молекул, залитых в
белковый матрикс.
6.
1959 г Дж.Д. Робертсон предположил, чтовсе клеточные мембраны построены по
одному принципу, и высказал концепцию
унитарной
(или
единообразной
мембраны). Эта модель во многом сходна
с классической моделью Дж.Даниелли:
основу мембраны составляет липидный
бислой, а нелипидные компоненты лежат
на поверхности связываясь с липидами за
счет электростатических и гидрофобных
взаимодействий.
В его модели нашла отражение
важная структурная особенность
– асимметрия.
7.
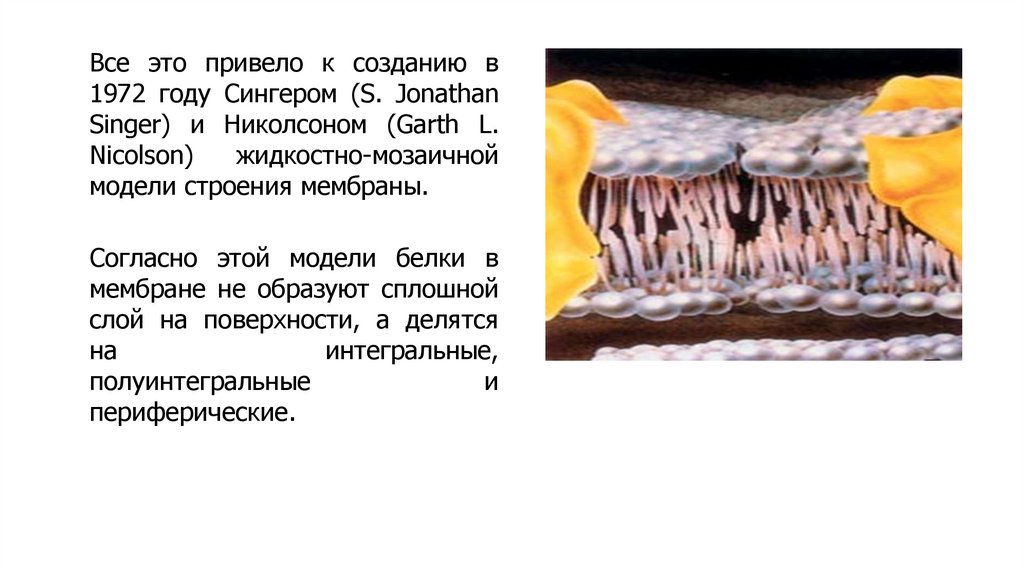
Все это привело к созданию в1972 году Сингером (S. Jonathan
Singer) и Николсоном (Garth L.
Nicolson)
жидкостно-мозаичной
модели строения мембраны.
Согласно этой модели белки в
мембране не образуют сплошной
слой на поверхности, а делятся
на
интегральные,
полуинтегральные
и
периферические.
8.
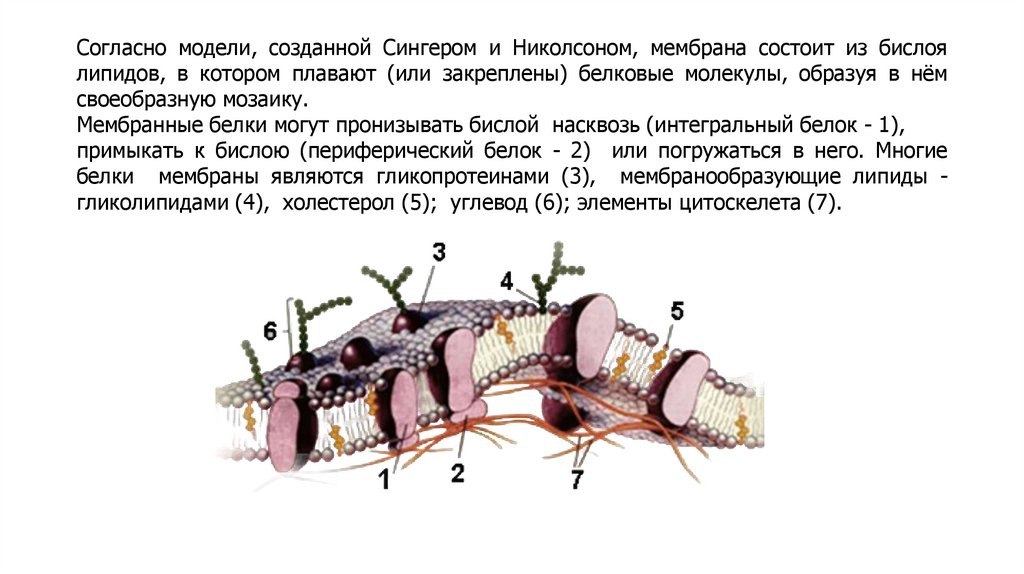
Согласно модели, созданной Сингером и Николсоном, мембрана состоит из бислоялипидов, в котором плавают (или закреплены) белковые молекулы, образуя в нём
своеобразную мозаику.
Мембранные белки могут пронизывать бислой насквозь (интегральный белок - 1),
примыкать к бислою (периферический белок - 2) или погружаться в него. Многие
белки мембраны являются гликопротеинами (3), мембранообразующие липиды гликолипидами (4), холестерол (5); углевод (6); элементы цитоскелета (7).
9.
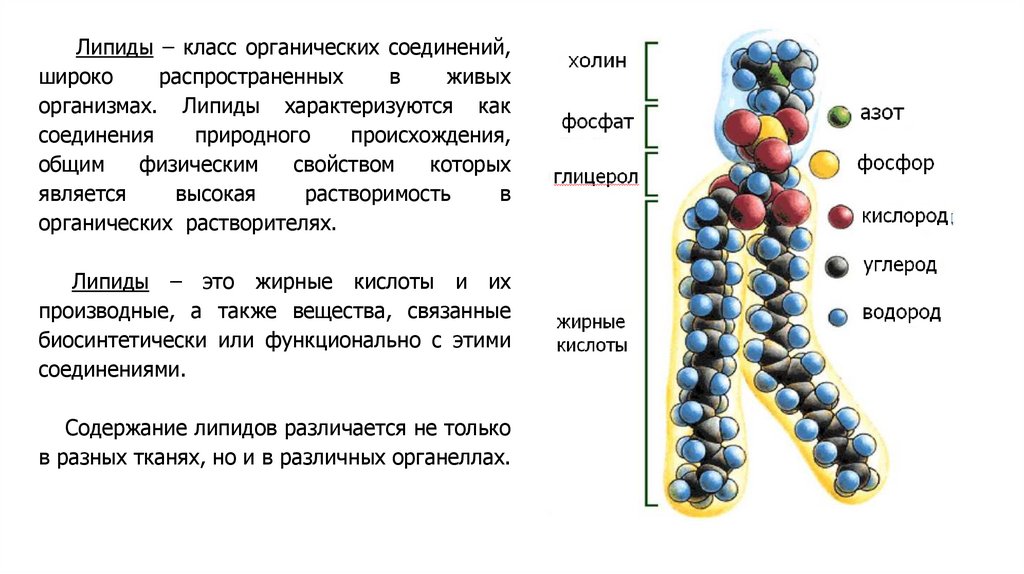
Липиды – класс органических соединений,широко
распространенных
в
живых
организмах. Липиды характеризуются как
соединения
природного
происхождения,
общим
физическим
свойством
которых
является
высокая
растворимость
в
органических растворителях.
Липиды – это жирные кислоты и их
производные, а также вещества, связанные
биосинтетически или функционально с этими
соединениями.
Содержание липидов различается не только
в разных тканях, но и в различных органеллах.
10.
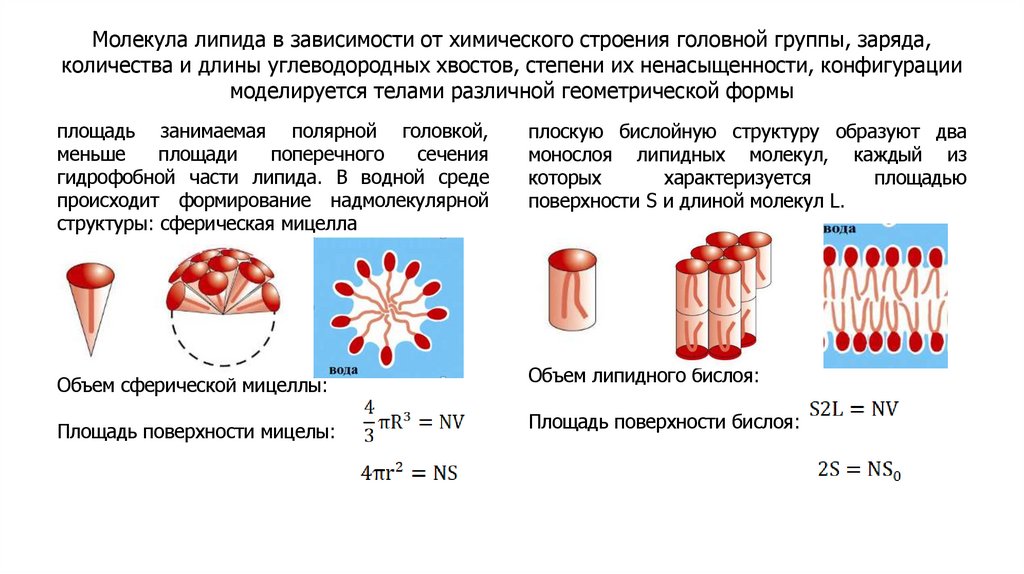
Молекула липида в зависимости от химического строения головной группы, заряда,количества и длины углеводородных хвостов, степени их ненасыщенности, конфигурации
моделируется телами различной геометрической формы
площадь занимаемая полярной головкой,
меньше
площади
поперечного
сечения
гидрофобной части липида. В водной среде
происходит формирование надмолекулярной
структуры: сферическая мицелла
плоскую бислойную структуру образуют два
монослоя липидных молекул, каждый из
которых
характеризуется
площадью
поверхности S и длиной молекул L.
Объем сферической мицеллы:
Объем липидного бислоя:
Площадь поверхности мицелы:
Площадь поверхности бислоя:
11.
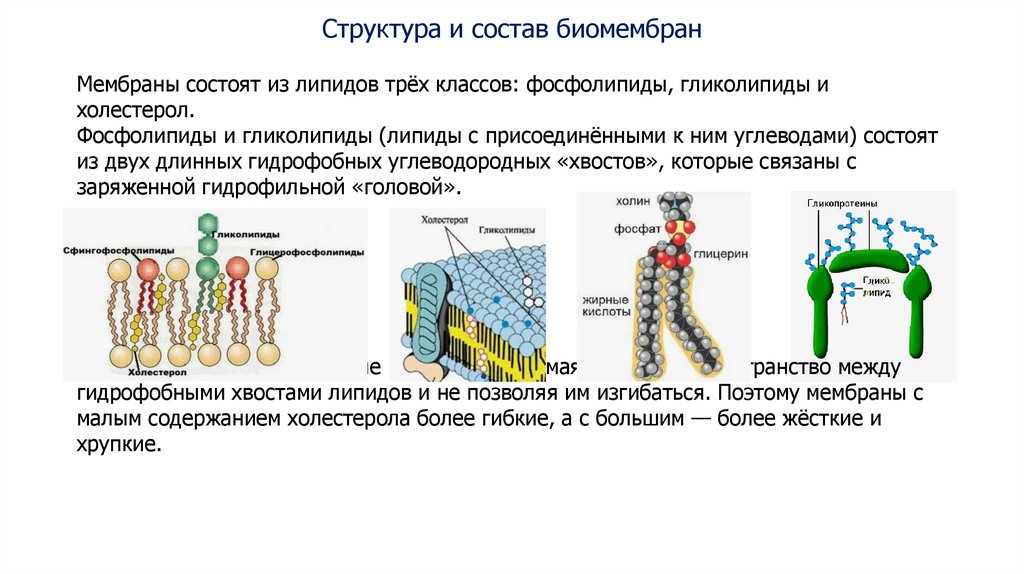
Структура и состав биомембранМембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и
холестерол.
Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят
из двух длинных гидрофобных углеводородных «хвостов», которые связаны с
заряженной гидрофильной «головой».
Холестерол придаёт мембране жёсткость, занимая свободное пространство между
гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с
малым содержанием холестерола более гибкие, а с большим — более жёсткие и
хрупкие.
12.
Гликолипиды - сложные липиды, образующиеся в результате соединения липидов суглеводами. Гликолипиды широко представлены в нервной ткани, в частности в ткани
мозга. Они локализованы преимущественно на наружной поверхности плазматической
мембраны, где их углеводные компоненты входят в число других углеводов клеточной
поверхности.
Холестери́ н — органическое соединение, природный жирный (липофильный) спирт,
содержащийся в клеточных мембранах всех живых организмов, за исключением
безъядерных (прокариоты). Нерастворим в воде, растворим в жирах и органических
растворителях.
Белки мембраны клеток представлены, в основном, гликопротеинами.
Различают: интегральные белки, проникающие через всю толщу мембраны и
периферические белки, прикрепленные только к поверхности мембраны, в основном, к
внутренней ее части.
Мембранные липиды – амфипатические молекулы, и в водной среде большинство из них
самопроизвольно образует бислои. Именно это свойство обусловливает самосборку
липидных бислоев в клетке и их способность самопроизвольно восстанавливаться при
повреждениях.
Различный липидный состав характерен и для разных мембран одной и той же
эукариотической клетки.
13.
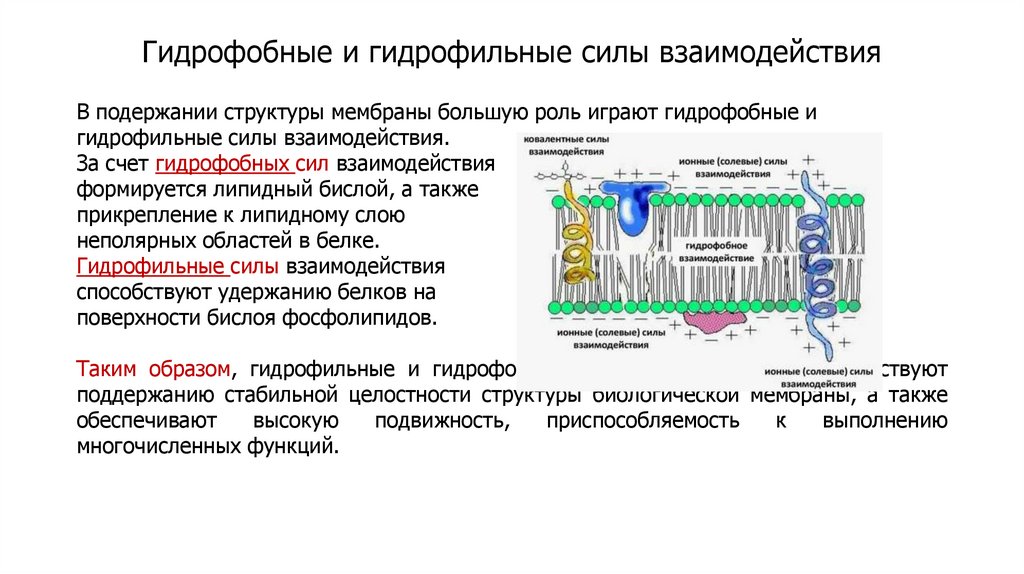
Гидрофобные и гидрофильные силы взаимодействияВ подержании структуры мембраны большую роль играют гидрофобные и
гидрофильные силы взаимодействия.
За счет гидрофобных сил взаимодействия
формируется липидный бислой, а также
прикрепление к липидному слою
неполярных областей в белке.
Гидрофильные силы взаимодействия
способствуют удержанию белков на
поверхности бислоя фосфолипидов.
Таким образом, гидрофильные и гидрофобные силы взаимодействия способствуют
поддержанию стабильной целостности структуры биологической мембраны, а также
обеспечивают
высокую
подвижность,
приспособляемость
к
выполнению
многочисленных функций.
14.
Свойства биологических мембран1.Замкнутость.
Липидные бислои (и мембраны) всегда самостоятельно замыкаются на себе с
образованием полностью ограниченных отсеков. При нарушении целостности
мембраны происходит ее «самосшивание».
2.Латеральная подвижность.
Компоненты мембраны могут перемещаться в пределах своего слоя.
Мембраны обладают свойствами двумерных жидкостей. По этой причине
модель строения биомембран называется жидкостно-мозаичной.
3.Асимметрия.
Наружная и внутренняя поверхности мембраны различаются по своему
составу:
-углеводные компоненты находятся с внешней поверхности плазмолеммы;
-многие белки расположены всегда только с наружной, а другие - только с
внутренней стороны;
Полярность (асимметрия) мембраны возникает на ранних стадиях ее
формирования и затем все время сохраняется.
15.
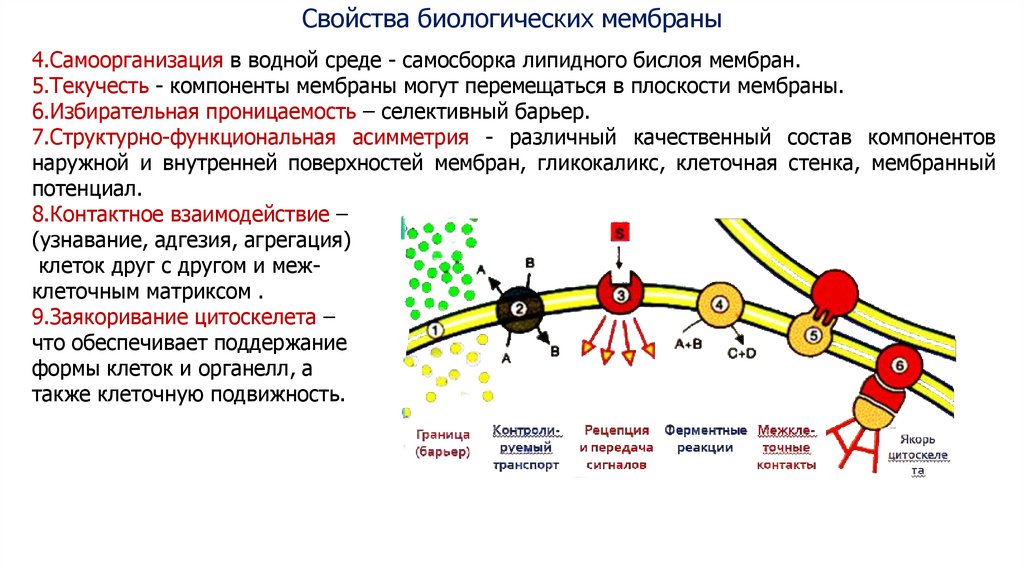
Свойства биологических мембраны4.Самоорганизация в водной среде - самосборка липидного бислоя мембран.
5.Текучесть - компоненты мембраны могут перемещаться в плоскости мембраны.
6.Избирательная проницаемость – селективный барьер.
7.Структурно-функциональная асимметрия - различный качественный состав компонентов
наружной и внутренней поверхностей мембран, гликокаликс, клеточная стенка, мембранный
потенциал.
8.Контактное взаимодействие –
(узнавание, адгезия, агрегация)
клеток друг с другом и межклеточным матриксом .
9.Заякоривание цитоскелета –
что обеспечивает поддержание
формы клеток и органелл, а
также клеточную подвижность.
16.
Функции биологических мембран1. Барьерная – обеспечивает регулируемый, пассивный и активный
обмен веществ; ограничение и обособлФение клеток и органелл.
2. Матричная – обеспечивает определенное взаимное расположение и
ориентацию мембранных белков, обеспечивает их оптимальное
взаимодействие.
3. Механическая – обеспечивает прочность и автономность клетки,
внутриклеточных структур.
4. Энергетическая – синтез АТФ на внутренних мембранах митохондрий
и фотосинтез в мембранах хлоропластов.
5. Транспортная - контролирует транспорт метаболитов и ионов с
целью поддержания внутриклеточного гомеостаза.
17.
Функции биологических мембран6. Информационная – восприятие внеклеточных информационных
сигналов и их передача внутрь клетки, а также инициация собственных
сигналов.
7. Ферментативный катализ реакций с неполярными субстратами –
биосинтез липидов, метаболизм неполярных ксенобиотиков (цитохром
Р450); реакции фотосинтеза и окислительного фосфорилирования
(дыхательная цепь митохондрий) .
8. Рецепторная – механическая, акустическая, обонятельная,
зрительная, химическая, терморецепция – мембранные
процессы)
18.
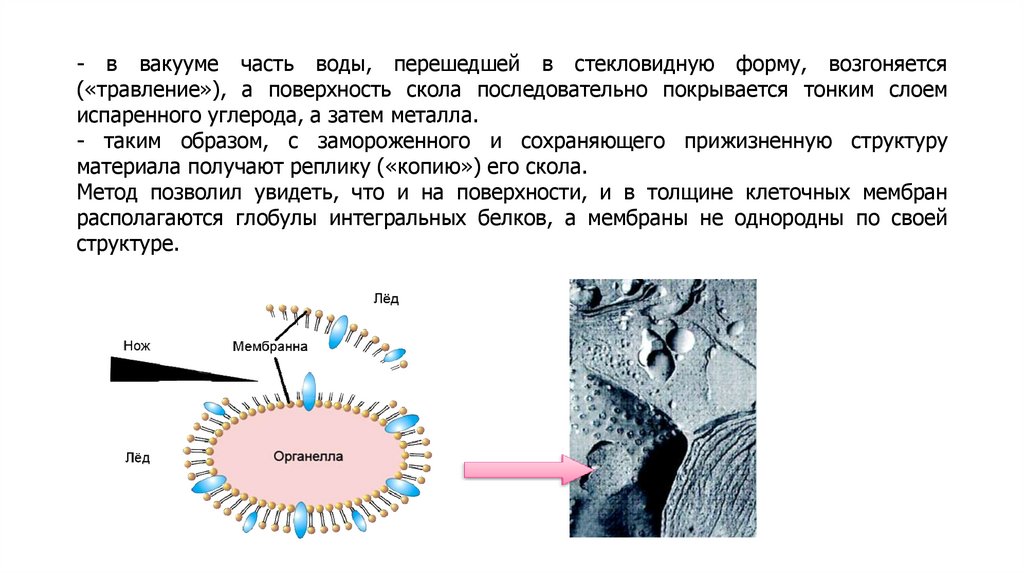
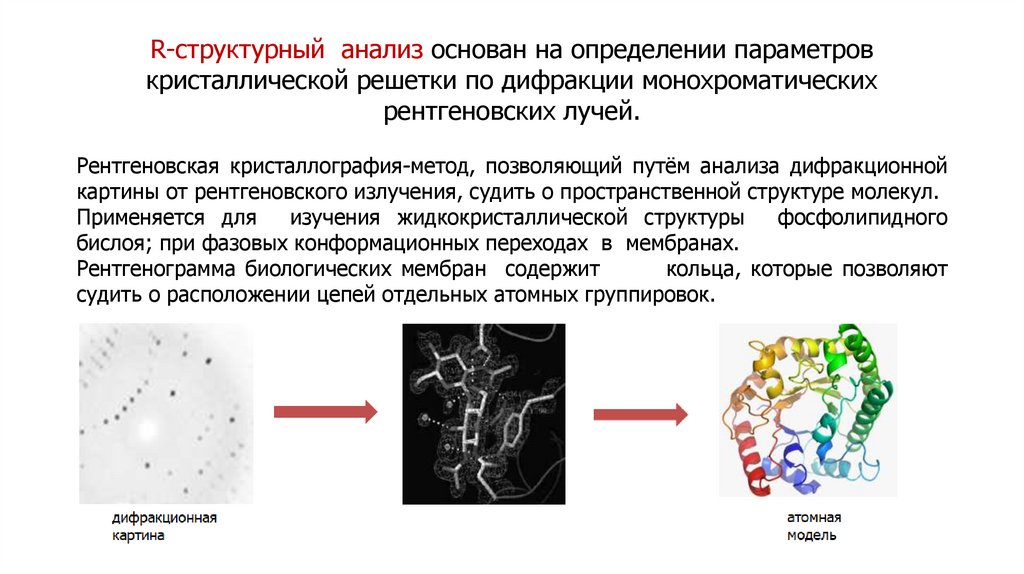
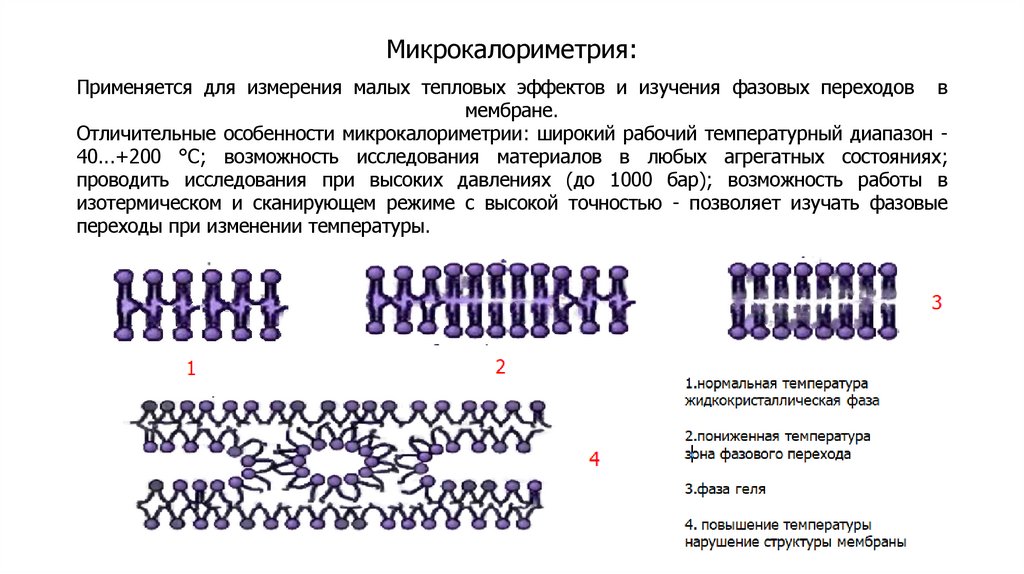
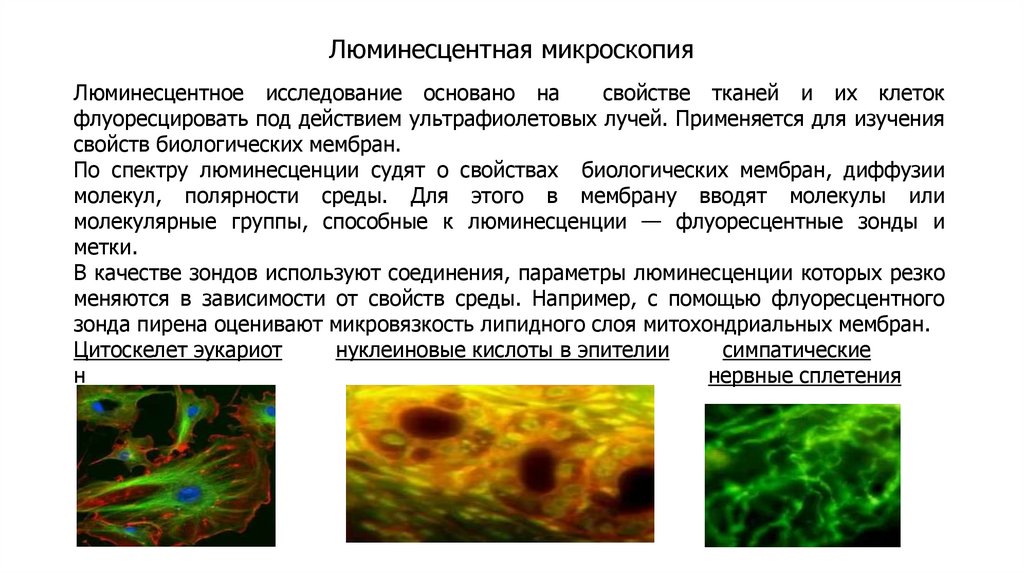
Методы изучения плазматических мембранВыделение мембранных фракций
Разрушение мембран
Осмотический шок (эритроциты), растирание с кварцевым
песком, размельчение гомогенизаторами.
Центрифугирование
Скорость осаждения пропорциональна массе частиц.
Разрушенные мембраны замыкаются в везикулы. Смесь
везикул разной природы, но с одинаковой плотностью,
называется фракцией микросом.