








































































 biology
biologySimilar presentations:










Сравнительная морфология живых организмов
1.
Учение А.Н.Северцова о филэмбриогенезах.Сравнительная морфология.
Смирнова С.Н.
2.
В процессе сравнительного изучения морфологииживых организмов на основах дарвинизма был открыт
целый ряд важных закономерностей эволюции
организмов. Однако эти открытия совершались
попутно, при изучении специальных вопросов
морфологии. Между тем развитие дарвинизма и
морфологии требовало более глубокого проникновения
в закономерности исторического развития и эволюции
мира живых организмов.
Для такого изучения Эрнст Геккель (нем.
биолог, основатель филогенетического направления
дарвинизма)
предложил
использовать
метод
тройственного
параллелизма,
т.е.
проводить
сопоставление данных палеонтологии, сравнительной
анатомии и эмбриологии при изучении эволюции
живых
организмов.
Э.Геккелем
(1866)
были
предложены два термина: «онтогенез» и «филогенез».
Эрнст Геккель
3.
Онтогенез - это индивидуальное развитие особи с момента еезарождения (формировании зиготы) и до конца жизни. В ходе
онтогенеза происходит развертывание генетической информации,
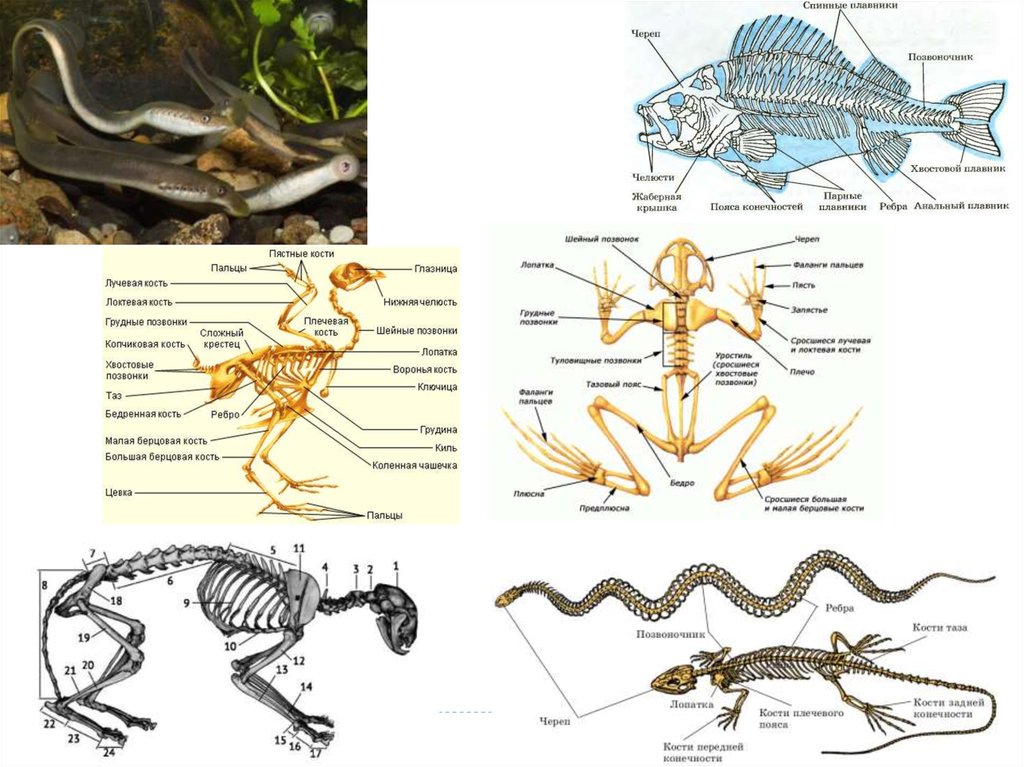
заложенной в гаметах. Т.к. все живые организмы разнообразны, то и
разнообразны их онтогенезы по продолжительности, темпам
развития и характеру дифференцировки тканей.
В историческом плане развития организмов их онтогенезы
менялись, приспосабливаясь к меняющимся условиям среды.
Наблюдалась тенденция к усложнению, а иногда и к упрощению
онтогенеза в ходе эволюции. Таким образом, онтогенез связан с
филогенезом.
4.
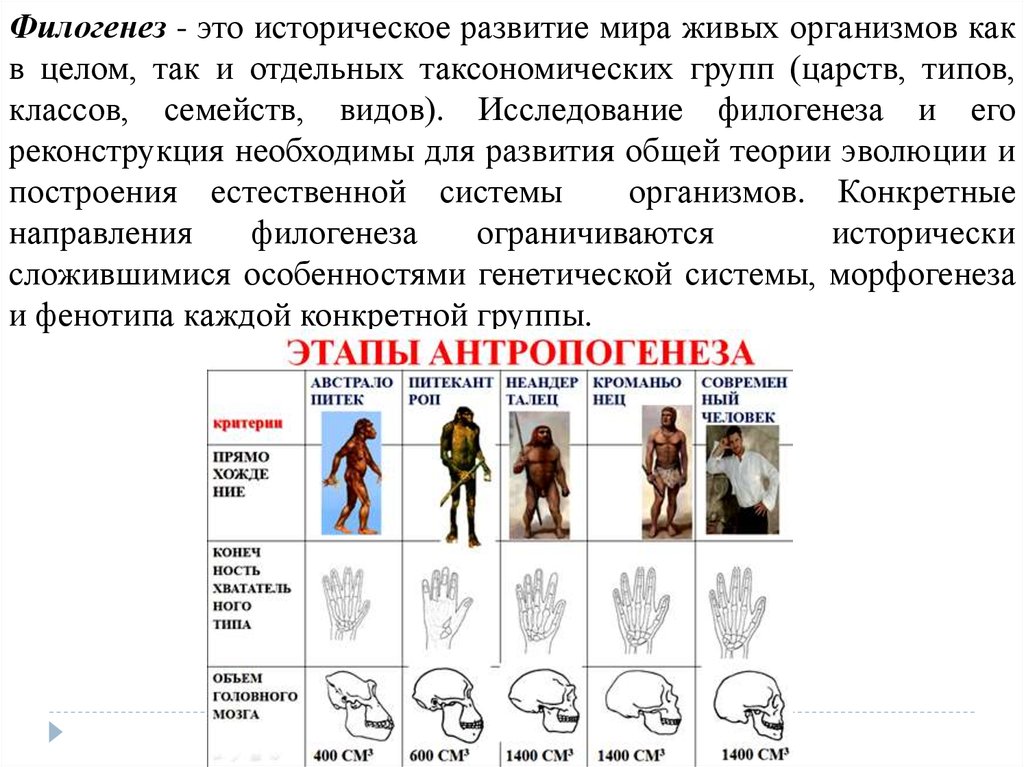
Филогенез - это историческое развитие мира живых организмов какв целом, так и отдельных таксономических групп (царств, типов,
классов, семейств, видов). Исследование филогенеза и его
реконструкция необходимы для развития общей теории эволюции и
построения естественной системы
организмов. Конкретные
направления
филогенеза
ограничиваются
исторически
сложившимися особенностями генетической системы, морфогенеза
и фенотипа каждой конкретной группы.
5.
Существуют следующие правила филогенеза :1-е - правило необратимости эволюции - невозможность
возврата любой группы организмов в состояние, пройденное
прежде.
2-е - правило прогрессивной специализации - если группа
организмов начала эволюционировать, то и в дальнейшем она
углубляет свою специализацию в наметившемся направлении.
3-е - правило происхождения новых групп организмов от
неспециализированных предков.
4-е - правило адаптивной радиации - разветвление
предкового ствола группы организмов в ходе приспособительной
эволюции на обособленные ветви. Филогенез любой группы
сопровождается ее дроблением на ряд более мелких стволов,
которые расходятся в разных направлениях от исходного уровня.
6.
Вопрос об отношении между эмбриональными признаками ипризнаками взрослых форм давно привлекал внимание биологов.
Еще И.Меккель (нем.зоолог) в 1821 году довольно близко подошел
к явлению повторения древних признаков предков в онтогенезе
потомков. На эти факты обратил внимание и К.М.Бэр
(рус.эмбриолог, 1828), который установил, что эмбрионы различных
животных имеют большее сходство между собой, чем взрослые
формы.
Ч.Дарвин
(1809-1882)
высоко
ценил
значение
эмбриологических фактов для раскрытия истории животного мира.
Он отмечал, что сходство в строении зародышей животных,
отличающихся во взрослом состоянии друг от друга,
свидетельствуют об общности их происхождения. Таким образом,
Ч.Дарвин первым высказал мысль о том, что онтогенез и
филогенез тесно связаны и взаимообусловлены.
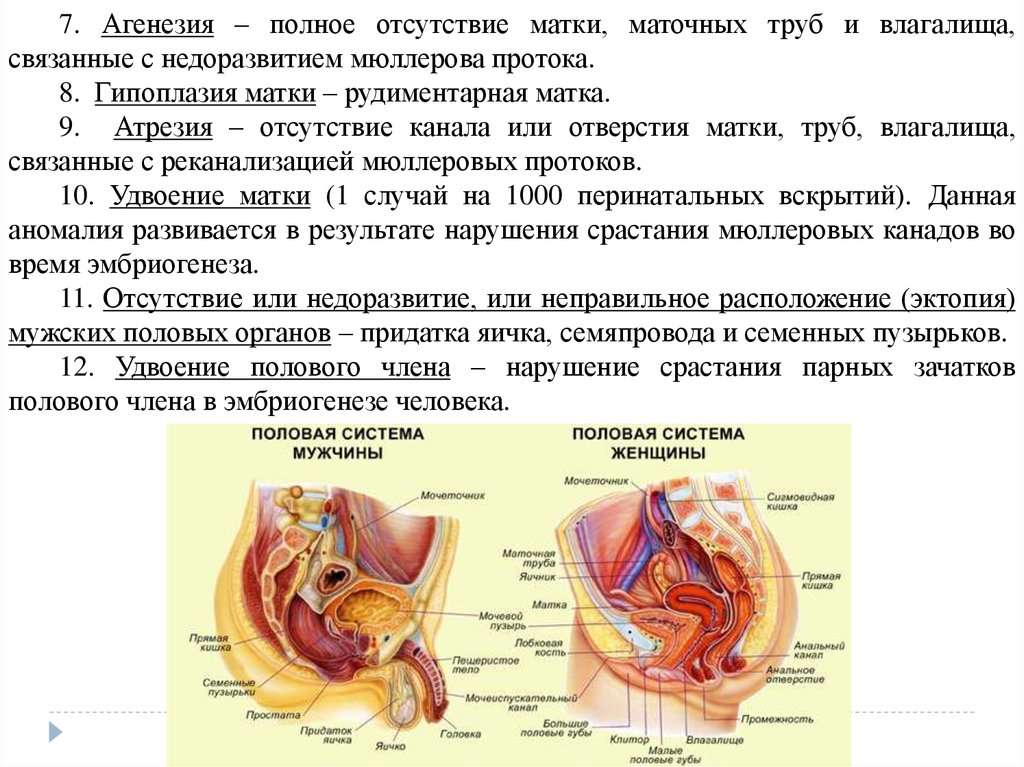
7.
Проблема связи между эмбриональным и историческим развитиеморганизмов снова была выдвинута современником Ч.Дарвина
немецким
зоологом
Ф.Мюллером
(1864),
который
дал
формулировку закона рекапитуляции, т.е. повторения древних
признаков предков в онтогенезе потомков.
Э.Геккель развил закон рекапитуляции Ф.Мюллера, дав ему
название
биогенетического
закона.
Он
сформулировал
биогенетический закон следующим образом: «Онтогенез есть
краткое и сжатое повторение филогенеза», т.е. развитие
зародыша - это сжатое и краткое повторение всех форм
возникающих из одного корня.
8.
Признаки предков, которые повторяются у зародыша в эмбриональном периоде,Геккель назвал палингенезами. У взрослого организма они исчезают. Само
явление повторения древних признаков предков в онтогенезе потомков он
назвал рекапитуляцией. В ходе эмбрионального развития отмечается явление
гетерохронии. Это сдвиги по времени закладки органов. Например, нервная
трубка у высших позвоночных (человека) закладывается раньше, чем у низших
позвоночных (рыб). Э.Геккелем также было
отмечено такое явление в
эмбриогенезе, как гетеротопия. Это изменение в процессе эволюции места
эмбриональной закладки того или иного органа. Например, закладка половых
желез у высших животных
осуществляется в мезодерме, а у низших
многоклеточных животных их закладка имеет место в экто- или энтодерме.
Таким образом, гетеротопии и гетерохронии - это пути эволюционных перестроек
онтогенеза. Итак, онтогенез и филогенез взаимообусловлены.
Биогенетический закон, выдвинутый Геккелем и Мюллером, казалось,
полностью объясняет соотношение онтогенеза и филогенеза. Однако ошибка
Э.Геккеля состояла в том, что он рассматривал эмбриональное развитие в отрыве
от развития всего организма. По Геккелю, эмбрион имеет свою историю, а
взрослая особь - свою. Т.е. палингенезы появляются и исчезают и у взрослого
организма не имеют значения для развития.
9.
Когда же ученые занялись изучением этого вопроса более детально и глубоко,выяснилось, что палингенезы имеют большое значение для формообразования.
Действительно, хорда в эмбриональном периоде у человека есть, а потом у
взрослого организма исчезает. Однако хорда, как оказалось, индуцирует
развитие нервной трубки, т.е. хорда выполняет формообразующую роль.
Биогенетический закон стал подвергаться критике еще и потому, что,
согласно Э. Геккелю, в онтогенезе организма повторяются целые
филогенетические стадии. Однако более поздние исследования онтогенеза
организмов многих видов показали, что рекапитуляция целых филогенетических
стадий невозможна. Рекапитулируют лишь отдельные признаки и процессы.
Возникла
необходимость
переосмысления
и
трансформации
предложенного Геккелем и Мюллером биогенетического закона. Это сделал
российский ученый, биолог и теоретик эволюционного учения Алексей Николаевич Северцов. За период с 1908 по 1934 год А. Н. Северцов со
своими сотрудниками провел обширные исследования развития различных частей
скелета позвоночных животных. Он убедился, что повторение признаков предков
в онтогенезе происходит достаточно часто и закономерно. Однако выяснилось,
что имеется много случаев, когда рекапитуляция ненаблюдаеся.
10.
В итоге А.Н. Северцов разработал теорию филэмбриогенезов, которая не толькообъясняет причины повторения или не повторения тех или иных признаков в
онтогенезе, но и раскрывает роль изменений онтогенеза в филогенезе животных.
Термином «филэмбриогенезы» А.Н. Северцов обозначил такие изменения
индивидуального развития, которые имеют филогенетическое значение, т.е.
отражаются на последующем историческом развитии потомков. Таким образом,
Северцов внес новый смысл в трактовку биогенетического закона. Онтогенез не
только следствие филогенеза, но и активное начало филогенеза. Основная мысль
теории филэмбриогенеза, по словам А.Н. Северцова, заключается в том, что
эволюционные изменения состоят в изменении хода онтогенетического развития.
И так, филэмбриогенезы по А.Н. Северцову это:
•Изменения, возникающие в процессе эмбрионального развития,
•Изменения, сохраняющиеся во взрослом состоянии,
•Изменения, наследующиеся потомками.
А.Н. Северцов обнаружил, что новые признаки, изменяющие
направления эволюции, могут появляться у эмбрионов на различных стадиях
развития: ранних, средних и поздних. От этого и зависит полнота повторения
признаков предков - рекапитуляция. Он выделил 3 основных способа изменения
онтогенеза:
архаллаксис,
девиация и
анаболия.
11.
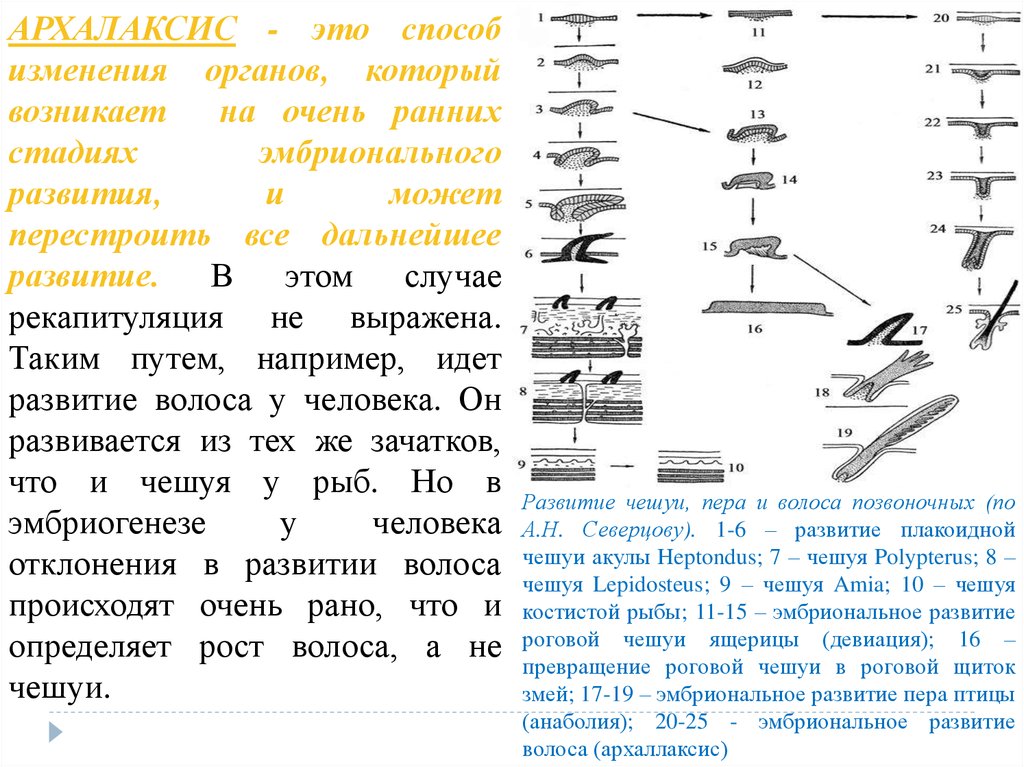
АРХАЛАКСИС - это способизменения органов, который
возникает
на очень ранних
стадиях
эмбрионального
развития,
и
может
перестроить все дальнейшее
развитие.
В этом
случае
рекапитуляция не выражена.
Таким путем, например, идет
развитие волоса у человека. Он
развивается из тех же зачатков,
что и чешуя у рыб. Но в
эмбриогенезе
у
человека
отклонения в развитии волоса
происходят очень рано, что и
определяет рост волоса, а не
чешуи.
Развитие чешуи, пера и волоса позвоночных (по
А.Н. Северцову). 1-6 – развитие плакоидной
чешуи акулы Heptondus; 7 – чешуя Polypterus; 8 –
чешуя Lepidosteus; 9 – чешуя Amia; 10 – чешуя
костистой рыбы; 11-15 – эмбриональное развитие
роговой чешуи ящерицы (девиация); 16 –
превращение роговой чешуи в роговой щиток
змей; 17-19 – эмбриональное развитие пера птицы
(анаболия); 20-25 - эмбриональное развитие
волоса (архаллаксис)
12.
13.
ДЕВИАЦИЯ - это отклонения в развитии органа на среднихстадиях эмбриогенеза. Примером девиации может служить
онтогенез роговых чешуй у рептилий. Первые стадии этого
процесса напоминают начало развития плакоидных чешуй акуловых
рыб. Рекапитуляция в этом примере наблюдается только на ранних
стадиях развития чешуи, а потом ее развитие у рептилий
отклоняется от пути развития чешуи рыб.
14.
АНАБОЛИЯ - это добавление новой стадии в конце морфогенеза какоголибо органа с соответствующим удлинением его онтогенетическогоразвития. До возникновения анаболии орган развивается так же, как и у
предка, т.е. имеет место рекапитуляция и проявление биогенетического
закона. Таким путем развиваются почки высших позвоночных, проходя
стадии предпочки, первичной и вторичной почек. Череп у птиц
закладывается как и у других позвоночных, но позже они срастаются, так
что швы между ними становятся незаметными. Путем анаболии идет
развитие пера у птиц.
И так, исходные стадии, при которых начинается эволюция органа,
различны при разных способах филэмбриогенеза. При архаллаксисе
зачатки органов возникают из совершенно индифферентной
эмбриональной ткани. Девиация возникает на стадии относительно
индифферентной эмбриональной ткани. При анаболии эволюция
органа начинается, когда основные черты животного уже
установились.
Филогенетически новообразования происходят именно путем
архаллаксиса. Архаллаксис - это наиболее быстрый способ эволюции.
Девиация же в этом отношении занимает промежуточное положение
между анаболией и архаллаксисом.
15.
Биологический прогресс и пути его достиженияА.Н.Северцов различал в природе два направления в эволюции:
биологический прогресс и биологический регресс.
БИОЛОГИЧЕСКИЙ ПРОГРЕСС - проявляется в том, что та или иная
группа животных быстро размножается, расселяется и захватывает новые ареалы,
распадаясь при этом на мелкие систематические подразделения. Непрерывно
возрастающее число особей данного вида служит важным признаком
биологически прогрессивной эволюции этого вида. И так, биологический
прогресс характеризуется тремя признаками:
•Численное увеличение особей данной систематической группы;
•Прогрессивное расселение и захват новых ареалов;
•Разделение
группы
на
соподчиненные
систематические
единицы
(разновидности, подвиды, виды и т.д.).
БИОЛОГИЧЕСКИ РЕГРЕССИВНАЯ ЭВОЛЮЦИЯ - представляет собой
противоположный процесс прогрессу. Она отличается тем, что у регрессирующей
группы число особей, составляющих вид, постепенно падает. Ареал постепенно
сужается. Иногда он принимает вид отдельных островков, разделенных большим
пространством. Число подчиненных таксономических групп постепенно
уменьшается. Все эти признаки характеризуют постепенное вымирание
животных.
16.
А.Н.Северцов пришел к выводу, что существуют следующие пути прогресса:•ароморфоз (морфофизиологический прогресс);
•идиоадаптация (морфофизиологическое приспособление);
•морфофизиологическая дегенерация;
•ценогенезы.
17.
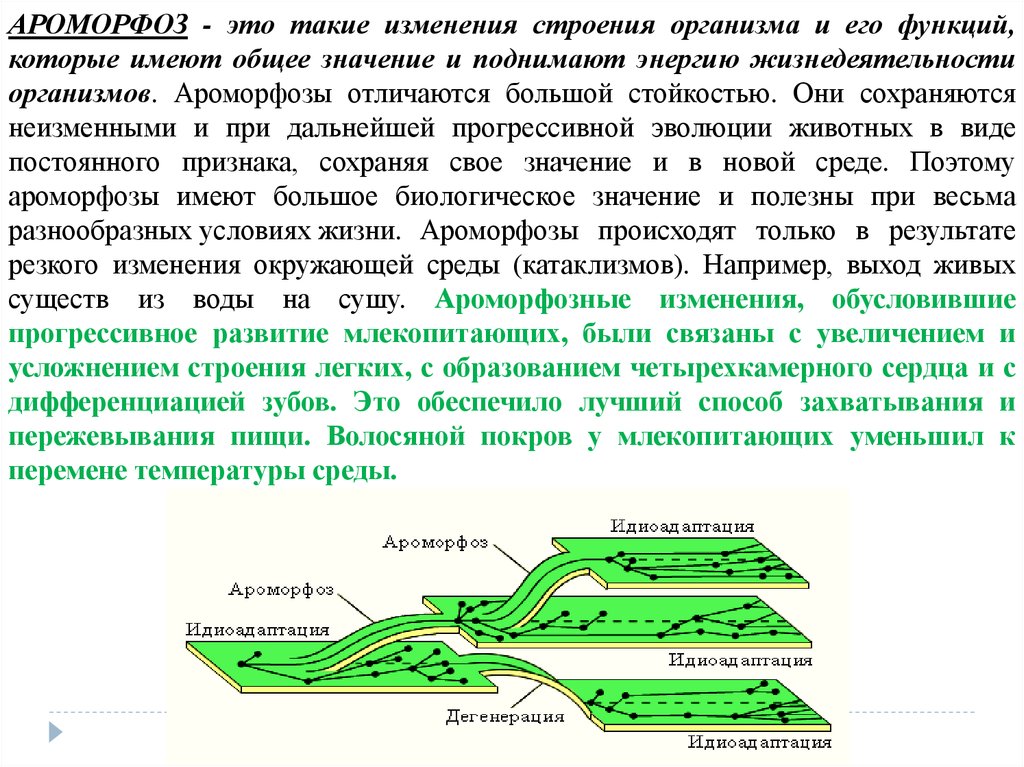
АРОМОРФОЗ - это такие изменения строения организма и его функций,которые имеют общее значение и поднимают энергию жизнедеятельности
организмов. Ароморфозы отличаются большой стойкостью. Они сохраняются
неизменными и при дальнейшей прогрессивной эволюции животных в виде
постоянного признака, сохраняя свое значение и в новой среде. Поэтому
ароморфозы имеют большое биологическое значение и полезны при весьма
разнообразных условиях жизни. Ароморфозы происходят только в результате
резкого изменения окружающей среды (катаклизмов). Например, выход живых
существ из воды на сушу. Ароморфозные изменения, обусловившие
прогрессивное развитие млекопитающих, были связаны с увеличением и
усложнением строения легких, с образованием четырехкамерного сердца и с
дифференциацией зубов. Это обеспечило лучший способ захватывания и
пережевывания пищи. Волосяной покров у млекопитающих уменьшил к
перемене температуры среды.
18.
ИДИОАДАПТАЦИИ - частные приспособления организма к определенномуобразу жизни в конкретных условиях внешней среды. Идиоадаптации не
сказываются существенно на общем уровне организации данной группы в
отличие от ароморфозов. Они обеспечивают адаптивную радиацию в пределах
одного уровня организации и бывают специфическими признаками видов, родов,
семейств. Т.е. при идиоадаптации нет повышения организации, а все сводится к
лучшему приспособлению к условиям среды в пределах прежнего, уже
существующего уровня организации.
Например, разные типы клювов у птиц в связи с использованием
различной пищи и способов ее добывания. К идиоадаптациям относятся
различные изменения окраски животных, включая случаи покровительственной и
защитной окраски. К ним также относятся и случаи мимикрии, связанные с
изменением не только окраски, но и формы тела. Например, бабочки,
иммитирующие листья растений, плоская форма тела камбалы и т.п. случаи.
Частным случаем идиоадаптации является специализация. Ее примерами
служат животные, приспособившиеся к жизни в почве, пещерах, на отвесных
скалах.
А.Н.Северцов считал, что идиоадаптации являются
самым
распространенным способом эволюции. Таким путем происходила эволюция
внутри каждого класса животного мира.
19.
20.
21.
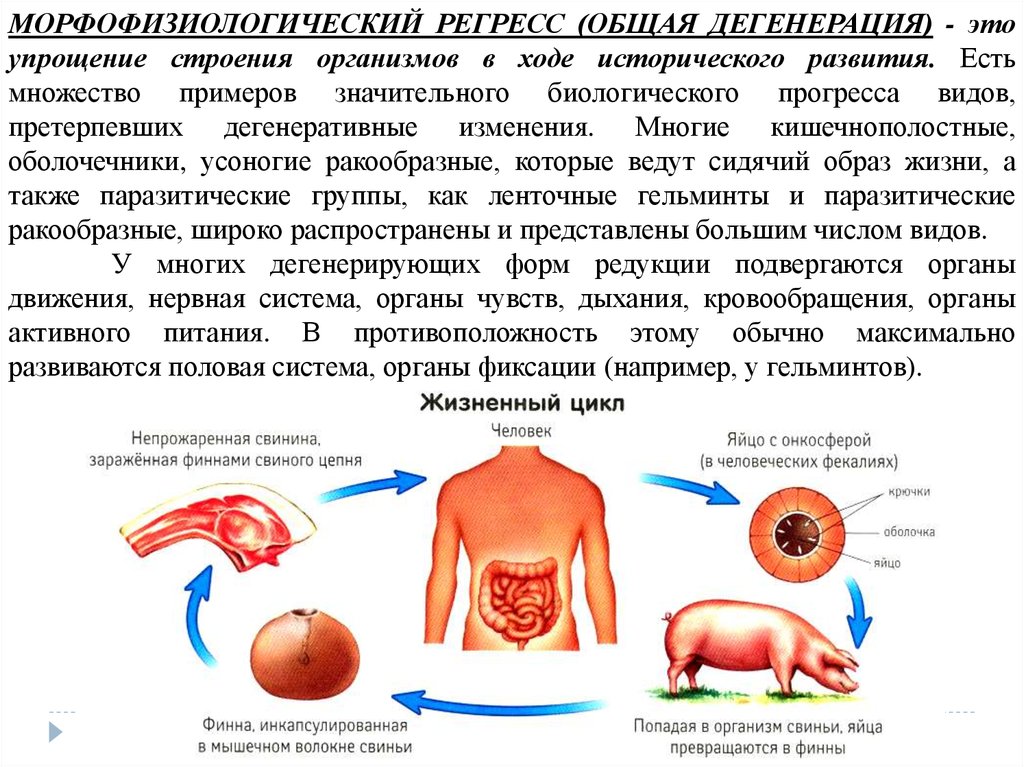
МОРФОФИЗИОЛОГИЧЕСКИЙ РЕГРЕСС (ОБЩАЯ ДЕГЕНЕРАЦИЯ) - этоупрощение строения организмов в ходе исторического развития. Есть
множество примеров значительного биологического прогресса видов,
претерпевших дегенеративные изменения. Многие кишечнополостные,
оболочечники, усоногие ракообразные, которые ведут сидячий образ жизни, а
также паразитические группы, как ленточные гельминты и паразитические
ракообразные, широко распространены и представлены большим числом видов.
У многих дегенерирующих форм редукции подвергаются органы
движения, нервная система, органы чувств, дыхания, кровообращения, органы
активного питания. В противоположность этому обычно максимально
развиваются половая система, органы фиксации (например, у гельминтов).
22.
Одно из самых необычных растений на нашей планете - африканская гиднора. Ее-то ирастением трудно назвать, уж очень она на гриб похожа. Кстати, из-за своей схожести с
грибами растение и получило свое название - Hydnora, что с греческого означает "гриб".
Гиднора распространена на востоке африканского континента и острове Мадагаскар, ведет
паразитический образ жизни, присасываясь к корням многих тропических деревьев.
Внешний вид ее очень своеобразный - лепестки соцветия толстые и мясистые, яркооранжевого цвета с внутренней стороны, бурая потрескавшаяся оболочка покрывает
растение внешне, края толстых лепестков покрыты многочисленными щитинками.
Большую часть своей жизни гиднора проводит под землей, и выходит на поверхность
после
того
как
выпадет
достаточное
количество
осадков.
23.
Клещи. Верхний ряд – акариевые клещи, слева направо: чесоточный клещ,почвенный клещ, двупятнистый паутинный клещ, водяной клещ. Нижний ряд,
слева направо: акариевые клещи (зерновой клещ), паразитические клещи
(таёжный клещ, отобиус, клещ-краснотелка)
24.
ЦЕНОГЕНЕЗЫ - это эмбриональные и личиночные приспособления, которыенепосредственно полезны развивающемуся организму, а затем исчезают.
Биологический прогресс видов происходит, таким образом, также и путем
возникновения различных полезных приспособлений, которые обеспечивают
выживание и развитие организмов в эмбриональном и личиночном периоде их
жизни.
А.Н.Северцов подразделил ценогенезы на 3 группы, взяв за основу
классификации функциональный принцип и принимая во внимание 3 основные
функции:
•защиту зародышей и личинок от врагов;
•питание эмбрионов;
•дыхание эмбрионов и личинок.
К группе защитных ценогенезов относятся приспособительные окраски
зародышей, пигментация яиц и личинок амфибий, прозрачность яиц и личинок
некоторых водных животных, зеленая окраска живущих на листьях личинок
насекомых.
К приспособлениям, обеспечивающих питание, относятся запас желтка в
яйцах, роговые челюсти у головастика и личинок других амфибий.
К эмбриональным и личиночным органам дыхания относятся жаберные
нити у личинок амфибий, сосудистые сплетения у эмбрионов костистых рыб,
сосуды аллантоиса рептилий, плацента у млекопитающих.
25.
Таким образом, ценогенотипические приспособления увеличивают числозародышей и личинок доживающих до взрослого состояния и тем самым
повышают численность особей данного вида. Ценогенезы тоже являются одним
из способов достижения биологического прогресса.
Рассматривая всю филогенетическую историю животного мира,
А.Н.Северцов пришел к заключению, что она шла путем чередования различных
способов достижения биологического прогресса.
26.
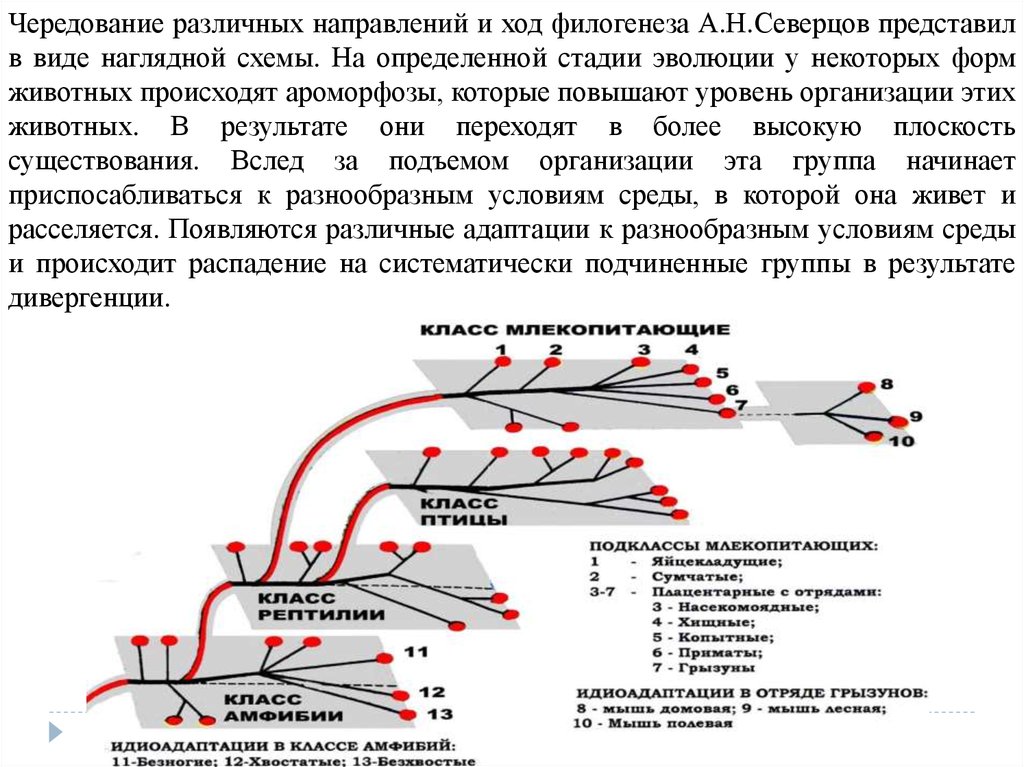
Чередование различных направлений и ход филогенеза А.Н.Северцов представилв виде наглядной схемы. На определенной стадии эволюции у некоторых форм
животных происходят ароморфозы, которые повышают уровень организации этих
животных. В результате они переходят в более высокую плоскость
существования. Вслед за подъемом организации эта группа начинает
приспосабливаться к разнообразным условиям среды, в которой она живет и
расселяется. Появляются различные адаптации к разнообразным условиям среды
и происходит распадение на систематически подчиненные группы в результате
дивергенции.
27.
Среда обитания и внешнее строение ланцетникаВо многих морях тропической и умеренной зон, в том числе и в Черном море, на
небольшой глубине в местах с чистым песчаным дном живут маленькие (длиной
4-8 см) полупрозрачные животные - ланцетники. Уплощенное с боков розоватое
тело ланцетника заострено на переднем и заднем концах. Хвостовой отдел
обрамлен кожной складкой - хвостовым плавником, напоминающим по форме
обоюдоострый хирургический инструмент - ланцет (отсюда название ланцетник). Большую часть времени ланцетник проводит зарывшись в песок и
выставив наружу передний конец тела, на котором находится рот, окруженный 1020 парами щупалец. Будучи потревоженным, ланцетник переплывает на
небольшое расстояние и снова зарывается в грунт.
28.
Хорда. Хордовые животныеВдоль спинной стороны тела ланцетника тянется хорда - плотный упругий стержень,
состоящий из особых тесно прилегающих друг к другу клеток. Хорда выполняет роль
внутреннего скелета. Она служит опорой внутренним органам и придает телу ланцетника
известную упругость, что имеет большое значение при зарывании в грунт. У ланцетника
хорда сохраняется в течение всей жизни.
Ланцетники лишены черепа, так как у них нет головного мозга. Поэтому их относят к
группе бесчерепных. Рыбы, земноводные, пресмыкающиеся, птицы и млекопитающие
обладают хордой только в личиночном состоянии или во время зародышевого развития. У
взрослых она заменяется хрящевым (хрящевые рыбы) или костным позвоночником,
состоящим из отдельных позвонков. Поэтому таких животных объединяют в группу
позвоночных.
Мышцы ланцетника
Мышцы ланцетника в виде двух лент расположены по сторонам от хорды. Ланцетник
может совершать довольно однообразные движения. Изгибая тело то в одну, то в другую
сторону, он плавает и зарывается в грунт.
Пищеварительная система ланцетника
На переднем конце тела находится окруженное предротовой воронкой со щупальцами
ротовое отверстие. С током воды мелкие планктонные организмы или пищевые частицы
через рот попадают в обширную глотку, прилипают к ее внутренней поверхности,
склеиваются слизью в комочки и направляются в кишечник, где перевариваются. Вода
проходит через жаберные щели в стенке глотки. Непереваренные остатки пищи удаляются
наружу через анальное отверстие на брюшке у начала хвостового плавника.
29.
Органы дыхания ланцетникаОрганы дыхания ланцетника - это жабры, с помощью которых он дышит
растворенным в воде кислородом. Дыхание осуществляется одновременно с
питанием. По бокам глотка прорезана множеством отверстий - жаберными
щелями. Ткани глотки вокруг жаберных щелей окружены сетью капилляров, в
которых совершается газообмен крови и воды. Из воды в кровь переходит
кислород, а из крови в воду - углекислый газ. Снаружи жаберные щели не видны,
так как они прикрыты кожными складками, защищающими их от попадания
песчинок.
Кровеносная система ланцетника
Кровеносная система замкнутая и состоит из двух главных кровеносных сосудов
— спинного и брюшного. По брюшному сосуду кровь, насыщенная углекислым
газом, течет вперед к жаберным капиллярам. Здесь она отдает углекислый газ,
обогащается кислородом и оттекает по брюшному сосуду. Сердца у ланцетника
нет, кровь движется по системе благодаря сокращениям стенок нескольких
кровеносных сосудов.
Органы выделения ланцетника
Органы выделения ланцетника имеют такое же строение, как у кольчатых червей,
но они расположены вдоль глотки и открываются в околожаберную полость.
30.
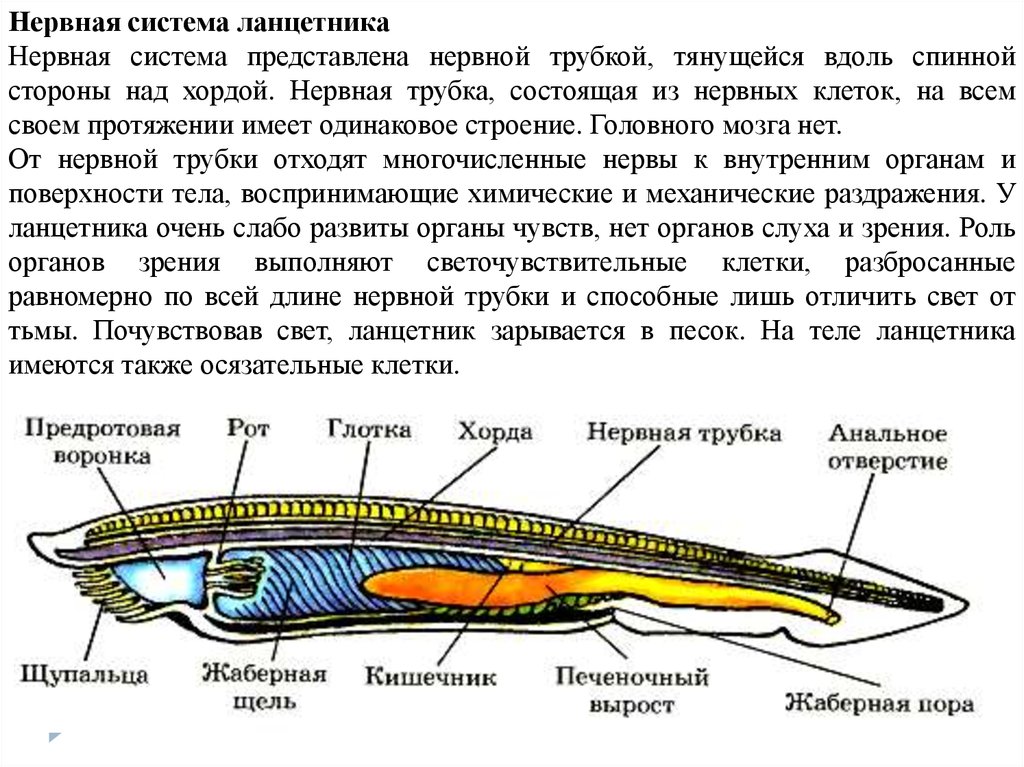
Нервная система ланцетникаНервная система представлена нервной трубкой, тянущейся вдоль спинной
стороны над хордой. Нервная трубка, состоящая из нервных клеток, на всем
своем протяжении имеет одинаковое строение. Головного мозга нет.
От нервной трубки отходят многочисленные нервы к внутренним органам и
поверхности тела, воспринимающие химические и механические раздражения. У
ланцетника очень слабо развиты органы чувств, нет органов слуха и зрения. Роль
органов зрения выполняют светочувствительные клетки, разбросанные
равномерно по всей длине нервной трубки и способные лишь отличить свет от
тьмы. Почувствовав свет, ланцетник зарывается в песок. На теле ланцетника
имеются также осязательные клетки.
31.
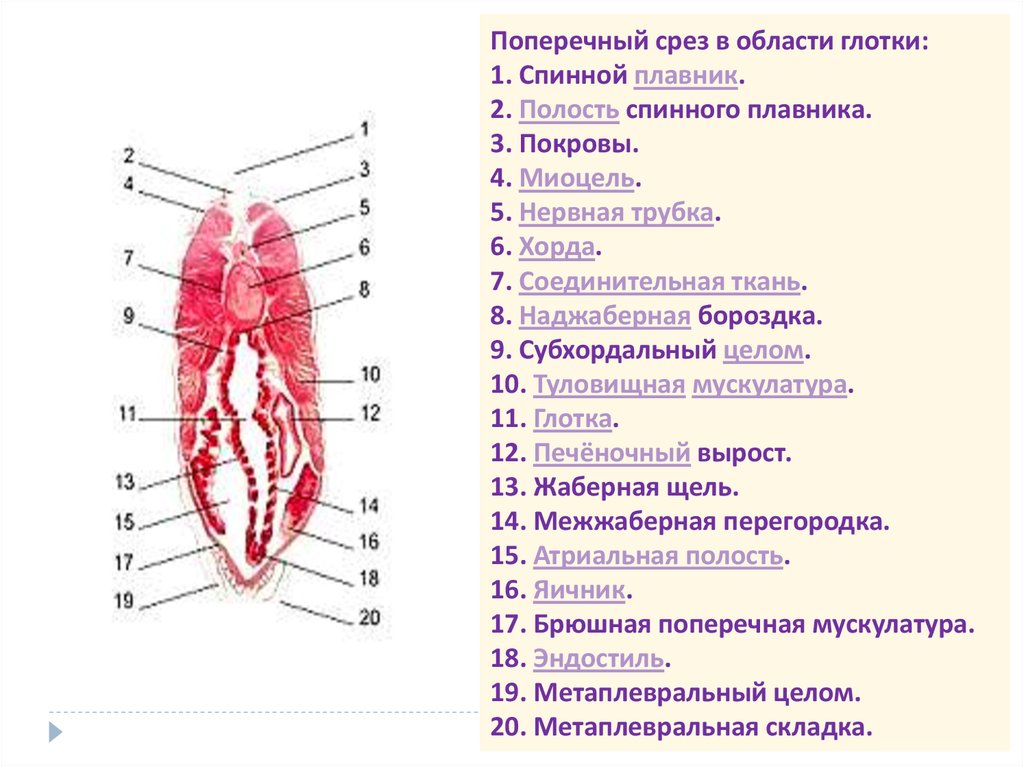
Поперечный срез в области глотки:1. Спинной плавник.
2. Полость спинного плавника.
3. Покровы.
4. Миоцель.
5. Нервная трубка.
6. Хорда.
7. Соединительная ткань.
8. Наджаберная бороздка.
9. Субхордальный целом.
10. Туловищная мускулатура.
11. Глотка.
12. Печёночный вырост.
13. Жаберная щель.
14. Межжаберная перегородка.
15. Атриальная полость.
16. Яичник.
17. Брюшная поперечная мускулатура.
18. Эндостиль.
19. Метаплевральный целом.
20. Метаплевральная складка.
32.
ФИЛОГЕНЕЗ ПОКРОВОВ ТЕЛА ХОРДОВЫХКожные покровы хордовых имеют эктодермальное (эпидермис) и
мезодермальное (дерма) происхождение. В связи с разной средой
обитания и образом жизни кожные покровы хордовых очень
разнообразны по строению и отличаются многообразием функций.
Функции кожи:
•защищает организм от вредных воздействий факторов внешней
среды
(механических,
химических,
проникновения
микроорганизмов);
•участвует в терморегуляции;
•является органом осязания (осязательные, температурные и
болевые рецепторы);
•депо крови;
• участвует в выделении продуктов диссимиляции (мочевины, солей
и др.);
•у некоторых животных участвует в дыхании.
В связи с разной средой обитания и образом жизни, кожа и ее
производные у хордовых разнообразны по строению и функциям.
33.
Основные направления эволюции покровов хордовых:1)дифференцировка на два слоя: наружный – эпидермис, внутренний – дерму и
увеличение толщины дермы;
1)от однослойного эпидермиса к многослойному;
2)дифференцировка дермы на 2 слоя – сосочковый и сетчатый;
3)появление подкожно-жировой клетчатки и совершенствование механизмов
терморегуляции;
4)от одноклеточных желез к многоклеточным;
6)дифференцировка различных производных кожи.
У низших хордовых (ланцетник) эпидермис однослойный, цилиндрический,
имеет железистые клетки, выделяющие слизь. Дерма (кориум) представлена тонким
слоем неоформленной соединительной ткани.
У низших позвоночных эпидермис становится многослойным. Нижний его
слой – ростковый (базальный), клетки его делятся и восполняют клетки
вышележащих слоев. Дерма имеет правильно расположенные волокна, сосуды и
нервы.
Производными кожи являются: одноклеточные (у круглоротых, рыб) и
многоклеточные (у земноводных) слизистые железы; чешуя: а) плакоидная у
хрящевых рыб, в развитии которой принимают участие эпидермис и дерма; б)
костная у костных рыб, которая развивается за счет дермы.
Плакоидная чешуя снаружи покрыта слоем эмали (эктодермального происхождения),
под которым находятся дентин и пульпа (мезодермального происхождения). Чешуя и
слизь выполняют защитную функцию.
34.
35.
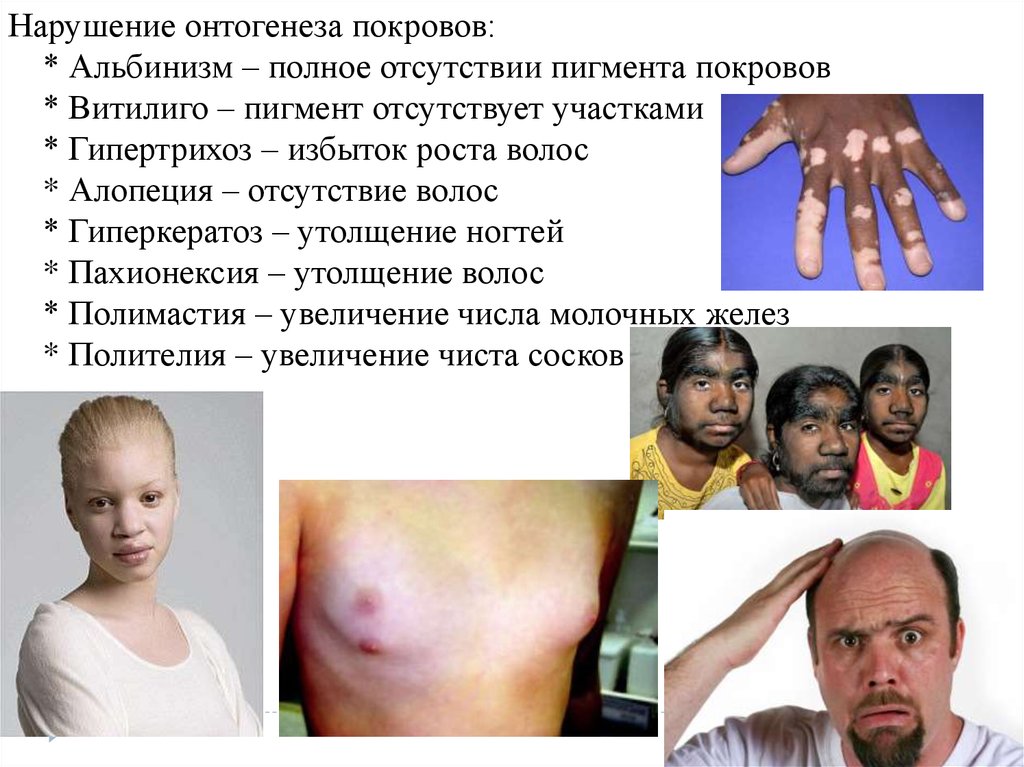
Нарушение онтогенеза покровов:* Альбинизм – полное отсутствии пигмента покровов
* Витилиго – пигмент отсутствует участками
* Гипертрихоз – избыток роста волос
* Алопеция – отсутствие волос
* Гиперкератоз – утолщение ногтей
* Пахионексия – утолщение волос
* Полимастия – увеличение числа молочных желез
* Полителия – увеличение чиста сосков
36.
Происхождение и функции скелета животных.Опорные
образования
у
беспозвоночных
животных,
представляющие им постоянную форму тела очень разнообразны.
Они имеют экто-, энто- и мезодермальное происхождение. У
позвоночных животных скелет имеет в основном мезодермальное
происхождение.
Скелет в организме животного выполняет разнообразные функции:
обеспечение постоянной формы тела;
пассивная часть опорно-двигательного аппарата;
защита от механических и других воздействий;
кроветворная функция.
37.
Осевой скелет в процессе эволюции претерпел большиеизменения.
У низших хордовых осевым скелетом является хорда, а у
высших она постепенно заменяется развивающимися позвонками. В
позвонках различают тело, верхние и нижние дуги.
Так у круглоротых хорда сохраняется в течении всей жизни, но
появляются закладки позвонков, представляющие собой небольшие
хрящевые образования, метамерно располагающиеся над хордой.
Они называются верхними дугами.
У примитивных рыб кроме верхних дуг появляются нижние
дуги, а у высших рыб и тела позвонков. Тела позвонков у
большинства рыб и вышестоящих животных формируется из ткани,
окружающей хорду, а также из оснований дуг. С телами позвонков
срастаются верхние и нижние дуги. Концы верхних дуг срастаются
между собой, образуя спинномозговой канал. На нижних дугах
появляются отростки, к которым прикрепляются ребра. У рыб два
отдела позвоночника – туловищный и хвостовой. Остатки хорды у
рыб сохраняются между телами позвонков.
38.
У земноводных на ранних стадиях развития хорда заменяетсяпозвоночником. В позвоночнике уже четыре отдела: шейный, грудной,
крестцовый и хвостовой. В шейном отделе всего один позвонок, грудной
состоит из пяти позвонков. К грудным позвонкам прикрепляются
небольшие ребра, заканчивающиеся свободно. Крестцовый отдел, как и
шейный, включает один позвонок, который является опорой для костей
таза и задних конечностей. Хвостовой отдел у бесхвостых амфибий
срастается в одну кость, а у хвостатых состоит из большого числа
позвонков.
У рептилий пять отделов в позвоночнике: шейный, грудной,
поясничный крестцовый и хвостовой. В шейном отделе у различных
видов пресмыкающихся разное число позвонков, но самое большое
восемь. Первый позвонок называется атлантом и имеет форму кольца, а
второй – эпистрофеем и имеет зубовидный отросток, на котором
вращается первый позвонок. В грудном отделе число позвонков не
постоянно, к ним прикрепляются ребра, большинство из которых
соединяется с грудиной, впервые образуя у высших животных грудную
клетку. В грудопоясничном отделе всего 22 позвонка, а в крестцовом два.
К позвонкам поясничного и крестцового отделов также прикрепляются
ребра. В хвостовом отделе число позвонков варьирует.
39.
У птиц позвоночный столб сходен с позвоночником рептилий,но обладает определенной специализацией в связи хвостовой.
Шейный отдел включает до 25-х позвонков, что обеспечивает
большую подвижность.
У млекопитающих в позвоночнике пять отделов: шейный
грудной, поясничный, крестцовый и хвостовой. В шейном отделе
семь позвонков, в грудном непостоянное число позвонков (от 9 до
24, но чаще 12-13). К грудным позвонкам присоединяются ребра,
большие количество из которых соединяется с грудиной.
Поясничный отдел включает от трех до девяти позвонков.
Крестцовые позвонки срослись, образовав крестец, а хвостовой
отдел позвоночника включает разное количество позвонков у
различных видов млекопитающих.
40.
41.
Аномалии и пороки развития скелета у человека.•Наличие ребер у нижнего шейного или у первых поясничных позвонков. В соответствии с
эволюцией позвоночных у человека во время эмбрионального развития во всех отделах
позвоночника закладываются ребра, но впоследствии они сохраняются только в грудном
отделе, а в других отделах ребра редуцируются. Но иногда у человека наблюдаются
подобные атавизмы.
•Наличие хвостовых позвонков. Во время эмбриогенеза у человека, как и позвоночных
животных, закладывается 8-11 хвостовых позвонков, затем они редуцируются и остается 45 недоразвитых позвонков, образуя копчик. Иногда проявляются атавистические признаки
в виде присутствия хвостового отдела позвоночника.
•Spina bifida – это нередкая аномалия, которая возникает при нарушении срастания
верхних дуг позвонков. Она чаще проявляется в пояснично-крестцовой области
позвоночника и в зависимости от глубины и протяженности расщепления может иметь
разную степень тяжести.
42.
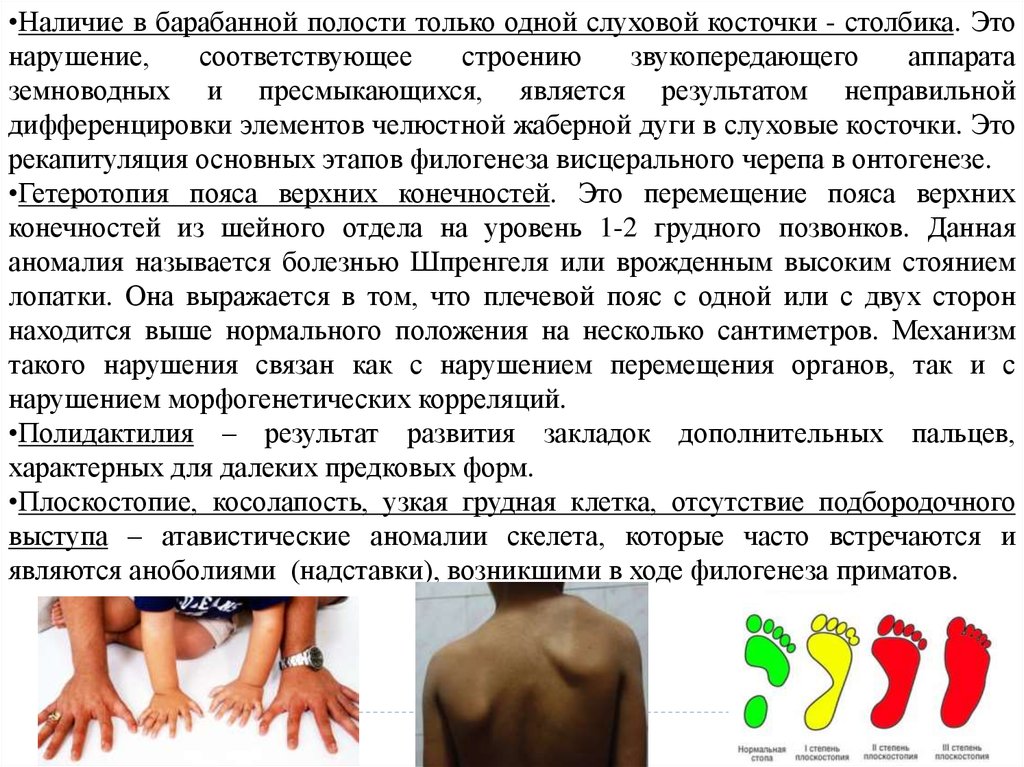
•Наличие в барабанной полости только одной слуховой косточки - столбика. Этонарушение,
соответствующее
строению
звукопередающего
аппарата
земноводных и пресмыкающихся, является результатом неправильной
дифференцировки элементов челюстной жаберной дуги в слуховые косточки. Это
рекапитуляция основных этапов филогенеза висцерального черепа в онтогенезе.
•Гетеротопия пояса верхних конечностей. Это перемещение пояса верхних
конечностей из шейного отдела на уровень 1-2 грудного позвонков. Данная
аномалия называется болезнью Шпренгеля или врожденным высоким стоянием
лопатки. Она выражается в том, что плечевой пояс с одной или с двух сторон
находится выше нормального положения на несколько сантиметров. Механизм
такого нарушения связан как с нарушением перемещения органов, так и с
нарушением морфогенетических корреляций.
•Полидактилия – результат развития закладок дополнительных пальцев,
характерных для далеких предковых форм.
•Плоскостопие, косолапость, узкая грудная клетка, отсутствие подбородочного
выступа – атавистические аномалии скелета, которые часто встречаются и
являются аноболиями (надставки), возникшими в ходе филогенеза приматов.
43.
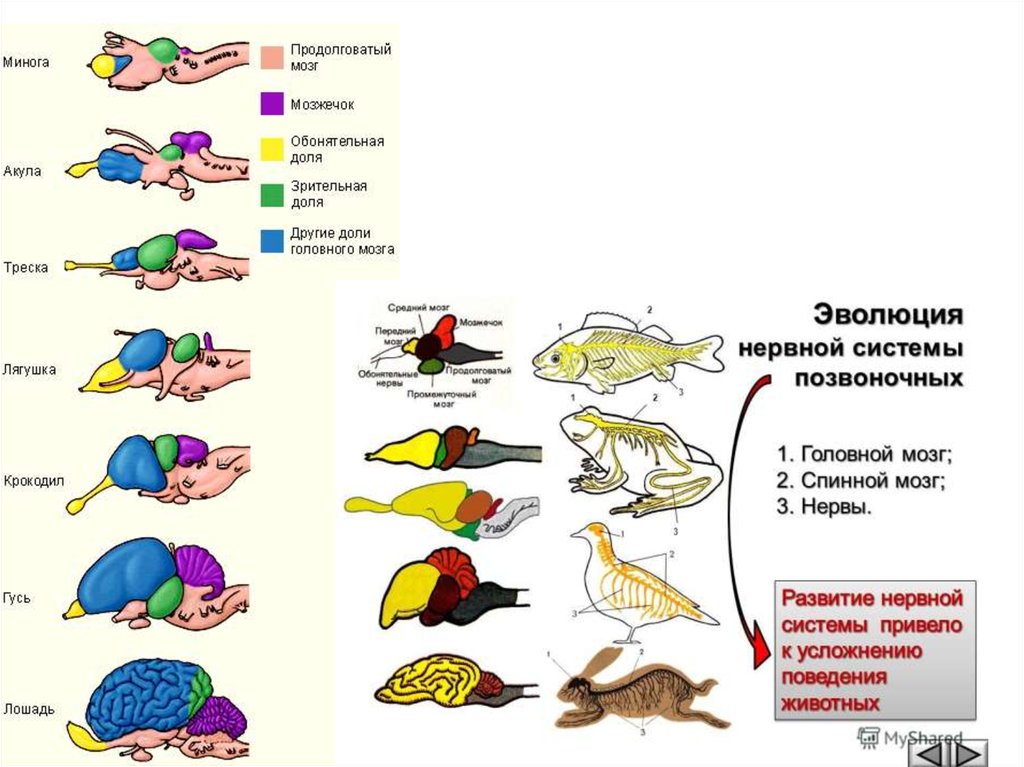
Эволюция нервной системы у хордовых животных.Нервная система у хордовых представлена нервной трубкой,
которая дифференцируется на головной и спинной мозг.
У низших хордовых нервная трубка имеет вид полой трубки
(невроцель) с отходящими от трубки нервами. У ланцетника в
головном отделе образуется небольшое расширение – зачаток
головного мозга. Это расширение получило название желудочка.
У высших хордовых на переднем конце нервной трубки
образуется три вздутия: передний, средний и задний пузыри. Из
первого мозгового пузыря образуется в дальнейшем передний и
промежуточный мозг, из среднего – средний, из заднего – мозжечок
и продолговатый мозг, переходящий в спинной.
У всех классов позвоночных животных мозг состоит из 5
отделов (передний, промежуточный, средний, задний и
продолговатый), но степень их развития неодинакова у животных
разных классов.
44.
Укруглоротых все отделы головного мозга расположены друг за другом вгоризонтальной плоскости. Продолговатый мозг непосредственно переходит в
спинной с центральным каналом в нутрии.
У рыб головной мозг более дифференцирован по сравнению с круглоротыми.
Объем переднего мозга увеличен, особенно у двоякодышащих рыб, но передний
мозг еще не разделен на полушария и функционально служит высшим
обонятельным центром. Крыша переднего мозга тонкая, она состоит только из
эпителиальных клеток и не содержит нервной ткани. В промежуточном мозге, с
которым связан эпифиз и гипофиз, расположен гипоталамус, являющийся
центром эндокринной системы. Наиболее развитым у рыб является средний мозг.
Хорошо выражены в нем зрительные доли. В области среднего мозга имеется
изгиб, характерный для всех вышестоящих позвоночных. Кроме того средний
мозг является анализирующим центром. Мозжечок, входящий в состав заднего
мозга, развит хорошо в связи со сложностью движения у рыб. Он представляет
собой центр координации движения, его размер варьирует в зависимости от
активности движения разных видов рыб. Продолговатый мозг обеспечивает связь
высших отделов головной мозга со спинным и содержит центры дыхания и
кровообращения.
Из головного мозга рыб выходит 10 пар черепно-мозговых нервов.
Такой тип мозга, в котором высшим центром интеграции является средний
мозг, называется ихтиопсидным.
45.
У амфибий нервная система по своему строению близка к нервной системедвоякодышащих рыб, но отличается значительным развитием и полным
разделением парных вытянутых
полушарий, а также слабым развитием
мозжечка, что обусловлено малой подвижностью амфибий и однообразием их
движений. Но у амфибий появилась крыша переднего мозга, называемая
первичным мозговым сводом – архипаллиумом. Число черепно-мозговых нервов,
как и у рыб, десять. И тип мозга тот же, т.е. ихтиопсидный.
Таким образом у всех анамний (круглоротых, рыб и амфибий) ихтиопсидный
тип головного мозга.
В строении головного мозга рептилий, относящихся к высшим позвоночным,
т.е. к амниотам, отчетливо выражены черты прогрессивной организации.
Значительное преобладание над другими отделами мозга получают полушария
переднего мозга. У их основания расположены крупные скопления нервных
клеток – полосатые тела. На латеральной и медиальной сторонах каждого
полушария появляются островки старой коры – архикортекс. Размеры среднего
мозга сокращаются, и он теряет значение ведущего центра. Анализирующим
центром становится дно переднего мозга, т.е. полосатые тела. Такой тип мозга
называется зауропсидный или стриарным. Мозжечок увеличен в размерах в
связи с многообразием движений пресмыкающихся. Продолговатый мозг
образует резкий изгиб, характерный для всех амниот. Из головного мозга выходит
12 пар черепно-мозговых нервов.
46.
Такой же тип мозга характерен и для птиц, но с некоторыми особенностями.Полушария переднего мозга относительно большие. обонятельные доли у птиц
развиты слабо, что указывает на роль обоняния в жизни птиц. В
противоположность этому средний мозг представлен крупными зрительными
долями. Хорошо развит мозжечок, из головного мозга выходит 12 пар нервов.
Головной мозг у млекопитающих достигает максимального развития.
Полушария настолько велики, что покрывают средний мозг и мозжечок. Особо
развита кора больших полушарий, площадь ее увеличена за счет извилин и
борозд. Кора имеет очень сложное строение и называется новой корой –
неокортекс. Появляется вторичный мозговой свод – неопаллиум. Спереди от
полушарий расположены крупные обонятельные доли. Промежуточный мозг, как
и у других классов, включает эпифиз, гипофиз и гипоталамус. Средний мозг
относительно мал, он состоит из четырех бугров – четыреххолмия. Передняя кора
связана со зрительным анализатором, задняя – со слуховым. Наряду с передним
мозгом сильно прогрессирует мозжечок. Из мозга выходит 12 пар черепномозговых нервов. Анализирующим центром является кора больших полушарий.
Такой тип мозга называется маммальным.
47.
48.
Аномалии и пороки развития нервной системы у человека.1. Ацефалия - отсутствие головного мозга, свода, черепа и лицевого скелета;
это нарушение связано с недоразвитием переднего отдела нервной трубки и
сочетается с дефектами спинного мозга, костей и внутренних органов.
2. Анэнцефалия - отсутствие больших полушарий и крыши черепа при
недоразвитии ствола мозга и сочетается с другими пороками развития. Эта
патология обусловлена незакрытием (дизрафия) головной части нервной трубки.
При этом не развиваются кости крыши черепа, а кости основания черепа
обнаруживают различные аномалии. Анэнцефалия не совместима с жизнью,
средняя частота 1/1500, при чем чаще у женских плодов.
3. Ателэнцефалия – остановка развития (гетерохрония) передней части
нервной трубки на стадии трех пузырей. В результате большие полушария и
подкорковые ядра не формируются.
4. Прозэнцефалия – конечный мозг делится продольной бороздой, но в
глубине оба полушария остаются связанными друг с другом.
5. Голопрозэнцефалия – конечный мозг не делится на полушария и имеет вид
полусферы с единой полостью (желудочком).
6. Алобарная прозэнцефалия – разделение конечного мозга только в задней
части, а лобные доли остаются неразделенными.
49.
7. Аплазия или гипоплазия мозолистого тела – полное или частичноеотсутствие сложной комиссуры мозга, т.е. мозолистого тела.
8.
Гидроэнцефалия - атрофия больших полушарий в сочетании с
гидроцефалией.
9. Агирия - полное отсутствие борозд и извилин (гладкий мозг) больших
полушарий.
10. Микрогирия - уменьшение числа и объема борозд.
11. Врожденная гидроцефалия - преграждение части желудочковой системы
мозга и ее выходов, она вызвана первичным нарушением развития нервной
системы.
12. Spina bifida - дефект замыкания и обособления от кожной эктодермы
спинального отдела нервной трубки. Иногда эта аномалия сопровождается
дипломиелией, при которой спинной мозг расщеплен на известном протяжении
на две части, каждая со своим центральным карманом.
13. Иниэнцефалия - редкая аномалия, несовместимая с жизнью, встречается
чаще у плодов женского пола. Это грубая аномалия затылка и головного мозга.
Головы повернута так, что лицо обращено кверху. Дорсально скальп
продолжается в кожу люмбодорсальной или сакральной области.
50.
Эволюция пищеварительной системы у хордовых животных.Эволюция пищеварительной системы у хордовых животных происходит в
следующих
направлениях:
удлинение
пути,
проходимого
пищей,
дифференцировка кишечной трубки, увеличение всасывающей поверхности,
развитие пищеварительных желез.
Пищеварительный канал хордовых почти целиком имеет энтодермальное
происхождение. Только небольшой ротовой отдел и задний отдел кишечника
произошли из эктодермы. Значительная часть переднего отдела кишечной трубки
многих хордовых преобразована в орган дыхания – жабры и легкие.
Пищеварительная система
низших хордовых представлена
слабо
дифференцированной кишечной трубкой. Например, у ланцетника это прямая
трубка с одним выростом, выполняющим роль печени и поджелудочной железы.
Большая часть кишечника занимает глотка, пронизанная многочисленными
жаберными щелями.
У позвоночных животных пищеварительная система усложняется. Это
выражается в дифференцировке пищеварительного канала на ротовую полость,
глотку, пищевод, желудок, тонкую и толстую кишки. Эти отделы у разных
классов животных развиты
неодинаково. Обособляются слюнные,
поджелудочная железа и печень.
51.
У большинства видов рыб и наземных классов, позвоночных внутренняяповерхность кишок образует складки слизистой оболочки, появляются ворсинки.
У рыб с появлением челюстей возникают многочисленные зубы и костные
пластинки, которые служат для схватывания и удерживания добычи. Желудок у
большинства рыб развит слабо, иногда он представляет собой
просто
мешковидное расширение.
Печень у рыб относительно круглая, из выростов кишок образуются
плавательный пузырь и поджелудочная железа.
У амфибий в связи с выходом на сушу появляются слюнные железы,
выделяющие секрет для смачивания пищи. На челюстях имеются мелкие
однородные зубы. В ротоглоточной полости происходит перекрест
пищеварительных и дыхательных путей. Хорошо обособлены желудок, тонкая и
толстая кишки, заканчивающиеся клоакой.
У рептилий зубы еще остаются преимущественно однородными
(гомодонтными), но уже начинается дифференцировка. Так ядовитые зубы змеи
отличаются от остальных зубов, происходит одновременно и преобразование
части слюнных желез в ядовитые. Между тонкой и толстыми кишками появляется
зачатки слепой кишки.
52.
У птиц пищеварительная система в связи с полетом сильно изменилась:исчезли челюсти и зубы, появился роговой клюв.
В пищеводе имеется
мешковидное образование - зоб. Желудок разделен на две части – железистую
(кардиальную) и мускулистую, которая предназначена для измельчения пищи.
Кишка птиц представлена длинной тонкой, двумя отростками слепой и короткой
толстой кишкой.
У млекопитающих наблюдается гетеродонтная зубная система, то есть
происходит дифференцировка зубов на резцы, клыки и коренные зубы. В области
глотки с боков образуется четыре жаберных кармана (пятый обычно не
развивается).
Из первого кармана образуется евстахиева труба и среднее ухо, из второго
тонзиллярный синус, из третьего и четвертого зобная и околощитовидная железы.
Желудок млекопитающих дифференцирован на отделы и содержит
пищеварительные железы различных типов. Усложняются и кишки, а именно
увеличивается длина толстой кишки по сравнению с другими классами,
развиваются червеобразный отросток и слепая кишка.
У всех млекопитающих кроме яйцекладущих
кишечник заканчивается
самостоятельным заднепроходным отверстием, отделенным промежностью от
мочеполового, а не клоакой.
Таким образом, на ранних этапах эволюции позвоночных,
пищеварительная система постепенно усложнялась, в ней появлялись новые
органы.
53.
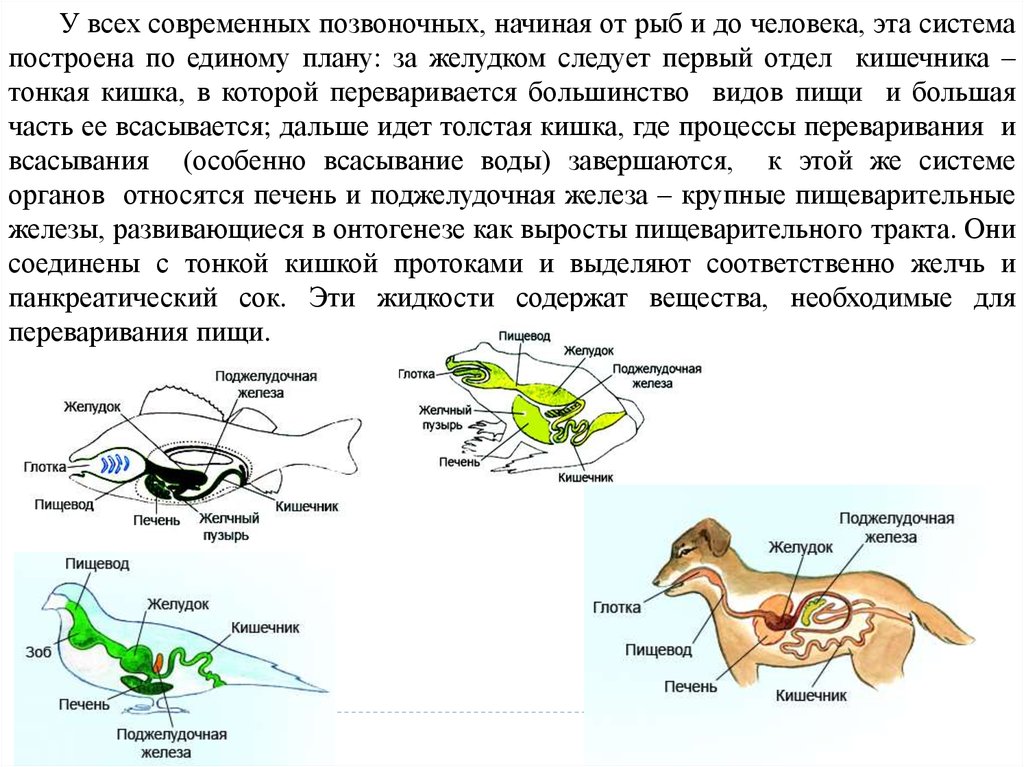
У всех современных позвоночных, начиная от рыб и до человека, эта системапостроена по единому плану: за желудком следует первый отдел кишечника –
тонкая кишка, в которой переваривается большинство видов пищи и большая
часть ее всасывается; дальше идет толстая кишка, где процессы переваривания и
всасывания (особенно всасывание воды) завершаются, к этой же системе
органов относятся печень и поджелудочная железа – крупные пищеварительные
железы, развивающиеся в онтогенезе как выросты пищеварительного тракта. Они
соединены с тонкой кишкой протоками и выделяют соответственно желчь и
панкреатический сок. Эти жидкости содержат вещества, необходимые для
переваривания пищи.
54.
Аномалии и пороки развития пищеварительной системы у человека.1. «Заячья губа» - нарушение клеточной адгезии (склеивания) верхней губы (частота
встречаемости 1:700-1000 новорожденных; несколько чаще у мальчиков).
2. «Волчья пасть» - несращение твердого неба (палатосхиз, частота встречаемости
1:2500 новорожденных, у девочек встречается чаще). Это нарушение бывает полным,
частичным, срединным, одно- и двусторонним, сквозным. Описаны также тяжелые
случаи, как сквозная расщелина верхней губы и неба – хейлогнатопалатосхиз. При
сквозных расщелинах имеется широкое сообщение между полостями носа и рта, что
редко затрудняет сосание, глотание и в последствии отразится на развитии речи.
3. Дефекты губ, которые подразделяются на:
•частичный (парциальный) дефект;
•отсутствие половины губы;
•почти полный (субтотальный) дефект, превышающие размеры половины губы;
•полный (тотальный) дефект, когда отсутствует вся губа;
•удвоенная губа (labium-duplex) – это складка слизистой оболочки, расположенная
параллельно красной каймы верхней губы, напоминая дополнительную губу. Это
явление встречается довольно часто, преимущественно у мужчин. Причиной служит
гипертрофия подслизистой клетчатки и слизистой оболочки желез губ, в результате
чего возникают складки слизистой оболочки, как правило, эти складки располагаются
по бокам губы, центральная же часть остается в пределах нормы.
•срединная расщелина нижней губы и нижней челюсти (частичная и полная форма).
Известны случаи одновременного расщепления верхней, нижней губы и нижней
челюсти.
55.
4. Макростомия (греч. macros – большой, stoma - рот) – поперечнаярасщелина лица. Частота встречаемости 1:80000. возникает вследствие
незаращения тканей верхней и нижней частей щеки, краев губ между собой, это
приводит к формированию большой ротовой щели.
5. Микростомия (micros – малый, stoma - рот) – чрезмерно уменьшенная
ротовая щель, является результатом избыточного срастания тканей краев губ и
щек. Обычно сочетается с тяжелыми пороками развития производных 1-ой
жаберной дуги.
6. Смещение углов рта (deviatia) – эта аномалия подразделяется на:
• смещение угла рта вниз;
• смещение угла рта вверх.
Эти анатомические изменения сопровождаются нарушением дикции,
истечением слюны, трудностями при приеме пище и оказании стоматологической
помощи.
7. Свищи нижней губы – нарушение обычно парное и располагаются на
красной кайме губ по обе стороны от средней линии. Представляют собой
протоки добавочных слизистых желез.
8. Двойной рот – крайне редкий порок ротовой полости, проявляющийся
добавочной ротовой щелью, открывающейся в добавочную ротовую полость
меньших размеров. При этом полости не сообщаются.
56.
9. Разнообразные аномалии и пороки развития языка – расщепление языка,ромбовидный, щелевидный (поверхность языка испещрена мелкими или глубокими
полосами, уходящими к основанию языка), складчатый язык. Описано формирование
большого (macros – большой, glossa - язык) или маленького (micros - малый, glossa язык) языка, добавочного языка или даже отсутствие (аглоссия) языка.
10. Пороки, связанные с развитием слюнных желез:
•аплазия – нарушение закладки и последующего развития слюнных желез;
•гипоплазия (недоразвитие) отдельных желез, в частности односторонняя гипоплазия
больших слюнных желез;
•дистопии или гетеротопии (dys – нарушение, heteros – иной, topos - место) –
аномалии положения желез.
11. Атавистические аномалии зубной системы, связанные с изменением количества
зубов и с нарушением их дифференцировки:
•адентия (а – отсутствие, odontos - зуб) – отсутствие зубов, которое может быть
полным или частичным. Полная адентия (отсутствии всех зубов) встречается редко;
•гомодонтная зубная система – все зубы одинаковые;
•трехбугористое строение коренных зубов вместо четырехбугористого.
•прорезывание сверхкоплектных зубов в ряду зубов или за его пределами.
•макро- или микродентия – формирование слишком крупных или слишком мелких
зубов, связанных с отклонениями нормального течения образования и
дифференцировки зубных зачатков.
57.
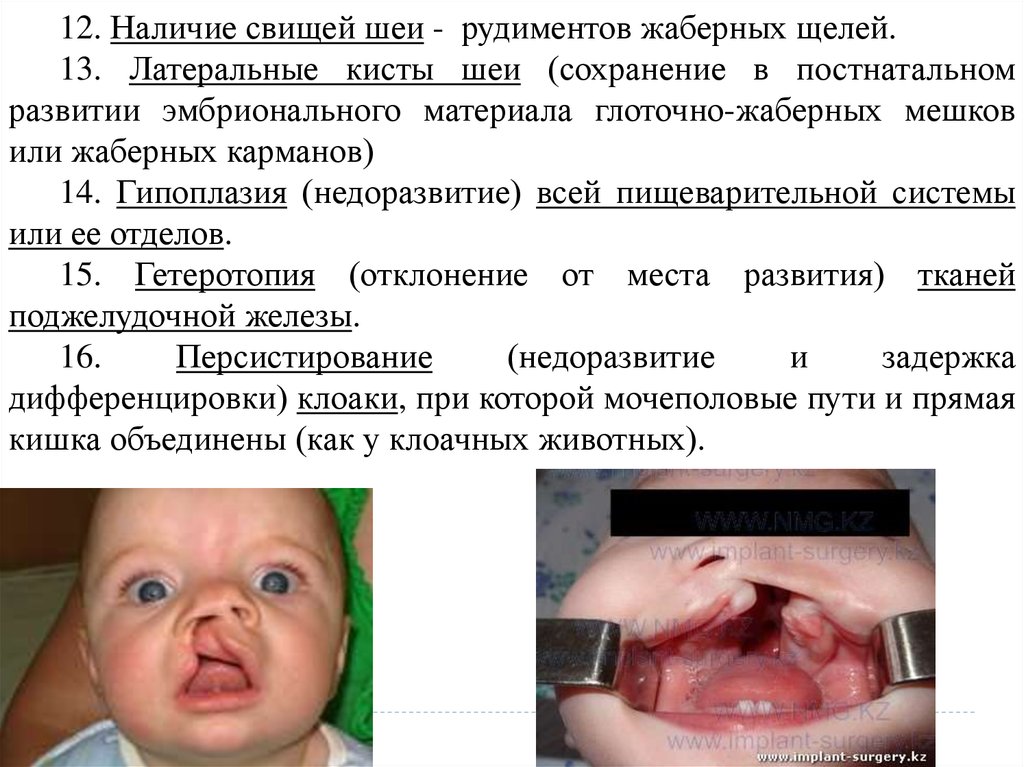
12. Наличие свищей шеи - рудиментов жаберных щелей.13. Латеральные кисты шеи (сохранение в постнатальном
развитии эмбрионального материала глоточно-жаберных мешков
или жаберных карманов)
14. Гипоплазия (недоразвитие) всей пищеварительной системы
или ее отделов.
15. Гетеротопия (отклонение от места развития) тканей
поджелудочной железы.
16.
Персистирование
(недоразвитие
и
задержка
дифференцировки) клоаки, при которой мочеполовые пути и прямая
кишка объединены (как у клоачных животных).
58.
Дыхательная система животных организмов имеет энтодермальное происхождение,так как по происхождению она связана с пищеварительной системой. Функцией
органов дыхания является газообмен между конкретным организмом и окружающей
средой.
Эволюция дыхательной системы у хордовых животных.
Дыхательная система всех хордовых животных и по происхождению, и
топографически связана с кишечником, то есть с энтодермой.
У водных хордовых функцию дыхания выполняют жаберные щели,
пронизывающие передний отдел кишечной трубки-глотки. У наземных хордовых
жаберные щели закрываются во время эмбрионального развития, а затем исчезают.
Функцию дыхания выполняют легкие, образующиеся из выпячивания кишечной
трубки.
Эволюция жаберного аппарата у хордовых выражалось в уменьшении числа
жаберных щелей при одновременном увеличении дыхательной поверхности путём
образования жаберных лепестков.
Эволюция лёгких шла в направлении обособления дыхательных путей и
увеличения дыхательной поверхности путем образования легких губчатого строения
со сложной системой разветвления внутриклеточных бронхов, заканчивающихся
пузырьками с ячеистыми клетками.
Наиболее примитивна дыхательная система у ланцетника, относящегося к
низшим хордовым (подтип Бесчерепные). Передний отдел кишечника (стенка глотки)
прободен жаберными щелями (до150 пар), которые открываются в артериальную
(околожаберную) полость.
59.
У круглоротых (подтип Позвоночные) органами дыхания являются такжежаберные щели, но их уже меньше (5-15 пар). Они сообщаются с передним
отделом кишечника и открываются наружу самостоятельными отверстьями.
Настоящие жабры появляются среди хордовых у рыб. Они представляют
собой тонкие складки слизистой оболочки глотки, лежащие на жаберных дугах и
снабжаемые венозной кровью через жаберные артерии, распадающиеся здесь на
капилляры. У рыб имеется 4-7 (чаще 5) жаберных мешков между жаберными
дужками. Тычинки, расположенные на выпуклой поверхности жаберных дужек,
препятствуют попаданию пищи из глотки в жабры.
Помимо жабр у рыб имеются добавочные органы дыхания, позволяющие им
использовать кислород воздуха. Таким органом у рыб является плавательный
пузырь. Стенки его богаты кровеносными сосудами, поэтому у некоторых
зарывающихся в ил рыб он может служить для газообмена. Плавательный пузырь
у большинства рыб развивается из дорзальных участков глотки и не является
гомологом легких. Только у кистеперых рыб плавательный пузырь образуется как
выпячивание вентральной части глотки и служит гомологом легких наземных
животных, так как легкие позвоночных развиваются из брюшной части
жаберного мешка.
У двоякодышащих рыб лёгкие есть, а плавательный пузырь отсутствует.
60.
У личинок амфибий, как и у рыб, органы дыхания представленыдревовидноветвящимися наружными жабрами. У большинства взрослых амфибий
появляются лёгкие в виде тонкостенных парных выростов брюшной глотки позади
последнего жаберного мешка. В связи с отсутствием грудной клетки и диафрагмы
воздух в них попадает из ротовой полости за счет глотательных движений,
осуществляющихся подбородочно-подъязычной мышцей. Материал жаберных дуг,
следующий за подъязычной дугой, частично входит в состав хрящей гортани, которая
появляется впервые у земноводных, являясь первым органом, относящимся к нижним
дыхательным путям. Лёгкие начинаются непосредственно от гортани. Они
крупноячеисты и имеют малую дыхательную поверхность, в связи с чем газообмен в
большей степени осуществляется через кожные покровы, которые пронизаны
большим количеством кровеносных капилляров и снабжены слизистыми железами.
У рептилий дыхательная система усложняется. Легкие у них уже мелко ячеистые, они содержат многочисленные ячеистые перекладины и обладают большой
дыхательной поверхностью. В дыхательных путях в связи с окончательным выходом
рептилий на сушу наблюдается прогресс: выделяются верхние дыхательные пути, хотя
и не окончательно отграниченные от ротовой полости - это носовая полость, а нижние
– гортань, трахея и бронхи. Впервые появляется диафрагма, которая в дыхании
принимает пассивное участие. Она либо частично разделяет грудную и брюшную
полость, либо лишена мышечных волокон.
Механизм дыхания осуществляется за счет сокращения межреберных мышц,
приводящих в движение грудную клетку.
61.
У птиц легкие представляют собой плотно-губчатые телапронизанные
разветвлениями бронхов, а не мешки, как у рептилий. Кроме того, они
дополняются в качестве резервуара для воздуха тонкостенными воздушными
мешками. Последние располагаются между всеми воздушными органами, между
мышцами, в полости кости и под костью.
При подъеме крыльев воздушные мешки через легкие
наполняются
воздухом, при опускании крыльев воздух через легкие выходит наружу. Таким
образом, во время полета у птиц осуществляется двойное дыхание. Во время
покоя птица дышит лишь путем расширения и сужения грудной клетки.
В нижней гортани помещается голосовой аппарат, обладающий сложной
мускулатурой.
Органы дыхания у всех млекопитающих характеризуются сложностью как
легких, имеющих альвеолярное строение, так и дыхательных путей. Идет
дальнейшее по сравнению с предыдущими классами позвоночных животных
разветвление бронхиального дерева.
Трахея делиться на бронхи, которые
ветвятся на бронхи второго, третьего и четвертого порядков и до самих мелких бронхиол, на разветвлениях которых находятся альвеолы, легочные пузырьки
имеющие в сумке огромную площадь (около 90 см²), в которых происходит
газообмен. Основная мышца, играющая важнейшую роль в акте дыхания,
является диафрагма. Дыхательные пути млекопитающих выстланы мерцательным
эпителием и полностью отделены от пищеварительной системы.
62.
63.
Аномалии и пороки развития дыхательной системы у человека.1. В эмбриогенезе человека отражается первоначальная связь
пищеварительной и дыхательной систем. В связи с этим становится понятным
появление у человека большой группы врожденных пороков развития пищевода и
трахеи типа эзофаготрахеальных свищей («жаберных щелей»).
2. Дизонтогенетические бронхолегочные кисты – округлая полоса в легких,
отграниченная от окружающей ткани
примитивно построенной стенкой
недифференцированного бронха. Эти аномалии могут быть объяснены
нарушением альвеолярной дифференцировки легочной ткани.
3. Кистозная гипоплазия – недоразвитие легкого. При этой патологии
недоразвитыми оказываются целые доли легкого, которые представляют собой
многочисленные полости, связанные с крупными бронхами и имеющие малую
поверхность. Газообмен в таких участках легкого резко ослаблен.
4. Гипоплазия диафрагмы – недоразвитие диафрагмы от небольших дефектов
в ее куполе до полной аплазии. Данная аномалия несовместима с жизнью и
встречается чаще вместе с другими множественными пороками развития.
64.
Происхождение и функции кровеносной системы.Функцией кровеносной системы является доставка ко всем органам тела
кислорода и питательных веществ, удаления из организма продуктов распада и
углекислоты, а также гуморальная функция.
Кровеносная система в основном имеет мезодермальное происхождение.
2. Эволюция кровеносной системы у хордовых животных.
У низших хордовых, в частности у ланцетника, кровеносная система замкнутая,
но сердца нет. Роль сердца выполняет брюшная аорта, от которой отходят приносящие
жаберные артерии, в количестве 100-150 пар, несущие венозную кровь. Проходя через
жаберные перегородки в неразветвленном виде кровь в артериях успевает окислиться
и через выносящие парные жаберные артерии уже артериальная кровь поступает в
корни спинной аорты, которые сливаются в непарную спинную аорту, от которой идут
сосуды, несущие питательные вещества и кислород ко всем частям тела. Венозная
кровь со спинной части собирается в передние и задние кардинальные вены, которые
сливаются в левой и правой кювьеровы протоки, а из них в брюшную аорту. Кровь от
брюшной стороны собирается в подкишечную вену, которая несет кровь в печень, где
она обеззараживается, а оттуда по печеночной вене также впадает в кювьеров проток и
далее брюшной сосуд.
65.
У высших хордовых, в частности у низших позвоночных, т.е. у круглоротых иу рыб, усложнение кровеносной системы выражено в появлении сердца, которое
имеет одно предсердие и один желудочек. В сердце бывает только венозная кровь.
Круг кровообращения один, в котором артериальная и венозная кровь не
смешиваются. Круговорот крови по телу сходен с кровеносной системой
ланцетника. От сердца венозная кровь идет к жабрам, где окисляется, и от них
окисленная (уже артериальная) кровь разносится по всему телу и по венам
возвращается к сердцу.
С выходом животных на сушу и с появлением легочного дыхания, появляется
второй круг кровообращения. Сердце получает не только венозную, но и
артериальную кровь, и поэтому дальнейшая эволюция кровеносной системы идет
по пути обособления двух кругов кровообращения. Это достигается делением
сердца на камеры.
У земноводных и рептилий трехкамерное сердце, которое не обеспечивает
полного разделения двух кругов кровообращения, поэтому еще происходит
смешение артериальной и венозной крови. Правда, у рептилий желудочек уже
разделен неполной перегородкой, а у крокодила четырех камерное сердце,
поэтому смешение артериальной и венозной крови наблюдается в меньшей
степени, чем у земноводных.
66.
У птиц и млекопитающих сердце полностью разделено на четыре камеры –два предсердия и два желудочка. Два круга кровообращения, артериальная и
венозная кровь не смешиваются.
Разберем эволюцию жаберных дуг у позвоночных животных.
У всех эмбрионов позвоночных животных впереди от сердца закладывается
непарная брюшная аорта, от которой отходят жаберные дуги артерий. Они
гомологичны артериальным дугам в кровеносной системе ланцетника. Но у них
число артериальных дуг небольшое и равняется числу висцеральных дуг. Так у
рыб их шесть. Первые две пары дуг у всех позвоночных испытывают редукцию,
т.е. атрофируются. Оставшиеся четыре дуги ведут себя следующим образом.
У рыб разделяются на приносящие к жабрам и выносящие из жабер
жаберные артерии.
Третья артериальная дуга у всех позвоночных, начиная с хвостатых амфибий,
превращается в сонные артерии и несет кровь к голове.
Четвертая артериальная дуга достигает значительного развития. Из нее у всех
позвоночных животных, опять же начиная с хвостатых амфибий, образуются
собственно дуги аорты. У амфибий и рептилий парные, у птиц правая дуга (левая
атрофируется), а у млекопитающих левая дуга аорты (правая атрофируется).
Пятая пара артериальных дуг у всех позвоночных, за исключением хвостатых
амфибий, атрофируется.
Шестая пара артериальных дуг теряет связь со спинной аортой, из нее
образуются легочные артерии.
67.
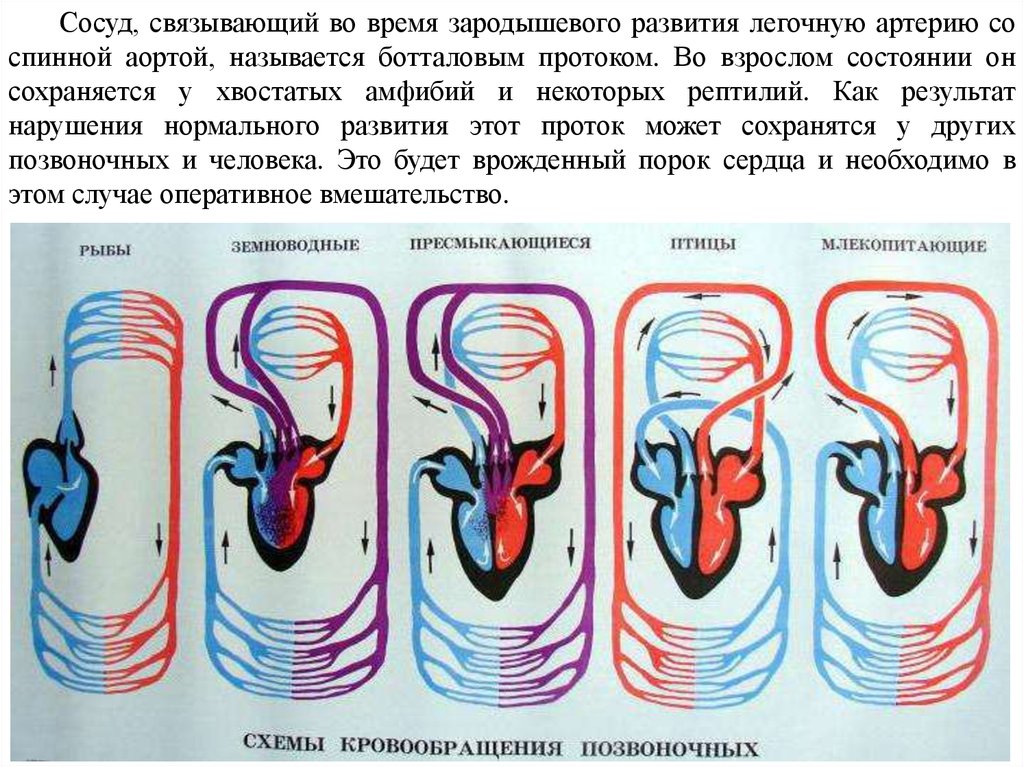
Сосуд, связывающий во время зародышевого развития легочную артерию соспинной аортой, называется ботталовым протоком. Во взрослом состоянии он
сохраняется у хвостатых амфибий и некоторых рептилий. Как результат
нарушения нормального развития этот проток может сохранятся у других
позвоночных и человека. Это будет врожденный порок сердца и необходимо в
этом случае оперативное вмешательство.
68.
1.Аномалии и пороки развития кровеносной системы у человека.На основании изучения филогенеза сердечно-сосудистой системы становится
понятным происхождения ряда аномалий и уродств у человека.
1. Шейная эктопия сердца – расположение сердца в области шеи. Сердце
человека развивается из парных закладок мезодермы, которые сливаются и
образуют единую трубку в области шеи. В процессе развития трубка смещается в
левую часть грудной полости. Если сердце задерживается в области
первоначальной закладки, то и возникает данный порок, при котором ребенок
обычно погибает сразу после рождения.
2. Дестрокардия (гетеротопия) – расположение сердца справа.
3. Двухкамерное сердце – остановка развития сердца на этапе двух камер
(гетерохрония). От сердца в этом случае отходит только один сосуд –
артериальный ствол.
4. Незаращение первичной или вторичной межпредсердной перегородки
(гетерохрония) в области овальной ямки, которая у эмбриона является
отверстием, а также полное их отсутствие приводит к образованию
трехкамерного сердца с одним общим предсердием (частота встречаемости 1:1000
рождений).
5. Незаращение межжелудочковой перегородки (гетерохрония) с частотой
встречаемости 2,5-5:1000 рождений. Редким пороком является ее полное
отсутствие.
69.
6. Персистирование (нарушение дифференцировки) артериального, илиботталлова, протока, представляющего собой часть корня спинной аорты между
4-й и 6-й парами артерий слева. Когда легкие не функционируют, у человека во
время зародышевого развития имеется ботталлов проток. После рождения проток
зарастает. Сохранение его ведет к серьезным функциональным нарушениям,
поскольку проходит смешанная венозная и артериальная кровь. Частота
встречаемости 0,5-1,2:1000 рождений.
7. Правая дуга аорты – самая частая аномалия жаберных дуг артерий. При
развитии происходит редукция левой дуги 4-й пары вместо правой.
8. Персистирование обеих дуг аорты 4-й пары, так называемое «Аортальное
кольцо» - у эмбриона человека иногда не происходит редукции правой артерии 4й жаберной дуги и корня аорты справа. В этом случае вместо одной дуги аорты
развиваются две дуги, которые обогнув трахею и пищевод, соединяются в
непарную спинную аорту. Трахея и пищевод оказываются в аортальном кольце,
которое с возрастом сжимается. Порок проявляется нарушением глотания и
удушением.
9. Персистирование первичного эмбрионального ствола. На определенной
стадии развития у эмбриона имеется общий артериальный ствол, который потом
разделяется спиральной перегородкой на аорту и легочной ствол. Если
перегородка не развивается, то общий ствол сохраняется. Это приводит к
смешению артериальной и венозной крови и обычно заканчивается смертью
ребенка.
70.
10. Транспозиция сосудов – нарушение дифференцировки первичногоаортального ствола, при котором перегородка приобретает не спиральную, а
прямую форму. В этом случае аорта будет отходить от правого желудочка, а
легочной ствол – от левого. Этот порок встречается с частотой 1:2500
новорожденных и несовместим с жизнью.
11. Открытый сонный проток – сохранение комиссуры между 3-й и 4-й
парами артериальных дуг (сонной артерией и дугой аорты). В результате
увеличивается кровоток в мозг.
12. Персистирование двух верхних полых вен. У человека аномалией
развития является наличие дополнительной верхней полой вены. Если обе вены
впадают в правое предсердие, аномалия клинически не проявляется. При
впадении левой вены в левое предсердие происходит сброс венозной крови в
большой круг кровообращения. Иногда обе полые вены впадают в левое
предсердие. Такой порок несовместим с жизнью. Данная аномалия встречается с
частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
13. Недоразвитие нижней полой вены – редкая аномалия, при которой отток
крови от нижней части туловища и ног осуществляется через коллатерали
непарной и полунепарной вен, являющихся рудиментами задних кардиальных
вен. Редко встречается атрезия (отсутствие) нижней полой вены (кровоток
осуществляется через непарные или верхнюю полую вену).
14. Отсутствие воротной системы печени.
71.
Выделительная система имеет мезодермальное происхождение, а точнееобразуется из нефротома сомитов. Она выполняет функцию удаления жидких
продуктов обмена веществ из организма.
Эволюция выделительной системы у хордовых животных.
Выделительная система у низших хордовых построена по типу нефридиев.
Так у ланцетника в области жаберных щелей метамерно расположено до 100 пар
нефридиев, один конец которых открывается во вторичную, а другой – в
околожаберную полость. Края целомического отверстия нефридиев (нефростом)
имеют множество соленоцитов – клеток похожих на терминальные клетки
протонефридиев. Следовательно, выделительные органы ланцетника имеют
характер как прото-, так и метанефридиев.
Далее эволюция выделительной системы у хордовых шла по пути перехода от
нефридиев низших хордовых к специальным органам – почкам, которые прошли
длительный путь развития.
У низших позвоночных (Anamnia) почки проходят две стадии: предпочки
(головная или pronefros) и первичной (туловищная или mesonefros). У высших
позвоночных (Amniota) развитие почек происходит в три стадии: предпочки,
первичной и вторичной (тазовой или metanefros).
72.
Структурной и функциональной единицей почек является нефрон, которыйтакже, как и почка, прошел длительный путь эволюции.
Почки закладываются, как уже уточнялось, в мезодерме, а именно в
нефротомах. Выделительная система позвоночных связана с органами половой
системы. Половые железы позвоночных, как правило, закладываются в виде
парных складок на вентральной поверхности мезонефросов. Зачаток гонад
состоит из утолщенного эпителия с большим количеством соединительной ткани.
Сначала мужские и женские половые железы имеют одинаковое строение.
Позднее происходит их специализация и возникает связь различными для
каждого вида частями выделительной системы, которые становятся половыми
протоками.
73.
У зародышей всех позвоночных животных закладывается головная почка илипредпочка. Она состоит из 6-12 нефронов, продукты выделения которых
собираются в общий мочеточник (парамезонефральный проток). Нефрон
предпочки состоит из воронки (нефростома), которая выстлана ресничками и
открывается в целом, и короткого прямого выделительного канала. Около воронок
в стенках полости тела образуются грушевидные выросты из клубочков
артериальных капилляров. Они отфильтровывают в целомическую полость как
продукты выделения, так и полезные вещества. Целомическая жидкости попадает
в воронки, канальцы и, собираясь в общий мочеточник, выводится в клоаку или
мочевое отверстие. Несовершенство нефронов предпочки заключается в
отсутствии прямой связи между кровеносной и выделительной системами, а
также в постоянном присутствии в целомической жидкости продуктов выделения.
Предпочка во взрослом состоянии функционирует только у миксин (класс
Круглоротые), а у всех остальных подвергается редукции (у зародыша человека
она сохраняется около 40 часов).
У анамний после редукции предпочки появляется первичная почка.
Первичная почка закладывается в туловищных сегментах тела. Она содержит
до нескольких сотен нефронов, продукты выделения которых собираются в
выделительные протоки. Нефрон первичной почки состоит из: воронки
(нефростома), которая выстлана ресничками и открывается в целом; почечного
тельца, которое состоит из двустенной капсулы Боумена–Шумлянского и
клубочка капилляров; извитого выделительного канала.
74.
Продукты выделения из капилляров клубочка отфильтровываются в полостькапсулы, по извитому канальцу собираются в мочеточник, мочевой пузырь и
выводятся через клоаку или мочевые отверстия.
Нефрон первичной почки характеризуется рядом прогрессивных изменений:
• появляется прямая связь между кровеносной и выделительной системами;
• увеличивается количество нефронов в почке;
• происходит удлинение и изменение формы извитого канальца, в результате
чего начинают осуществляться процессы обратного всасывания нужных веществ,
и происходит концентрация мочи;
• уменьшается количество продуктов выделения в целоме.
У низших позвоночных (у миног из класса Круглоротые, у рыб и у амфибий)
первичная почка функционирует в течении всей жизни как орган выделения.
У высших позвоночных (рептилий, птиц и млекопитающих) в том числе и
человека первичная почка редуцируется.
У самок амниот часть канальцев первичной почки сохраняется в виде
незначительных рудиментов epoophron и paraophron, а из остатков предпочки и
мочеточника развивается яйцевод, который дифференцируется на отделы, а
именно маточные трубы, матку и влагалище.
75.
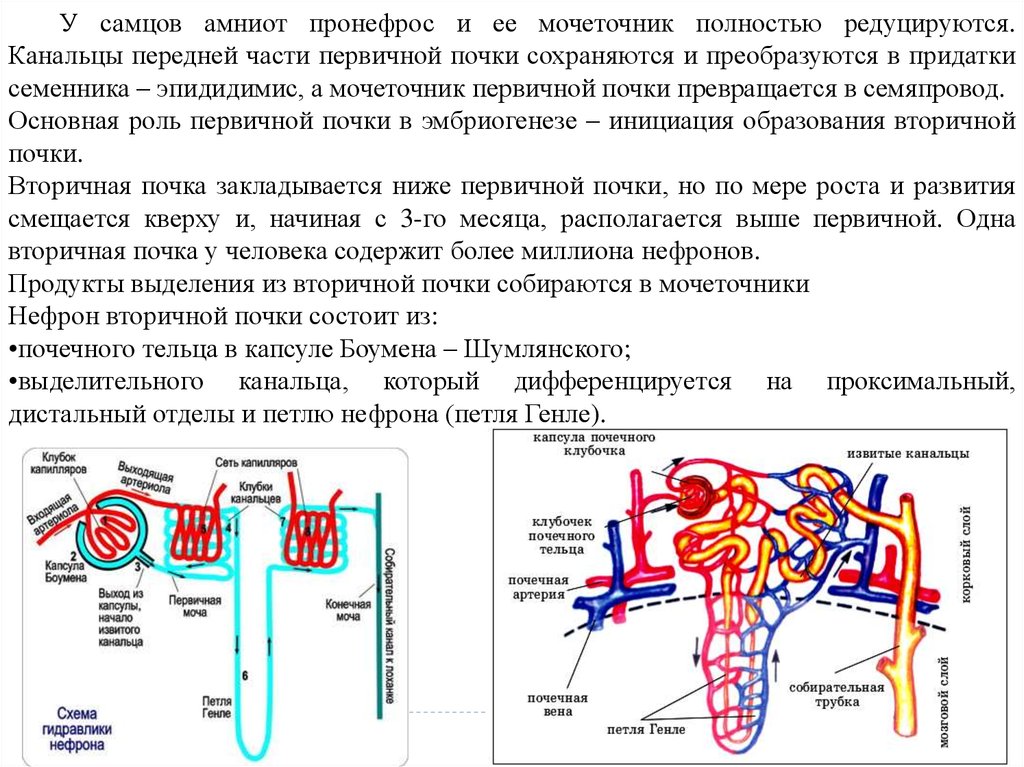
У самцов амниот пронефрос и ее мочеточник полностью редуцируются.Канальцы передней части первичной почки сохраняются и преобразуются в придатки
семенника – эпидидимис, а мочеточник первичной почки превращается в семяпровод.
Основная роль первичной почки в эмбриогенезе – инициация образования вторичной
почки.
Вторичная почка закладывается ниже первичной почки, но по мере роста и развития
смещается кверху и, начиная с 3-го месяца, располагается выше первичной. Одна
вторичная почка у человека содержит более миллиона нефронов.
Продукты выделения из вторичной почки собираются в мочеточники
Нефрон вторичной почки состоит из:
•почечного тельца в капсуле Боумена – Шумлянского;
•выделительного канальца, который дифференцируется на проксимальный,
дистальный отделы и петлю нефрона (петля Генле).
76.
Продукты выделения поступают в нефрон путем фильтрации крови вкапсулах. Образуется первичная моча, у человека она составляет 170-180 литров в
сутки. В извитых почечных канальцах первичная моча концентрируется за счет
реабсорбции – обратного всасывания нужных веществ и образование вторичной
мочи. Вторичная моча (1,7-1,8 литра в сутки у взрослого человека) собирается в
мочеточники. Они образуются из боковых выростов мочеточника первичной
почки.
Таким образом, в эволюции животных можно выделить три типа
выделительных систем: протонефридии, метанефридии, почки. Эволюция этой
системы в ряду позвоночных идет в направлении, во-первых, увеличения более
тесной связи с кровеносной системой, во-вторых, увеличения выделительной
поверхности за счет увеличения числа нефронов и, в-третьих, совершенствования
строения самого нефрона, который теряет связь с целомической полостью,
удлиняет почечный каналец и создает механизм обратного всасывания.
Процесс формирования мочеполовой системы позвоночных является ярким
примером субстиции органов.
Субстиция – способ преобразования органов, при котором более ранние
закладки органа после появления последующих редуцируются.
77.
78.
1.Аномалии и пороки развития выделительной системы у человека.Сложность формирования почек делает возможность появления отклонений
от нормального процесса. Аномалии развития почек бывают различными.
1. «Опущение почки» - одна из почек может не подняться из области таза в
поясничную область и остаться там, где закладывалась, т.е. в области таза.
2. «Подковообразная почка» - при низком положении почек и срастании их
нижними полюсами получается подковообразная почка.
3. Образование общей почечной массы – нарушение, при котором обе почки
могут оказаться по одну сторону от средней линии и срастись в общую почечную
массу.
4. Наличие третьей почки – число почек может быть больше или меньше
нормального. Очень редко встречается третья почка, лежащая на позвоночном
столб между двумя или ниже какой-либо из них.
5. Арения – врожденное отсутствие обеих почек, часто сочетается с другими
врожденными пороками. Дети нежизнеспособны.
6. Агенезия почек – врожденное отсутствие одной из почек.
7. Гипоплазия почек – врожденное уменьшение массы и объема почек, может
быть одно- и двусторонним. При односторонней гипоплазии наблюдается
гипертрофия второй нормальной почки.
8. Поликистоз почек – двустороннее увеличение почек с формированием
кист. Различают крупнокистозные и мелкокистозные почки.
79.
Крупнокистозные почки характеризуются образованием в корковом слоебольшого числа крупных кист с прозрачным содержимым, между кистами
сохраняются участки нормальной почечной паренхимы. С такими почками больные
могут доживать до 40-50 лет.
9. Агенезия (отсутствие), атрезия (врожденное отсутствие или аномальное
строение), стеноз (сужение просвета), эктопия (ненормальное расположение) устьев
лоханок и мочеточников.
10. Удвоение мочеточников – частичное расщепление мочеточника или полное –
два мочеточника с двумя лоханками и двумя устьями в мочевом пузыре.
11. Агенезия (отсутствие) мочевого пузыря.
12. Полное или неполное удвоение мочевого пузыря – при удвоении мочевого
пузыря продольная перегородка делит пузырь на две самостоятельные части. Каждая
часть пузыря имеет либо самостоятельное сообщение с отдельным для каждой из них
мочеиспускательным каналом (т.е. имеется двойная уретра), или обе половины пузыря
открываются в одну уретру, либо открывается в уретру только одна половина пузыря.
13. Расщепление мочеиспускательного канала – это нарушение может
располагаться на его нижней (гипоспадия) или верхней (эписпадия) поверхности. Все
пороки мочевыводящих путей ведут к нарушению оттока мочи и к появлению
осложнений (гидронефроз, хронический пиелонефрит и др.). необходимо
хирургическое вмешательство.
14. Эписпадия – врожденная аномалия, при которой нарушенное отверстие
уретры находится на верхней поверхности полового члена, встречается очень редко.
80.
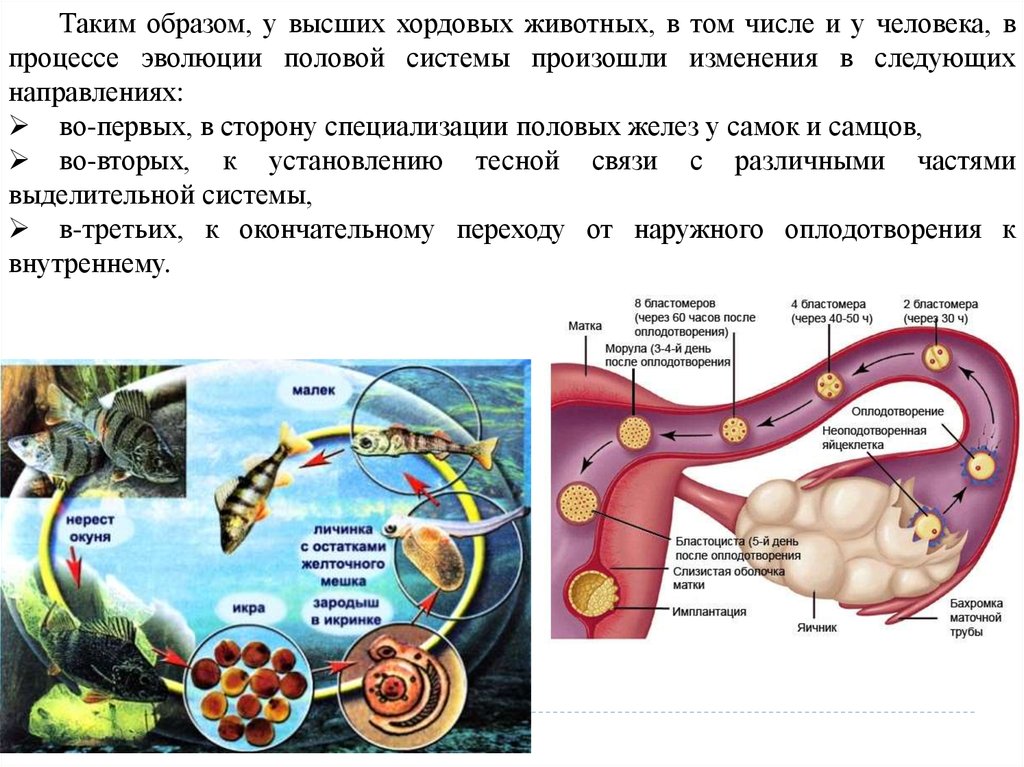
Размножение, или репродукция – это одно из основных свойств,характеризующих жизнь. Под размножением понимают способность организмов
воспроизводить себе подобных. Размножение – необходимое условие
существования вида и преемственности генераций внутри вида.
Органы размножения у всех животных имеют мезодермальное
происхождение, хотя первичные половые клетки обосабливаются в онтогенезе
значительно раньше, чем образуется третий зародышевый листок. Очень рано в
эмбриогенезе происходит дифференцировка на соматические и половые клетки.
Развитие половой системы тесно связано с развитием выделительной
системы, которая закладывается раньше половой.
Эволюция половой системы у хордовых животных.
Ходовые животные за исключением оболочников являются раздельнополыми.
Половые железы у разных полов низших хордовых очень сходны по строению, а
половой диморфизм у мужских и женских особей слабо выражен.
Так у ланцетника половые железы у самцов и самок имеют вид пузырьков,
метамерно располагающихся в стенках протоков. Половые клетки попадают в
околожаберную полость через разрыв стенок, откуда с током воды через атриопор
выходят наружу.
81.
Половые железы у всех позвоночных развиваются в виде парных складокчасти нефрогонатома, в области ножек сомита. Половые складки вдаются в
полость тела и оказываются подвешенными на брыжейке. Первичные половые
клетки обособляются у зародышей на стадии гаструляции. Вначале они
обнаруживаются в составе презумптивной эктодермы головного конца эмбриона,
затем попадают в энтодерму, откуда перемещаются в половые складки. Здесь
дифференцирующийся эпителий половых желез, включающий в себя первичные
половые клетки, объединяется с соединительнотканной стромой в виде шнуров.
Такая гонада индифферентна в половом отношении и может развиваться как в
семенник, так и в яичник в зависимости от генетического и эпигенетического
факторов дифференцировки полов. Индифферентность развивающейся половой
железы называется первичным гермафродитизмом. Он эволюционно связан,
вероятно, с гермафродитизмом древних предков позвоночных. Доказательством
этого может служить наличие у наиболее примитивных позвоночных –
круглоротых – половых желез, передняя часть которых является яичником, а
задняя – семенником. У хрящевых рыб строение половых желез самок и самцов в
значительной степени сходно. У самок некоторых земноводных на протяжении
всей жизни сохраняется рудиментный участок индифферентной половой железы.
Да и зародыш человека до определенного возраста имеет недифференцированные
половые железы.
82.
Но, все же, начиная с костистых рыб и далее у всех остальных позвоночных,в норме наблюдаются отличия половых органов у самок и самцов. У самок
яичник всегда имеет фолликулярное строение, а в семенниках появляются
семявыносящие каналы, по которым зрелые сперматозоиды выходят во внешнюю
среду. У немногих живородящих рыб оплодотворение внутреннее, а в основном у
рыб оплодотворение наружное, поэтому половые железы сильно разрастаются в
связи с выделением большого количества половых продуктов.
Амфибии, как и рыбы, анамнии. У них, как и у рыб, чаще наружное
оплодотворение, только у хвостатых амфибий оплодотворение внутреннее.
Продуцируют амфибии также большое количество половых продуктов.
У всех высших позвоночных, т.е. амниот (у рептилий, птиц и
млекопитающих) оплодотворение внутреннее. У рептилий и птиц половые
железы закладываются как парные органы, но затем развиваются неравномерно.
У самок рептилий лучше развивается правый яичник, а у птиц – левый. У птиц
яйцевод дифференцирован на отделы: передняя часть его выделяет белковые
вещества, а задняя – вещества, необходимые для формирования скорлупы.
Рептилии и птицы – клоачные животные.
83.
У млекопитающих происходит дальнейшее усложнение половой системы.Только половая система однопроходных млекопитающих (утконос и ехидна)
напоминают половую систему рептилий и птиц (откладывание яиц, наличие
клоаки). Усложнение половой системы млекопитающих связано прежде всего с
развитием приспособлений для вынашивания детеныша в матке и питания их
через плаценту. Матка у самок образуется за счет срастания расширенных частей
задней части яйцеводов. Противоположные концы яйцеводов открываются
воронками в брюшную полость рядом с яичниками. У самцов большинства видов
млекопитающих семенники выходят наружу и лежат в мошонке. Появляется у
них сложная система придаточных желез (семенные пузырьки, предстательная
железа) и копулятивные органы.
Половые железы у млекопитающих развиваются, как и других позвоночных,
впереди закладок почек, но затем перемещаются в тазовую область. Яичники
остаются в задней части брюшной полости, а семенники в норме опускаются из
брюшной полости в мошоночную, вместе с обволакивающей их складкой
брюшины.
Закладка половой системы у обоих полов млекопитающих происходит на
стадии 3-4 недель в виде половых валиков – утолщений целомического эпителия
на поверхности первичных почек и протекает в начальных стадиях развития
одинаково. От валиков в первичную почку врастают тяжи (половые шнурки) с
зачатковыми клетками, называемыми гонобластами.
84.
Дифференцировка индифферентной половой железы по мужскому илиженскому типу начинается на 6-8 недели. При развитии по мужскому типу
половые шнуры превращаются в семенные канальцы, а гонобласты образуют
сперматогенный эпителий. При развитии по женскому типу происходит рост
половых шнуров, образующих корковую часть яичника, а мозговая часть яичника
образуется из мезенхимальных элементов остатков тела первичной почки
(мезонефроса).
В эмбриогенезе всех позвоночных выводящие протоки половых желез тесно
связанны с выделительными канальцами.
При развитии предпочки вдоль тела, от головного конца к клоаке
закладывается пронефрический канал, по которому продукты диссимиляции из
нефронов поступают во внешнюю среду. При развитии первичной почки этот
канал либо расщепляется на два канала, идущих параллельно, либо второй канал
образуется в продольном утолщении стенки первого. Один канал называется
вольфовым каналом, а другой мюллеровым. Вольфов канал вступает в связь с
каналами первичной почки. Мюллеров канал срастается передним концом с
одним из нефронов предпочки и образует яйцевод, открывающийся передним
концом в целом широкой воронкой, а задним – впадающий в клоаку.
85.
У всех позвоночных животных, вне зависимости от пола, обязательноформируются как вольфов, так и мюллеров протоки, но судьба их различна у
разных животных в зависимости от пола, и принадлежности к тому или иному
классу.
У самок рыб и земноводных вольфов канал выполняет функцию
мочеточников, а мюллеров - яйцеводов. У самцов мюллеров канал редуцируется,
а вольфов выполняет роль мочеточника и семяпровода. Семенные канальца при
этом впадают в почку, а сперматозоиды при оплодотворении поступают в воду
вместе с мочой.
У амниот, т.е. у рептилий, птиц и млекопитающих, судьба вольфова и
мюллерова каналов несомненно иная. Вольфов канал у самцов выполняет в
основном функцию семяизвергательного канала, только отщепившийся от его
каудальной части проток выполняет роль мочеточника вторичной почки. В
семяпровод открываются протоки придаточных половых желез – предстательной
и семенных пузырьков. Мюллеров канал у самцов редуцируется, а вольфов канал
за исключением отщепившегося от его каудальной части протока, выполняющего
роль мочеточника, становится семяпроводом. У самок мюллеров канал выполняет
роль яйцеводом, а вольфов канал за исключением отщепившегося от его
каудальной части протока, выполняющего роль мочеточника, редуцируется.
86.
У плацентарных млекопитающих мюллеров канал дифференцируется насобственно яйцевод, матку и влагалище.
У яйцекладущих и сумчатых млекопитающих мюллеров канал сохраняет
парность как все элементы половой системы, но в дальнейшей эволюции происходит
срастание мюллеровых протоков с образованием одного влагалища и матки, которая
может быть двойной, как у многих грызунов, либо двураздельной, как у хищных, либо
двурогой насекомоядных и китообразных, либо простой как у приматов и человека.
Соответственно дифференцировкам мюллерова канала, у пресмыкающихся и
млекопитающих
развиваются
копулятивные
органы.
У
большинства
пресмыкающихся, а также у сумчатых они парные. У плацентарных, имеющих одно
влагалище, копулятивный орган непарный, но образуется он в результате срастания
парных зачатков.
В эмбриогенезе человека также закладываются парные вольфовы и мюллеровы
каналы. Позже в зависимости от пола происходит их редукция, но со своеобразными
особенностями. Рудимент мюллерова канала у мужчин располагается в
предстательной железе и называется мужской маточкой – utriculus masculinus.
Канальцы передней части первичной почки у них вступают в связь с семенниками и
преобразуются в придатки семенника – эпидидимис. Иногда кроме канальцев,
превращающихся в эпидидимис, остается несколько рудиментальных канальцев
туловищной почки – парадидимис, склонных к патологическому перерождению. У
плодов женского пола возможно нарушение редукции вольфовых каналов, которые
располагаются по бокам от влагалища. Это нарушение может привести к образованию
кист и злокачественных опухолей.
87.
Таким образом, у высших хордовых животных, в том числе и у человека, впроцессе эволюции половой системы произошли изменения в следующих
направлениях:
во-первых, в сторону специализации половых желез у самок и самцов,
во-вторых, к установлению тесной связи с различными частями
выделительной системы,
в-третьих, к окончательному переходу от наружного оплодотворения к
внутреннему.
88.
Аномалии и пороки развития половой системы.1. Ovotestis – нарушение дифференцировки половых желез, в результате чего в
половой железе сочетаются элементы семенника и яичника. У детей, имеющих
ovotestis обнаруживаются признаки гермафродитизма и в наружных половых органах.
2. Крипторхизм – неопущение яичек в мошонку из брюшной полости у плодов
мужского пола (к 8-му месяцу внутриутробного развития). Яички обычно в этом
случае бывают недоразвитыми, а часть семявыносящих каналов заменена
соединительной тканью. Для предотвращения бесплодия мальчикам в раннем возрасте
необходимо хирургическое низведение яичек в мошонку.
3. Utriculum masculinus (мужская маточка) – рудимент мюллерова канала у
мужчин, располагающийся в предстательной железе.
В норме во время эмбриогенеза у организмов мужского пола закладываются
вольфовы и мюллеровы каналы. Позже мюллеровы каналы у них редуцируются. Если
они не редуцируются полностью, тогда возникает данная аномалия.
4. Эпидидимис – придаток семенника, образованный у организмов мужского пола
за счет вступления в связь канальцев первичной почки (mezonephros)с семенниками.
5. Парадидимис – рудиментарные канальцы туловищной почки (каудальнее
эпидидимиса), которые склонны к патологическому перерождению.
6. Нарушение редукции вольфова протока у плодов женского пола. Эта аномалия
опасна возможностью образования кист и злокачественных перерождений.
89.
7. Агенезия – полное отсутствие матки, маточных труб и влагалища,связанные с недоразвитием мюллерова протока.
8. Гипоплазия матки – рудиментарная матка.
9. Атрезия – отсутствие канала или отверстия матки, труб, влагалища,
связанные с реканализацией мюллеровых протоков.
10. Удвоение матки (1 случай на 1000 перинатальных вскрытий). Данная
аномалия развивается в результате нарушения срастания мюллеровых канадов во
время эмбриогенеза.
11. Отсутствие или недоразвитие, или неправильное расположение (эктопия)
мужских половых органов – придатка яичка, семяпровода и семенных пузырьков.
12. Удвоение полового члена – нарушение срастания парных зачатков
полового члена в эмбриогенезе человека.