




























 medicine
medicineSimilar presentations:










Imunitatea adaptativa. Generalitati. Antigenele
1.
IMUNITATEA ADAPTATIVĂ.ANTIGENELE.
2.
Imunitatea adaptativă / dobândită secaracterizează prin:
1.
2.
3.
4.
5.
6.
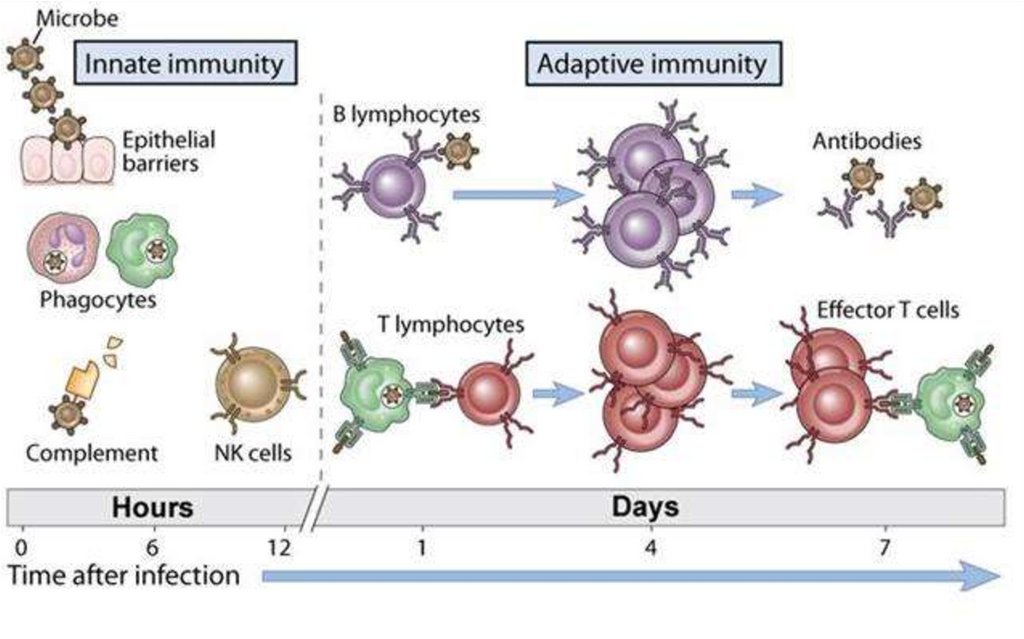
Dezvoltare lentă și manifestare tardivă (câteva zile,
săptămâni dupa contactul cu un antigen)
Specificitate faţă de antigen (capacitatea de a
recunoaște și raspunde specific la numeroase substanțe
străine, inclusiv agenți infecțioși)
Diversitatea răspunsurilor imune elaborate
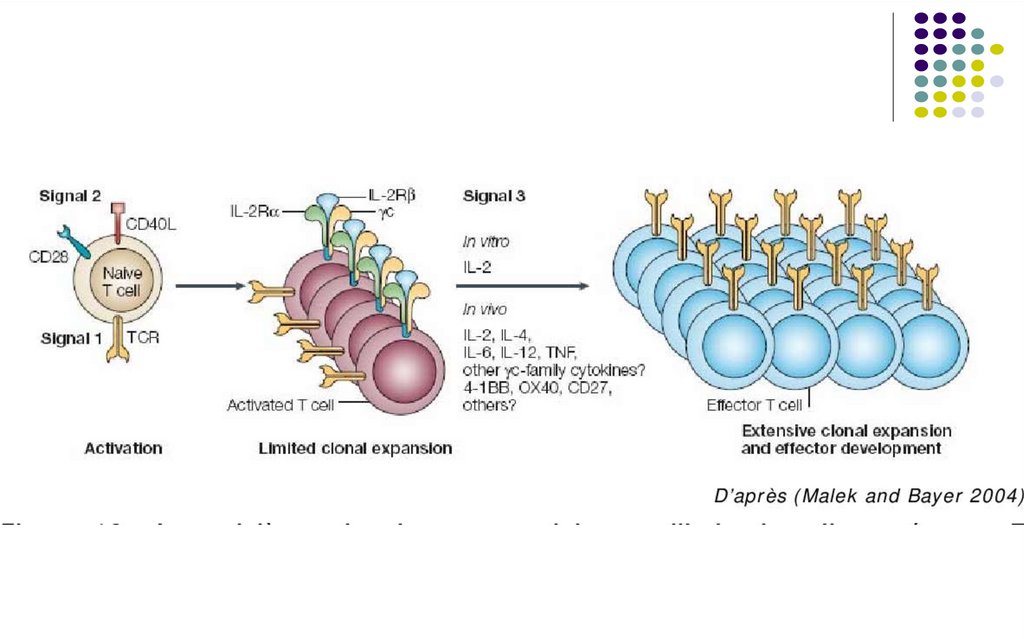
Expansiunea clonală a limfocitelor activate de antigene
Memorie imunologică (capacitatea de a elabora răspuns
mai rapid, mai intens și eficace la întâlniri repetate cu un
antigen)
Toleranţă (lipsa reactivităţii) faţă de antigenele proprii
3.
TIPURILE IMUNITĂȚII ADAPTATIVE1.
2.
Naturală
- Activă (postinfecţioasă)
- Pasivă (Ac materni: transplacentar, prin
laptele matern)
Artificială
- Activă (postvaccinală)
- Pasivă (administrarea Ac / seruri imune sau
a limfocitelor)
4.
Imunitatea dobândită poate fi:Antibacteriană, antivirală, antimicotică, antitoxică,
antitumorală, antitransplant
În funcţie de mecanismele reacţiilor imune
-
-
Imunitate umorală, exercitată prin intermediul unor
glicoproteine numite anticorpi (Ac, Ig), produse de
limfocitele B. Fiind secretate în sange și lichide biologice
neutralizează și elimină microbii extracelulari și toxinele lor.
Imunitate celulară, eficientă in eliminarea paraziților
intracelulari și a celulelor tumorale. Este exercitată prin
intermediul limfocitelor T (citotoxicitate directă, activarea
macrofagelor).
5.
6.
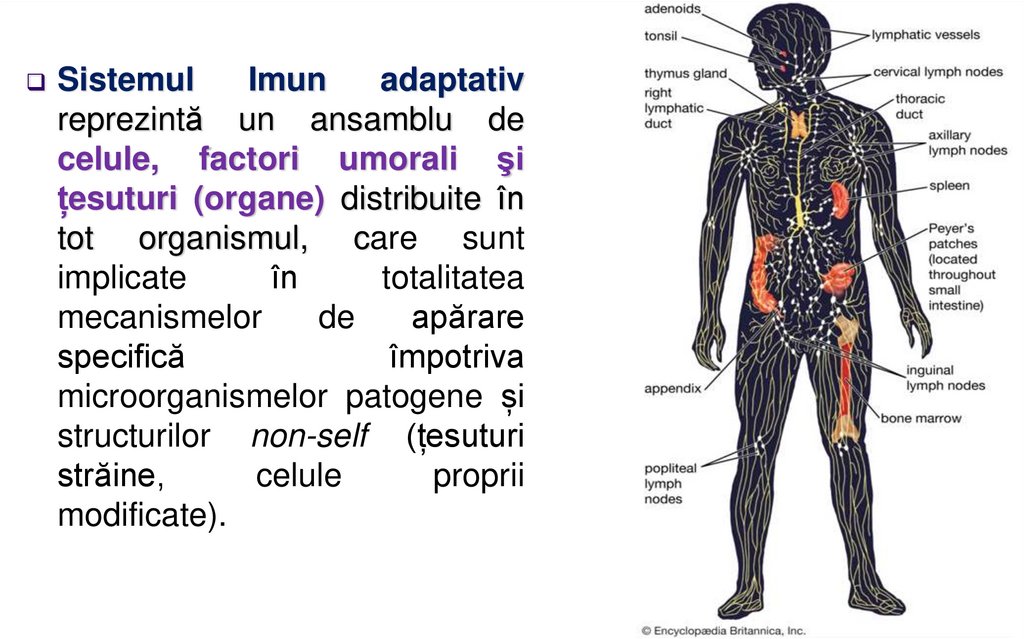
SistemulImun
adaptativ
reprezintă un ansamblu de
celule, factori umorali şi
țesuturi (organe) distribuite în
tot organismul, care sunt
implicate
în
totalitatea
mecanismelor
de
apărare
specifică
împotriva
microorganismelor patogene și
structurilor non-self (țesuturi
străine,
celule
proprii
modificate).
7.
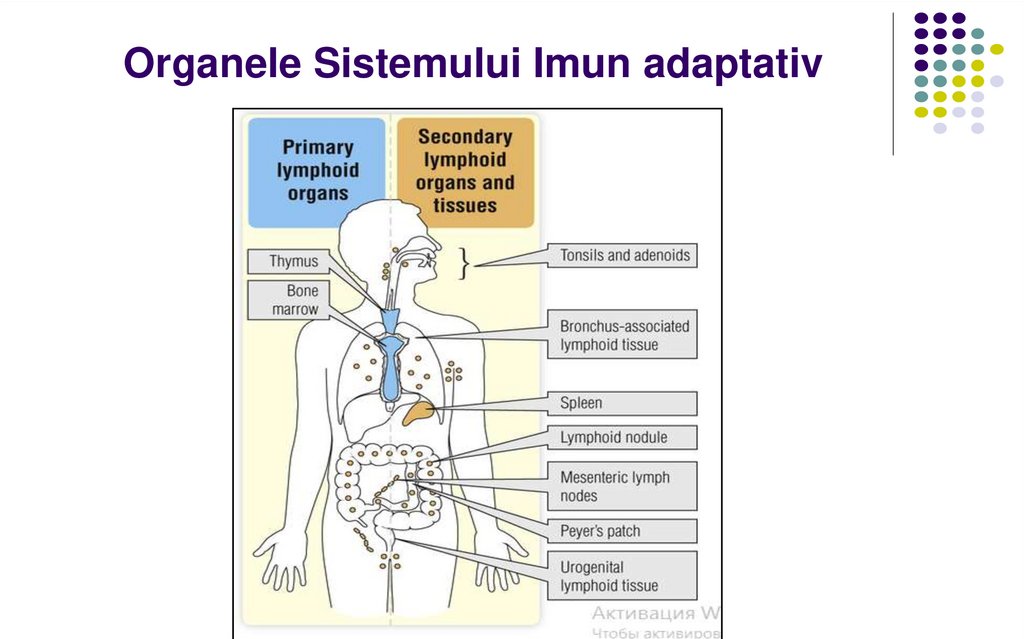
Organele Sistemului Imun adaptativ8.
Organele centrale (primare) ale SI –măduva osoasă şi timusul la vertebrate
(ficatul în perioada embrionară).
Apar primele în timpul vieţii embrionare.
Rolul – sursa celulelor-stem, “instruirea”,
maturizarea şi selecția celulelor
imunocompetente (limfocitele T şi B).
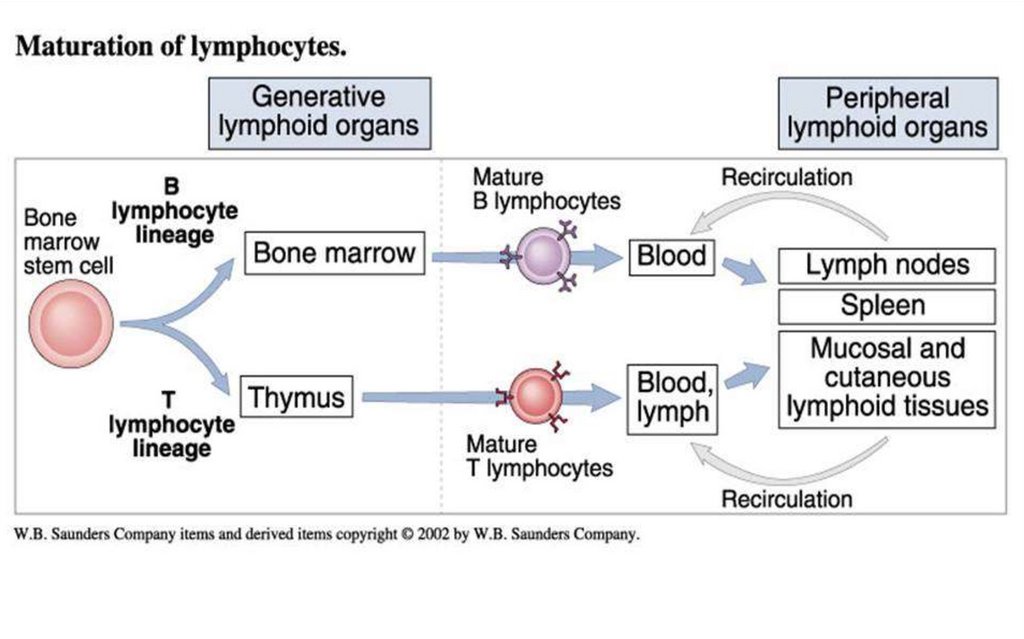
9.
În măduva osoasă se află celulele-stem, pluripotente,precum și precursori ai T şi B-limfocitelor. De asemenea,
măduva osoasă este și sediul unde celulele B se
diferențiază, dobândesc receptori specifici pentru
antigen (BCR), și unde celulele B autoreactive sunt
eliminate prin selecție negativă.
Timusul este un organ limfoid primar implicat în maturarea
celulelor T. Pre-T limfocitele migrează în timus, unde va
avea loc instruirea, diferențierea şi maturizarea lor,
devenind capabile să recunoască specific un singur Ag
prin achiziţionarea unor receptori specifici (TCR).
După părăsirea organelor centrale limfocitele nu mai revin
aici, ele se stabilesc în organele limfoide secundare,
recirculând prin sânge, limfă.
10.
11.
Organele periferice (secundare) ale SIÎn ele se realizează contactul dintre antigen, celulele
prezentatoare de antigen ( CPA) şi celulele
imunocompetente, cu inducerea unui raspuns imun.
Organele limfoide secundare sunt poziționate strategic
pentru a permite captarea de antigen la diferite porți de
intrare, cum ar fi tegumentul, sângele sau membranele
mucoase.
Ganglionii limfatici (raspuns imun contra Ag ce
penetrează prin tegument / mucoase și sunt transportate
cu limfa)
Splina (raspuns imun contra Ag transportate cu sângele)
Țesutul limfoid asociat mucoaselor (MALT): amigdalele,
plăcile Peyer, apendicele, etc.
12.
CELULELE SISTEMULUI IMUN ADAPTATIV1.
2.
3.
Celulele prezentatoare de Ag (CPA) –
macrofagele, celulele dendritice, limfocitele B, dar
și orice celulă nucleată a organismului. CPA
realizează captarea, procesarea și prezentarea Ag
asociate cu molecule CMH limfocitelor T.
Limfocitele T, B (celule imunocompetente,
capabile să recunoască Ag, cu activarea,
proliferarea și diferențierea lor ulterioară în celule
efectoare)
Celulele efectoare (plasmocite, Th (helper), Tc
(citotoxic) - eliminarea microbilor și altor structuri
străine, Treg).
13.
Limfocitul reprezintă celula centrală a sistemului imunadaptativ.
Limfocitele T și B poartă pe suprafața lor receptori specifici
(TCR, BCR), care permit fiecărei celule să reacționeze cu un
anumit antigen. Aceasta constituie baza specificității
imunologice.
În imunitatea adaptativă, receptorii specifici agenților patogeni
sunt „dobândiți” pe parcursul vieții organismului (în timp ce în
imunitatea înnăscută receptorii specifici agenților patogeni sunt
deja codificați în genom).
Limfocitele se divid după stimularea antigenică și formează
clone celulare cu specificitate identică.
Unele limfocite au durată de viață lungă reținând informația
antigenică timp îndelungat. Aceasta este baza memoriei
imunologice.
14.
15.
Recunoaștere specifică a structurilor străine(discriminare self/non-self) în cadrul imunității
adaptative
În timp ce imunitatea înnăscută este capabilă să
discrimineze între diferite clase de
microorganisme, de exemplu, exercitând un
răspuns diferit la o infecție bacteriană sau virală,
sistemul imun adaptativ este capabil să distingă
tulpini foarte asemănătoare ale aceluiași
microorganism, prin recunoașterea structurilor
numite antigene (Ag).
16.
ANTIGENELEANTIGENE – substanţe străine (non-self) de natură endo- sau
exogenă capabile să declanşeze un răspuns imun specific.
Se disting:
Ag de origine infecţioasă (bacterii, virusuri, fungi,
protozoare, etc)
Ag de origine neinfecţioasă (alimente, medicamente, polen,
substanţe chimice, cosmetice, țesuturi, etc.)
Proprietăţile de bază ale Ag:
1.
Antigenitatea (specificitatea) – capacitatea Ag de a
interacţiona specific cu Ac sau cu receptorul pentru Ag
complementar al limfocitelor sensibilizate
2.
Imunogenitatea – capacitatea Ag de a fi recunoscut ca străin
(non-self) şi de a induce răspuns imun specific
17.
Antigenele care posedă ambele caractere sunt Agcomplete.
Ag complete:
Sunt substanţe străine
Au greutate moleculară de peste 10 kDa
Au structură chimică complexă (proteine,
polizaharide, LPZ, etc)
Sunt formate din gruparea carrier (transportator) şi
epitopi (determinante antigenice)
18.
19.
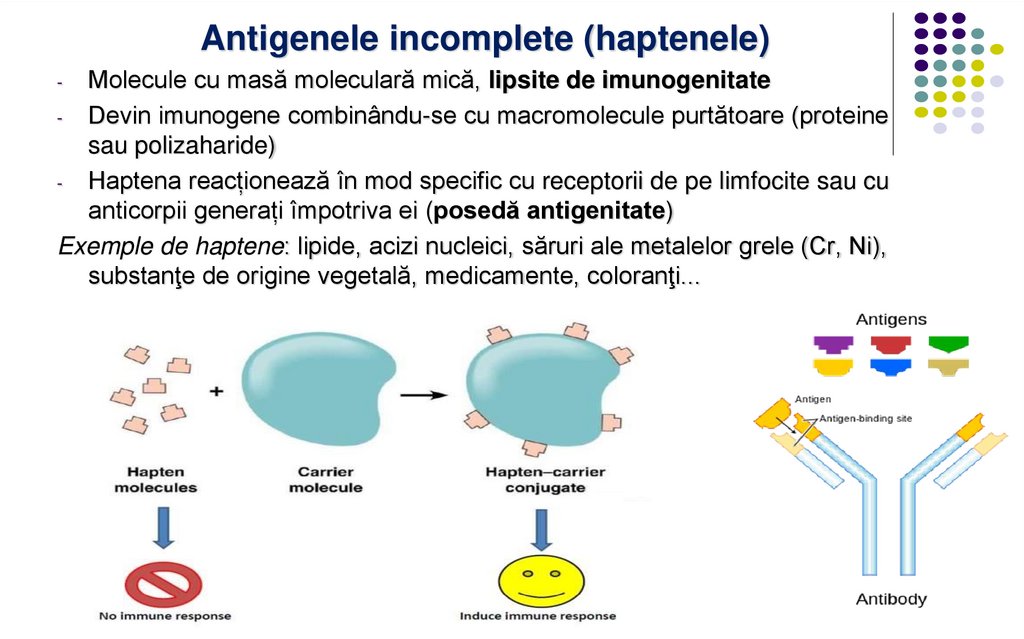
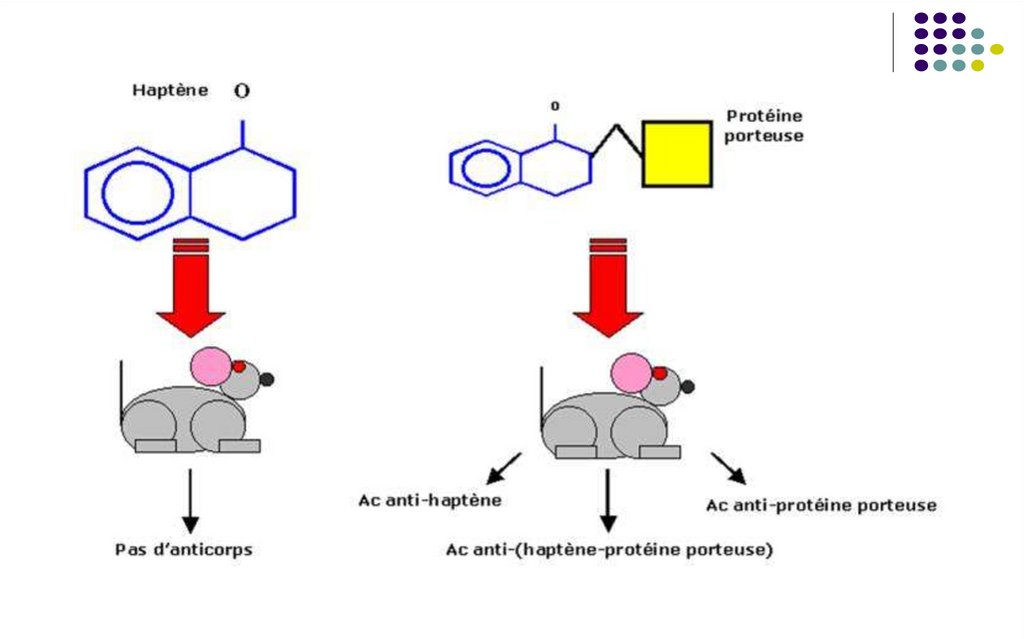
Antigenele incomplete (haptenele)Molecule cu masă moleculară mică, lipsite de imunogenitate
- Devin imunogene combinându-se cu macromolecule purtătoare (proteine
sau polizaharide)
- Haptena reacționează în mod specific cu receptorii de pe limfocite sau cu
anticorpii generați împotriva ei (posedă antigenitate)
Exemple de haptene: lipide, acizi nucleici, săruri ale metalelor grele (Cr, Ni),
substanţe de origine vegetală, medicamente, coloranţi...
-
20.
21.
Tipurile de antigeneAg solubile: proteine plasmatice, toxine, enzime,
hormoni, etc
- Ag corpusculare (celulare, figurate): celule, bacterii,
paraziţi, etc.
- Exoantigene, provenite din exteriorul organismului și
procesate în fagolizozomi (bacterii, fungi,
protozoare...) .
- Endoantigene, provenite din interiorul organismului,
reprezintă proteine proprii modificate sau proteine
virale sintetizate de celulele macroorganismului,
procesate în proteazomi.
-
22.
În funcţie de provenienţă se deosebesc:- Ag heterofile – Ag comune mai multor specii de animale.
Ex.: Ag polizaharidice Forssman, prezente în hematiile
de cal, câine, oaie, cobai; sistemul Rh al eritrocitelor se
întâlneşte la om şi maimuţele Macaccus rhesus, etc
- Ag heteroloage (xeno-Ag, hetero-Ag) – Ag provenite din
organismul altei specii
- Izo-Ag (alo-Ag) – Ag de grup în cadrul unei specii (sistemul
AB0 şi Rh, clasele de Ig)
- Antigene leucocitare/de histocompatibilitate (Human
leukocyte antigen - HLA). Antigenele specifice unui individ.
Corespund moleculelor CMH, participă la recunoașterea
Ag și elaborarea răspunsului imun.
- Autoantigene – Ag proprii unui organism, devenite
imunogene în anumite condiţii (cristalinul ochiului, ţesutul
nervos, țesuturi denaturate etc).
23.
Alergen – orice substanță care provoacă o reacție dehipersensibilitate sau răspuns imun anormal la persoane care au
fost sensibilizate la ele.
Antigenele tumorale sunt markeri aflați la suprafața tumorilor:
antigene asociate tumorilor (TAA), antigene specifice tumorilor
(TSA), neoantigene sau antigene oncogene.
Superantigen – molecule proteice particulare (enterotoxinele
stafilococice, toxina şocului toxic stafilococic, toxina exfoliativă a
stafilococilor, nucleocapsida virusului rabic), capabile să ducă la
activarea policlonală a celulelor T și eliberarea masivă de citokine.
SuperAg provoacă reacţii imunopatologice.
24.
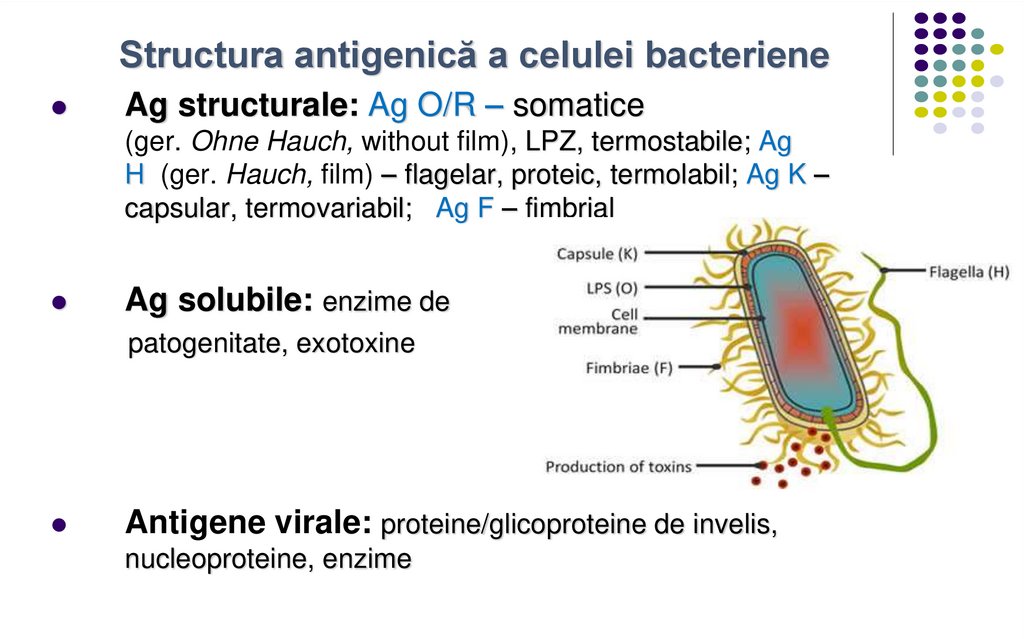
Structura antigenică a celulei bacterieneAg structurale: Ag O/R – somatice
(ger. Ohne Hauch, without film), LPZ, termostabile; Ag
H (ger. Hauch, film) – flagelar, proteic, termolabil; Ag K –
capsular, termovariabil; Ag F – fimbrial
Ag solubile: enzime de
patogenitate, exotoxine
Antigene virale: proteine/glicoproteine de invelis,
nucleoproteine, enzime
25.
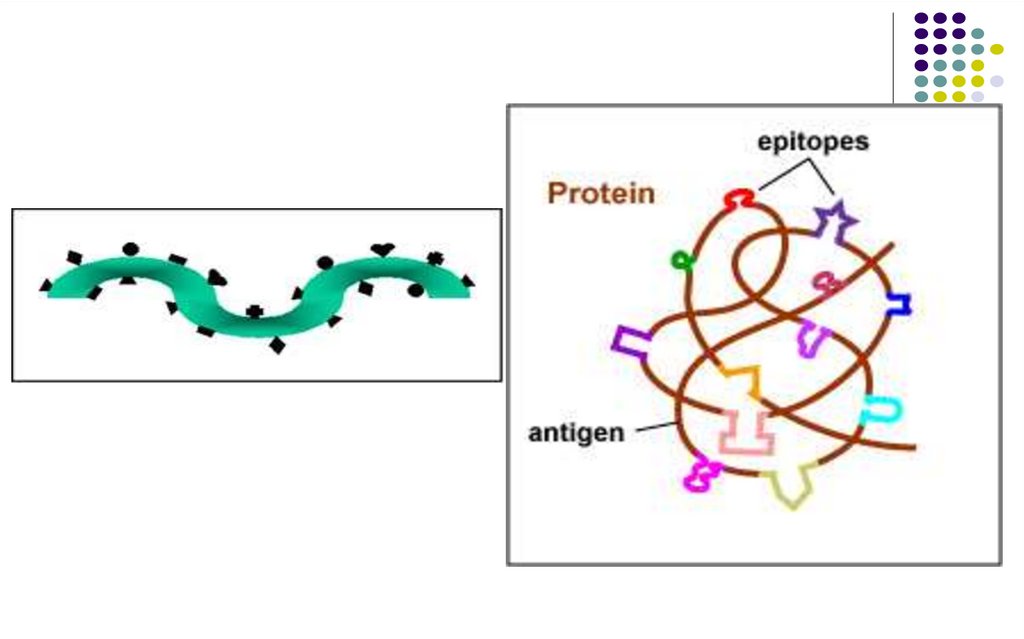
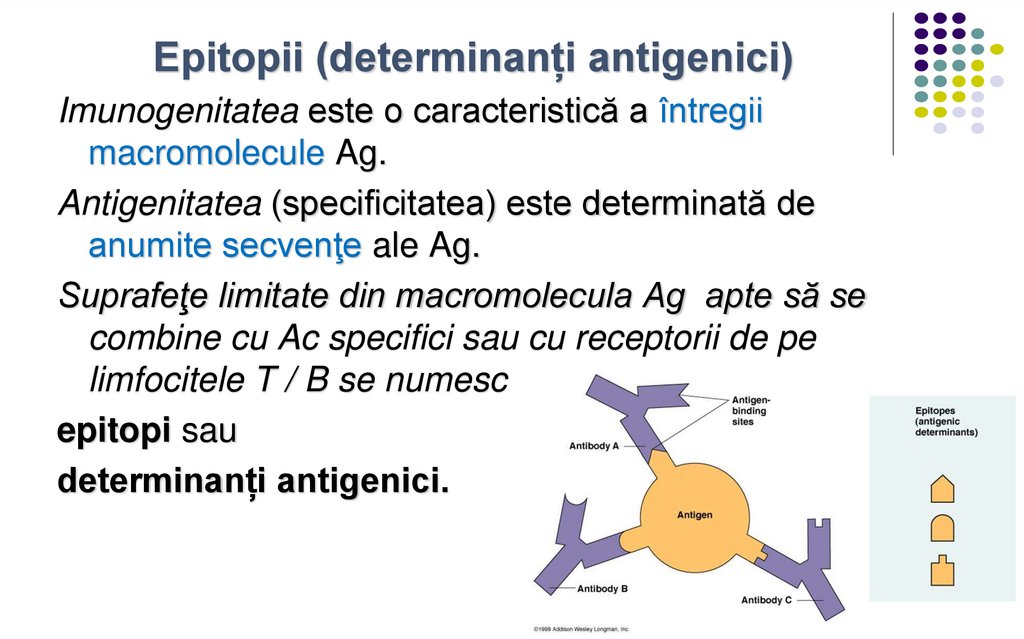
Epitopii (determinanți antigenici)Imunogenitatea este o caracteristică a întregii
macromolecule Ag.
Antigenitatea (specificitatea) este determinată de
anumite secvenţe ale Ag.
Suprafeţe limitate din macromolecula Ag apte să se
combine cu Ac specifici sau cu receptorii de pe
limfocitele T / B se numesc
epitopi sau
determinanți antigenici.
26.
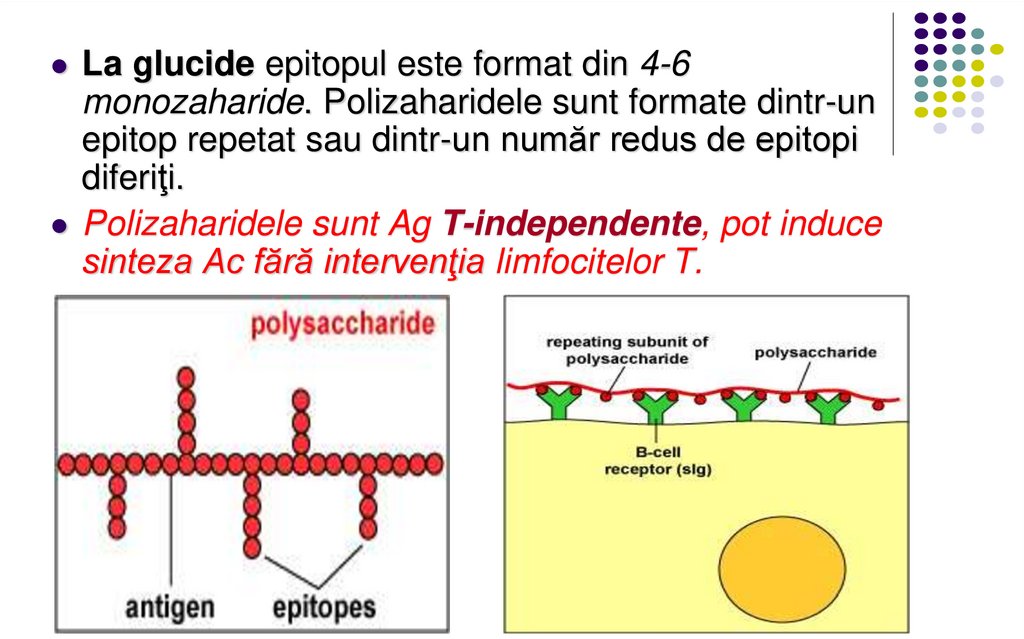
La glucide epitopul este format din 4-6monozaharide. Polizaharidele sunt formate dintr-un
epitop repetat sau dintr-un număr redus de epitopi
diferiţi.
Polizaharidele sunt Ag T-independente, pot induce
sinteza Ac fără intervenţia limfocitelor T.
27.
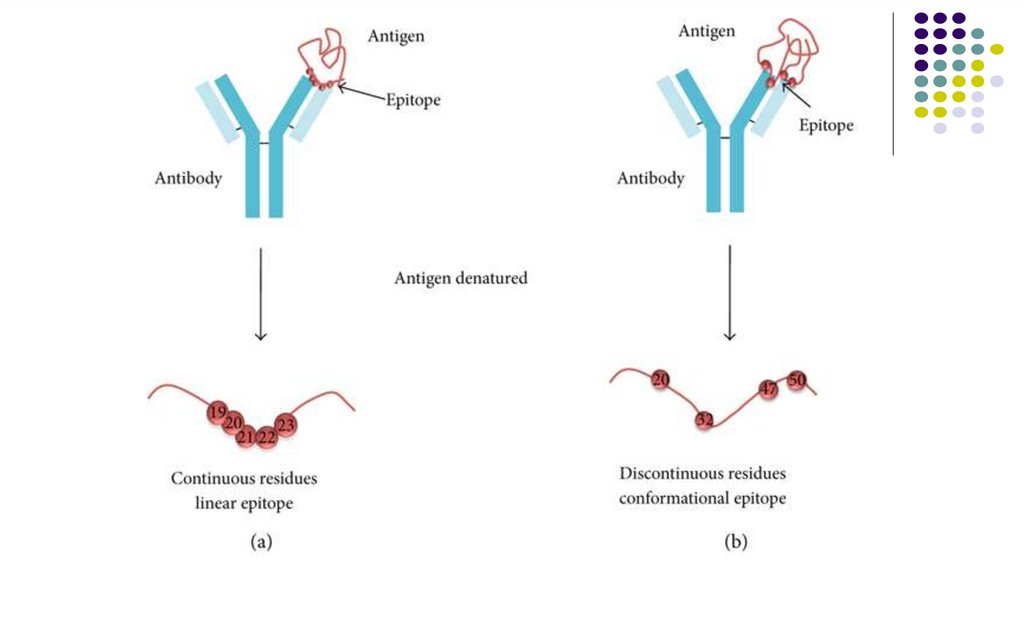
Epitopul proteic este constituit din câţivaaminoacizi (AA).
-
-
Epitopii lineari (secvenţiali) sunt determinaţi de
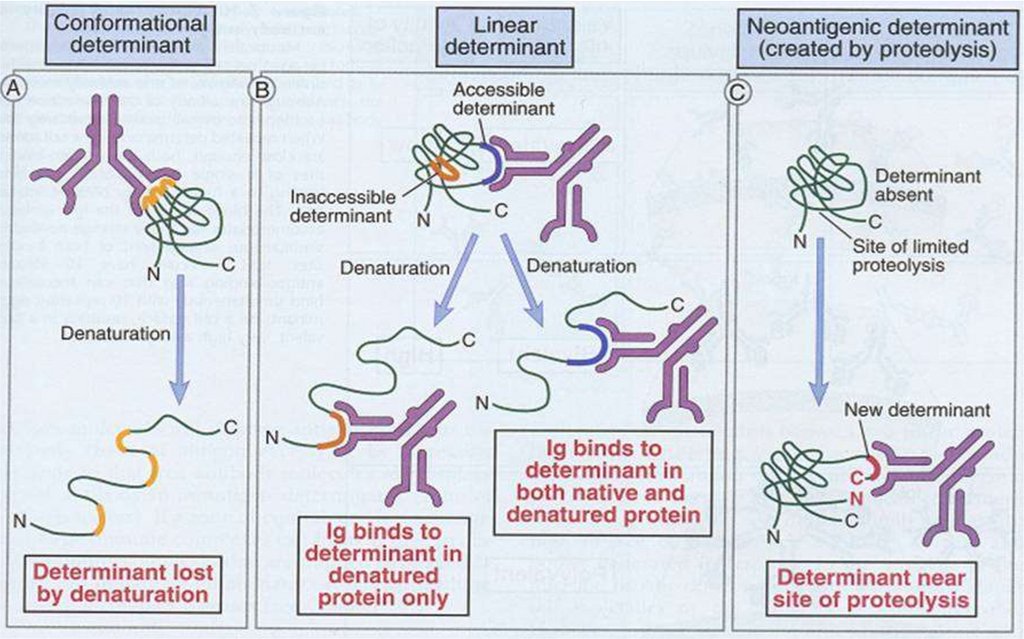
structura primară a moleculei proteice (8-30 AA).
Nu se modifică la denaturarea proteinei.
Epitopii conformaţionali sunt determinaţi de
structura secundară sau terţiară a moleculei
proteice (juxtapoziţia în spaţiu a AA situaţi la
distanţă). Se modifică la denaturarea proteinei.
28.
29.
30.
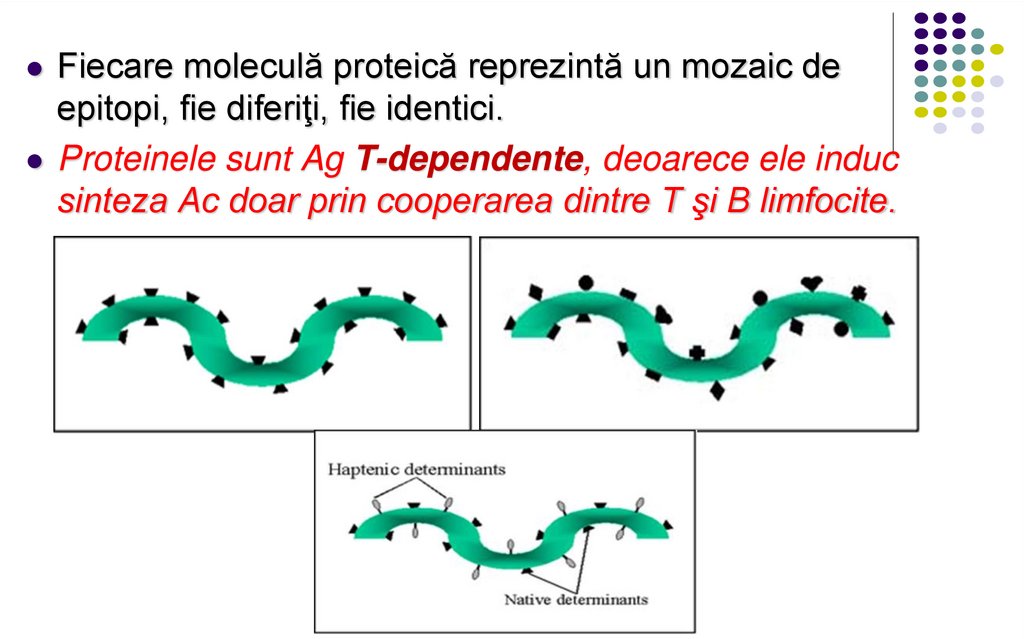
Fiecare moleculă proteică reprezintă un mozaic deepitopi, fie diferiţi, fie identici.
Proteinele sunt Ag T-dependente, deoarece ele induc
sinteza Ac doar prin cooperarea dintre T şi B limfocite.
31.
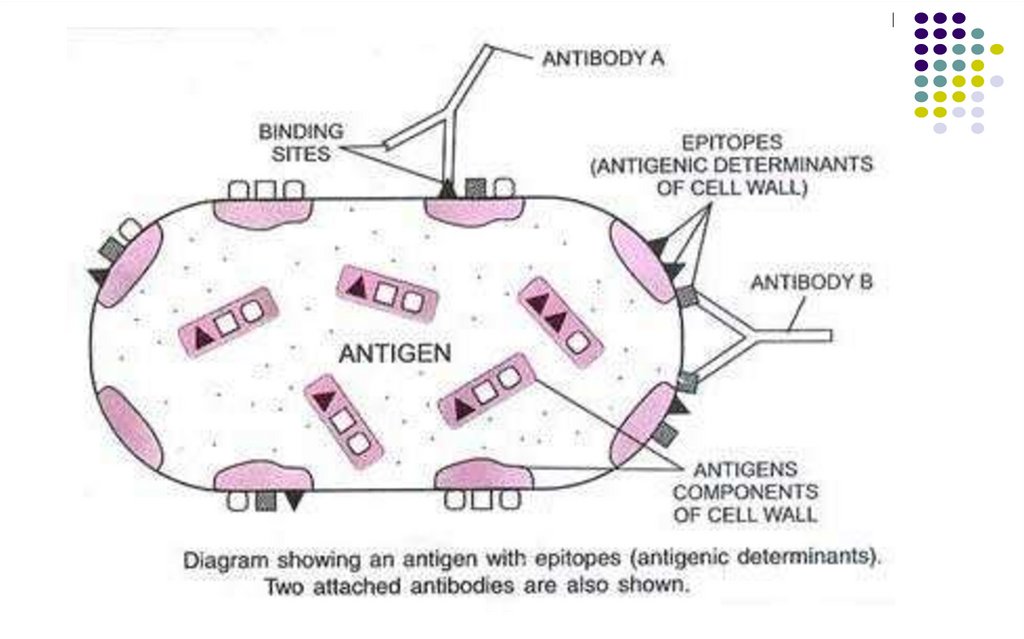
Numărul de epitopi de pe o moleculă imunogenă reprezintăvalenţa Ag.
Un Ag polivalent conține multipli epitopi în compoziția sa
(proteine, polizaharide, LPZ) și pot provoca răspunsuri din
partea unei varietăți de limfocite specifice. O celulă bacteriană
poate conține peste 100 de epitopi.
Unii determinanți antigenici sunt mai eficienți decât alții în
declanșarea unui răspuns imun, probabil deoarece sunt
prezente un număr mai mare de limfocite receptive.
Este posibil ca două sau mai multe substanțe diferite să aibă
un epitop comun. În aceste cazuri, componentele imune
induse de un antigen sunt capabile să reacționeze cu toate
celelalte antigene care poartă același epitop. Astfel de
antigene sunt cunoscute sub numele de antigene cu
reactivitate încrucișată.
32.
33.
Ig prezintă glicoproteine din fracţia γ-globulinelor. Sedisting Ig membranare (BCR) şi Ig solubile (secretate) –
Ac ca atare.
Ac (Ig) sunt molecule glicoproteice produse de
plasmocitele derivate din limfocitele B activate de Ag.
Ac circulă în sânge şi pătrund în ţesuturi. Sunt eficienţi
contra bacteriilor, toxinelor microbiene, helminților şi
virusurilor în poziţie extracelulară.
Ac au proprietatea de a recunoaşte şi a se combina cu
Ag complementar, atât in vivo, cât şi in vitro.
34.
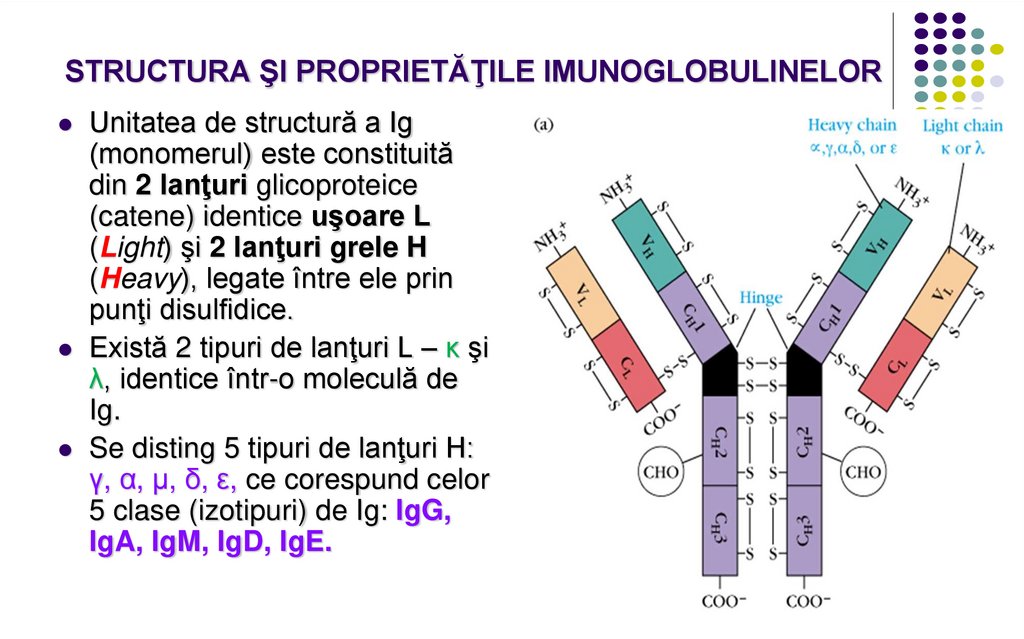
STRUCTURA ŞI PROPRIETĂŢILE IMUNOGLOBULINELORUnitatea de structură a Ig
(monomerul) este constituită
din 2 lanţuri glicoproteice
(catene) identice uşoare L
(Light) şi 2 lanţuri grele H
(Heavy), legate între ele prin
punţi disulfidice.
Există 2 tipuri de lanţuri L – κ şi
λ, identice într-o moleculă de
Ig.
Se disting 5 tipuri de lanţuri H:
γ, α, µ, δ, ε, ce corespund celor
5 clase (izotipuri) de Ig: IgG,
IgA, IgM, IgD, IgE.
35.
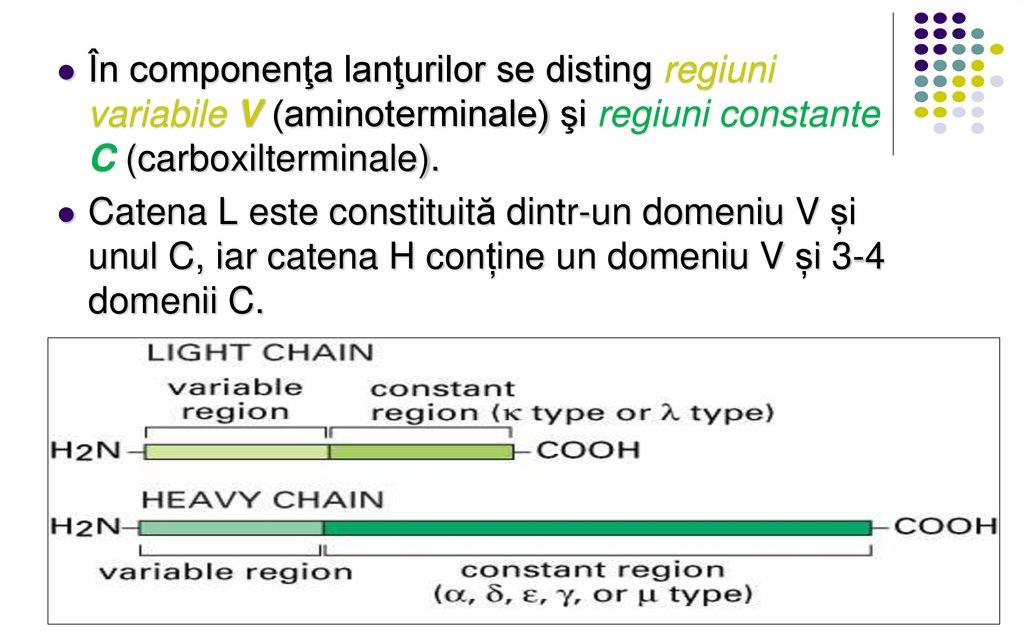
În componenţa lanţurilor se disting regiunivariabile V (aminoterminale) şi regiuni constante
C (carboxilterminale).
Catena L este constituită dintr-un domeniu V și
unul C, iar catena H conține un domeniu V și 3-4
domenii C.
36.
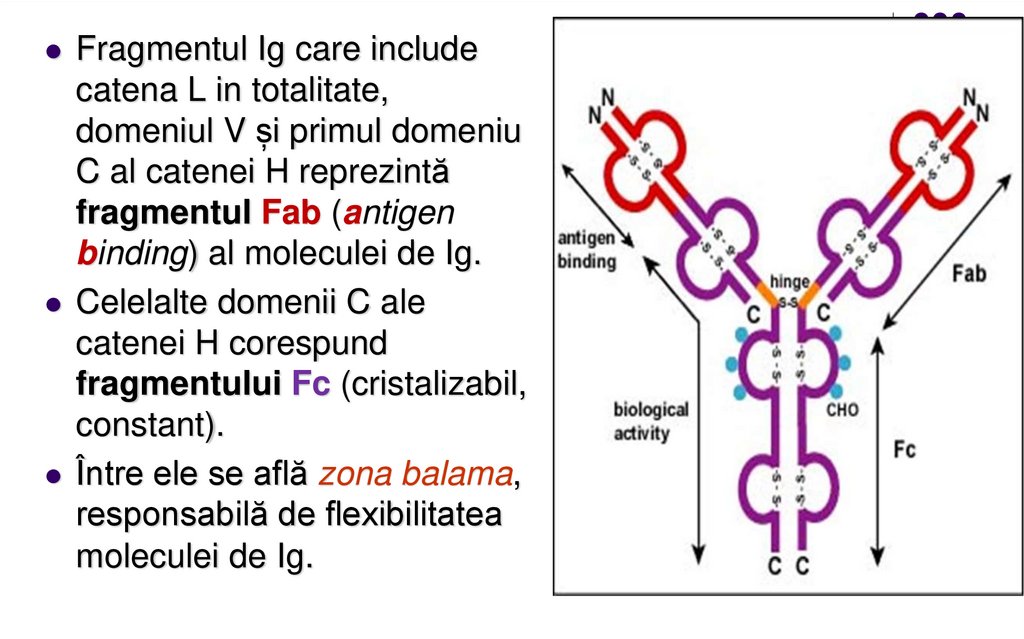
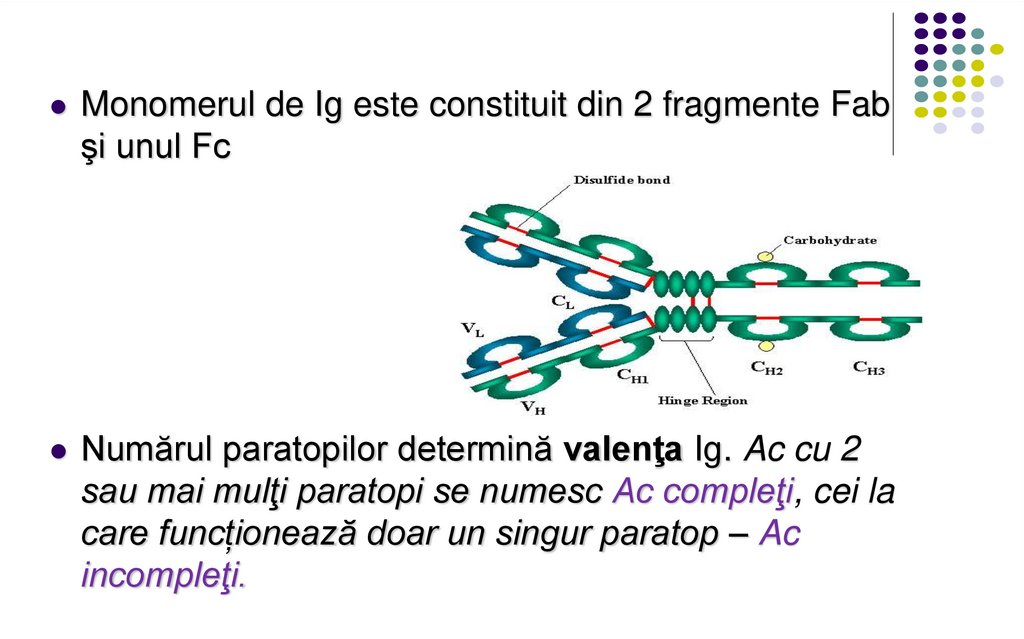
Fragmentul Ig care includecatena L in totalitate,
domeniul V și primul domeniu
C al catenei H reprezintă
fragmentul Fab (antigen
binding) al moleculei de Ig.
Celelalte domenii C ale
catenei H corespund
fragmentului Fc (cristalizabil,
constant).
Între ele se află zona balama,
responsabilă de flexibilitatea
moleculei de Ig.
37.
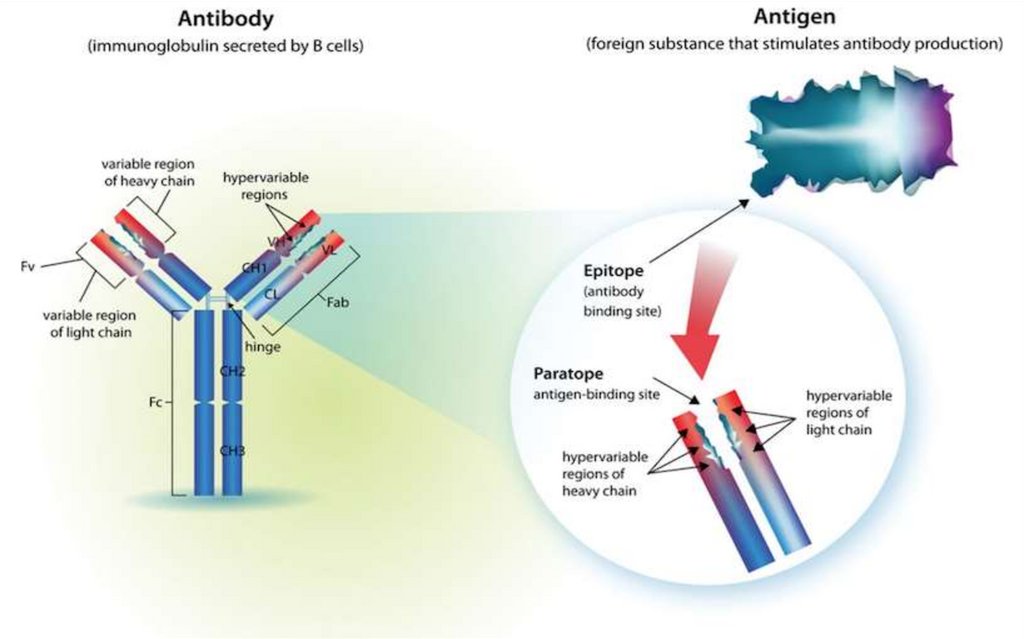
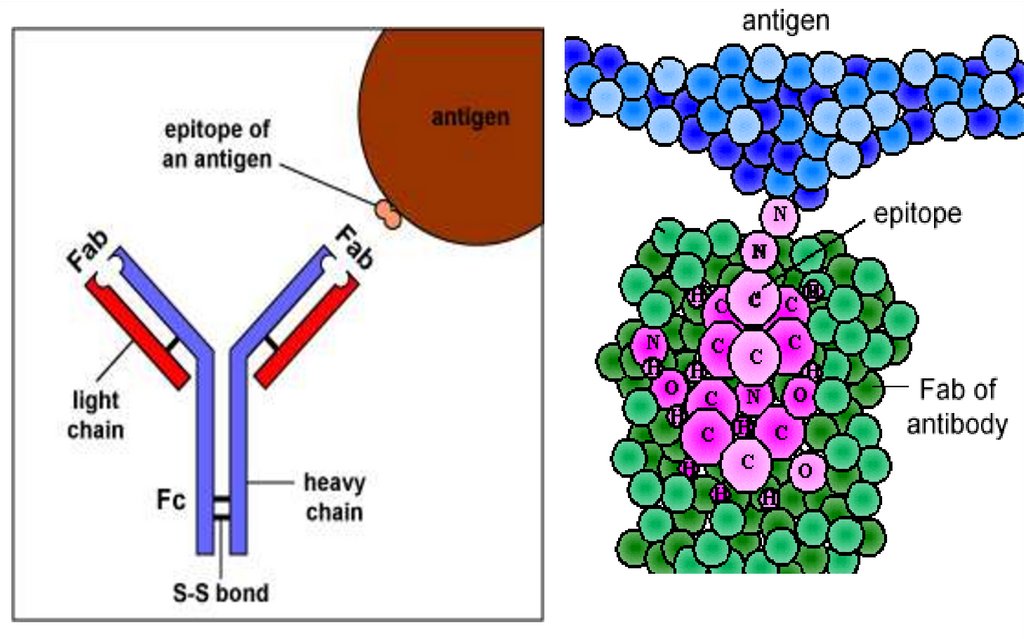
Fiecare regiune variabila VH si VL afragmentului Fab contine zone
hipervariabile, care formeaza o cavitate
tridimensională - paratopul (centrul activ)
moleculei de Ig.
Paratopul va reacţiona specific cu epitopul
(determinanta antigenică) Ag
corespunzător, constituit din 5-7 AA sau 34 reziduuri glucidice.
38.
39.
40.
Fragmentul Fc este purtător de receptori şiresponsabil de activitatea biologică a Ig:
-Transportul transplacentar al unor Ig
-Fixarea pe diferite celule (mastocite, bazofile,
eozinofile, fagocite, limfocite, etc.)
-Capacitatea de a fixa unele fracții ale
complementului (C1, C3b)
-Capacitatea de fixare a proteinei A a stafilococilor
-Defineşte clasele şi subclasele de Ig (specificitatea
antigenică a catenei H)
41.
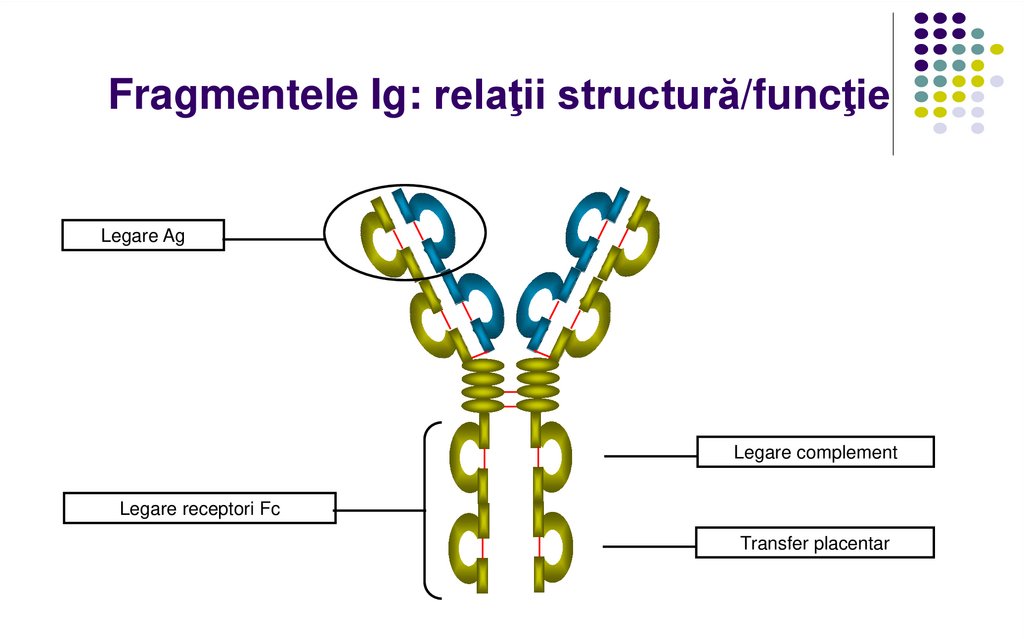
Fragmentele Ig: relaţii structură/funcţieLegare Ag
Legare complement
Legare receptori Fc
Transfer placentar
42.
Monomerul de Ig este constituit din 2 fragmente Fabşi unul Fc
Numărul paratopilor determină valenţa Ig. Ac cu 2
sau mai mulţi paratopi se numesc Ac compleţi, cei la
care funcționează doar un singur paratop – Ac
incompleţi.
43.
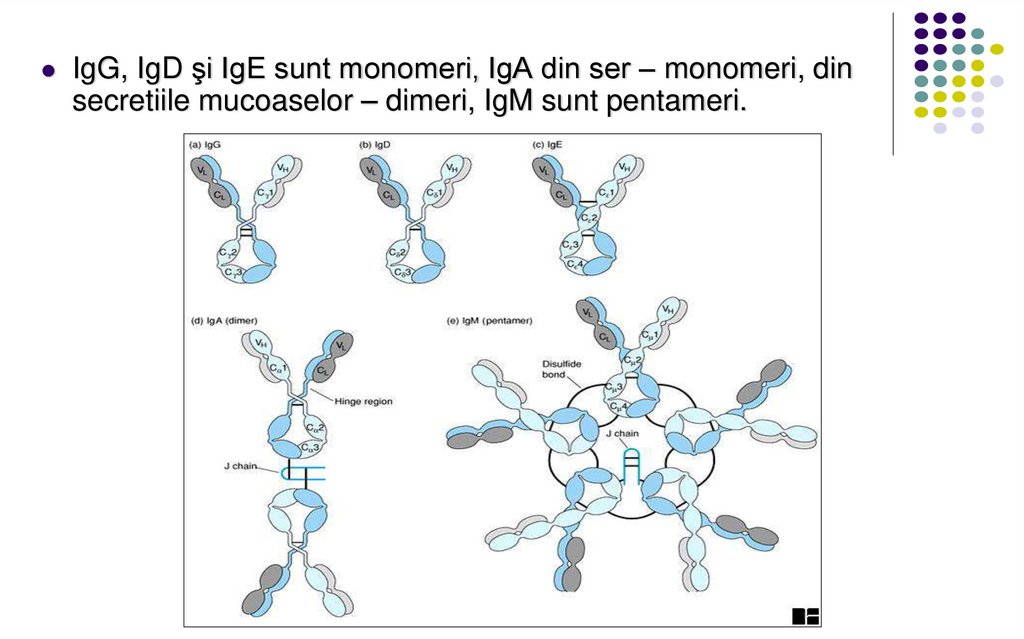
IgG, IgD şi IgE sunt monomeri, IgA din ser – monomeri, dinsecretiile mucoaselor – dimeri, IgM sunt pentameri.
44.
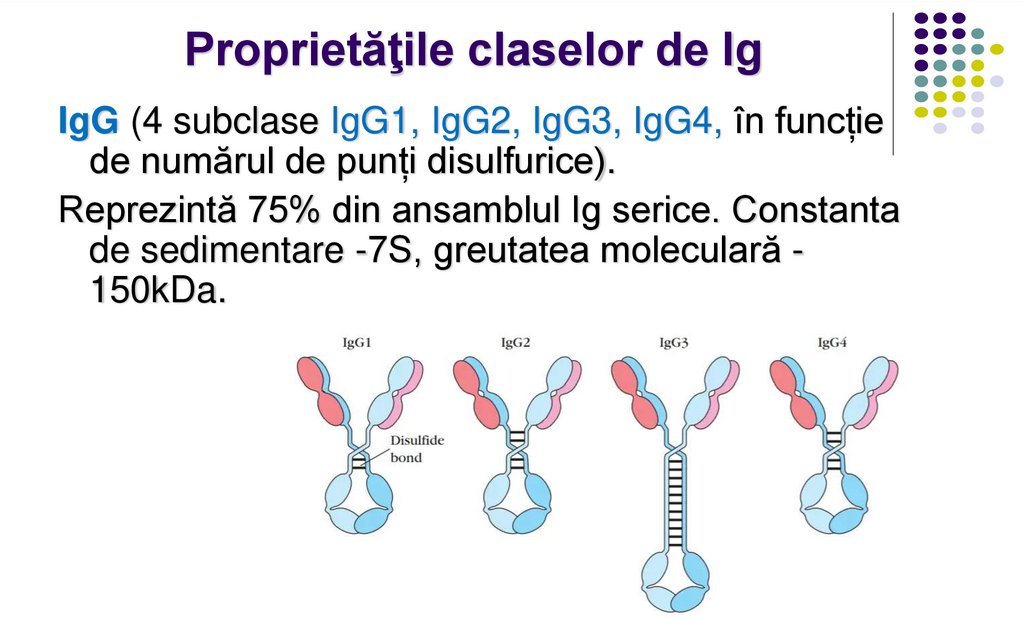
Proprietăţile claselor de IgIgG (4 subclase IgG1, IgG2, IgG3, IgG4, în funcție
de numărul de punți disulfurice).
Reprezintă 75% din ansamblul Ig serice. Constanta
de sedimentare -7S, greutatea moleculară 150kDa.
45.
Funcțiile biologice ale IgGIgG sunt unicele Ig capabile să traverseze bariera placentară
(IgG3, IgG1, IgG4, IgG2) și traversează activ bariera
endotelială.
Fc al IgG are centre de fixare a complementului (activarea C
pe cale clasică), a macrofagelor şi neutrofilelor (rol în
opsonizare), a proteinei A a stafilococilor.
Timpul de înjumătățire 21 de zile.
Manifestă activitate opsonizantă (IgG3, IgG1, IgG4),
neutralizantă (IgG1, IgG3, IgG4), de activare a
complementului (IgG3, IgG1, IgG2).
IgG predomină în infecții cronice și asigură imunitatea postinfecțioasă și postvaccinală.
46.
Ig M – 5-6% din totalul Ig. Pentamer.Constanta de sedimentare - 19S, greutatea
moleculară - 900kDa.
Se distrug sub acţiunea mercapto-etanolului sau
cisteinei.
Timpul de înjumătățire – 5 zile.
Fixează si activează complementul pe cale clasică.
Nu traversează placenta, prezenţa Ig M la nou-născut
denotă infecţie intrauterină.
Sunt primele care apar după un stimul antigenic
primar şi indică un proces infecţios acut.
47.
48.
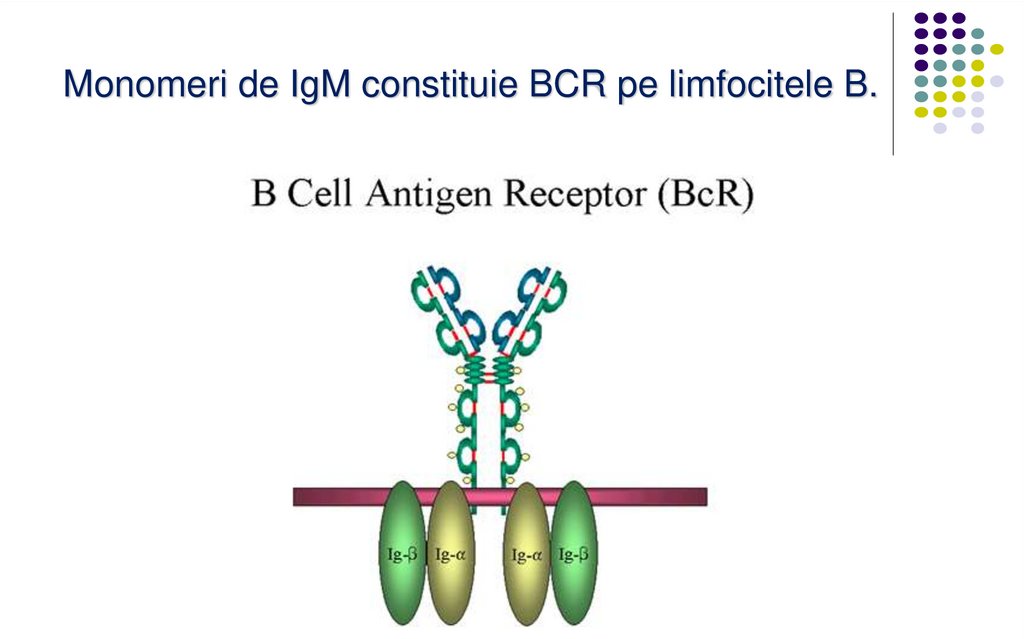
Monomeri de IgM constituie BCR pe limfocitele B.49.
Ig A – 15% din totalul Ig. CS -7S, GM – 160 kDa.Bogate în glucide.
Se disting 2 subclase: IgA1 (80-90% - serice) şi IgA2
(10-20% - secreții).
Semi-viaţa – 6 zile, nu traversează placenta, nu activează
complementul pe cale clasică.
- Ig A serice (monomeri) – 6% din totalul Ig serice.
Agregate de IgA pot activa complementul pe cale
alternativă.
- Ig A secretoare (sIg A) – salivă, lacrimi, colostrum, lapte,
secreţii gastro-intestinale, nazale, bronhice. Dimer.
Asigură protecţia mucoaselor, blocând ataşarea
bacteriilor şi virusurilor la receptorii mucoaselor.
50.
51.
Ig D – 0,2% din totalul Ig. CS – 6,5S, GM – 170 kDa.Semi-viaţa – 3 zile.
Rolul – receptor pentru Ag (BCR) pe LB; posibil participă la
eliminarea limfocitelor B care produc autoAc (Ac
autoreactivi)
Ig E – 0,002 - 0,01%, CS – 7,9S, GM – 185 kDa. Semi-viaţa
– 2-3 zile. Nu traversează placenta, nu fixează
complementul. Termolabile (inactivate la 56°C în 30 min).
Se pot fixa pe suprafaţa mastocitelor şi bazofilelor,
determinând degranularea lor cu eliberarea unor amine
vazo-active (consecinţa - şoc anafilactic, dereglări
alergice).
Eficiente în afecţiuni parazitare (opsonizarea helminţilor şi
protozoarelor, efect chimiotactic pozitiv pentru eozinofile).
52.
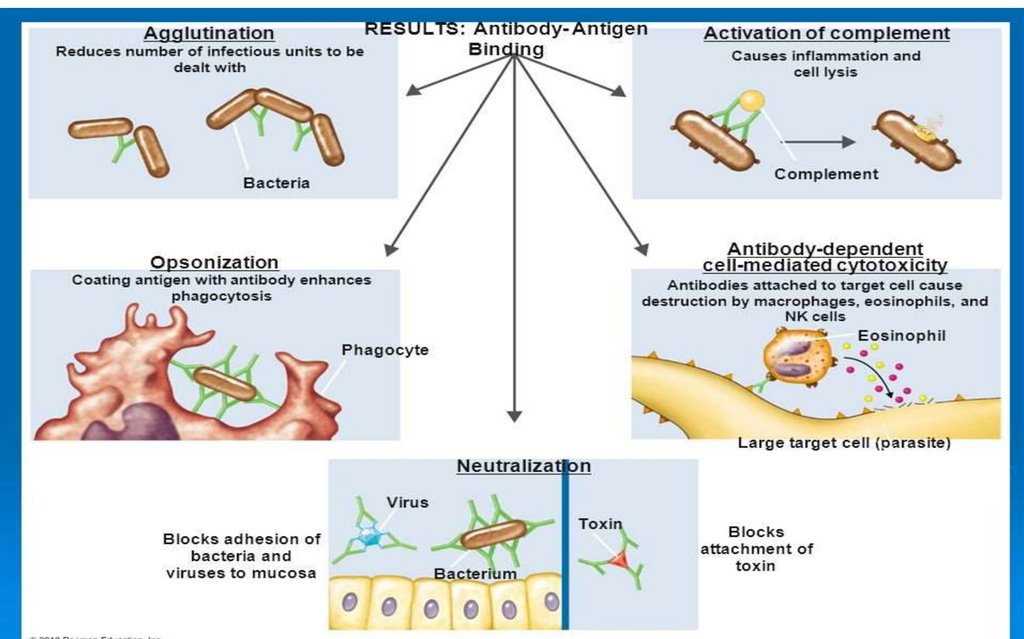
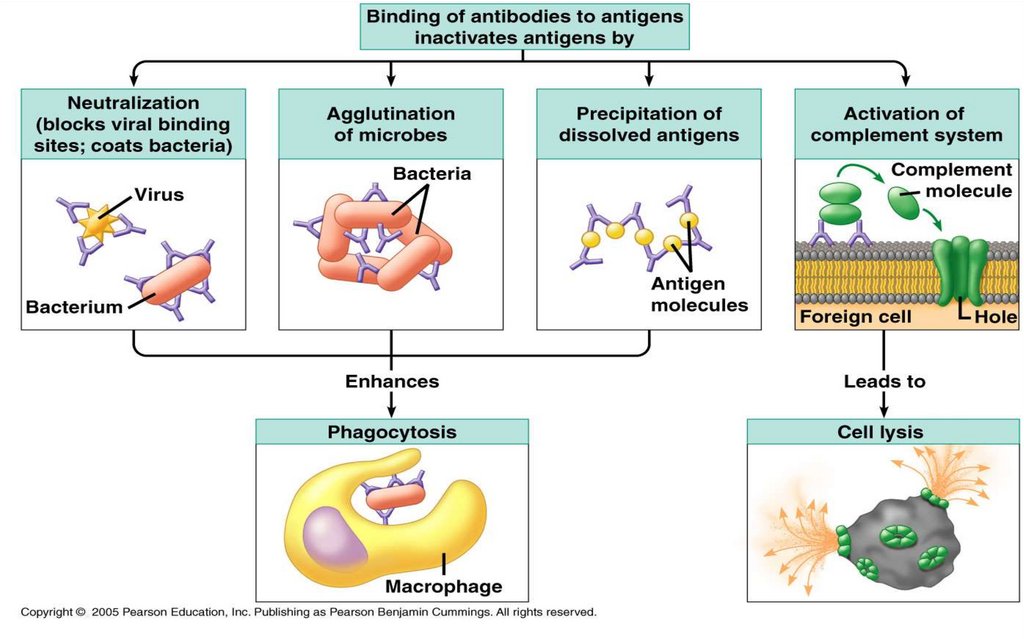
Efectul interacţiunii dintre Ag şi Ac in vivoDepinde de natura Ag şi de tipul de Ac şi se poate manifesta prin:
Aglutinarea bacteriilor (IgM, IgG)
Opsonizarea bacteriilor şi intensificarea fagocitozei (IgG, IgA)
Activarea complementului pe cale clasică determinând citoliză (inclusiv
bacterioliză), opsonizare prin C3b, sau stimularea inflamației prin C3a, C5a
(IgM, IgG)
Neutralizare - blocarea atașării toxinelor/enzimelor, adeziunii bacteriilor,
virusurilor la receptorii celulari (IgM, IgG, IgA)
Citotoxicitatea mediată celular anticorp-dependentă – ADCC
Ac se leagă de patogeni sau celulele infectate, inducând degranularea celulelor
citotoxice:
- IgG stimulează degranularea celulelor NK (consecință - distrugerea celulelor
infectate cu virus sau tumorale)
- IgE stimulează degranularea eozinofilelor (distrugerea helminților)
Degranularea mastocitelor – IgE se leagă de mastocite prin Fc, iar prin Fab
interacționează cu Ag (induce reacții anafilactice)
53.
54.
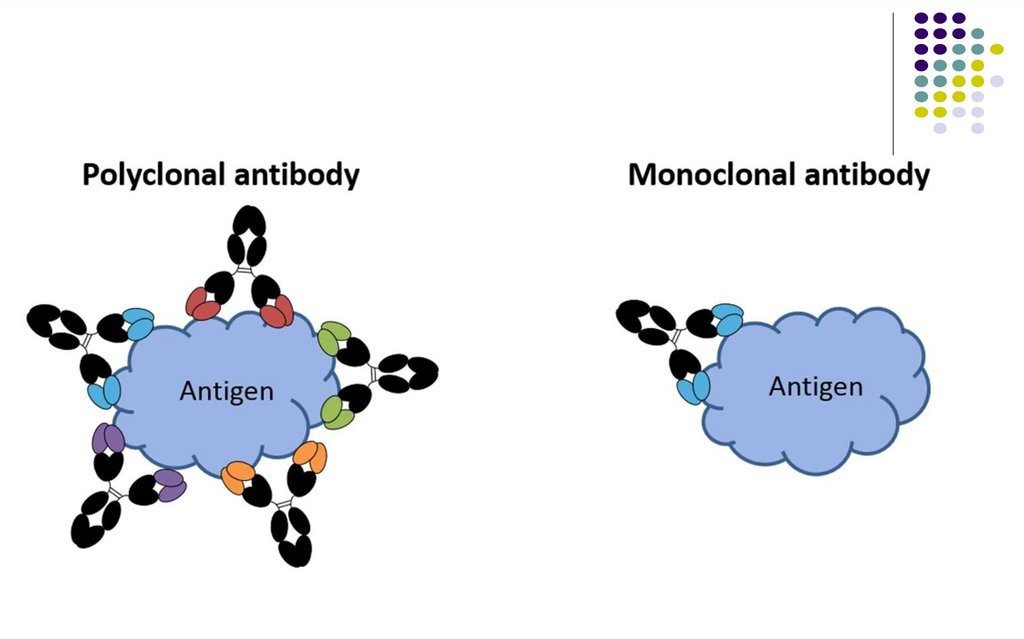
55.
Anticorpii policlonali sunt un grup de anticorpiproduși de către diferite clone de plasmocite și
care au afinitate pentru același antigen, dar se
leagă de epitopi diferiți.
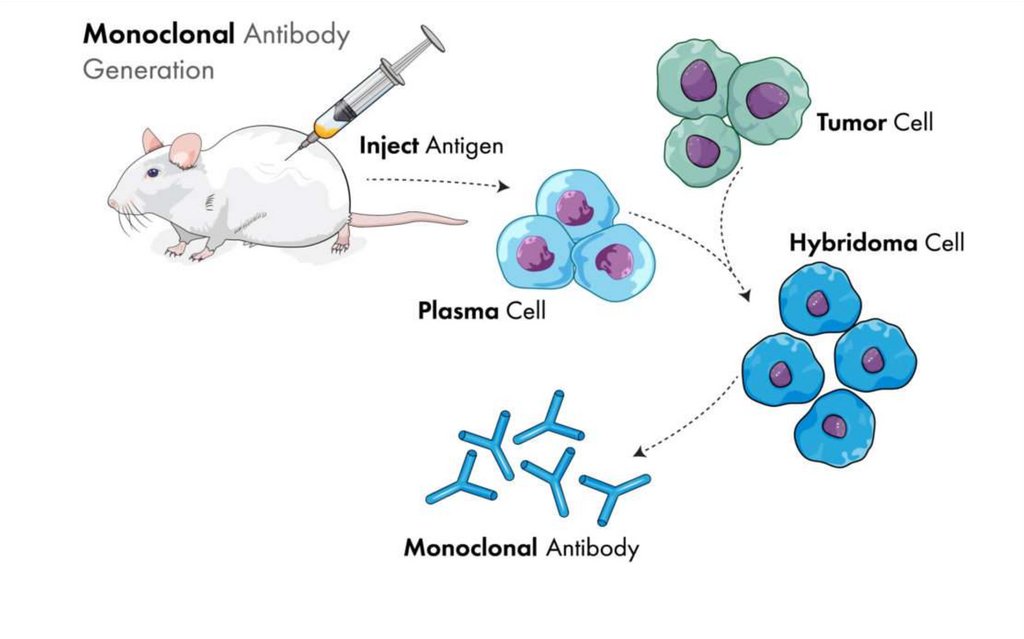
Anticorpii monoclonali sunt un set de anticorpi
identici produși de către o singură clonă de
plasmocite și care recunosc un singur tip de
epitop al unui singur antigen specific.
Provin din hibridoame de origine murină, umană
sau chimerică.
56.
57.
58.
Utilizarea practică a anticorpilorTratamentul și profilaxia unor maladii (boli
infecțioase, procese autoimune,
inflamatorii, cancer...)
Diagnostic (seroidentificare,
serodiagnostic)
Cercetare (imunohistochimie, purificarea
unor structuri, identificarea unor molecule)