









")




")





. СТЕРОЛЫ (СТЕРИНЫ) – стероидные спирты")



")




 и кавеолы")








 chemistry
chemistrySimilar presentations:


")






")
Химия липидов. Строение и функции биологических мембран. Лекция 5
1.
КАФЕДРА МЕДИЦИНСКОЙ БИОХИМИИИ БИОФИЗИКИ
БИОХИМИЯ ЧЕЛОВЕКА
Лекция 5
ХИМИЯ ЛИПИДОВ.
СТРОЕНИЕ И ФУНКЦИИ
БИОЛОГИЧЕСКИХ МЕМБРАН
лектор – Емельянов Виктор Владимирович,
к.м.н., доцент, зав. кафедрой медицинской биохимии и биофизики
30 сентября 2025 г.
2. ЛИПИДЫ:
• Природные органические соединения• Плохо растворимы в воде
• Хорошо растворимы в неполярных
органических растворителях
• Содержат длинные углеводородные
радикалы
3. КЛАССИФИКАЦИЯ ЛИПИДОВ
ЛИПИДЫОМЫЛЯЕМЫЕ
O
C
O
НЕОМЫЛЯЕМЫЕ
СЛОЖНЫЕ ЭФИРЫ
ТЕРПЕНЫ
ТРИГЛИЦЕРИДЫ, ВОСКИ,
ЭФИРЫ СТЕРОИДОВ
СТЕРОИДЫ
ФОСФОЛИПИДЫ
ЖИРНЫЕ К-ТЫ, ВЫСШИЕ СПИРТЫ,
УГЛЕВОДОРОДЫ
ГЛИКОЛИПИДЫ
O
C N
H
ЦЕРЕБРОЗИДЫ,
ГАНГЛИОЗИДЫ
4. ВЫСШИЕ ЖИРНЫЕ КИСЛОТЫ
• Карбоновые кислоты с числом атомовуглерода 10 и более
• Четное число атомов углерода
• Неразветвленные боковые радикалы
• Насыщенные, моно- и
полиненасыщенные
• Ненасыщенные в цис-конфигурации
• Двойные связи не ранее 9 атома
5.
6. ВАЖНЕЙШИЕ ЖИРНЫЕ КИСЛОТЫ
Тривиальноеназвание
Число атомов С,
число и
положение
F-связей
Пальмитиновая
16 : 0
Стеариновая
18 : 0
Олеиновая
18 : 1; 9
ω-9
Линолевая
18 : 2; 9,12
ω-6
Линоленовая
18 : 3; 9,12,15
ω-3
Арахидоновая
20 : 4; 5,8,11,14
ω-6
Структурная формула
COOH
COOH
10
13 12
16 15
15 14
13 12
12
11
10
9
COOH
9
COOH
10
9
9
8
COOH
6
5
COOH
7. ω-3 ω-6
8. ЦИС – ТРАНС ИЗОМЕРИЯ
COOHH C
C
H
COOH
COOH
HOOC
C
H
C
H
9. ЦИС – ТРАНС ИЗОМЕРИЯ
Транс-ненасыщенные ЖК повытянутой форме
углеводородных «хвостов»
аналогичны насыщенным ЖК
Цис-ненасыщенные ЖК имеют
изогнутые углеводородные
«хвосты» и занимают больше
места, чем «хвосты» насыщенных
ЖК
H
H
H
H
COOH
COOH
COOH
COOH
CH3
10. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
1. Структурные компоненты омыляемыхлипидов.
2. Энергетический материал.
3. Пластический материал.
4. Сигнальные молекулы: производные
жирных кислот – ЭЙКОЗАНОИДЫ простагландины, тромбоксаны,
лейкотриены и др.
11. ТРИГЛИЦЕРИДЫ (триацилглицериды, триацилглицеролы)
OO
R2
C
CH2 O C
O
C
R1
O
H
CH2 O C
R3
O
O
O
O
O
O
ЖИРЫ – твердые (кроме рыбьего),
МАСЛА – жидкие (кроме пальмового и масла какао)
12. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
1. Резерв ЖК.2. Теплоизоляция.
3. Механическая
амортизация.
4. Депонирование
жирорастворимых
витаминов.
13. ФОСФОЛИПИДЫ
• I. ФОСФАТИДОВЫЕ КИСЛОТЫ.• II. ФОСФАТИДЫ.
1. Фосфатидилэтаноламины (кефалины).
2. Фосфатидилхолины (лецитины).
3. Фосфатидилсерины.
4. Фосфатидилинозитолы.
• III. СФИНГОЛИПИДЫ.
• IV. ПЛАЗМАЛОГЕНЫ.
• V. КАРДИОЛИПИНЫ.
14. СТРУКТУРА ФОСФАТИДА
OO
CH2O C R1
R2 C O C H O
CH2O P O CH2 CH2
OH
ФОСФАТИДОВАЯ КИСЛОТА
CH
+ 3
N CH3
CH3
АМИНОСПИРТ ХОЛИН
15. СТРУКТУРА ФОСФАТИДА
OC
O
C
O
CH2
O
C
O
O
CH2
H
P
O
CH2
O
+
CH2
N
H3C
CH3
H3C
Неполярный хвост
Фосфатидилхолин ( лецитин)
Полярная
часть
16. ФОСФАТИДИЛХОЛИН (МОДЕЛЬ СТЮАРТА)
R1R2
ГИДРОФИЛЬНАЯ
ОБЛАСТЬ
Х
17. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
1. Структурное: главная составнаячасть мембран, липосом,
липопротеинов.
2. Регуляторное.
3. Пластическое.
18. ФОСФОЛИПИДЫ В ВОДЕ самоорганизуются без добавления эмульгатора
Ламеллярная (бислойная) структура19. СФИНГОЛИПИДЫ
сфингозинH3C (CH2)12 C
H
OH
C
CH CH NH C R
H
остаток
фосфорной
кислоты
O
CH2
O
O
жирная кислота
P
_
O
O
CH2 CH2 N+ CH3
CH3
CH3
холин
20. ГЛИКОЛИПИДЫ
сфингозинH3C (CH2)12 C
H
OH
C
CH CH NH C R
H
жирная кислота
CH2OH
O
ОН
галактоза
O
OН
OH
O CH2
ГЛИКОЛИПИДЫ
21. Кардиолипин
22. СТЕРОИДЫ – производные циклопентанпергидрофенантрена (стерана, гонана). СТЕРОЛЫ (СТЕРИНЫ) – стероидные спирты
21 CH3
11
HC
CH3
CH2 CH2 CH2 CH
CH3
CH3
17
CH3
3
HO
5
• СТЕРОИДНЫЙ СПИРТ ХОЛЕСТЕРИН
(холестерол) – ОСНОВНОЙ СТЕРОИД В
ОРГАНИЗМЕ ЖИВОТНЫХ
23. Нумерация атомов в молекулах стероидов
Растительные стерины(фитостерины):
Эргостерин,
Ситостерин,
Стигмастерин и др.
24. ЭФИРЫ - ЗАПАСНАЯ ФОРМА ХОЛЕСТЕРИНА
CH3HC
CH3
17
CH3
3
O
C=O
CH2
CH2
CH2
CH
CH3
CH3
25. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
1. Структурное: компоненты мембран.2. Регуляторная: стероидные гормоны,
витамин D.
3. Пластическое: желчные кислоты.
26. СТРОЕНИЕ МЕМБРАНЫ (жидкостно-мозаичная модель Зенгера и Николсона, 1972)
27.
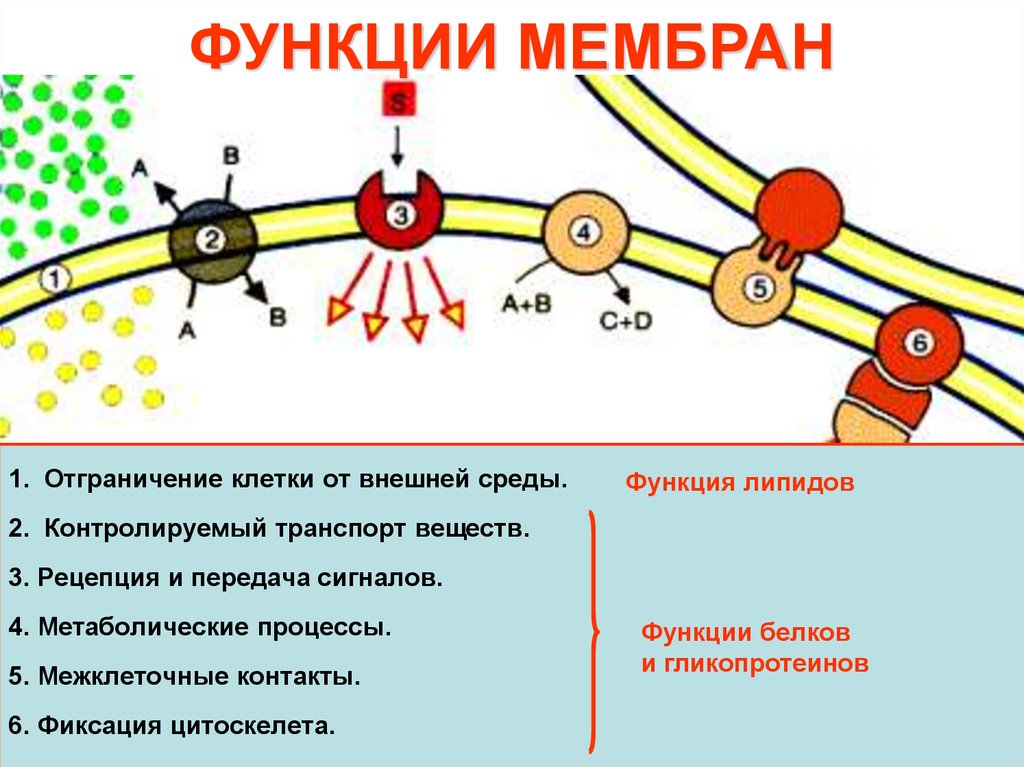
ФУНКЦИИ МЕМБРАН1. Отграничение клетки от внешней среды.
Функция липидов
2. Контролируемый транспорт веществ.
3. Рецепция и передача сигналов.
4. Метаболические процессы.
5. Межклеточные контакты.
6. Фиксация цитоскелета.
Функции белков
и гликопротеинов
28.
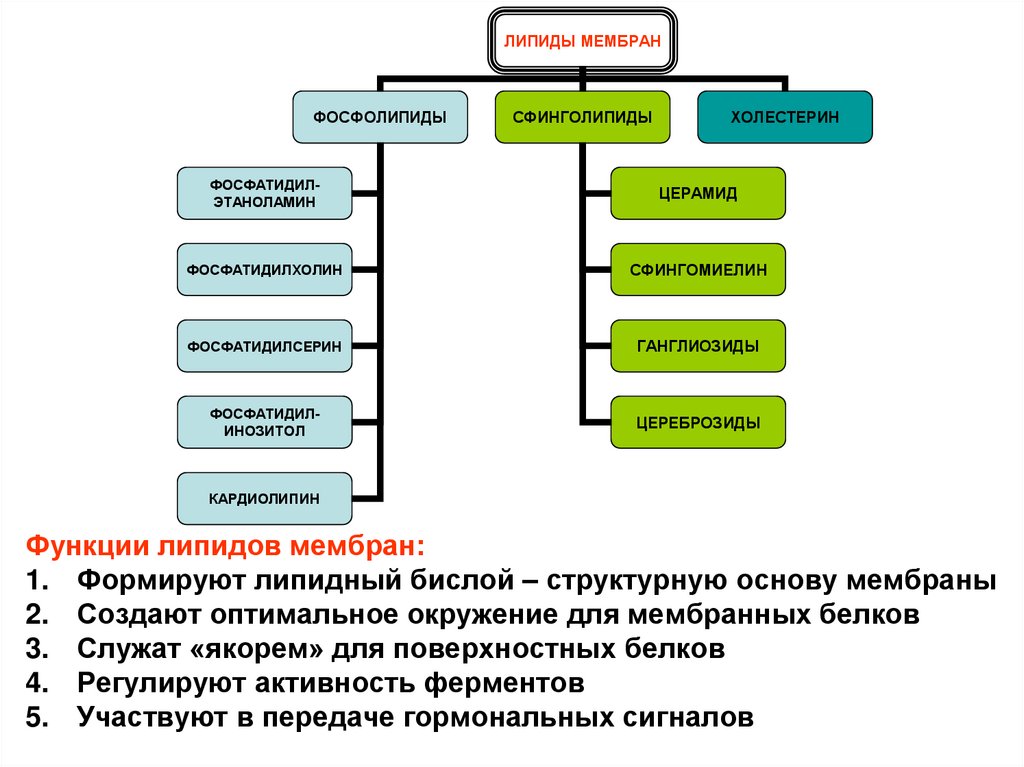
ЛИПИДЫ МЕМБРАНФОСФОЛИПИДЫ
СФИНГОЛИПИДЫ
ХОЛЕСТЕРИН
ФОСФАТИДИЛЭТАНОЛАМИН
ЦЕРАМИД
ФОСФАТИДИЛХОЛИН
СФИНГОМИЕЛИН
ФОСФАТИДИЛСЕРИН
ГАНГЛИОЗИДЫ
ФОСФАТИДИЛИНОЗИТОЛ
ЦЕРЕБРОЗИДЫ
КАРДИОЛИПИН
Функции липидов мембран:
1. Формируют липидный бислой – структурную основу мембраны
2. Создают оптимальное окружение для мембранных белков
3. Служат «якорем» для поверхностных белков
4. Регулируют активность ферментов
5. Участвуют в передаче гормональных сигналов
29. ЛИПИДЫ МЕМБРАН
Отношение ФОСФОЛИПИДЫ / ХОЛЕСТЕРИН(2-3:1) характеризует микровязкость мембран.
Максимальное содержание ХС в плазматической
мембране, занчительно меньше его в мембранах
клеточных органелл
30.
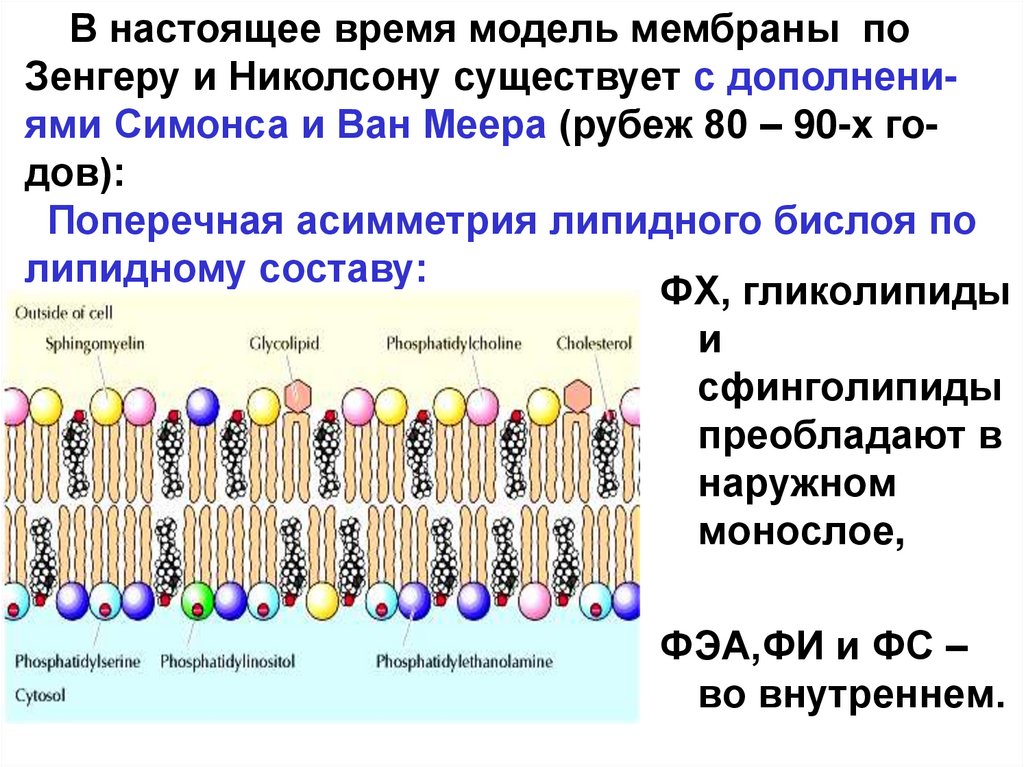
В настоящее время модель мембраны поЗенгеру и Николсону существует с дополнениями Симонса и Ван Меера (рубеж 80 – 90-х годов):
Поперечная асимметрия липидного бислоя по
липидному составу:
ФХ, гликолипиды
и
сфинголипиды
преобладают в
наружном
монослое,
ФЭА,ФИ и ФС –
во внутреннем.
31.
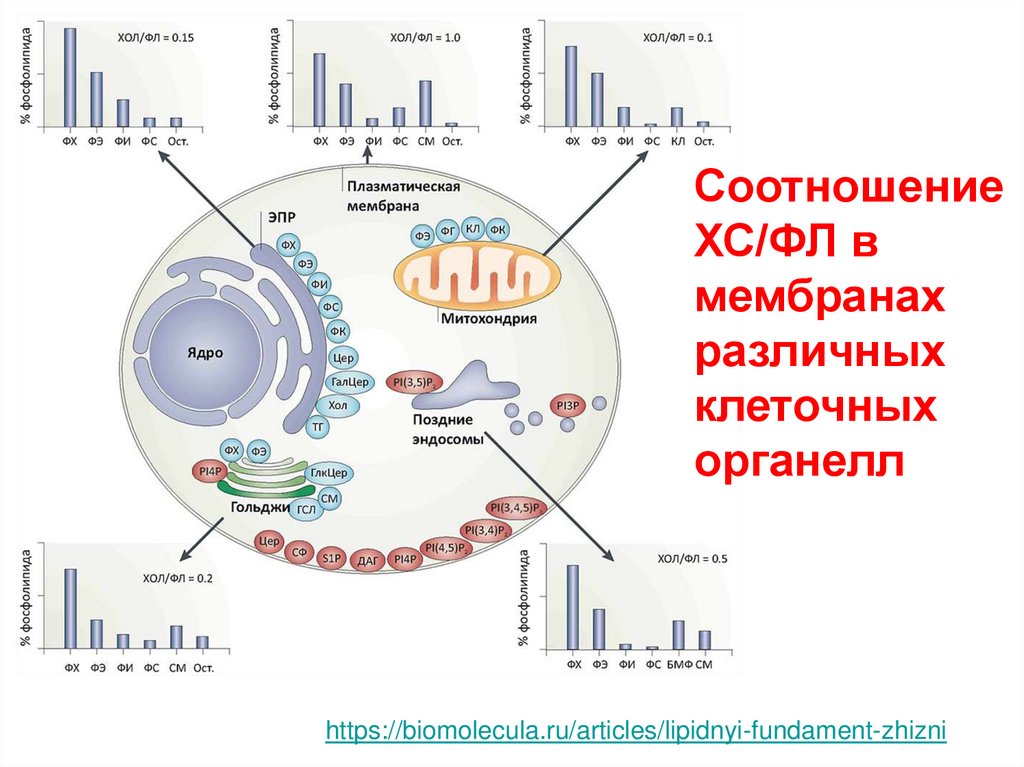
СоотношениеХС/ФЛ в
мембранах
различных
клеточных
органелл
https://biomolecula.ru/articles/lipidnyi-fundament-zhizni
32. Липиды перемещаются в пределах мембраны:
• кинки,• вращения,
• латеральная
диффузия
• флип-флоп.
33.
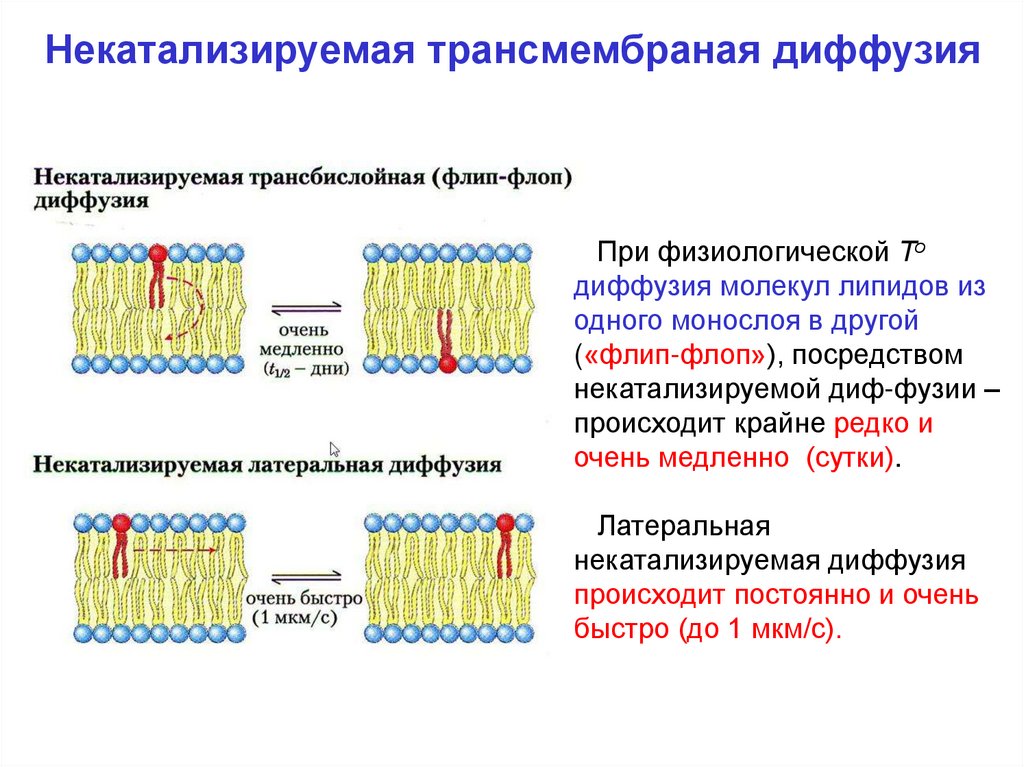
Некатализируемая трансмембраная диффузияПри физиологической То
диффузия молекул липидов из
одного монослоя в другой
(«флип-флоп»), посредством
некатализируемой диф-фузии –
происходит крайне редко и
очень медленно (сутки).
Латеральная
некатализируемая диффузия
происходит постоянно и очень
быстро (до 1 мкм/с).
34.
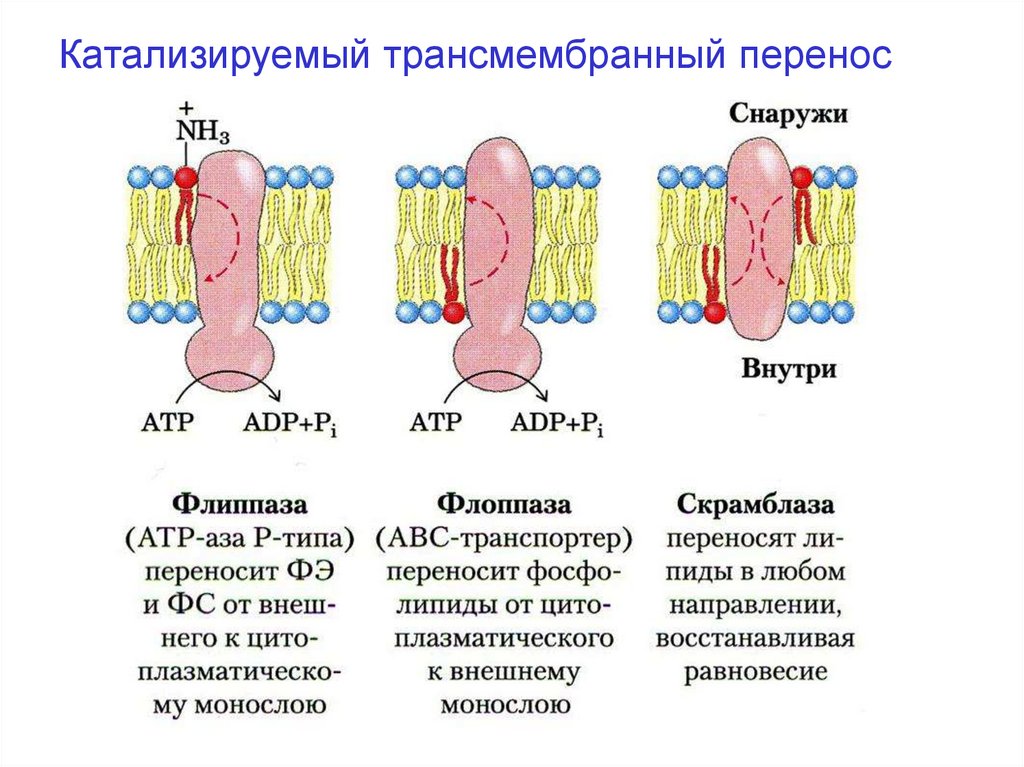
Катализируемый трансмембранный перенос35.
Поперечная асимметрия липидного бислоявозможна благодаря селективным энергозависимым переносчикам липидов. К их числу
относится семейство белков (флоппазы,
флиппазы и скрэмблазы), которые облегчают
перемещение молекул липидов поперёк бисля
мембраны (катализируемый трансмембранный
перенос):
А. Флиппазы. Катализируют перенос ФЭА и ФС
из внешнего монослоя во внутенний. Перенос 1
молекулы ФЛ требует затраты 1 моле-кулы АТФ.
По структуре флиппазы родственны транспорным
АТФазам.
36.
Б. Флоппазы. Перемещают ФЛ в обратномнаправлении – из внутреннего монослоя во
внешний.
В. Скрамблазы. Переносят через липидный
бислой любые ФЛ вдоль градиента концентрации, не требуют АТФ, но активируются в
присутствии ионов Са2+.
37. Липидные рафты (плоты, плотики) и кавеолы
• Особая фракция мембранных липидов, богатаяХС и сфинголипидами с насыщенными ЖК.
• Играют роль в процессах рецепции,
синаптической передачи, эндо- и экзоцитоза
38.
2. Рафты и сигнальные платформы.Представления о рафтах в липидной фазе цитоплазматических мембран были сформированы Симонсом, Ван
Меером и Айконеном на рубеже 80-90-х годов про-шлого
столетия.
Рафты - (10 – 200 нм) небольшие микродомены цитоплазматической мембраны, содержащие холестерол,
гликолипиды и сфингомиелин. Сохраняя свой липидный
состав, рафты «плавают» в глицерофосфолипидном
«озере» (латеральная диффузия), подобно плотам (от
англ. «raft» - плот).
Рафты четко отграничены от их глицерофосфолипидного
окружения в пределах мембранного бислоя и не
смешиваются с ним, имеют большую степень упорядоченности.
39.
В составе рафтов типично присутствие рецепторов,обладающих собственной тирозинкиназной активностью и
других белков, участвующих в передаче внешнего сигнала
внутрь клетки (10 – 15 различных мемб-ранных белков).
В зависимости от типа клеток, рафты могут занимать 20%
- 50% поверхности плазматической мембраны.
При воздействии на клетку биологических (гормон) и
физических факторов происходит слияние рафтов в более
крупные
липидные
макродомены
–
сигнальные
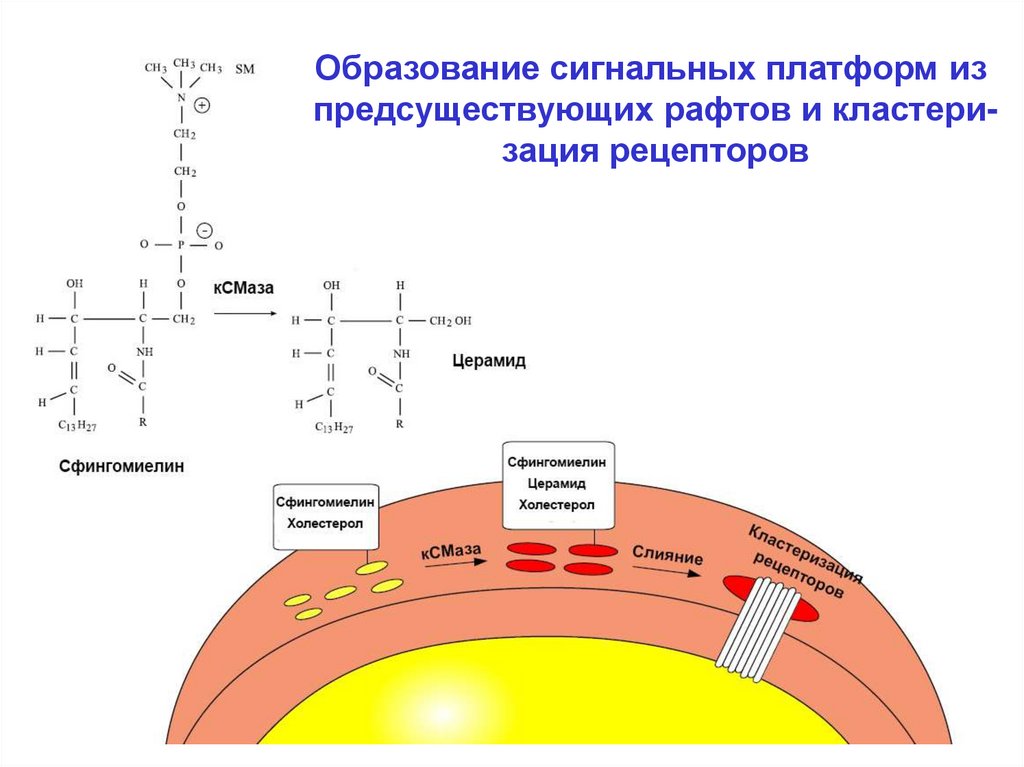
«платформы». Активация рецептора сопряжено с активацией кислой СМазы. Мембранный сфингомиелин
превращается в церамид. Резкое увеличение содержания
церамида в составе рафта заставляет их сливаться,
образуя платформу.
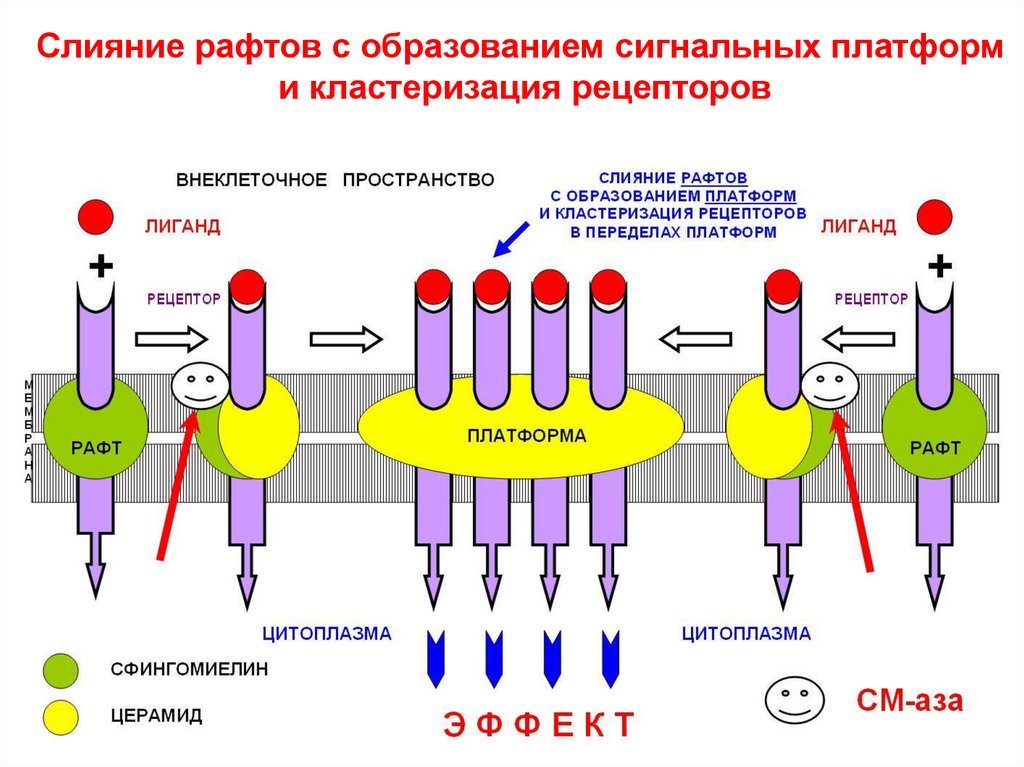
40.
В пределах образующейся сигнальной платформы втечение нескольких секунд происходит кластеризация
рецепторов, что является эффективным способом
усиления внешнего регуляторного сигнала и облегче-ния
его проведения внутрь клетки.
В состав платформы могут входить: адренорецеп-тор, Gбелок, аденилатциклаза, протеинкиназа А и
протеинфосфатаза PP2 и др.
В составе платформы содержатся молекулы, образующие высокоинтегрированную сигнальную единицу.
Платформа способна инициировать и завершить ответ
клетки на внешний сигнал.
41.
Образование сигнальных платформ изпредсуществующих рафтов и кластеризация рецепторов
42.
Слияние рафтов с образованием сигнальных платформи кластеризация рецепторов
43.
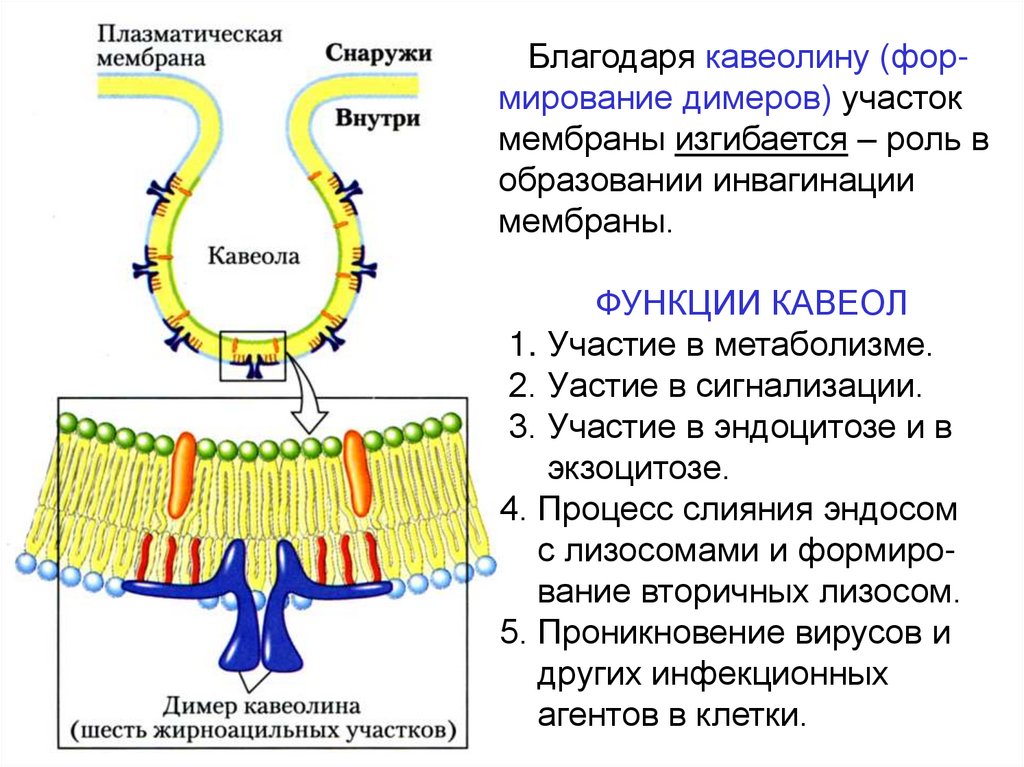
Благодаря кавеолину (формирование димеров) участокмембраны изгибается – роль в
образовании инвагинации
мембраны.
ФУНКЦИИ КАВЕОЛ
1. Участие в метаболизме.
2. Уастие в сигнализации.
3. Участие в эндоцитозе и в
экзоцитозе.
4. Процесс слияния эндосом
с лизосомами и формирование вторичных лизосом.
5. Проникновение вирусов и
других инфекционных
агентов в клетки.
44. БЕЛКИ МЕМБРАН
• СоотношениеБЕЛКИ/ ЛИПИДЫ = 1:1
• Локализация:
интегральные,
погруженные,
периферические
• Функции:
ферменты,
рецепторы,
транспортеры
45. Интегральные белки мембран
Внутримембранный домен трансмембранного белка имеет строениеα-спирали и содержит много гидрофобных остатков
Правило «положительные остатки - внутри» - внутриклеточный домен
несет много Arg, Lys, His
Внеклеточный домен гликозилирован
46. Интегральные белки мембран
• Мотив «β-бочонок» в супервторичной стуктуретипичен для белков, образующих мембранные
поры (каналы)
• Отсутствие воды в липидном слое облегчает
образование межцепочечных водородных
связей и способствует формированию βскладчатых листов
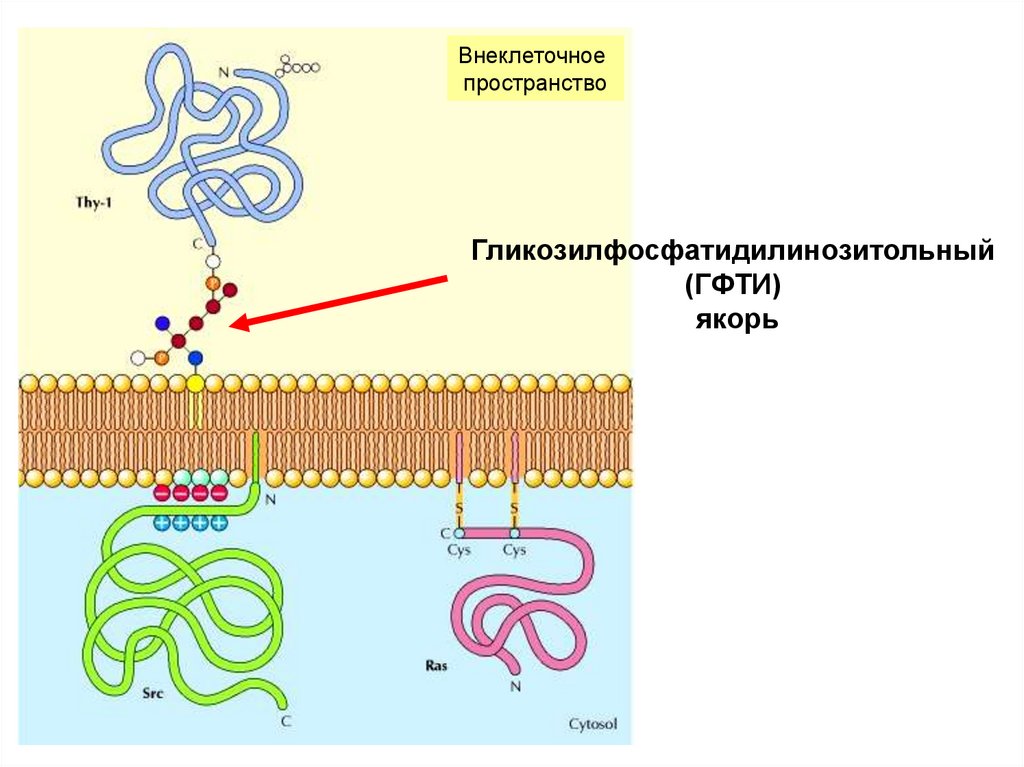
47. Ковалентное связывание с липидами – заякоривание периферических белков в мембране
липидный компонент присоединяется к белку N – амидной(миристоилированные, пальмитоилированные)
или S-тиоэфирной связями (пренилированные белки)
48.
Внеклеточноепространство
Гликозилфосфатидилинозитольный
(ГФТИ)
якорь
49.
БЛАГОДАРЮ ЗАВНИМАНИЕ!