






























 biology
biologySimilar presentations:


")

")





Эволюционная биология развития. Генетика развития дрозофилы. (Тема 2)
1.
ЭВОЛЮЦИОННАЯ БИОЛОГИЯ РАЗВИТИЯТема 2
Генетика развития дрозофилы
2.
Муха-дрозофила (Drosophilamelanogaster) - излюбленный
объект генетиков со времен
Томаса Ханта Моргана,
который начал ее изучать в
1909 году.
3.
Развитие дрозофилы имеет несколькоособенностей, нехарактерных для
большинства животных (кроме насекомых).
Во-первых, у нее центролецитальное яйцо с
желтком внутри и поверхностным слоем
чистой цитоплазмы, в котором после
дробления располагаются ядра (такое
дробление называется поверхностным).
Такой зародыш очень долго - до стадии
нескольких тысяч ядер - остается
синцитием, т.е. единой многоядерной
клеткой.
* pole cells - полярные клетки, т.е. первичные половые
клетки
4.
Пока зародыш остается синцитием, сигнальные молекулы могут простодиффундировать по общей цитоплазме, без необходимых большинству других
животных промежуточных звеньев в виде передачи от клетки к клетке и
срабатывания мембранных рецепторов со вторичными посредниками.
5.
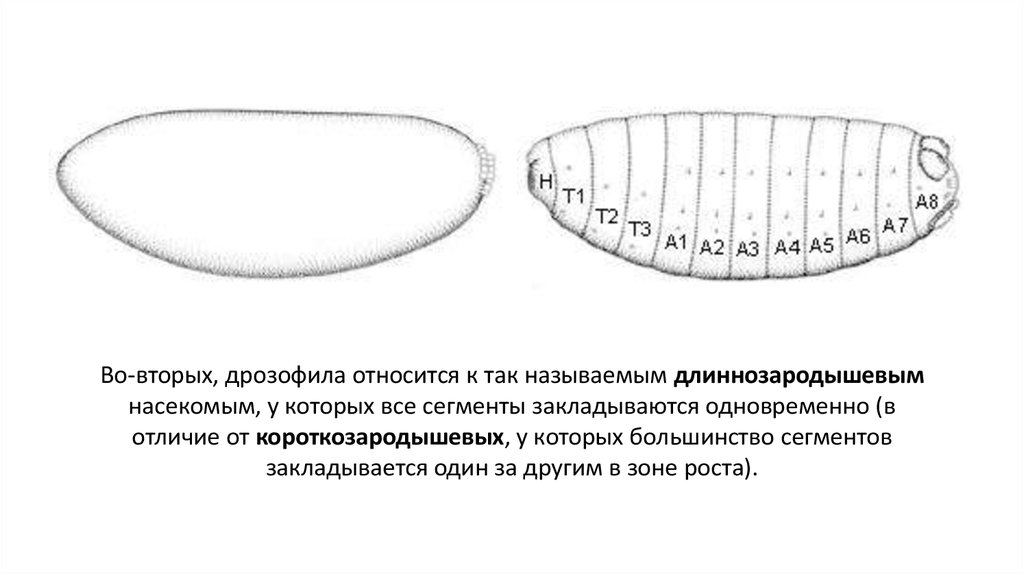
Во-вторых, дрозофила относится к так называемым длиннозародышевымнасекомым, у которых все сегменты закладываются одновременно (в
отличие от короткозародышевых, у которых большинство сегментов
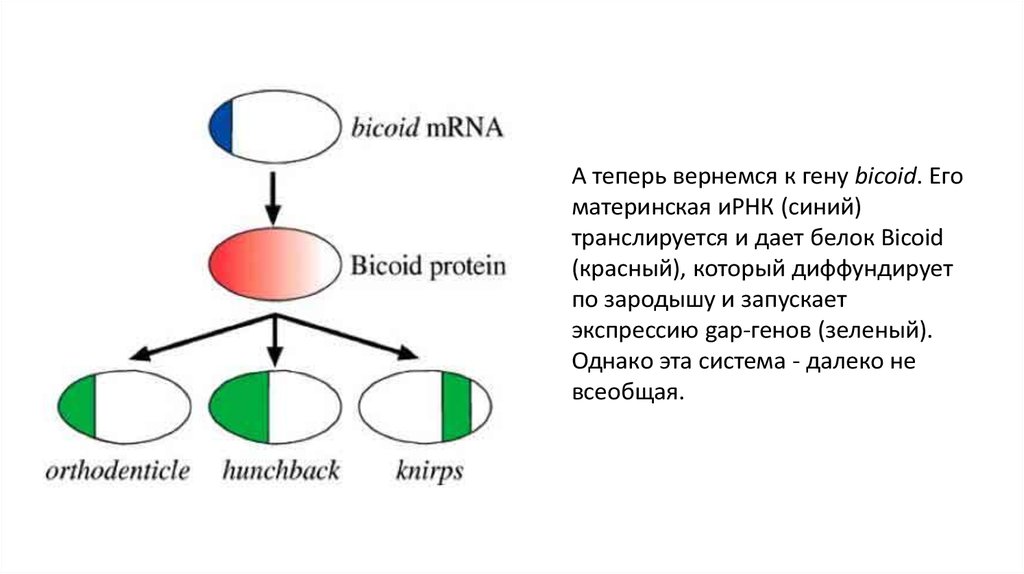
закладывается один за другим в зоне роста).
6.
Центролецитальное яйцо, поверхностное дробление, длиннозародышевость признаки, несвойственные большинству животных. Это чисто эмбриональныеспециализации, сложившиеся у продвинутых насекомых (обратим внимание на
положение двукрылых на древе). Они затрудняют сравнение дрозофилы с другими
Metazoa, но парадоксальным образом упрощают само ее изучение.
7.
Способ созревания яйцеклетки, который использует дрозофила, называетсянутриментарным. В развитии участвуют несколько трофоцитов (клетки-кормилки,
nurse cells), которые синтезируют вещества, нужные зародышу, и переправляют их в
яйцеклетку через цитоплазматические мостики.
8.
У дрозофилы известна мутацияпод названием bicoid (букв.
«двухвостость»). Мутантная
личинка лишена головы, но имеет
два задних конца тела. При этом у
самой личинки ген bicoid может
быть совершенно нормальным.
Фенотипический эффект
определяется состоянием гена
bicoid не у данной личинки, а у
мухи - ее матери. Это ген с
материнским эффектом.
9.
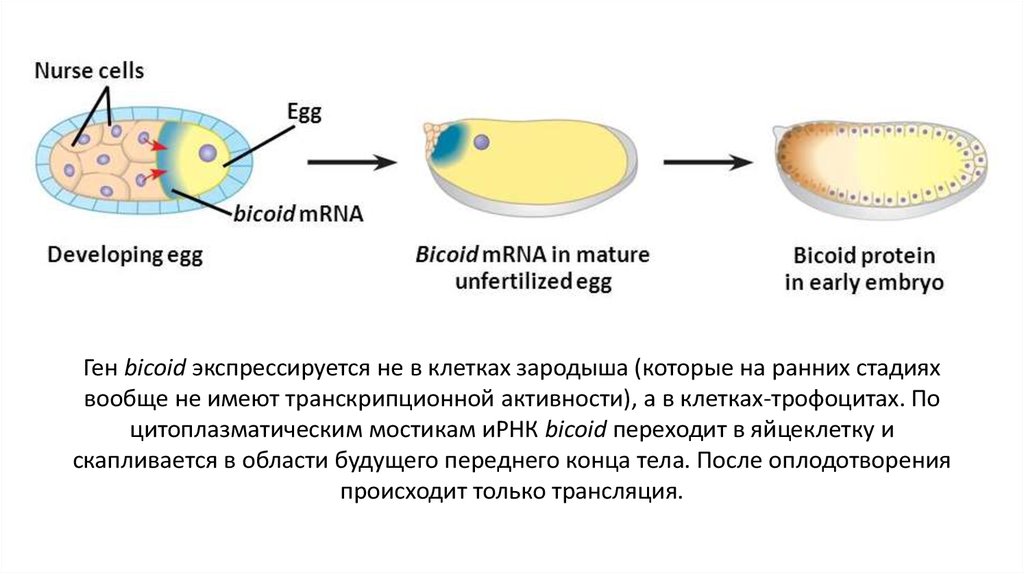
Ген bicoid экспрессируется не в клетках зародыша (которые на ранних стадияхвообще не имеют транскрипционной активности), а в клетках-трофоцитах. По
цитоплазматическим мостикам иРНК bicoid переходит в яйцеклетку и
скапливается в области будущего переднего конца тела. После оплодотворения
происходит только трансляция.
10.
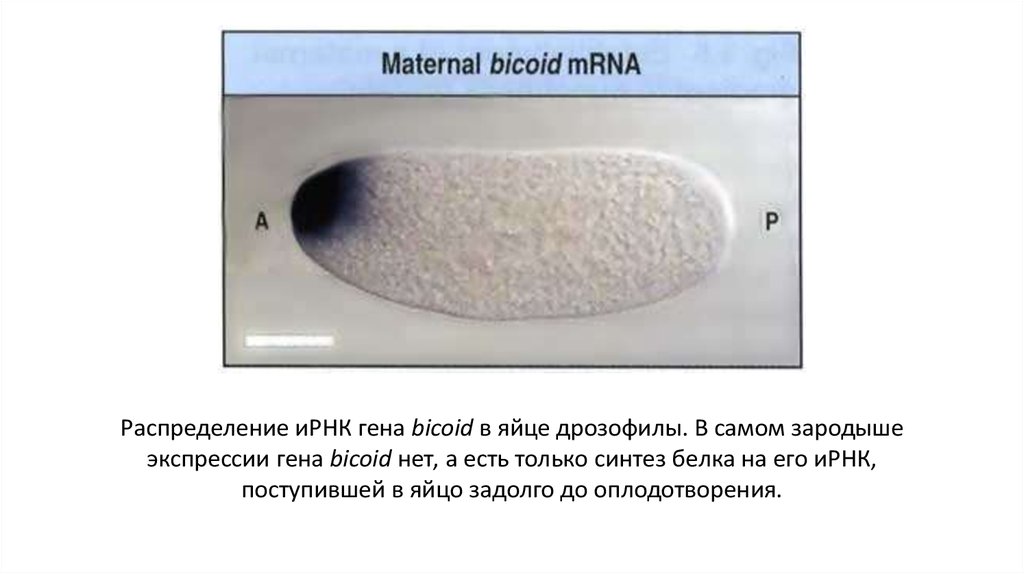
Распределение иРНК гена bicoid в яйце дрозофилы. В самом зародышеэкспрессии гена bicoid нет, а есть только синтез белка на его иРНК,
поступившей в яйцо задолго до оплодотворения.
11.
После трансляции градиент иРНК bicoidтрансформируется в градиент белка
bicoid, который является типичным
морфогеном. Градиент белка гораздо
менее крутой - это означает, что он
диффундирует эффективнее (или его
диффузия меньше тормозится).
12.
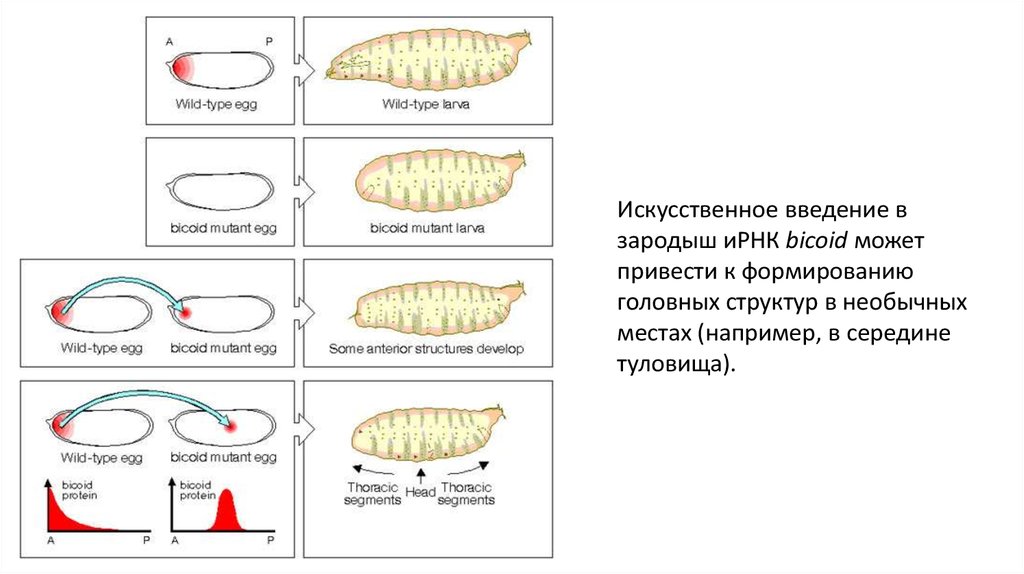
Искусственное введение взародыш иРНК bicoid может
привести к формированию
головных структур в необычных
местах (например, в середине
туловища).
13.
Увеличение числа копийгена bicoid в материнском
геноме (это можно сделать
через искусственные
дупликации) приводит к
тому, что все большая часть
зародыша становится
головой. Это аналог модели
французского флага, только
с двумя “цветами”.
14.
Следующую ступень генетической регуляции после генов материнского эффектаобразуют gap-гены (gap genes). Их особенности:
Gap-гены экспрессируются уже в клетках самого зародыша, как только их ядра
становятся синтетически активны.
Область экспрессии каждого такого гена - полоса, соответствующая нескольким
будущим сегментам.
Белки - продукты генов материнского эффекта являются факторами
транскрипции, запускающими экспрессию gap-генов.
Если какой-то из gap-генов не работает, у зародыша будут отсутствовать те
сегменты, которые должны развиться в области его экспрессии. Вот почему эти
гены так называются (gap - разрыв, провал).
15.
Области экспрессии нескольких gapгенов. Дальше мы рассмотрим вкачестве примера экспрессию гена
hunchback, продукт которого
маркирует головные сегменты.
Название гена (“горбун”) связано с
тем, что он влияет также и на
формирование груди.
* Строго говоря, hunchback - еще и ген
материнского эффекта, но это пока неважно.
16.
Экспрессия гена hunchback запускается продуктом гена bicoid: bicoid → hunchbackЭксперименты с дупликациями гена bicoid показывают, что чем больше доза bicoid,
тем сильнее экспрессия hunchback.
17.
Случай посложнее: ген Krüppel . Это типичный gap-ген. Он экспрессируетсяполосой посредине тела, его мутация приводит к отсутствию у личинки
грудных и передних брюшных сегментов.
18.
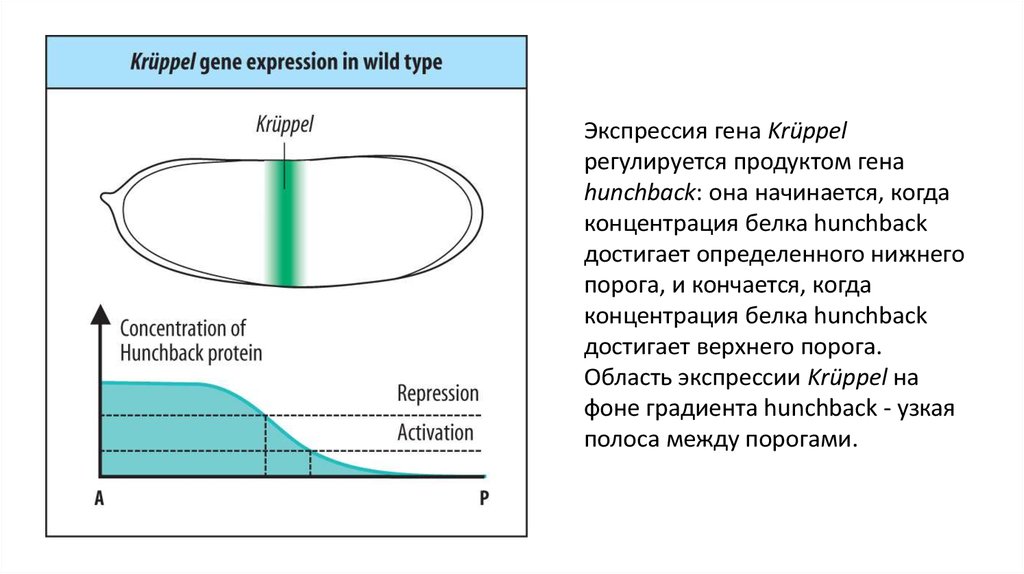
Экспрессия гена Krüppelрегулируется продуктом гена
hunchback: она начинается, когда
концентрация белка hunchback
достигает определенного нижнего
порога, и кончается, когда
концентрация белка hunchback
достигает верхнего порога.
Область экспрессии Krüppel на
фоне градиента hunchback - узкая
полоса между порогами.
19.
Следующий уровень регуляции - гены парного правила (pair rule). Это группа генов,каждый из которых экспрессируется или во всех четных, или во всех нечетных
будущих сегментах. Их экспрессия — периодическая, в отличие от апериодических
gap-генов, которые служат для них факторами транскрипции.
* На самом деле здесь не сегменты, а парасегменты, но это пока неважно.
20.
Экспрессия двух генов парного правила: even-skipped (нечетные сегменты, синий) иfushi tarazu (четные сегменты, коричневый). В каждой клетке экспрессируется или
только eve, или только ftz.
21.
Генная сеть с участием пяти генов парногоправила, включая еще не названные oddskipped (odd), runt (run) и hairy (h). Мы
видим, что гены парного правила могут
действовать на экспрессию друг друга иногда их даже делят на первичные (run, h,
eve) и вторичные (ftz, odd). Кроме того,
схема показывает, что продукты генов eve и
ftz имеют противоположные функции:
первый - репрессор, второй - активатор.
Внимание: запоминать названия отдельных
генов не надо! Нам важны только принципы
работы подобных сетей.
22.
Следующий уровень - гены сегментарной полярности (segment polarity) периодические гены, которые экспрессируются или в передней, или в задней частикаждого сегмента. Факторами транскрипции для них служат продукты генов парного
правила. В развитии дрозофилы происходит пересегментация, поэтому, например,
ген engrailed экспрессируется в передних частях парасегментов, которые становятся
задними частями настоящих сегментов.
23.
Сложности, касающиеся геновсегментарной полярности
дрозофилы: (1) пересегментация, (2)
специфические изгибы зародыша
около огромного внутреннего
желтка - на некоторых стадиях он
буквально складывается пополам,
запутывая наблюдателя. На картинке
показаны области экспрессии двух
генов-антагонистов - engrailed и
wingless.
24.
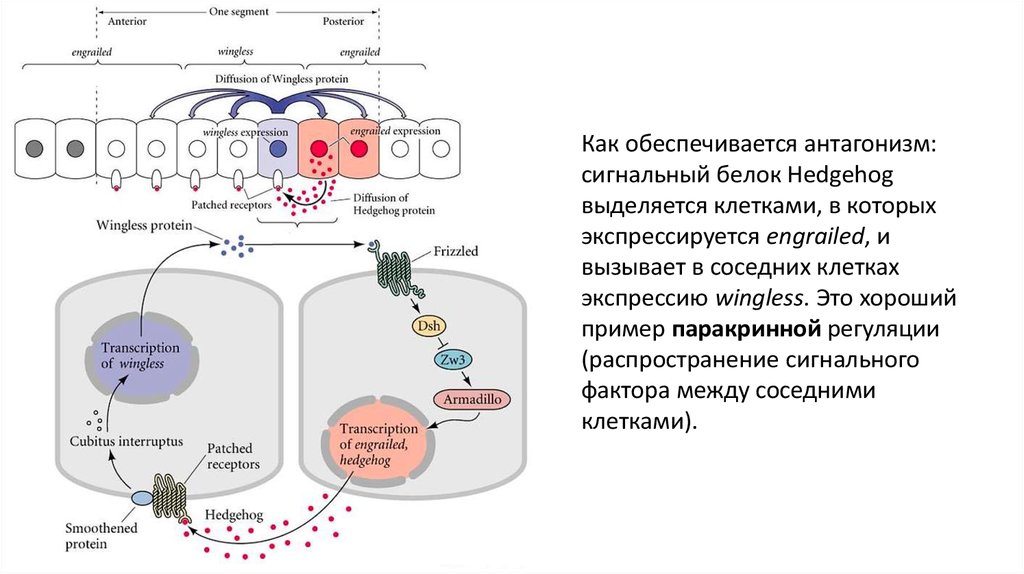
Как обеспечивается антагонизм:сигнальный белок Hedgehog
выделяется клетками, в которых
экспрессируется engrailed, и
вызывает в соседних клетках
экспрессию wingless. Это хороший
пример паракринной регуляции
(распространение сигнального
фактора между соседними
клетками).
25.
Именно на стадии, когдаэкспрессируются гены
сегментарной полярности,
зародыши всех членистоногих
больше всего похожи друг на
друга. Это - горлышко “песочных
часов”, или филотипическая
стадия. На картинке зародыш не
дрозофилы, а какого-то
короткозародышевого насекомого
(возможно, кузнечика).
26.
Чего не хватает системе сегментациипосле того, как заработали гены
сегментарной полярности? Не хватает
генов, которые будут контролировать
специализацию сегментов, отличая,
например, 1-й грудной сегмент от 2-го,
а тот и другой от брюшных. Для этого
должен существовать еще один
уровень экспрессии, представленный
апериодическими генами
сегментарной идентичности.
Факторами транскрипции для них
служат gap-гены. Большинство
показанных здесь генов (но не все)
относятся к семейству Hox.
27.
Итог: общая схема уровнейэкспрессии генов развития
дрозофилы. О некоторых
указанных здесь генах (но не обо
всех) будет сказано дальше.
28.
На этой стадии у нас должно возникнутьмного вопросов. Например, вот такой вопрос.
Откуда берется периодичность экспрессии
генов парного правила, если из генов,
экспрессирующихся до этого, периодическим
не является ни один? Получается, что
периодичность возникает de novo,
самоорганизуется.
29.
Например, вот как включаетсяген even-skipped в 3-м сегменте.
Зависимость следующая: ЕСЛИ в
некоторой точке есть продукты
bicoid и hunchback, но нет
продуктов giant и Krüppel, ТО в
этой точке должен
экспрессироваться eve. Причем
эта функция работает только в 3м сегменте, в других eve
включается иначе. “Если бы это
была компьютерная программа,
она выглядела бы так, будто ее
писал какой-то идиот”. Но из
этого беспорядка рождается
регулярность.
30.
Модель включения гена eve в разных сегментах под влиянием gap-генов. Тупаястрелочка от gap-гена означает, что данная зона экспрессии eve активна там, где
продукта этого gap-гена нет.
31.
Теперь у нас есть общая модель, в рамках которой можнососредоточиться на том или ином частном сюжете. Рассмотрим один
такой сюжет, в котором фигурирует еще не упоминавшийся ген nanos.
Nanos — ген материнского эффекта, который:
Имеет максимум концентрации продукта на заднем конце зародыша.
Необходим для образования брюшка.
Обладает свойством блокировать трансляцию материнской порции
иРНК гена hunchback, которая равномерно распределена по яйцу.
* Когда-то эта функция nanos считалась основной, на самом деле она не единственная, но
другие функции этого гена нам сейчас неважны, потому что связаны не с сегментацией, а с
созреванием половых клеток.
32.
Nanos - типичный генматеринского эффекта, только
его иРНК транспортируется по
микротрубочкам не на
передний конец будущего тела
(как иРНК bicoid), а на задний.
Заодно здесь показана
материнская порция иРНК
hunchback, распределенная по
зародышу равномерно.
33.
Продукт гена nanos ингибирует трансляцию гена hunchback там, где продуктпоследнего не нужен, то есть в задней части тела. При этом hunchback все равно
экспрессируется в ядрах зародыша, как обычный gap-ген. Зачем нужна его
дополнительная материнская доза, которую приходится блокировать таким
сложным способом?
34.
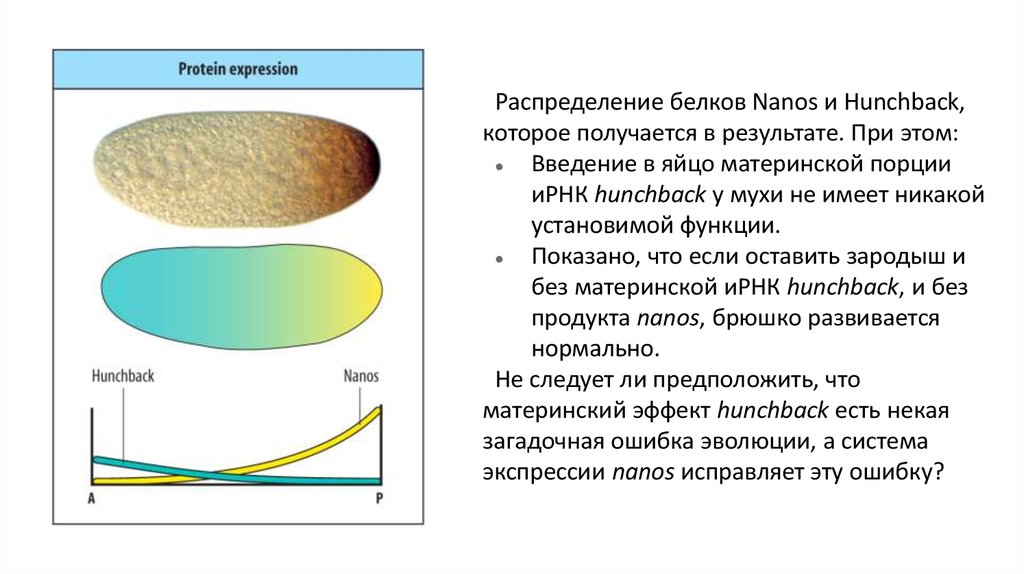
Распределение белков Nanos и Hunchback,которое получается в результате. При этом:
Введение в яйцо материнской порции
иРНК hunchback у мухи не имеет никакой
установимой функции.
Показано, что если оставить зародыш и
без материнской иРНК hunchback, и без
продукта nanos, брюшко развивается
нормально.
Не следует ли предположить, что
материнский эффект hunchback есть некая
загадочная ошибка эволюции, а система
экспрессии nanos исправляет эту ошибку?
35.
А теперь вернемся к гену bicoid. Егоматеринская иРНК (синий)
транслируется и дает белок Bicoid
(красный), который диффундирует
по зародышу и запускает
экспрессию gap-генов (зеленый).
Однако эта система - далеко не
всеобщая.
36.
Ген bicoid есть не у всех животных и даже не у всех насекомых. Он есть только укруглошовных двукрылых (Cyclorrapha), то есть у мух в строгом смысле. Например, у
комара и даже у слепня его нет.
37.
Как же была устроена разметка тела, когда гена bicoid еще не было?У двукрылых, не относящихся к Cyclorrapha, его функцию выполняют два gap-гена:
hunchback и orthodenticle. У дрозофилы мы видим остаток старой функции
hunchback, когда он еще был геном материнского эффекта:
Mother → hunchback
Потом к нему добавился bicoid:
Mother → bicoid + hunchback
А потом геном материнского эффекта остался bicoid, а hunchback переместился на
следующий уровень экспрессии:
Mother → bicoid → hunchback
Вот она, та эволюционная ошибка, для исправления которой понадобилась
экспрессия гена nanos. Материнская порция продукта hunchback – не что иное, как
рудимент старой функции этого гена.
38.
У всех двукрылых есть ген Hox3 — одиниз генов сегментарной идентичности,
имеющий в данном случае две
дополнительные функции:
Экспрессия в качестве гена
материнского эффекта,
маркирующего передний конец
тела.
Экспрессия во внезародышевых
тканях (амнион, сероза).
Дальше происходит разделение
функций (как в морфологии). Ген Hox3
дуплицируется и дает два гена: ген
zerknüllt, который экспрессируется во
внезародышевых тканях, и уже
известный нам ген bicoid.
39.
В теории эволюции есть понятие «ценогенез» (от греч. kainós - новый) приспособление организма, возникающее на стадии зародыша, плода или личинкии не сохраняющееся у взрослой особи. В сюжете с эволюцией гена bicoid меняется
генетический контроль закладки сегментов на ранних стадиях онтогенеза, но сама
сегментация взрослого животного остается совершенно неизменной (у мухи и
комара она абсолютно идентична). Это пример ценогенеза на молекулярном
уровне. Изучением таких явлений и занимается эволюционная биология развития.
40.
Понятия, введенные в этой теме: трофоциты, гены с материнским эффектом, gapгены, гены парного правила, гены сегментарной полярности, гены сегментарнойидентичности, модель “песочных часов”, филотипическая стадия, ценогенез.