

























 biology
biologySimilar presentations:

")

")
")





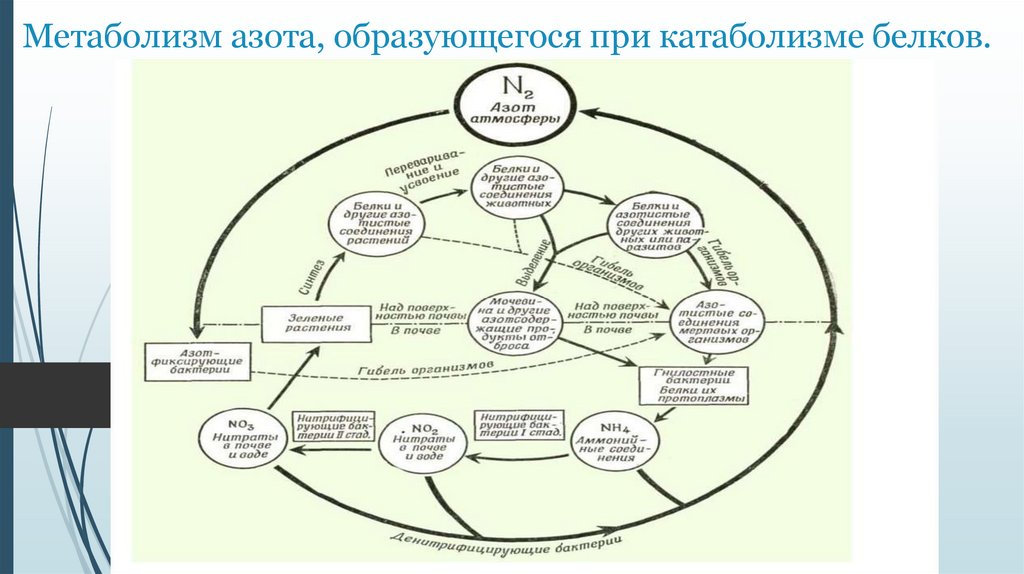
Метаболизм азота, образующегося при катаболизме белков. Круговорот азота в природе (лекция № 7)
1.
Метаболизм азота, образующегося при катаболизме белков.2.
Лекция №7.Вопросы лекции:
1. Метаболизм азота, образующегося при катаболизме белков.
2. Орнитиновый цикл.
3. Патология азотистого обмена.
4. Пути выведения аминного азота из живых организмов.
3.
4.
Роль азота в природеАзот — одно из самых распространенных веществ в биосфере, узкой оболочке Земли, где
поддерживается жизнь.
Основная часть атмосферного азота находится в свободной форме, при которой два
атома азота соединены вместе, образуя молекулу азота — N2.
Из-за того, что связи между двумя атомами очень прочные, живые организмы не
способны напрямую использовать молекулярный азот — его сначала необходимо
перевести в «связанное» состояние.
В процессе связывания молекулы азота расщепляются, давая возможность отдельным
атомам азота участвовать в химических реакциях с другими атомами, например с
кислородом, и таким образом мешая им вновь объединиться в молекулу азота.
Связь между атомами азота и другими атомами достаточно слабая, что позволяет
живым организмам усваивать атомы азота. Поэтому связывание азота — чрезвычайно
важная часть жизненных процессов на нашей планете.
5.
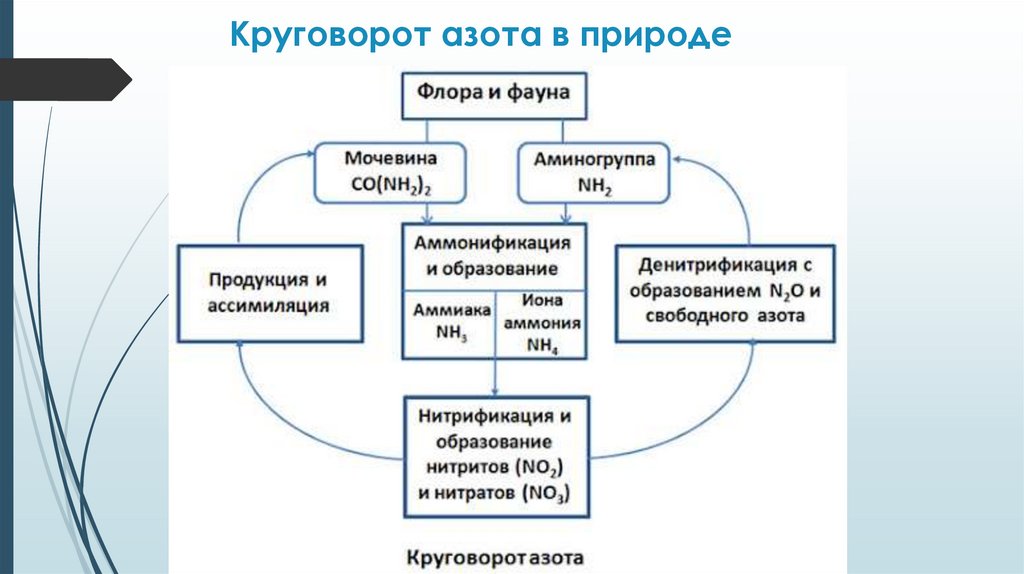
Круговорот азота в природе6.
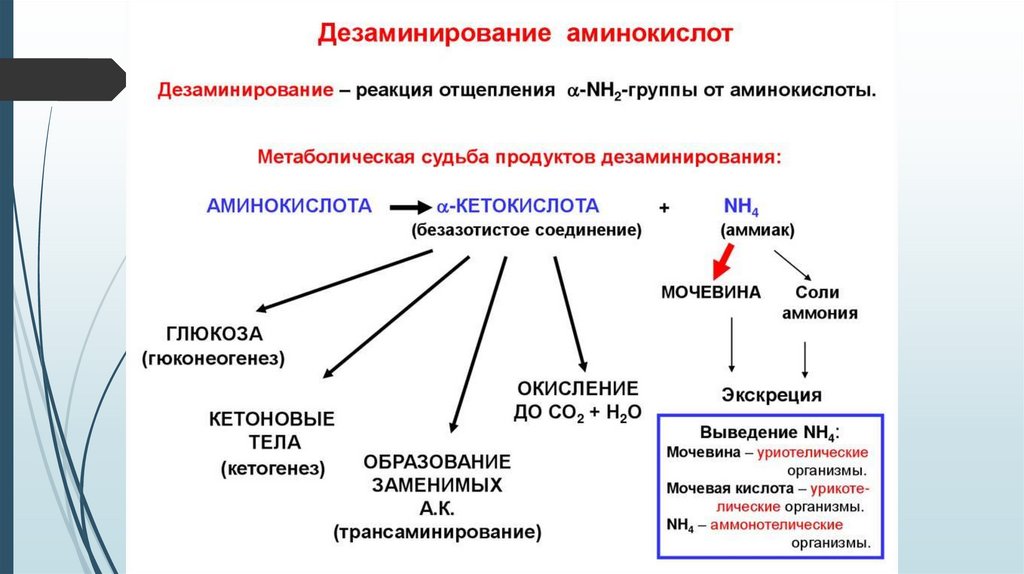
Условия для окислительное расщепление аминокислот:1. Если аминокислоты, освобождающиеся при обновлении белков, не
используются для синтеза новых белков, то они подвергаются
окислительному расщеплению.
2. Если организм с пищей получает больше аминокислот, чем ему
необходимо для синтеза белка, то избыточное количество
расщепляется. ( аминокислоты не откладываются про запас).
3. Во время голодания или при сахарном диабете, т.е. когда
углеводов нет или когда их утилизация нарушена, тогда в качестве
топлива используются белки.
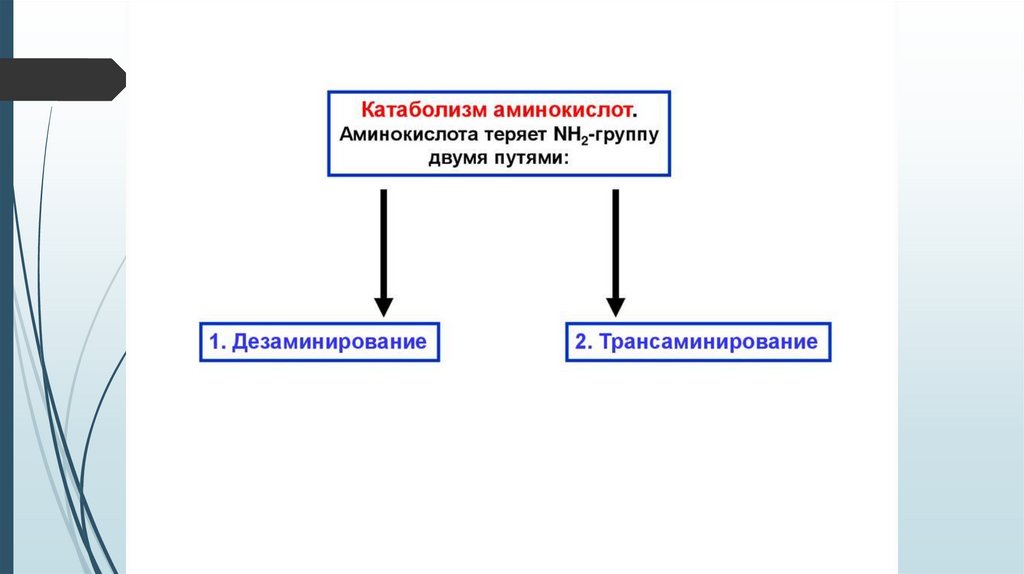
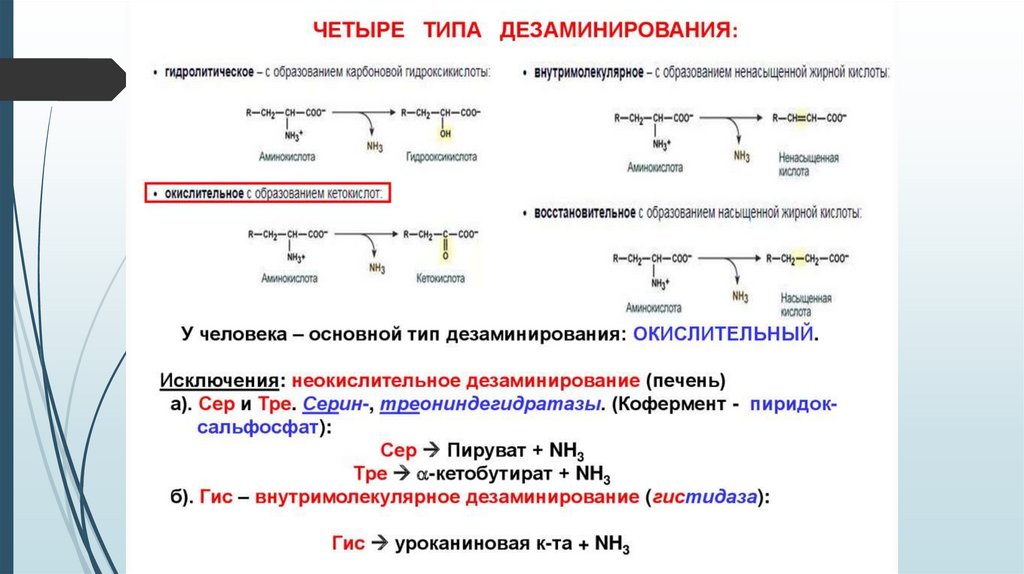
1 стадия: удаление α-аминогруппы
- переаминирование (трансаминирование);
- дезаминирование;
2 стадия: распад углеродного скелета
7.
8.
9.
10.
11.
12.
13.
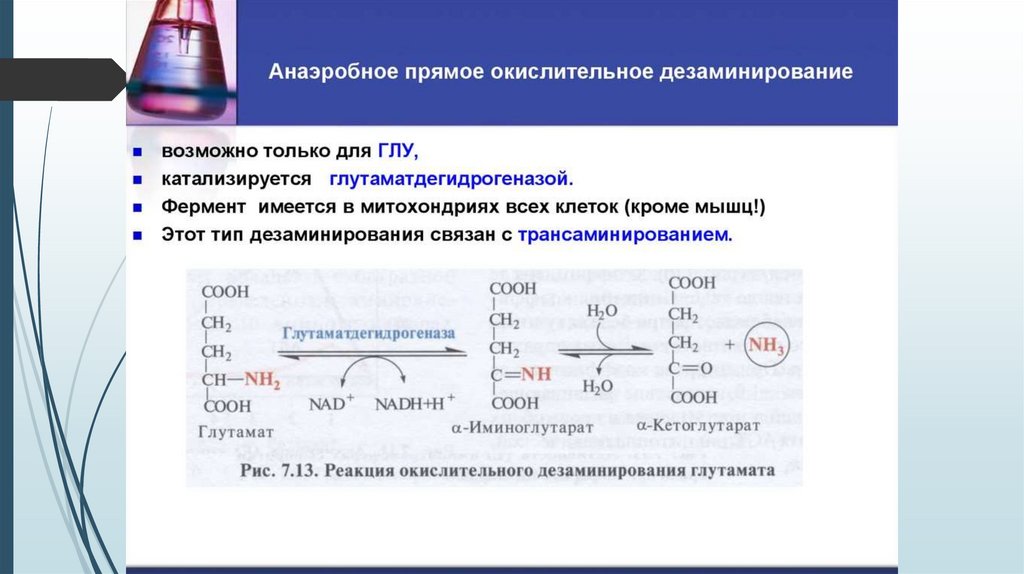
Роль глутаминовой кислотыявляется одной из транспортных форм аминного
азота в гепатоциты,
способна реагировать
обезвреживая его.
со
свободным
аммиаком,
повышает устойчивость организма к гипоксии
является нейромедиаторной аминокислотой, одной из важных
представителей класса «возбуждающих аминокислот». В медицине
применение
глутаминовой
кислоты
оказывает
незначительное
психостимулирующее, возбуждающее действие, что используют в лечении
ряда заболеваний нервной системы.
Глутаминовая
кислота
(E620)
и
её
соли
(
Е621,
Е622, Е623, Е624, Е625) используются как усилитель вкуса во многих
пищевых
продуктах
14.
15.
16.
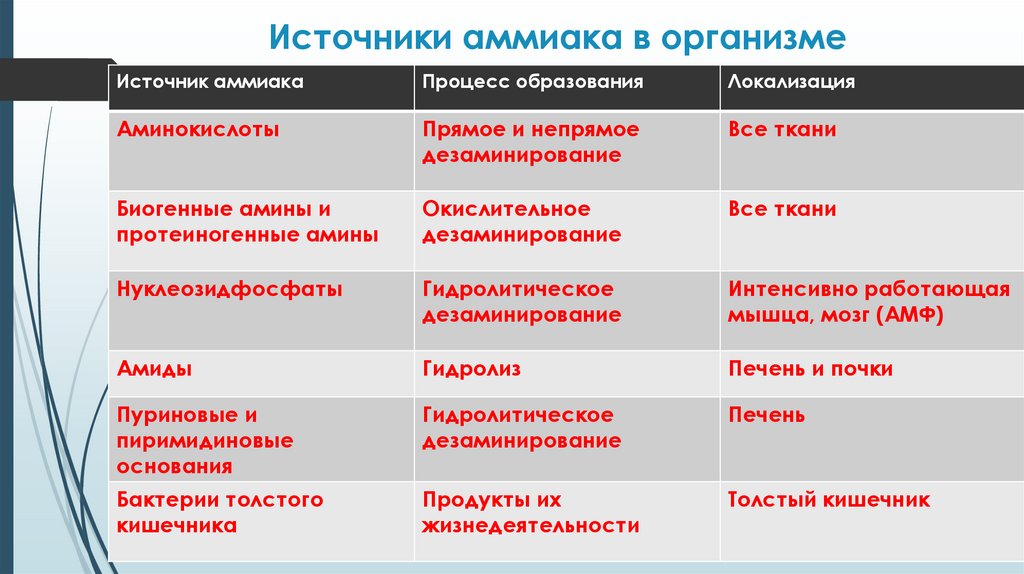
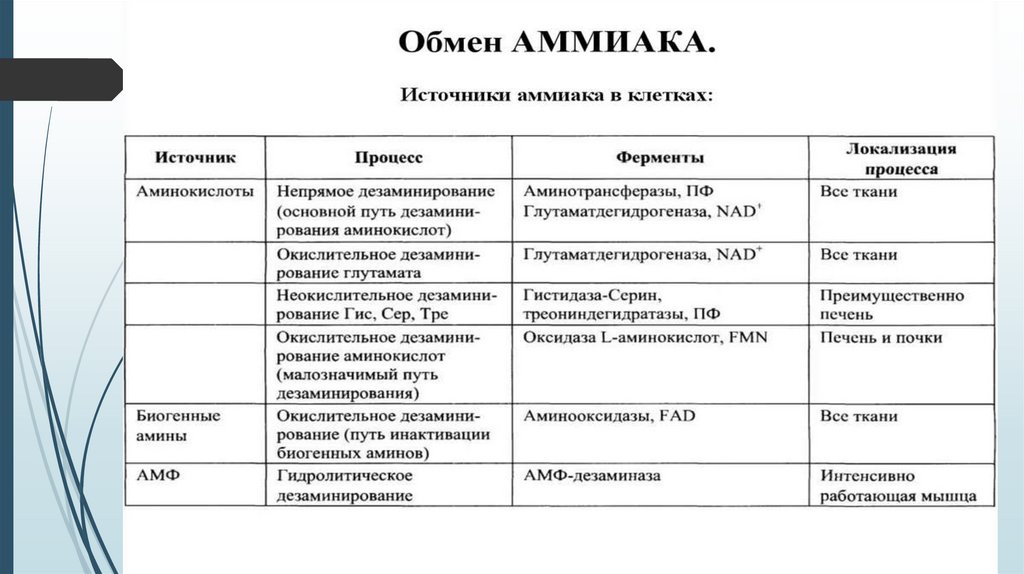
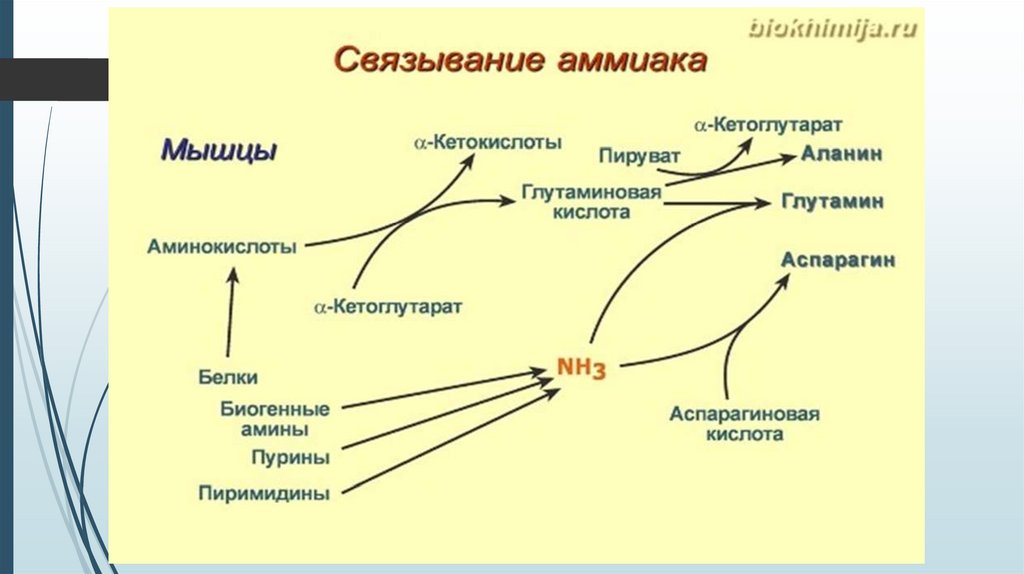
Источники аммиака в организмеИсточник аммиака
Процесс образования
Локализация
Аминокислоты
Прямое и непрямое
дезаминирование
Все ткани
Биогенные амины и
протеиногенные амины
Окислительное
дезаминирование
Все ткани
Нуклеозидфосфаты
Гидролитическое
дезаминирование
Интенсивно работающая
мышца, мозг (АМФ)
Амиды
Гидролиз
Печень и почки
Пуриновые и
пиримидиновые
основания
Гидролитическое
дезаминирование
Печень
Бактерии толстого
кишечника
Продукты их
жизнедеятельности
Толстый кишечник
17.
18.
Прифизиологических
значениях рН молекула
NН3 легко превращается в
ион
аммония
NН4+,
который
не
способен
проникать
через
биологические мембраны
и задерживается в клетке.
19.
20.
Пути утилизации аммиака1
2
3
• Восстановительное аминирование альфа кетокислот
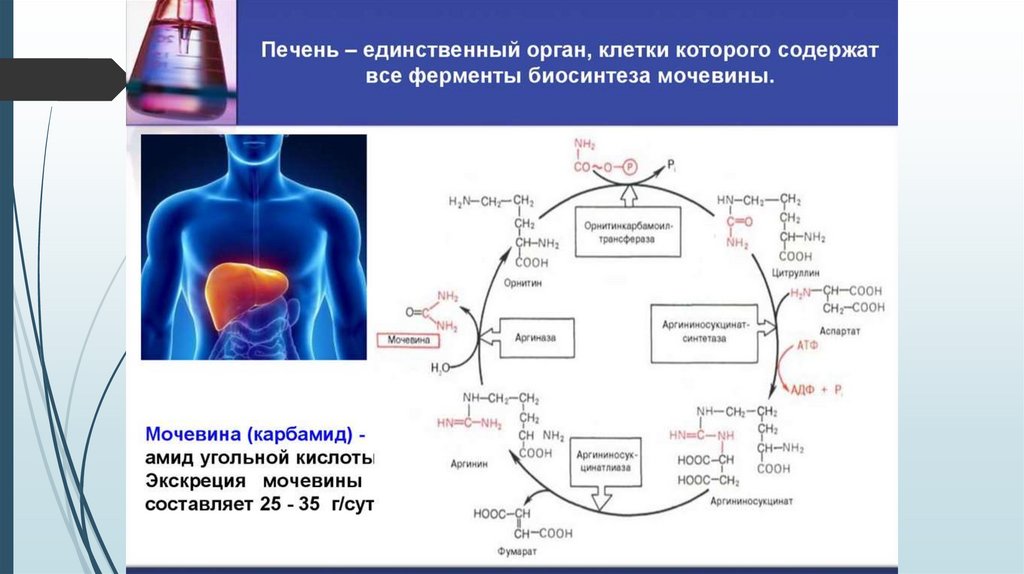
• Образование мочевины ( 25 г/сут) ( печень)
• Образование солей аммония 0,5 г/сут ( почки)
• Образование глутамина и аспарагина ( мышцы, мозг)
4
21.
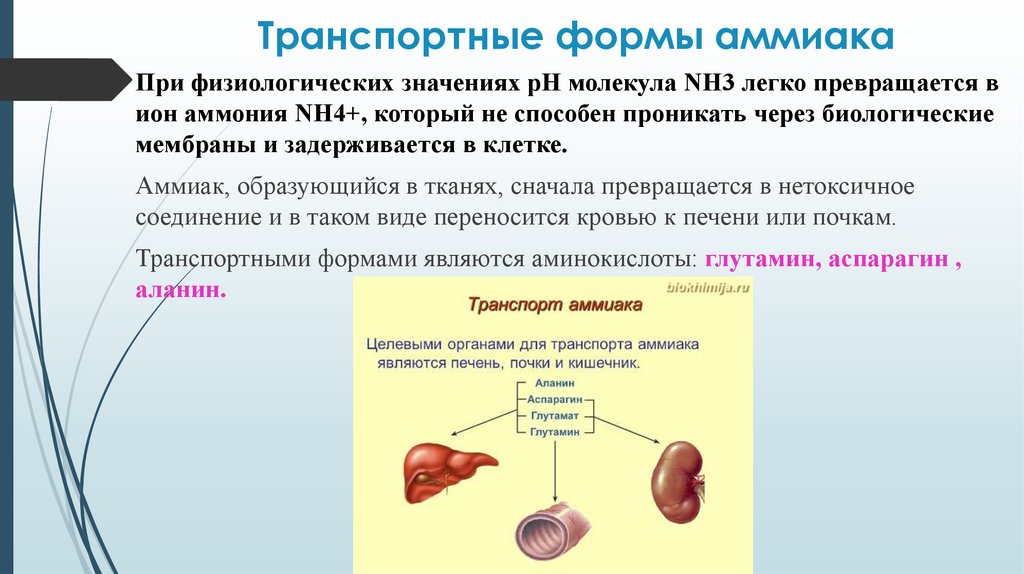
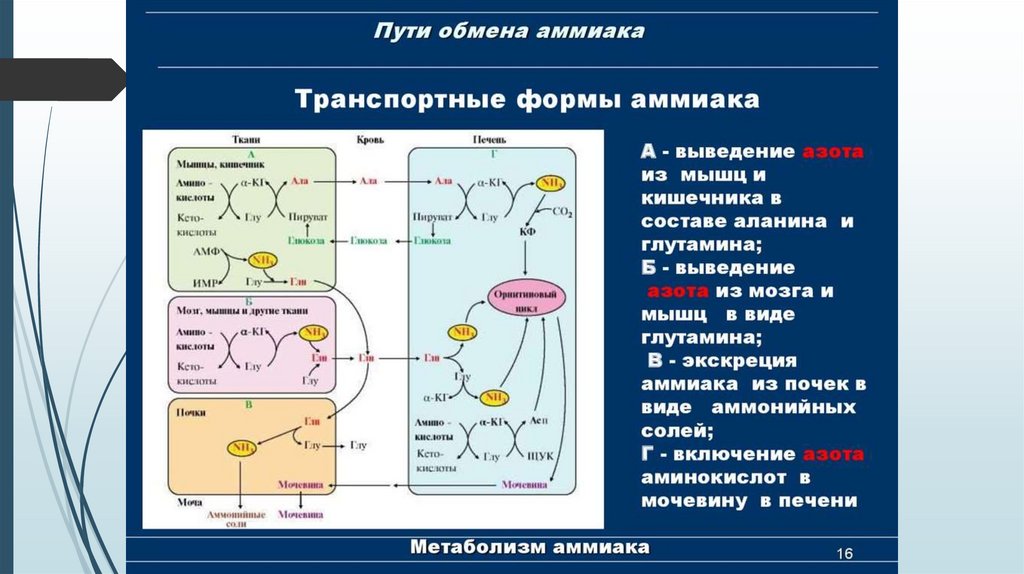
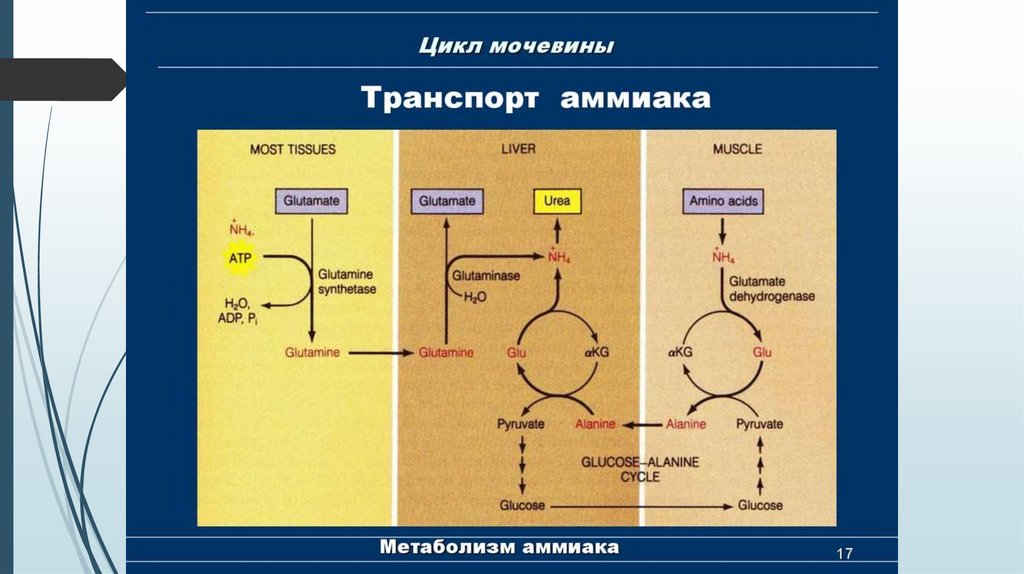
Транспортные формы аммиакаПри физиологических значениях рН молекула NН3 легко превращается в
ион аммония NН4+, который не способен проникать через биологические
мембраны и задерживается в клетке.
Аммиак, образующийся в тканях, сначала превращается в нетоксичное
соединение и в таком виде переносится кровью к печени или почкам.
Транспортными формами являются аминокислоты: глутамин, аспарагин ,
аланин.
22.
23.
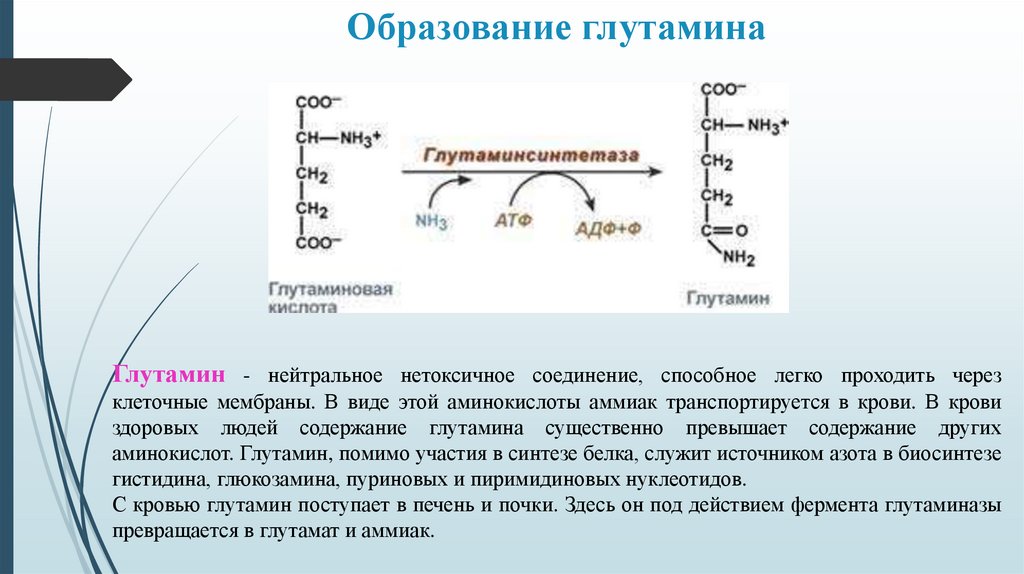
Образование глутаминаГлутамин - нейтральное нетоксичное соединение, способное легко проходить через
клеточные мембраны. В виде этой аминокислоты аммиак транспортируется в крови. В крови
здоровых людей содержание глутамина существенно превышает содержание других
аминокислот. Глутамин, помимо участия в синтезе белка, служит источником азота в биосинтезе
гистидина, глюкозамина, пуриновых и пиримидиновых нуклеотидов.
С кровью глутамин поступает в печень и почки. Здесь он под действием фермента глутаминазы
превращается в глутамат и аммиак.
24.
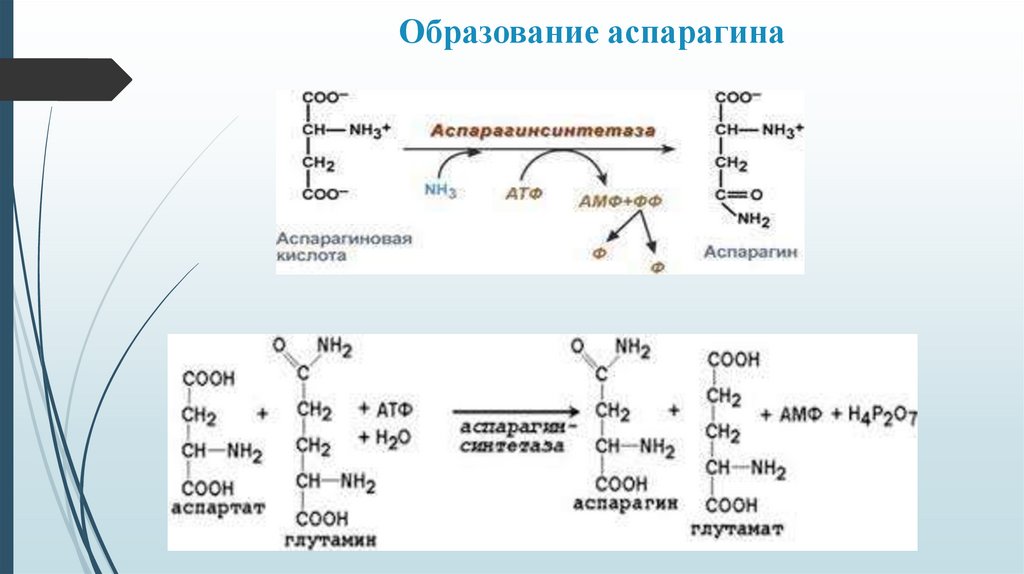
Образование аспарагина25.
Образование аланинаАланин является транспортной формой аммиака, которая образуется преимущественно в
мышцах. При интенсивной физической нагрузке источниками аммиака служат реакции
дезаминирования аминокислот и аденозинмонофосфата (АМФ).
1. Сначала аммиак превращается в аминогруппу
глутамата в реакции восстановительного
аминирования 4.
2. Образовавшийся глутамат переносит затем свою
α-аминогруппу на пируват, всегда имеющийся в
достаточном количестве, поскольку это продукт
протекающего в мышцах гликолиза.
3. Аланин (нейтральная аминокислота, не несущая
суммарного заряда при значениях рН, близких к 7)
выходит из клеток и доставляется кровью к печени.
Здесь он под действием аланинаминотрансферазы
передаёт свою аминогруппу α-кетоглутарату, в
результате чего образуется глутамат.
4. Глутамат дезаминируется с
образованием α-кетоглутарата и
аммиака,
который
в
печени
превращается в мочевину.
26.
27.
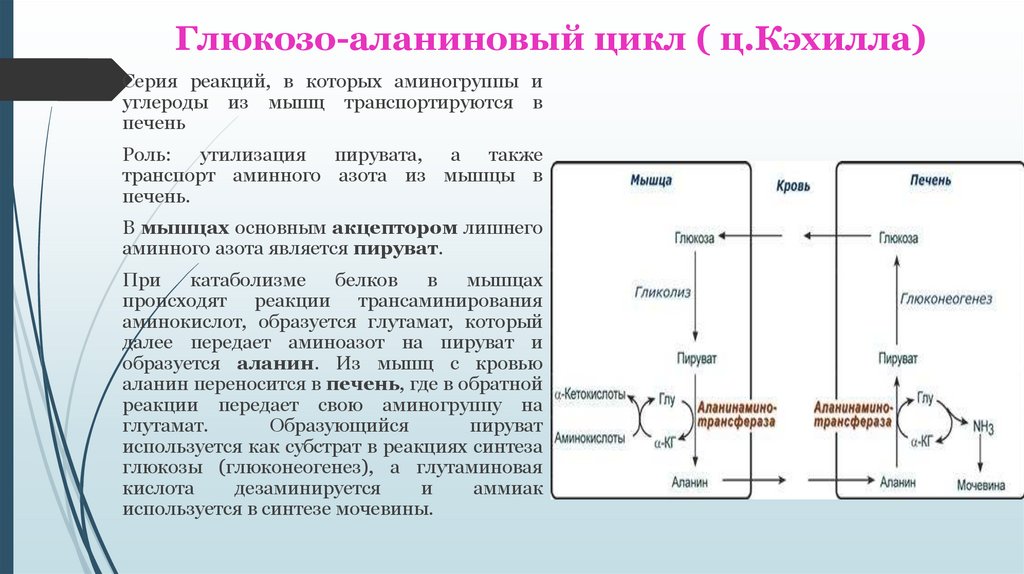
Глюкозо-аланиновый цикл ( ц.Кэхилла)Серия реакций, в которых аминогруппы и
углероды из мышц транспортируются в
печень
Роль: утилизация пирувата, а также
транспорт аминного азота из мышцы в
печень.
В мышцах основным акцептором лишнего
аминного азота является пируват.
При
катаболизме
белков
в
мышцах
происходят реакции трансаминирования
аминокислот, образуется глутамат, который
далее передает аминоазот на пируват и
образуется аланин. Из мышц с кровью
аланин переносится в печень, где в обратной
реакции передает свою аминогруппу на
глутамат.
Образующийся
пируват
используется как субстрат в реакциях синтеза
глюкозы (глюконеогенез), а глутаминовая
кислота
дезаминируется
и
аммиак
используется в синтезе мочевины.
28.
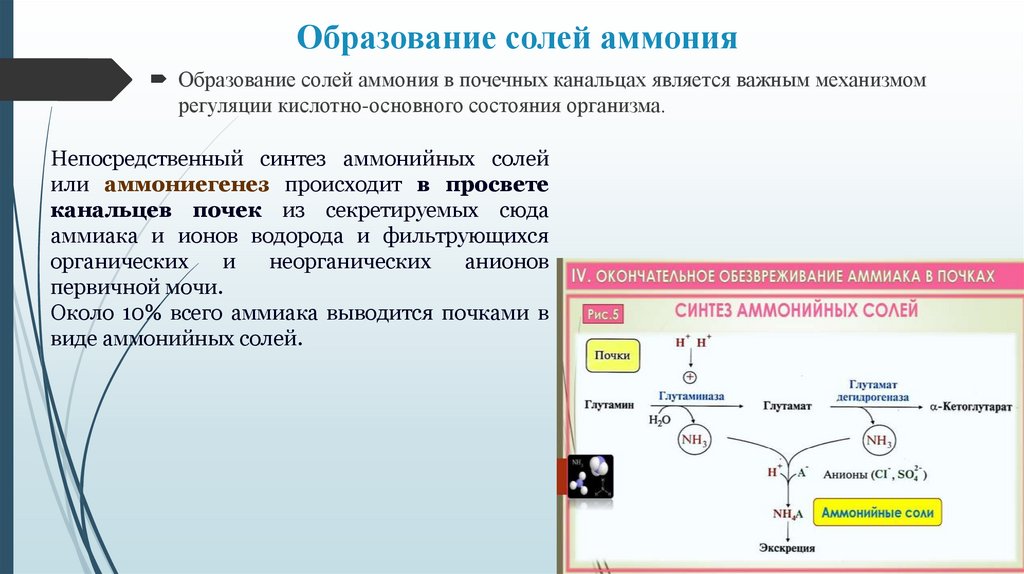
Образование солей аммонияОбразование солей аммония в почечных канальцах является важным механизмом
регуляции кислотно-основного состояния организма.
Непосредственный синтез аммонийных солей
или аммониегенез происходит в просвете
канальцев почек из секретируемых сюда
аммиака и ионов водорода и фильтрующихся
органических
и
неорганических
анионов
первичной мочи.
Около 10% всего аммиака выводится почками в
виде аммонийных солей.
29.
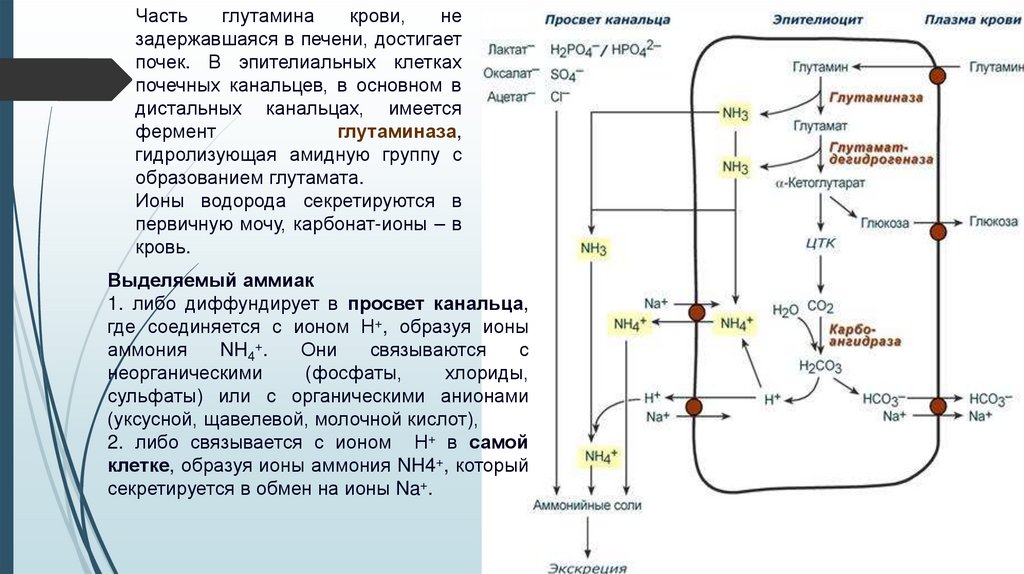
Частьглутамина
крови,
не
задержавшаяся в печени, достигает
почек. В эпителиальных клетках
почечных канальцев, в основном в
дистальных канальцах, имеется
фермент
глутаминаза,
гидролизующая амидную группу с
образованием глутамата.
Ионы водорода секретируются в
первичную мочу, карбонат-ионы – в
кровь.
Выделяемый аммиак
1. либо диффундирует в просвет канальца,
где соединяется с ионом Н+, образуя ионы
аммония
NH4+.
Они
связываются
с
неорганическими
(фосфаты,
хлориды,
сульфаты) или с органическими анионами
(уксусной, щавелевой, молочной кислот),
2. либо связывается с ионом Н+ в самой
клетке, образуя ионы аммония NH4+, который
секретируется в обмен на ионы Na+.
30.
31.
32.
33.
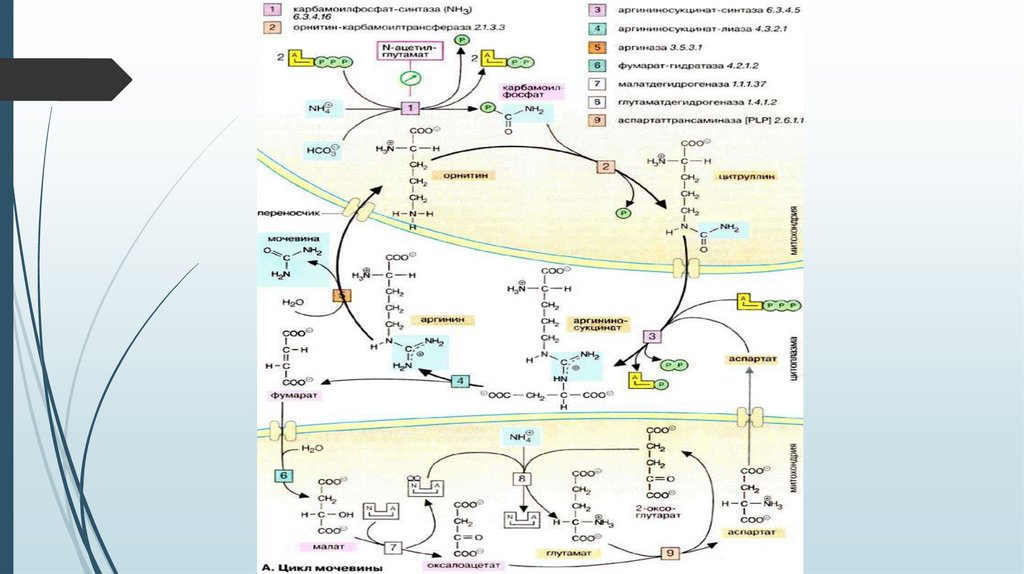
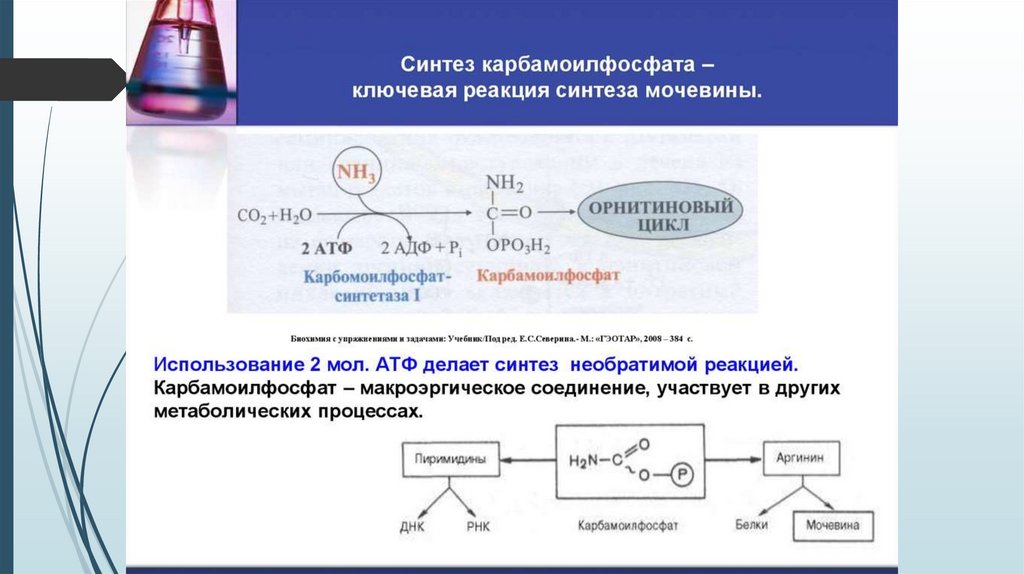
Орнитиновый цикл образования мочевины34.
35.
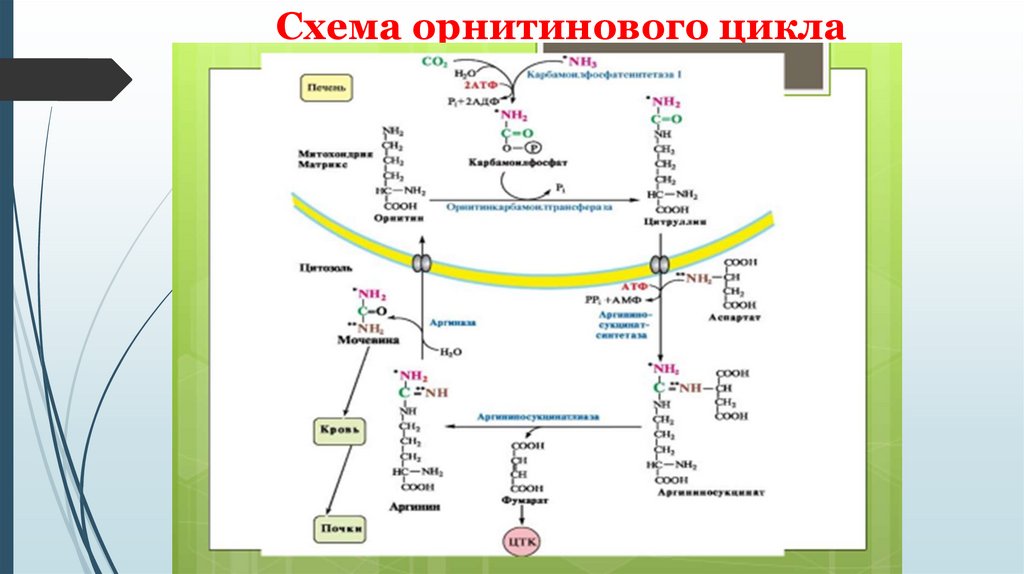
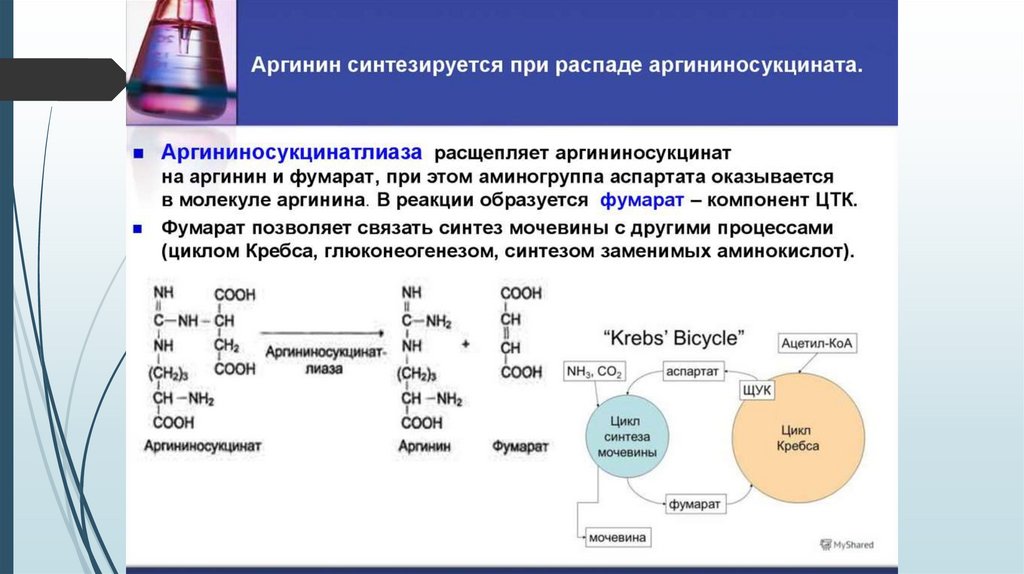
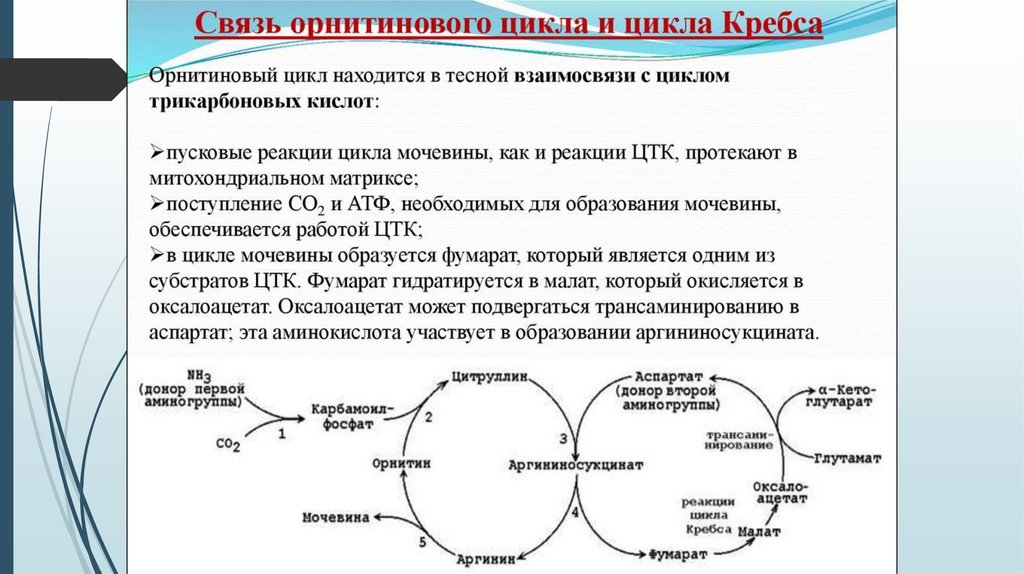
Связь орнитинового цикла с ЦТКОрнитиновый цикл находится в тесной взаимосвязи с циклом
трикарбоновых кислот:
пусковые реакции цикла мочевины, как и реакции ЦТК, протекают в
митохондриальном матриксе;
поступление СО2 и АТФ, необходимых для образования мочевины,
обеспечивается работой ЦТК;
в цикле мочевины образуется фумарат, который является одним из
субстратов ЦТК. Фумарат гидратируется в малат, который в свою
очередь окисляется в оксалоацетат. Оксалоацетат может подвергаться
трансаминированию в аспартат; эта аминокислота участвует в
образовании аргининосукцината.
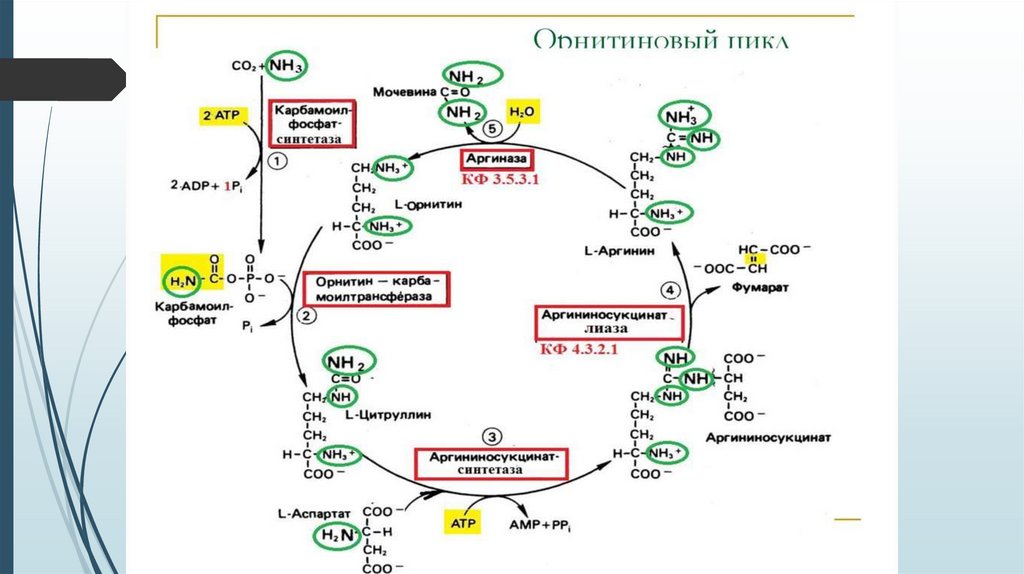
36.
Схема орнитинового цикла37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
Регуляция цикла мочевины1. осуществляется главным образом на уровне карбамоилфосфатсинтетазы,
которая малоактивна в отсутствие своего аллостерического активатора - Nацетил-глутамата. Концентрация последнего зависит от концентрации его
предшественников (ацетил-КоА и глутамата), а также аргинина, который
является аллостерическим активатором N-ацетилглутаматсинтазы.
2. концентрация ферментов орнитинового цикла зависит от содержания белка в
пищевом рационе:
при переходе на диету, богатую белком, в печени повышается синтез
ферментов орнитинового цикла.
при возвращении к сбалансированному рациону концентрация ферментов
снижается.
в условиях голодания, когда усиливается распад тканевых белков и
использование аминокислот как энергетических субстратов, возрастает
продукция аммиака, концентрация ферментов орнитинового цикла
увеличивается.