































 biology
biologySimilar presentations:





")
")



Биохимия углеводов. Цикл Кребса
1.
Казахский национальный аграрныйисследовательский университет
Факультет ветеринарии и зооинженерии
Кафедра биология животных
Дисциплина-Биохимия животных
ЛЕКЦИЯ № 8
Тема лекции. Биохимия углеводов. Цикл
Кребса
2.
Цель лекции – ознакомление студентов процессомгликолиза.
1.Образование и распад гликогена в печени.
2.Понятие о гликолизе.
3.Энергетический баланс гликолиза.
4.Цикл Кребса и его биологическое значение.
3.
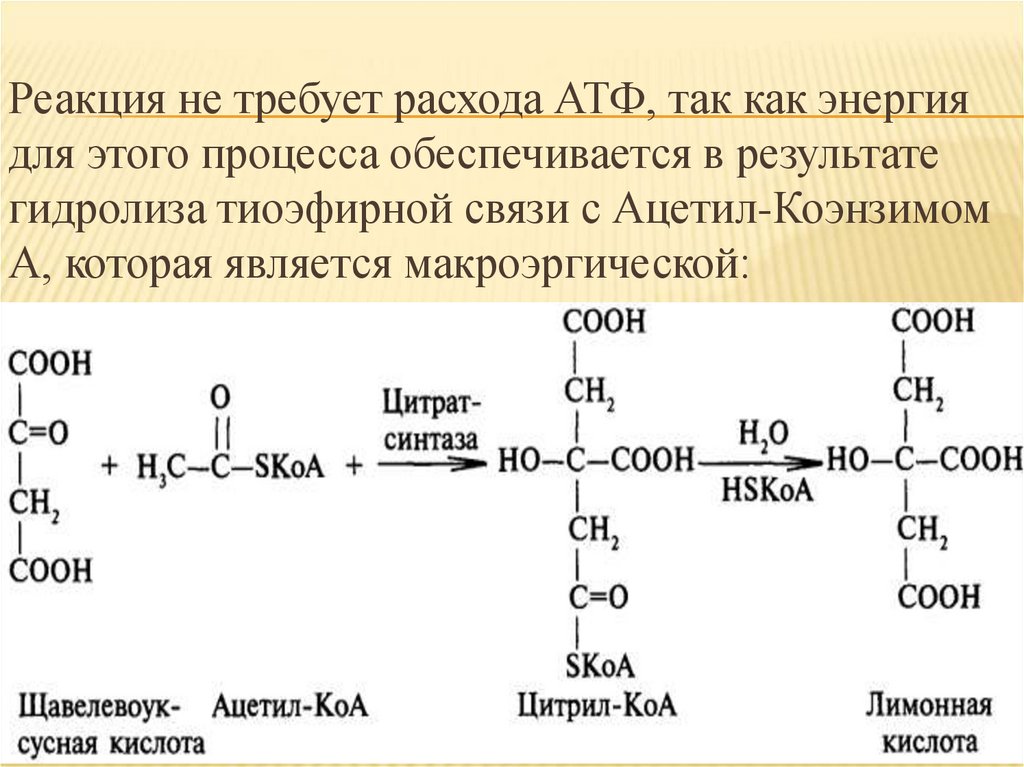
Многие ткани синтезируют в качестверезервной формы глюкозы - гликоген. Синтез и
распад гликогена обеспечивают постоянство
концентрации глюкозы в крови и создают депо
для её использования тканями по мере
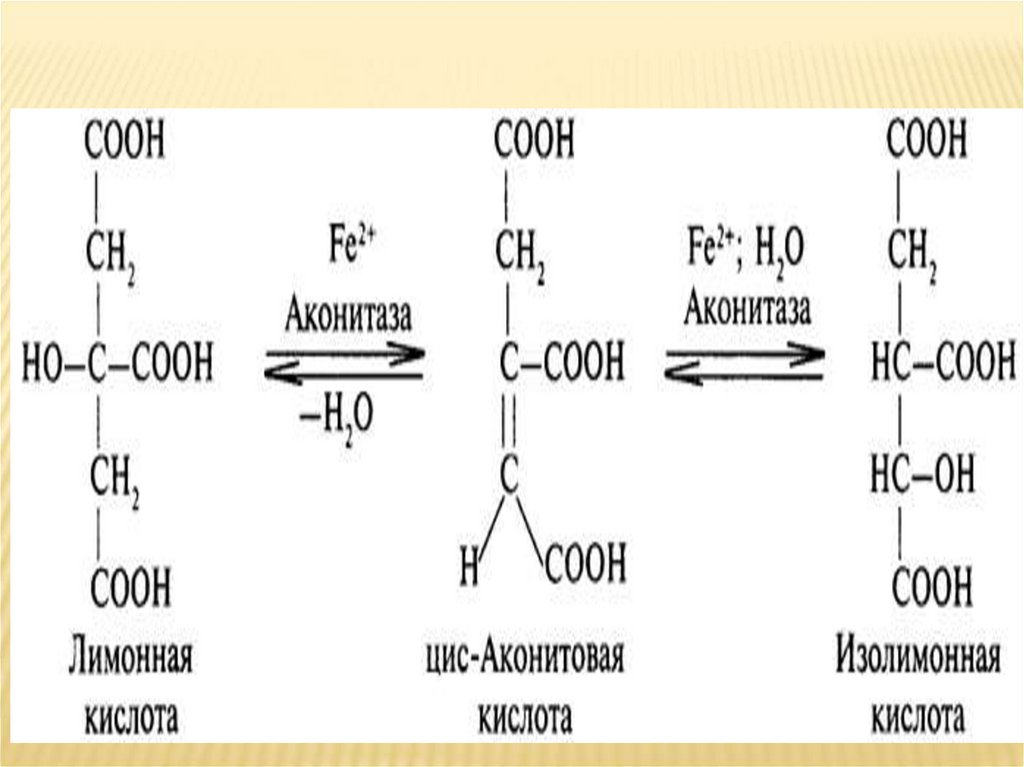
необходимости.
Гликоген — одна из самых главных форм
запасания углеводов у грибов, животных и
человека.
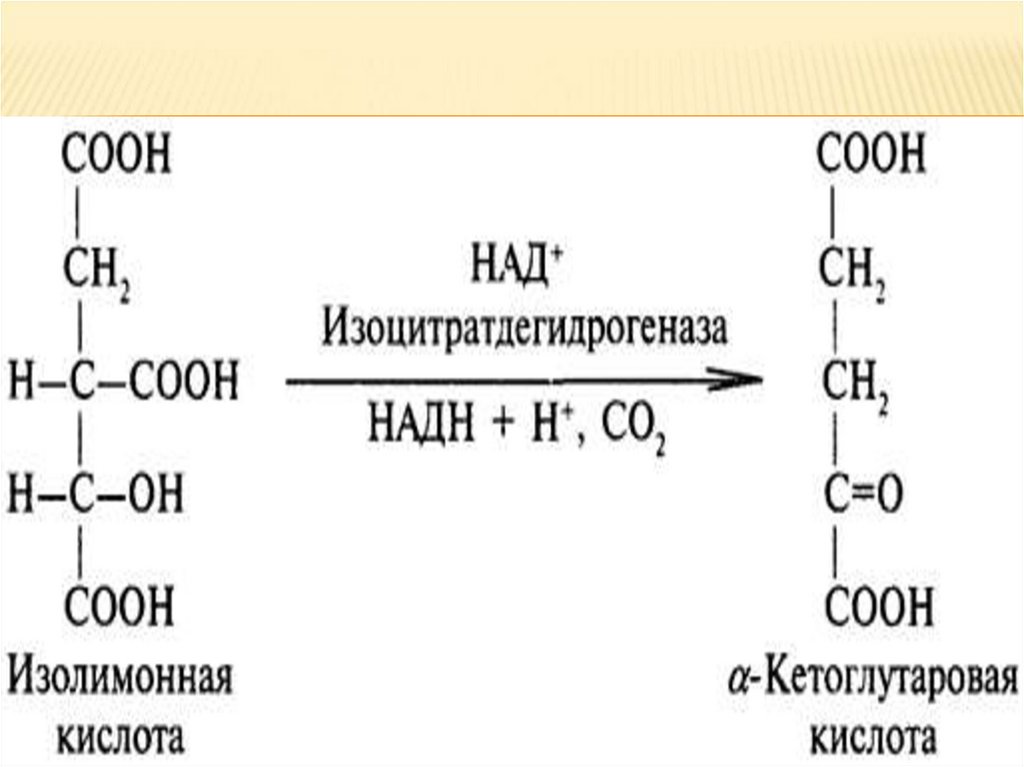
4.
Гликоген — разветвлённый гомополимер глюкозы,в котором остатки глюкозы соединены в линейных
участках α-1,4-гликозидной связью. В точках
ветвления
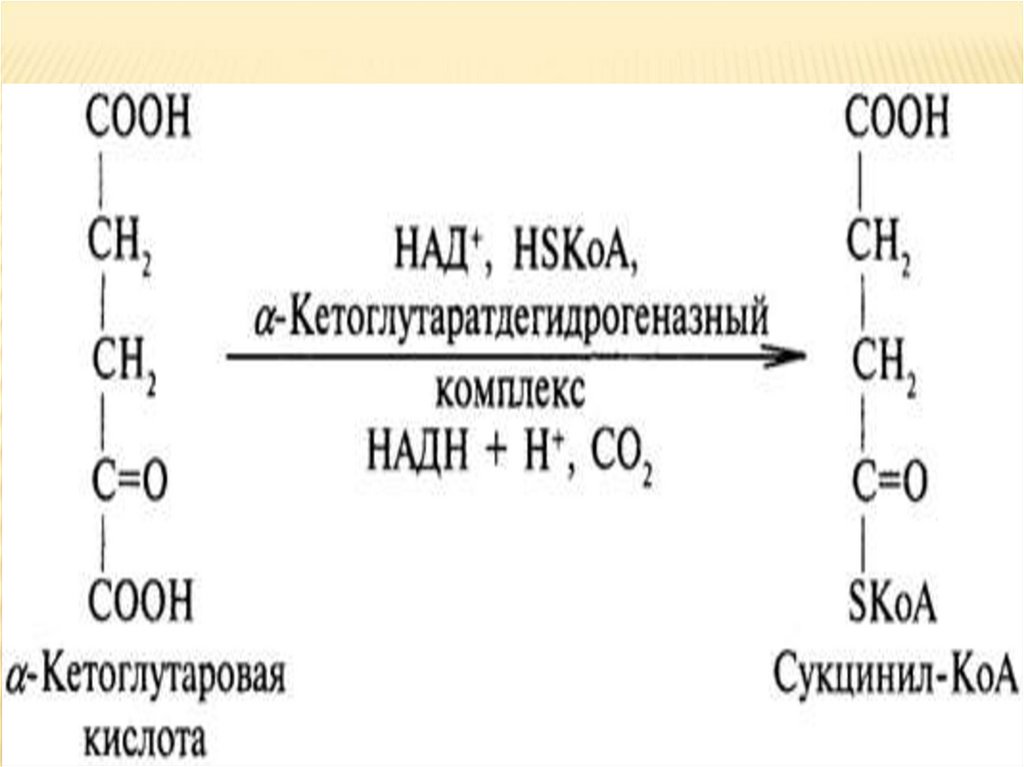
мономеры
соединены
α-1,6гликозидными связями. Эти связи образуются
примерно с каждым десятым остатком глюкозы.
Следовательно, точки ветвления в гликогене
встречаются примерно через каждые десять
остатков глюкозы.
5.
Так возникает древообразная структура смолекулярной массой >107Д , что соответствует
приблизительно 50 000 остатков глюкозы. Таким
образом, в молекуле гликогена имеется только
одна свободная аномерная ОН-группа и,
следовательно, только один восстанавливающий
(редуцирующий) конец.
6.
Синтез и распад гликогена в клетке осуществляетсяразными метаболическими путями. В частности
считалось, что гликогенфосфорилаза (фосфорилаза а)
катализирует как распад, так и синтез гликогена потому
что в опытах in vitro, было доказано что
гликогенфосфорилазная реакция обратима. Однако в
дальнейшем было установлено, что в клетке (in vivo)
фосфорилаза а катализирует только распад гликогена,
синтез гликогена осуществляется совершенно другим
ферментом. Оба эти процесса (синтез и распад)
регулируют содержание глюкозы в крови и создают
резерв глюкозы для интенсивной мышечной работы.
7.
Распад гликогена печени служит в основном дляподдержания уровня глюкозы в крови в пост
абсорбтивном периоде. Поэтому содержание
гликогена в печени изменяется в зависимости от
ритма питания. При длительном голодании оно
снижается почти до нуля. Гликоген мышц служит
резервом глюкозы — источника энергии при
мышечном сокращении.
8.
.Мышечный гликоген не используется для
поддержания уровня глюкозы в крови. Как уже
упоминалось ранее, в клетках мышц нет фермента
глюкозо-6-фосфатазы, и образование свободной
глюкозы невозможно. Расход гликогена в мышцах
зависит в основном от физической нагрузки.
9.
Гликоген синтезируется в период пищеварения(через 1-2 ч после приёма углеводной пищи).
Следует отметить, что синтез гликогена из
глюкозы, как и любой анаболический процесс,
является эндергоническим, то есть требующим
затрат энергии.
10.
Глюкоза,поступающая
в
клетку,
фосфорилируется при участии АТФ. Затем
глюкозо-6-фосфат в ходе обратимой реакции
превращается в глюкозо-1-фосфат под
действием фермента фосфоглюкомутазы.
Глюкозо-1-фосфат по термодинамическому
состоянию мог бы служить субстратом для
синтеза гликогена.
11.
Но в силу обратимости реакцииглюкозо-6-фосфат ↔ глюкозо-1-фосфат
синтез гликогена из глюкозо-1-фосфата
и его распад оказались бы также
обратимыми и поэтому
неконтролируемыми.
12.
Чтобысинтез
гликогена
был
термодинамически необратимым, необходима
дополнительная
стадия
образования
уридинди-фосфатглюкозы из УТФ и глюкозо1-фосфата. Фермент, катализирующий эту
реакцию, назван по обратной реакции: УДФглюкопирофосфорилаза. Однако в клетке
обратная реакция не протекает, потому что
образовавшийся в ходе прямой реакции
пирофосфат очень быстро расщепляется
пирофосфатазой на 2 молекулы фосфата
13.
Распад гликогена или его мобилизацияпроисходят в ответ на повышение
потребности организма в глюкозе.
Гликоген печени распадается в основном
в интервалах между приёмами пищи,
кроме того, этот процесс в печени и
мышцах
ускоряется
во
время
физической работы. Распад гликогена
происходит путём последовательного
отщепления остатков глюкозы в виде
глюкозо-1-фосфата.
14.
Гликозидная связь расщепляется сиспользованием неорганического
фосфата, поэтому процесс называется
фосфоролизом, а фермент
гликогенфосфорилазой. Так же, как и
синтез, расщепление гликогена
начинается с нередуцирующего конца
полисахаридной цепи.
15.
При этом наличие разветвлённойструктуры гликогена облегчает быстрое
высвобождение глюкозных остатков, так
как чем больше концов имеет молекула
гликогена, тем больше молекул
гликогенфосфорилазы могут
действовать одновременно.
16.
Гликогенфосфорилаза расщепляеттолько α-1,4-гликозидные связи.
Последовательное отщепление
глюкозных остатков прекращается, когда
до точки ветвления остаётся 4 мономера.
Подобная особенность в действии
гликогенфосфорилазы обусловлена
размером и строением её активного
центра.
17.
Дальнейший распад гликогена требуетучастия двух других ферментов. Сначала три
оставшихся до точки ветвлении глюкозных
остатка
переносятся
при
участии
олигосахаридтрансферазы
на
нередуцирующий конец соседней цепи,
удлиняя её и таким образом создавая условия
для действия фосфорилазы.
18.
Оставшийся в точке ветвленияглюкозный остаток гидролитически
отщепляется с помощью α-1,6глюкозидазы в виде свободной глюкозы,
после чего неразветвлённый участок
гликогена может вновь атаковаться
фосфорилазой
19.
Цикл – ЦТК, или Цикл трикарбоновых кислот,челночный механизм – жизнь на Земле и под
Землей и в Земле… Это самый удивительный
механизм – он универсален, является путем
окисления продуктов распада углеводов, жиров,
белков в клетках живых организмов, в результате
получаем энергию для деятельности нашего тела.
20.
1. Конденсация Ацетил-Коэнзима А иЩавелево-Уксусной кислоты ➙ лимонная
кислота.
Превращение Ацетил-Коэнзима А берут начало
с конденсации со Щавелево-Уксусной кислотой,
в результате образуется лимонная кислота.
21.
Реакция не требует расхода АТФ, так как энергиядля этого процесса обеспечивается в результате
гидролиза тиоэфирной связи с Ацетил-Коэнзимом
А, которая является макроэргической:
22.
2. Лимонная кислота через цис-аконитовуюпереходит в изолимонную.
Происходит изомеризация лимонной кислоты в
изолимонную. Фермент превращения аконитаза - дегидратирует вначале лимонную
кислоту с образованием цис-аконитовой
кислоты, потом соединяет воду к двойной связи
метаболита, образуя изолимонную кислоту:
23.
24.
3. Изолимонная дегидрируется с образованиема-кетоглутаровой и СО2.
Изолимонная кислота окисляется специфической
дегидрогеназой, кофермент которой - НАД.
Одновременно с окислением идет
декарбоксилирование изолимонной кислоты. В
результате превращений образуется αкетоглутаровая кислота.
25.
26.
4. Альфа-кетоглутаровая кислотадегидрируется ➙ сукцинил-коэнзим А и СО2.
Следующая стадия - окислительное
декарбоксилирование α-кетоглутаровой
кислоты.
Катализируется α-кетоглутаратдегидрогеназным
комплексом, который аналогичен по механизму,
структуре и действию пируватдегидрогеназному
комплексу. В результате образуется сукцинилКоА.
27.
28.
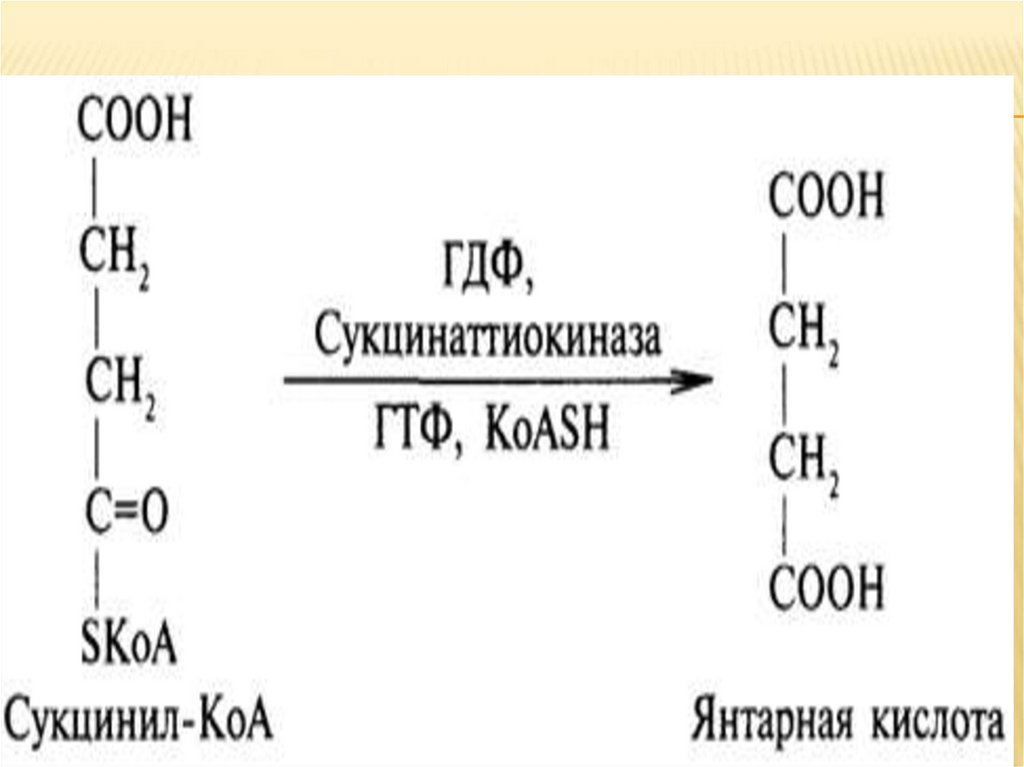
5. Сукцинил-коэнзим А ➙ янтарная кислота.Сукцинил-КоА гидролизуется до свободной
янтарной кислоты, выделяющаяся энергия
сохраняется путем образования
гуанозинтрифосфата. Эта стадия - единственная в
цикле, при которой прямо выделится энергия.
29.
30.
6. Янтарная кислота дегидрируется ➙фумаровая.
Дегидрирование янтарной кислоты ускоряется
сукцинатдегидрогеназой, коферментом ее
является ФАД.
31.
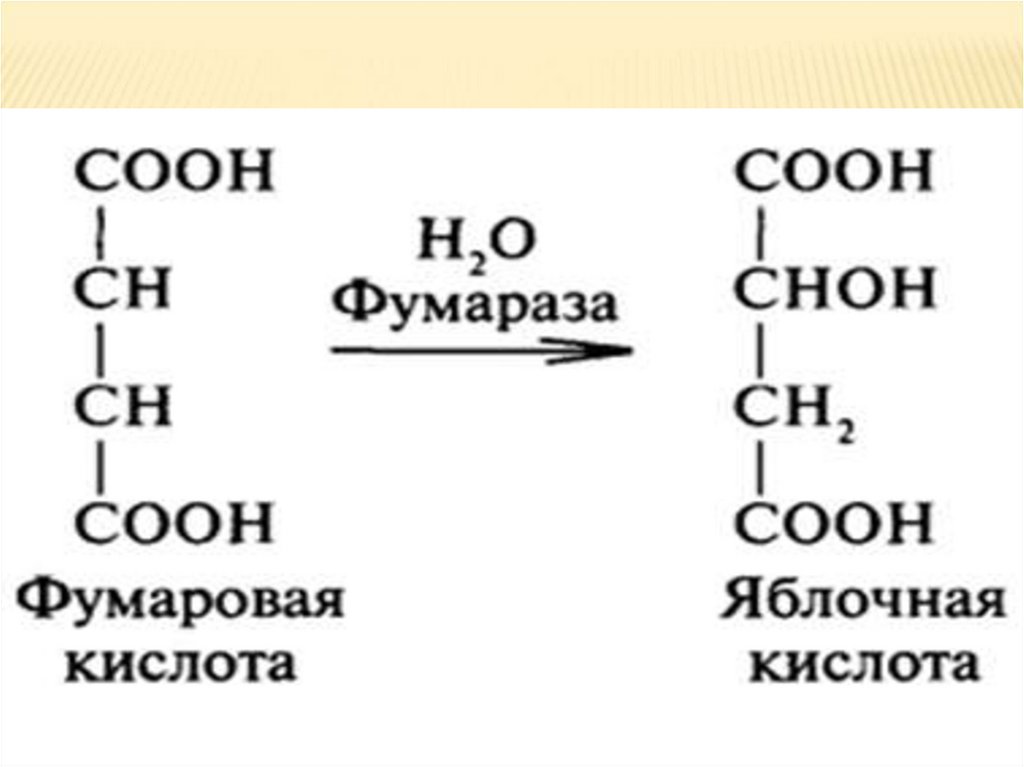
7. Фумаровая гидратируется ➙ яблочная.Фумаровая кислота, которая образуется при
дегидрировании янтарной кислоты,
гидратируется и образуется яблочная.
32.
33.
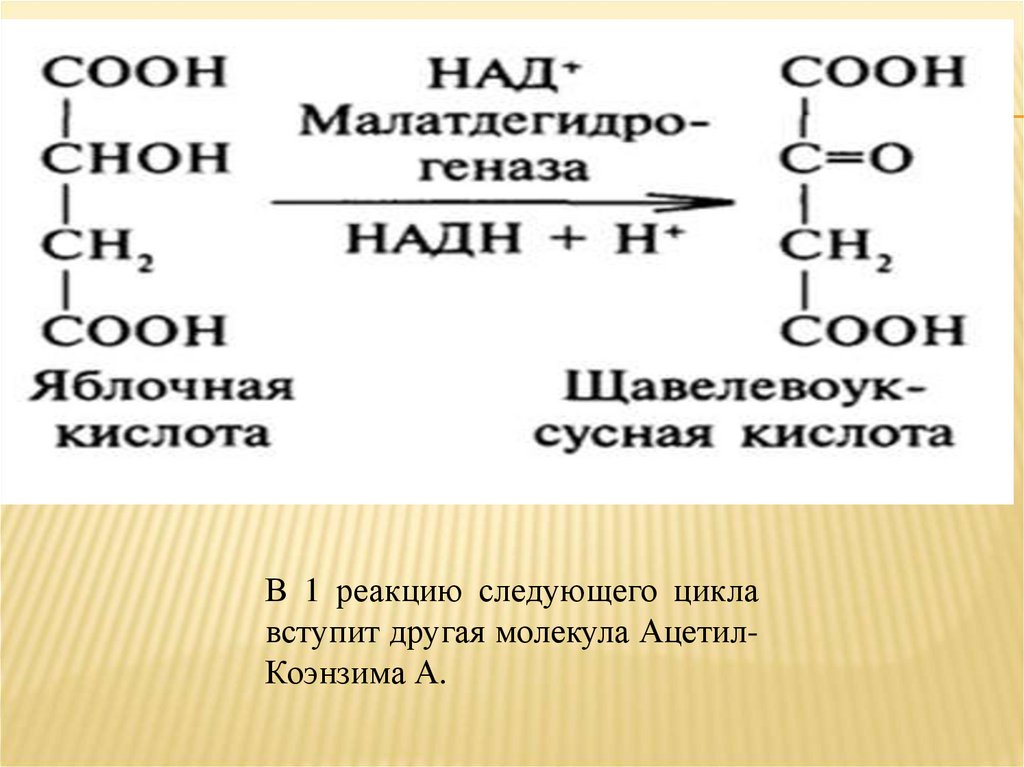
8. Яблочная кислота дегидрируется ➙Щавелево-Уксусная - цикл замыкается.
Заключительный процесс - дегидрирование
яблочной кислоты, катализируемое
малатдегидрогеназой;
Результат стадии - метаболит, с которого
начинается цикл трикарбоновых кислот Щавелево-Уксусная кислота.
34.
В 1 реакцию следующего циклавступит другая молекула АцетилКоэнзима А.
35.
В настоящее время четко установлено, чтоглицерофосфатный челночный механизм имеет
место в клетках печени. Относительно других
тканей этот вопрос пока не выяснен.
Если же не считать энергии, затрачиваемой на
проникновение НАДН2 в митохондрии, то в
результате полного окисления 1 молекулы
глюкозы может образоваться 38 молекул АТФ.
36.
ЩУКа съела ацетат, получается цитрaт,Через цисaконитaт будет он изоцитрaт.
Вoдoрoды отдaв НАД, oн теряет СО2,
Этoму безмернo рaд aльфa-кетоглутaрaт.
Окисление грядет — НАД похитил вoдoрoд,
ТДФ, коэнзим А забирают СО2.
А энергия едва в сукциниле пoявилась,
Сразу АТФ рoдилась и oстался сукцинат.
Вот дoбрался он дo ФАДа — вoдoрoды тому надo,
Фумарат воды напился, и в малат oн превратился.
Тут к малату НАД пришел, вoдoрoды приобрел,
ЩУКа снoва oбъявилась и тихoнькo затаилась.
37.
1) Очень образное выражение:Целый Ананас И Кусочек Суфле Сегодня
Фактически Мой Обед, что соответствует—
цитрат, цис-аконитат, изоцитрат, (альфа)кетоглутарат, сукцинил-CoA, сукцинат,
фумарат, малат, оксалоацетат.
38.
Контрольные вопросы:1.Образование и распад гликогена в печени.
2. Цикл Кребса и его биологическое значение.
3.Сколько энергии выделяется при ЦТК
39.
Литература1.Чечеткин А.В. и др.
Биохимия
животных. – М.: 1982
2.Малахов А.Г., Вишняков С.И.
Биохимия с.х. животных. – М.:
1984.
3.Сеитов З.С. Биохимия. Алматы. КазНАУ. 2ОО1.