


























 biology
biologySimilar presentations:


")







Механизм действия гормонов
1.
КАЗАХСКИЙ НАЦИОНАЛЬНЫЙ АГРАРНЫЙУНИВЕРСИТЕТ
Дисциплина-Биохимия
ЛЕКЦИЯ № 5
Тема лекции: Механизм действия гормонов
Лектор – Саримбекова С.Н.
2.
Цель: Ознакомить студентов с механизмомдействия гормонов
План:
1.Механизм действия стероидных гормонов.
2. Гормон роста (соматотропин)
3. Тиреоидные гормоны
4. Тиреоидные гормоны в крови
3.
Гормоны – это продукты внутреннейсекреции, которые вырабатываются
специальными железами или отдельными
клетками, выделяются в кровь и разносятся
по всему организму в норме вызывая
определенный биологический эффект.
Сами гормоны непосредственно не влияют
на какие-либо реакции клетки. Только
связавшись с определенным, свойственным
только ему рецептором вызывается
определенная реакция.
4.
Гормоны имеют различную химическуюструктуру. Это приводит к тому, что они
имеют
разные
физические
свойства.
Гормоны
разделяют
на
водои
жирорастворимые.
Принадлежность
к
какому-то из этих классов обуславливает их
механизм действия. Это объясняется тем, что
жирорастворимые гормоны могут спокойно
проникать через клеточную мембрану,
которая состоит преимущественно из бислоя
липидов, а водорастворимые этого не могут.
5.
В связи с этим рецепторы(Р) для водо- ижирорастворимых
гормонов
имеют
различное место локализации (мембрана и
цитоплазма). Связавшись с мембранным
рецептором гормон вызывает каскад
реакций в самой клетке, но никак не влияет
на генетический материал. Комплекс
цитоплазматического Р и гормона может
воздействовать на ядерные рецепторы и
вызывать
изменения
в
генетическом
аппарате, что ведет к синтезу новых белков.
Рассмотрим это поподробнее.
6.
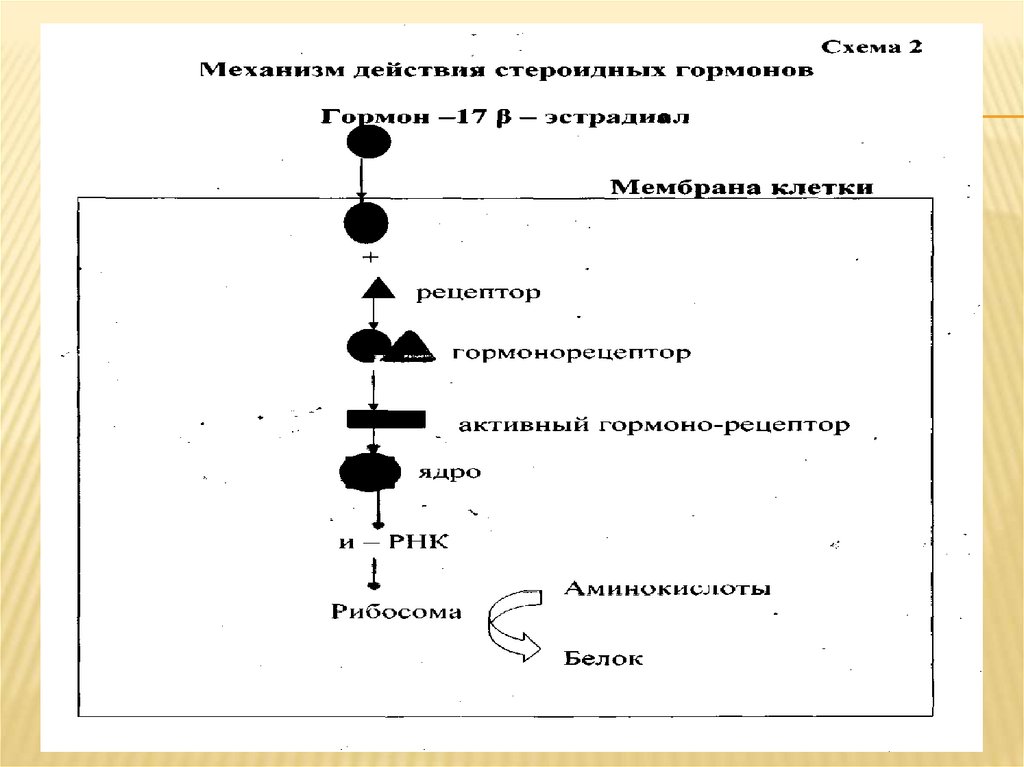
Механизм действия стероидных (жирорастворимых) гормоновПроникновение стероида (С) в клетку
Образование комплекса СР. Все Р стероидных гормонов представляют
собой глобулярные белки примерно одинакового размера, с очень
высоким сродством связывающие гормоны
Трансформация СР в форму, способную связываться ядерными
акцепторами [СР] Любая клетка содержит всю генетическую
информацию. Однако при специализации клетки большая часть ДНК
лишается возможности быть матрицей для синтеза иРНК. Это
достигается путем сворачивания вокруг белков гистонов, что ведет к
препятствию транскрипции. В связи с этим генетический материал
клетки
можно
разделить
на
ДНК
3-х
видов:
1.транскрипционно
неактивная
2.постоянно
экспрессируемая
3.индуцируемая гормонами или другими сигнальными молекулами.
Связывание
[СР]
с
хроматиновым
акцептором.
Следует отметить, что этот этап действия С полностью не изучен и
имеет ряд спорных моментов. Считается что [СР] взаимодействует со
специфическими участками ДНК так, что это дает возможность РНКполимеразе вступить в контакт к определенным доменам ДНК.
7.
Клетки-мишени - это клетки, которыеспецифически взаимодействуют с гормонами с
помощью специальных белков-рецепторов. Эти
белки-рецепторы располагаются на наружной
мембране клетки, или в цитоплазме, или на ядерной
мембране и на других органеллах клетки.
Биохимические механизмы передачи сигнала от
гормона в клетку-мишень.
Любой белок-рецептор состоит, минимум из двух
доменов (участков), которые обеспечивают
выполнение двух функций:
узнавание гормона;
преобразование и передачу полученного сигнала в
клетку.
8.
Один из доменов белка-рецептора имеет в своемсоставе участок, комплементарный какой-то части
сигнальной молекулы. Процесс связывания
рецептора с сигнальной молекулой похож на
процесс образования фермент-субстратного
комплекса и может определяется величиной
константы сродства.
Большинство рецепторов изучены недостаточно,
потому что их выделение и очистка очень сложные,
а содержание каждого вида рецепторов в клетках
очень низкое. Гормоны взаимодействуют со своими
рецепторами физико-химическим путем. Между
молекулой гормона и рецептором формируются
электростатические и гидрофобные
взаимодействия.
9.
При связывании рецептора с гормономпроисходят конформационные изменения белкарецептора и комплекс сигнальной молекулы с
белком-рецептором активируется. В активном
состоянии он может вызывать специфические
внутриклеточные реакции в ответ на принятый
сигнал. Если нарушен синтез или способность
белков-рецепторов связываться с сигнальными
молекулами, возникают заболевания эндокринные нарушения.
10.
Есть три типа таких заболеваний.1.Связанные с недостаточностью синтеза
белков-рецепторов.
2.Связанные с изменением структуры рецептора
- генетические дефекты.
3.Связанные с блокированием белковрецепторов антителами.
11.
В зависимости от строения гормонасуществуют два типа взаимодействия. Если
молекула гормона липофильна, (например,
стероидные гормоны), то она может
проникать через липидный слой наружной
мембраны клеток-мишеней. Если молекула
имеет большие размеры или является
полярной, то ее проникновение внутрь
клетки невозможно. Поэтому для
липофильных гормонов рецепторы
находятся внутри клеток-мишеней, а для
гидрофильных - рецепторы находятся в
наружной мембране.
12.
Для получения клеточного ответа нагормональный сигнал в случае гидрофильных
молекул действует внутриклеточный механизм
передачи сигнала. Это происходит с участием
веществ,
которых
называют
вторыми
посредниками. Молекулы гормонов очень
разнообразны
по
форме,
а
"вторые
посредники" - нет.
Надежность передачи сигнала обеспечивает
очень высокое сродство гормона к своему
белку-рецептору.
13.
Это циклические нуклеотиды (цАМФ ицГМФ), инозитолтрифосфат, кальцийсвязывающий белок - кальмодулин, ионы
кальция, ферменты, участвующие в синтезе
циклических нуклеотидов, а также
протеинкиназы - ферменты
фосфорилирования белков. Все эти вещества
участвуют в регуляции активности
отдельных ферментных систем в клеткахмишенях.
14.
Существует два главных способа передачисигнала в клетки-мишени от сигнальных
молекул с мембранным механизмом
действия:
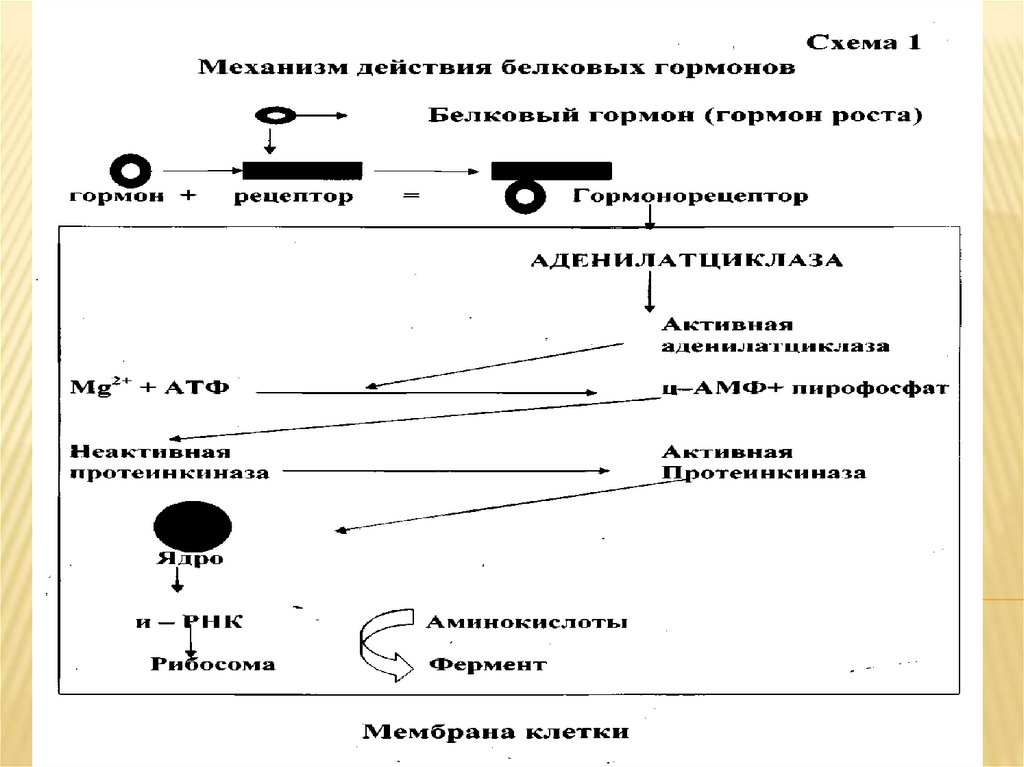
1. Аденилатциклазная (или
гуанилатциклазная) системы;
2. Фосфоинозитидный механизм.
15.
Аденилатциклазная система.Основные компоненты: мембранный белокрецептор, G-белок, фермент аденилатциклаза,
гуанозинтрифосфат, протеинкиназы.
Кроме того, для нормального
функционирования аденилатциклазной
системы, требуется АТФ.
Белок-рецептор, G-белок, рядом с которым
располагаются ГТФ и фермент
(аденилатциклаза) встроены в мембрану клетки.
16.
До момента действия гормона эти компонентынаходятся в диссоциированнном состоянии, а
после образования комплекса сигнальной
молекулы с белком-рецептором происходят
изменения конформации G-белка. В результате
одна из субъединиц G-белка приобретает
способность связываться с ГТФ.
Комплекс "G-белок-ГТФ" активирует
аденилатциклазу. Аденилатциклаза начинает
активно превращать молекулы АТФ в ц-АМФ.
17.
ц-АМФ обладает способностью активироватьособые ферменты - протеинкиназы, которые
катализируют реакции фосфорилирования
различных белков с участием АТФ. При этом в
состав белковых молекул включаются остатки
фосфорной кислоты. Главным результатом этого
процесса фосфорилирования является
изменение активности фосфорилированного
белка. В различных типах клеток
фосфорилированию в результате активации
аденилат-циклазной системы подвергаются
белки с разной функциональной активностью.
18.
В результате реакции фосфорилирования белкимогут становятся функционально активными
или неактивными.
Такие процессы будут приводить к изменениям
скорости биохимических процессов в клеткемишени.
Активация аденилатциклазной систтемы длится
очень короткое время, потому что G-белок после
связывания с аденилатциклазой начинает
проявлять ГТФ-азную активность. После
гидролиза ГТФ G-белок восстанавливает свою
конформацию и перестает активировать
аденилатциклазу. В результате прекращается
реакция образования ц-АМФ.
19.
Когда останавливается образование цАМФ,реакции фосфорилирования в клетке
прекращаются не сразу: пока продолжают
существовать молекулы цАМФ - будет
продолжаться и процесс активации протеинкиназ.
Для того, чтобы прекратить действие цАМФ, в
клетках существует специальный фермент фосфодиэстераза, который катализирует реакцию
гидролиза 3',5'-цикло-АМФ до АМФ.
20.
Некоторые вещества, обладающиеингибирующим действием на
фосфодиэстеразу, (например, алкалоиды
кофеин, теофиллин), способствуют
сохранению и увеличению концентрации
цикло-АМФ в клетке. Под действием этих
веществ в организме продолжительность
активации аденилатциклазной системы
становится больше, т. е. усиливается действие
гормона.
21.
Инозитолтрифосфат - это вещество, котороеявляется производным сложного липида инозитфосфатида. Оно образуется в результате
действия специального фермента фосфолипазы "С", который активируется в
результате конформационных изменений
внутриклеточного домена мембранного белкарецептора.
Этот фермент гидролизует фосфоэфирную
связь в молекуле фосфатидил-инозитол-4,5бисфосфата и в результате образуются
диацилглицерин и инозитолтрифосфат.
22.
Образование диацилглицерина иинозитолтрифосфата приводит к увеличению
концентрации ионизированного кальция внутри
клетки. Это приводит к активации многих
кальций-зависимых белков внутри клетки, в
том числе активируются различные
протеинкиназы. И здесь, как и при активации
аденилатциклазной системы, одной из стадий
передачи сигнала внутри клетки является
фосфорилирование белков, которое в приводит
к физиологическому ответу клетки на действие
гормона.
23.
В работе фосфоинозитидного механизмапередачи сигналов в клетке-мишени
принимает участие специальный кальцийсвязывающий белок - кальмодулин. Это
низкомолекулярный белок (17 кДа), на 30 %
состоящий из отрицательно заряженных
аминокислот (Глу, Асп) и поэтому способный
активно связывать Са+2. Одна молекула
кальмодулина имеет 4 кальций-связывающих
участка.
24.
После взаимодействия с Са+2 происходятконформационные изменения молекулы
кальмодулина и комплекс "Са+2-кальмодулин"
становится способным регулировать активность
(аллостерически угнетать или активировать)
многие ферменты - аденилатциклазу,
фосфодиэстеразу, Са+2,Мg+2-АТФазу и
различные протеинкиназы.
25.
Таким образом, в роли "вторых посредников"для передачи сигналов от гормонов в клеткахмишенях могут быть:
1. циклические нуклеотиды (ц-АМФ и ц-ГМФ);
ионы Са;
2. комплекс "Са-кальмодулин";
3. диацилглицерин;
4. инозитолтрифосфат.
26.
27.
28.
Контрольные вопросы:1.Что такое гормоны?
2. Где синтезируются гормоны?
3. Что такое клетки мишени?
4. Опишите механизм действия стероидных
гормонов?
29.
1. Руководство к практическим занятиям побиологической химии. Под ред. Тапбергенова С.О.
2012 г.
2. Биохимия. Под ред. Е.С.Северина. 2009 г.
3. Сеитов З.С. Биохимия : Өзгертіліп,
толықтырылып үшінші рет басылып шығуы / З. С.
Сеитов. - Алматы : "ЭВЕРО", 2012. - 570 с. : ил.
3. Биохимия сұрақтары мен жауаптары. Биохимия
в вопросах и ответах: учебное пособие / Т. С.
Сейтембетов, Б. И. Төлеуов. - Алматы :
ТОО"ЭВЕРО", 2010 г.