




")





























 biology
biologySimilar presentations:




")





Цитология старения. Окислительная теория старения
1. ЦИТОЛОГИЯ СТАРЕНИЯ
Окислительная теория старения2.
Aging is defined when two criteriaare met.
First, the probability of death at any
point in time increases with the
age of the organism. This
statistical definition applies from
yeast to mammals and reflects the
progressive nature of aging.
Second, characteristic changes in
phenotype occur in all individuals
over time due to the limiting
processes.
3.
Impressive progress havebeen done in the framework
of two main paradigms of
gerontology —
programmed
and
stochastic aging.
4. Теории и гипотезы старения, альтернативные теломерной
1. Окислительная теория (Дэнхем Харман, 1956)
2. Репарационные теории
3. Теория SIRT- белков
4. Теория феноптоза и запрограммированного
старения
5. Митохондриальные теории
6. Надклеточные теории:
Эндокринные (нейроэндокринная гипоталамус)
Андростероновая (надпочечники)
Мелатониновая (эпифиз)
Инволюции половых желез
7. Иммунные теории
8. Мозговые теории
5.
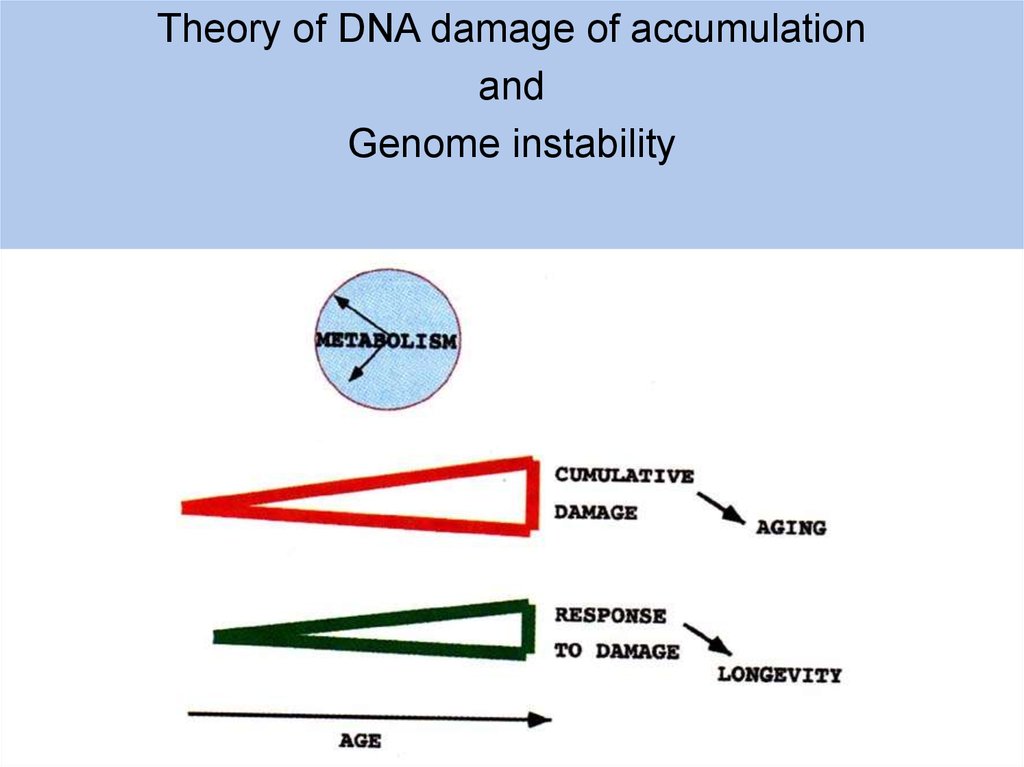
Theory of DNA damage of accumulationand
Genome instability
6. Essential Features of Aging in Saccharomyces cerevisiae
7. Дэнхем Харман Denham Harman (February 14, 1916 November 25, 2014)
Дэнхем ХарманDenham Harman (February 14, 1916 November 25, 2014)
• B 1954 году впервые
высказал идею о том, что в
организме могут протекать
реакции с образованием
свободных радикалов, и что
эти реакции лежат в основе
многих патологических
процессов.
8.
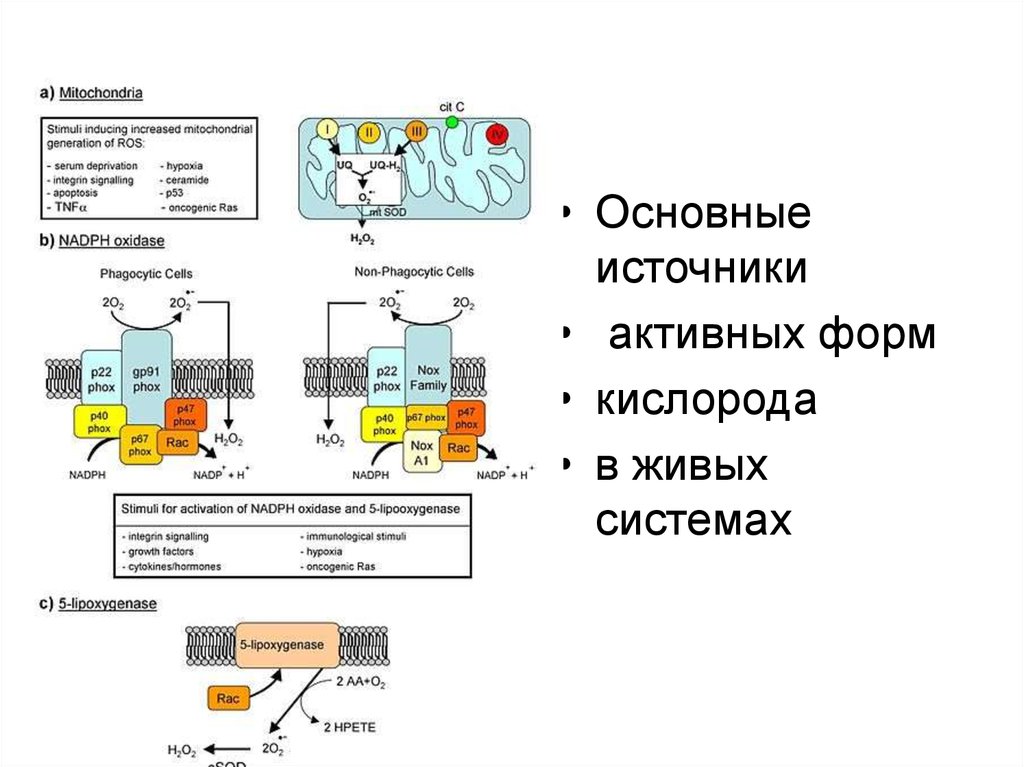
• Основныеисточники
• активных форм
• кислорода
• в живых
системах
9.
Free radical theory or the theory of reactiveoxygen species (ROS) of aging
(Harman 1981)
ROS:
• superoxide anion
• hydrogen peroxide
• hydroxyl radical
ROS have been shown to cause molecular
damage relatively indiscriminately to proteins,
lipids, and nucleic acids.
10. Дыхательная цепь митохондрий
Комплекс I (НАДН дегидрогеназа)
окисляет НАД-Н, отбирая у него
два электрона и перенося их на
растворимый в липидах убихинон,
который внутри мембраны
диффундирует к комплексу III.
Вместе с этим, комплекс I
перекачивает 2 протона и 2
электрона из матрикса в
межмембранное пространство
митохондрии.
Комплекс II (Сукцинат
дегидрогеназа) не перекачивает
протоны, но обеспечивает вход в
цепь дополнительных электронов за
счёт окисления сукцината.
Комплекс III (Цитохром
bc1 комплекс) переносит электроны
с убихинона на два
водорастворимыхцитохрома с,
расположенных на внутренней
мембране митохондрии. Убихинон
передаёт 2 электрона, а цитохромы
за один цикл переносят по одному
электрону. При этом туда также
переходят 2 протона убихинона и
перекачиваются комплексом.
Комплекс IV (Цитохром c оксидаза)
катализирует перенос 4 электронов
с 4 молекул цитохрома на O2и
перекачивает при этом 4 протона в
межмембранное пространство.
Комплекс состоит из цитохромов a и
a3, которые, помимо гема, содержат
ионы меди.
Кислород, поступающий в
митохондрии из крови, связывается
с атомом железа в геме цитохрома
a3 в форме молекулы O2. Каждый из
атомов кислорода присоединяет по
два электрона и два протона и
превращается в молекулу воды.
11. Слабость окислительной теории старения
• Согласно окислительной теории, старение происходит врезультате накопления в клетках повреждений, вызываемых
активными формами кислорода (АФК).
• Нет сомнения в том, что окислительные повреждения ускоряют
старение, однако окислительная теория старения сама по себе
не в состоянии ответить на следующие вопросы:
• 1. Почему окислительные повреждения, вызываемые
активными формами кислорода, не накапливаются в ходе
созревания гамет, а если накапливаются, то куда они исчезают
при образовании зиготы?
• 2. Почему окислительные повреждения не накапливаются в
трансформированных клетках, а если накапливаются, то почему
не препятствуют пролиферации этих клеток? Тем более, что в
трансформированных клетках образуется не меньше, а больше
активных форм кислорода, чем в норме.
• На эти вопросы отвечает наличие теломеразы в этих клетках.
12.
• Вне всякого сомнения, АФК способныускорять старение как in vitro,так и in
vivo, но в огромной массе
накопленных данных о роли АФК в
старении трудно выявить собственно
механизм, с помощью которого
происходит это ускорение старения.
Единственным таким механизмом
представляется именно ускорение
укорочения теломер вызываемыми
АФК повреждениями в теломерной
ДНК
13. Сближение окислительной и теломерной теорий старения
В последние годы появились доказательства того, что окислительные повреждения
вызывают укорочение теломер, (Von Zglinicki et al, 2009, 2011). Т.о., можно думать, что
теломерная и окислительная теории старения объединились в единую теорию.
Это, однако, не совсем так:
Даже самые мощные антиоксиданты (SkQ В.П.Скулачева) не в состоянии остановить
старение, а могут лишь несколько замедлить его, и что существенно – именно и
исключительно за счет замедления укорочения теломер (Von Zglinicki et al, 2009) ,
причем не только в делящихся клетках, при репликации, но и в неделящихся.
Если бы совсем не было окислительных повреждений, теломеры все равно
укорачивались бы при каждом митозе, ибо так устроен механизм репликации ДНК
(Оловников, 1971), и старение, пусть медленнее, все равно прогрессировало бы, что
мы и наблюдаем при действии любых антиоксидантов. А вот если теломеры не
укорачиваются, что имеет место в трансформированных клетках, т.к. там
экспрессируется ТЕРТ, старение действительно останавливается, несмотря на
повреждающее действие активных форм кислорода, даже более интенсивное, чем в
здоровых клетках.
Т.о., правильно будет считать, что укорочение теломер – единственный механизм
старения, а кислородные повреждения могут старение ускорять, дополнительно
укорачивая теломеры, наряду с рядом других факторов – ионизирующей радиацией,
ошибками репарации, эпигенетическими явлениями и даже психологическим
стрессом. Но это – не механизмы старения. (Пример Наполеона).
Соответственно, активные формы кислорода могут ускорять старение, но только в
пределах той доли старения, которая вызвана окислительными повреждениями
именно теломер.
14. О принципиальной невозможности существенного увеличения видовой продолжительности жизни человека Старение - побочный, хотя и неизбежный
О принципиальной невозможностисущественного увеличения видовой
продолжительности жизни человека
Старение - побочный, хотя и неизбежный
результат самогő индивидуального
развития (онтогенеза). Чтобы замедлить его,
необходимо замедлить индивидуальное
развитие. Это невозможно (или очень
трудно), да и вряд ли нужно. Что же нужно и
возможно?
15. Можно ли замедлить старение?
• С помощью антиоксидантов, или инымиспособами можно в принципе, не увеличивая
видовую продолжительность жизни человека,
замедлить сам процесс старения, т.е.
одряхления (senescence). В результате
человек не будет жить дольше, но будет
дольше «оставаться молодым», т.е.
сохранять умственную и физическую
активность. Этой проблемой занимается
коллектив во главе с академиком
В.П.Скулачевым в МГУ.
16. Нестареющие организмы
Существует мнение, что некоторые виды животных не стареют –
именно те, которые растут на протяжении всей жизни. Это –
многие пресмыкающиеся, рыбы и некоторые другие виды.
В последние годы обнаружено, что во всех клетках этих животных
на протяжении всей жизни экспрессируется ТЕRТ. В результате
они, хотя и стареют (со временем экспрессия ТЕRТ ослабевает),
но очень медленно.
Очень велико своеобразие механизмов старения у насекомых и у
некоторых примитивных водных организмов – гидр, морских звезд
и т.п.
Единственный действительно нестареющий организм – это амеба.
Амебы могут безгранично размножаться как в культуре, так и в
природе, если для этого есть соответствующие условия. Но что
особенно интересно – амебы являются также единственной
группой организмов, не имеющих полового размножения. Они
размножаются только вегетативно – простым делением.
Это исключение – сочетание отсутствия старения с отсутствием
полового размножения - открывает важнейшую, на мой взгляд,
закономерность бытия:
Природа дала живым существам возможность бессмертия, но
не индивидуального (для человека – не личного), а в
поколениях.
17.
• Голый землекоп (лат. Heterocephalus glaber) —небольшой роющий грызун семейства землекоповых.
Вид отличается уникальными для млекопитающих
особенностями: сложной социальной организацией
колонии,холоднокровностью, иммунитетом к раку в
целом, нечувствительностью к некоторым формам
боли (термическим ожогам и химическим ожогам
кислотами), выносливостью к высоким
концентрациям CO2. Является самым долгоживущим
из грызунов (до 28 лет).
18.
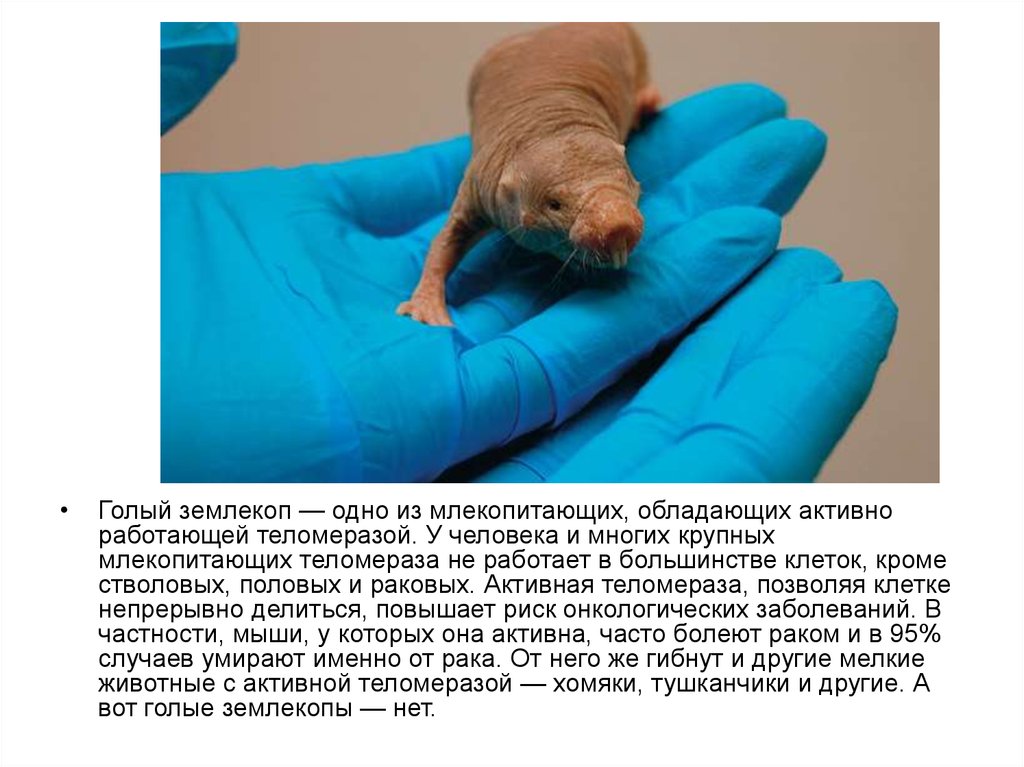
Голый землекоп — одно из млекопитающих, обладающих активно
работающей теломеразой. У человека и многих крупных
млекопитающих теломераза не работает в большинстве клеток, кроме
стволовых, половых и раковых. Активная теломераза, позволяя клетке
непрерывно делиться, повышает риск онкологических заболеваний. В
частности, мыши, у которых она активна, часто болеют раком и в 95%
случаев умирают именно от рака. От него же гибнут и другие мелкие
животные с активной теломеразой — хомяки, тушканчики и другие. А
вот голые землекопы — нет.
19.
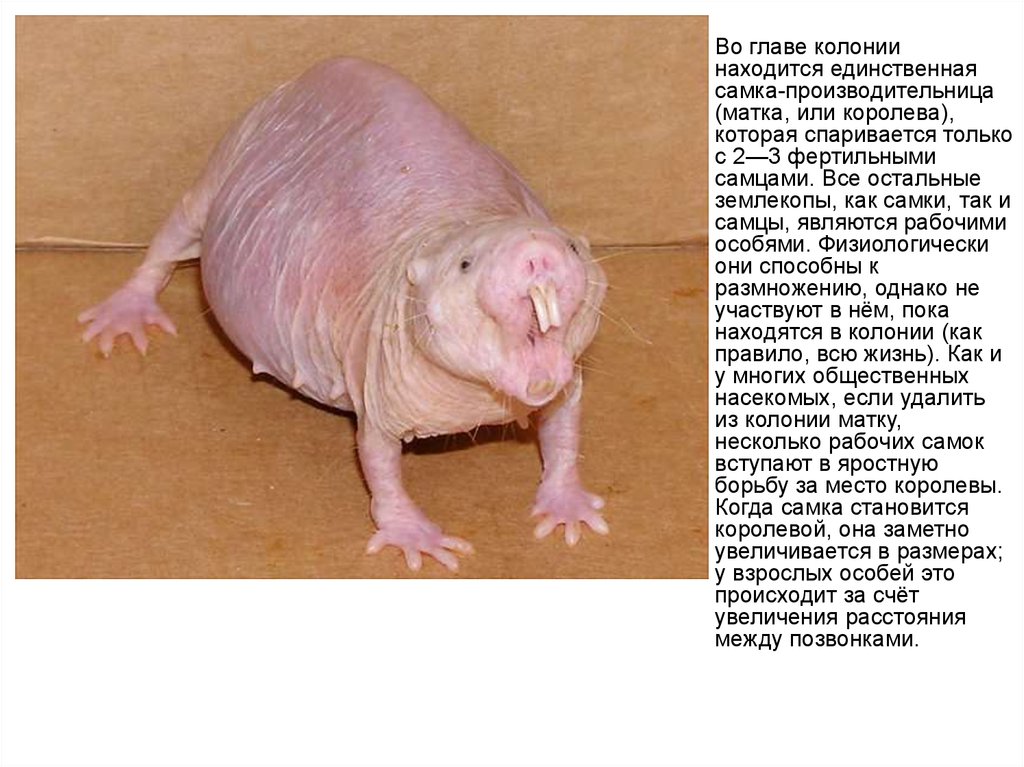
Во главе колонии
находится единственная
самка-производительница
(матка, или королева),
которая спаривается только
с 2—3 фертильными
самцами. Все остальные
землекопы, как самки, так и
самцы, являются рабочими
особями. Физиологически
они способны к
размножению, однако не
участвуют в нём, пока
находятся в колонии (как
правило, всю жизнь). Как и
у многих общественных
насекомых, если удалить
из колонии матку,
несколько рабочих самок
вступают в яростную
борьбу за место королевы.
Когда самка становится
королевой, она заметно
увеличивается в размерах;
у взрослых особей это
происходит за счёт
увеличения расстояния
между позвонками.
20.
21. Гиалуроновая кислота
У голых землекопов даже гиалуроновая кислота оказалась необычной.
Каждая молекула этого вещества у них была в пять раз больше, чем у
других грызунов и даже человека. Исследователи выяснили, что
ферменты, ответственные за разрушение сахаров в организме
землекопов, не очень активны, из-за чего эти вещества могут
накапливаться в межклеточном матриксе в очень крупных дозах. Такое
количество гиалуроновой кислоты эти грызуны приобрели в процессе
эволюции для того, чтобы их кожа стала достаточно эластичной и не
разрывалась, когда животное пытается пролезть по узкому проходу.
22. Tian, Azpurua, Hine, Vaidya, Myakishev-Rempel, Ablaeva, Mao, Nevo, Vera Gorbunova & Andrei Seluanov 2013.High-molecular-mass hyaluronan mediates the cancer resistance of the naked mole rat. Nature
Tian, Azpurua, Hine, Vaidya, Myakishev-Rempel, Ablaeva, Mao,Nevo, Vera Gorbunova & Andrei Seluanov 2013.High-molecularmass hyaluronan mediates the cancer resistance of the naked
mole rat. Nature
• Доказательство 510 кратного
превышения
размеров НА у
голого землекопа
23.
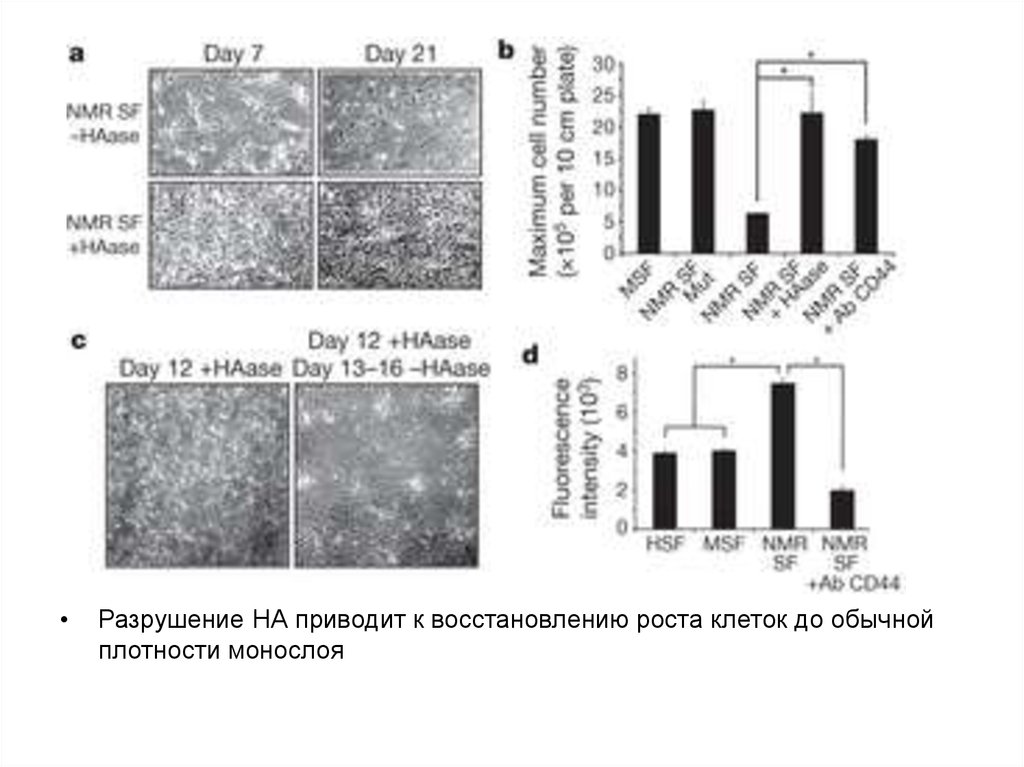
Разрушение НА приводит к восстановлению роста клеток до обычной
плотности монослоя
24.
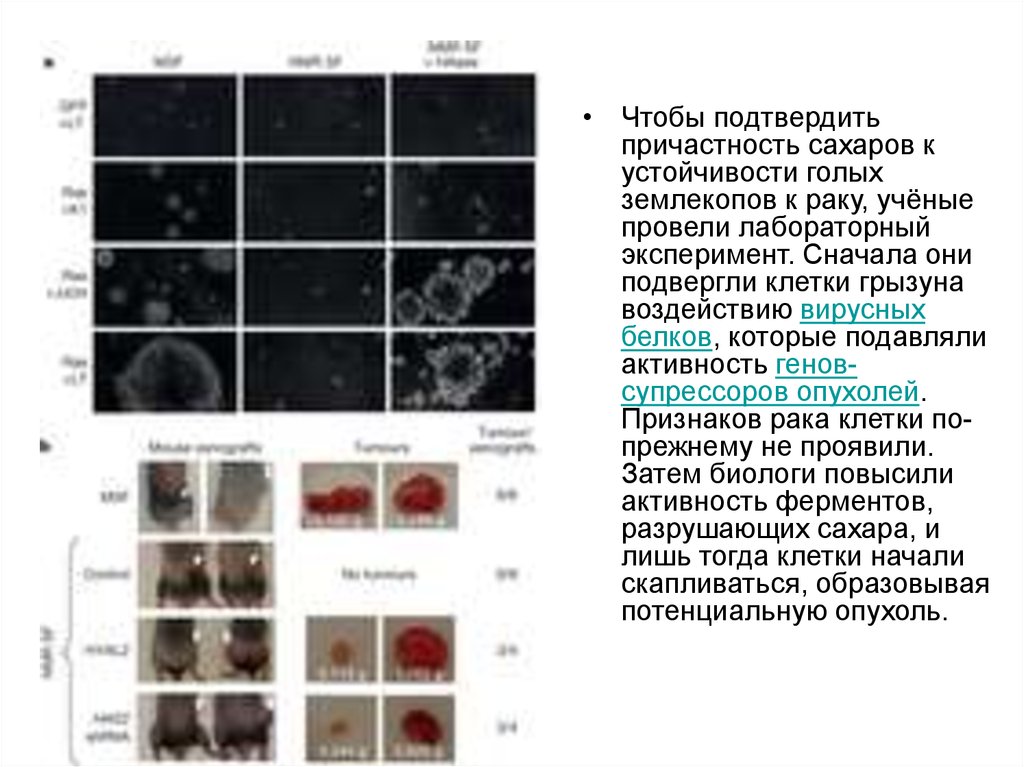
• Чтобы подтвердитьпричастность сахаров к
устойчивости голых
землекопов к раку, учёные
провели лабораторный
эксперимент. Сначала они
подвергли клетки грызуна
воздействию вирусных
белков, которые подавляли
активность геновсупрессоров опухолей.
Признаков рака клетки попрежнему не проявили.
Затем биологи повысили
активность ферментов,
разрушающих сахара, и
лишь тогда клетки начали
скапливаться, образовывая
потенциальную опухоль.
25. Снижение калорийности питания и его экспериментальные модели
26. Карликовые мыши Эймса
Одной из интересных моделей для изучения
старения являются карликовые мыши Эймса
(Mattison, 2000; Bartke et al., 2001). Они являются
гомозиготными аутосомально-рецессивными
мутантами, несущими единичные точечные мутации
в Prophet pit-1 гене. Карликовые мыши Эймса живут
на 50-65 % дольше (самцы и самки соответственно),
чем мыши "дикого" типа (Brown-Borg et al., 1996;
Brown-Borg, Rakoczy, 2000). Эта модель - один из
первых примеров способности единичного гена
значительно продлевать жизнь у млекопитающих.
Аутосомальные рецессивные мутации ведут к
гипопитуитаризму, проявляющемуся в снижении
синтеза и секреции гормона роста (ГР), пролактина и
тиреоидстимулирующего гормона (ТСГ). У этих
мышей снижен уровень инсулинподобного фактора
роста 1 (IGF-1) и инсулина в крови, повышена
чувствительность к инсулину и понижена
температура тела (Mattison, 2000; Bartke et al,, 2001).
Как самцы, так и самки карликовых мышей
бесплодны, у них выражена иммунодепрессия
(Mattison, 2000). Показано, что у этих мышей в печени
снижен уровень глутатиона и аскорбиновой кислоты и
увеличена активность каталазы по сравнению с
контролем, что проявляется большей устойчивостью
к окислительному стрессу (Brown-Borg et al., 1996;
Brown-Borg, Rakoczy, 2000; Mattison, 2000; Bartke,
Turyn, 2001). Частота возникновения спонтанных
опухолей у карликовых и нормальных мышей
одинакова (Mattison, 2000). Однако карликовые мыши
живут значительно дольше, чем нормальные, и
возможно, что опухоли развиваются у них позднее.
27.
Мыши, гомозиготные по мутации "little"
(lit/lit), имеют изолированный дефицит
гормона роста, обусловленный потерей
функции гена, кодирующего рецептор
гормона роста высвобождающего
гормона (GHRH). У этих мышей
замедлена скорость роста и несколько
уменьшены размеры тела. Однако в
отличие от карликовых мышей по
неизвестной причине они продолжают
медленно расти в зрелом возрасте, и в
целом различие в весе тела между ними
и их сиблингами дикого типа становятся
незначительными (Bartke et al., 2001).
Мыши lit/lit живут существенно дольше
нормальных мышей, если их содержать
на низкожировой диете для
предотвращения ожирения.
выключением гена рецептора
IGF-1 у мышей. Гомозиготы (Igf1/-) умирали при рождении.
Однако у самок гетерозиготных
мышей с частично
нокаутированным геном
рецептора IGF-1 (Igf1r+/-)
наблюдали увеличение средней
продолжительности жизни на 33
% по сравнению с самками
дикого типа (р<0.001), тогда как у
самцов - лишь на 16%. У этих
мышей не наблюдалось
карликовости; основной обмен,
температура тела, потребление
корма, физическая активность и
фертильность у них не
отличались от контроля. Также не
отличалась от контроля и частота
развития спонтанных опухолей. В
то же время Igf1r+/- мыши
отличались большей
устойчивостью к оксидативному
стрессу, чем контроль дикого
типа (Igf1r+/+) (Holzenberger et al.,
2003).
28. Р66shc
Предполагается, что p66shc вовлечен в феноптоз, то есть
запрограммированную гибель организма, обусловленную
массовым апоптозом в жизненно важных органах в
результате воздействия активных форм кислорода
(Skulachev, 2000). Полагают, что активные формы
кислорода вызывают окисление фосфотидил-серина
внутреннего слоя клеточной плазматической мембраны и
приводят к появлению этого фосфолипида на наружном
слое мембраны, что распознается специальным
рецептором и вызывает фосфорилирование серинового
остатка p66shc. Фосфорилированный по серину p66shc
блокирует митоз и запускает апоптоз. Массовый апоптоз
приводит к феноптозу и, следовательно, к сокращению
продолжительности жизни организма. Представляется
весьма важным то обстоятельство, что р53 вовлечен в
реализацию влияния АФК на p66shc и, напротив,
стимулирует продукцию АФК при реализации феноптоза
(Skulachev, 2002).
Направленная мутация гена p66shc у мышей привела к
развитию устойчивости к вызванному паракватом
окислительному стрессу, индуцирующему продукцию
анионов супероксида, и увеличению продолжительности
жизни животных на 30% (Migliaccio et al,, 1999). У p66shc-/мышей выявлено снижение системного и тканевого
окислительного стресса, атерогенеза и апоптоза в
эндотелии аорты (Napoli et al., 2003).
29. Рапамицин и метформин
Ингибирование TOR–сигналинга генетическим или фармакологическим
вмешательством удлиняет продолжительность жизни у
беспозвоночных, включая дрожжей, нематод и дрозофил. Однако было
неизвестно, может ли ингибирование сигнального пути mTOR
увеличить продолжительность жизни у млекопитающих. Авторы статьи
в Nature показали,
что рапамицин, ингибитор mTOR, удлиняет среднюю и максимальную
продолжительность жизни как у мужских, так и женских особей мышей
при начале кормления на 600 день жизни. Рапамицин привел к
увеличению продолжительности жизни на 14% для женских особей и
на 9% для мужских. Эффект наблюдался в трех независимых
экспериментах в разных лабораториях (Дэвид Харрисон, Лаборатория
Джексона, Рэнди Стронг, Университет Техаса, Ричард Миллер,
Университет Мичигана).
В независимом анализе рапамицин, скармливаемый мышам, начиная с
270 дня жизни, также увеличил выживаемость как мужских, так и
женских особей. Такие результаты дал промежуточный анализ,
проведенный в средней точке выживаемости. Рапамицин может
продлить жизнь путем отсрочки смерти от рака, замедления
механизмов старения,
30.
Программируемая клеточнаясмерть
31. Caenorhabditis elegans
Biochemical model for the activation of
programmed cell death. (A) In normal cell, not
undergoing apoptosis, CED-4 is bound to the
sufrace of mitochondria by its interaction with
CED-9. (B) THe apoptosis activator EGL-1 binds
to CED-9, which leads to the release of CED-4
and its successive binding to the pro-caspase
CED-3 (C). The procaspase pro CED-3
undergoes an autoproteolytic activation and
forms an active executioner caspase CED-3.
32. Пути апоптоза
Apoptotic pathways
are genetically very
conserved. It has
been shown, that Bid
and Bcl-2 are highly
homologous in
sequence and in
function to Egl-1 and
Ced-9. Bcl-2
therefore functions as
a regulator and
Bid/Bax proteins
function as regulators
of Bcl-2. The
mammalian caspase
9 and Apaf-1 also
show homology to
Ced-4 and Ced-3
in C. elegans. The
activation of
procaspase-9 is
similar to the
activation of Ced-3,
but there is also a
need of Cytochrome
C involvement.
33. Mitochondrial theory of aging
34. В.П.Скулачев
35. Работы В.П.Скулачёва
SkQ1 – это эффективный антиоксидант. Его задача – даление из митохондрий избытка
АФК, возникающего в процессе старения. На сердечных митохондриях in vitro было
показано, что SkQ1 может восстанавливаться дыхательной цепью, то есть служить
возобновляемым антиоксидантом многократного действия. Его свойства изучались под
руководством академика РАН Владимира Скулачева с участием сотрудников Института
физико-химической биологии и ряда факультетов МГУ, а также более чем 30
исследовательских институтов в России, Швеции, США и на Украине.
Результаты показали, что SkQ1способен тормозить апоптоз клеток HeLa и фибробластов
человека, вызванный Н2О2, увеличивать медианную продолжительность жизни у грибов
Podospora anserina, рачков Ceriodaphnia affinis, дрозофил, а у мышей – до двух раз.
На мышах и крысах было показано, что SkQ1 замедляет развитие таких признаков
старения, как инволюция тимуса и фолликулярных клеток селезенки, уменьшение
соотношения лимфоцитов и нейтрофилов в крови, остеопороз, катаракта, ретинопатия,
глаукома, облысение, поседение, исчезновение эстральных циклов у самок и полового
влечения у самцов, гипотермия и т. д.
Исследователи пришли к выводу, что этот антитиоксидант снижает смертность на ранних
и средних этапах старения и предотвращает появление большой группы старческих
дефектов
36. Концепция Михаила Батина Фонд «Наука против старения»
1. Многим подопытным животным удалось существенно продлить жизнь.
Абсолютный рекорд (1998 – 2003 гг.) в продлении жизни мышей принадлежит
Анджею Бартке из Школы медицины университета Южного Иллинойса. Его
мышь прожила 1819 дней – вдвое больше обычной лабораторной мыши. В
пересчете на возраст человека – это около 180 - 200 лет. У подопытных
животных был выключен ген рецептора гормона роста. По мнению ученых
мышь прожила так долго в первую очередь из-за того, что у неё был меньший
уровень инсулина и глюкозы. Также было показано, что долгоживущие
карликовые мыши Эймса сохраняют высокий уровнь когнитивных функций
(обучение и память) в пожилом возрасте.
...
В январе 2008 года Валтер Лонго с коллегами из университета Южной
Калифорнии на основе дрожжевого грибка создали генетическимодифицированный штамм, способный жить в 10 раз дольше.
В 2008 году Роберт Шмуклер Райс с коллегами из Арканзасского медицинского
университета (Литтл-Рок, США) продлили жизнь червя нематоды в десять раз.
Исходя из общности биологических законов, мы можем говорить о возможности
существенного продления жизни человека.
37.
2.В природе существуют животные с пренебрежимым старением.
У таких организмов смертность с возрастом не растёт, а иногда даже падает: жемчужница,
морской ёж, гренландский кит, некоторые черепахи. Принято называть этих животных
нестареющими. В основном они умирают по физическим причинам или от голода.
Одним из самых интересных млекопитающих в этом списке является голый землекоп небольшой грызун, обитающий в некоторых странах Африки.
В апреле 2002 года недалеко от местечка Мтито Анджел в Кении умер самец голого
землекопа, впервые пойманный в 1974 году в возрасте примерно 1 года. Это означает, что
он прожил более 28 лет. Такая продолжительность жизни является рекордом среди
грызунов! Любой представитель отряда грызунов, обладающий такой же массой тела, живет
в среднем не более 3х-4х лет, то есть в 7-8 раз меньше.
Но, помимо того, что голые землекопы живут колоссально долго, они обладают еще одной
крайне необычной особенностью. Дело в том, что ученые не обнаружили у представителей
вида онкологических заболеваний. В 2009 году исследователи из Университета Рочестера
(США) Вера Горбунова и Андрей Селуянов обнаружили у голых землекопов ген, который
делает беспрепятственное деление клеток в их организме невозможным. Благодаря гену
p16, клеточное деление останавливается тогда, когда клеток может стать больше, чем
нужно для нормальной жизнедеятельности животного. Эффект гена р16 настолько
значителен, что даже когда экспериментаторы пытались вызвать у голых землекопов
мутагенез, они добивались лишь некоторого изменения клеточного роста, в то время как у
обычных мышей от такого воздействия развивался рак.
38.
3. Учёным удалось обратить назад развитие клетки человеческого
организма, увеличив потенциал генома.
Это дает возможность, в том числе, выращивать новые собственные органы и
ткани человека. Большое число ученых, среди которых Такаси Цудзи, Макото
Асашимо, Лари Беновиц, Элизабед Джонс, Николас Дейл, Пол Шарп, Роберт
Лангер, Лаура Никласон, Энтони Атала, Робет Ланза, Паоло Маккиарини, Шинья
Яманака, своими достижениями в области регенеративной медицины и
развитии клеточных технологий фактически произвели революции в медицине.
К примеру, Паоло Маккиарини из Университетской клиники Барселоны удалось
вырастить новую трахею пациентке больной, страдающей от последствий
туберкулеза. В результате чего она была избавлена от пересадки легких.
4. Несмотря на достижения в области продления жизни, внимание
общества, финансирование исследований находятся на крайне низком
уровне.
То есть, даже при таком положении вещей наука добивается выдающихся
результатов. Очевидно, что они будут гораздо выше, если этой области будет
уделяться больше внимания, не говоря уже о том – если она станет
приоритетной для государств и общества.
39.
5. Аргументация против продления жизни сводится в основном к
нецелесообразности увеличения срока жизни человека и
зачастую носит абсурдный характер.
Так, практически всегда люди, столкнувшись с идей радикального
продления жизни, начинают рассуждать о перенаселении. При этом
оппоненты, не понимают, что прежде всего отказывают себе в
существовании, ни в коей мере не решая проблему ограниченности
ресурсов на земле. Или представляют себе продление жизни в виде
лежащего больного, не осознавая, что цель науки – максимально
увеличить потенциал жизнеспособности человека.
Я привожу эти аргументы, чтобы усилить интерес в обществе к идее
радикального продления жизни. Я рассказываю, как сейчас обстоят
дела с этой задачей у себя блоге: m-batin.livejournal.com. Буду рад
новым френдам. Считаю, что если блог, рассказывающий о том, как
радикально продлить жизнь человека, будет одним из самых
популярных в ЖЖ, это будет означать, что данная задача включена в
общественную повестку дня. Соответственно шансы на
финансирование отрасли возрастают. Спасибо за внимание. Михаил
Батин.
40.
• Итак, продлить жизнь сверх положенногочеловеку лимита скорее всего невозможно, да и
не нужно.
У природы нет плохой погоды,
Ход времен нельзя остановить.
Осень жизни, как и осень года,
Надо, не скорбя, благословить.
Эльдар Рязанов
«Служебный роман»