

















































")


























































 biology
biologySimilar presentations:










Фазы и этапы фотосинтеза
1. ФАЗЫ И ЭТАПЫ
ФОТОСИНТЕЗА2.
Фотосинтез - это сложныймногоступенчатый
окислительновосстановительный процесс.
Он состоит из двух типов
реакций.
Одни
из
них
происходят только на свету, а
для осуществления других
свет не требуется. Поэтому в
фотосинтезе
принято
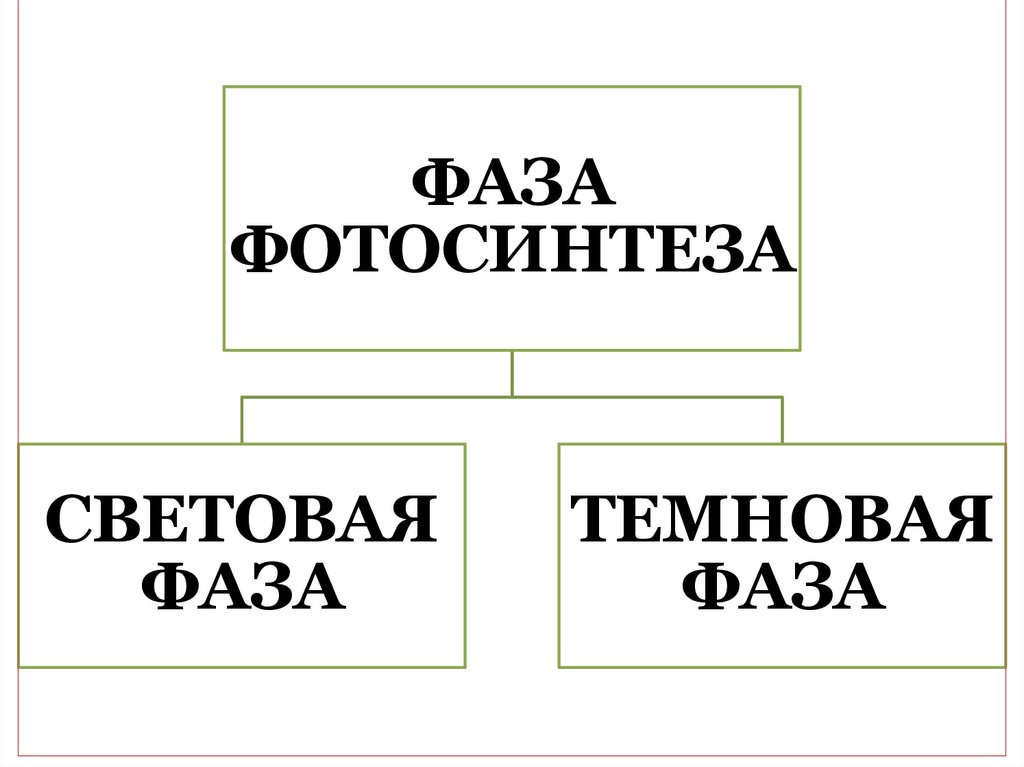
выделять две фазы:
3.
ФАЗАФОТОСИНТЕЗА
СВЕТОВАЯ
ФАЗА
ТЕМНОВАЯ
ФАЗА
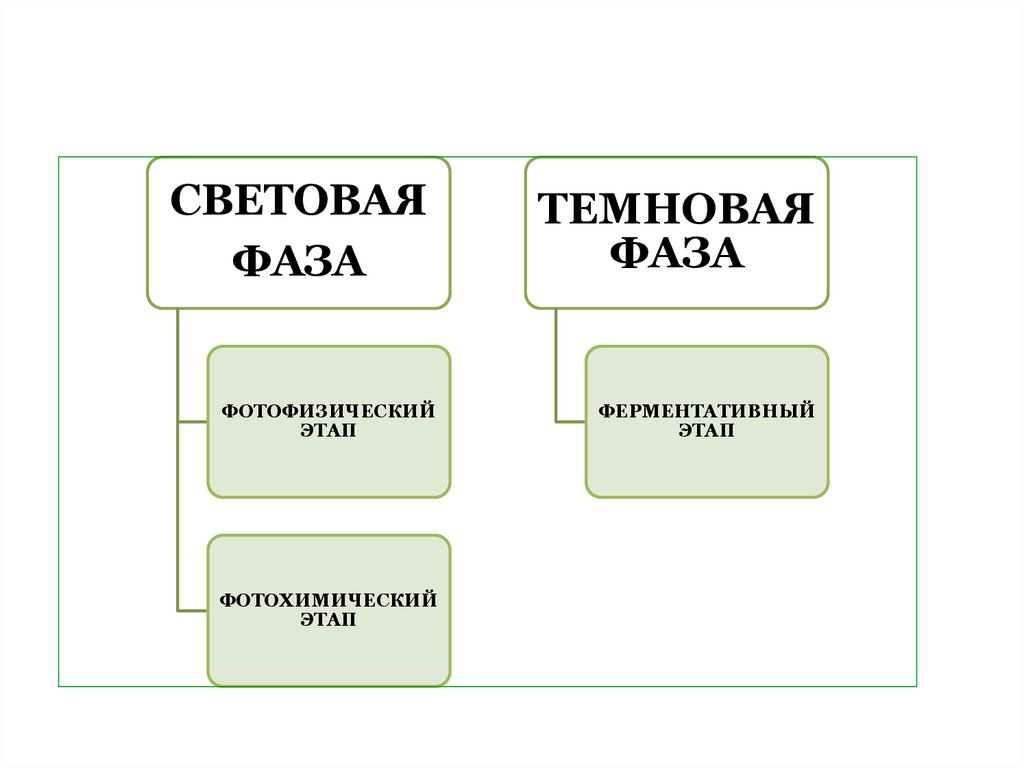
4. В зависимости от осуществляемых в ходе фотосинтеза процессов, в нем выделяют три этапа:
ЭТАПЫФОТОСИНТЕЗА
ФОТОФИЗИЧЕСКИЙ
ФОТОХИМИЧЕСКИЙ
ФЕРЕМЕНТАТИВНЫЙ
5.
СВЕТОВАЯФАЗА
ТЕМНОВАЯ
ФАЗА
ФОТОФИЗИЧЕСКИЙ
ЭТАП
ФЕРМЕНТАТИВНЫЙ
ЭТАП
ФОТОХИМИЧЕСКИЙ
ЭТАП
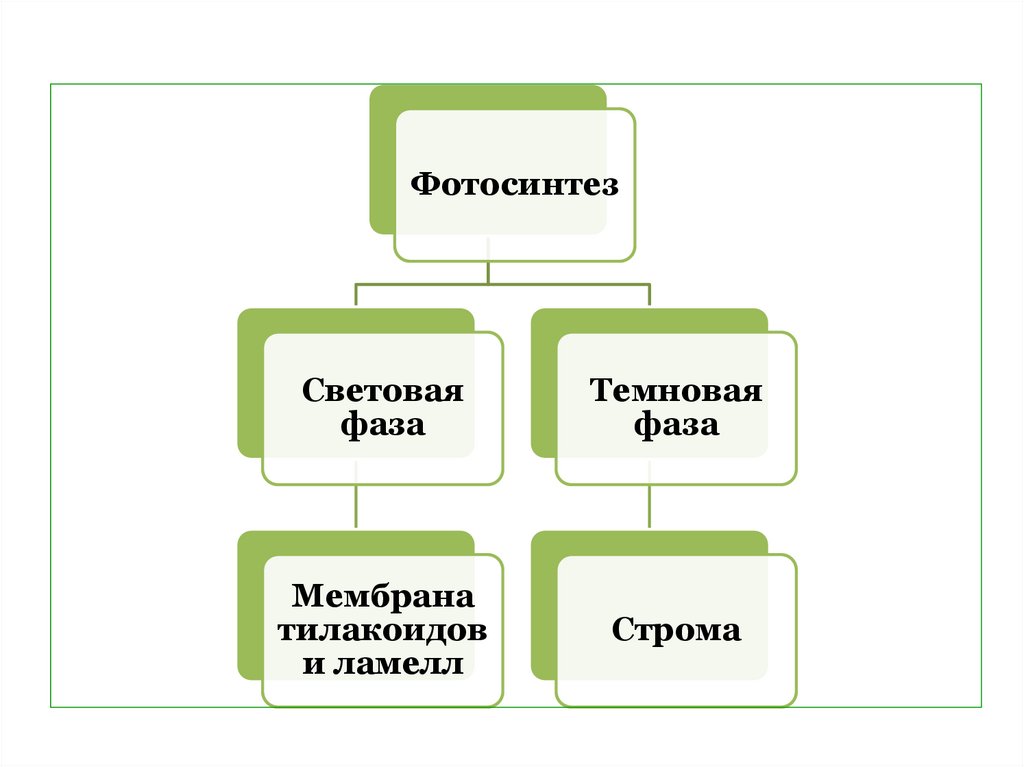
6.
ФотосинтезСветовая
фаза
Темновая
фаза
Мембрана
тилакоидов
и ламелл
Строма
7. СВЕТОВАЯ ФАЗА
ФОТОСИНТЕЗА8. ФОТОФИЗИЧЕСКИЙ ЭТАП
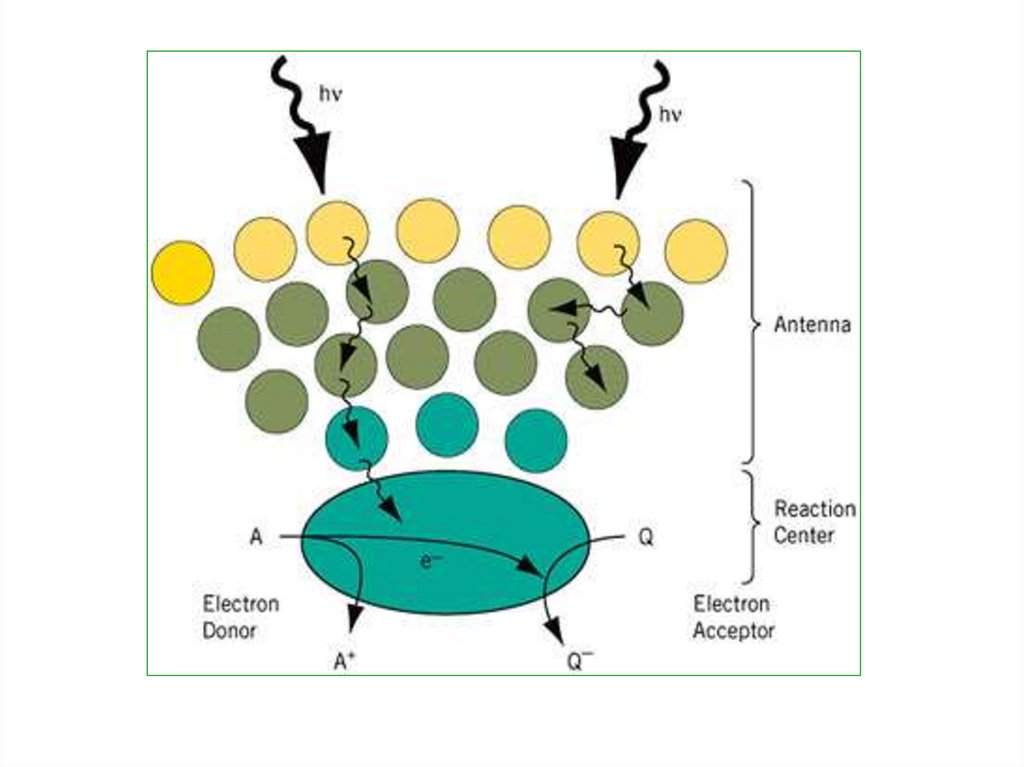
Сущностьфотофизического
этапа заключается в том, что
кванты света поглощаются
пигментами-антенами ССК и
переводят их молекулы в
возбужденное состояние.
9.
Затем эта энергия резонанснымпутем
передается
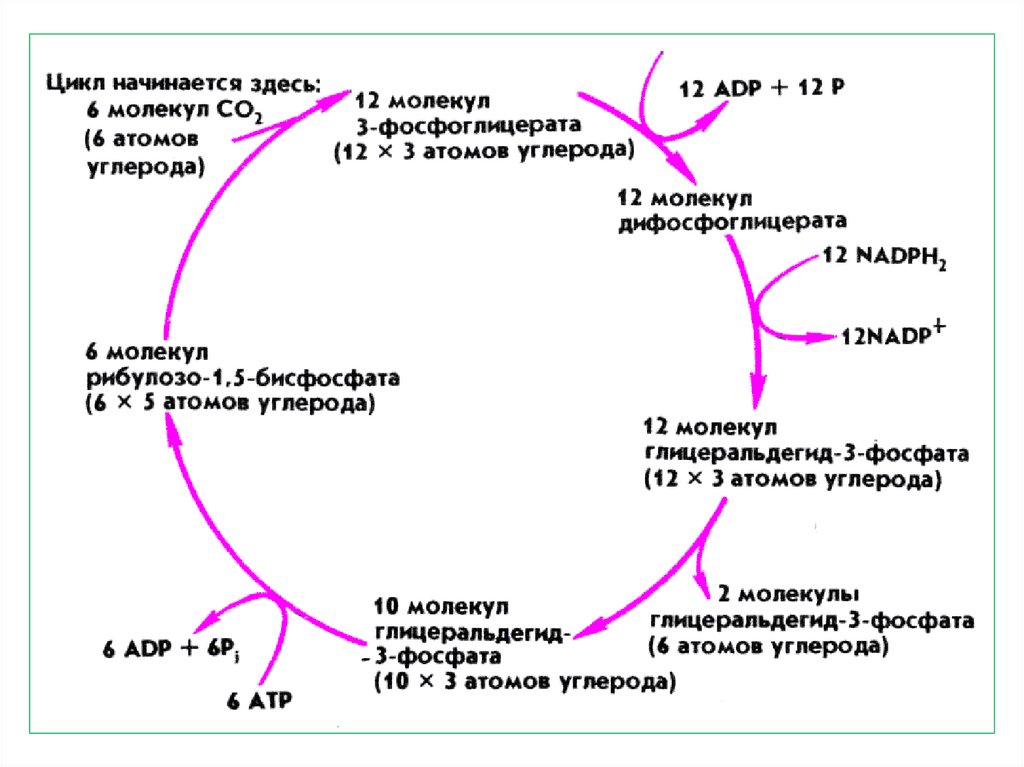
через
ряд
пигментов ССК фотосистемы на
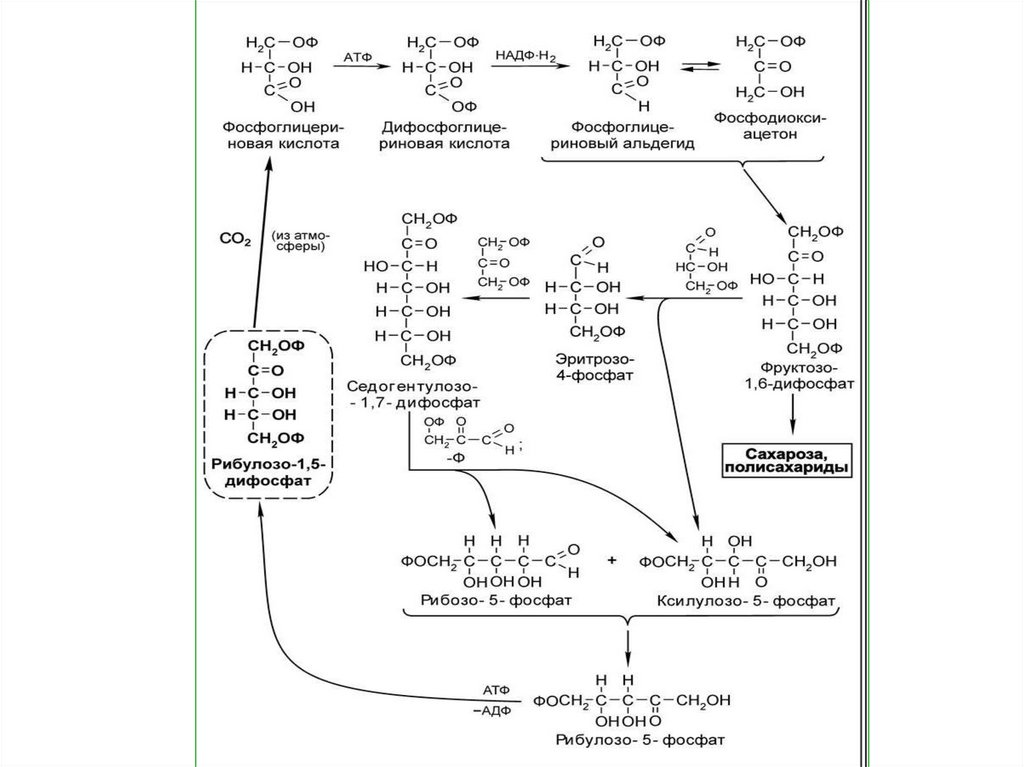
хлорофилл-ловушку, входящую в
реакционный
центр
фотосистемы.
Хлорофиллловушка
затем
принимает
непосредственное
участие
в
фотохимическом процессе.
10.
11.
12.
При поглощении квантасвета молекулой какоголибо вещества электрон в
такой молекуле переходит
на
более
высокий
энергетический уровень.
13. Поглощение света подчиняется следующим законам:
• 1. Каждый квант света активируеттолько одну молекулу.
• 2. Вся энергия кванта поглощается
одним электроном, который при
этом поднимается на более высокий
энергетический уровень.
• 3. Только поглощенный свет может
производить химическое действие.
14.
Электрон,кроме
того,
что
находится
на
определенной
орбитали и вращается вокруг
ядра, обладает еще и спином
(вектором магнитного момента),
характеристикой, которую можно
трактовать
как
направление
вращения электрона вокруг своей
оси.
Спин
электрона
может
принимать два значения.
15.
Когдав
молекуле
все
электроны
расположены
попарно, их суммарный
спин равен 0. Это основное
синглетное состояние (S0).
16.
Припоглощении
света
электроны
переходят
на
орбитали с более высоким
уровнем энергии, и при этом
имеются
две
возможности:
если электроны не меняют
спина, то это приводит к
возникновению первого или
второго
синглетного
состояния (S1*, S2*);
17.
если же один из электроновменяет
спин, то такое
состояние
называют
триплетным (Т1*).
18.
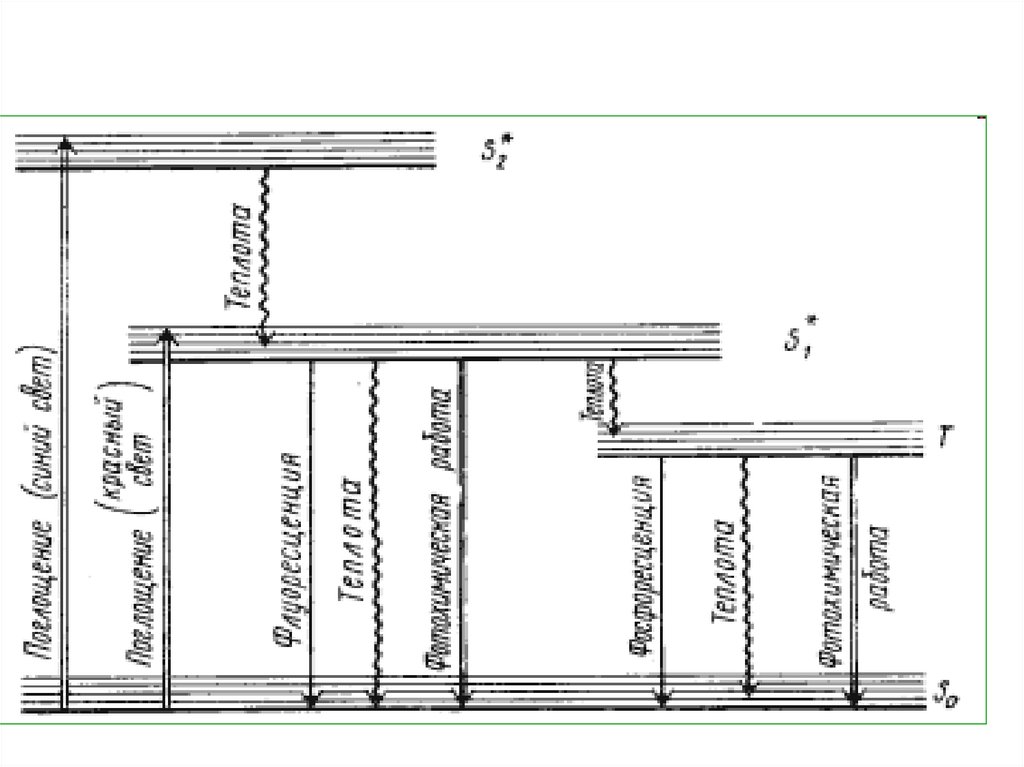
Наиболее высокий уровеньэнергии в S2*. На него
электрон переходит под
влиянием
синефиолетовых лучей. В S1*
электроны
переходят,
поглощая более мелкие
кванты красного света.
19.
Времяжизни
на
S2*
чрезвычайно мало. Затем
электрон переходит из S2*
в
S1*.
Такой
переход
сопровождается
потерей
части энергии в виде
тепла. Время жизни в S1*
больше.
20.
Наибольшее время жизниэлектрона
в
Т1*.
Из
возбужденного S1* или Т1*
состояния
молекула
хлорофилла
может
переходить
в
основное
состояние. При этом теряется
часть энергии.
21.
22. Потеря энергии может происходить:
• 1) путем выделения энергии в видесвета
(флуоресценция
и
фосфоресценция) или в виде тепла;
• 2) путем переноса электрона на
другую молекулу пигмента;
• 3) путем затрачивания энергии на
фотохимические процессы (потеря
электрона и присоединение его к
акцептору, образование АТФ и
НАДФ . Н + Н+).
23.
В любом из указанных случаевмолекула пигмента переходит
на основной энергетический
уровень. Энергия, испускаемая
в виде флуоресценции или
тепла, в дальнейшем клеткой
не может быть использована.
24. ФОТОХИМИЧЕСКИЙ ЭТАП
Фотохимические реакции - этореакции, в которых энергия света
преобразуется
в
энергию
химических
связей
АТФ.
Одновременно
под
действием
света
происходит
разложение
воды с выделением кислорода и
образование
восстановленного
НАДФ.
25.
Сущность этих реакций выясненав 1954-58 гг. Арноном. Синтез АТФ
при фотосинтезе сопровождается
потоком
электронов:
нециклическим
или
циклическим.
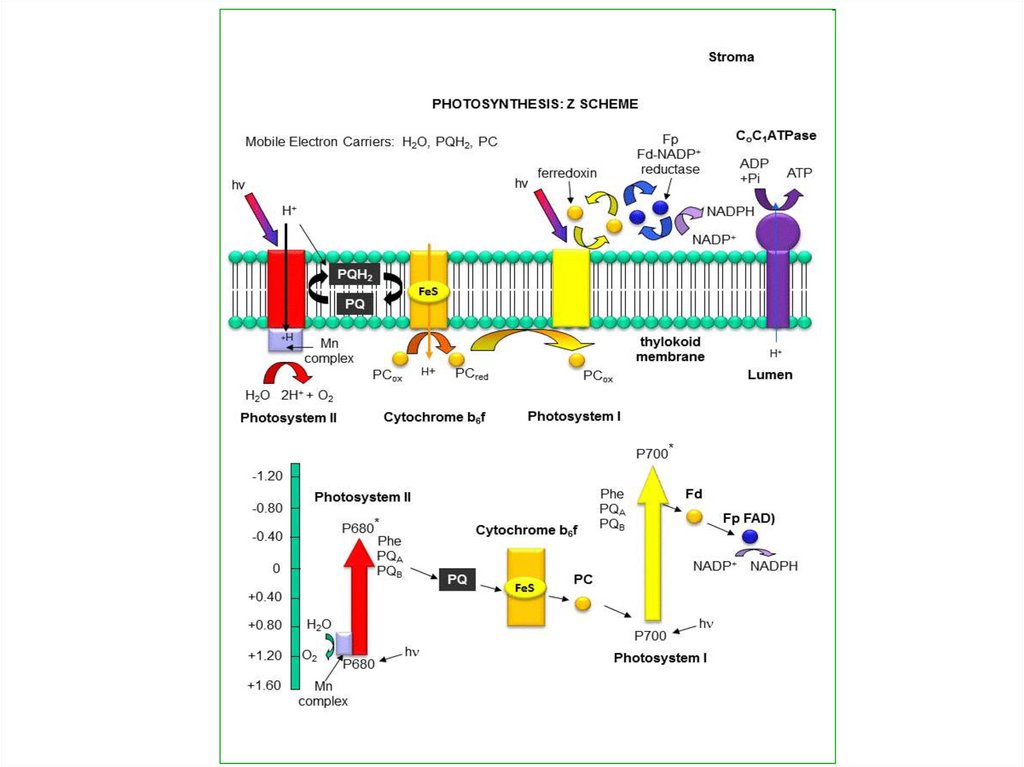
26. НЕЦИКЛИЧЕСКОЕ ФОСФОРИЛИРОВАНИЕ
Вреакциях
нециклического
фосфорилирования принимают
участие две фотосистемы.
Обе фотосистемы работают
одновременно,
улавливая
кванты света.
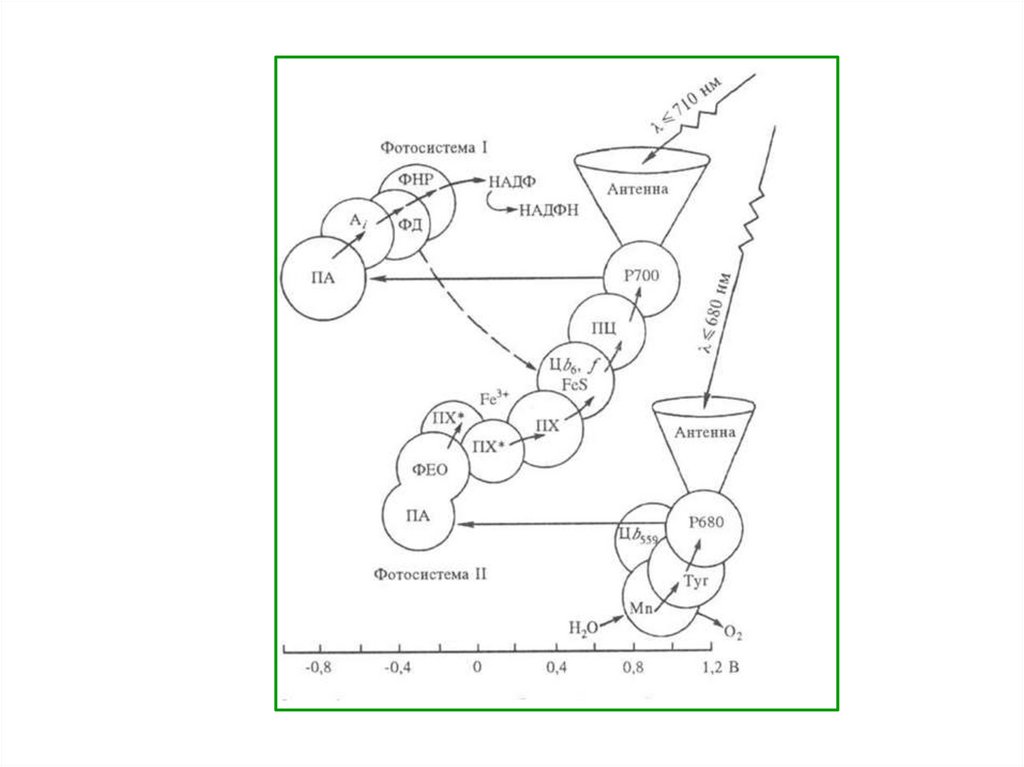
27. НЕЦИКЛИЧЕСКОЕ ФОСФОРИЛИРОВАНИЕ
Энергиякванта
света,
улавливаемая
в
ходе
фотофизического
этапа
пигментами-антенами,
передается хлорофиллу-ловушке
Р700,
расположенному
в
реакционном центре ФС I.
28.
В результате этого электронв молекуле Р700 переходит на
более
высокий
энергетический
уровень
(S1*).
В
возбужденном
состоянии Р700 легко отдает
электрон,
при
этом
окисляясь.
29.
Электронпринимает
неидентифицированный
переносчик и передает его
на железосерные белки.
Затем электрон поступает
на
белок
ферредоксин,
содержащий
два
атома
железа, и через различные
флавопротеины
переносится на НАДФ.
30.
Когдадва
электрона
присоединяются на НАДФ,
из
водной
среды,
окружающей тилакоидные
мембраны,
к
нему
присоединяются свободные
протоны. Образуется НАДФ
·
Н
+
Н+,
который
используется в темновой
фазе.
31.
Таким образом, Р700 окислен,у
него
образовалась
электронная
дырка.
Он
получает электрон для ее
заполнения от хлорофилла
Р680,
входящего
в
реакционный центр ФС II.
32.
Под влиянием кванта светавозбужденный
электрон
молекулы Р680 передается в
ЭТЦ на молекулу феофитина,
затем
на
пластихиноны,
переносящие как электроны,
так
и
протоны,
отсюда
электроны
передаются
на
железосерный белок, а затем
на
цитохром
f
(группа
цитохромов с).
33.
Следующим переносчикомслужит
медьсодержащий
белок
пластоцианин
(содержит два атома меди).
От пластоцианина электрон
заполняет
электронную
дырку
у
Р700.
Перенос
электрона по ЭТЦ от ФС II к
ФС
I
сопровождается
синтезом АТФ.
34.
Теперь электронная дыркаобразовалась
в
Р680.
Он
заполняет
ее
за
счет
присоединения электрона от
молекулы
воды.
Отрыв
электрона
от
воды
происходит под действием
света.
35.
Молекула воды распадается,образуя
протоны
и
кислород.
Реакция
фотоокисления
воды
называется фотолизом.
hn
2 Н2О
4 е + 4 Н+ + О2
36. Механизм фотолиза заключается в следующих превращениях молекулы воды:
• Н2 ОН+
+ ОН-
• ОН-
ОН +
е
• 4 ОН
2 Н2О + О2
• 4 Н2О
• 2 Н2 О
4 Н+ + 4 е + 2 Н2О + О2
4 Н+ + 4 е + О 2
37.
Дляфотоокисления
воды
необходимы
в
качестве
катализаторов
ионы
марганца,
хлора
и
кальция.
38.
Процессфосфорилирования
АДФ,
сопряженный
с
нециклическим транспортом
электронов по ЭТЦ, называют
сопряженным
мембранным
нециклическим
фотосинтетическим
фосфорилированием.
39.
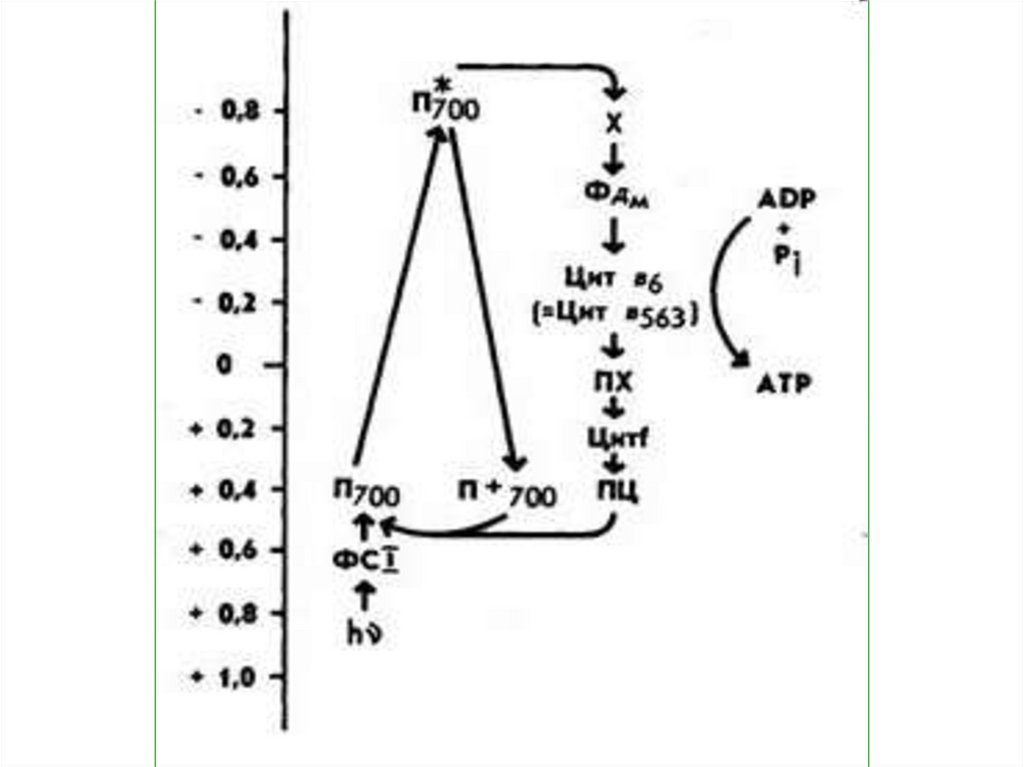
40. ЦИКЛИЧЕСКОЕ ФОСФОРИЛИРОВАНИЕ
В циклическомфосфорилировании принимает
участие только ФС I. Начинается
оно также, как и нециклическое
фосфорилирование, но от
ферредоксина электрон через ряд
промежуточных переносчиков
возвращается к хлорофиллу Р700.
41.
От ферредоксина электронпередается к цитохром b6,
затем
переходит
на
пластихион.
От
пластихинона
электрон
поступает к цитохрому f, от
цитохрома f передается на
пластоцианин, а уже от
пластоцианина - молекуле
хлорофилла Р700.
42.
• В то время, как электронтранспортируется по этому
циклическому пути, его
энергия используется на
присоединение фосфата к
АДФ с образованием АТФ.
43.
Такое фосфорилированиеназывается сопряженным
мембранным
циклическим
фотосинтетическим
фосфорилированием.
44.
Так как в циклическомфосфорилировании
отсутствует
фотолиз,
а
поэтому
не
образуется
кислород, то теоретически
оно может идти и при
закрытых устьицах.
45.
46.
47.
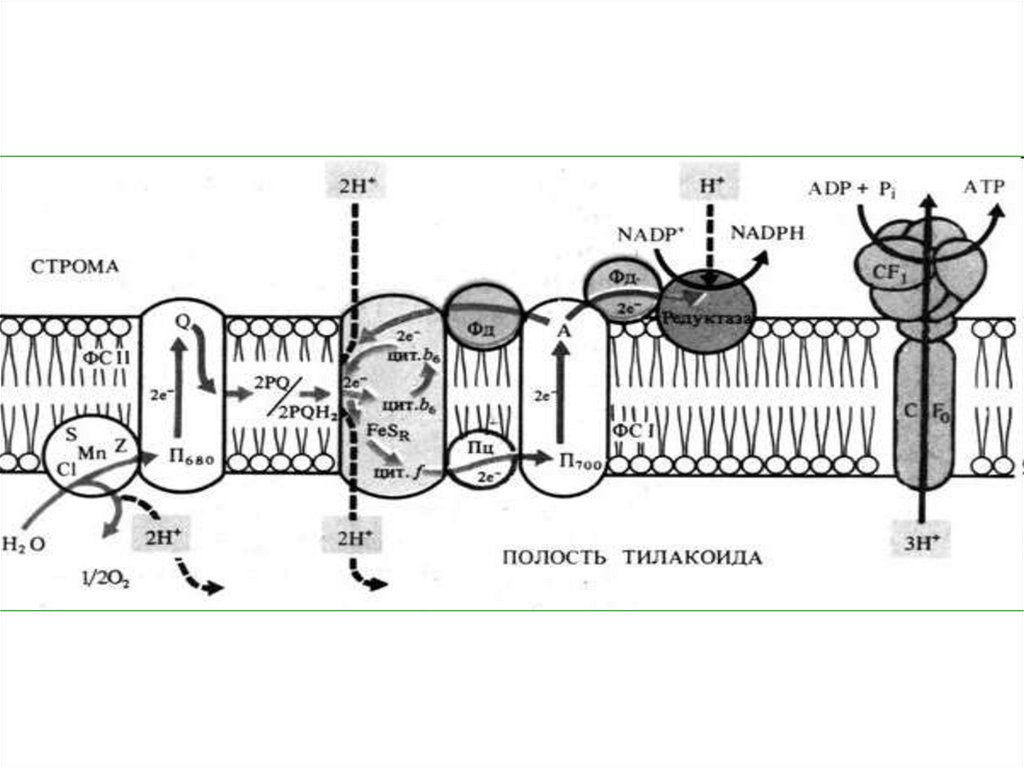
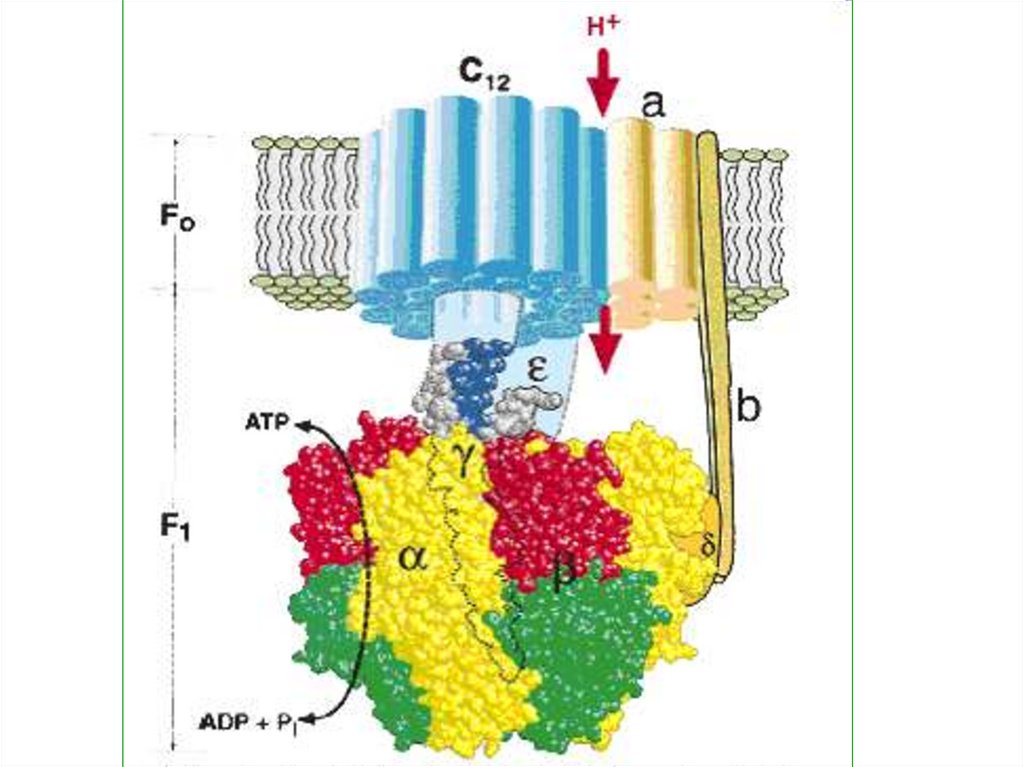
48. СИНТЕЗ АТФ
СинтезАТФ
во
время
фотосинтеза происходит за
счет химиоосмоса. Ионы
водорода на внутренней
стороне
мембраны
тилакоидов
образуют
протонный резервуар.
49.
Вмембране
находятся
сложные
ферментные
комплексы АТФ-синтетазы,
состоящие из двух частиц
(факторов) – F0 и F1. F0частица
пронизывает
мембрану насквозь, образуя
в ней канал.
50.
На наружной сторонемембраны с ней соединена
сферическая F1-частица.
51.
Водородныеионы
поступают
в
протонный
резервуар
за
счет
однонаправленного
переноса их на внутреннюю
сторону
тилакоидной
мембраны пластихинонами
из
стромы
и
за
счет
фотолиза воды.
52.
По обе стороны мембранывозникает
разность
электрохимических
потенциалов.
Она
разрешается
за
счет
пассивного
прохождения
ионов
водорода
из
протонного
резервуара
через канал АТФ-синтетазы
в строму хлоропласта.
53.
Энергияпередвижения
этих
ионов
идет
на
фосфорилрование, то есть
синтез АТФ из АДФ и
фосфата.
54.
55.
• На каждые 2 электрона,передаваемые
по
ЭТЦ,
внутри
тилакоида
накапливается
4
иона
водорода.
• На
каждые
3
иона
водорода, возвращающихся
в строму, синтезируется 1
молекула АТФ.
56.
Оченьмногие
авторы
начинают
описание
нециклического
фосфорилирования
с
работы
второй
фотосистемы.
57.
Поэтомув
заданиях
на
последовательность
процессов
световой фазы следует давать в таком
виде:
1) возбуждение
хлорофила
и
перемещение электронов;
2)
фотолиз
воды
и
образование
электронов;
3) синтез молекул АТФ;
4) соединение водорода с НАДФ и
образование восстановленного НАДФ.
58. ТЕМНОВАЯ ФАЗА
ФОТОСИНТЕЗА59.
Темновая фаза фотосинтеза этокомплекс
ферментативных реакций, в
течение которых происходит
восстановление
поглощенного
растением
углекислого газа за счет
продуктов световой фазы АТФ и НАДФ ∙ Н + Н+.
60.
Осуществляетсятемновая
фаза в строме хлоропласта.
Совокупность
темновых
реакций получила название
цикла Кальвина по имени
его открывателя Мелвина
Кальвина.
61. Мелвин Кальвин (1911-1997)
АмериканскийБиохимик.
Нобелевская
премия по химии
1961 г.
за исследование
усвоения
двуокиси
углерода
растениями
62.
Темновыереакции
осуществляются в три
основные стадии.
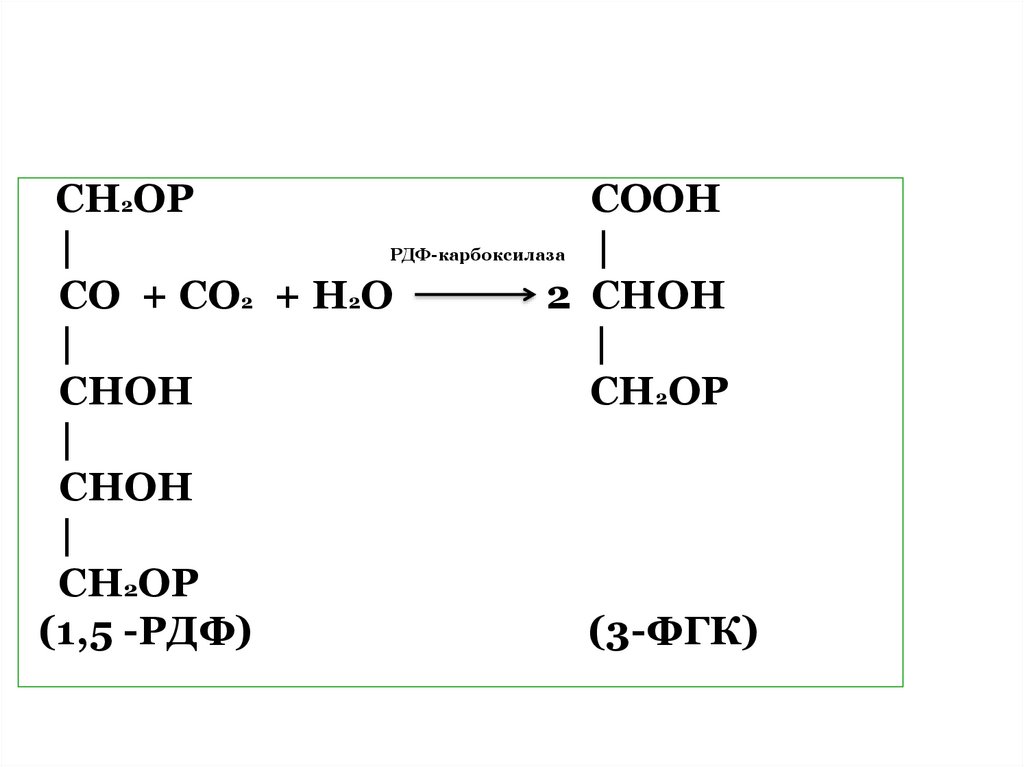
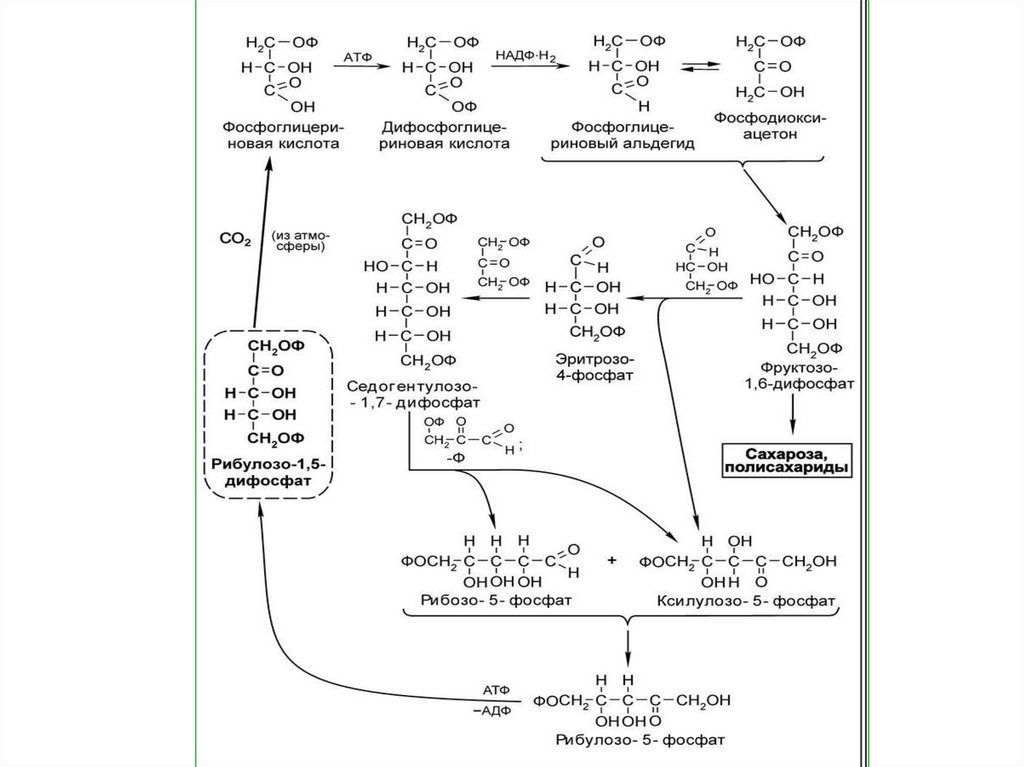
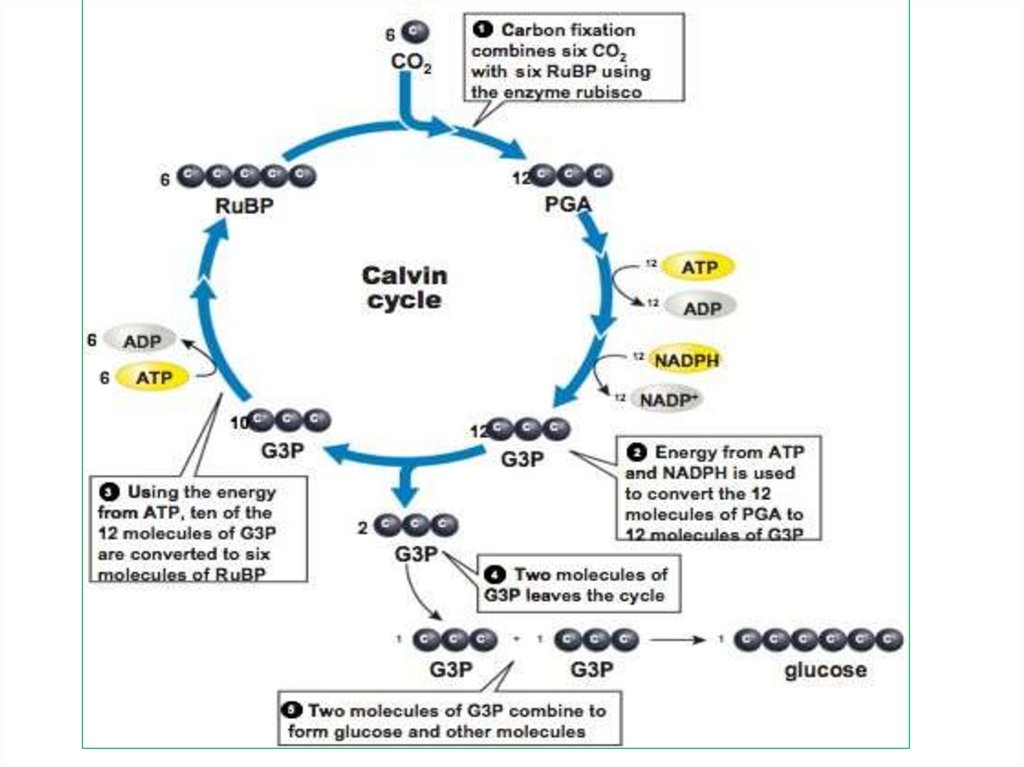
63. 1. Карбоксилирование
Поддействием
фермента
рибулозодифосфаткарбоксила
зы
(РДФ-карбоксилазы)
происходит связывание СО2 с
имеющимся
в
строме
пятиуглеродым соединением
1,5
рибулозодифосфатом
(рибулозобифосфатом (РДФ).
64.
СН2ОРСООН
|
РДФ-карбоксилаза
|
СО + СО2 + Н2О
2 СНОН
|
|
СНОН
СН2ОР
|
СНОН
|
СН2ОР
(1,5 -РДФ)
(3-ФГК)
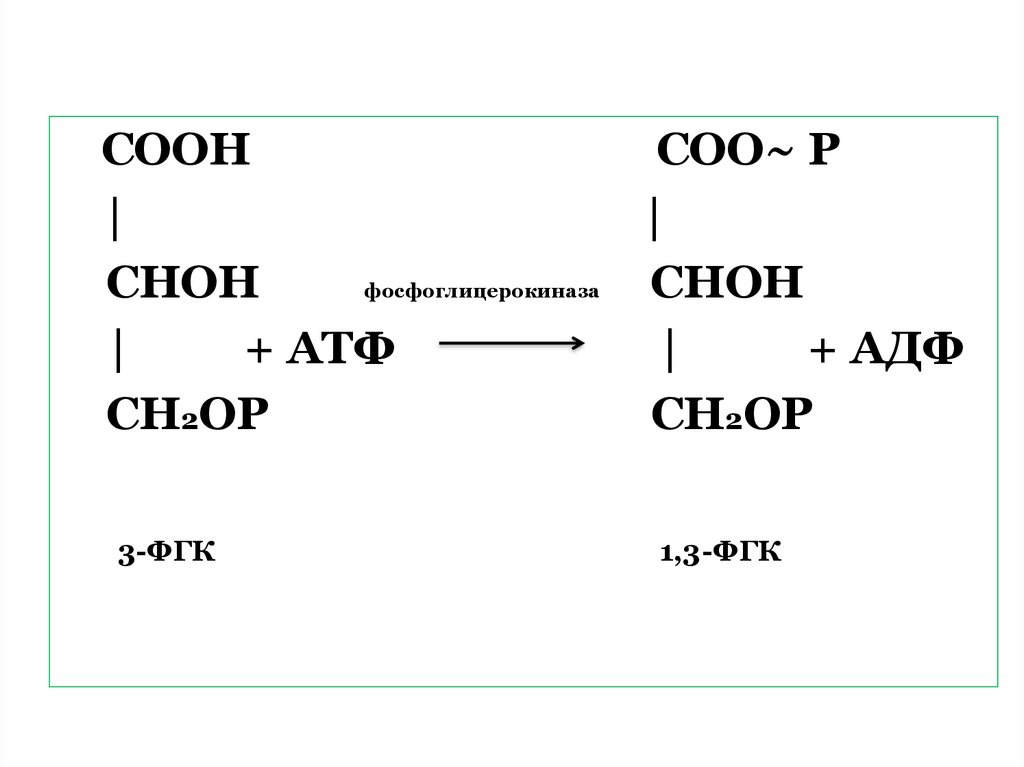
65. 2. Восстановление ФГК
Требуетучастия
продуктов
световой фазы фотосинтеза: АТФ
и НАДФ ∙ Н + Н+.
Сначала
происходит
реакция
фосфорилирования
3-ФГК
с
образованием
1,3-диФГК
с
участием
фермента
фосфоглицерокиназы.
66.
СООНСОО~ Р
|
|
СНОН
фосфоглицерокиназа
СНОН
|
+ АТФ
|
+ АДФ
СН2ОР
СН2ОР
3-ФГК
1,3-ФГК
67.
Затем1,3
ди-ФГК
восстанавливается за счет
НАДФ ∙ Н + Н+ и образуется
фосфоглицероальдегид (ФГА,
триозофосфат - ТФ).
68.
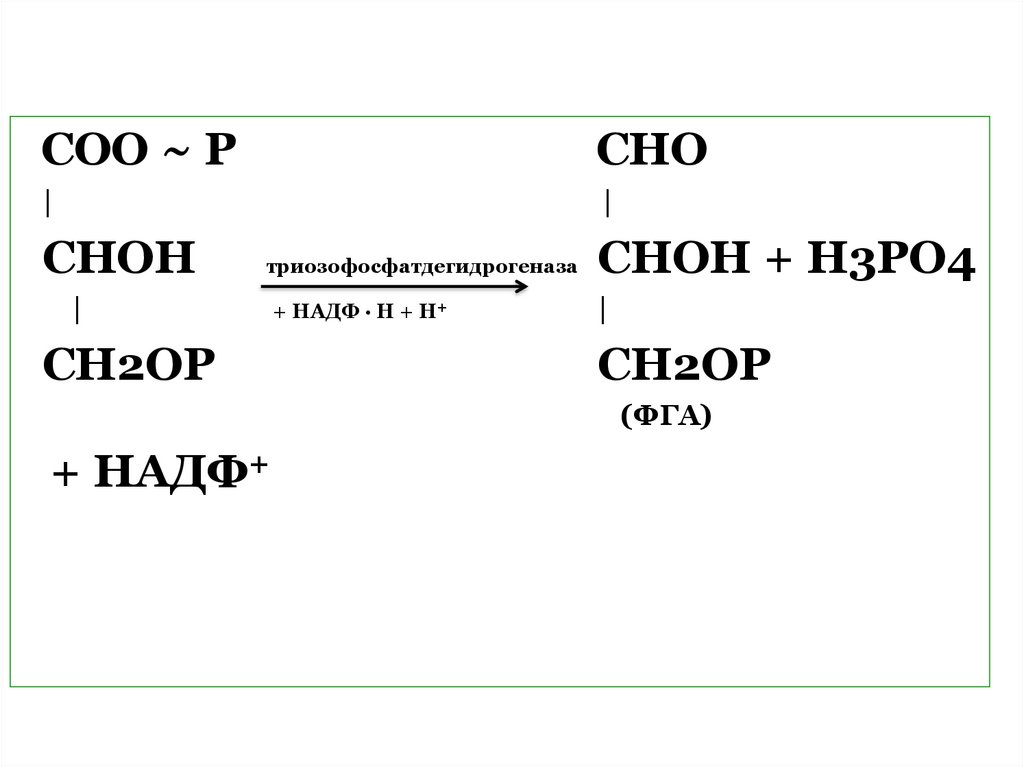
СОО ~ РСНО
|
|
CНОН
триозофосфатдегидрогеназа
СНОН + Н3РО4
+ НАДФ ∙ Н + Н+
|
|
СН2ОР
СН2ОР
(ФГА)
+ НАДФ+
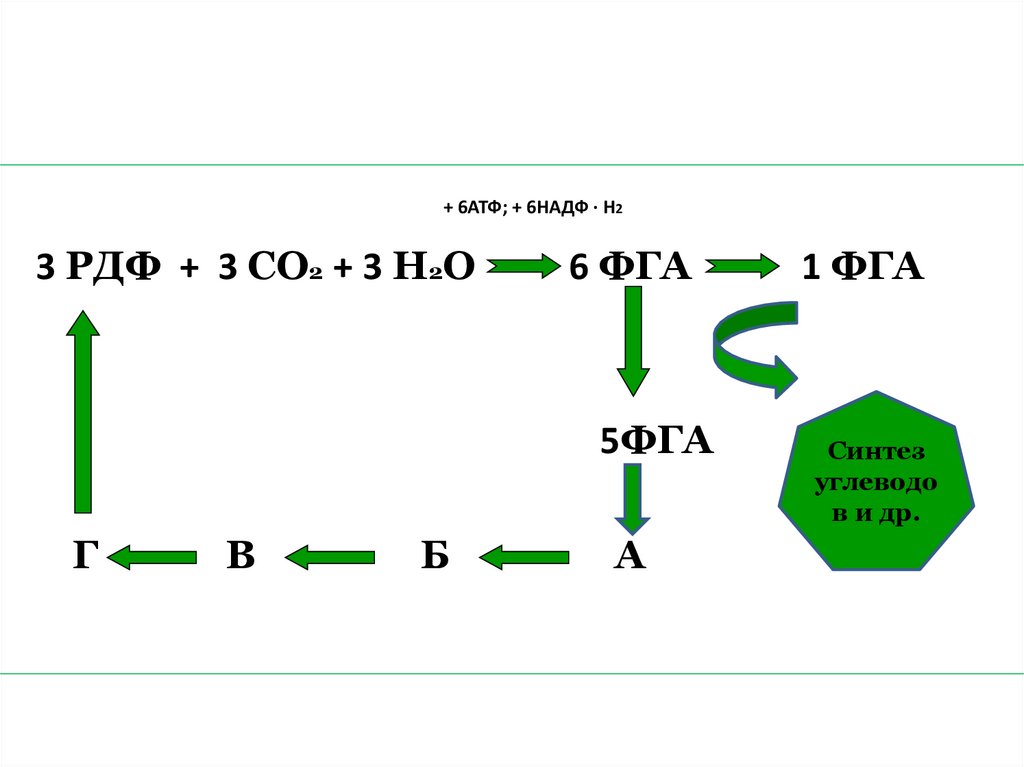
69. 3. Регенерация акцептора.
• Из каждых шести молекул ФГА 5молекул
используется
на
регенерацию
акцептора,
в
результате чего образуется 3
молекулы РДФ. Этот процесс идет
через образование 4-, 5-, 6-, 7углеродных соединений. Одна
молекула ФГА выходит из цикла и
идет на синтез углеводов и других
органических веществ.
70.
+ 6АТФ; + 6НАДФ ∙ Н23 РДФ + 3 СО2 + 3 Н2О
6 ФГА
5ФГА
Г
В
Б
А
1 ФГА
Синтез
углеводо
в и др.
71.
72.
При прохождении двух такихциклов
из
12
молекул
образовавшегося
ФГА
две
молекулы выходят, образуя
одну молекулу фруктозо-6фосфата, которая переходит
во фруктозу, а затем в глюкозу
или другие гексозы.
73.
Всепромежуточные
продукты цикла Кальвина
являются
фосфорилированными
соединениями.
В
таком
состоянии
они
наиболее
реакционноспособны.
74. Суммарное уравнение темновых реакций:
6 СО2 +12 НАДФ ∙ Н +12 Н++18 АТФ +11 Н2Офруктозо-6-Р + 12 НАДФ + 18 АДФ +17Фн
75.
76.
77.
78.
Цикл Кальвина называютС3-циклом, т.к. первый
продукт цикла является
С3-соединением.
79.
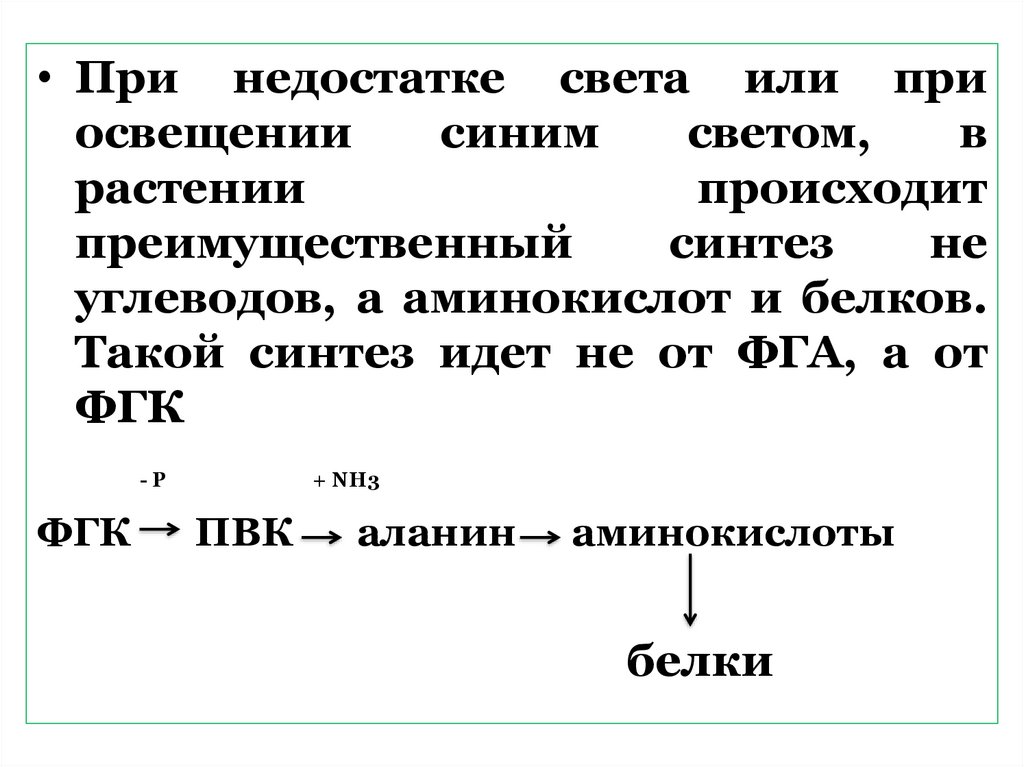
• При недостатке света или приосвещении
синим
светом,
в
растении
происходит
преимущественный
синтез
не
углеводов, а аминокислот и белков.
Такой синтез идет не от ФГА, а от
ФГК
-Р
ФГК
+ NН3
ПВК
аланин
аминокислоты
белки
80.
Под влиянием синего светаувеличивается проницаемость
мембран хлоропластов для
ФГК, и она выходит из
хлоропласта в цитоплазму и
используется там для синтеза
аминокислот.
81.
Под влиянием красного светаувеличивается
синтез
крахмала, так как в этом
случае
проницаемость
мембран хлоропласта для ФГК
низкая,
она
в
основном
остается в хлоропластах и
идет на синтез крахмала.
82.
БЕЛКИКРАХМАЛ
83. ЦИКЛ ХЭТЧА-СЛЭКА
• Цикл Кальвина - основной, но неединственный путь восстановления СО2.
Австралийские ученые Маршалл Хэтч и
Карл Роджер Слэк, а также российский
ученый Юрий Соломонович Карпилов
установили, что у некоторых растений,
главным
образом
тропических
и
субтропических, в том числе у кукурузы,
сахарного тростника, сорго, просо и др.
фотосинтез идет по несколько иному пути.
84.
85.
Маршалл Хэтч (р. 1932)Карл Роджер Слэк
(1937 – 2016)
86.
Юрий Соломонович Карпилов(1931 – 1937)
Советский фитофизиолог,
доктор
биологических
наук,
профессор,
первооткрыватель
С4пути.
87.
Вэтом
случае
первым
продуктом карбоксилирования
является
четырехуглеродное
соединение
ЩУК.
Такие
растения стали называть С4растениями, в отличие от С3растений,
у
которых
первым
продуктом является трехуглеродная
ФГК.
88.
Акцептором углекислого газау
С4-растений
является
трехуглеродный
фосфоенолпируват
(ФЕП).
СО2 при участии фермента
ФЕП-карбоксилазы
фиксируется
этим
соединением с образованием
ШУК.
89.
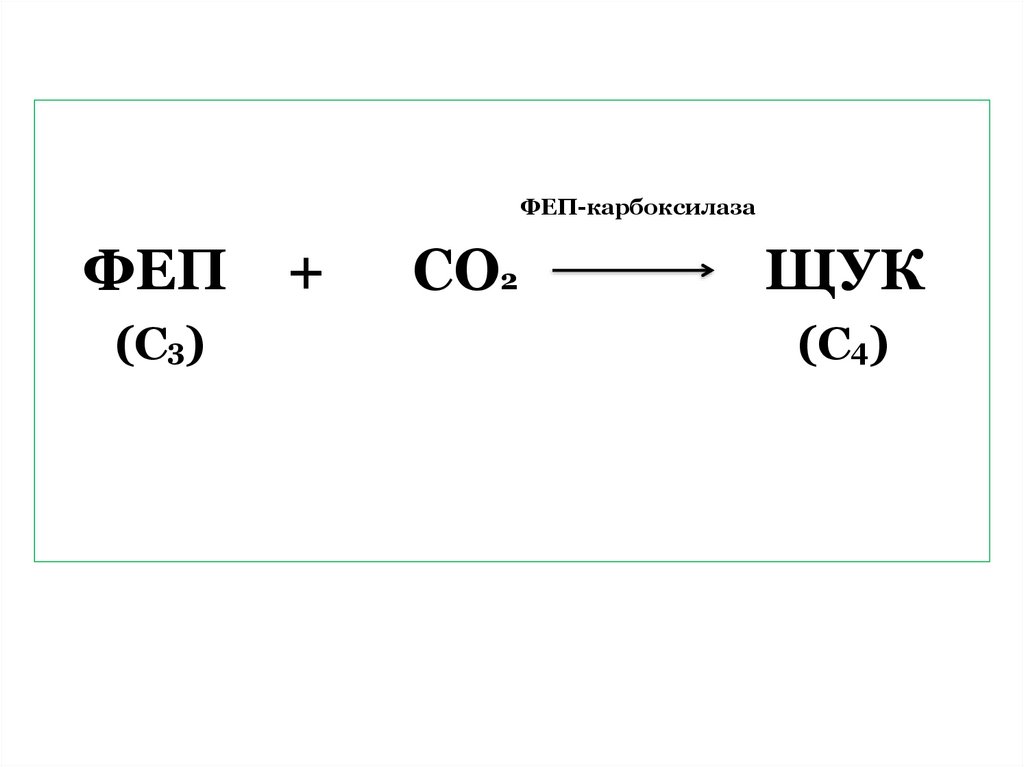
ФЕП-карбоксилазаФЕП
(С3)
+
СО2
ЩУК
(С4)
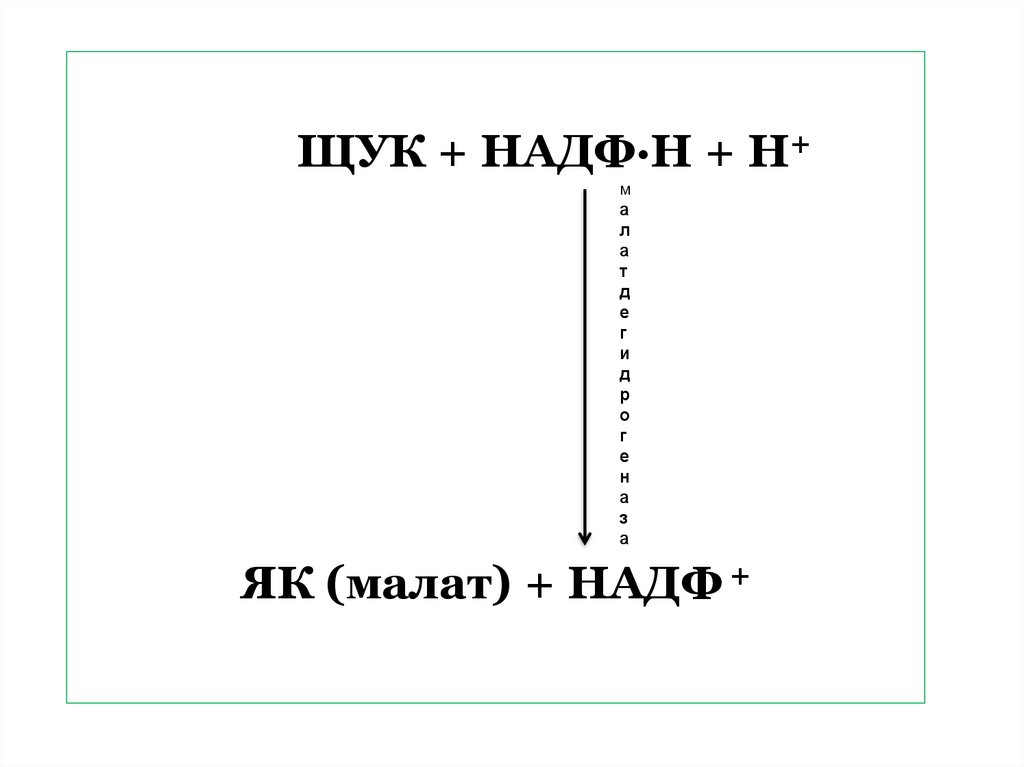
90.
Затем ЩУК под действиемфермента
малатдегидрогеназы
при
участии НАДФ ∙ Н + Н+,
продукта световой фазы,
восстанавливается
до
яблочной кислоты (малата).
91.
ЩУК + НАДФ∙Н + Н+м
а
л
а
т
д
е
г
и
д
р
о
г
е
н
а
з
а
ЯК (малат) + НАДФ +
92.
Черезплазмодесмы
в
клеточных
стенках
ЯК
переходит
в
хлоропласты
клеток обкладки проводящих
пучков. Там она используется
для образования СО2 и ПВК
путем декарбоксилирования.
При этом также образуется
НАДФ ∙ Н + Н+.
93.
Затем СО2уже повторно
фиксируется
РДФкарбоксилазой в обычном С3пути, где также используется и
НАДФ ∙ Н + Н+ , а ПВК
возвращается
в
клетки
мезофилла,
где
фосфорилируется с участием
АТФ, превращаясь в ФЕП и
цикл замыкается.
94. А- мезофилл листа Б- обкладка проводящего пучка
95.
Унекоторых
растений
образовавшаяся
ЩУК
в
процессе восстановительного
аминирования
с
участием
аспартаттрансферазы
превращается
в
аспарагиновую
кислоту,
и
декарбоксилированию
в
клетках обкладки проводящих
пучков
подвергается
аспарагиновая кислота.
96.
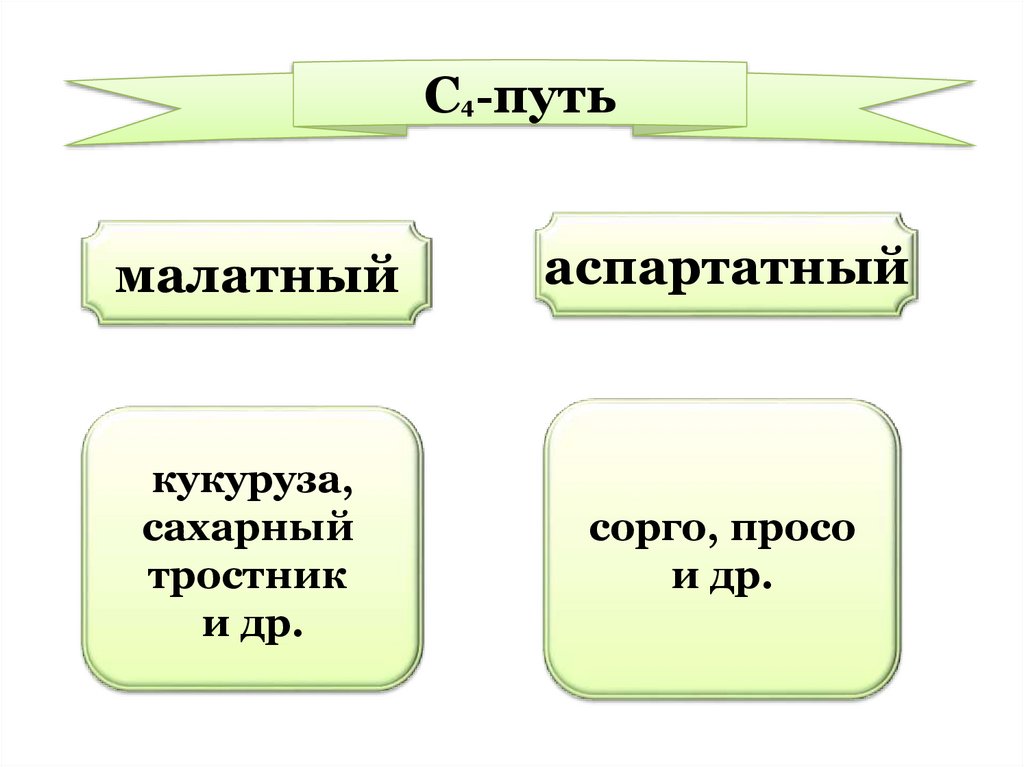
В зависимости от того,какая
органическая
кислота: ЯК (малат) или
аспарагиновая (аспартат) декарбоксилируется,
С4растения делятся на два
типа:
97.
C4-путьмалатный
аспартатный
кукуруза,
сахарный
тростник
и др.
сорго, просо
и др.
98.
У С4-растений хлоропластыв
клетках
мезофилла
мелкие гранальные, а в
клетках
обкладки
проводящих
пучков
пластиды крупные, часто
лишенные гран.
99.
В клетках мезофилла попреимуществу
идет
нециклическое
фосфорилирование
и
образуется НАДФ ∙ Н + Н+,
необходимый для цикла
Кальвина.
100.
В хлоропластах клеток обкладкиидет
только
циклическое
фосфорилирование.
Такое
разделение
типов
фосфорилирования связано с тем,
что
к
хлоропластам
клеток
обкладки,
расположенным
в
глубине
листа,
проникает
преимущественно
длинноволновый свет, который не
поглощается ФС II.
101.
Основнаяфункция
С4цикла - концентрация СО2
для С3-пути. Ночью у С4растений
происходит
образование органических
кислот,
а
днем
они
поступают
в
обкладку
проводящих пучков, где
происходит цикл Кальвина.
102.
Такимобразом,
фотосинтез может идти
днем
при
закрытых
устьицах,
что
предохраняет растение
от потери воды.
103.
ФЕП-карбоксилазафиксирует
СО2
очень
активно,
поэтому
концентрация
СО2
в
клетках обкладки намного
выше, чем в окружающей
среде.
104.
Это очень важно, т.к. С4растения обитают в условияхповышенных
температур,
когда
растворимость
СО2
значительно снижена.
105. ФОТОДЫХАНИЕ
106.
Фиксация углерода по С4путиимеет
еще
ряд
преимуществ. Растения С3пути
характеризуются
высокой
интенсивностью
процесса,
получившего
название фотодыхание.
107.
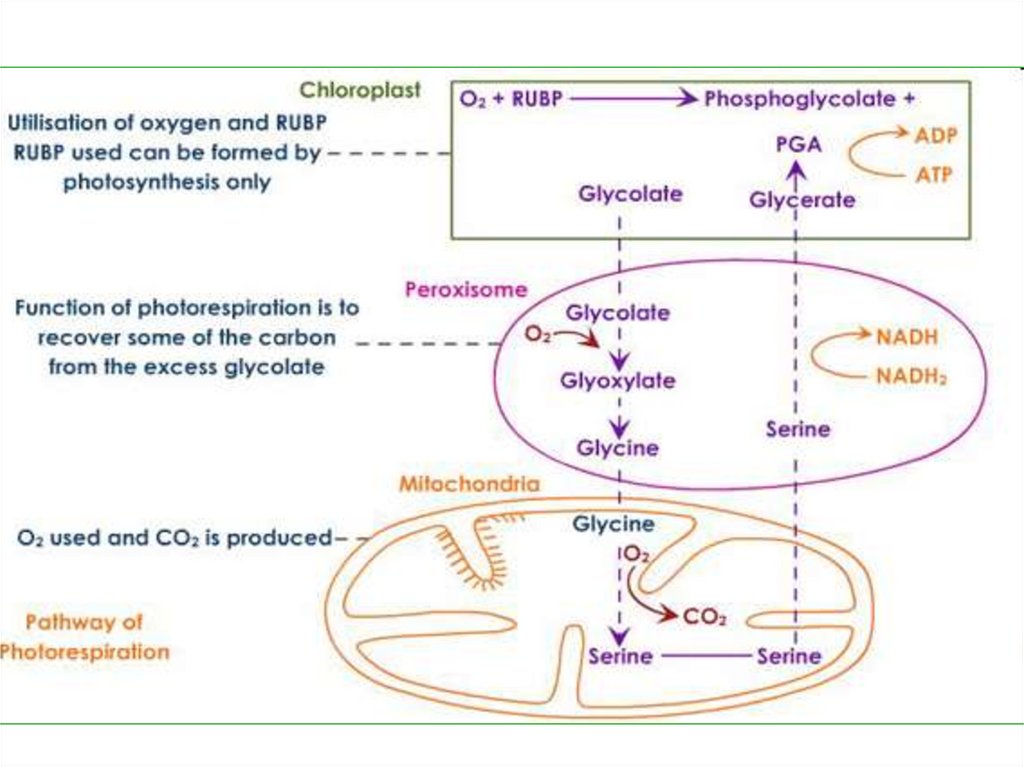
Фотодыхание - поглощениекислорода
и
выделение
углекислого газа на свету с
использованием
промежуточных
продуктов
цикла
Кальвина.
Этот
процесс никак не связан с
клеточным дыханием.
108.
ФерментРДФ-карбоксилаза
может реагировать не только с
СО2, но и с кислородом. При
взаимодействии
РДФ
с
кислородом образуется одна
молекула
трехуглеродной
фосфоглицериновой кислоты и
одна молекула двухуглеродной
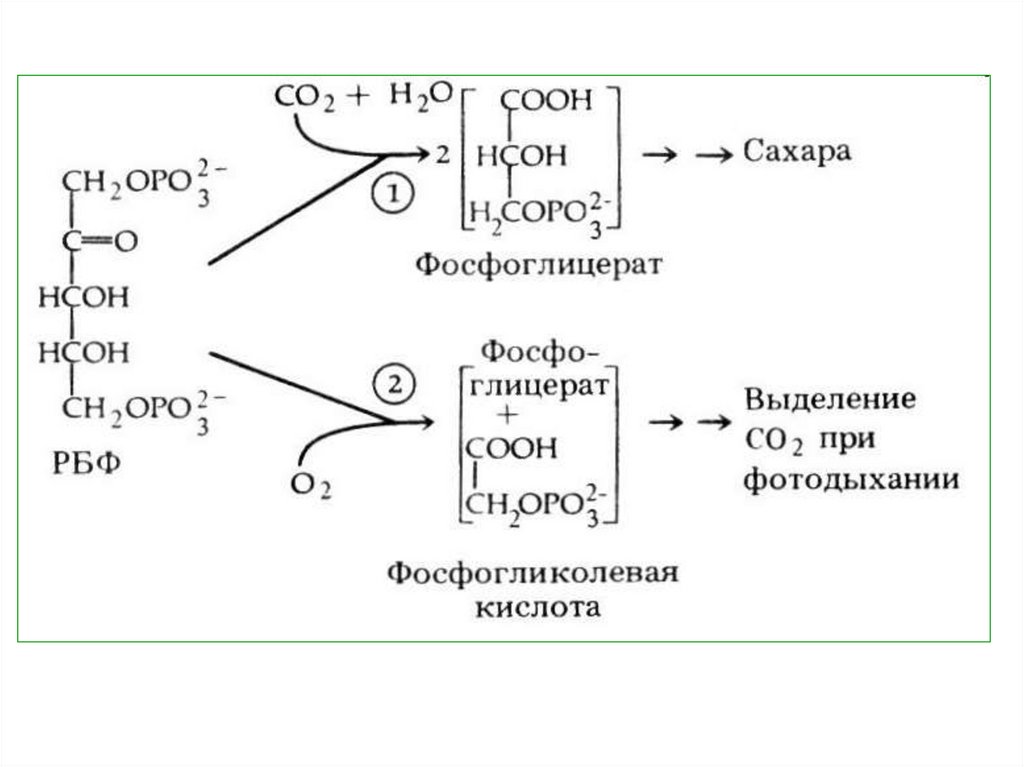
фосфогликолевой кислоты.
109.
РДФ-карбоксилаза(РДФ-оксигеназа)
РДФ
+
О
2
(С5)
ФГК + фосфогликолевая кислота
(С3)
(фосфогликолат – С2)
110.
ФГК идет на синтез органическихсоединений в цикле Кальвина, а
фосфогликолевая
кислота
транспортируется
через
мембрану хлоропласта. Во время
транспортировки
она
дефосфорилируется,
превращаясь
в
гликолевую
кислоту, которая поступает в
перокисому.
111.
Здесь гликолат реагирует скислородом воздуха, образуя
пероксид
водорода
и
глиоксиловую
кислоту.
Пероксид водорода разлагается
каталазой,
а
глиоксалат
в
результате трансаминирования
превращается в аминокислоту
глицин, которая поступает в
митохондрию.
112.
Здесь из двух молекул глицинасинтезируется одна молекула
серина, образуется аммиак и
восстанавливается НАД. СО2
чаще
всего
выделяется
в
атмосферу, т.е. теряется для
растения. НАД ∙ Н + Н+
поступает в ЭТЦ митохондрии
для синтеза АТФ.
113.
Серинвозвращается
в
пероксисому, где после ряда
превращений
образует
ФГК,
которая может поступать в С3цикл. Аммиак не выделяется в
среду, а связывается в виде
глутамина. Таким образом, в
фотодыхании
участвуют
хлоропласты, пероксисомы и
митохондрии.
114.
115.
Присравнительно
высоких
концентрациях
СО2
у
С3растений происходит фиксация
СО2 в цикле Кальвина. При
более низких концентрациях
СО2,
приближающихся
к
атмосферным
(0,045
%)
в
хлоропласте
образуется
значительное
количество
фосфогликолевой кислоты.
116.
117.
Интенсивноидущее
фотодыхание может снизить
продуктивность
фотосинтеза на 30 %, т.к.
при
фотодыхании
происходит не связывание, а
потеря СО2.
118.
Фотодыхание - процесснеобходимый растениям.
Оно является источником
метаболитов, необходимых
для различных синтезов и
поставляет
восстановленный НАД в
ЭТЦ.
119.
Считают,что
избыток
продуктов
цикла
Кальвина
усиливает
фотодыхание,
а
недостаток
активизирует
фотосинтез.
Следовательно,
соотношение фотосинтеза и
фотодыхания
выполняет
в
растении регуляторную роль.
120.
УС4-растений
интенсивность
фотодыхания
очень
низкая,
потому
что
у
них
высокая
концентрация
СО2
в
клетках
обкладки. К тому же в них
понижена
концентрация
кислорода, т.к. в хлоропластах
этих
клеток
идет
только
циклическое фосфорилирование,
а значит и не идет фотолиз, в
результате которого образуется
кислород.
121.
Образующийсяпри
фотодыхании СО2 у С4-растений
тут же используется в клетках
мезофилла ФЕП-карбоксилазой,
поэтому С4-растения
характеризуются низкой
потерей СО2 в ходе
фотодыхания.
122.
Интенсивностьфотодыхания
у
С3растений возрастает при
повышения температуры
и
освещенности.
Оптимальная температура
для них 20-25 С.
123.
Т.к.у
С4-растений
фотодыхание
почти
отсутствует,
они
могут
эффективнее,
чем
С3растения использовать свет и
высокую температуру.
124.
Т.к. образование продуктовфотосинтеза у С4-растений
происходит в хлоропластах,
расположенных
около
проводящих пучков, то это
благоприятствует
оттоку
ассимилянтов
и
как
следствие
повышает
интенсивность фотосинтеза.
125.
ЭволюционноС4-путь
возник позднее С3-пути и
характерен
только
для
цветковых растений.
126. МЕТАБОЛИЗМ ПО ТИПУ ТОЛСТЯНКОВЫХ
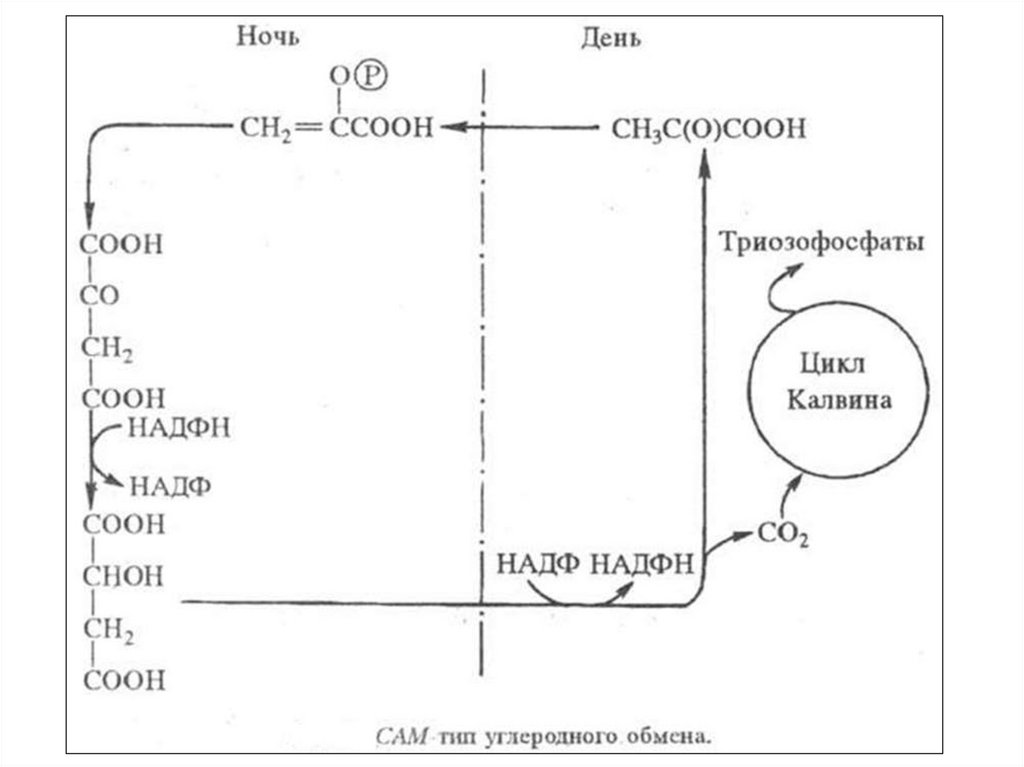
(САМ - crassulaean acid metabolism)127.
•Усуккулентных
растений
устьица
днем
закрыты
для
уменьшения транспирации. Они
поглощают ночью, когда устьица
широко
открыты,
большое
количество СО2.
128.
• Этот СО2 используется как иу
С4-растений
для
карбоксилирования ФЕП с
образованием ЩУК, которая
восстанавливается до ЯК. ЯК
накапливается в вакуоли,
поэтому в течение ночи
клеточный сок становится
все более кислым.
129.
• Утром она транспортируется вхлоропласты, где происходит ее
окислительное
декарбоксилирование,
что
приводит к образованию ПВК и
СО2 и восстановлению НАДФ+.
СО2 и НАДФ ∙ Н + Н+
используются
в
цикле
Кальвина,
а
ПВК,
фосфорилируется
с
образованием ФЕП.
130.
• Онв
результате
ряда
превращений дает ФГК,
которая поступает в цикл
Кальвина
и
служит
источником
образования
сахаров.
131.
• ФЕП,необходимый
для
акцептирования
СО2
образуется ночью в ходе
гликолиза. Таким образом,
С3-цикл у растений типа
толстянковых идет днем
при закрытых устьицах, что
предотвращает
потери
воды.