



















")




















 ЭТАП КЛЕТОЧНОГО ДЫХАНИЯ")











")









 ЦЕПИ")



")



















")






 biology
biologySimilar presentations:










Клеточный катаболизм (энергетический обмен в клетке)
1. КЛЕТОЧНЫЙ КАТАБОЛИЗМ
(ЭНЕРГЕТИЧЕСКИЙОБМЕН В КЛЕТКЕ)
2.
Катаболизм - это процессрасщепления
органических
веществ,
в
результате
которого
энергия
органических
соединений
передается на АТФ и таким
образом в дальнейшем может
быть
использована
в
метаболических
процессах
клетки.
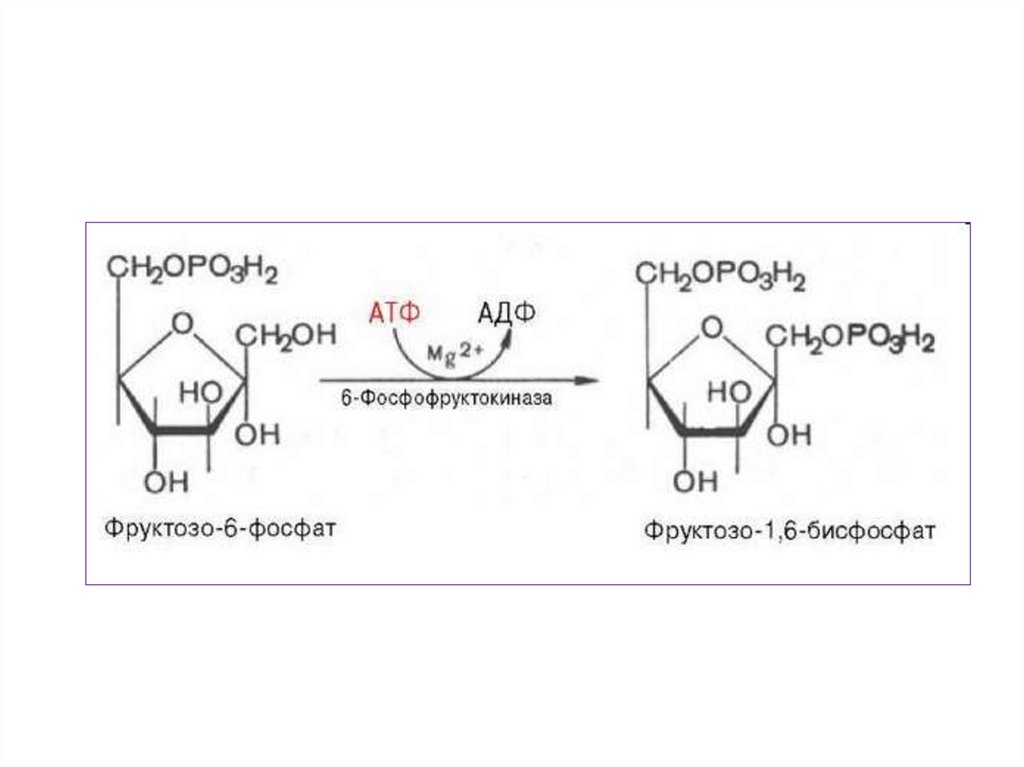
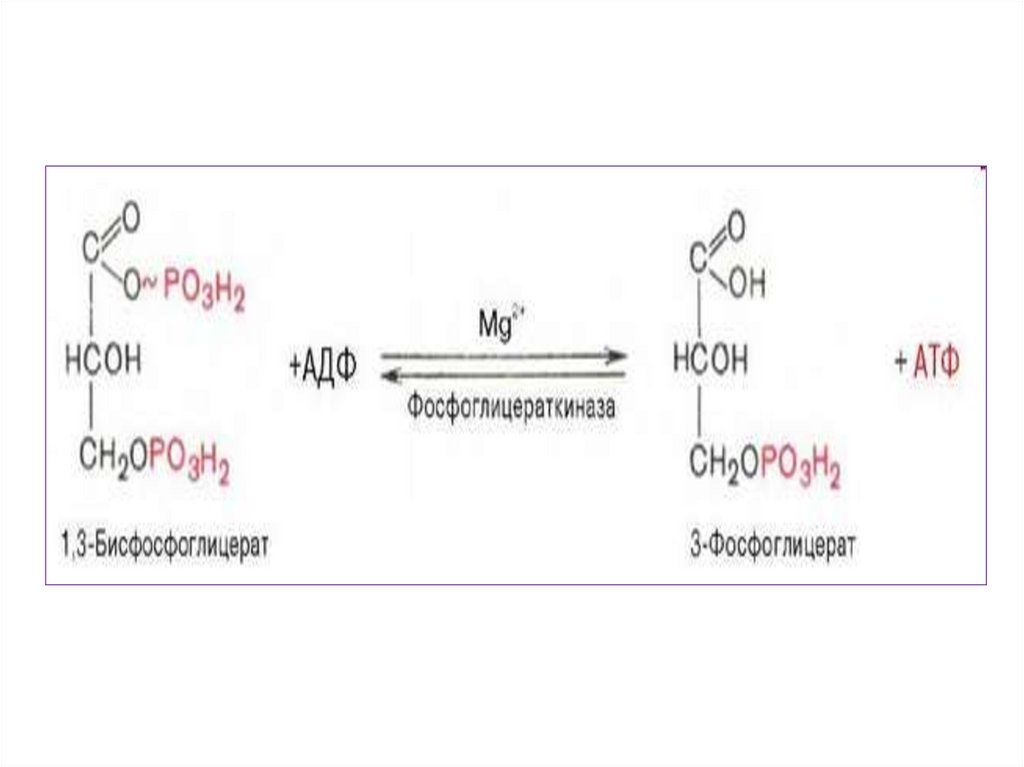
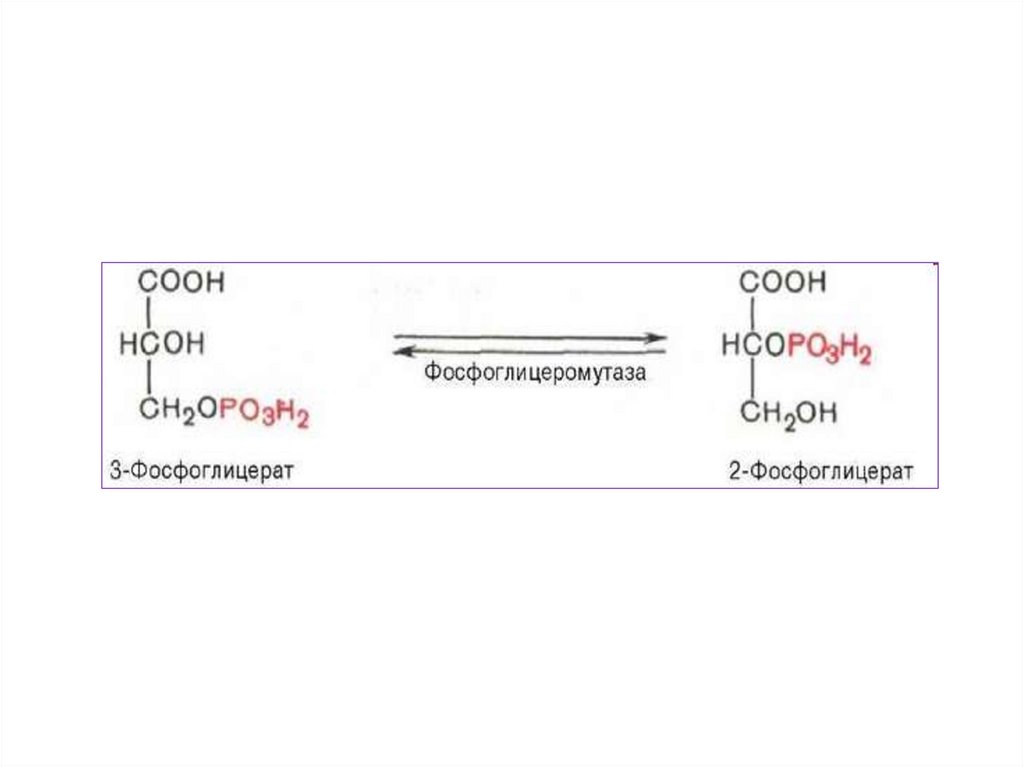
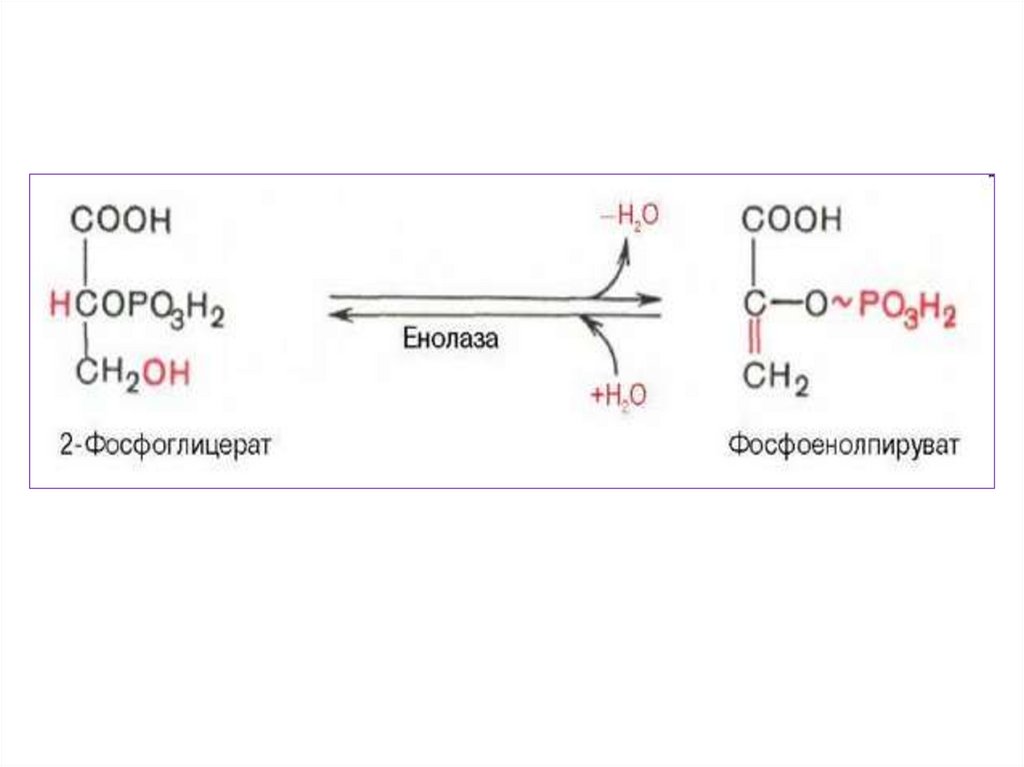
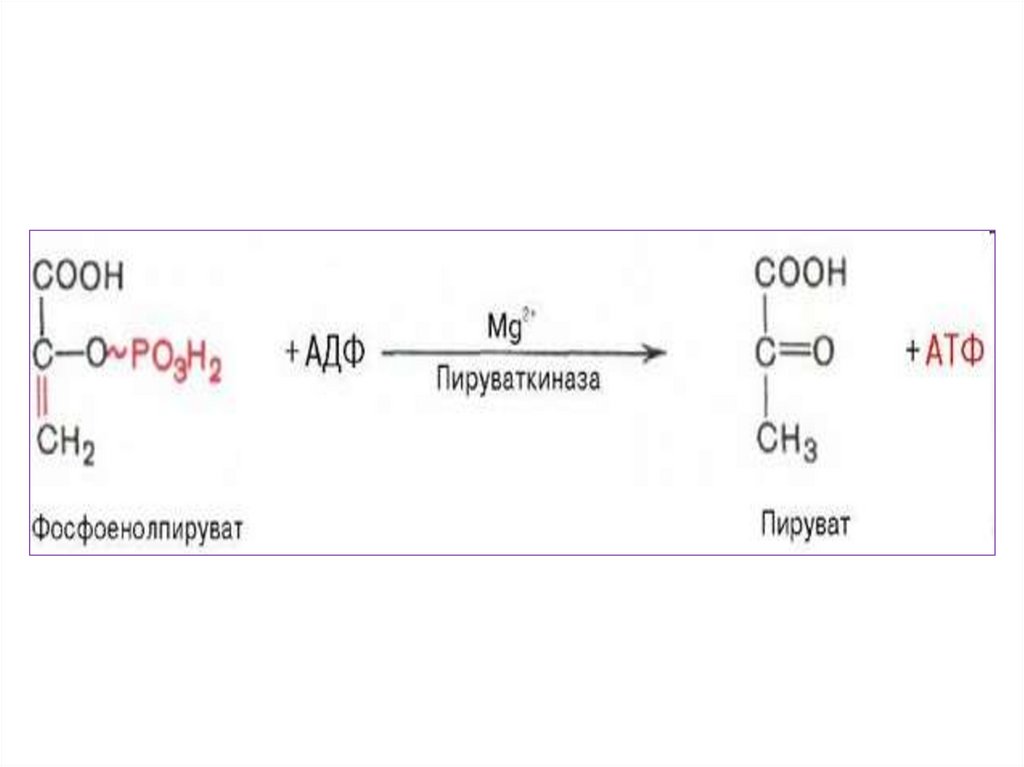
3.
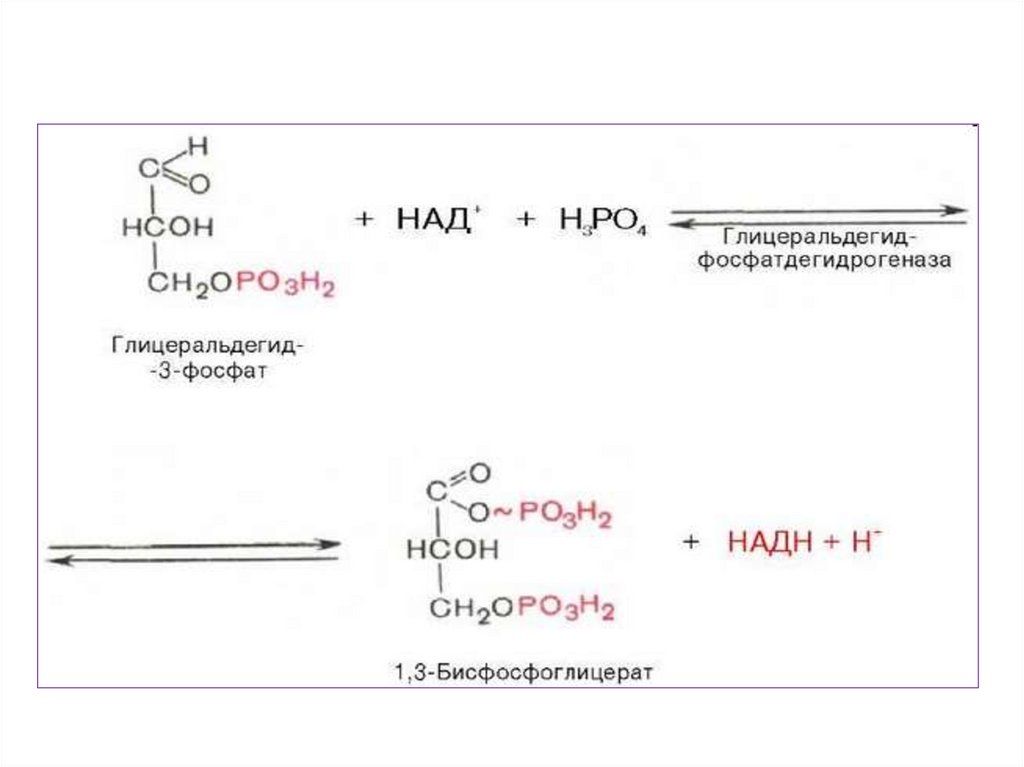
• Для осуществления катаболическихпроцессов
необходим
подготовительный этап.
• В ходе подготовительного этапа
происходит
расщепление
органических соединений до более
простых органических веществ.
АТФ на данном этапе не образуется.
• Подготовительный
этап
осуществляется в лизосомах.
4.
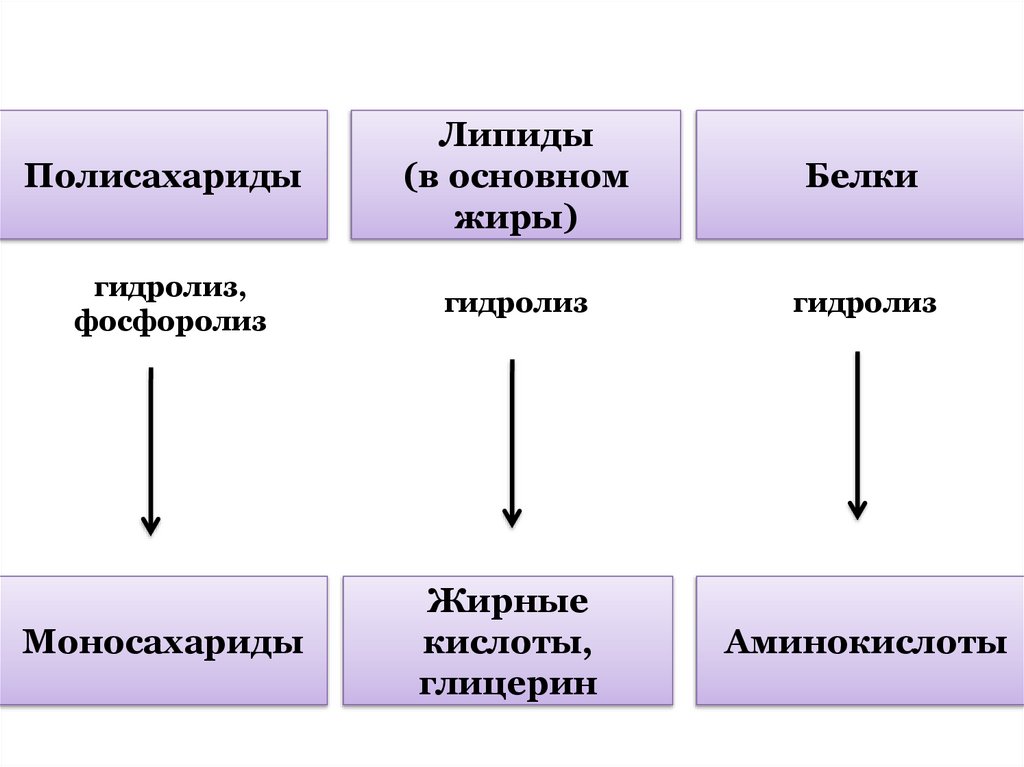
ПолисахаридыЛипиды
(в основном
жиры)
Белки
гидролиз,
фосфоролиз
гидролиз
гидролиз
Моносахариды
Жирные
кислоты,
глицерин
Аминокислоты
5.
Собственно катаболическиепроцессы
это
окислительный
распад
органического субстрата до
простых
неорганических
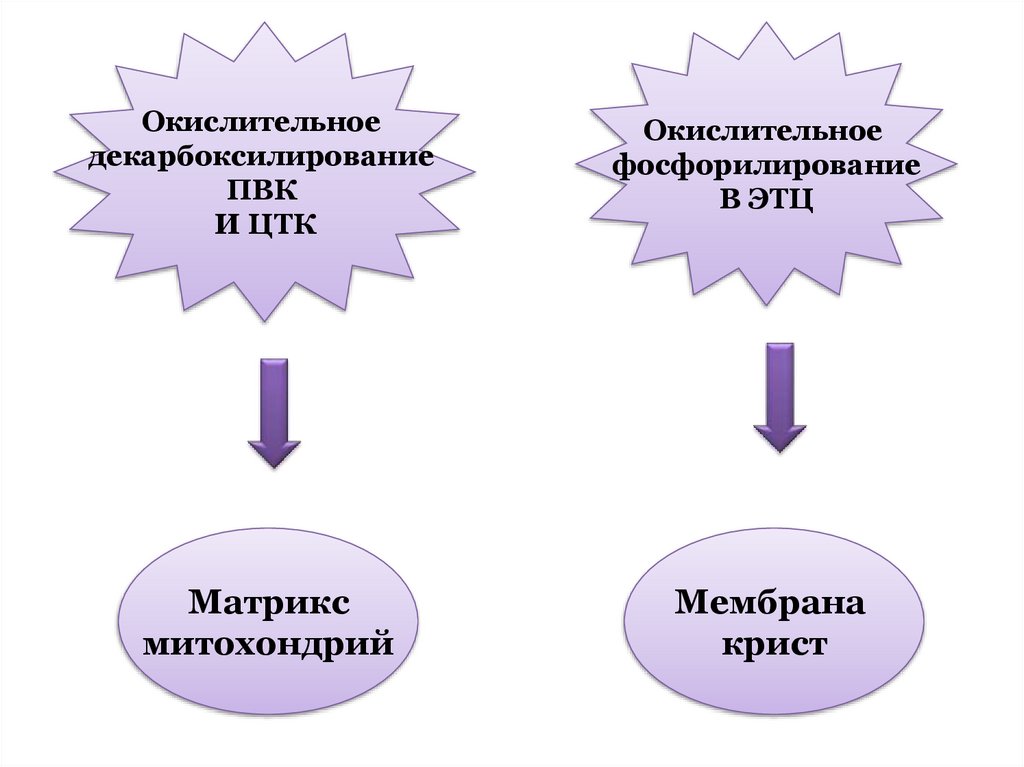
(полный распад) или простых
органических
(неполный
распад)
веществ
с
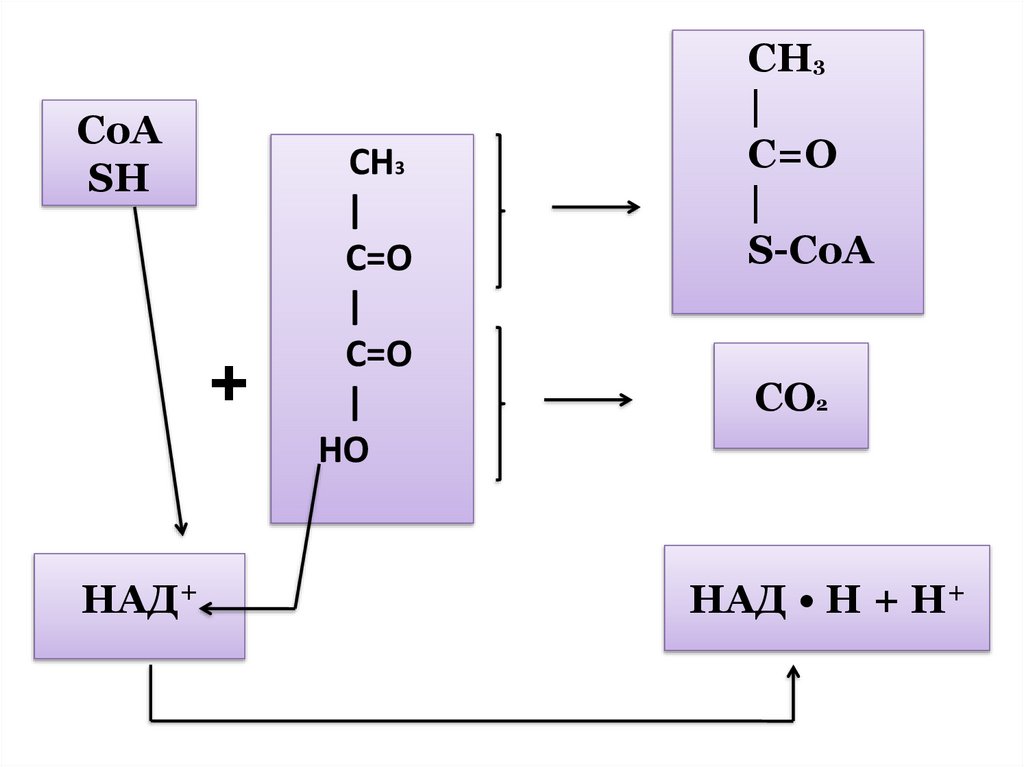
образованием
некоторого
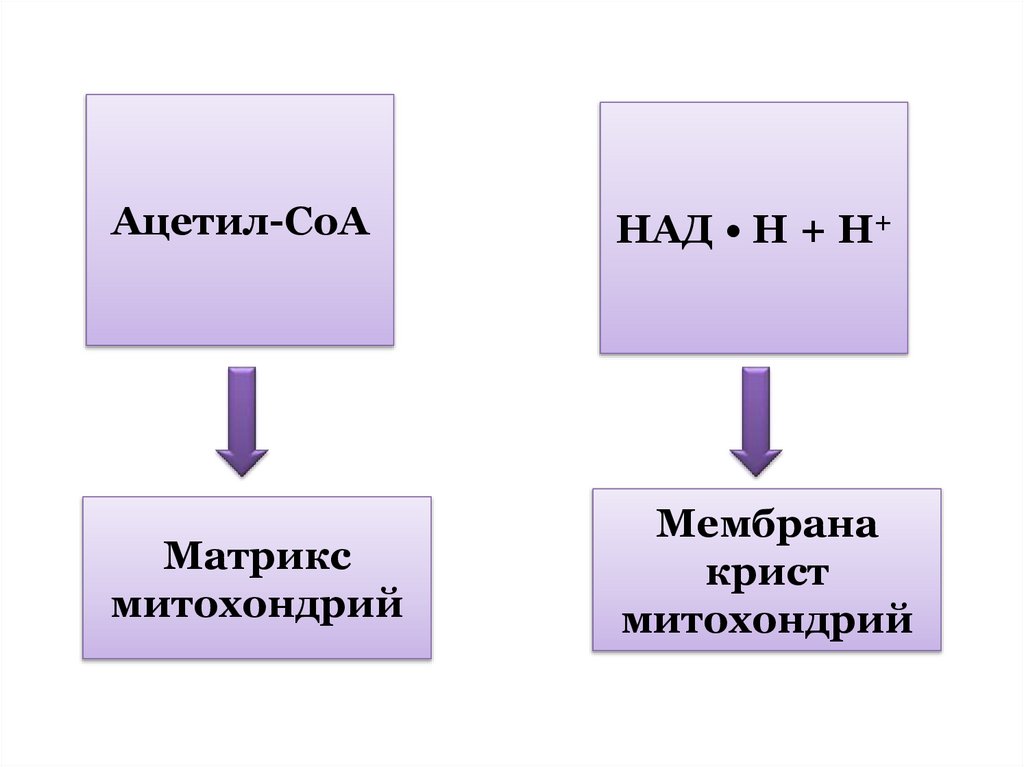
количества АТФ.
6.
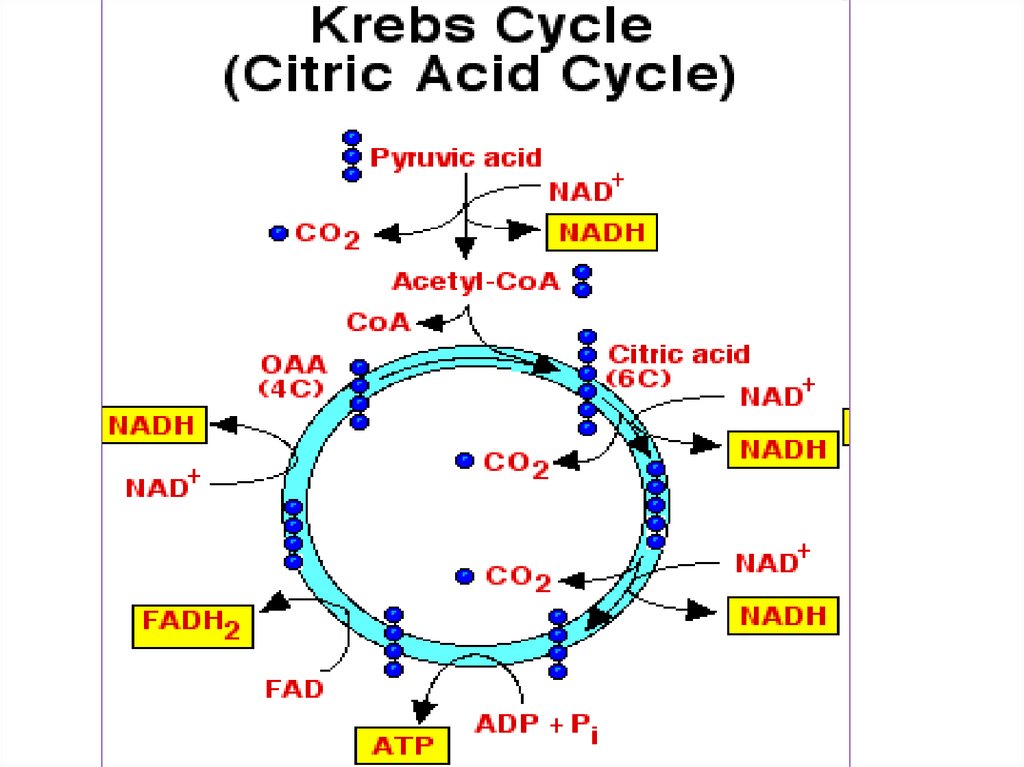
• Путь катаболических процессовзависит
от
присутствия
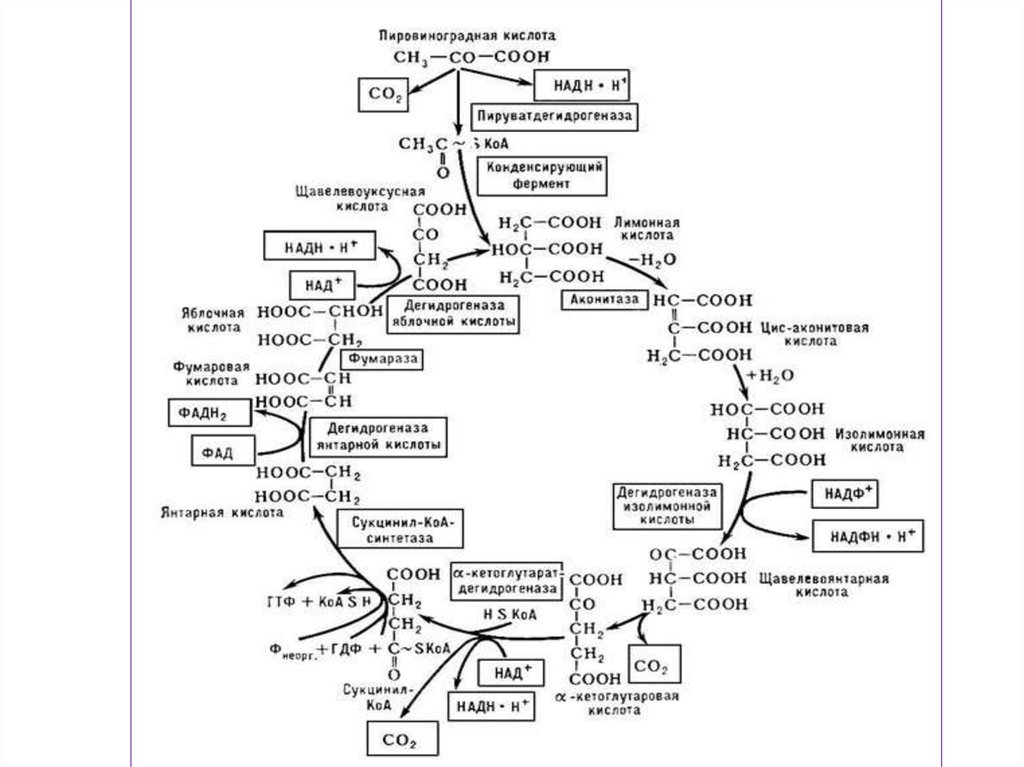
или
отсутствия в окружающей среде
кислорода.
• При
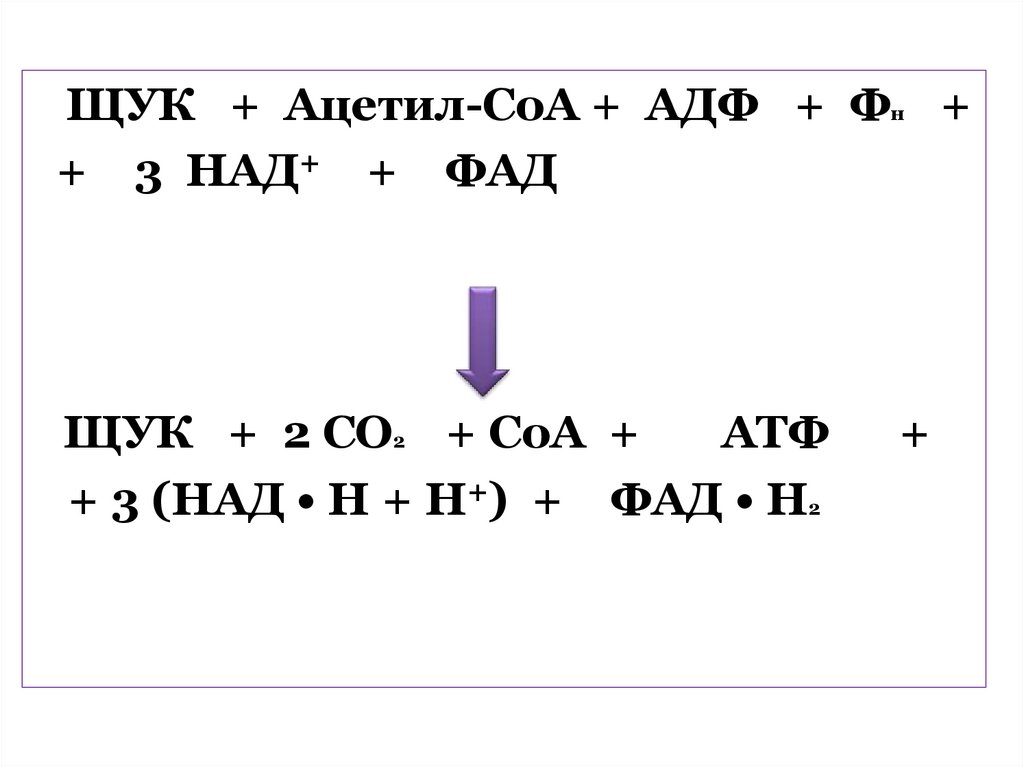
достаточном
количестве
кислорода
происходит
полный
окислительный распад субстрата до
неорганических веществ (СО2 и
Н2О).
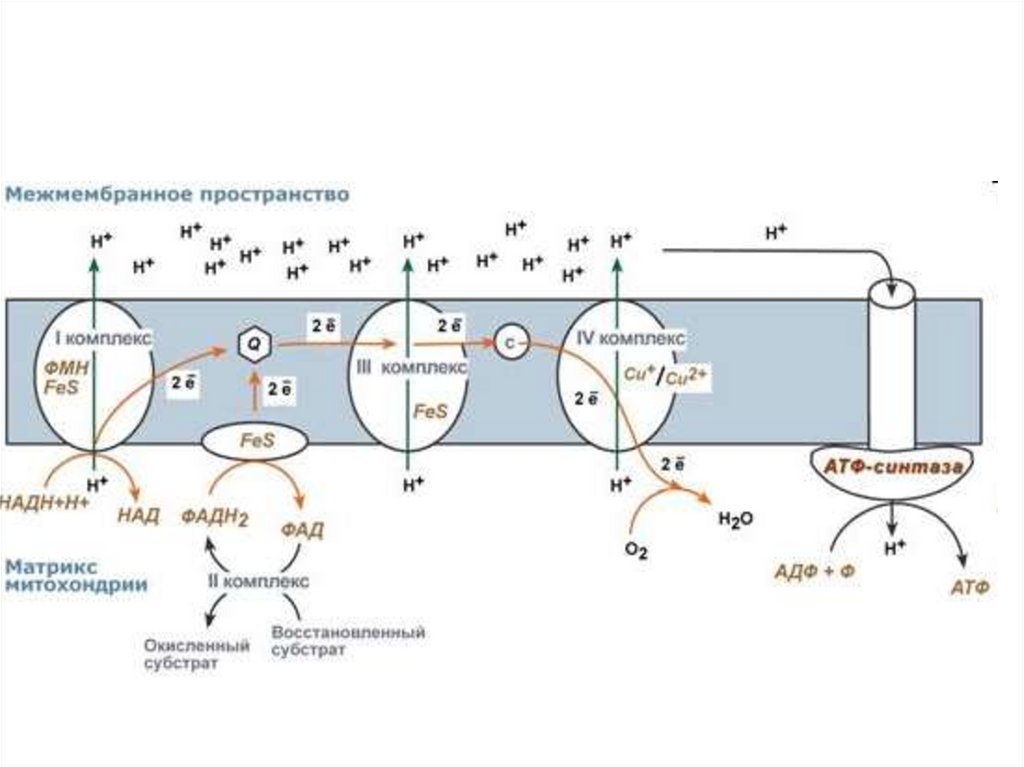
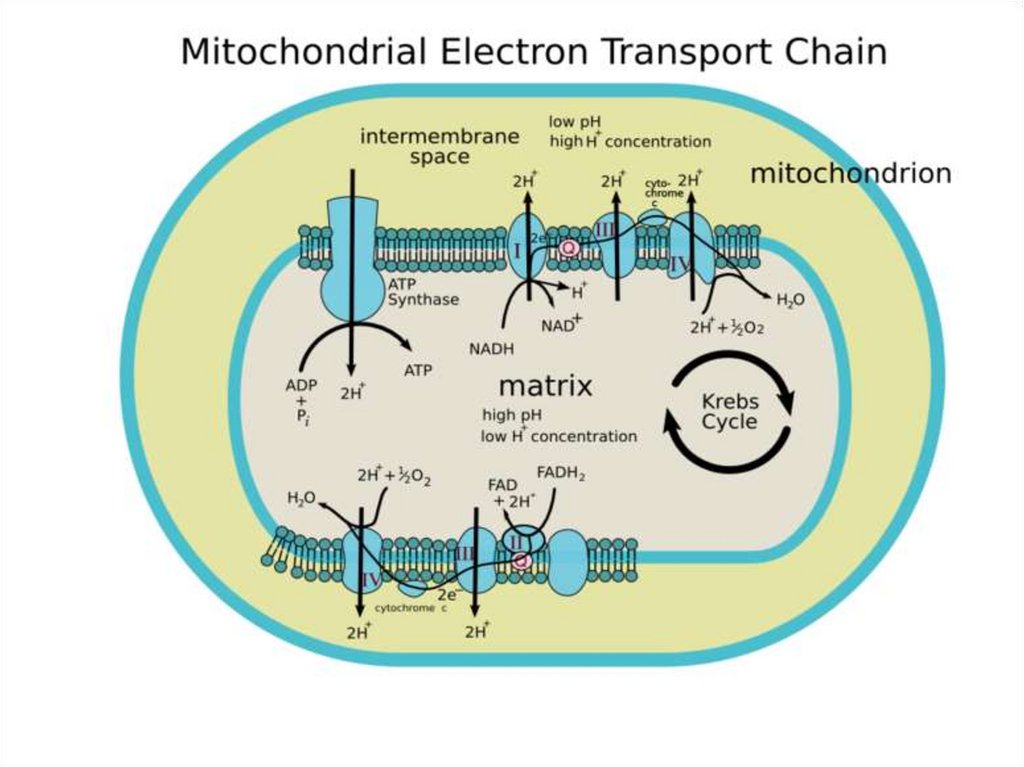
При
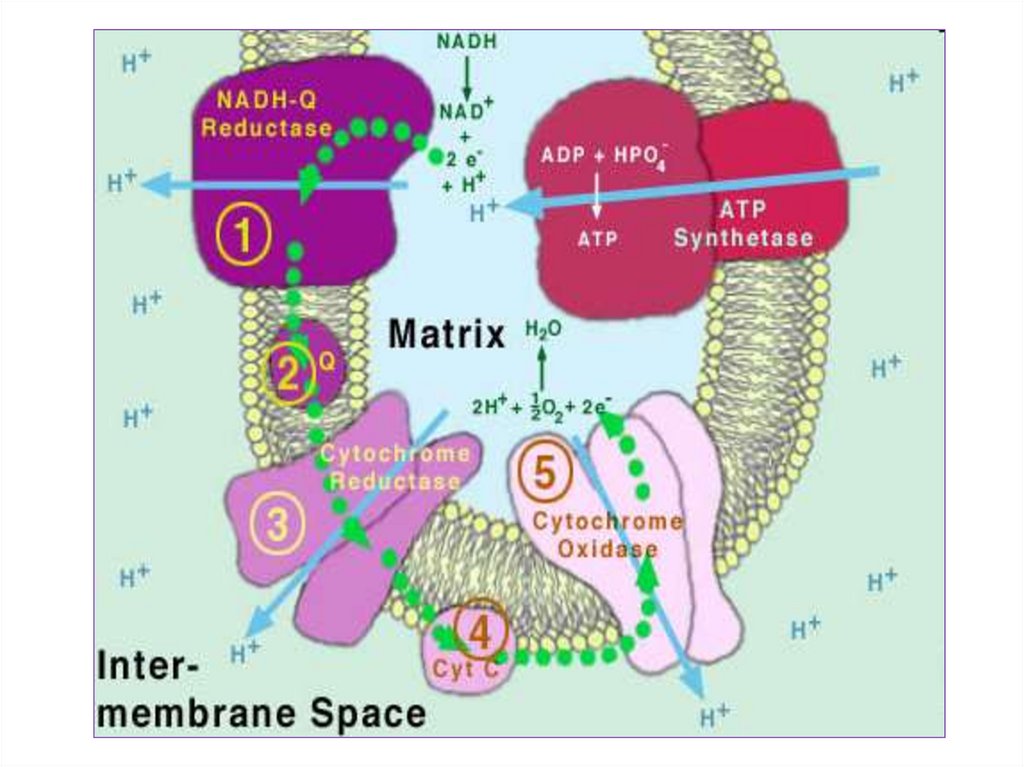
этом
образуется
довольно большое количество АТФ.
7.
При отсутствии или недостаткекислорода
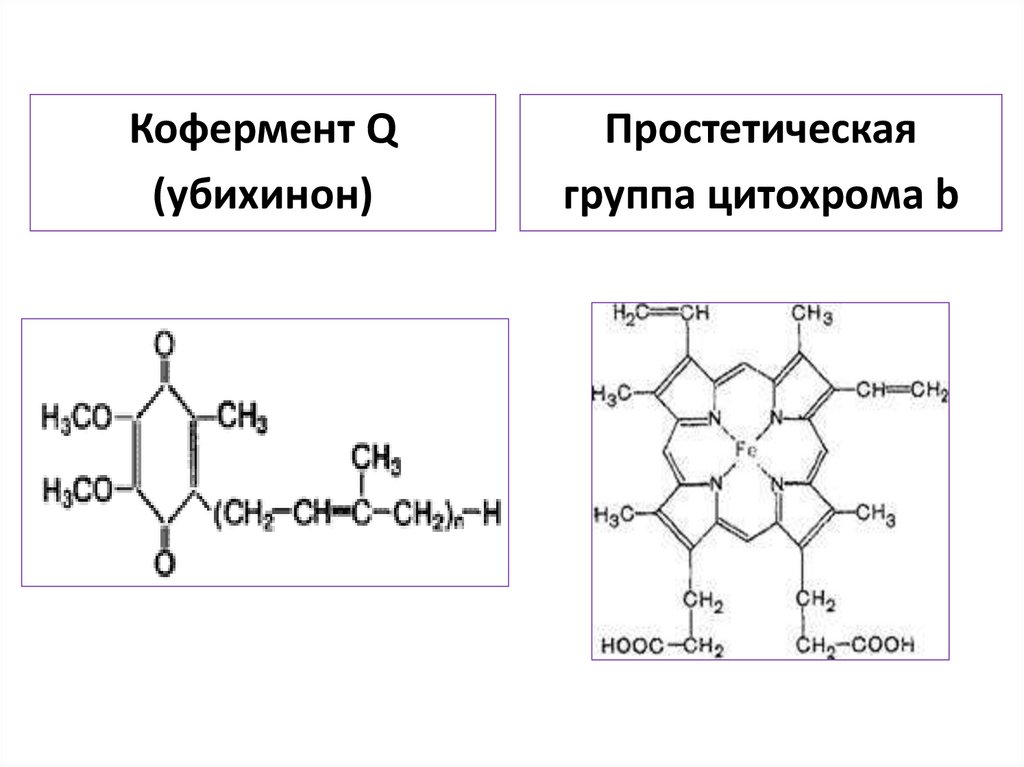
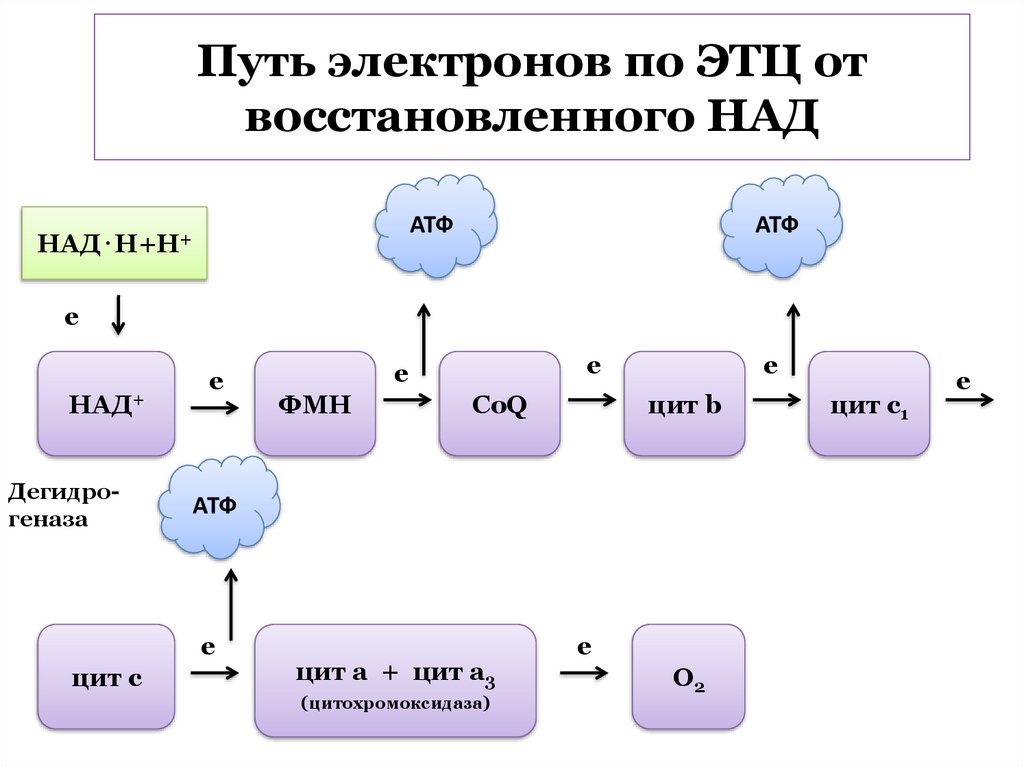
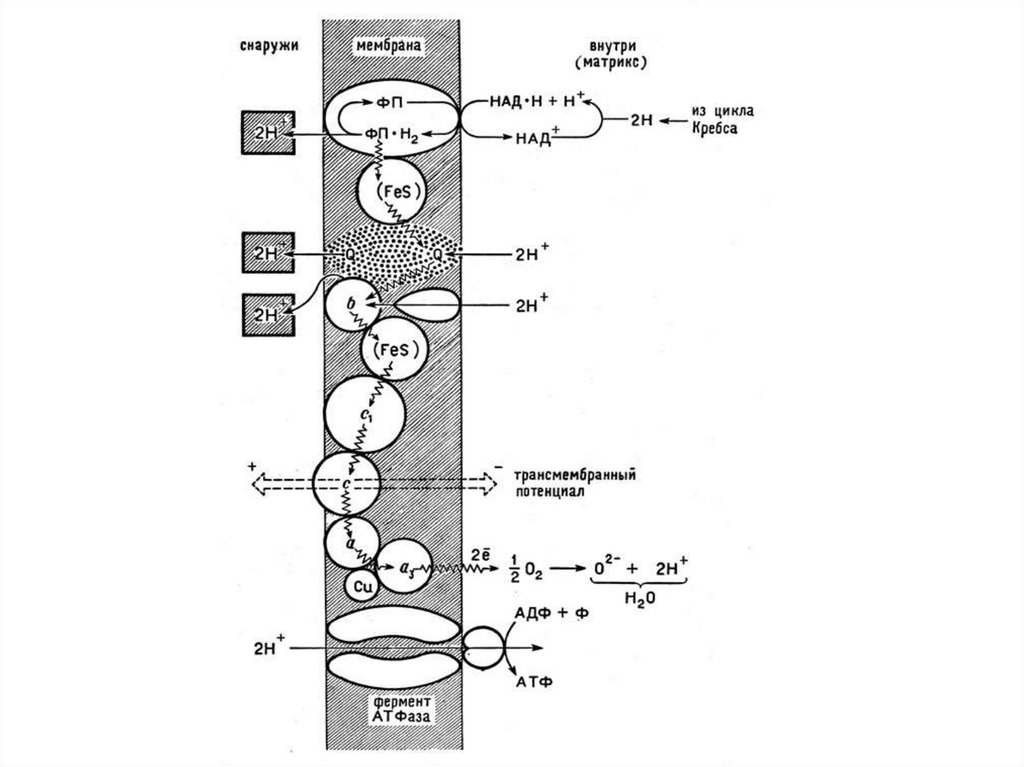
осуществляется
неполный
окислительный
распад субстрата до простых
органических веществ. АТФ
при этом образуется намного
меньше,
чем
при
полном
распаде.
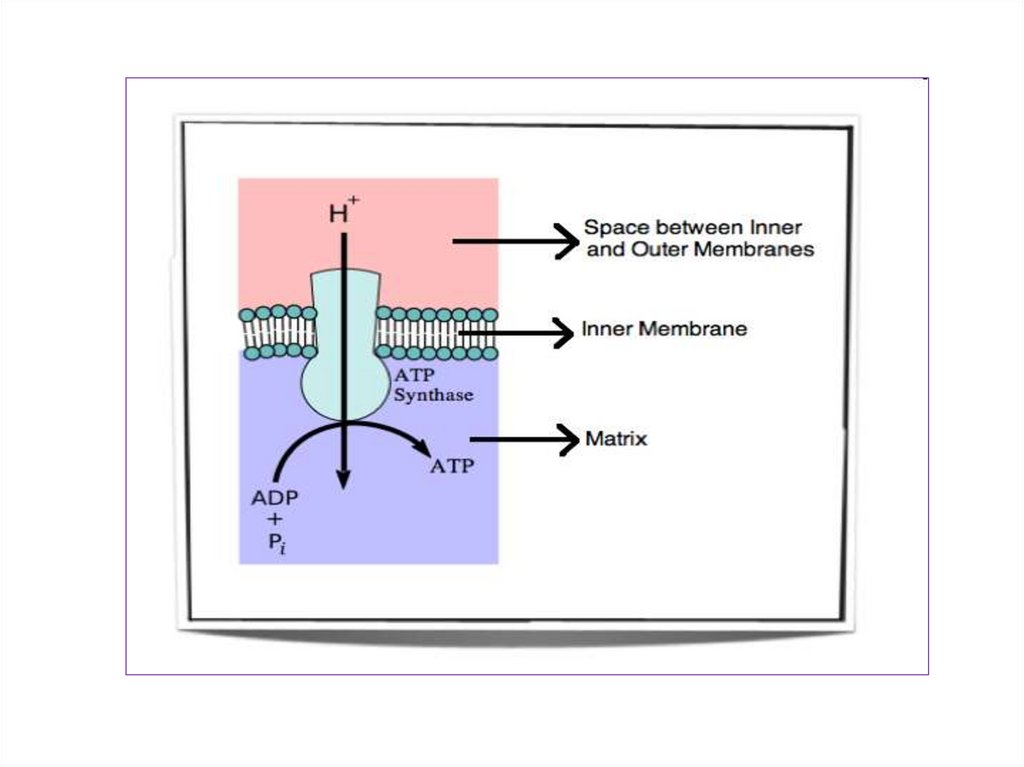
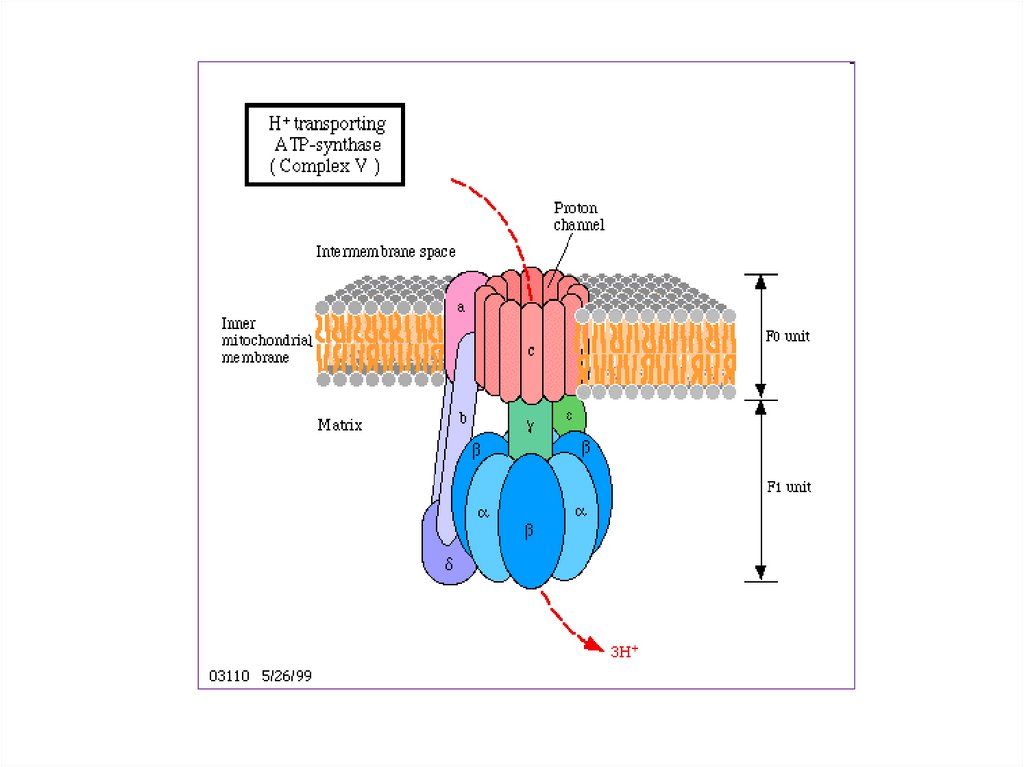
8.
Полныйокислительный
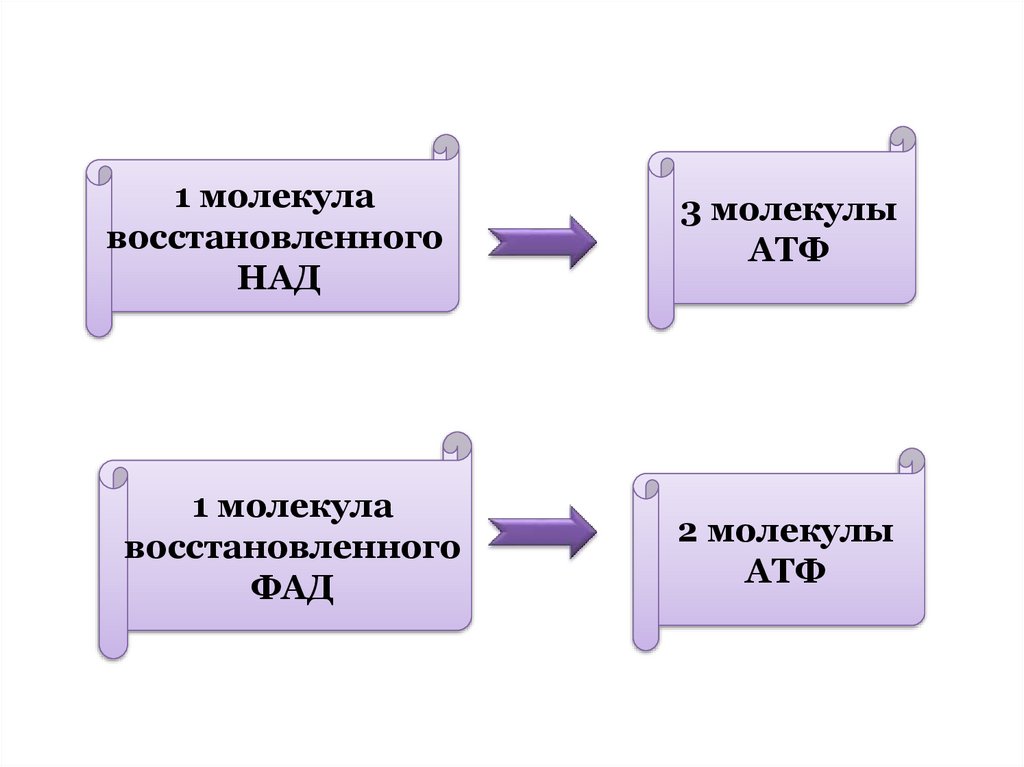
распад субстрата называют
аэробным
клеточным
дыханием,
неполный
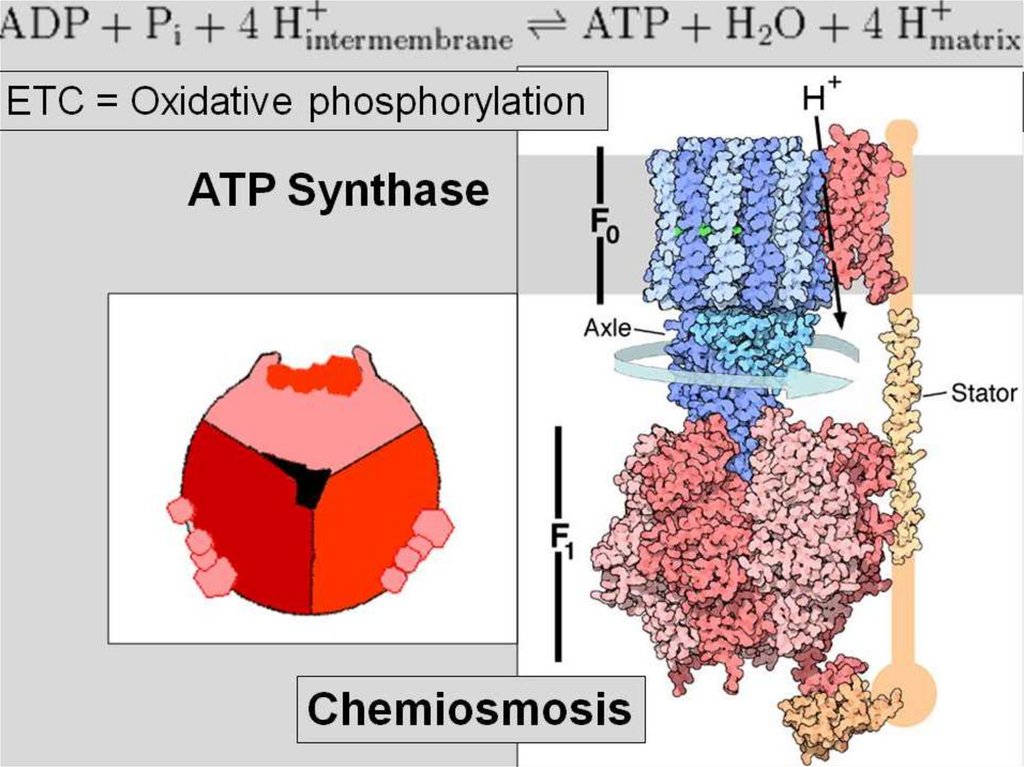
распад
анаэробным
клеточным дыханием.
9.
В более узком смысле словапод клеточным дыханием
понимают
только
аэробный
путь
катаболизма, а анаэробный
окислительный
распад
субстрата
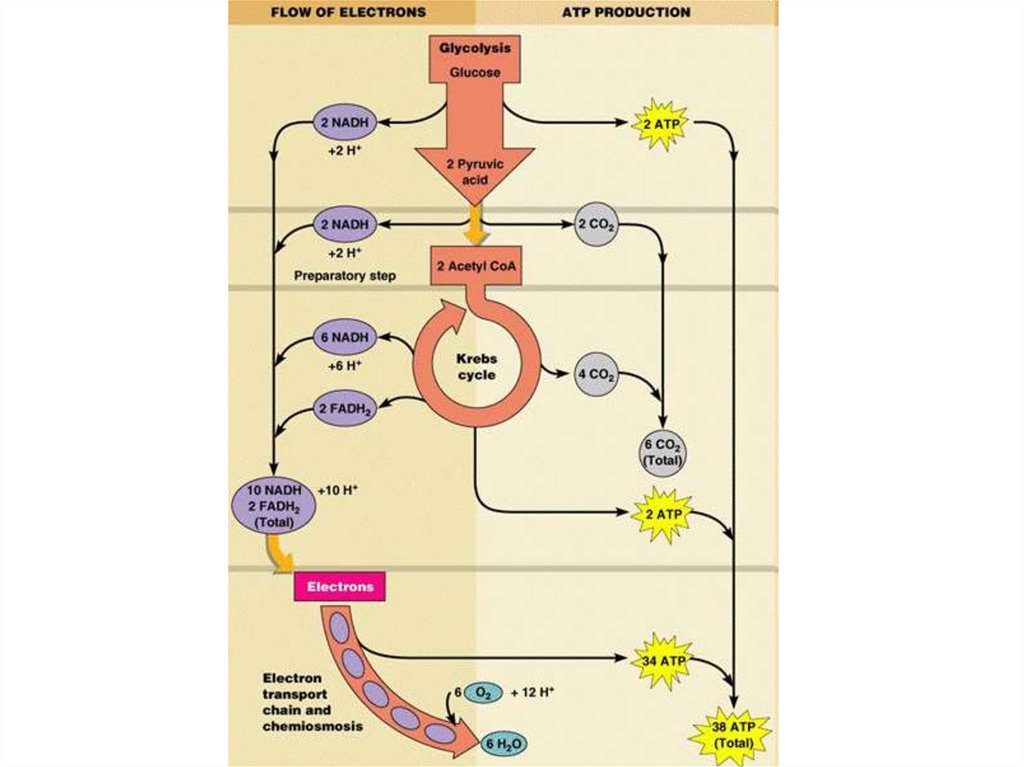
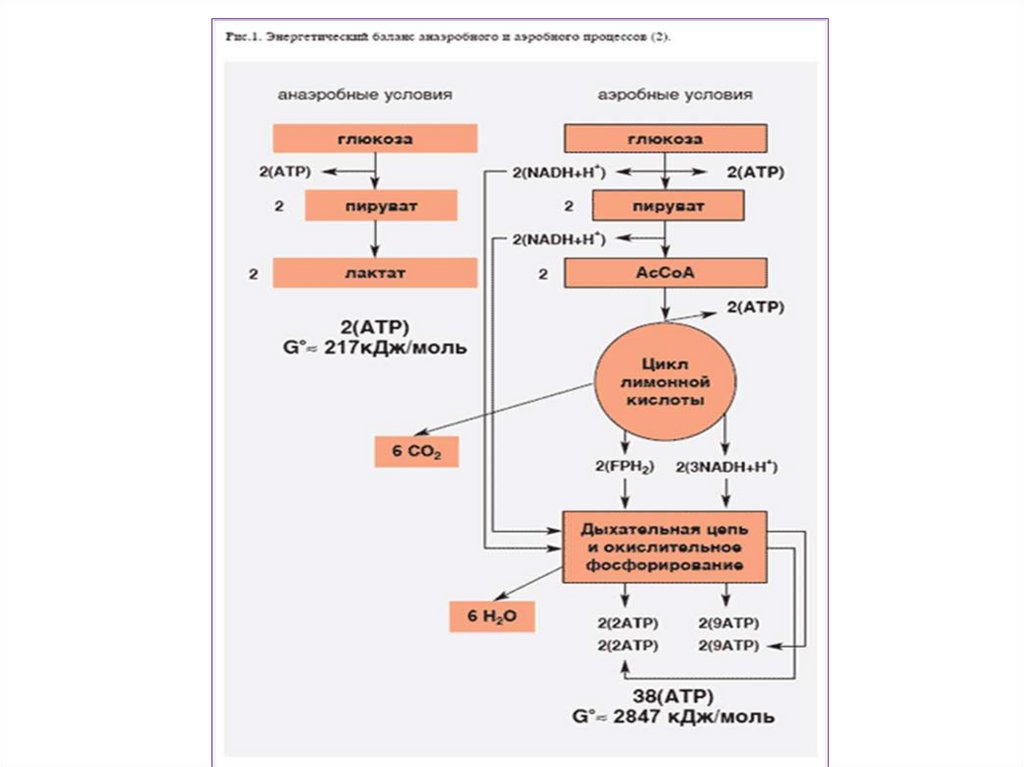
называют
брожением.
10. Классификация живых организмов по типу энергетического обмена
ОрганизмыАэробные
(аэробы)
Анаэробные
(анаэробы)
Клеточное дыхание
Брожение
Большинство
животных и
растений, часть
микроорганизмов
Облигатные
Факультативные
Живут только в
бескислородной среде
Предпочитают
бескислородные условия,
но могут жить и в
присутствии кислорода
Только некоторые
бактерии,
например
клостридии
Например,
дрожжи, цепни и
пр.
11. Типы реакций биологического окисления
• 1. Прямое окисление молекулярнымкислородом
А + О2
АО2
• 2. Реакции с переносом электронов
Fe2+
Fе3+ + е
• 3.Дегидрирование
АН2 + В
А + ВН2
12.
Вживой
клетке
атомы
водорода,
отнимаемые
от
субстрата,
передаются
на
специальные
носители
производные нуклеотидов:
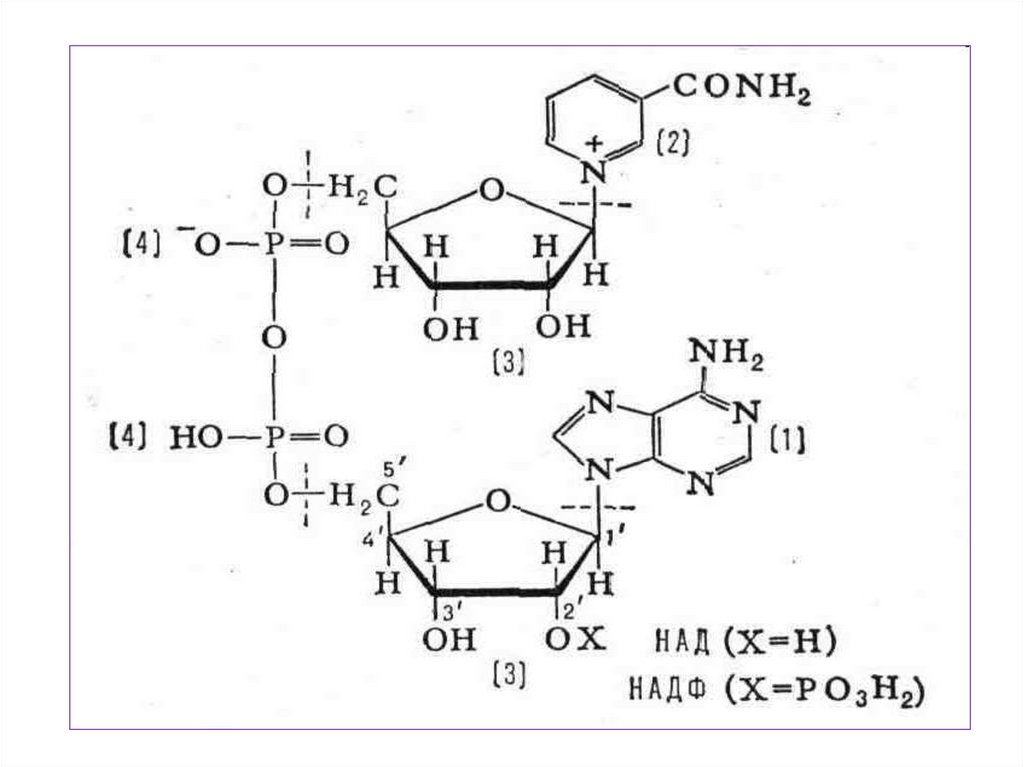
• 1. Никотинамидадениндинуклеотид НАД+(NAD+).
• 2.Никотинамидадениндинуклеофосфат
– НАДФ+ (NADP+).
13.
14.
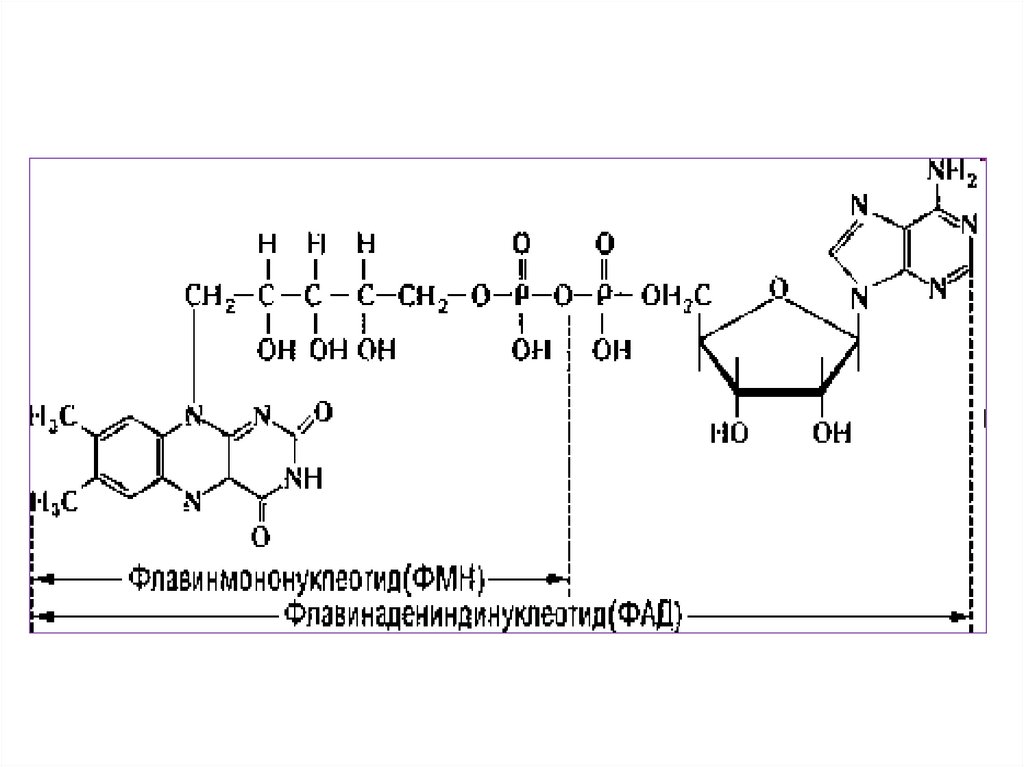
• 3. Флавинадениндинуклеотид- ФАД (FAD).
• 4. Флавинмононуклеотид ФМН (FMN).
15.
16.
Этисоединения
являются
коферментами
ферментов
дегидрогеназ,
катализирующих
отщепление
водорода.
Они
способны
присоединять
водородные атомы и переходить в
восстановленную форму - НАД . Н +
Н+ (NAD . H + H+), НАДФ . Н + Н+
(NADP . H + H+), ФАД . Н2 (FAD . H2),
ФМН . Н2, (FMN . Н2).
17.
Водород в таких комплексахнаходится в энергетически
реакционноспособном
состоянии и в дальнейшем
используется клеткой для
получения
энергии
и
восстановления субстратов.
18. АЭРОБНОЕ КЛЕТОЧНОЕ ДЫХАНИЕ
19.
Суммарное уравнение клеточногодыхания имеет следующий вид:
Органические молекулы + О2
СО2
+ Н2 О
+
Е
20.
Точнотаким
же
уравнением
описываются и процессы горения,
но в отличие от горения при
дыхании
энергия
выделяется
постепенно, малыми порциями в
ходе
последовательных
многоступенчатых
ферментативных
превращений
субстрата, что позволяет клетке
удерживать часть выделившейся
энергии в форме АТФ.
21.
Это было бы невозможным втом случае, если бы энергия
выделялась бурно, как это
происходит при горении или
взрыве.
22.
Окислительному распаду в клетке могутподвергаться различные органические
вещества - моносахариды, жирные
кислоты,
аминокислоты.
Но
значительно
преобладает
окислительный
распад
глюкозы,
который включает в себя два основных
этапа:
• 1. Бескислородный, то есть анаэробный
этап, именуемый гликолизом.
• 2. Кислородный, то есть аэробный этап
23.
Кислородный этап состоит из трехстадий:
• 1. Окислительное
декарбоксилирование
пировиноградной кислоты (ПВК).
• 2. Цикл трикарбоновых кислот
(ЦТК).
• 3. Окислительное
фосфорилирование в
электронтранспортной цепи (ЭТЦ).
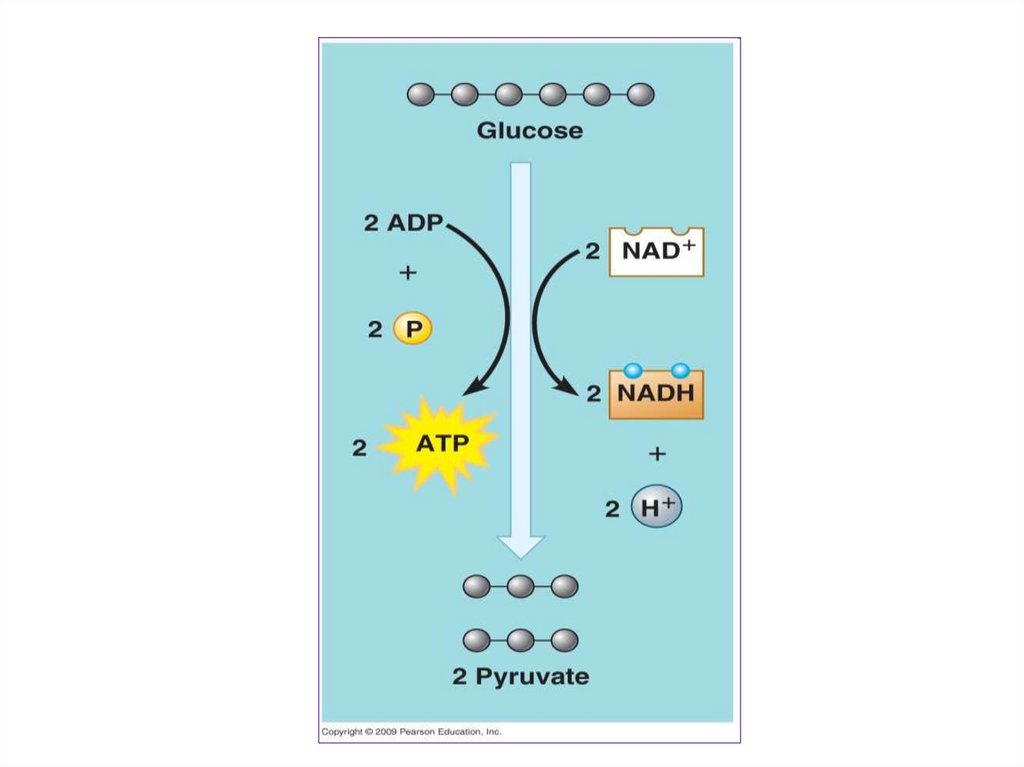
24. ГЛИКОЛИЗ (АНАЭРОБНЫЙ ЭТАП)
25.
Гликолиз - это многоступенчатыйферментативный окислительный
распад глюкозы до двух молекул
пировиноградной
кислоты.
Гликолиз
происходит
в
цитоплазме
и
не
требует
присутствия
кислорода.
Ферменты,
катализирующие
реакции гликолиза, присутствуют
в
растворенном
виде
в
цитоплазме.
26.
Когда молекула глюкозы в ходепоследовательно
идущих
реакций
окисляется,
часть
выделившейся
энергии
запасается
в
виде
АТФ.
Согласно
II
закону
термодинамики
часть
химической
энергии
рассеивается в виде тепла.
27.
У птиц и млекопитающих тепло,выделяемое при дыхании, тем или
иным
образом
сохраняется,
поэтому температура их тела в
холодное
время
года
выше
температуры окружающей среды. У
растений
скорость
дыхания
относительно невелика, поэтому
выделенное тепло не влияет на
температуру самого растения.
28.
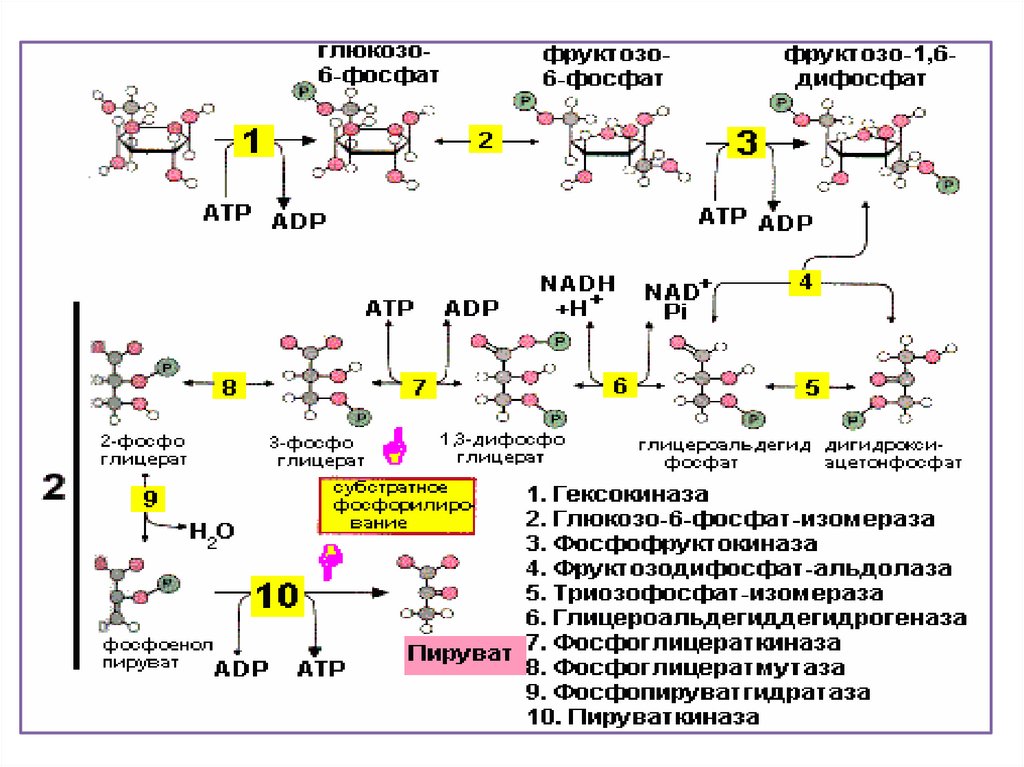
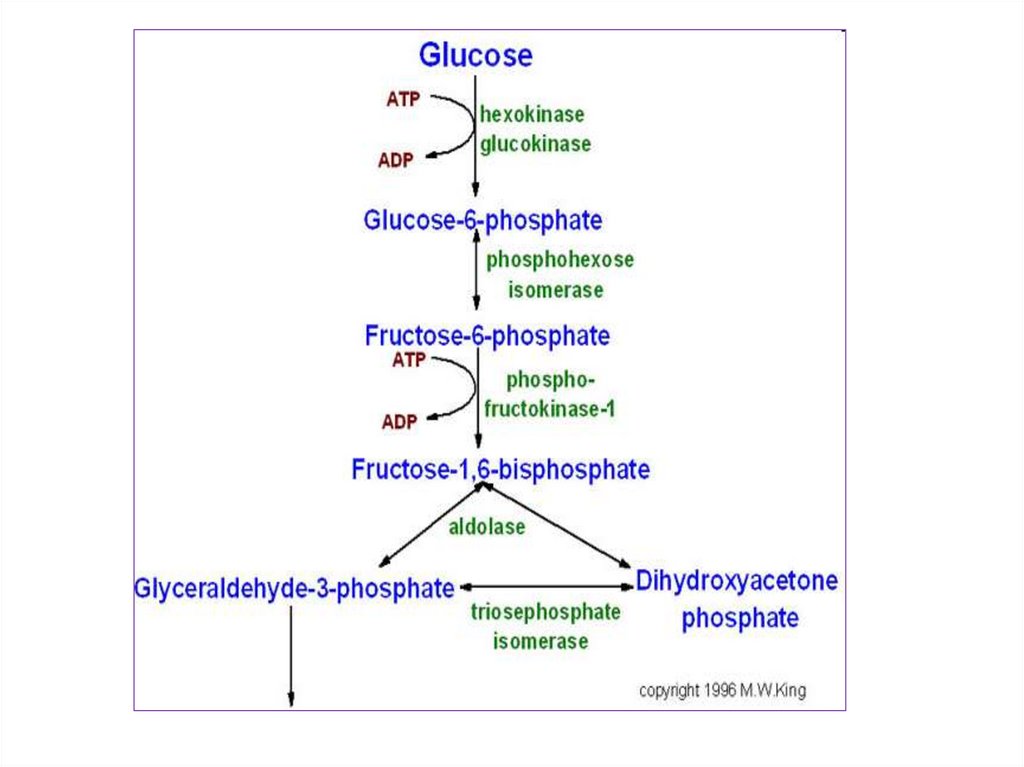
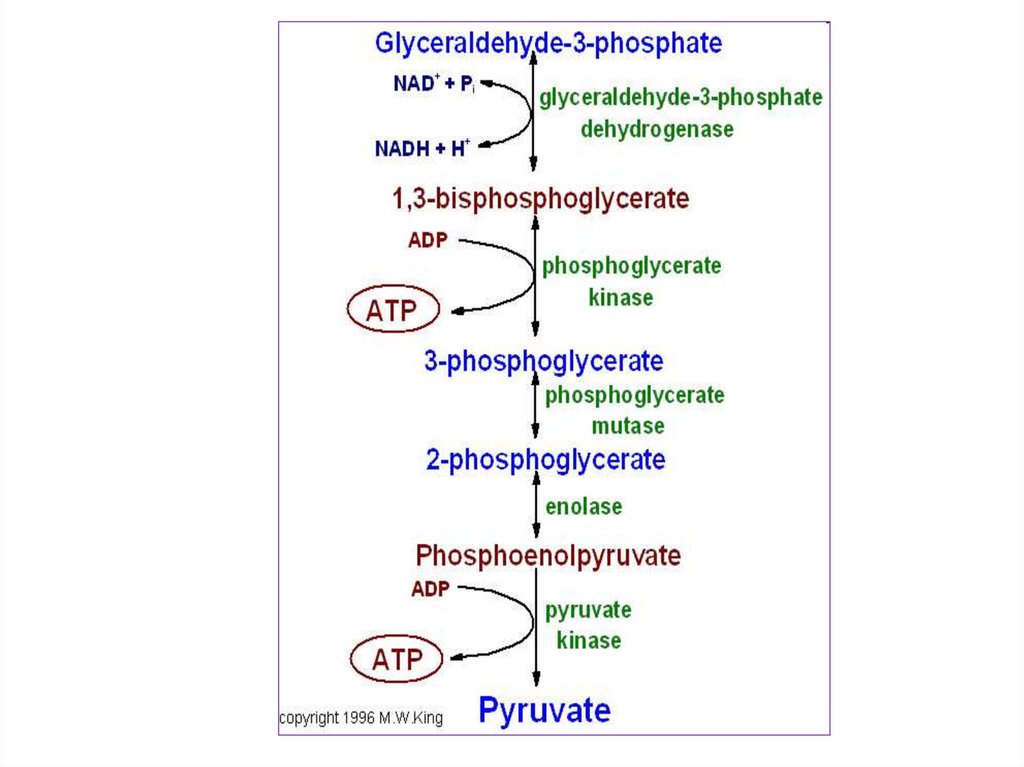
Гликолиз состоит из десятипоследовательных реакций.
29.
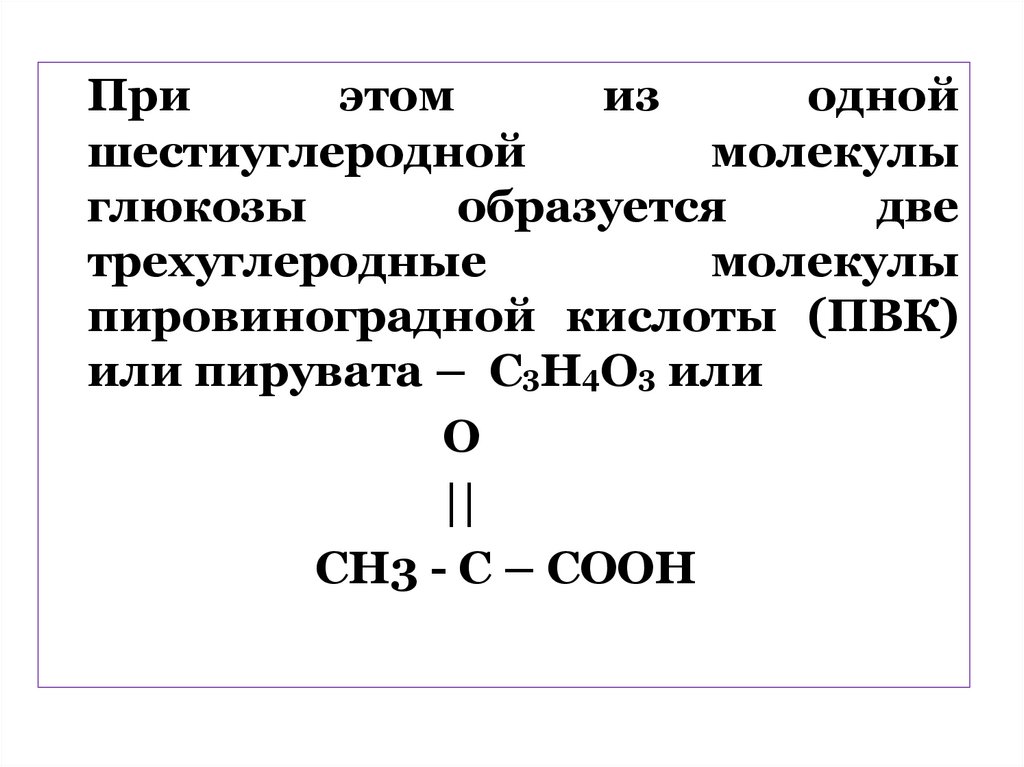
Приэтом
из
одной
шестиуглеродной
молекулы
глюкозы
образуется
две
трехуглеродные
молекулы
пировиноградной кислоты (ПВК)
или пирувата – C3Н4О3 или
О
||
СН3 - С – СООН
30.
Молекулаглюкозы
при
гликолизе теряет четыре атома
водорода. Акцептором атомов
водорода
выступают
две
молекулы НАД+. Присоединяя
водород, окисленная форма
НАД+
переходит
в
восстановленную – НАД . Н +
Н+. Образуется две таких
молекулы.
31.
Крометого,
происходит
образование четырех молекул АТФ
путем
субстратного
фосфорилирования. Так как на
первых стадиях гликолиза две
молекулы АТФ расходуются, то
чистый выход АТФ при гликолизе
составляет
две
молекулы
в
пересчете
на
одну
молекулу
глюкозы.
32. Энергетический выход гликолиза
Выделившаясяэнергия
60%
рассеивается
в виде тепла
40%
запасается в
форме АТФ
33.
Суммарное уравнение гликолизаимеет следующий вид:
С6Н12О6 + 2 АДФ + 2 Фн + 2 НАД+
2 С3Н4О3+ 2 АТФ + 2 (НАД . Н + Н+ )
34.
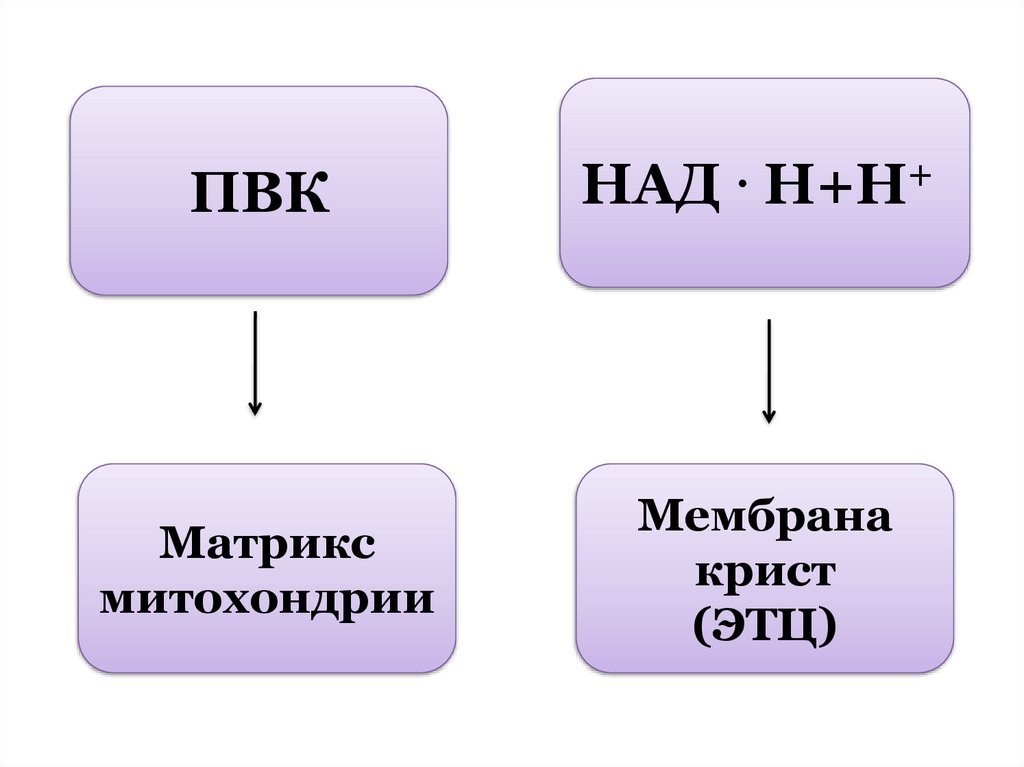
ПВКНАД . Н+Н+
Матрикс
митохондрии
Мембрана
крист
(ЭТЦ)
35.
Гликолиз - это последовательностьреакций общая для процесса
дыхания и брожения. Во время
гликолиза
синтезируется
небольшое
количество
АТФ.
Многие виды бактерий, грибов,
некоторые
мелкие
беспозвоночные
способны
существовать,
довольствуясь
только
АТФ,
полученном
в
процессе гликолиза.
36.
У большинства же организмовпроисходит полное окисление
глюкозы, а поэтому есть еще и
цикл трикарбоновых кислот и
электронтранспортная цепь,
поставляющие
клетке
во
много раз больше энергии,
нежели гликолиз.
37.
У организмов, у которыхесть ЦТК и ЭТЦ (аэробных
организмов),
главное
назначение
гликолиза
состоит
в
том,
чтобы
вырабатывать
ПВК
для
ЦТК.
38.
Гликолизфилогенетически
наиболее
древний
способ
добывания энергии, и для него не
нужно специальных органоидов.
Вероятно, он возник до появления
кислорода в атмосфере Земли и
формирования
клеточных
органелл.
39.
Первые живые организмы наЗемле были анаэробными, так как
в атмосфере Земли отсутствовал
кислород.
Они
вырабатывали
энергию путем гликолиза. Когда
же концентрация кислорода в
атмосфере
достигла
1
%
от
современного
состояния
атмосферы,
стал
возможен
переход от гликолиза на путь
аэробного клеточного дыхания.
40.
Концентрация кислорода, прикоторой стал возможен этот
переход,
получила
название
пастеровской точки, а сама
возможность
переключения
с
гликолитического пути на путь
аэробного клеточного дыхания
была
названа
эффектом
Пастера.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
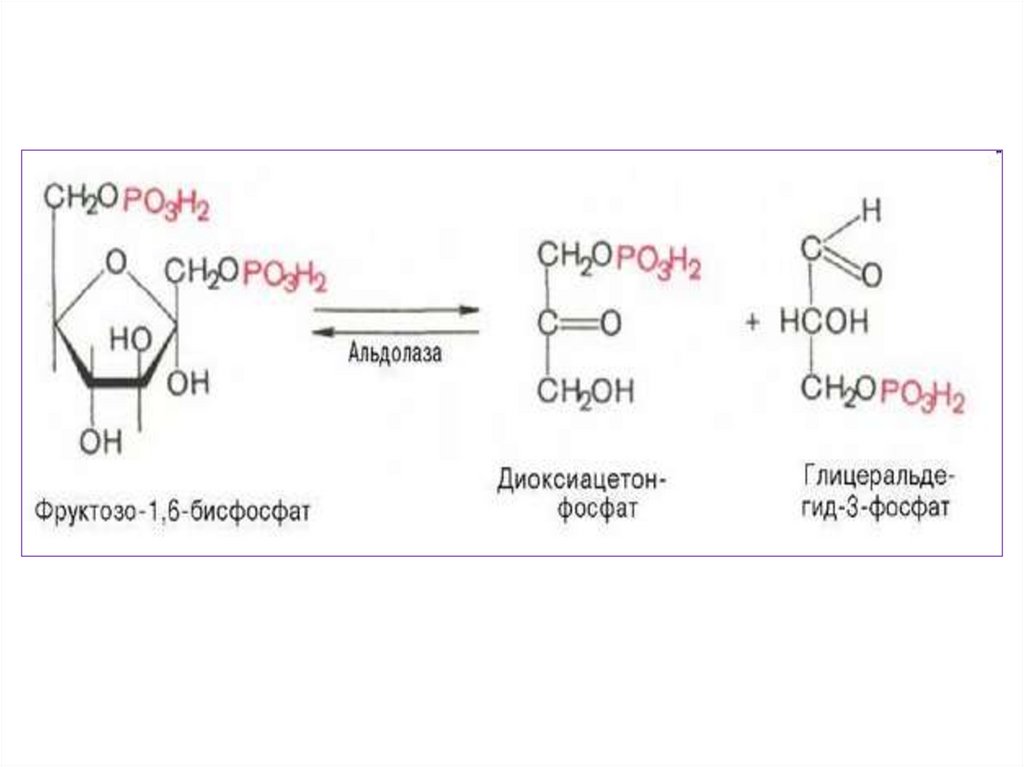
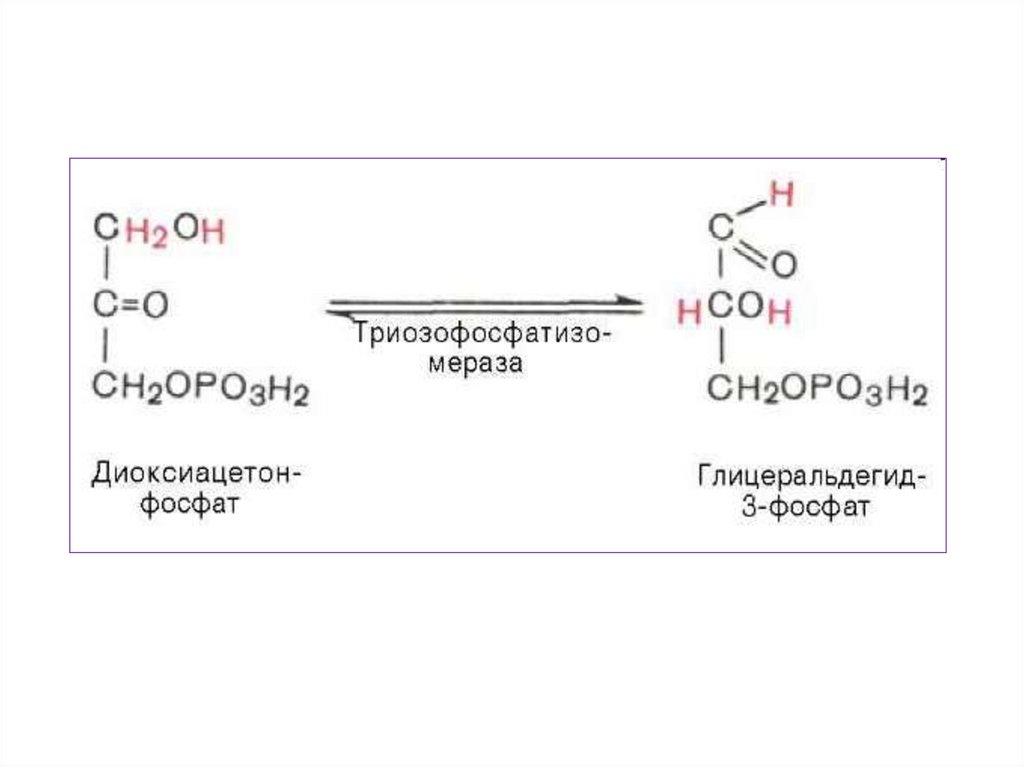
Так как из одной молекулы глюкозыобразуются две различные триозы фосфоглицероальдегид
и
фосфодиоксиацетон,
а
затем
фосфодиоксиацетон в большинстве
случаев
переходит
в
фосфоглицероальдегид
и
дальнейшим
превращениям
подвергаются две молекулы ФГА, то
общий итог превращения ФГА
следует удвоить.
53.
54.
Условно гликолиз можно разбитьна два этапа. На первом этапе,
который
заканчивается
образованием
ФДА
и
ФГА,
происходит активация глюкозы и
расщепление гексозы, на втором
этапе
синтез
АТФ
путем
субстратного фосфорилирования,
окисление
и
восстановление,
образование ПВК.
55.
56.
57.
Таккак
в
ходе
гликолиза
происходит распад гексозы на
две
триозы,
то
гликолиз
называют
дихотомическим
путем
распада.
Кроме
дихотомического
распада
глюкозы
существует
ее
апотомический
распад,
осуществляемый
в
пентознофосфатном пути.
58. Значение гликолиза
• 1. В процессе гликолиза идетмедленное освобождение энергии,
часть
которой
запасается
в
макроэргических
связях
синтезируемых молекул АТФ и
может использоваться для работы
клеток.
• 2.
Гликолиз
дает
некоторое
количество
восстановленного
НАД, который поступает затем в
ЭТЦ митохондрии, в результате
чего
клетка
получает
дополнительное количество АТФ.
59.
• 3. ПВК, образованная в ходе гликолиза, вмитохондриях в цикле трикарбоновых
кислот
дает
восстановленные
коферменты, которые пройдя через
дыхательную цепь митохондрии дают
основную часть АТФ.
• 4.
Гликолиз
обогащает
клетку
промежуточными продуктами обмена
веществ.
Например
фосфоглицероальдегид
может
превращаться в глицерин, необходимый
для
синтеза
жиров,ПВК
может
аминироваться
с
образованием
аминокислоты аланина и т.п.
60.
• 5. Гликолиз в какой-то степениспособен регулировать течение
других процессов в клетке.
Например,
образование
из
каждой молекулы гексозы двух
молекул триоз может увеличить
осмотическое давление в клетке,
что в свою очередь повлияет на
поступление воды в клетку.
61. КИСЛОРОДНЫЙ (АЭРОБНЫЙ) ЭТАП КЛЕТОЧНОГО ДЫХАНИЯ
62.
Данныйэтап
осуществляется
в
митохондриях и требует
обязательного присутствия
достаточного
количества
кислорода. Он включает в
себя следующие стадии:
63.
1. Окислительноедекарбоксилирование ПВК
2. Цикл трикарбоновых
кислот (ЦТК)
3. Окислительное
фосфорилирование в ЭТЦ
64.
Окислительноедекарбоксилирование
ПВК
И ЦТК
Окислительное
фосфорилирование
В ЭТЦ
Матрикс
митохондрий
Мембрана
крист
65. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПВК
66.
Некоторые авторы называютэту стадию аэробного этапа
клеточного
дыхания
активацией
уксусной
кислоты.
Окислительное
декарбоксилирование
ПВК
заключается в отщеплении
углекислого
газа
с
одновременным окислением
путем дегидрирования.
67.
Происходитс
участием
пируватдекарбоксилазы,
являющейся
сложной
мультифакторной системой и
включающей 3 фермента и 5
коферментов.
68.
Попавшая в матрикс митохондрииПВК реагирует с находящимся здесь
соединением - коферментом А (или
коэнзимом А: КоА, или СоА). Это
большая органическая молекула,
состоящая из остатков АДФ,
2-аминоэтандиола и пантотеоновой
кислоты (витамина В3).
69. Кофермент А
70.
При этом от ПВК отщепляетсяатом С и два атома О в виде
молекулы СО2. Связанной с СоА
остается
двухуглеродная
ацетильная
группа.
Это
соединение называется ацетилкофермент
А
(ацетил-СоА).
Одновременно
НАД+
присоединяет два атома Н.
71.
СоАSH
+
НАД+
CН3
|
C=O
|
C=O
|
HO
CH3
|
C=O
|
S-CoA
СO2
НАД • Н + Н+
72.
Ацетил-СоАНАД • Н + Н+
Матрикс
митохондрий
Мембрана
крист
митохондрий
73.
При окислении жирных кислот иаминокислот
тоже
образуется
ацетил-СоА. Таким образом в ЦТК
могут поступать молекулы ацетилСоА,
полученные
из
разных
источников. Если энергетическим
субстратом
служат
жирные
кислоты, то, начиная с концевой
карбоксильной
группы,
происходит
последовательное
отщепление двухуглеродных групп
с образованием молекул ацетилСоА.
74. ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ
75.
Циклтрикарбоновых
(более
точное название - трикарбоновых
и дикарбоновых) кислот, или цикл
лимонной кислоты, или цикл
Кребса
(назван
по
имени
открывшего его в 1937 г. немецкого
биохимика Ганса Кребса, позднее
удостоенного
за
эту
работу
Нобелевской
премии)
также
осуществляется
в
матриксе
митохондрий.
76. Ганс Адольф Кребс (1900-1981)
Немецкий биохимик. Автортрудов по обмену углеводов.
Описал основные реакции
аэробного окисления - цикл
трикарбоновых кислот (цикл
Кребса) и орнитиновый цикл
синтеза мочевины. Лауреат
Нобелевской премии 1953 г. в
области физиологии и
медицины.
77.
Ацетил-СоАсоединяется
с
четырехуглеродной
щавелевоуксусной
кислотой
(ЩУК) и передает ей свою
двухуглеродную
ацетильную
группу. В результате образуется
шестиуглеродная
лимонная
кислота. Сам СоА при этом
высвобождается
и
может
присоединять новую ацетильную
группу.
78.
Под действием ряда ферментов отлимонной кислоты поочередно
отщепляется два углеродных атома
из шести в виде молекул СО2, и
оставшееся
четырехуглеродное
соединение
после
ряда
последовательных
превращений
образует новую молекулу ЩУК.
79.
МолекулаЩУК
вновь
присоединяет ацетильную группу
от ацетил-СоА, превращаясь в
новую
молекулу
лимонной
кислоты, и цикл повторяется.
Единственное
условие
непрерывного повторения цикла
- это подача новых молекул
ацетил-СоА.
80.
На разных этапах цикла отсубстрата
отщепляются атомы
водорода
и
передаются
их
акцепторам - НАД и ФАД, которые
при этом восстанавливаются. В
каждом
обороте
цикла
восстанавливаются 3 молекулы
НАД и 1 молекула ФАД, а также
синтезируется 1 молекула АТФ.
Синтез АТФ происходит путем
субстратного фосфорилирования.
81.
ЩУК + Ацетил-СоА + АДФ + Фн ++ 3 НАД+ + ФАД
ЩУК + 2 СО2 + СоА +
АТФ
+ 3 (НАД • Н + Н+) + ФАД • Н2
+
82.
Так как в гликолизе каждаямолекула глюкозы дает 2 молекулы
ПВК, которые затем вступают в
ЦТК, то количество в нем веществ в
пересчете
на
одну
молекулу
глюкозы
удваивается.
Поэтому
каждая молекула глюкозы дает на
этой
стадии
6
молекул
восстановленного НАД, 2 молекулы
восстановленного
ФАД
и
2
молекулы АТФ.
83.
Еще2
молекулы
восстановленного
НАД
образуются в ходе окислительного
декарбоксилирования
ПВК
и
образования ацетил-СоА.
Таким образом общий итог ЦТК,
включая подготовку к нему,
составляет на одну молекулу
глюкозы - 2 АТФ, 8 (НАД • Н + Н+)
и 2 ФАД • Н2.
84.
Побочнымпродуктом
ЦТК
является СО2. При окислительном
декарбоксилировании ПВК каждая
ее молекула дает 1 молекулу СО2 и
еще 2 молекулы образуются в
самом цикле. В итоге 1 молекула
ПВК даст 3 молекулы СО2, а в
пересчете на 1 молекулу глюкозы
выделяется 6 молекул СО2.
85. Значение ЦТК в клетке
• 1) дает некоторое количествоАТФ, которое может
запасаться и расходоваться на
нужды клетки;
• 2) образует значительный
запас энергии в форме
восстановленных
коферментов;
86.
• 3) из промежуточных продуктовЦТК
синтезируется
большое
количество
веществ (например,
ацетил-СоА
предшественник
стероидных
гормонов,
холестерина и жирных кислот,
сукцинил-СоА - предшественник
гема в молекулах гемоглобина и
миоглобина и т.д.).
87.
88.
89. ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ В ЭЛЕКТРОНТРАНСПОРТНОЙ (ДЫХАТЕЛЬНОЙ) ЦЕПИ
90.
• Осуществляется на внутреннеймембране митохондрий.
• Это
превращение
энергии
восстановленных НАД и ФАД,
запасенной
в
процессе
гликолиза и цикла Кребса, в
энергию связей АТФ.
91.
Входе
этого
процесса
электроны от восстановленных
коферментов передаются по
многоступенчатой
цепи
переноса электронов (ЭТЦ) к их
конечному
акцептору
молекулярному кислороду.
92.
При переходе электронов соступени
на
ступень
высвобождается энергия, которая
служит для фосфорилирования
АДФ
в
АТФ.
Поэтому
этап
называют
окислительным
фосфорилированием. Его открыл
в 1931 г. советский биохимик
Владимир
Александрович
Энгельгардт.
93. Владимир Александрович Энгельгардт (1894-1984)
советскийбиохимик, один из
основоположников
молекулярной
биологии в СССР
94.
В ходе гликолиза из каждоймолекулы глюкозы образуется 2
молекулы АТФ, еще 2 молекулы
АТФ дает ЦТК. Основная же
масса
энергии
сосредотачивается
в
восстановленных коферментах.
Реализуется эта энергия при
переходе
электронов
и
протонов по ЭТЦ.
95.
Компоненты ЭТЦ находятся навнутренней
мембране
митохондрий.
Переносчики
представляют собой ферменты,
коферментами которых служат
НАД+, ФАД и ФМН. Затем
в
цепи
располагается
кофермент Q (убихинон), а
завешается ЭТЦ различными
цитохромами.
96.
КоферментQ
переносит
электроны и протоны, а все
остальные
переносчики
только
электроны.
Цитохромы
это
гемисодержащие
белки
(содержат гем), а цитохромы
а содержат медь.
97.
Известно около 20 различныхцитохромов,
делящихся
на
4
главных класса - a, b, c, d.
Поступающие
к
цитохромам
электроны
восстанавливают
железо,
переводя
его
из
трехзарядного
состояния
в
двухзарядное.
Конечным
акцептором электронов служит
кислород.
98.
Кофермент Q(убихинон)
Простетическая
группа цитохрома b
99.
100.
Каждыйвосстановленный
кофермент передает в ЭТЦ на
молекулу переносчика два атома
водорода и возвращается обратно.
Молекулы,
принимающие
водородные атомы, находятся на
внутренней стороне внутренней
мембраны (мембраны крист). Они
переносят атомы водорода через
мембрану
на
ее
наружную
поверхность и передают их второму
переносчику.
101.
Этотпереносчик
может
передавать в обратную сторону
только электроны,
а
протоны
поступают
в
протонный
резервуар
(Н+резервуар),
расположенный
около наружной поверхности
мембраны
крист
(в
межмембранном пространстве
митохондрии).
102.
Теперьперенесенные
на
внутреннюю поверхность мембраны
электроны
уже
не
могут
соединиться с протонами. Здесь их
принимает следующий переносчик,
способный переносить целые атомы
водорода.
Он
получает
их
в
результате
присоединения
полученных электронов к новым
протонам, которые образуются при
диссоциации воды.
103.
• Электроны совершают еще рядпереходов через мембрану в
обоих направлениях, причем
только
переход
изнутри
наружу
происходит
в
сопровождении протонов. Все
эти переходы поставляют в
общей сложности 64 протона
на каждую молекулу глюкозы.
104.
Конечнымакцептором
электронов является кислород,
который принимает электроны
и протоны (образовавшиеся
опять-таки
за
счет
диссоциации воды) и образует
новую молекулу воды
1/2 О2 + 2е + 2Н+
Н2О
105.
Энергиязапасенных
протонов Н+- резервуара
используется для синтеза
АТФ.
Во
внутренней
мембране
митохондрий
есть каналы, через которые
протоны возвращаются из
протонного
резервуара
обратно в матрикс.
106.
Каждый такой канал содержитбольшой ферментный комплекс АТФ-синтетазу, которая состоит из
двух
основных
компонентов
(факторов) – F0 и F1.
F0
пронизывает
мембрану
насквозь.
F1
большая
сферическая частица из девяти
субъединиц, соединенная с
F0частицей со стороны матрикса.
107.
Визолированной
форме
F1компонент
не
может
синтезировать АТФ из АДФ и
неорганического
фосфата,
но
способен гидролизовать АТФ до
АДФ, то есть работает как АТФ-аза.
Но обычная его функция в
контакте с фактором F0 обратная синтез АТФ.
108.
1 молекулавосстановленного
НАД
3 молекулы
АТФ
1 молекула
восстановленного
ФАД
2 молекулы
АТФ
109.
Протоныпреодолевают
электрохимический градиент,
существующий
между
наружной
и
внутренней
сторонами мембраны крист,
когда проходят пассивным
путем через частицы F0 и F1.
110.
• Электрохимический градиентслагается
из
двух
компонентов: химического и
электрического.
• Химический
градиент
обусловлен
разностью
концентраций протонов по
разные стороны мембраны.
111.
• Электрический градиент, илиразность потенциалов
возникает за счет накопления
противоположных зарядов по
разные стороны внутренней
мембраны.
112.
• Наружная сторона заряжаетсяположительно
вследствие
накопления
протонов,
а
внутренняя
поверхность,
обращенная
к
матриксу,
приобретает
отрицательный
заряд,
обусловленный
присоединением электронов к
молекулярному кислороду
О2 + е
О2-
113.
• Свободная энергия, котораявыделяется при прохождении
протонов через канал
АТФ-синтетазы,
идет
на
синтез АТФ. Синтез АТФ на
внутренней
мембране
митохондрий
называют
сопряженным мембранным
окислительным
фосфорилированием.
114.
115.
116.
117. Строение и работа митохондрий
а
митохондрии
(указаны
стрелкой), видимые в световом
микроскопе; б - ультраструктура
митохондрий:
1
митохондриальный матрикс, 2 внутренняя
митохондриальная
мембрана, 3 - межмембранное
пространство,
4
внешняя
митохондриальная мембрана; в общая схема функционирования
митохондрий:
при
переносе
электронов в цепи окисления в
межмембранном
пространстве
накапливаются протоны и при
достижении
определенного
потенциала
возвращаются
в
матрикс; энергия этого потенциала
тратится на синтез АТФ
118.
Механизм окислительногофосфорилирования,
названный
хемосмосом,
был предложен английским
биохимиком
Питером
Митчеллом, удостоенным в
1978
г.
Нобелевской
премии.
119. Питер Митчелл (1920-1992)
Английскийбиохимик, автор
теории
хемосмоса,
лауреат
Нобелевской
премии 1978 г.
120.
121.
Путь электронов по ЭТЦ отвосстановленного НАД
АТФ
НАД . Н+Н+
АТФ
е
НАД+
Дегидрогеназа
е
ФМН
CoQ
е
цит b
АТФ
е
цит с
е
е
цит а + цит а3
(цитохромоксидаза)
е
O2
цит с1
е
122. Путь электронов по ЭТЦ от восстановленного ФАД
АТФФАД . Н2
АТФ
е
е
е
ФАД
цит а + цит а3
CoQ
(цитохромоксидаза)
е
цит b
е
O2
е
цит с1
е
цит с
123.
124.
125.
• В пересчете на одну молекулуглюкозы в ЭТЦ образуется 34
молекулы АТФ.
Суммарное уравнение
окислительного
фосфорилирования в ЭТЦ можно
выразить следующим образом
12 Н2 + 6 О2 + 34 АДФ + 34Фн
12 Н2О
+
34 АТФ
126.
Таким образом, в кислородном(аэробном)
этапе
клеточного
дыхания образуется 36 молекул
АТФ. Из них 2 молекулы дает ЦТК и
34
молекулы
окислительное
фосфорилирование в ЭТЦ. В ходе
гликолиза образуется
2
молекулы
АТФ.
Поэтому
суммарный
итог
клеточного
дыхания составит
38 молекул АТФ.
127.
Гликолиз
2 АТФ
2 НАД • Н+ 2Н+
в ЭТЦ 3 х 2 = 6 АТФ | 2 АТФ + 6 АТФ = 8АТФ
Ц Т К (вместе с подготовкой)
2 АТФ
8 НАД • Н + 8Н+
в ЭТЦ 8 х 3 = 24 АТФ
2 ФАД • Н2
в ЭТЦ 2 х 2 = 4 АТФ | 2 АТФ + 24 АТФ + 4 АТФ
= 30 АТФ
Итого:
38 АТФ
128.
• Суммарное уравнение клеточногодыхания выглядит следующим
образом:
С6Н12О6 + 6 О2 + 38 АДФ + 38 Фн
6 СО2 + 6 Н2О + 38 АТФ
• При этом в виде АТФ запасается 55
% энергии, освобождаемой при
расщеплении глюкозы, а 45 %
рассеивается в виде тепла.