





































 biology
biologySimilar presentations:









")
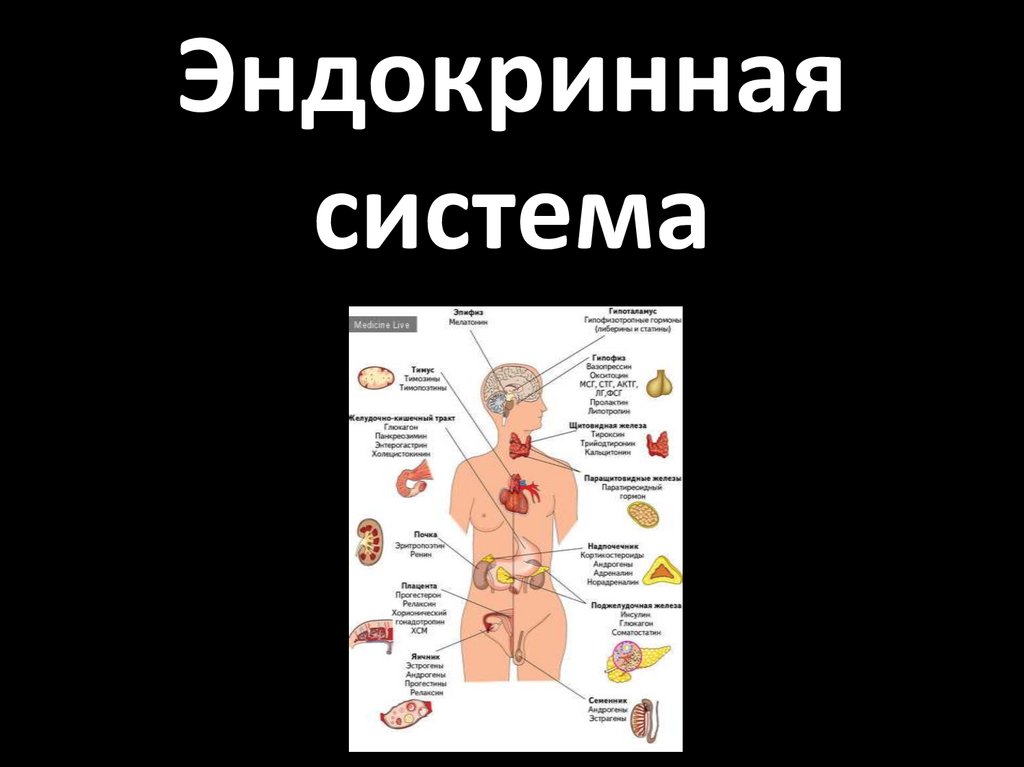
Эндокринная система
1.
Эндокриннаясистема
2.
3.
4.
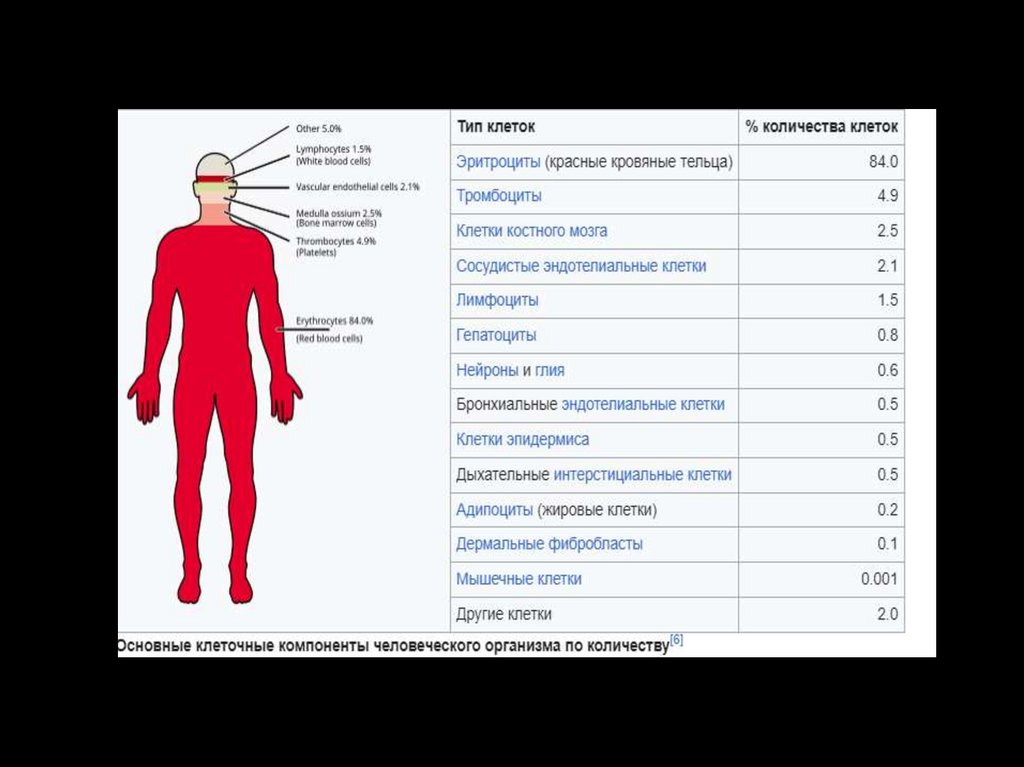
Регуляторные системыОбщее количество клеток в организме – 35-100 трлн. (10¹³)
Общее количество нейронов – 100 млрд (109)
Общее количество иммунных клеток – 1-2 трлн. ( 1010)
Общее количество эндокринных клеток - ?
5.
ЭндокринологияА. Бертольд
Начало
этой
науке
положили
опыты
немецкого
физиолога
Адольфа
Бертольда,
которому в 1849 году
удалось установить, что
при
пересадке
кастрированному петуху в
брюшную
полость
семенников другого петуха
у первого исчезают все
последствия кастрации.
Профессор
БроунСекар создал учение о
железах
внутренней
Броун-Секар секреции,
превратившееся
в
самостоятельную науку —
эндокринологию.
6.
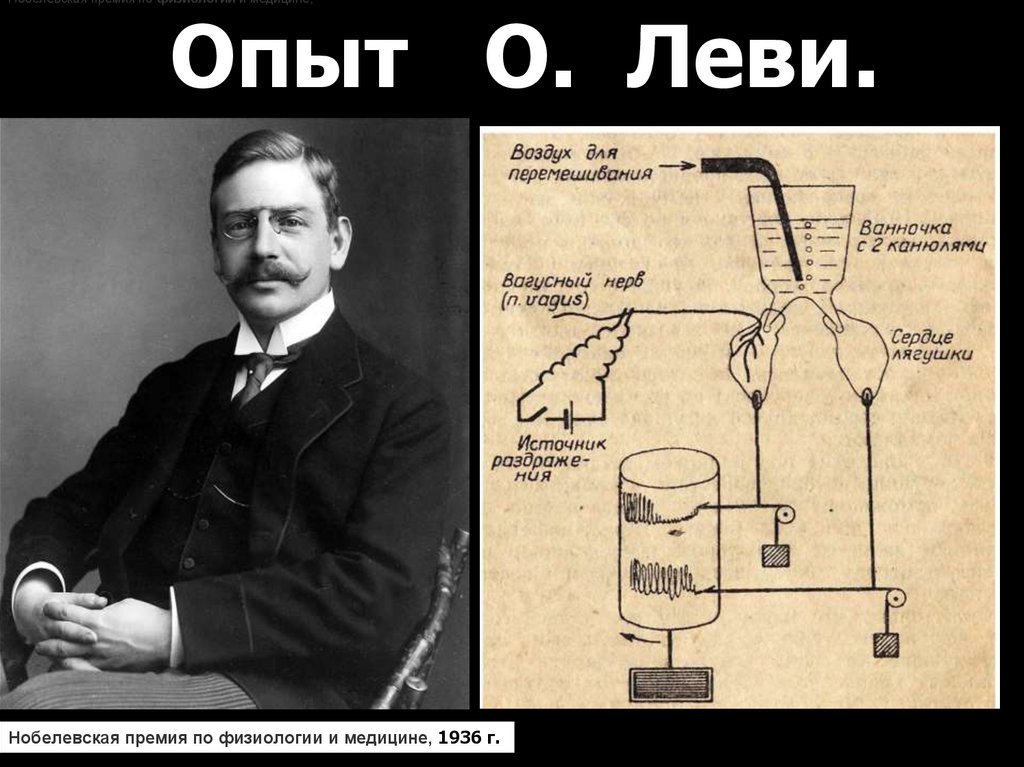
7.
Нобелевская премия по физиологии и медицине,Опыт О. Леви.
Нобелевская премия по физиологии и медицине, 1936 г.
8.
Что такое гормоны?Гормоны
- (от греч. hormáo - привожу в
движение,
побуждаю),
биологически
активные
вещества, вырабатываемые железами внутренней
секреции, и выделяемые ими непосредственно в
кровь.
Термин «гормоны» введён англ. физиологами У.
Бейлиссом и Э. Старлингом в 1902 г.
Гормоны разносятся кровью и влияют на
деятельность органов, изменяя физиологические и
биохимические реакции путём активации или
торможения ферментативных процессов.
Известно более 60 различных
гормонов,
выделяемых
эндокринными
железам
млекопитающих и человека.
9.
Физиологическое действиегормонов направлено на:
• 1)
обеспечение
гуморальной,
т.е.
осуществляемой
через
кровь,
регуляции
биологических процессов.
• 2) поддержание целостности и постоянства
внутренней среды.
• 3) регуляцию процессов роста, созревания и
репродукции.
10.
Эффекты действия гормонов1. Метаболическое
действие
–
изменяют
проницаемость мембран клетки и активность
транспортных систем; активность ферментов,
индуцируют
или
подавляют
процессы
транскрипции.
2. Морфологическое
действие
–
стимуляция
формообразовательных
процессов,
дифференцировки, роста.
3. Кинетическое
действие
–
запускают
определённую функцию исполнительных органов
(например, окситоцин вызывает сокращение матки).
11.
4.Корригирующее
действие
–
изменяет
интенсивность функции органа (нормализующий
эффект гормонов, когда их влияние направлено на
восстановление измененного или даже нарушенного
процесса).
5. Реактогенное действие – способность гормона
менять реактивность ткани к действию гормонов или
медиаторов.
12.
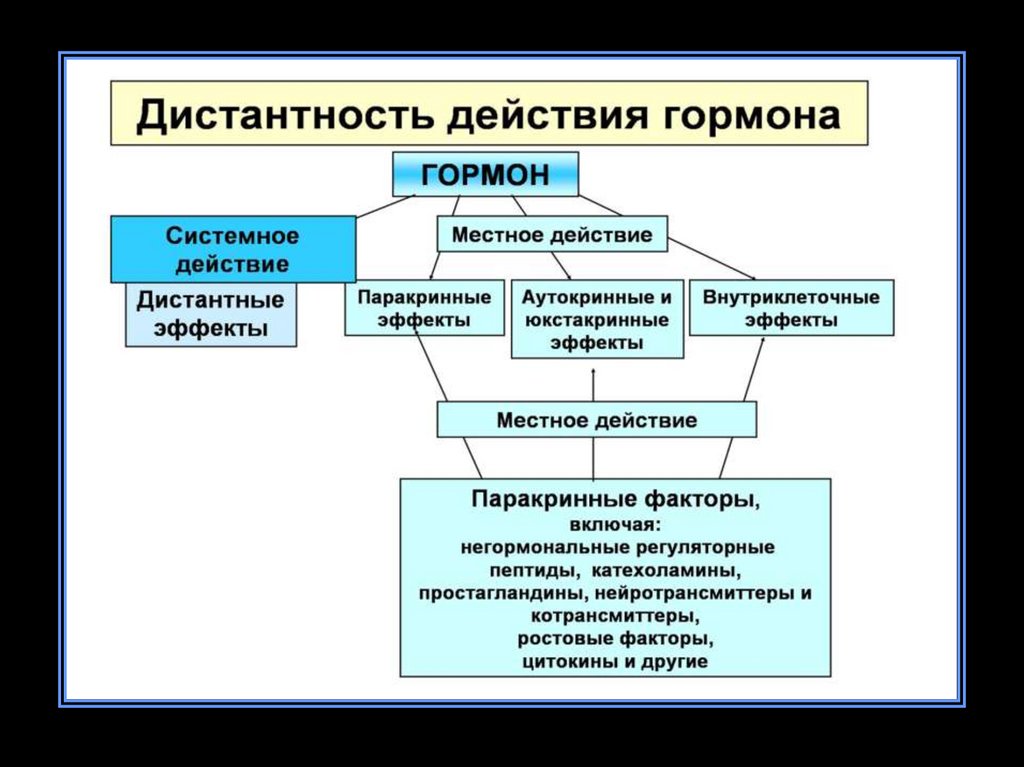
Классическим гормонам присущ рядпризнаков:
• Дистантность действия – синтез в железах внутренней
секреции, а регуляция отдаленных тканей.
• Избирательность действия.
• Строгая специфичность действия.
• Кратковременность действия.
• Действуют в очень низких концентрациях, под контролем
ЦНС и регуляция их действия осуществляется по типу
обратной связи.
• Действуют опосредованно через белковые рецепторы и
ферментативные системы.
13.
Гормональный балансВ нормальном состоянии существует строгий баланс
между активностью эндокринных желез, состоянием
нервной системы и ответом тканей-мишеней.
Любое нарушение в каждом из этих звеньев быстро
приводит к отклонениям от нормы.
Избыточная или недостаточная продукция гормонов
служит причиной различных заболеваний.
14.
Регуляция уровня гормонов ворганизме
• Изменение концентрации метаболитов в клеткахмишенях по механизму отрицательной обратной
связи подавляет синтез гормонов, действуя либо
на эндокринные железы, либо на гипоталамус.
• Существуют эндокринные железы для которых
отсутствует регуляция тропными гормонами –
пара-щитовидная
железа,
ренинальдостероновая система и поджелудочная
железа.
Они контролируются нервными влияниями или
концентрацией определенных веществ в крови.
15.
Организация нервно-гормональнойрегуляции
• Существует строгая
иерархия или
соподчиненность
гормонов.
• Поддержание уровня
гормонов в организме в
большинстве случаев
обеспечивает механизм
отрицательной обратной
связи.
16.
Транспорт гормоновосуществляется кровью в
комплексе с белками
Часть этих белков - специфические транспортные протеины
(например, транскортин - связывает гормоны коры
надпочечников), часть - неспецифические (например, γглобулины).
Образование комплексов - процесс обратимый.
Часть гормонов связана в крови с форменными элементами,
в частности с эритроцитами.
17.
Образование связанной формыгормонов :
1. предохраняет организм от избыточного накопления
в крови свободных гормонов (и, следовательно, влияния на
ткани).
2. связанная форма гормона является его
физиологическим резервом.
3. связывание с белками способствует защите
гормона от разрушения ферментами, т.е. продлевает его
жизнь.
4. препятствует фильтрации гормонов через
почечные клубочки и тем самым удерживает их в организме.
18.
Классификация эндокринныхструктур:
I. Центральные регуляторные образования эндокринной системы:
– гипоталамус (нейросекреторные ядра);
– гипофиз (аденогипофиз и нейрогипофиз);
– эпифиз.
II. Периферические эндокринные железы:
– щитовидная железа;
– околощитовидные железы;
– надпочечники (корковое и мозговое вещество).
III. Органы, объединяющие эндокринные и неэндокринные
функции:
– гонады (половые железы - семенники и яичники);
– плацента;
– поджелудочная железа.
IV. Одиночные гормонпродуцирующие клетки, апудоциты
(АПУД-система).
Y. Гормоны, продуцируемые скелетными мышцами:
- миокины
19.
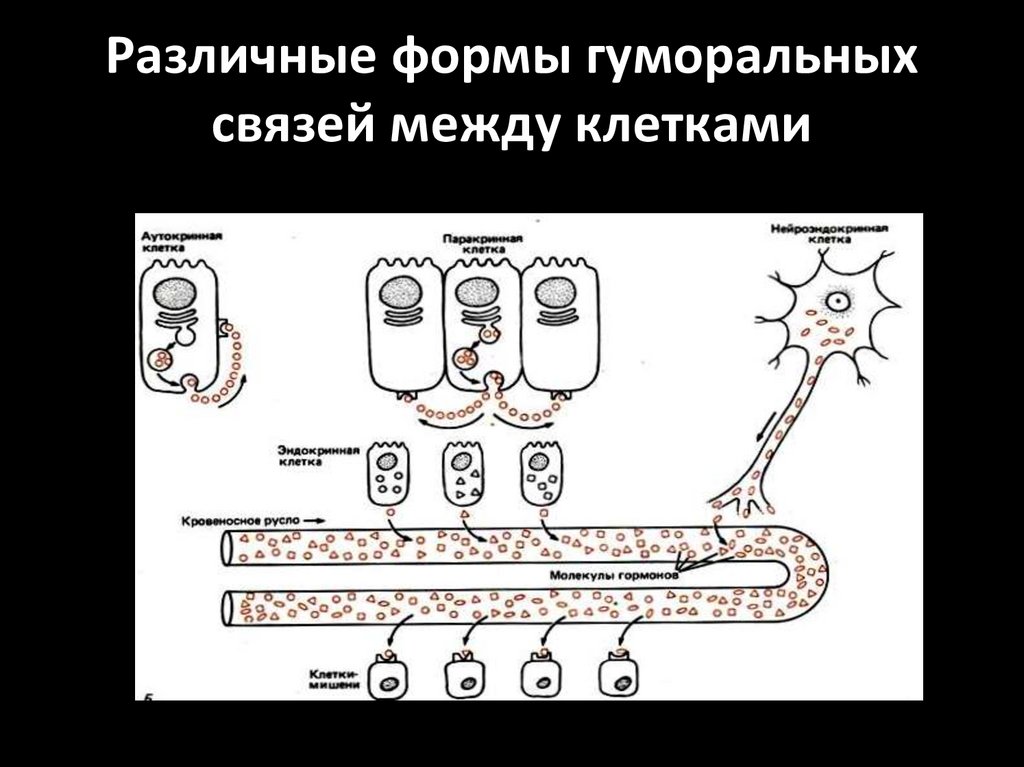
Различные формы гуморальныхсвязей между клетками
20.
21.
Особенности группированияразных типов эндокринных клеток:
• Эндокринные клетки неэндокринных
органов: энтероциты ЖКТ, эндокринные
нейроны гипоталамуса.
• Эндокринные клетки желез смешанной
секреции: семенников, яичников,
поджелудочной железы.
• Эндокринные клетки эндокринных желёз:
надпочечников, щитовидной железы,
паращитовидных желез, аденогипофиза,
эпифиза.
22.
Биологический смыслинтеграции эндокринных
клеток в эндокринную железу:
создание более совершенного
механизма управления потоками
информации и эффекторными
функциями.
23.
Классификация гормонов• по химическому строению;
• по биологическим функциям;
• по механизму действия;
24.
1. По химической структурегормоны, подразделяют на:
1. пептиды,
2. производные аминокислот,
3. стероиды,
4. производные арахидоновой
кислоты.
25.
Пептидные гормонысекретируются путем экзоцитоза,
- относятся к полярным веществам, поэтому не
могут проникать через мембраны,
- рецепторы встроены в мембрану клетки–
мишени, а передачу сигнала к внутриклеточным
структурам
осуществляют
вторичные
посредники.
-
- Гормоны гипоталамуса; гормоны гипофиза;
гормоны поджелудочной железы - инсулин,
глюкагон;
гормоны
щитовидной
и
паращитовидной
желез
–
соответственно
кальцитонин и паратгормон.
26.
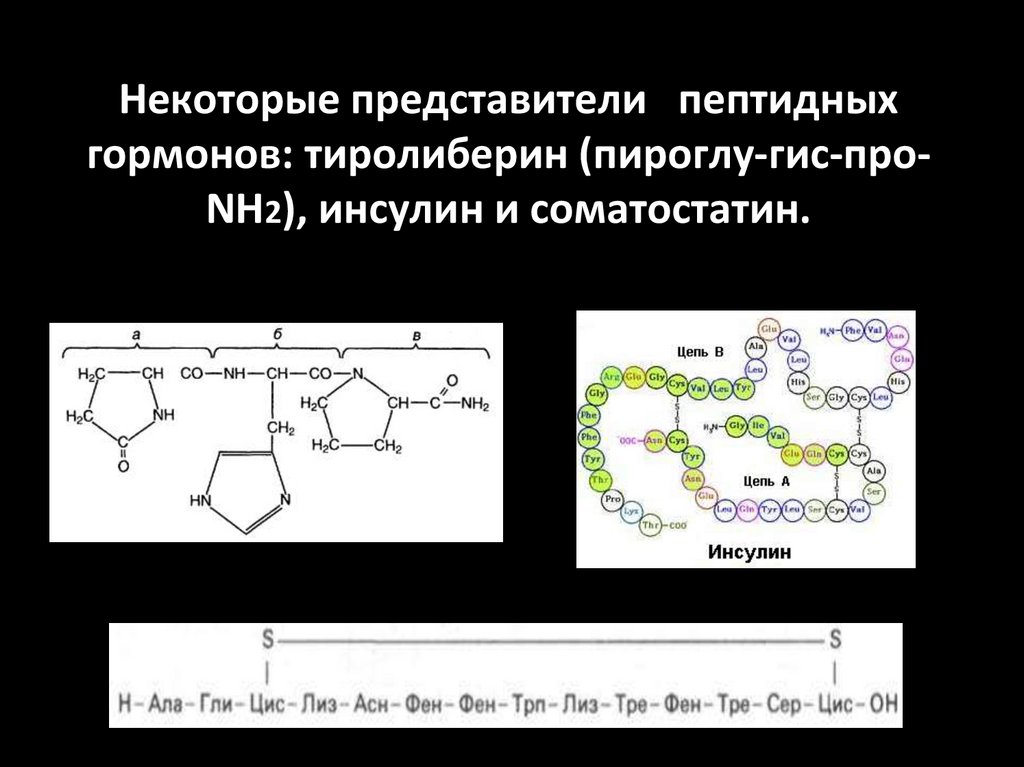
Некоторые представители пептидныхгормонов: тиролиберин (пироглу-гис-проNН2), инсулин и соматостатин.
27.
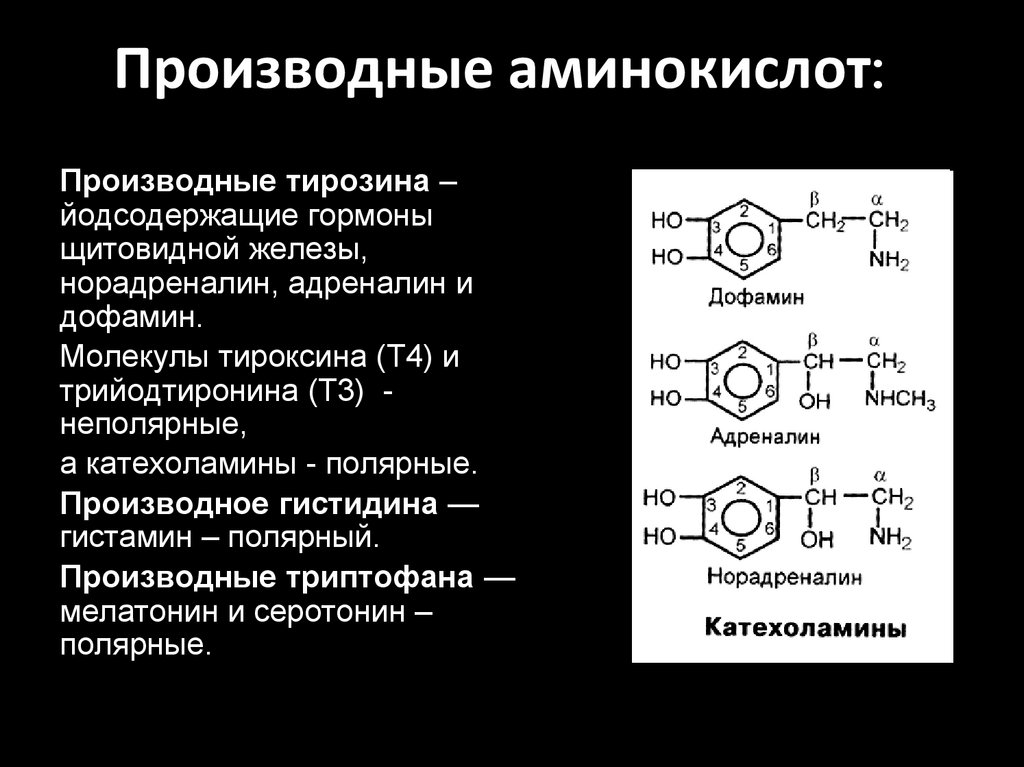
Производные аминокислот:Производные тирозина –
йодсодержащие гормоны
щитовидной железы,
норадреналин, адреналин и
дофамин.
Молекулы тироксина (T4) и
трийодтиронина (Т3) неполярные,
а катехоламины - полярные.
Производное гистидина —
гистамин – полярный.
Производные триптофана —
мелатонин и серотонин –
полярные.
28.
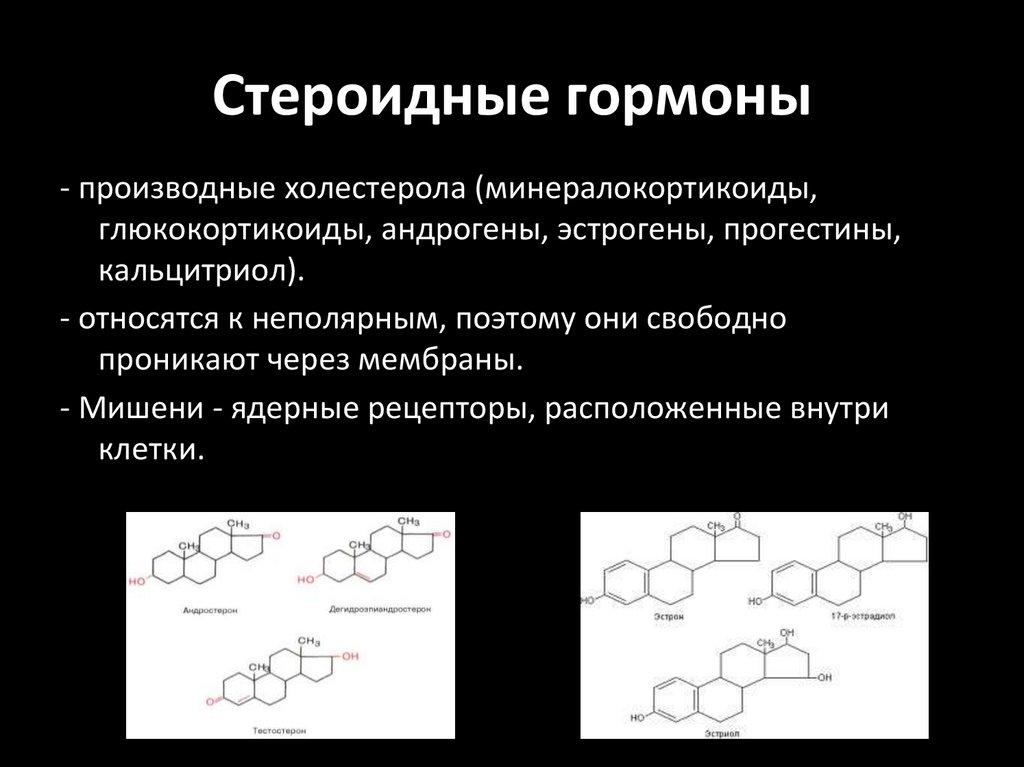
Стероидные гормоны- производные холестерола (минералокортикоиды,
глюкокортикоиды, андрогены, эстрогены, прогестины,
кальцитриол).
- относятся к неполярным, поэтому они свободно
проникают через мембраны.
- Мишени - ядерные рецепторы, расположенные внутри
клетки.
29.
Производные арахидоновойкислоты (эйкозаноиды)
Предшественником всех эйкозаноидов является
арахидоновая кислота.
Они делятся на 3 группы:
простагландины,
лейкотриены,
тромбоксаны.
• Эйкозаноиды - локальные гормоны
— широко распространенная группа сигнальных
веществ, которые образуются почти во всех ,клетках
организма, действуют как локальные биорегуляторы
путем связывания с мембранными рецепторами в
непосредственной близости от места их синтеза как на
синтезирующие их клетки (аутокринное действие), так
и на соседние клетки (паракринное действие).
30.
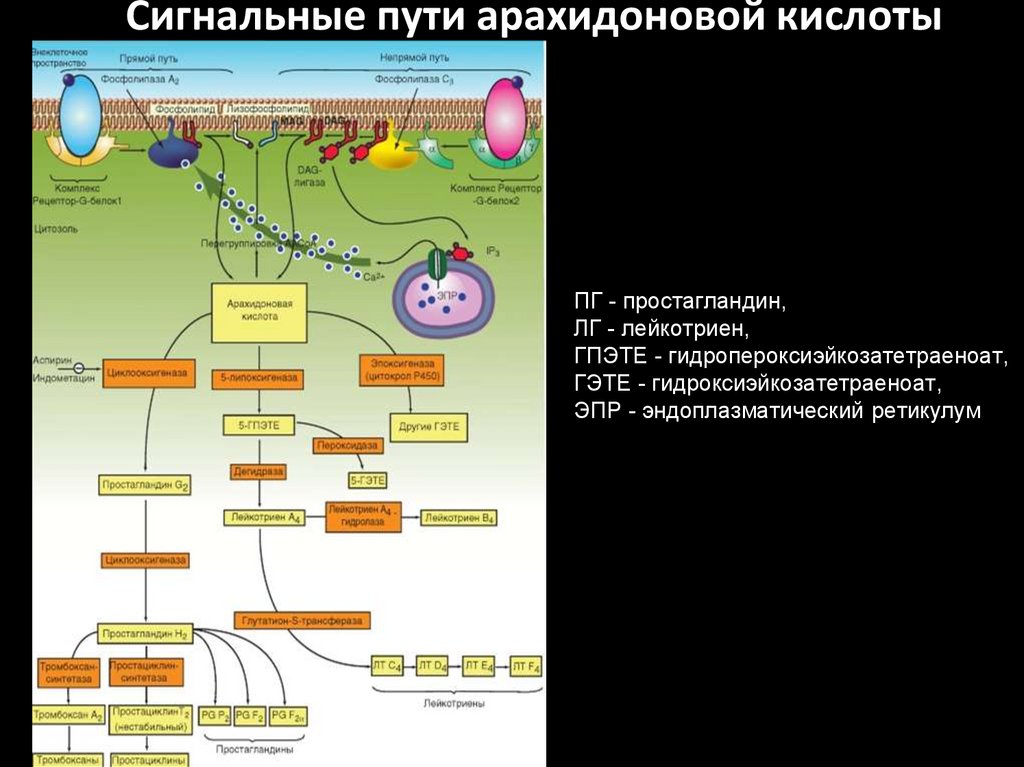
Сигнальные пути арахидоновой кислотыПГ - простагландин,
ЛГ - лейкотриен,
ГПЭТЕ - гидропероксиэйкозатетраеноат,
ГЭТЕ - гидроксиэйкозатетраеноат,
ЭПР - эндоплазматический ретикулум
31.
Характеристика разных группэйкозаноидов
Простагландины (Pg) — синтезируются практически
во всех клетках, кроме эритроцитов и лимфоцитов.
Выделяют следующие группы простагландинов: A,
B, C, D, E, F.
Функции простагландинов сводятся к изменению
тонуса гладких мышц: бронхов, мочеполовой и
сосудистой систем, желудочно-кишечного тракта; при
этом направленность изменений различна в
зависимости от типа простагландинов и условий. Они
также влияют на температуру тела.
Простациклины являются подвидом простагландинов
(PgI),
но
дополнительно
обладают
особой
функцией — ингибируют агрегацию тромбоцитов и
обусловливают вазодилатацию. Особенно активно
синтезируются в эндотелии сосудов миокарда, матки,
слизистой желудка.
32.
Тромбоксаны и лейкотриены• Тромбоксаны (Tx) образуются в тромбоцитах,
стимулируют их агрегацию и вызывают сужение
мелких сосудов.
• Лейкотриены (Lt) активно синтезируются в лейкоцитах,
в клетках лёгких, селезёнки, мозга, сердца.
Выделяют 6 типов лейкотриенов: A, B, C, D, E, F.
В лейкоцитах они стимулируют подвижность,
хемотаксис и миграцию в очаг воспаления.
Кроме этого, вызывают сокращение мускулатуры
бронхов в дозах в 100—1000 раз меньших, чем
гистамин.
33.
2. биологические функции гормоновГОРМОНЫ
Тропные гормоны гипофиза,
либерины и статины гипоталамуса.
РЕГУЛИРУЕМЫЕ ПРОЦЕССЫ
Синтез и секреция гормонов
эндокринных желез.
Альдостерон, вазопрессин.
Водно-солевой обмен.
Паратгормон, кальцитонин,
кальцитриол.
Обмен кальция и фосфатов.
Эстрогены, андрогены, гонадотропные
гормоны.
Репродуктивная функция.
Инсулин, глюкагон, адреналин,
кортизол,тироксин,соматотропин.
Обмен углеводов, липидов,
аминокислот.
34.
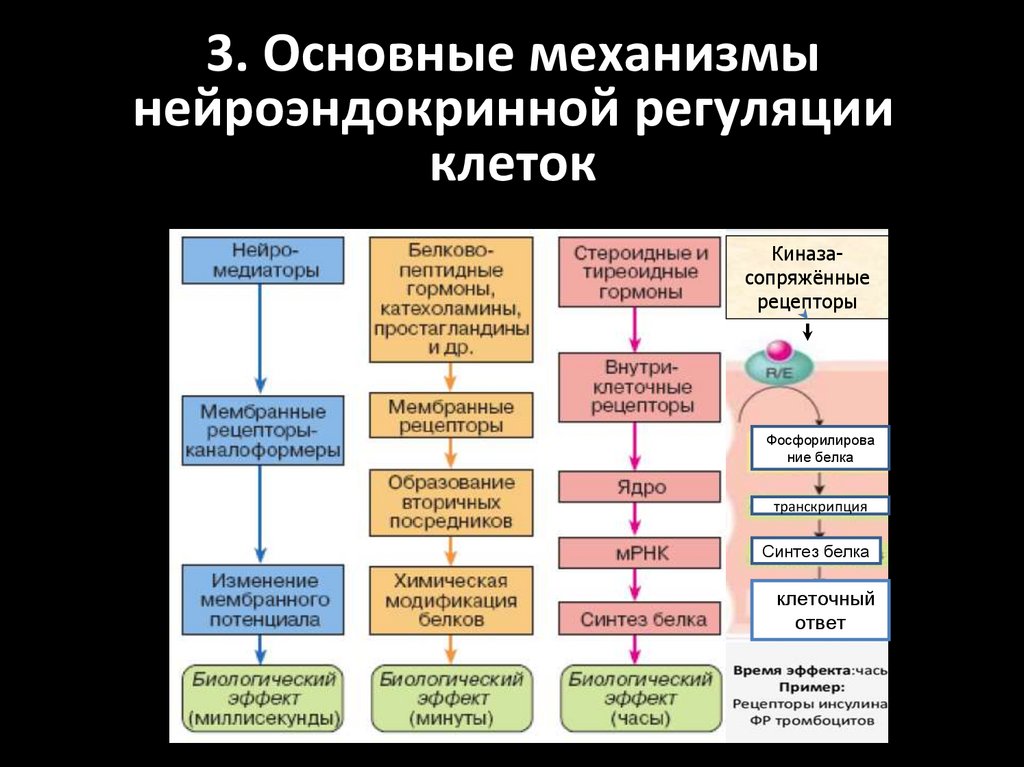
3. Основные механизмынейроэндокринной регуляции
клеток
Киназасопряжённые
рецепторы
Фосфорилирова
ние белка
транскрипция
Синтез белка
Кклеточный
ответ
35.
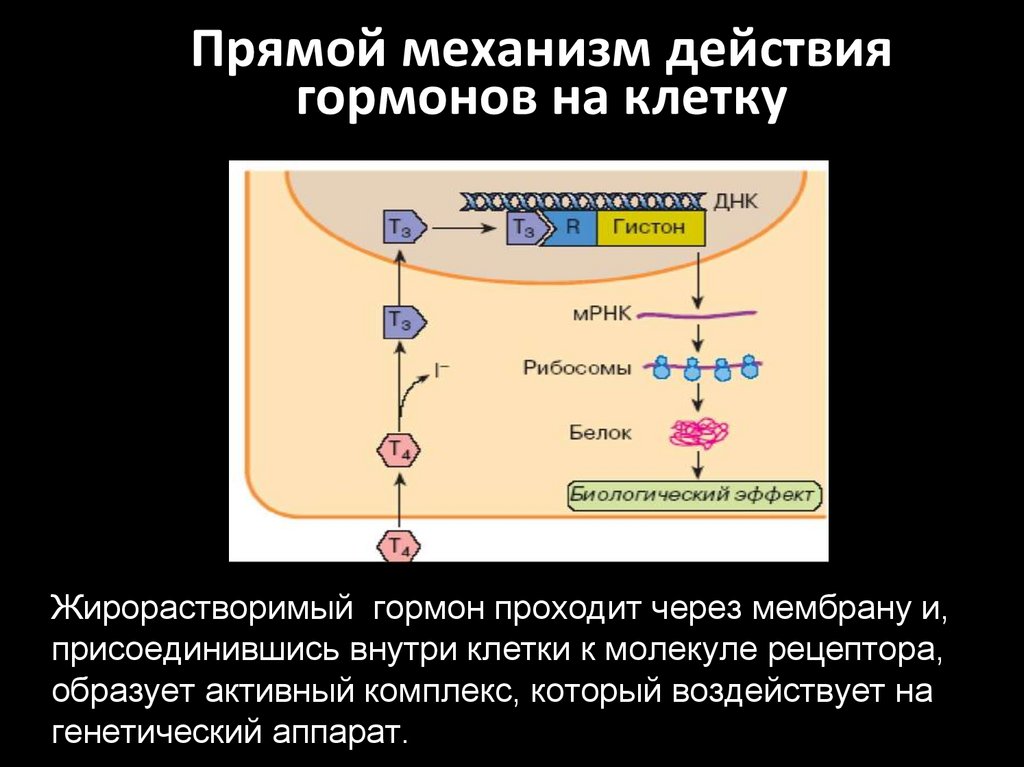
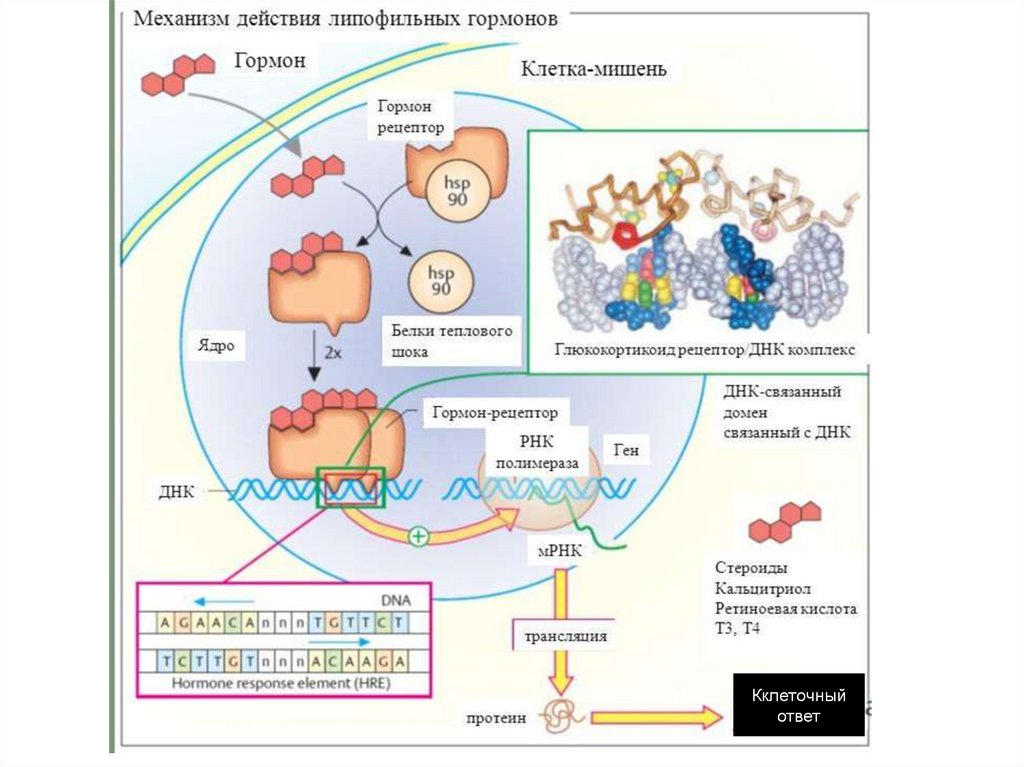
Прямой механизм действиягормонов на клетку
Жирорастворимый гормон проходит через мембрану и,
присоединившись внутри клетки к молекуле рецептора,
образует активный комплекс, который воздействует на
генетический аппарат.
36.
37.
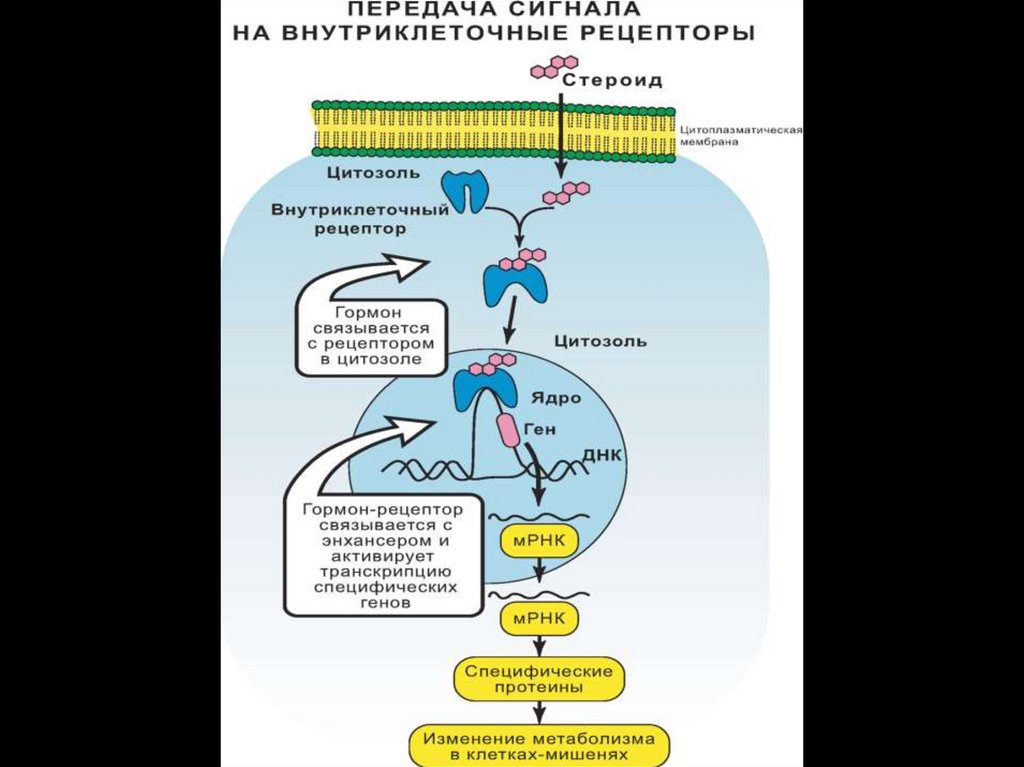
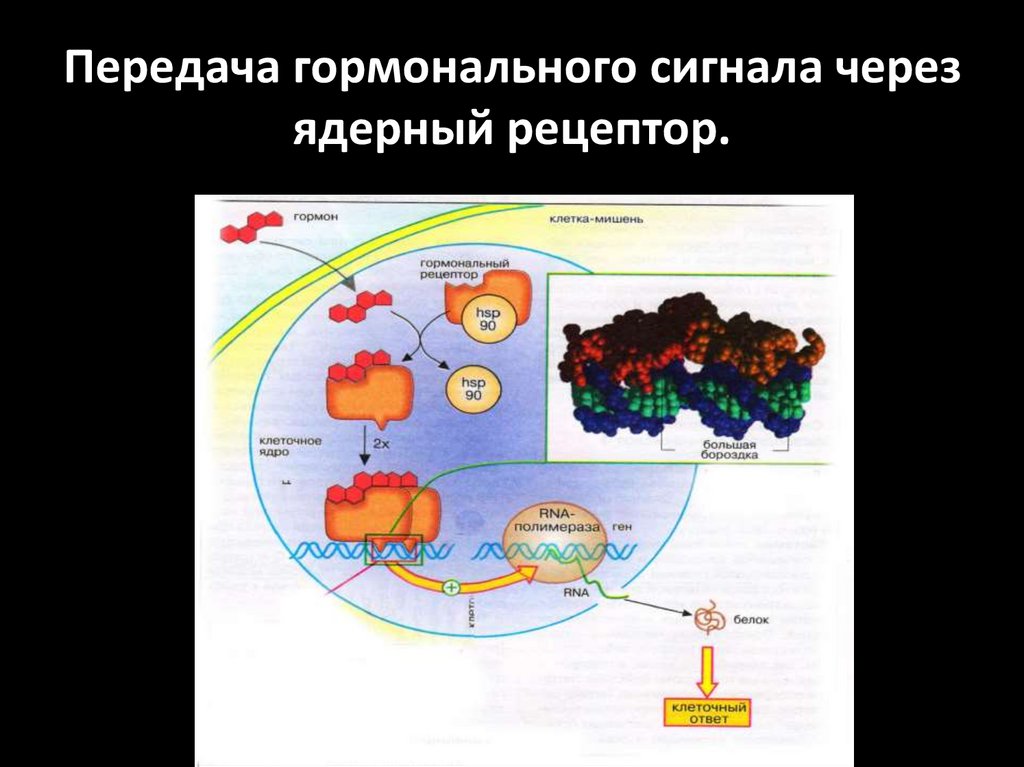
Ядерные рецепторыЯдерные рецепторы - это ДНК-связывающие факторы
транскрипции, локализованные в ядре.
В отсутствие лиганда основная часть рецепторов
стероидов находиться в цитоплазме.
Рецепторы стероидов связываются в цитоплазме с
белками
теплового
шока
(Hsp),
которые
препятствуют транспорту рецептора через ядерную
мембрану.
Активность
ядерных
рецепторов
регулируется
фосфорилированием.
38.
Передача гормонального сигнала черезядерный рецептор.
39.
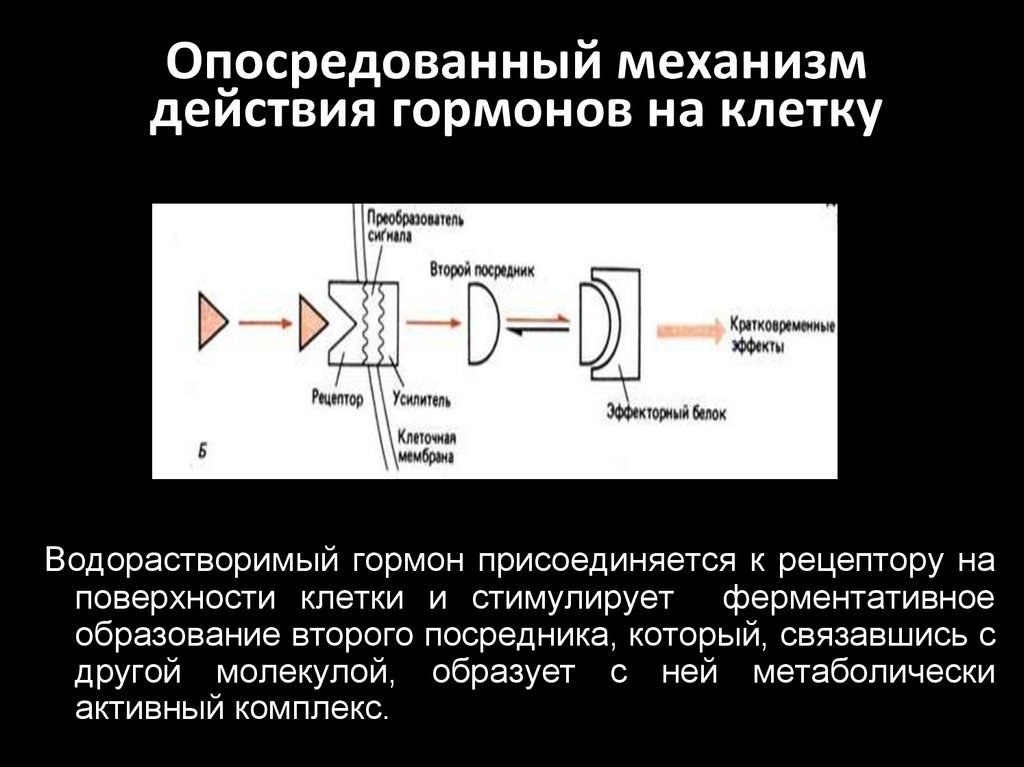
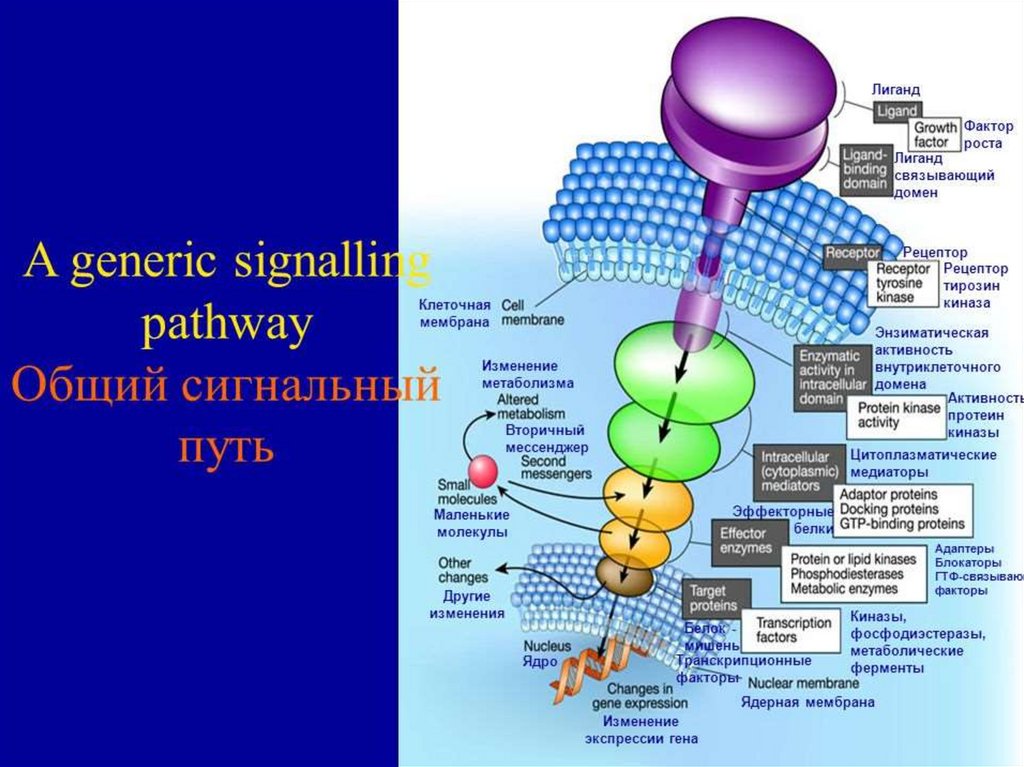
Опосредованный механизмдействия гормонов на клетку
Водорастворимый гормон присоединяется к рецептору на
поверхности клетки и стимулирует ферментативное
образование второго посредника, который, связавшись с
другой молекулой, образует с ней метаболически
активный комплекс.
40.
Характерной особенностью действия этихгормонов является быстрый эффект –
обусловленный
активацией
уже
существующих ферментов и других белков.
41.
Вторичныемессенджеры
42.
Требования предъявляемыевторичному мессенджеру:
1. небольшая молекулярная масса (чтобы с высокой
скоростью диффундировать в цитоплазме),
2. должен легко проникать через мембрану,
3. должен быстро расщепляться, а в случае Са2+
откачиваться.
43.
КЛАССИЧЕСКИЕ ВТОРИЧНЫЕМЕССЕНДЖЕРЫ
• цАМФ (циклический аденозинмонофосфат);
• цГМФ (циклический гуанозинмонофосфат);
• ИФ3 (инозитолтрифосфат);
• ДИАЦИЛГЛИЦЕРОЛ (ДАГ)
• ИОНЫ КАЛЬЦИЯ (Са2+)
• ОКСИД АЗОТА (NO)
• МОНООКСИД УГЛЕРОДА (СО)
• СУЛЬФИД ВОДОРОДА (SH)
44.
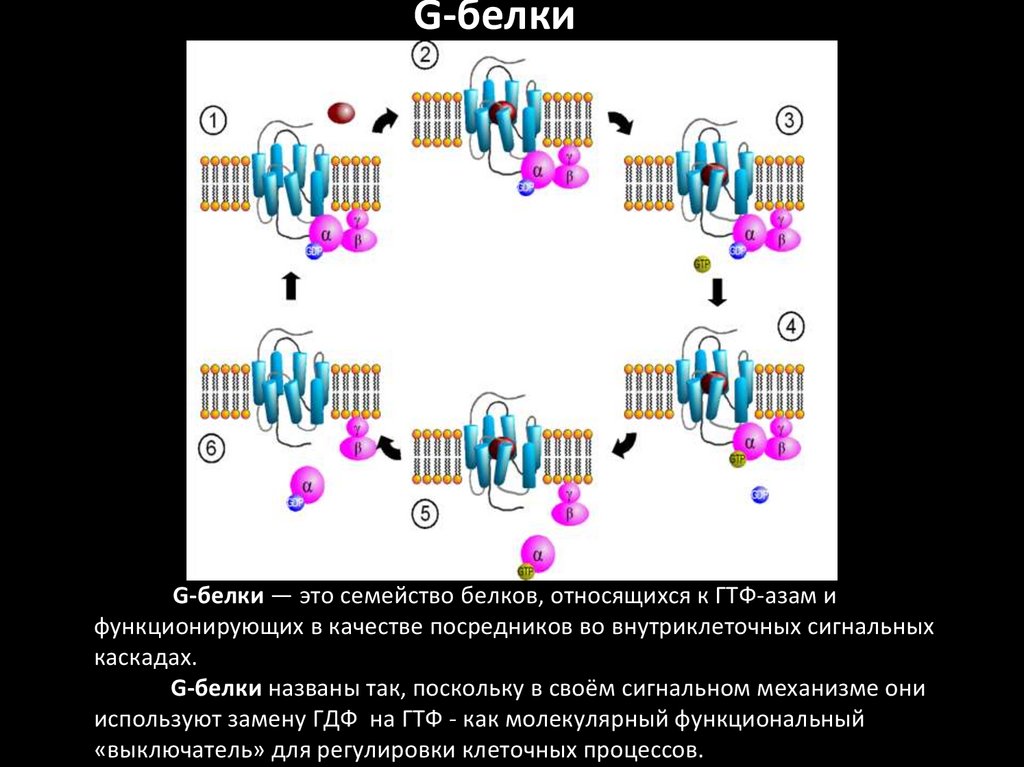
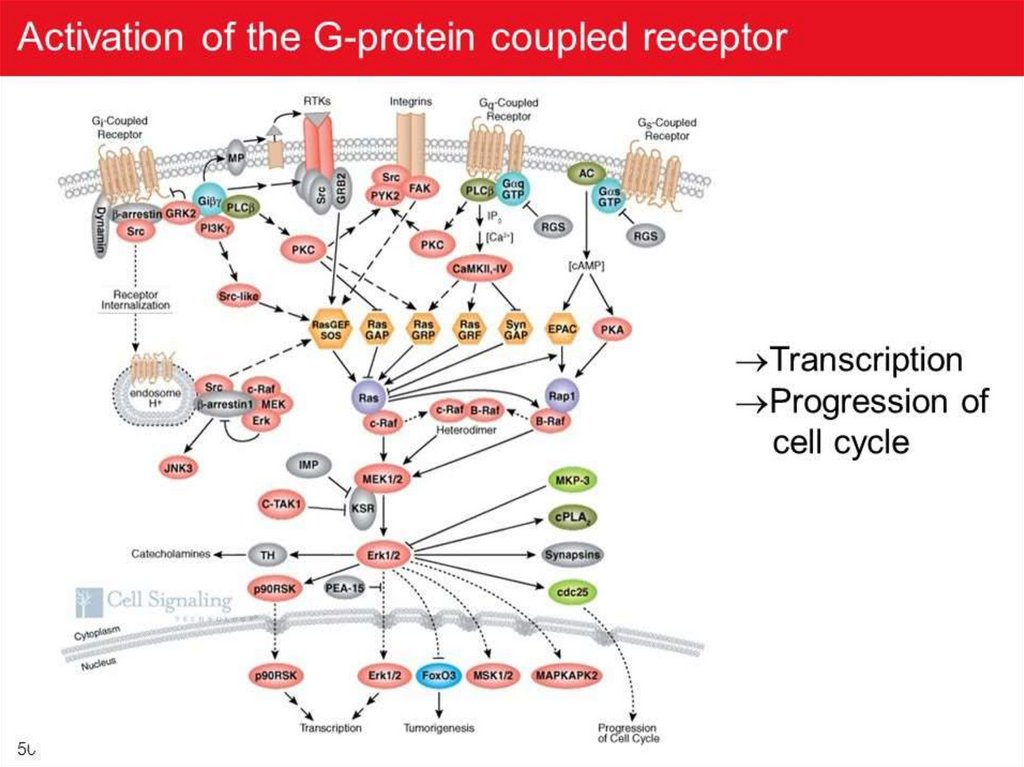
G-белкиG-белки — это семейство белков, относящихся к ГТФ-азам и
функционирующих в качестве посредников во внутриклеточных сигнальных
каскадах.
G-белки названы так, поскольку в своём сигнальном механизме они
используют замену ГДФ на ГТФ - как молекулярный функциональный
«выключатель» для регулировки клеточных процессов.
45.
G-белкиG-белки делятся на две основных группы — гетеротримерные
(«большие») и «малые».
Гетеротримерные G-белки — это белки, состоящие из трёх
субъединиц: альфа(α), бета (β) и гамма (γ).
Малые G-белки — это белки из одной полипептидной цепи
(массой 20 —25 кДа), она гомологична α-субъединице
гетеротримерных G-белков. Относятся к суперсемейству
Ras малых ГТФаз.
Обе группы G-белков участвуют во внутриклеточной
сигнализации.
46.
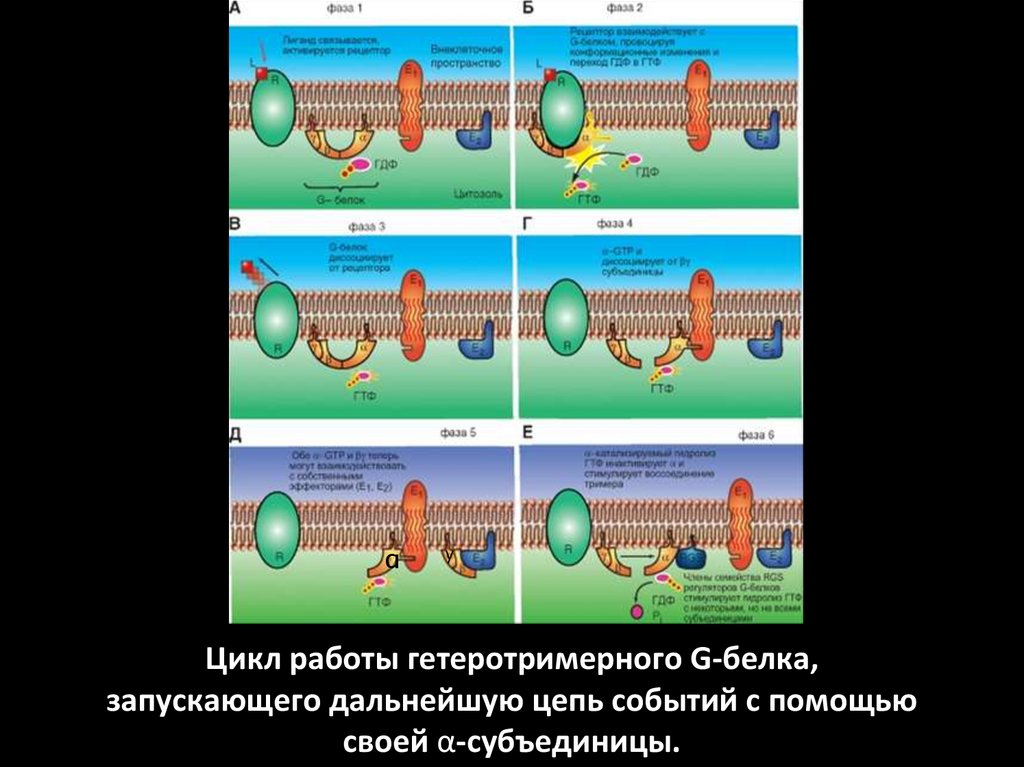
αy
Цикл работы гетеротримерного G-белка,
запускающего дальнейшую цепь событий с помощью
своей α-субъединицы.
47.
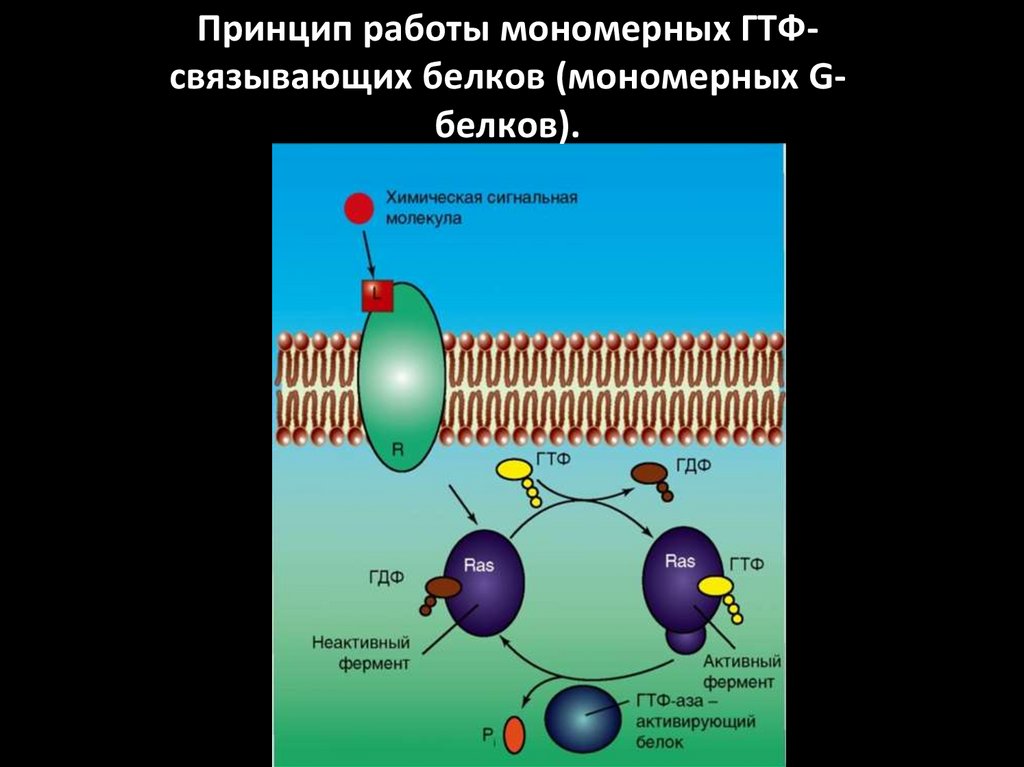
Принцип работы мономерных ГТФсвязывающих белков (мономерных Gбелков).48.
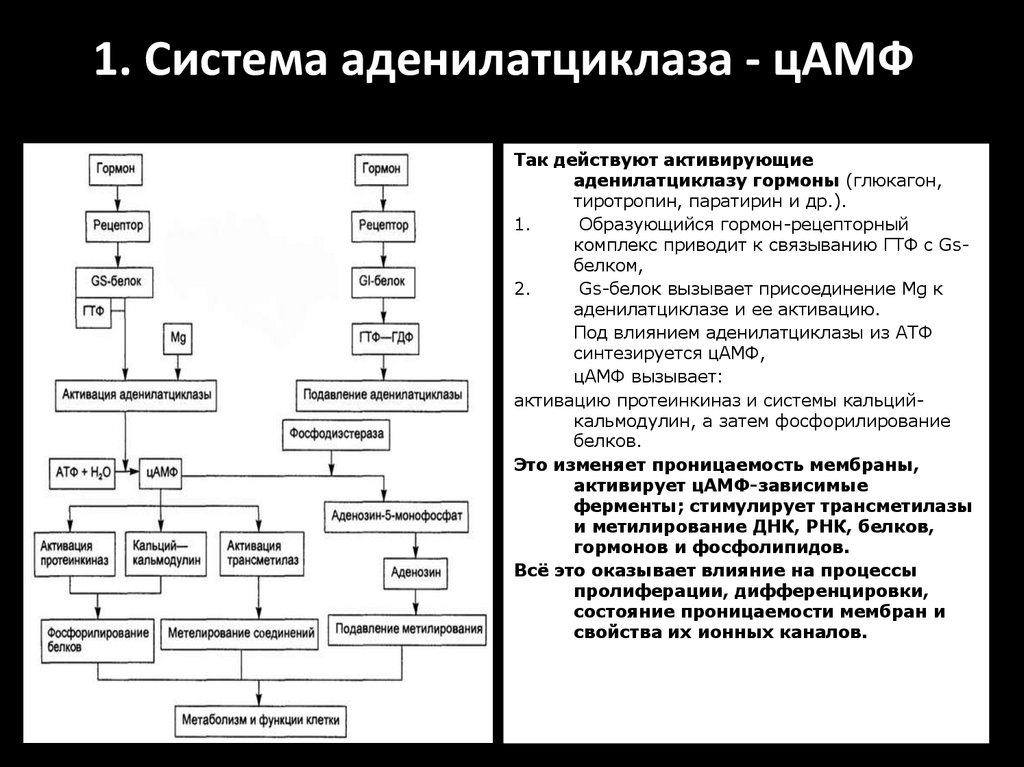
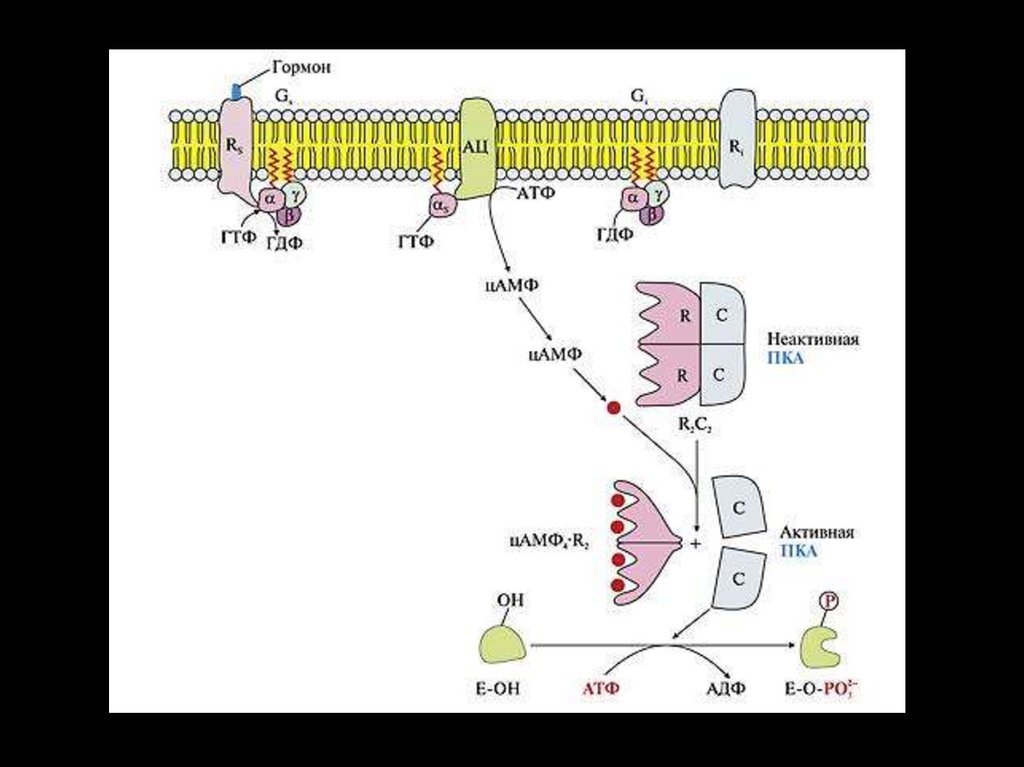
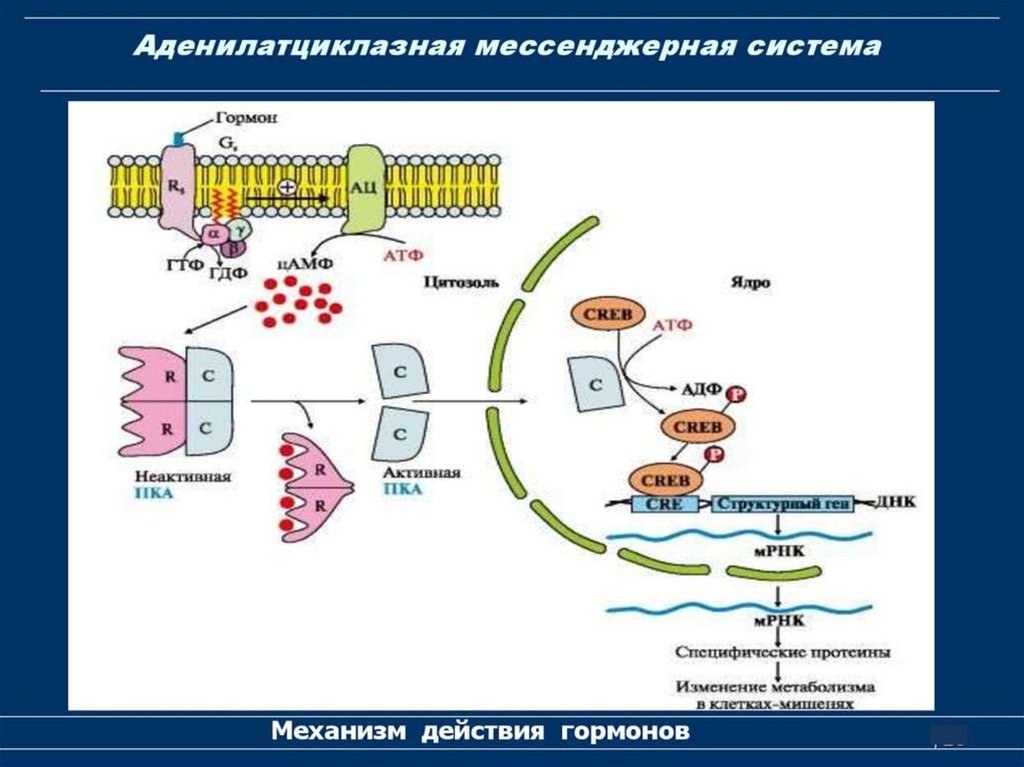
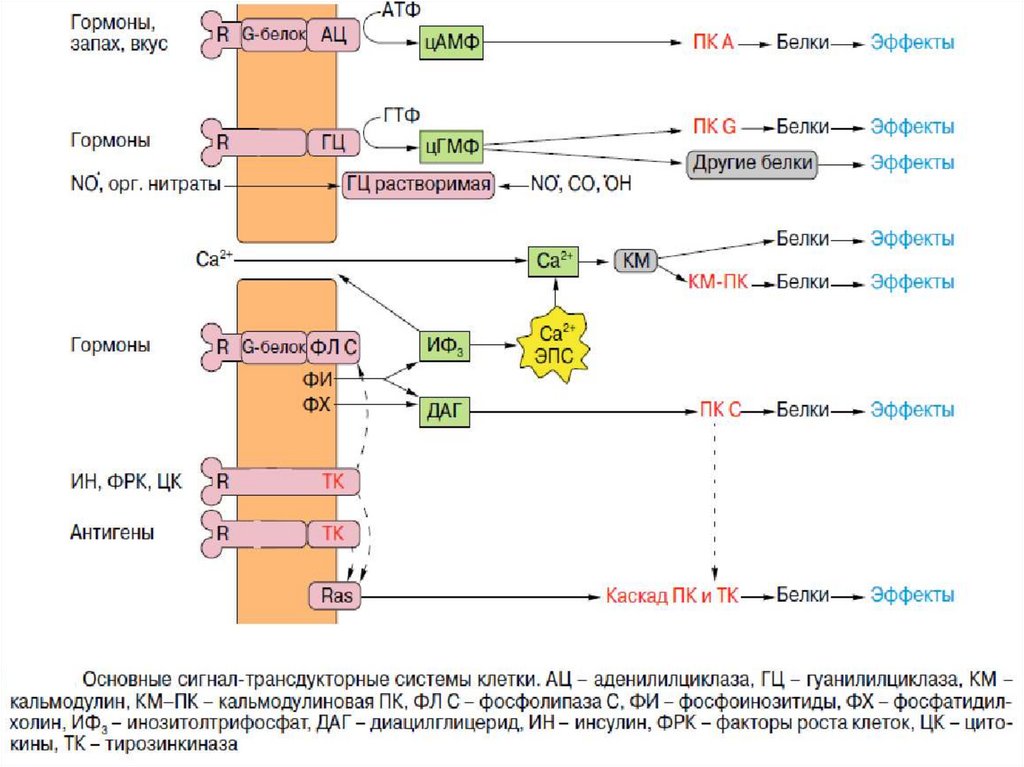
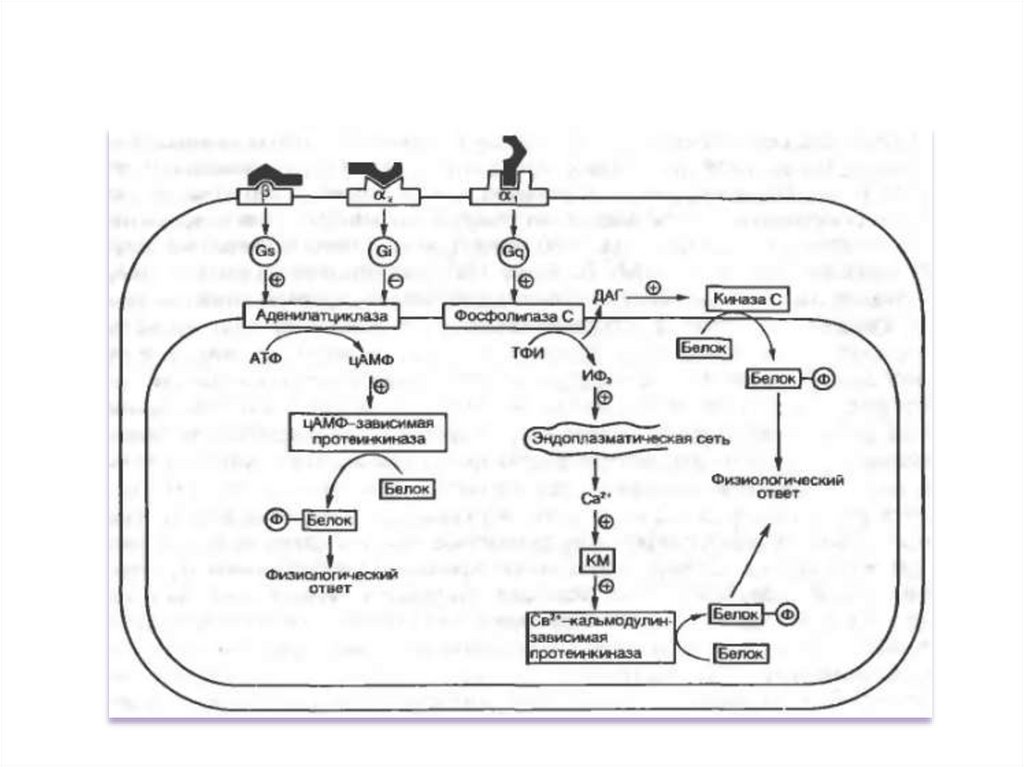
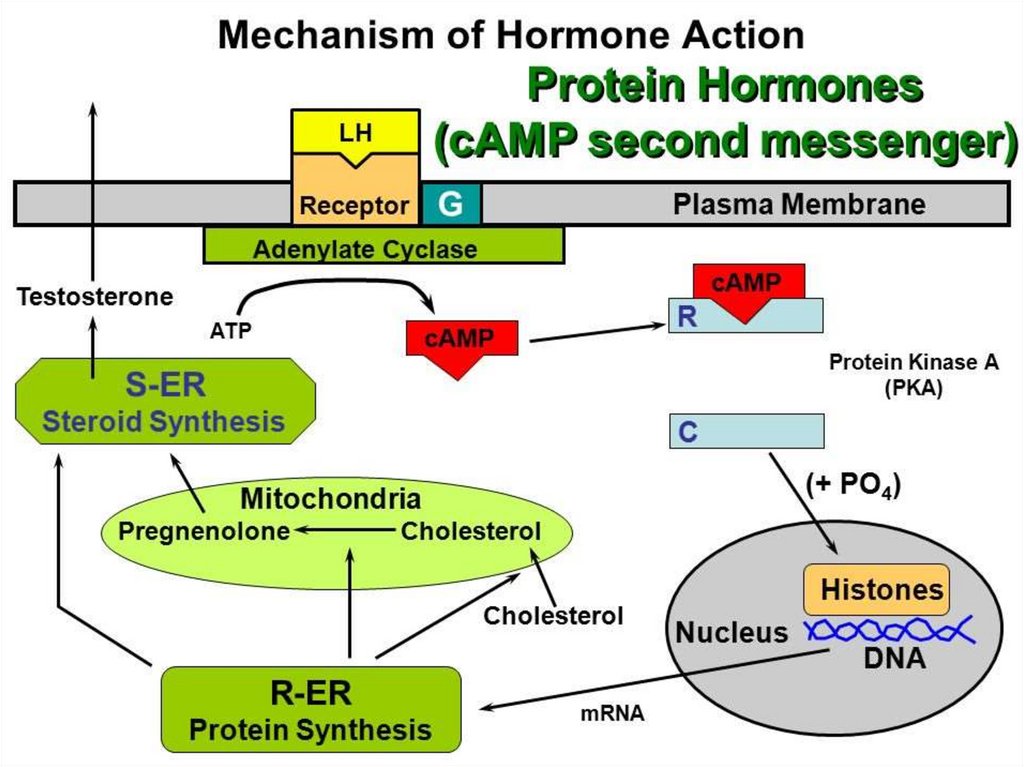
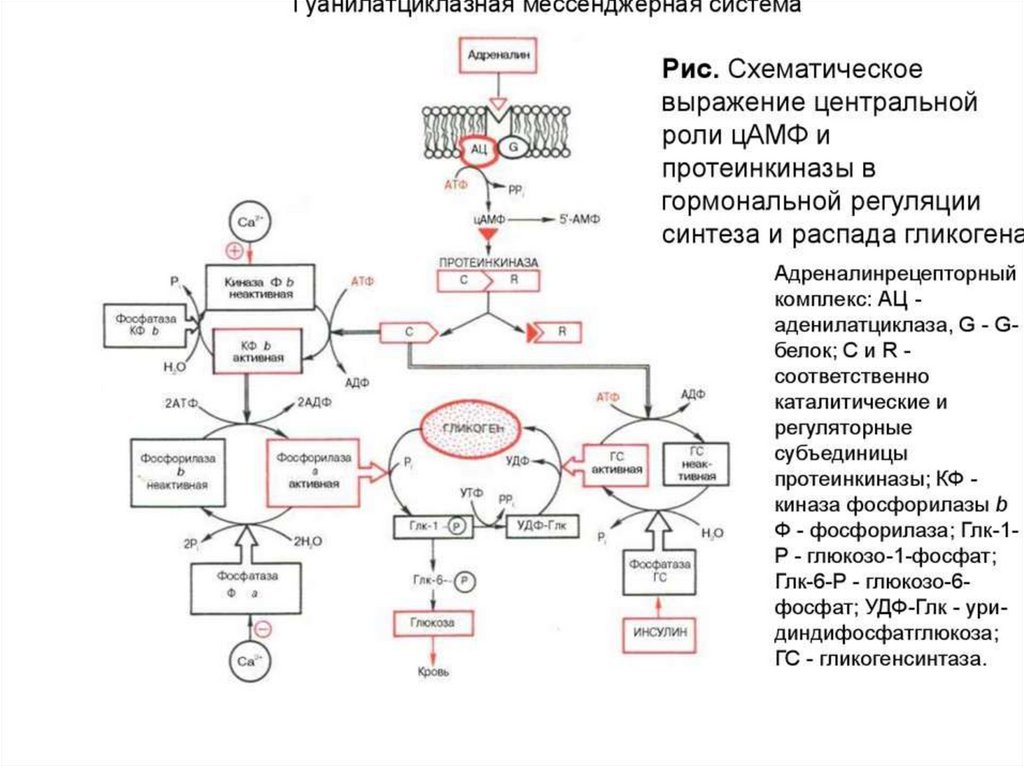
1. Система аденилатциклаза - цАМФТак действуют активирующие
аденилатциклазу гормоны (глюкагон,
тиротропин, паратирин и др.).
1.
Образующийся гормон-рецепторный
комплекс приводит к связыванию ГТФ с GSбелком,
2.
GS-белок вызывает присоединение Mg к
аденилатциклазе и ее активацию.
3.
Под влиянием аденилатциклазы из АТФ
синтезируется цАМФ,
4.
цАМФ вызывает:
активацию протеинкиназ и системы кальцийкальмодулин, а затем фосфорилирование
белков.
Это изменяет проницаемость мембраны,
активирует цАМФ-зависимые
ферменты; стимулирует трансметилазы
и метилирование ДНК, РНК, белков,
гормонов и фосфолипидов.
Всё это оказывает влияние на процессы
пролиферации, дифференцировки,
состояние проницаемости мембран и
свойства их ионных каналов.
49.
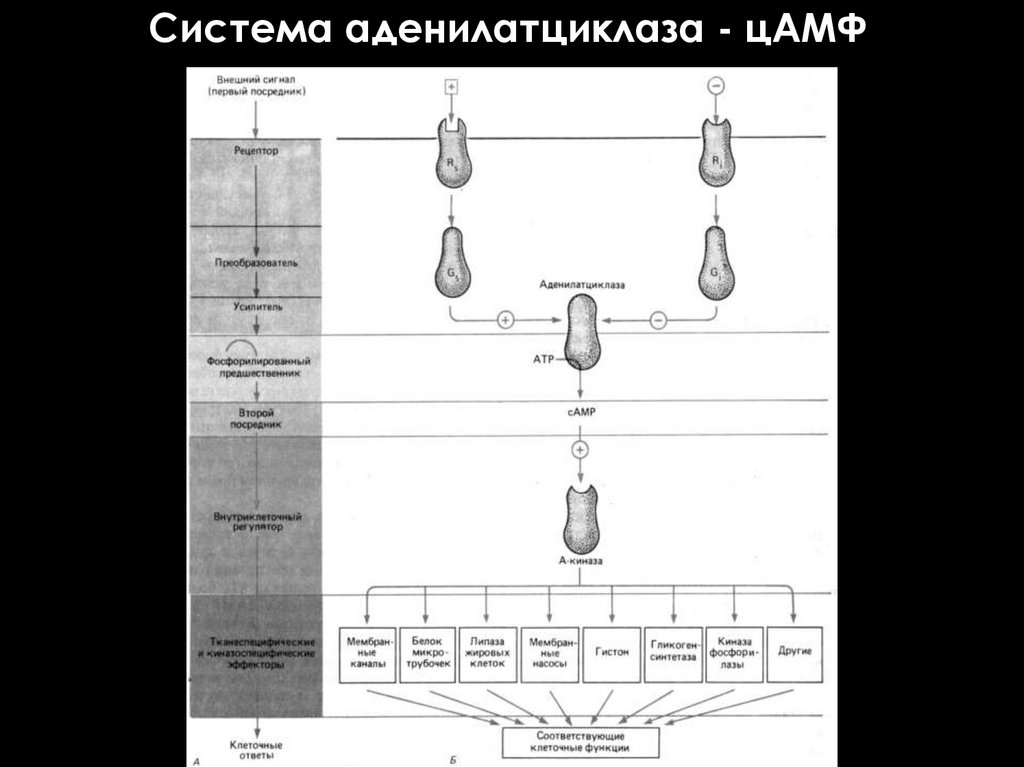
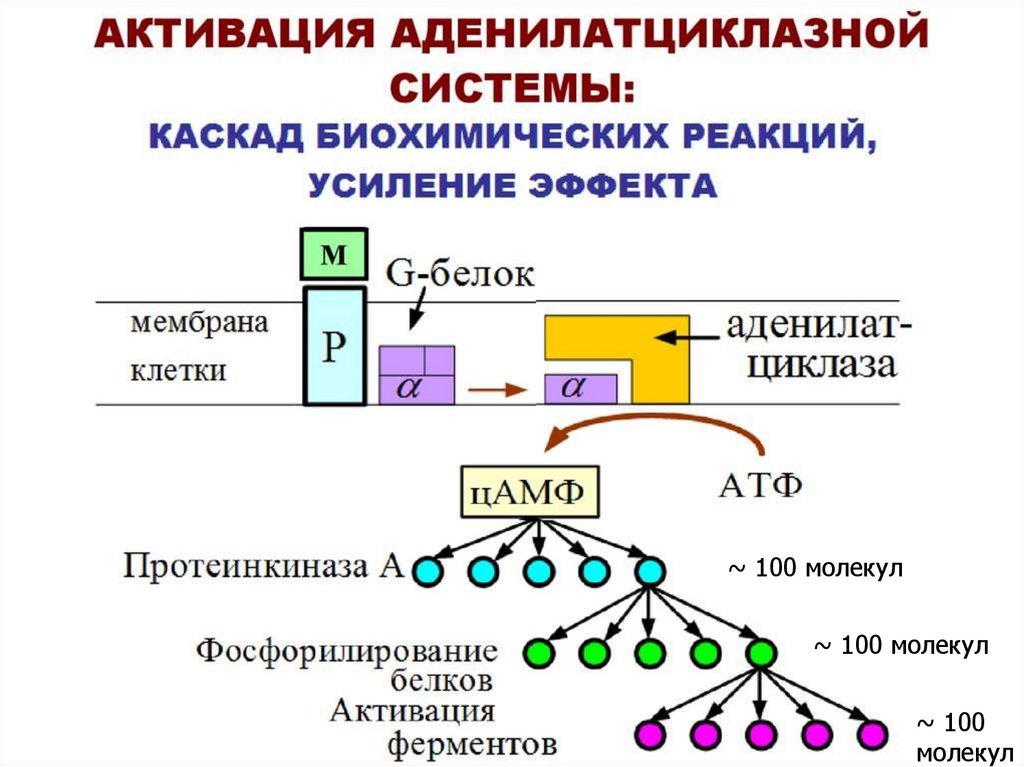
Система аденилатциклаза - цАМФ50.
51.
~ 100 молекул~ 100 молекул
~ 100
молекул
52.
53.
Холера – это инфекция, вызывающая диарею, котораяможет быстро привести к обезвоживанию организма и
смерти.
Диарея возникает в результате действия токсина,
продуцируемого
холерным
вибрионом.
Компонент
холерного токсина проникает в клетку и катализирует
ковалентное связывание АДФ-рибозы с α-субъединицей
GS –белка. В результате GS
стойко активируется и
постоянно стимулирует аденилатциклазу.
Вследствие этого концентрация цАМФ постоянно повышена.
Щёточная кайма мембраны, смотрящая в просвет тонкой
кишки, содержит хлорные каналы, которые открываются,
когда уровень цАМФ повышается.
Постоянная активация этих Cl- -каналов приводит к выводу
в просвет кишки CL-, Na+, и воды. Результатом этого
является постоянная водная диарея.
54.
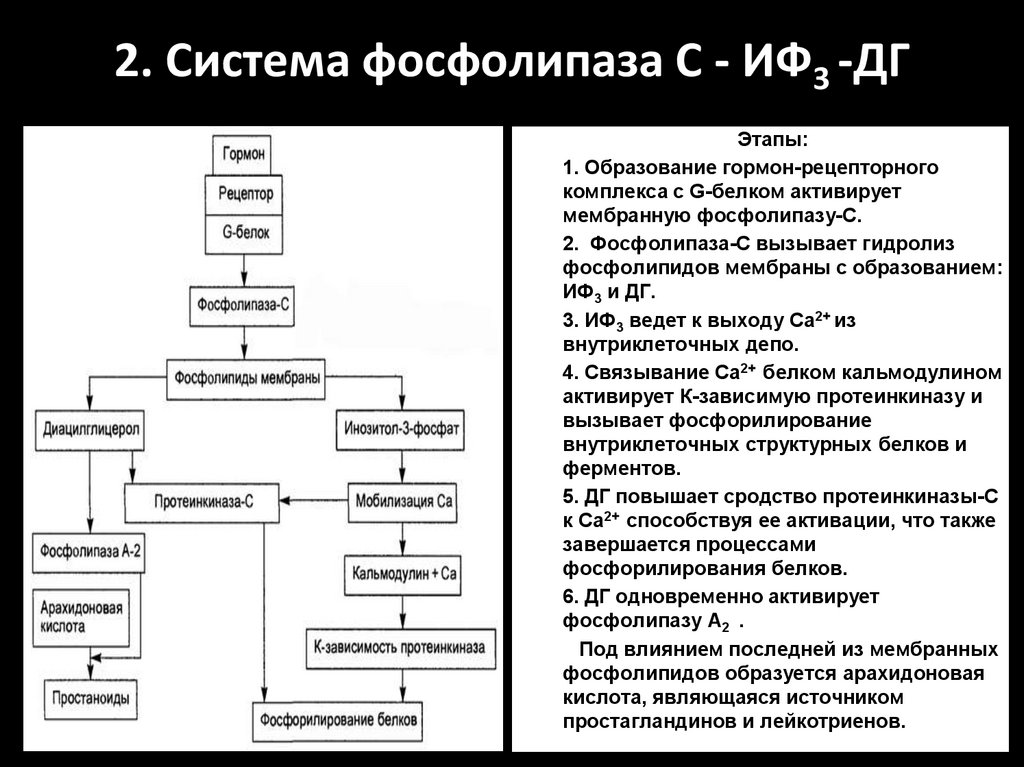
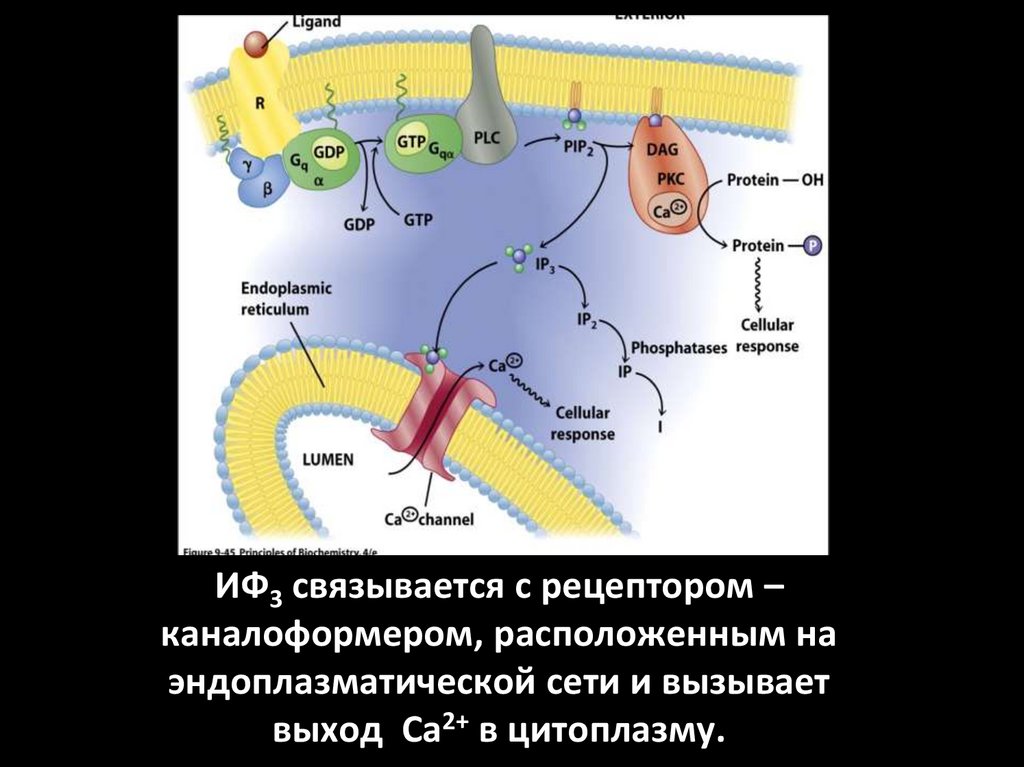
2. Система фосфолипаза С - ИФ3 -ДГ1.
2.
3.
4.
5.
6.
7.
Этапы:
1. Образование гормон-рецепторного
комплекса с G-белком активирует
мембранную фосфолипазу-С.
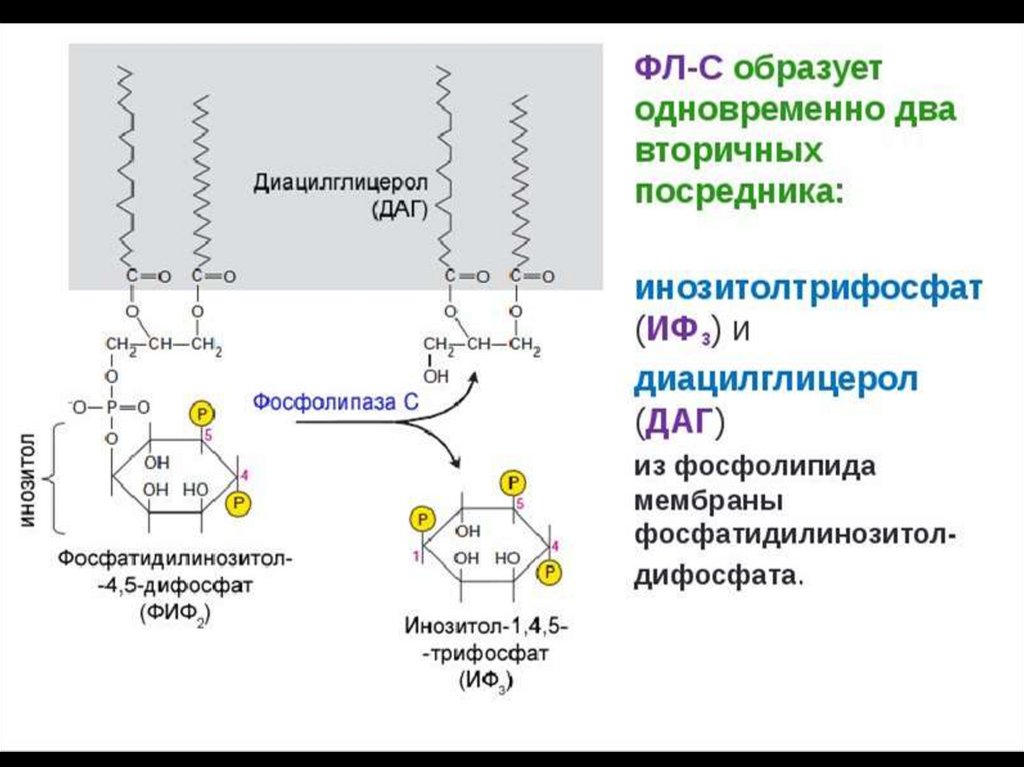
2. Фосфолипаза-С вызывает гидролиз
фосфолипидов мембраны с образованием:
ИФ3 и ДГ.
3. ИФ3 ведет к выходу Са2+ из
внутриклеточных депо.
4. Связывание Са2+ белком кальмодулином
активирует К-зависимую протеинкиназу и
вызывает фосфорилирование
внутриклеточных структурных белков и
ферментов.
5. ДГ повышает сродство протеинкиназы-С
к Са2+ способствуя ее активации, что также
завершается процессами
фосфорилирования белков.
6. ДГ одновременно активирует
фосфолипазу А2 .
Под влиянием последней из мембранных
фосфолипидов образуется арахидоновая
кислота, являющаяся источником
простагландинов и лейкотриенов.
55.
56.
ИФ3 связывается с рецептором –каналоформером, расположенным на
эндоплазматической сети и вызывает
выход Са2+ в цитоплазму.
57.
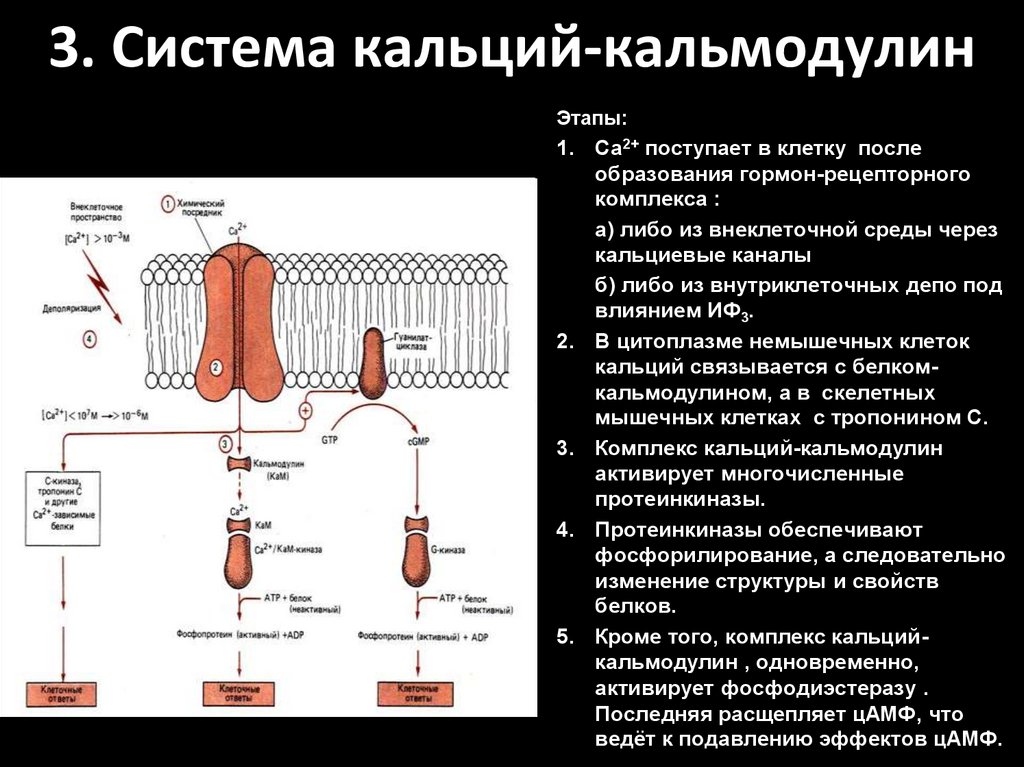
3. Система кальций-кальмодулинЭтапы:
1. Са2+ поступает в клетку после
образования гормон-рецепторного
комплекса :
а) либо из внеклеточной среды через
кальциевые каналы
б) либо из внутриклеточных депо под
влиянием ИФ3.
2. В цитоплазме немышечных клеток
кальций связывается с белкомкальмодулином, а в скелетных
мышечных клетках с тропонином С.
3. Комплекс кальций-кальмодулин
активирует многочисленные
протеинкиназы.
4. Протеинкиназы обеспечивают
фосфорилирование, а следовательно
изменение структуры и свойств
белков.
5. Кроме того, комплекс кальцийкальмодулин , одновременно,
активирует фосфодиэстеразу .
Последняя расщепляет цАМФ, что
ведёт к подавлению эффектов цАМФ.
58.
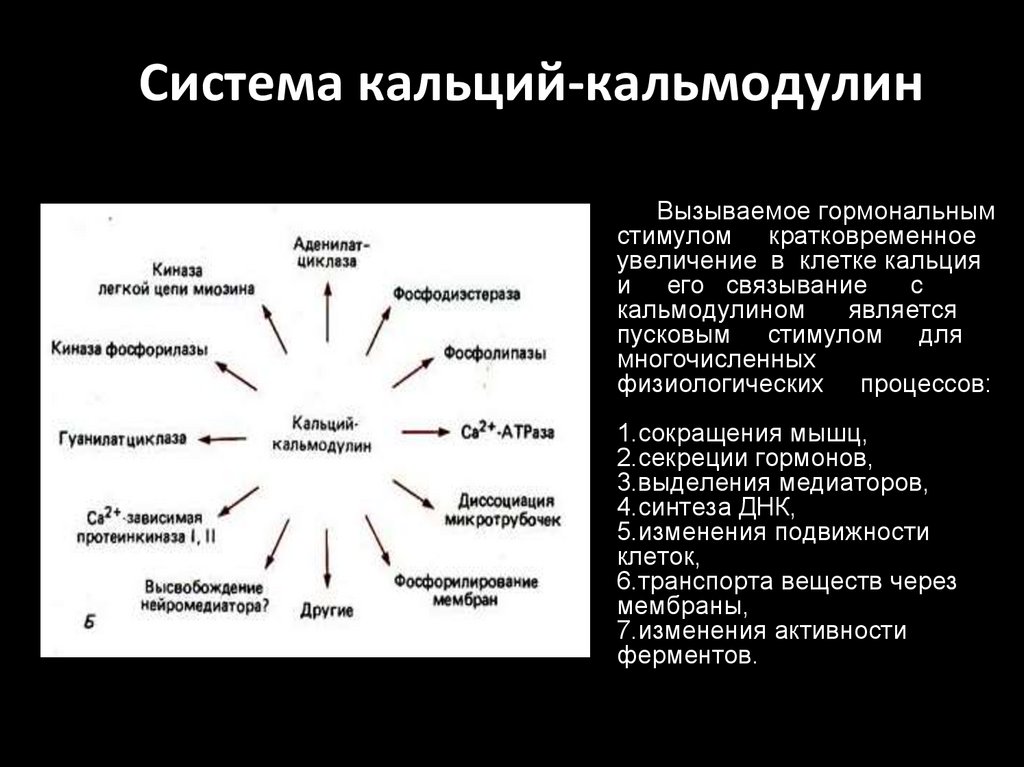
Система кальций-кальмодулинВызываемое гормональным
стимулом кратковременное
увеличение в клетке кальция
и его связывание
с
кальмодулином
является
пусковым стимулом для
многочисленных
физиологических процессов:
1.сокращения мышц,
2.секреции гормонов,
3.выделения медиаторов,
4.синтеза ДНК,
5.изменения подвижности
клеток,
6.транспорта веществ через
мембраны,
7.изменения активности
ферментов.
59.
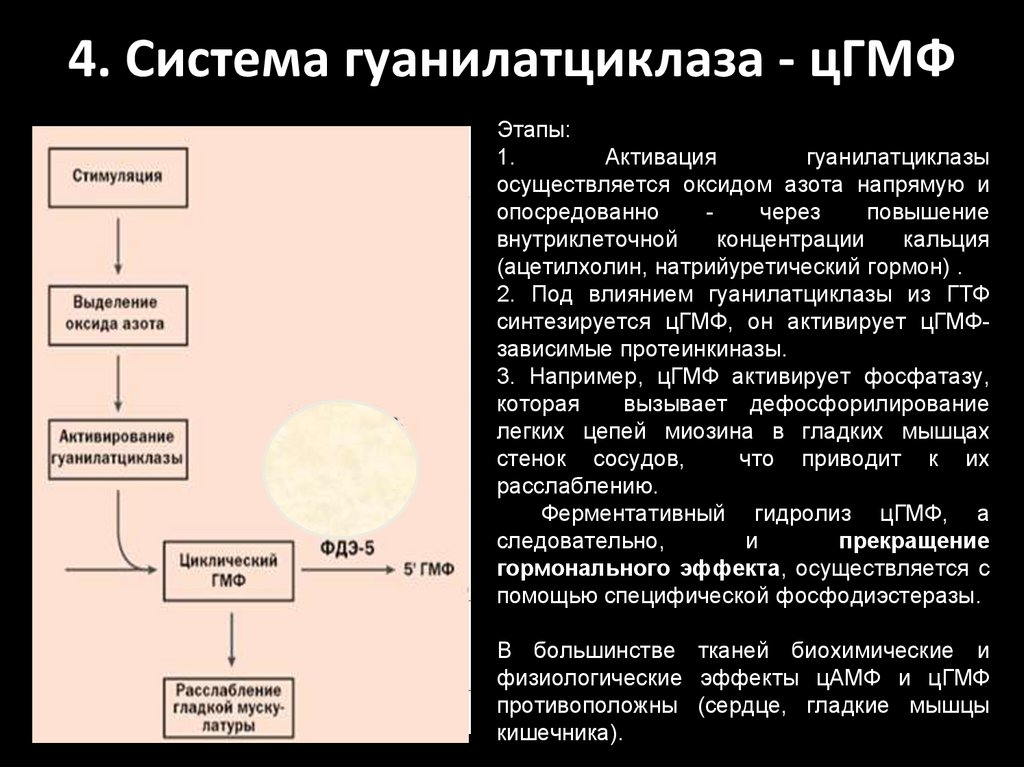
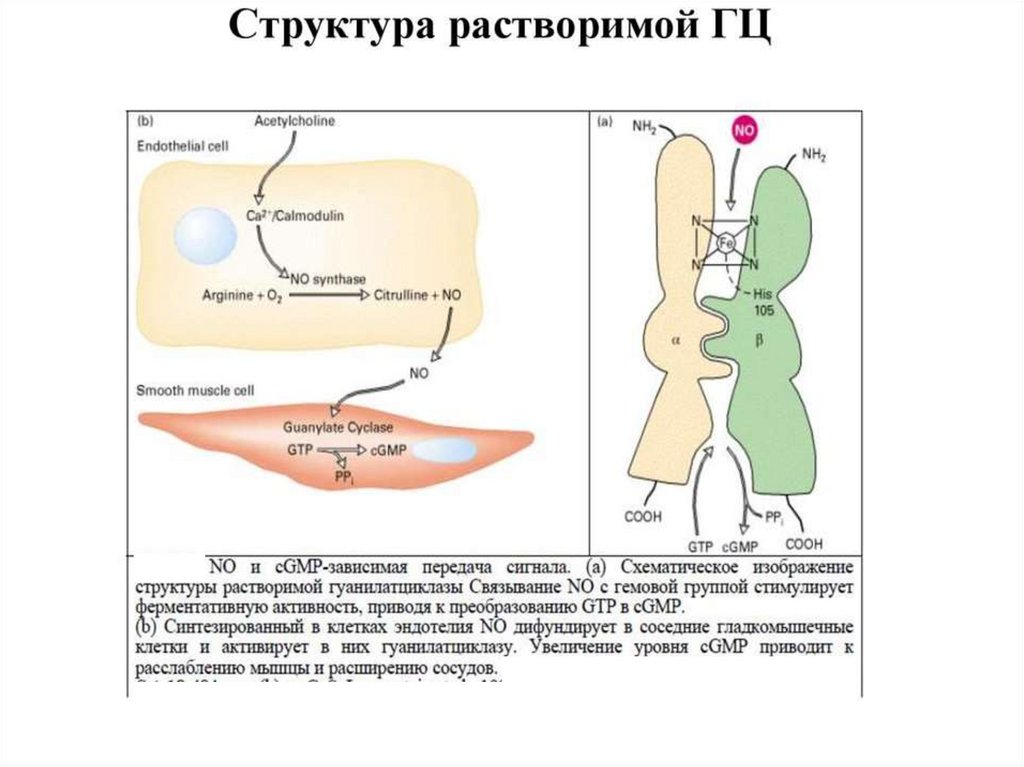
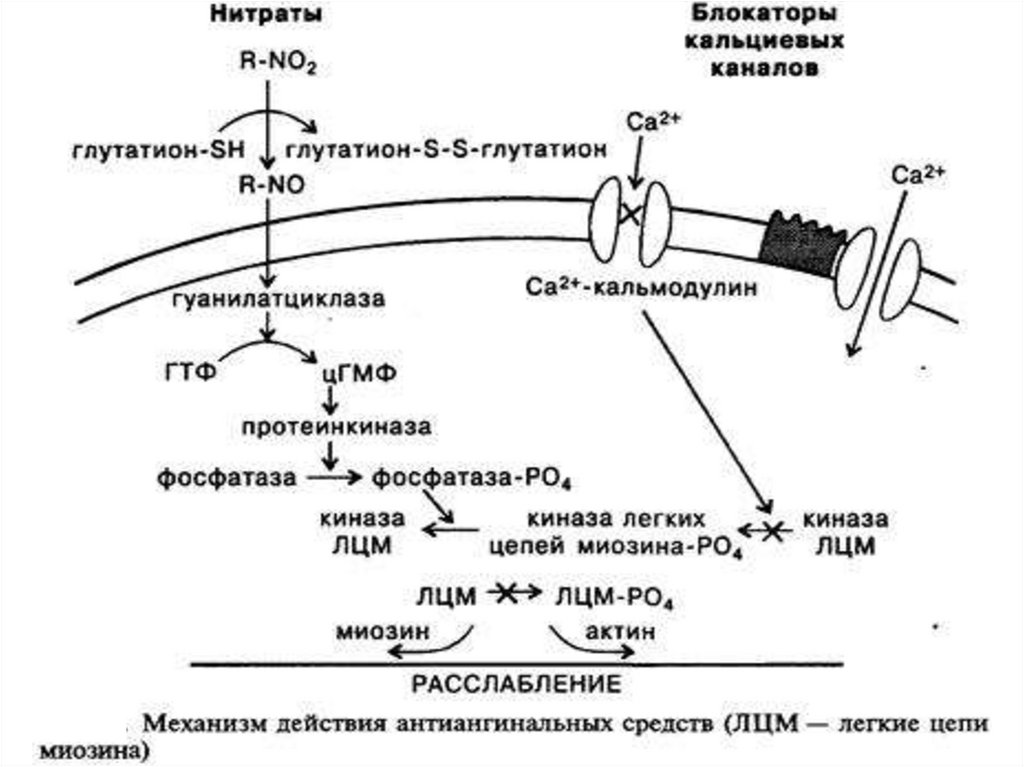
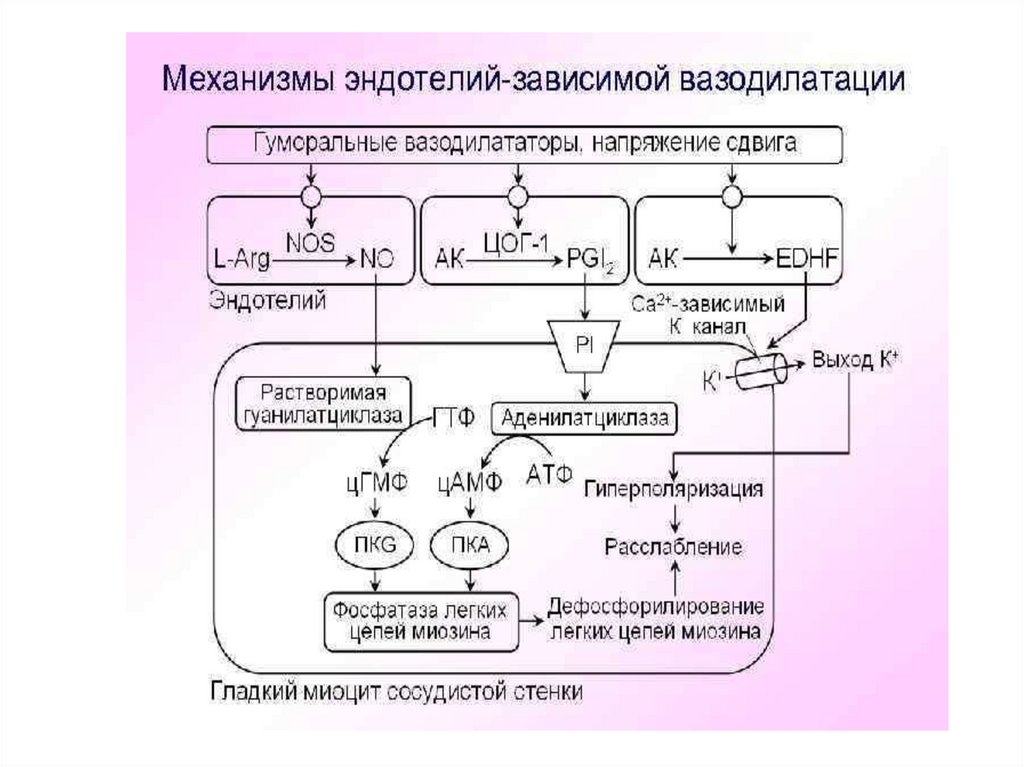
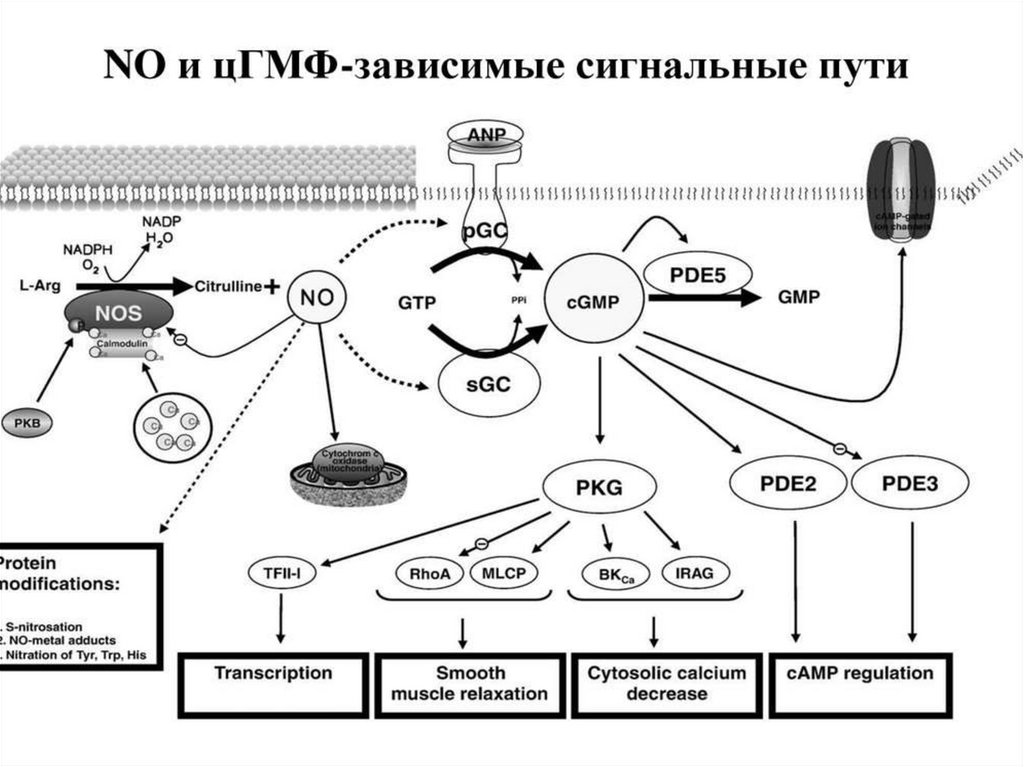
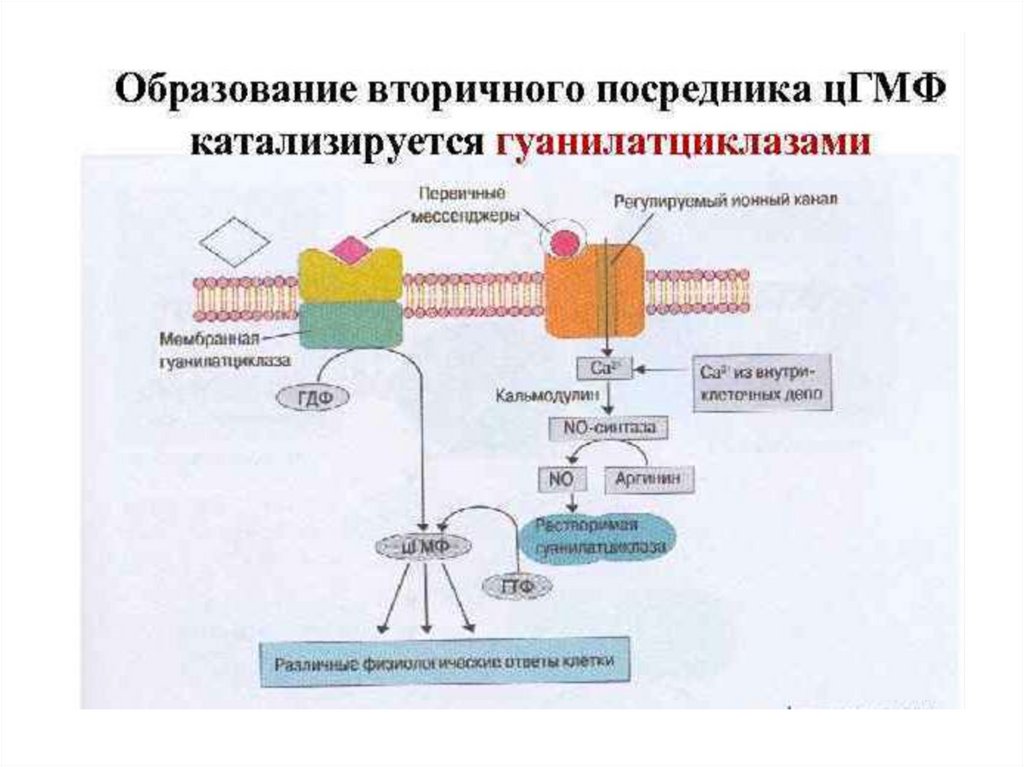
4. Система гуанилатциклаза - цГМФЭтапы:
1.
Активация
гуанилатциклазы
осуществляется оксидом азота напрямую и
опосредованно
через
повышение
внутриклеточной
концентрации
кальция
(ацетилхолин, натрийуретический гормон) .
2. Под влиянием гуанилатциклазы из ГТФ
синтезируется цГМФ, он активирует цГМФзависимые протеинкиназы.
3. Например, цГМФ активирует фосфатазу,
которая
вызывает дефосфорилирование
легких цепей миозина в гладких мышцах
стенок сосудов,
что приводит к их
расслаблению.
Ферментативный гидролиз цГМФ, а
следовательно,
и
прекращение
гормонального эффекта, осуществляется с
помощью специфической фосфодиэстеразы.
В большинстве тканей биохимические и
физиологические эффекты цАМФ и цГМФ
противоположны (сердце, гладкие мышцы
кишечника).
60.
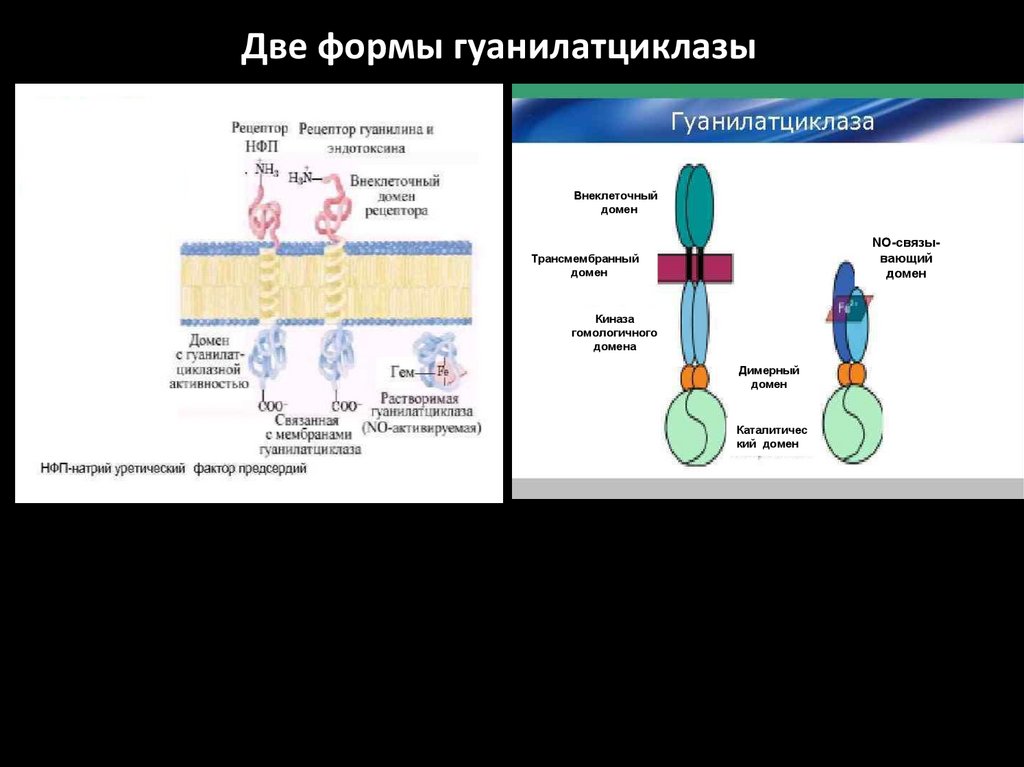
Гуанилатциклаза (ГЦ)1.ГЦ катализирует образование вторичного посредника цГМФ из
ГТФ.
2.В клетке имеется 2 формы ГЦ – мембраносвязанная (мГЦ) и
растворимая (рГЦ).
3.Соотношение форм ГЦ в тканях разное: в клетках кишечника –
90% мГЦ, в лёгких и печени – 20% мГЦ.
4.рГЦ – димер (α и β-сб.), простетическая группа – гем, активируется NO и АФК.
5.мГЦ – трансмембранный гликопротеин, относится к рецепторам,
сопряжённым с ферментативной активностью. Различают 3
изоформы
мГЦ,
которые
активируются
различными
регуляторами: предсердным натрийуретическим фактором,
натрийуретическим пептидом мозга, кишечным пептидом
гуанилином.
6.В клетках выявлено 3 типа эффекторных белков, с которыми
взаимодействует цГМФ: цГМФ-зависмая ПК (ПКG), цГМФрегулируемые
ионные
каналы,
цГМФ-зависимая
фосфодиэстераза.
61.
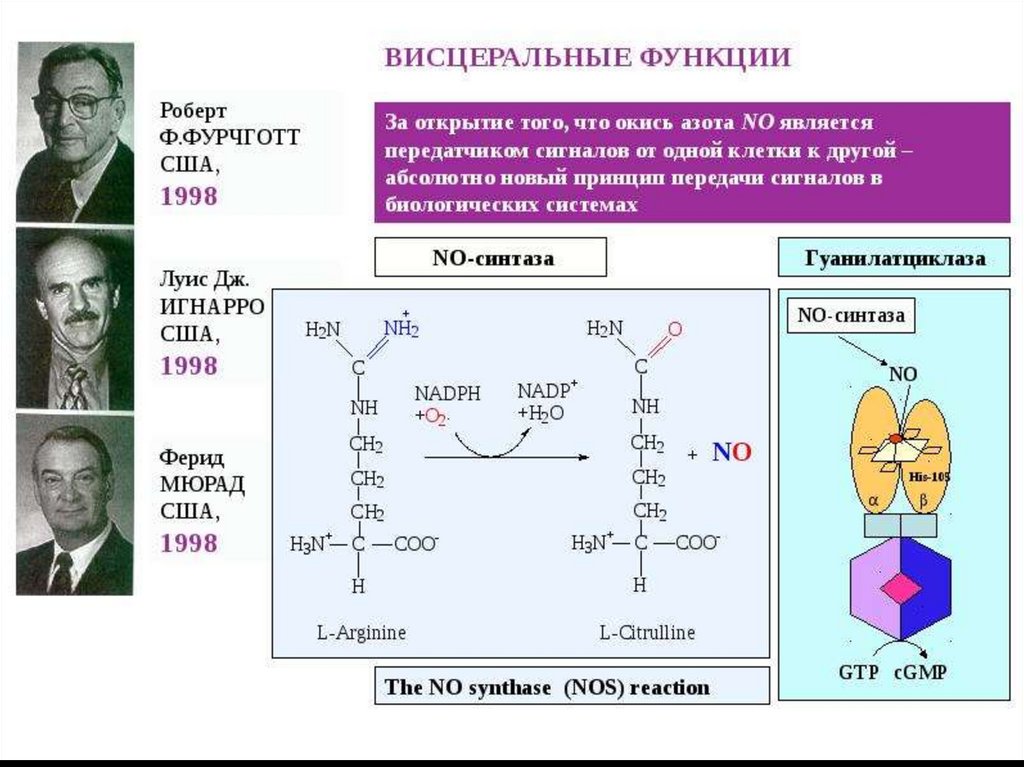
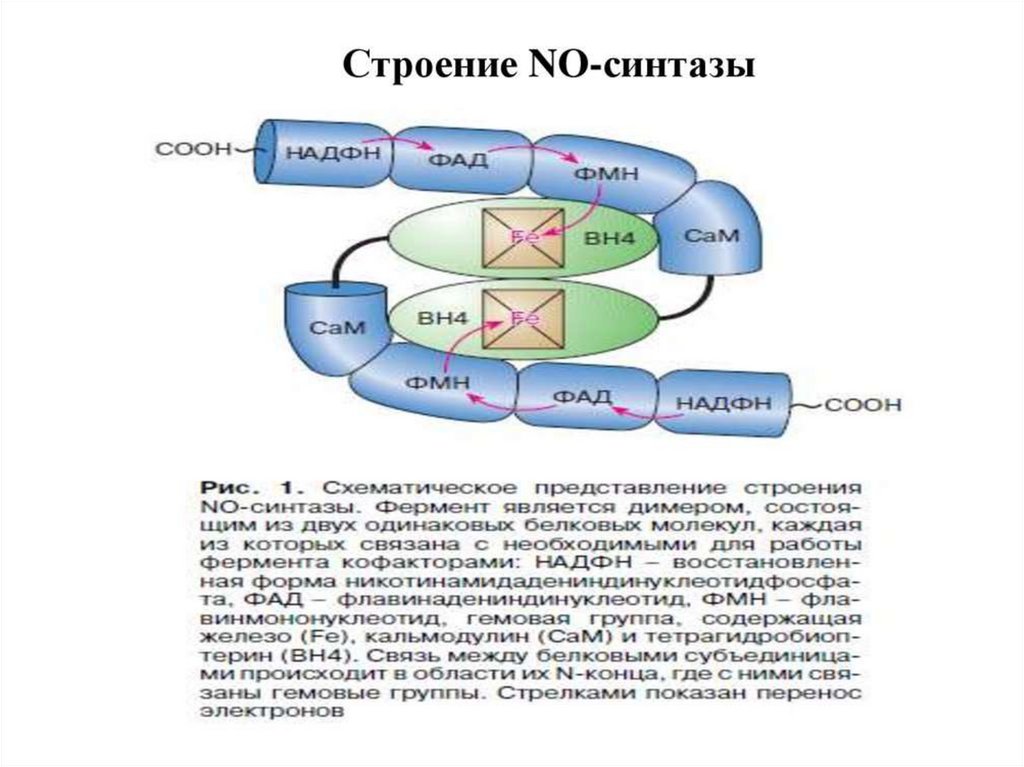
NOкак вторичный мессенджер
Образуется при окислении L-аргинина ферментом NOсинтазой (NOS).
Данный фермент
присутствует: в нервной ткани,
эндотелии сосудов, тромбоцитах и других тканях.
62.
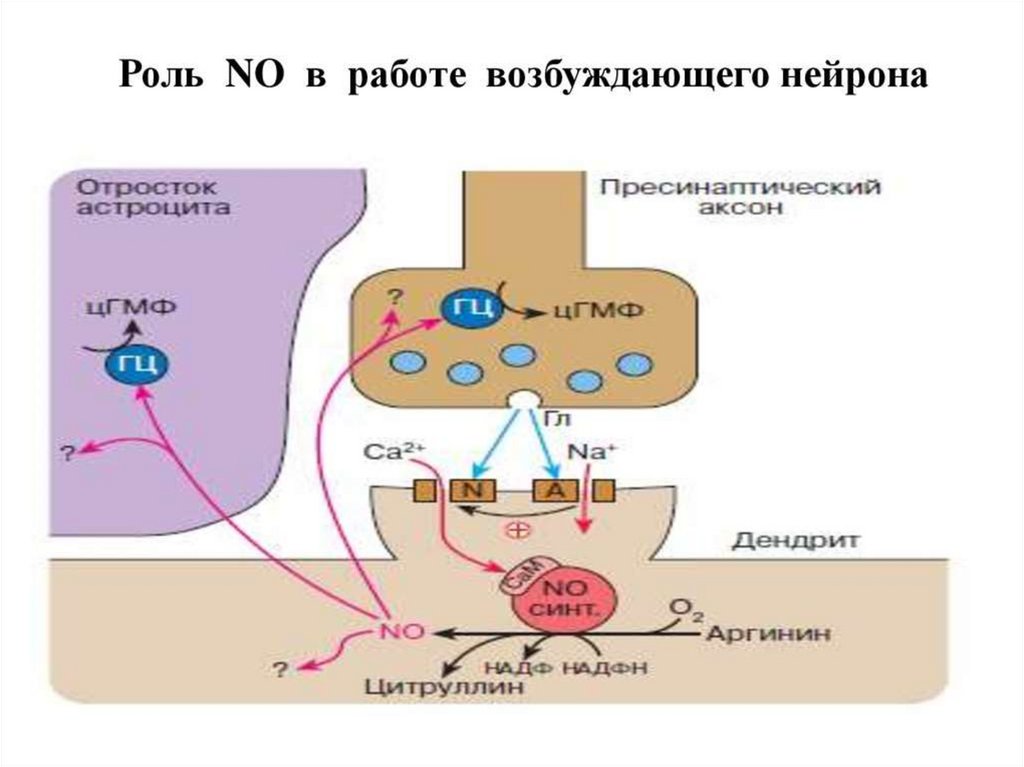
Механизм действияВ клетках-мишенях, например, эндотелиальных
клетках: NO взаимодействует с входящим в
активный центр гуанилатциклазы ионом железа,
способствуя тем самым быстрому образованию
цГМФ.
Увеличение концентрации цГМФ в клетках
вызывает активацию цГМФ-зависимых киназ, что
в конечном итоге приводит к расслаблению ГМК
сосудов.
63.
64.
Две формы гуанилатциклазыВнеклеточный
домен
NО-связывающий
домен
т
Трансмембранный
домен
Киназа
гомологичного
домена
Димерный
домен
Каталитичес
кий домен
65.
66.
67.
Взаимосвязи вторичных посредниковМежду вторичными посредниками устанавливаются
различные взаимоотношения:
1) равнозначное участие, когда разные посредники
необходимы для полноценного гормонального эффекта;
2) один из посредников является основным, а другой лишь
способствует реализации эффектов первого;
3) посредники действуют последовательно: например, ИФ3 –
обеспечивает освобождение кальция, ДГ - облегчает
взаимодействие кальция с протеинкиназой С;
4) посредники дублируют друг друга для обеспечения
избыточности с целью надежности регуляции;
5) посредники являются антагонистами, т. е. один из них
включает реакцию, а другой — тормозит (например, в
гладких мышцах сосудов ИФ3 и кальций реализуют их
сокращение, а цАМФ — расслабление).
68.
69.
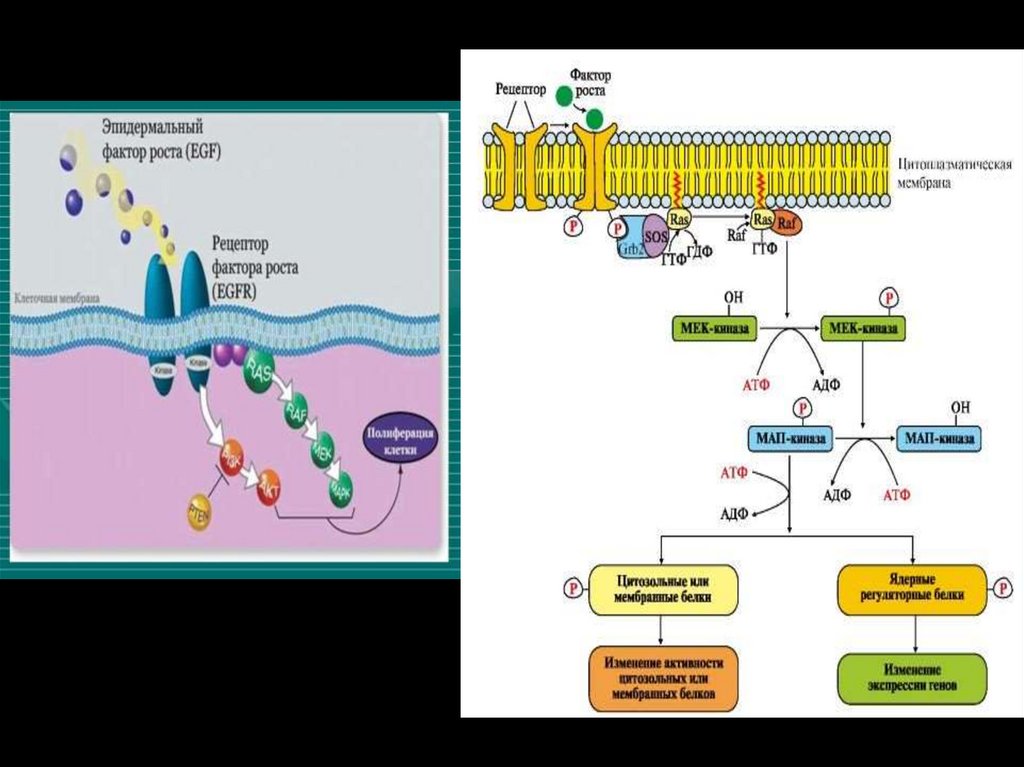
малые G белки(низкомолекулярные ГТФазы)
Опосредуют стимуляцию клетки факторами роста, а также
ее злокачественное перерождение.
Малые GTP-азы распадаются на следующие семейства:
ras-белки (онкобелки)
rho/rac-белки (цитоскелет, форма, движение клеток)
rab-белки(везикулярный транспорт)
ran-белки(ядерный транспорт)
Члены каждой группы имеют сходные функциональные
характеристики.
70.
71.
72.
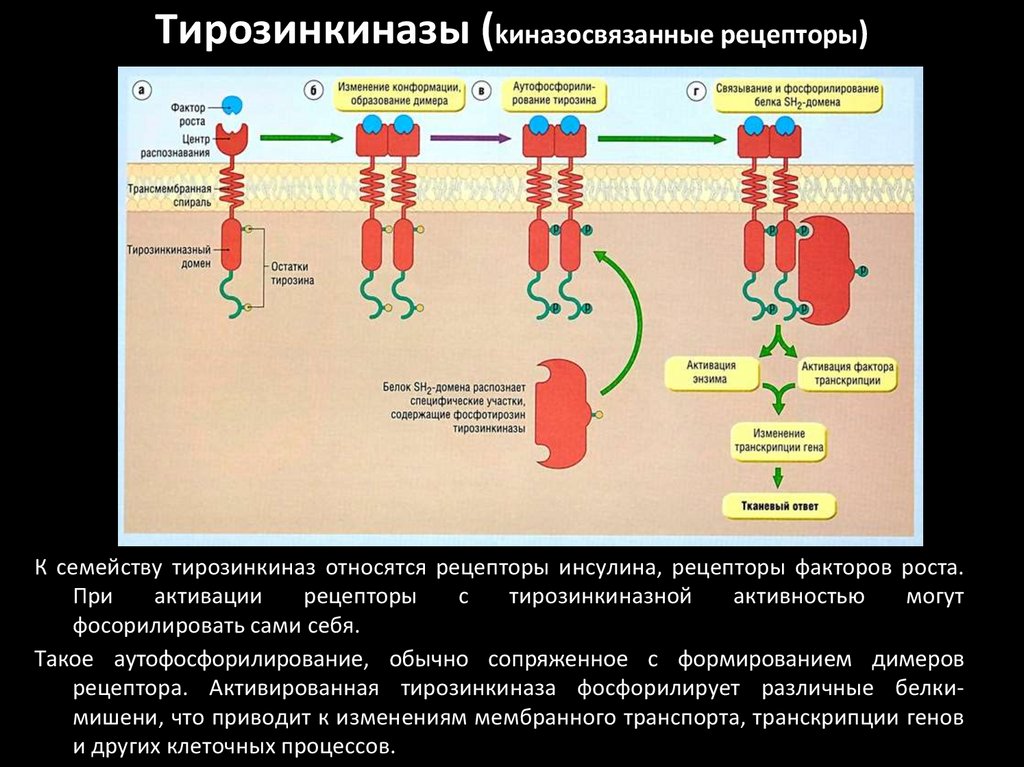
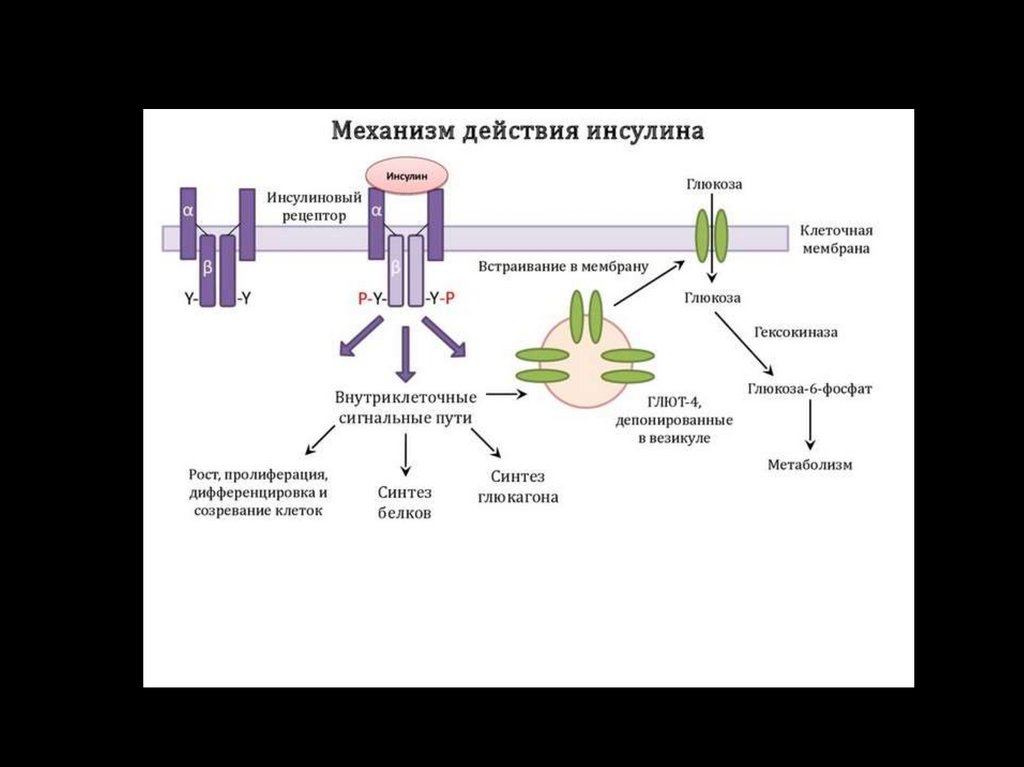
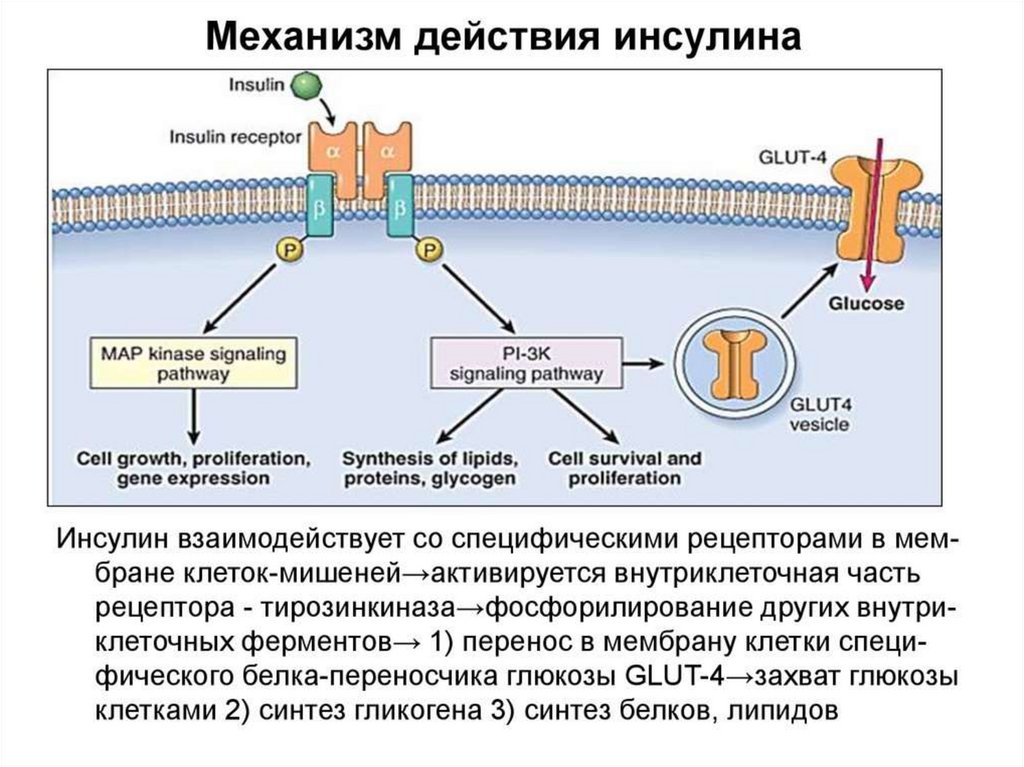
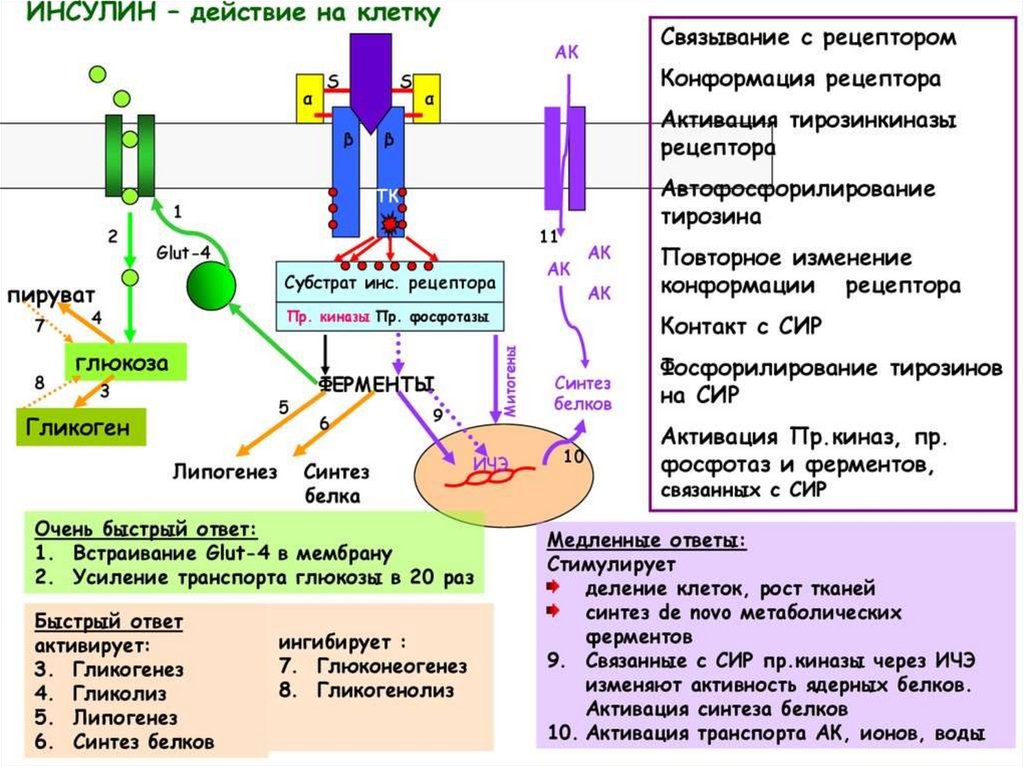
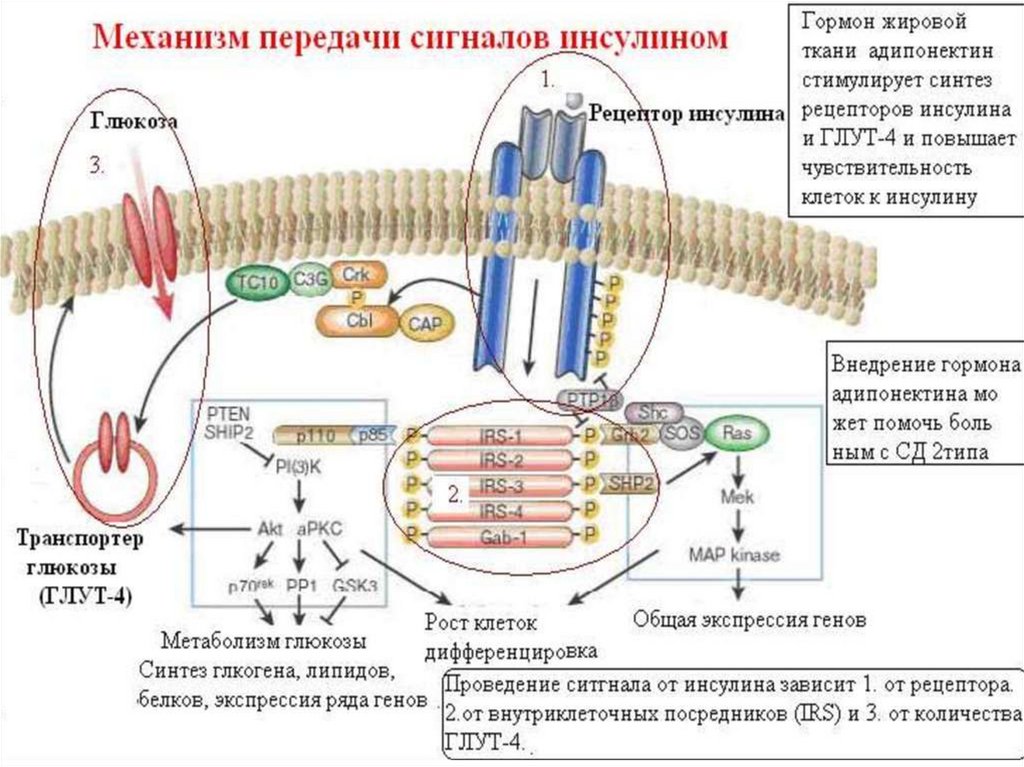
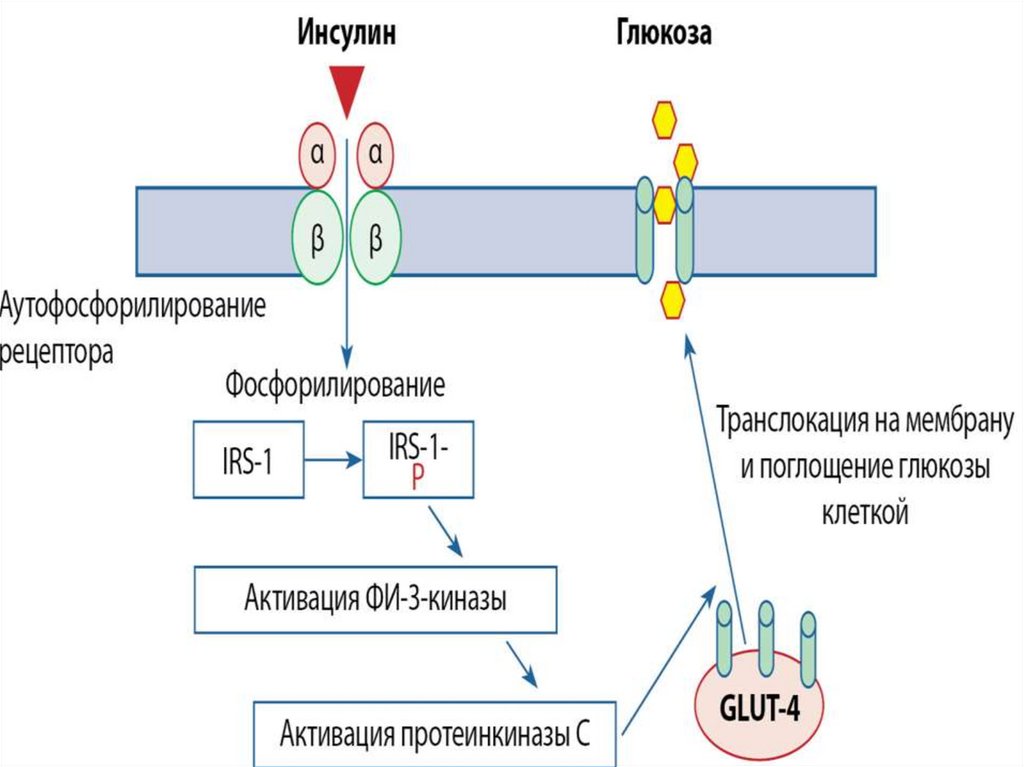
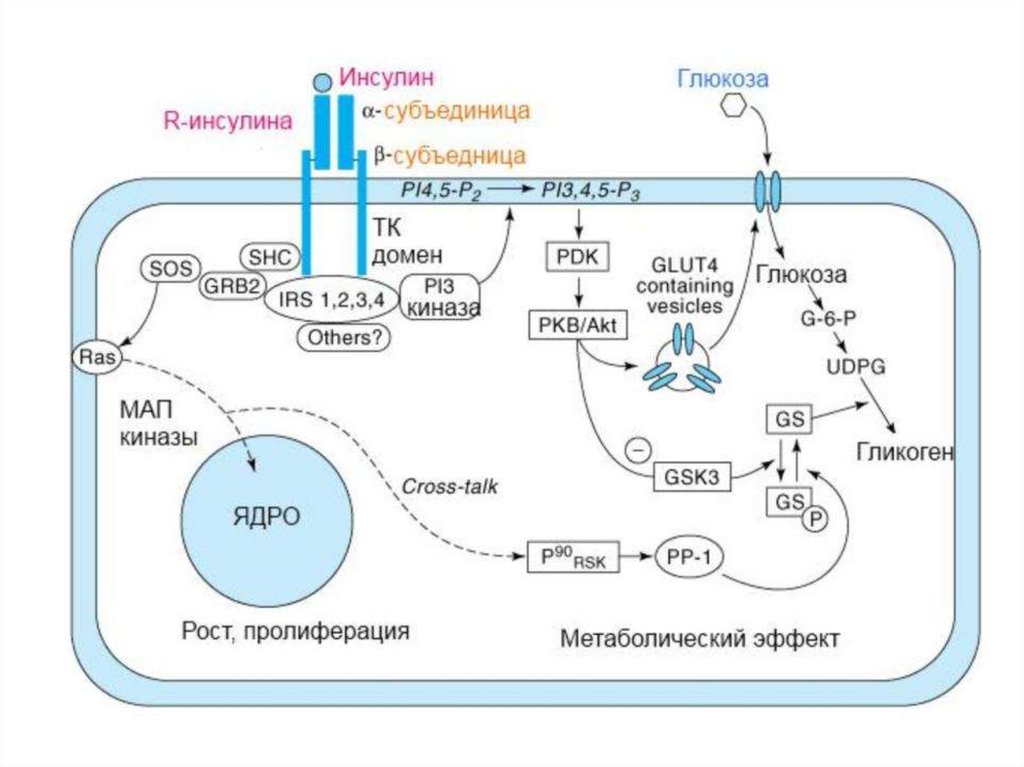
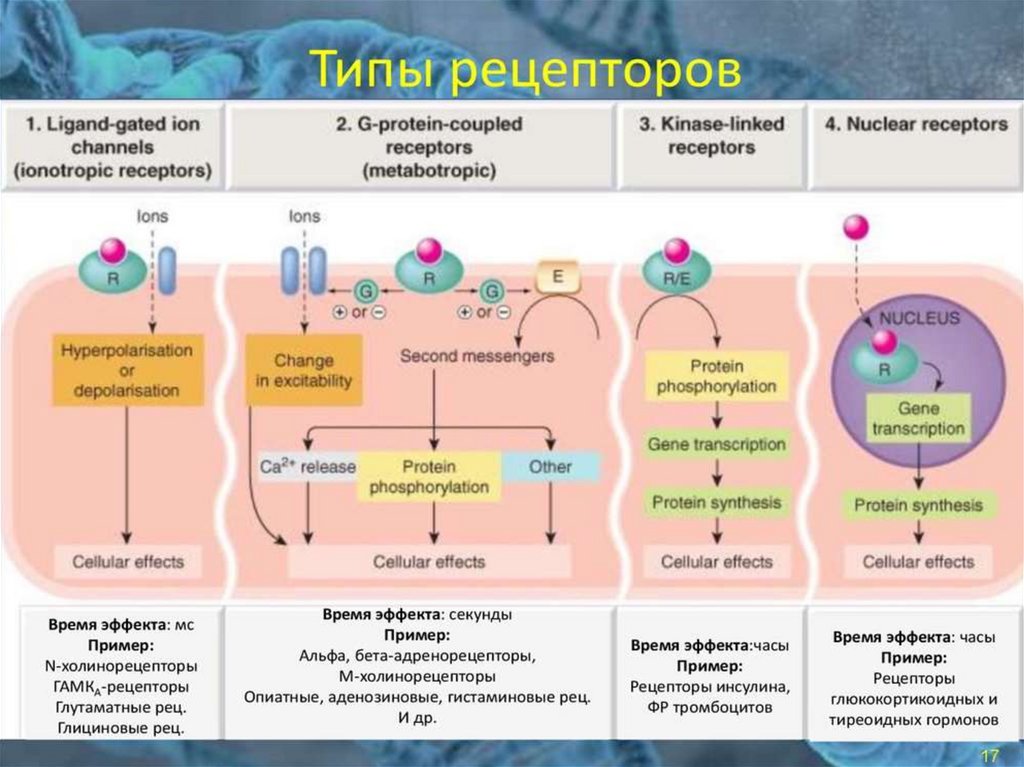
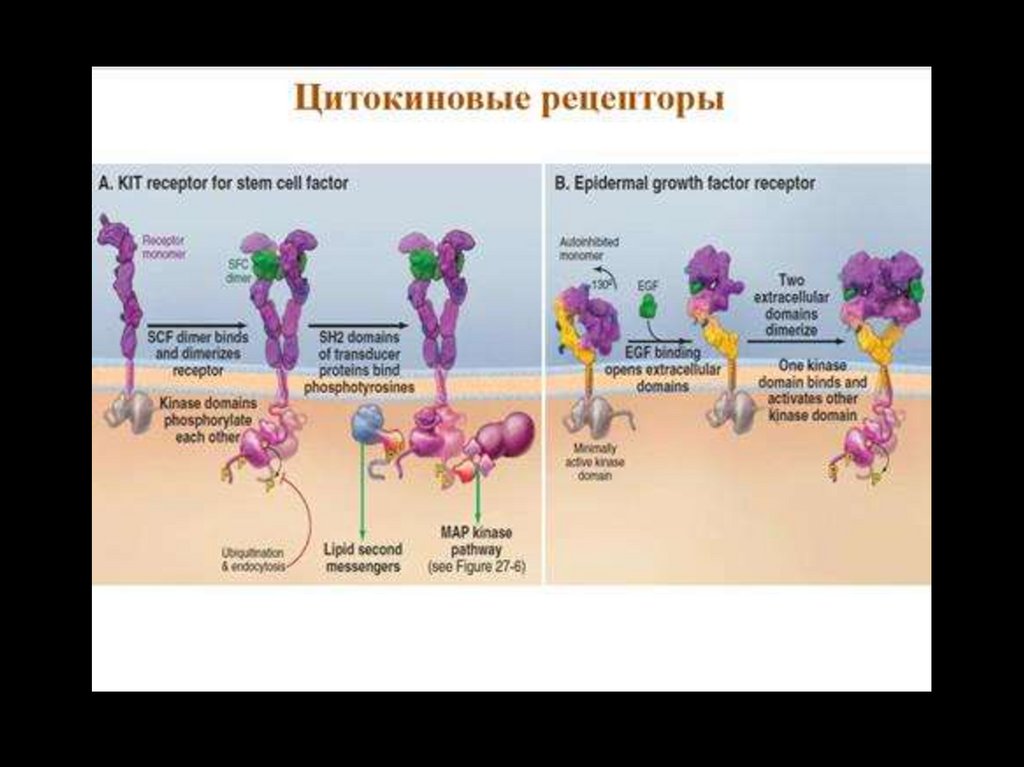
Тирозинкиназы (kиназосвязанные рецепторы)К семейству тирозинкиназ относятся рецепторы инсулина, рецепторы факторов роста.
При
активации
рецепторы
с
тирозинкиназной
активностью
могут
фосорилировать сами себя.
Такое аутофосфорилирование, обычно сопряженное с формированием димеров
рецептора. Активированная тирозинкиназа фосфорилирует различные белкимишени, что приводит к изменениям мембранного транспорта, транскрипции генов
и других клеточных процессов.
73.
74.
75.
76.
77.
Спасибо за внимание78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
Кклеточныйответ