


























 biology
biologySimilar presentations:




")





Генетика развития растений. Развитие листа
1.
Генетика развития растенийРазвитие =
рост + дифференцировка
Развитие клетки,
ткани, органа
(морфогенез)
2.
Развитиелиста
3.
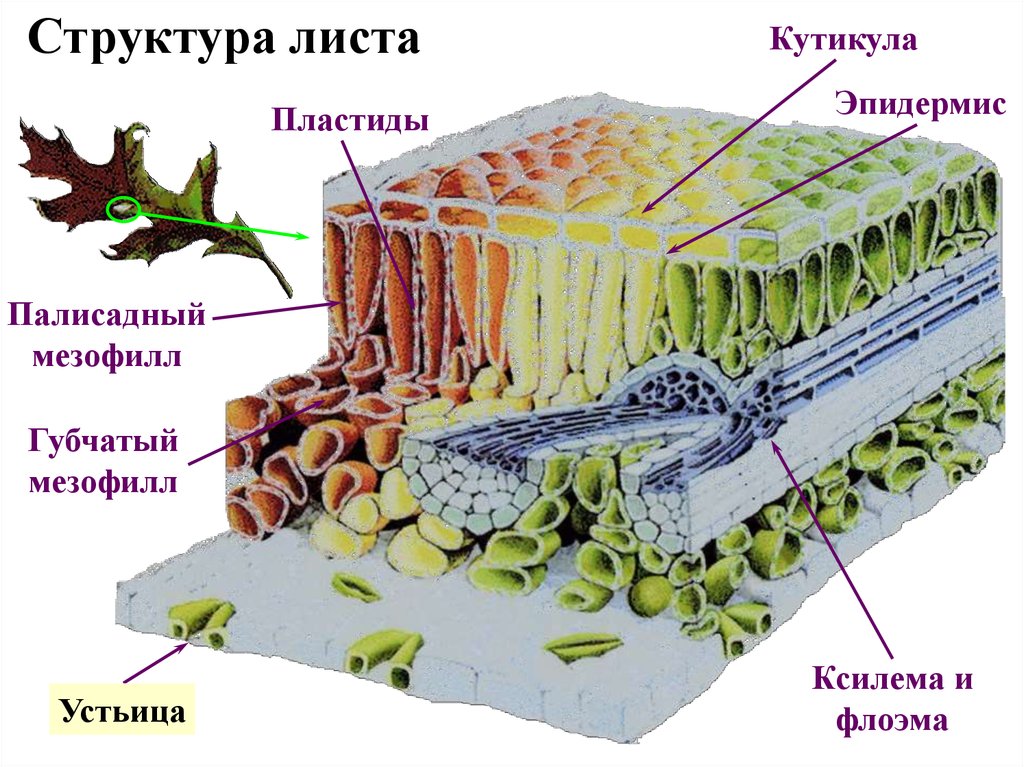
Структура листаПластиды
Кутикула
Эпидермис
Палисадный
мезофилл
Губчатый
мезофилл
Строма
Устьица
Ксилема и
флоэма
4.
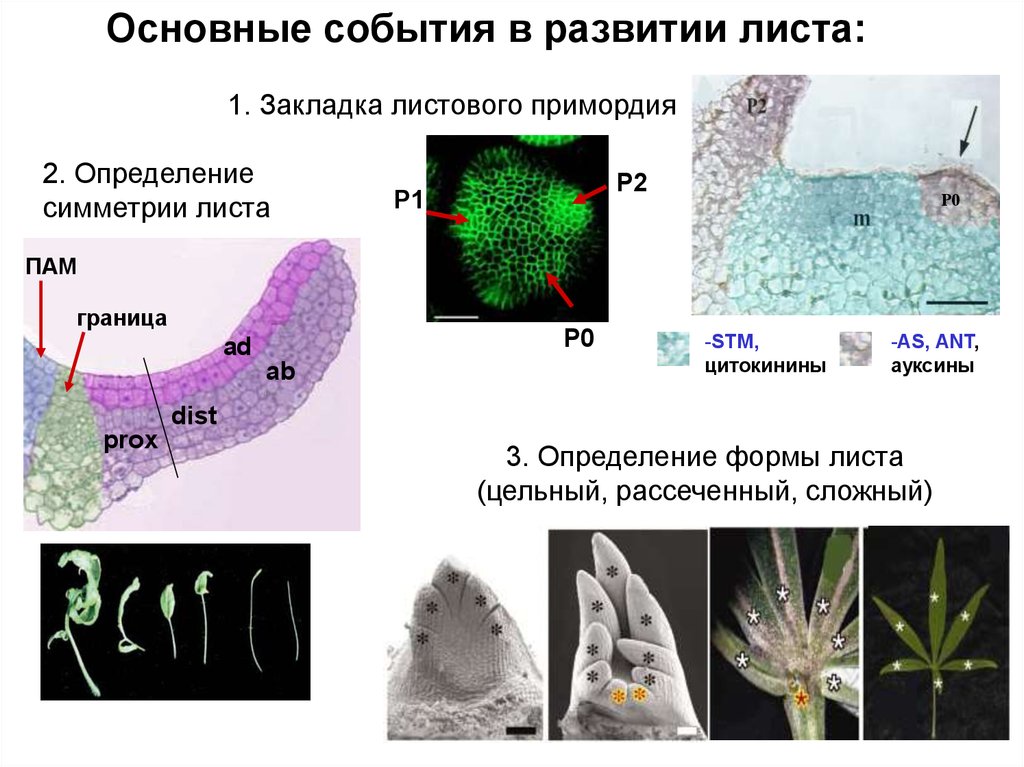
Основные события в развитии листа:1. Закладка листового примордия
2. Определение
симметрии листа
P2
P1
Р0
ПАМ
граница
P0
ad
ab
-STM,
цитокинины
-AS, ANT,
ауксины
dist
prox
3. Определение формы листа
(цельный, рассеченный, сложный)
5.
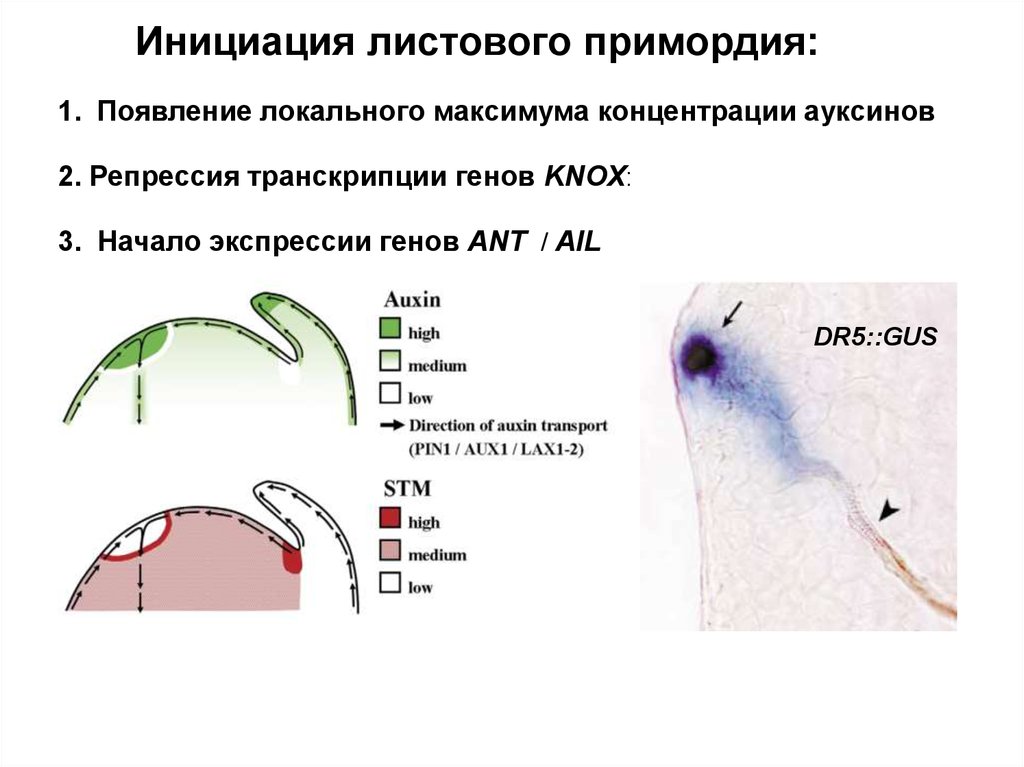
Инициация листового примордия:1. Появление локального максимума концентрации ауксинов
2. Репрессия транскрипции генов KNOX:
3. Начало экспрессии генов ANT / AIL
DR5::GUS
6.
Роль генов KNOX в регуляции развития листьев• В норме гены
KNOX
класса
I
никогда
не
экспрессируются в
листовых зачатках
STM
• KNAT1 и KNAT2
предположительно
играют основную
роль в развитии
листа
экспрессия KNOX
пролиферация клеток
• Возможно
возобновление
экспрессии KNOX генов
на более поздних стадиях
развития листа:
формирование сложных листьев
7.
Гены ASYMMETRIC LEAF (AS1 и AS2)– репрессоры KNOX генов в листовых примордиях
• Продукты генов AS –
Транскрипционные
факторы
семейства MYB (AS1)
семейства LOB (AS2)
STM
AS1, AS2
• Фенотипы
мутантов:
пролиферация
клеток
листового
зачатка,
нарушение
симметрии
листа
• Ортологи гена AS1 Arabidopsis:
RS2 (Zea mais),
PHAN (Anthirrinum majus)
8.
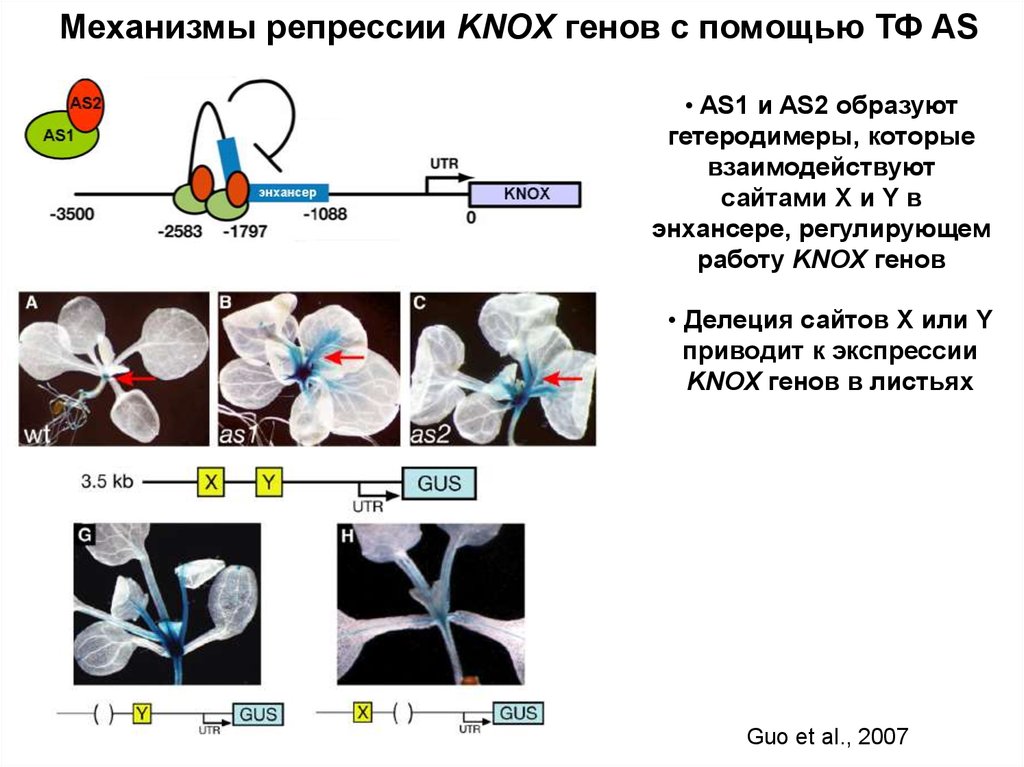
Механизмы репрессии KNOX генов с помощью ТФ AS• AS1 и AS2 образуют
гетеродимеры, которые
взаимодействуют
сайтами X и Y в
энхансере, регулирующем
работу KNOX генов
• Делеция сайтов Х или Y
приводит к экспрессии
KNOX генов в листьях
Guo et al., 2007
9.
Эпигенетические механизмы репрессии KNOX генов1. Кратковременная репрессия (только инициация примордия)
AS2
HDAC
AS1
HIRA
Ac
Ac
Деацетилирование
гистонов:
Гистоновый шаперон
HIRA,
взаимодействующий с
гистон-деацетилазами
(HDAC) и димерами ТФ
AS1-AS2
ring 1a, 1b
2. Долговременная репрессия (инициация и развитие)
1. Метилирование гистонов:
Белок CURLY LEAF (CLF) в
составе Polycomb
комплекса
2. Убиквитинирование
гистонов:
Белок LHP, Убиквитинлигазы RING1a и 1b
10.
Пролиферация клеток листового примордия: генAINTEGUMENTA (ANT)
ауксины
?
ANT
Циклины D ?
пролиферация
клеток
YABBY
Продукт гена ANT – ТФ
семейства APETALA2
?
симметрия
листа
ant
WT
35S::ANT
WT 35S::ANT
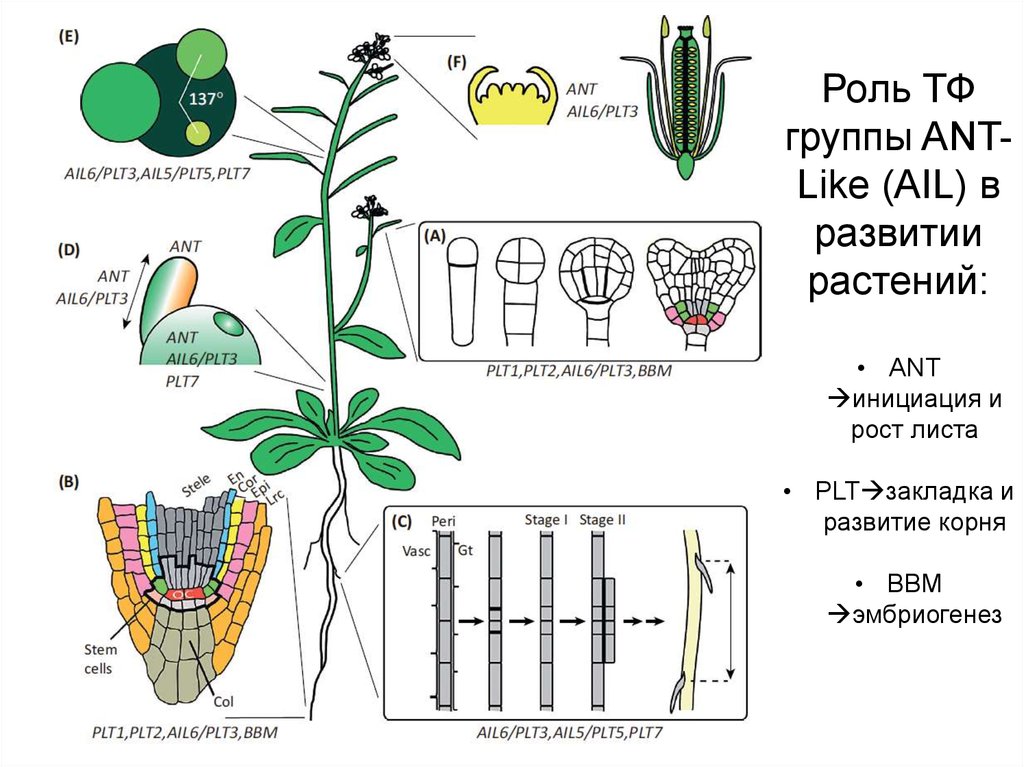
11.
Роль ТФгруппы ANTLike (AIL) в
развитии
растений:
• ANT
инициация и
рост листа
• PLT закладка и
развитие корня
• BBM
эмбриогенез
12.
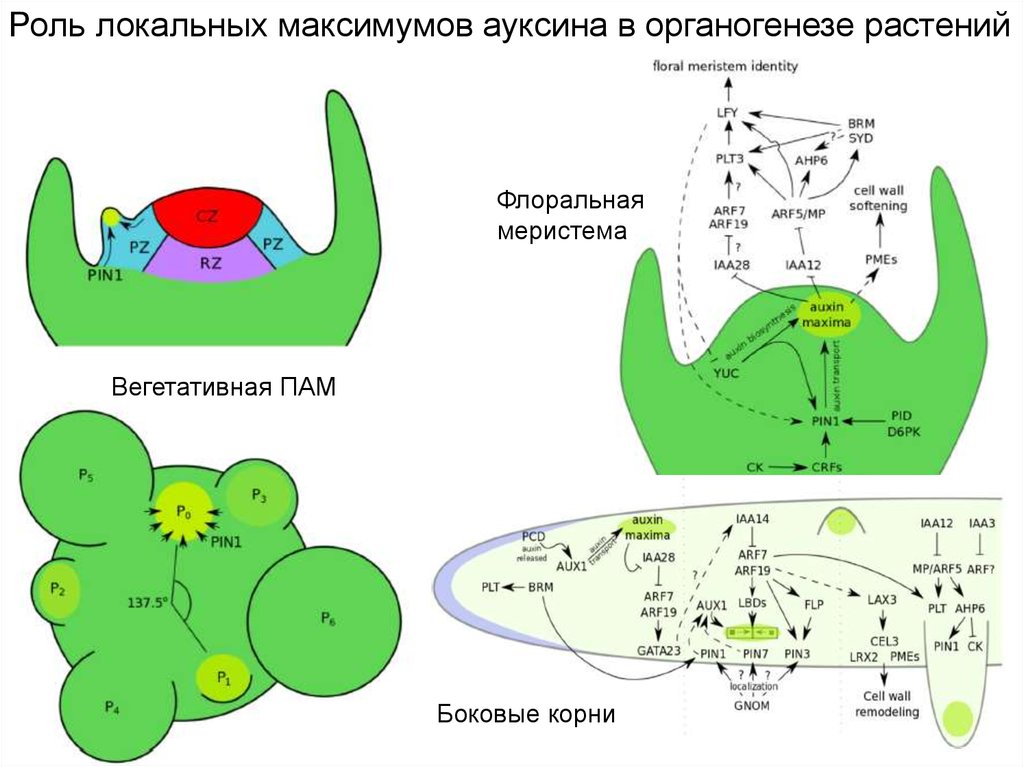
Ауксины – ключевые регуляторы инициации листаЛокальные максимумы
концентрации ауксинов
Транспорт
ауксинов в ПАМ
Рецепция и
передача сигнала
ауксинов
Белки PIN
Синтез ауксинов
в молодых
листьях
ТФ ARF
гены Н-АТФаз,
гены экспансинов
растяжение клеток
изменение
направления
деления
ген AS2
репрессия генов
KNOX
дифференцировка
клеток
(Auxin
Response
Factors)
???
экспрессия
генов PIN,
VND, CND
ограничение зон
экспрессии AS и
ANT
отделение
примордиев от ПАМ
13.
Роль локальных максимумов ауксина в органогенезе растенийФлоральная
меристема
Вегетативная ПАМ
Боковые корни
14.
Полярный транспорт ауксинов (ПАТ) и инициация листаНаправление ПАТ задают белки
PIN, обеспечивающие выход
ауксинов из клетки в
определенном направлении
Для PIN белков
характерна
полярная
локализация на
мембране
PIN
Мутанты pin
(pinformed)
формируют
очень мало
листьев и
цветков
15.
Полярный транспорт ауксинов и инициация листаГены PIN1 и STM имеют
комплементарный
характер экспрессии в
ПАМ
P2
pDR5::3XVENUS-N7
P1
P0
pPIN1::GFP
Направления ПАТ
в листовых
примордиях
pDR5::3XVENUS-N7
pPIN1::GFP
16.
Ауксины контролируют развитие проводящей системы листаDR5::GUS
Canalisation hypothesis:
Локальный
максимум
ауксинов
Создание канала ПАТ:
Включение
Транспорт ИУК
экспресcии PIN1
по каналу от
в группе клеток
максимума
«под»
максимумом
Клетки канала
ПАТ
дифференцируются в сосуды
17.
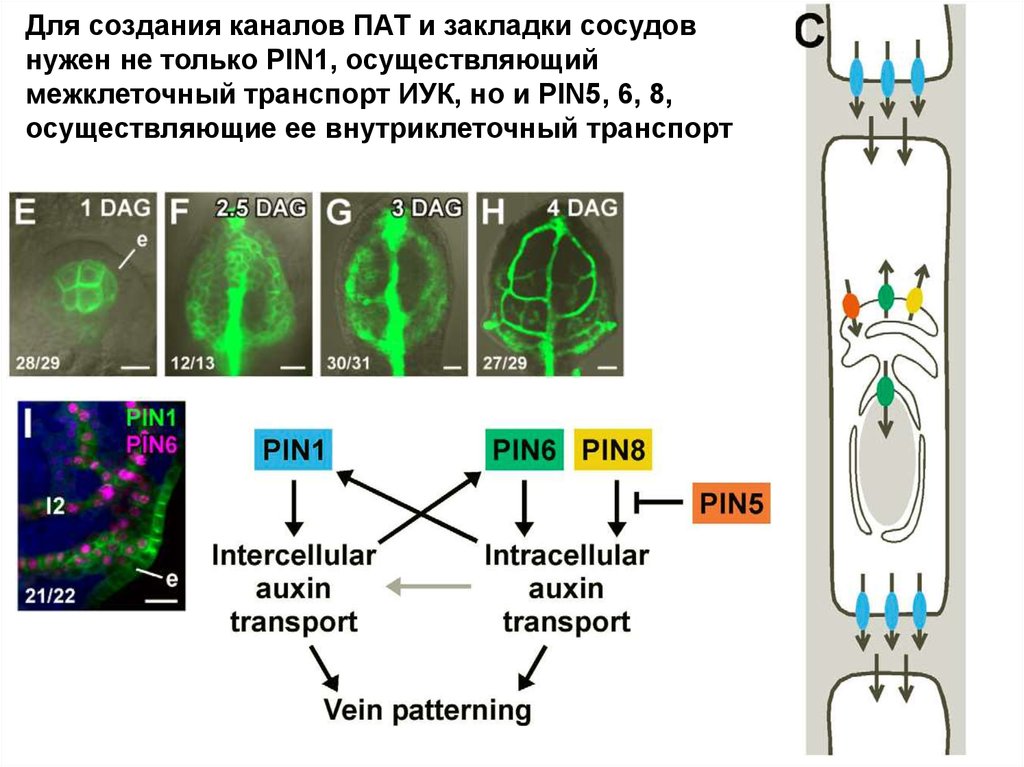
Для создания каналов ПАТ и закладки сосудовнужен не только PIN1, осуществляющий
межклеточный транспорт ИУК, но и PIN5, 6, 8,
осуществляющие ее внутриклеточный транспорт
18.
АуксиныAS2
Репрессия KNOX
???
Ограничение зон экспрессии
AS1, AS2, ANT
WT
KNAT1::GUS
MS0
ANT
20mM
TIBA
AS1
mp pin1
19.
Локальнаяиндукция
экспрессии
гена EXP1
Закладка
листовых
примордиев
Формирование
листьев
Ауксины стимулируют закладку
листьев через активацию
мембранных Н-АТФаз и
экспансинов
20.
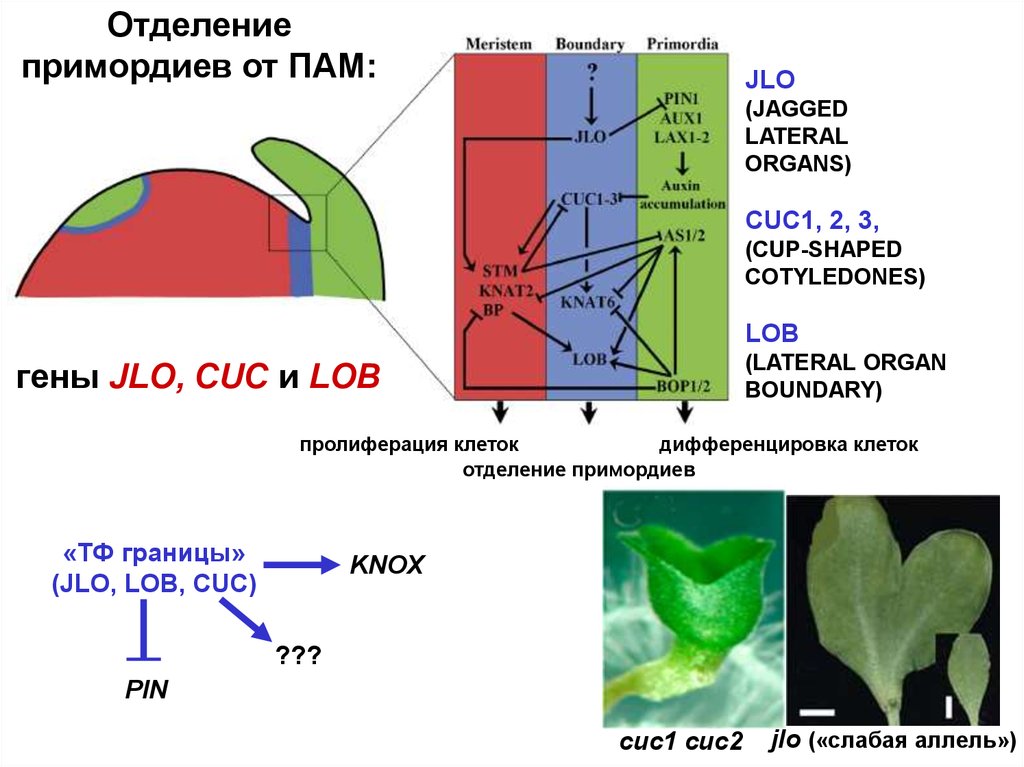
Отделениепримордиев от ПАМ:
JLO
(JAGGED
LATERAL
ORGANS)
CUC1, 2, 3,
(CUP-SHAPED
COTYLEDONES)
LOB
(LATERAL ORGAN
BOUNDARY)
гены JLO, CUC и LOB
пролиферация клеток
дифференцировка клеток
отделение примордиев
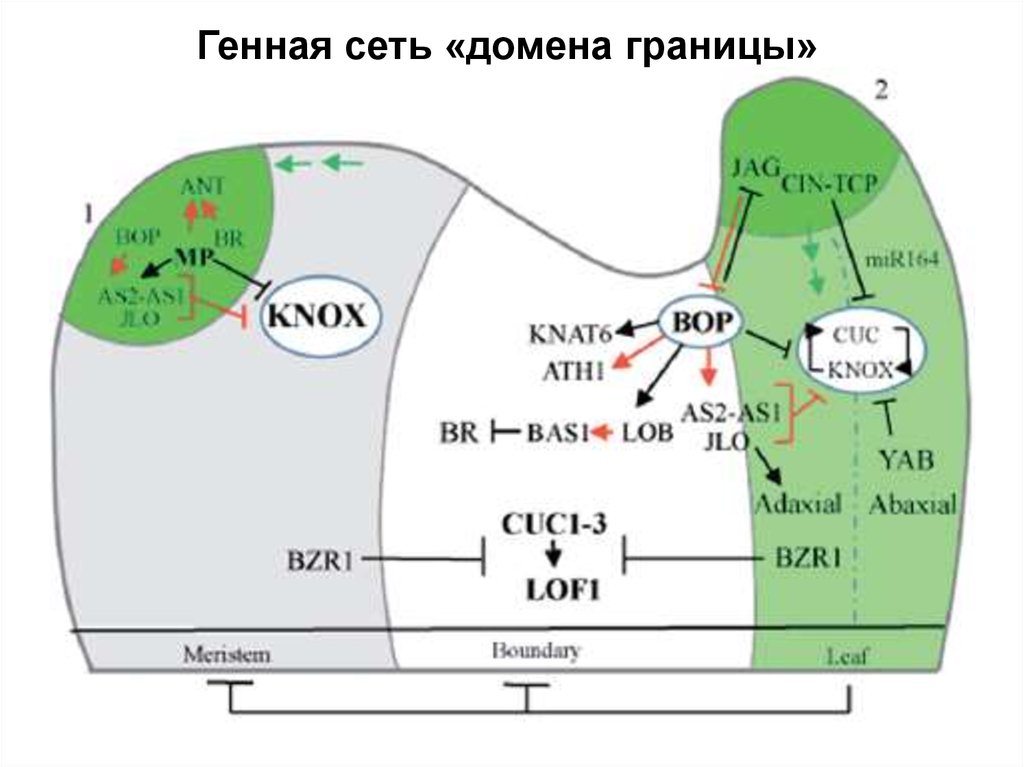
«ТФ границы»
(JLO, LOB, CUC)
KNOX
???
PIN
cuc1 cuc2
jlo («слабая аллель»)
21.
Генная сеть «домена границы»22.
«ТФ границы» важны на всех этапах развития растенияЗакладка ПАМ в
эмбриогенезе
Закладка флоральной
меристемы
Закладка пазушных
меристем
Закладка лопастей
сложного или
рассеченного листа
Закладка органов цветка (завязь)
23.
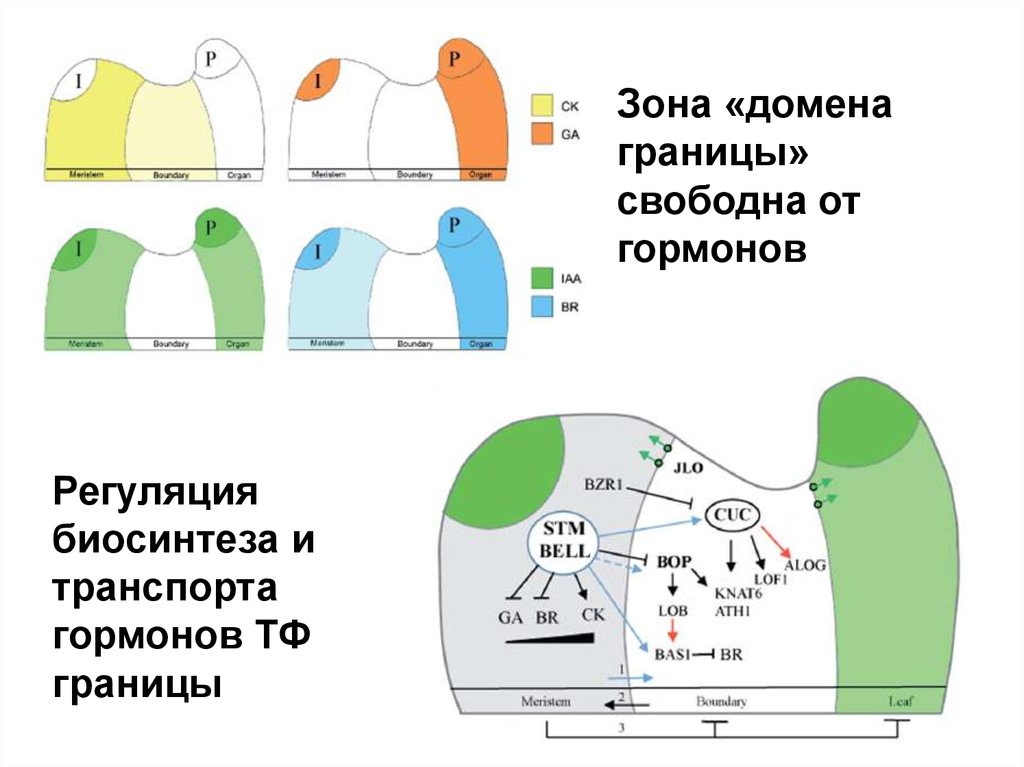
Зона «доменаграницы»
свободна от
гормонов
Регуляция
биосинтеза и
транспорта
гормонов ТФ
границы
24.
leafИммунолокализация:
гистон H4
(маркер
деления
клеток)
CUC2
leaflet
Домен границы
есть как в листовых
примордиях (А), так
и в примордиях
компонентов
сложного листа (В)
25.
Определение симметрии листа –разметка доменов
Идентичность разных доменов контролируют определенные ТФ
26.
Идентичность проксимального и дистального доменовлиста - гены BLADE ON PETIOLE (ВОР) и JAGGED (JAG)
ad
ab
prox
dist
BOP2
ВОР1
ВОР2
KNOX
JAG
LOB
bop1
WT
bop1
WT
JAG
27.
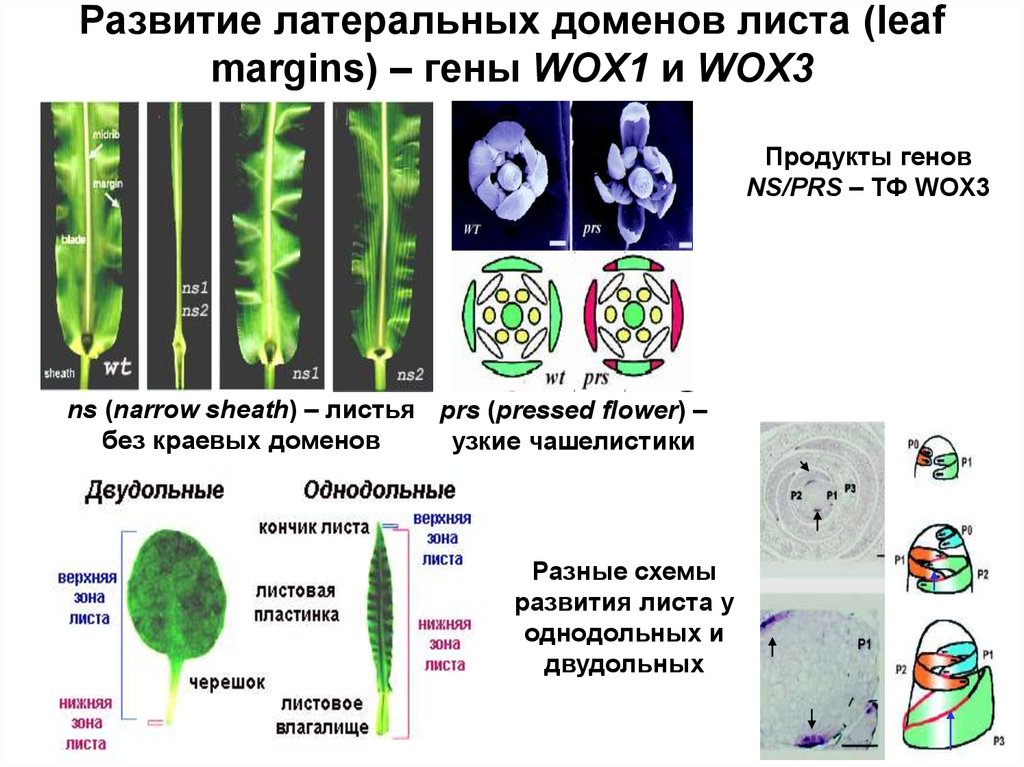
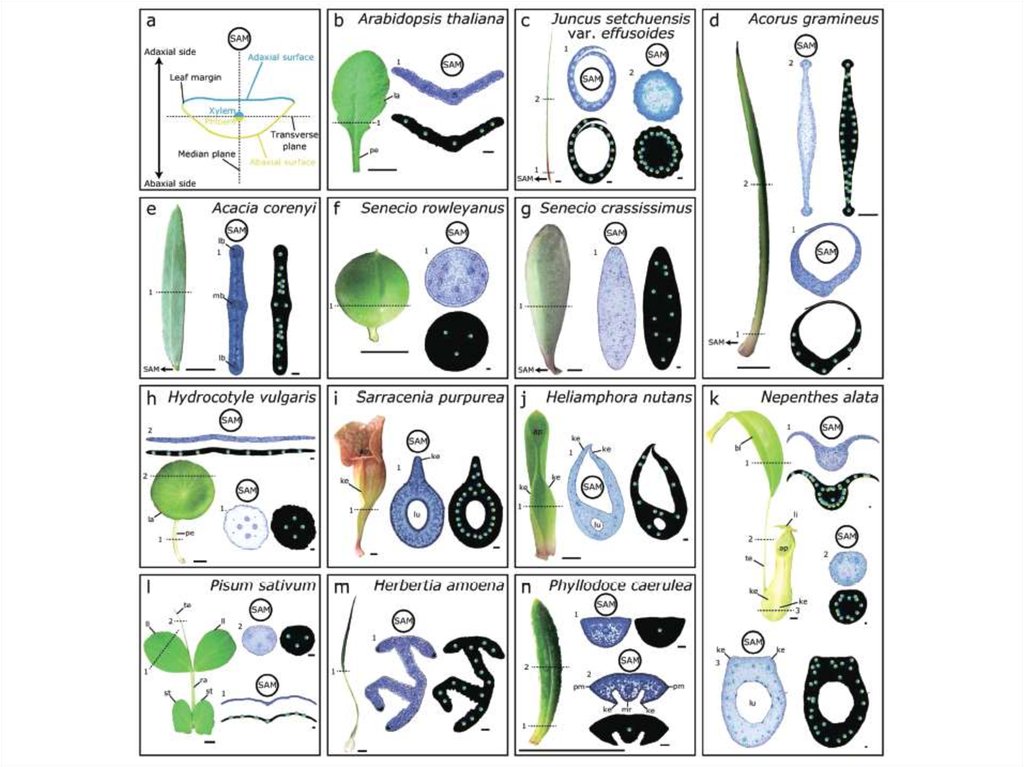
Развитие латеральных доменов листа (leafmargins) – гены WOX1 и WOX3
Продукты генов
NS/PRS – ТФ WOX3
ns (narrow sheath) – листья prs (pressed flower) –
без краевых доменов
узкие чашелистики
Разные схемы
развития листа у
однодольных и
двудольных
28.
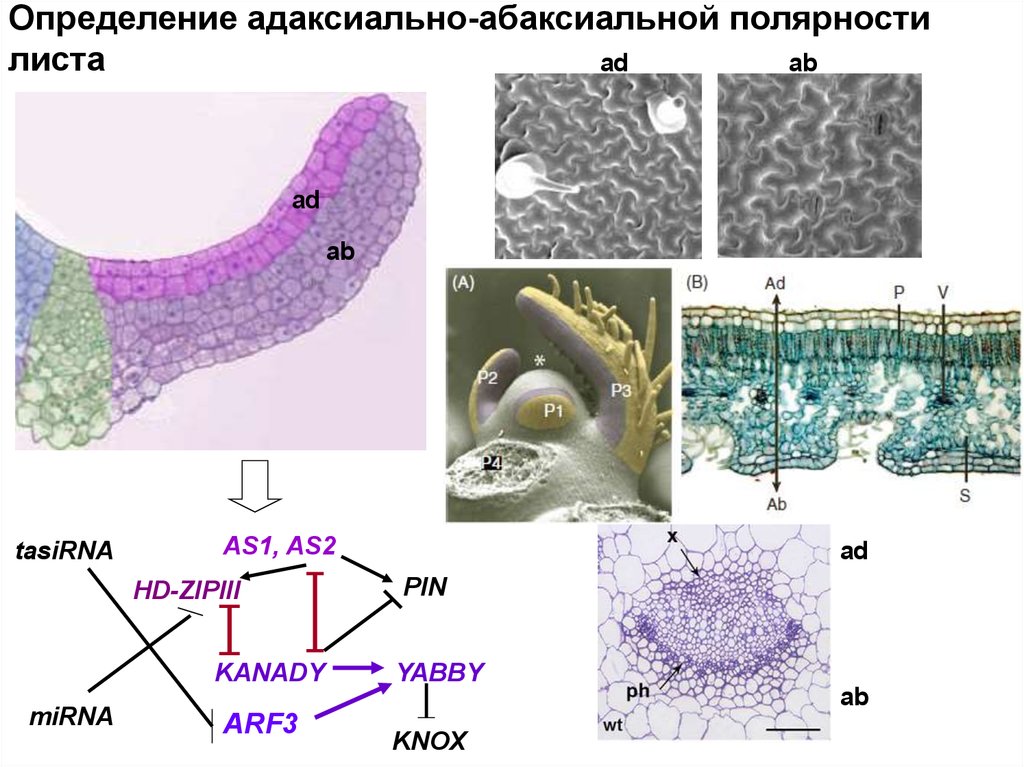
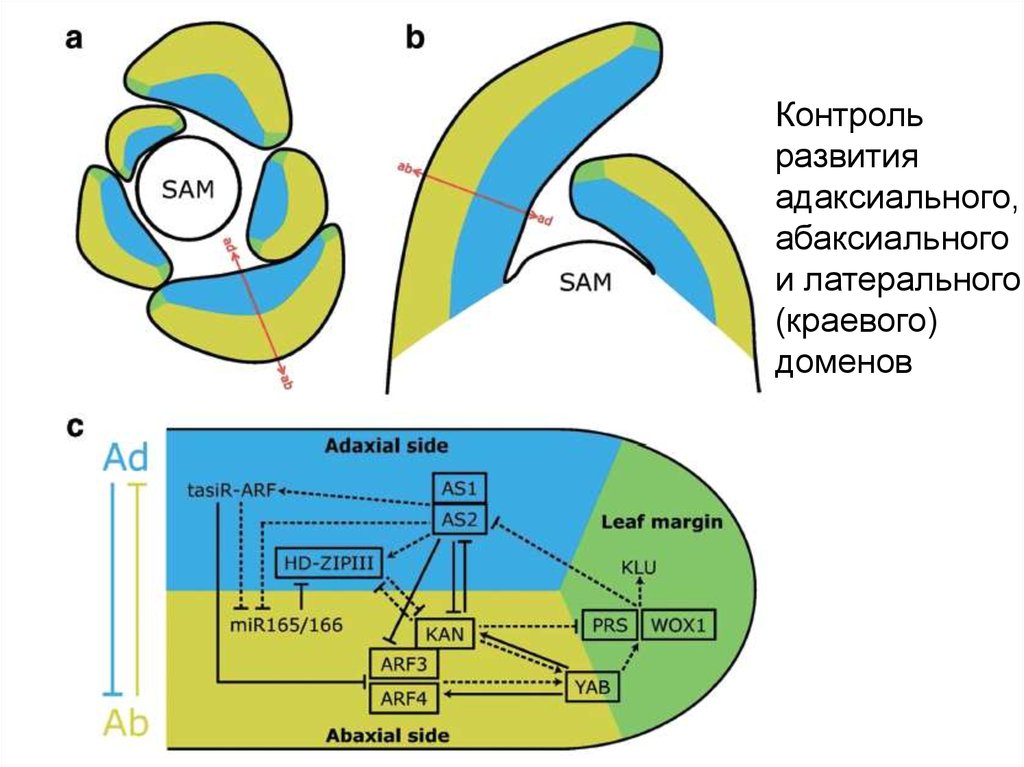
Определение адаксиально-абаксиальной полярностилиста
ad
ab
ad
ab
ad
tasiRNA
AS1, AS2
HD-ZIPIII
KANADY
miRNA
ad
PIN
YABBY
ab
ARF3
KNOX
29.
Контрольразвития
адаксиального,
абаксиального
и латерального
(краевого)
доменов
30.
Форма листа зависитот соотношения
ad и ab сторон
Регуляция ad-ab
полярности листа:
Взаимодействие
разных семейств
ТФ и миРНК
31.
32.
HD-ZIPIII:PHABULOSA (PHB),
PHAVOLUTA (PHV),
REVOLUTA (REV)
KANADY:
KANADY 1-4 (KAN1-KAN4)
(ТФ семейства GARP)
phb phv
kan1-4
p
phb
Le-yab
YABBY:
YABBY2 (YAB2),
YABBY3 (YAB3),
FILAMENTOUS FLOWER
(FIL)
HD-ZIPIII
KANADY, YABBY
x
PHB
33.
Антагонизм транскрипционных факторов KAN иHD-ZIPIII в контроле распределения белка PIN1
закладка и развитие
листьев
транспорт ауксинов
HD-ZIPIII
PIN
KANADY
34.
Другие регуляторы адаксиально-абаксиальнойполярности листа
tasiRNA3 ad
ТФ AS1, AS2 ad
ТФ ANT ab
ТФ ARF3/4
ab
miRNA165/166 ab
35.
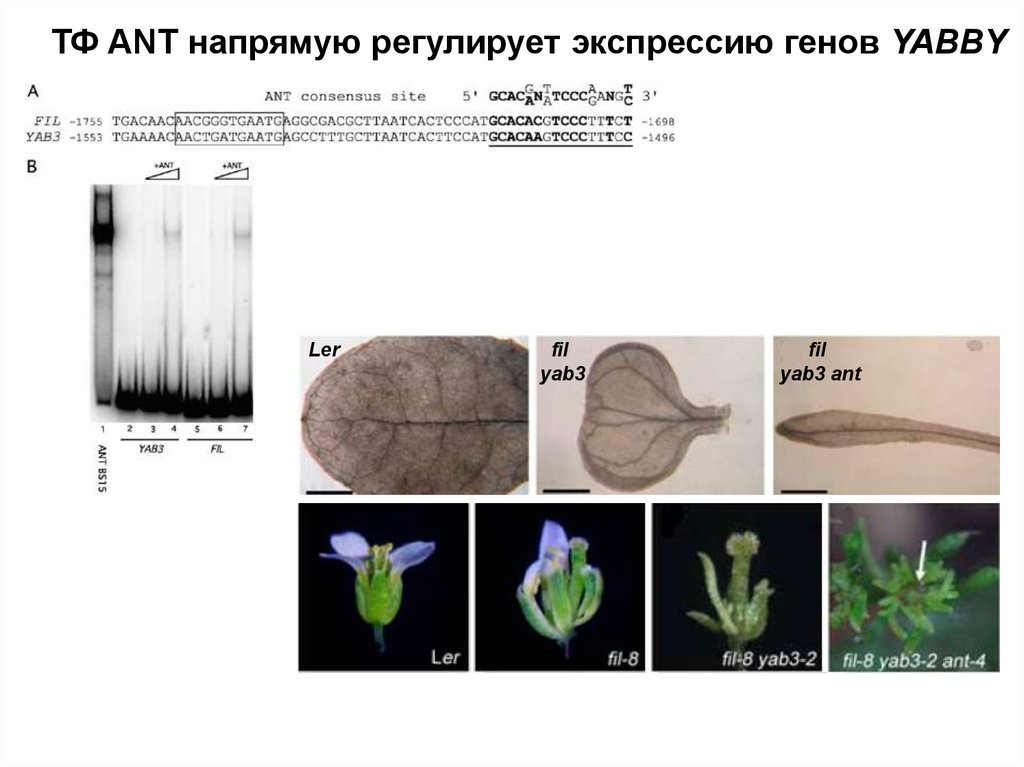
ТФ ANT напрямую регулирует экспрессию генов YABBYLer
fil
yab3
fil
yab3 ant
36.
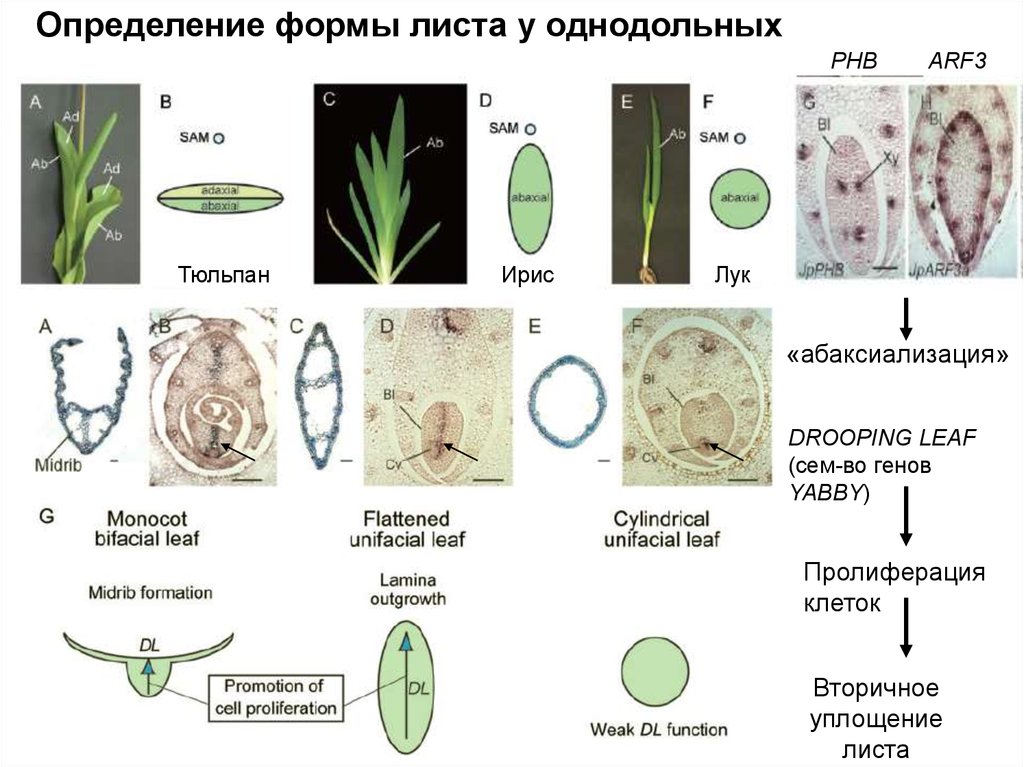
Определение формы листа у однодольныхPHB
Тюльпан
Ирис
ARF3
Лук
«абаксиализация»
DROOPING LEAF
(сем-во генов
YABBY)
Пролиферация
клеток
Вторичное
уплощение
листа
37.
МикроРНК в определении ad-ab полярностиtasiR3a – репрессор ab стороны. Мишень:
транскрипты ARF3
miR166 – репрессор ad стороны. Мишень:
транскрипты НD-ZIPIII
38.
Другие миРНК,контролирующие развитие
листа
39.
При развитии лопастей сложного листадействуют те же механизмы, что при закладке
листовых примордиев в ПАМ
PIN
KNOX
EXP
PIN
miR164
1.
2.
3.
4.
Возобновление экспрессии KNOX генов
Создание локального максимума концентрации ауксинов
Определение границы
Определение полярности
40.
Для развития сложного листа необходимовозобновление экспрессии генов KNOX в примордии
WT
WT
35S::KNAT1
35S::LeT6
41.
Для развития сложного листа необходимовозникновение локальных максимумов концентрации
ауксинов в краевых доменах
PIN
транспорт ауксинов
локальный максимум
концентрации ауксинов
активация экспансинов,
рост клеток,
изменение направления деления
EXP
закладка лопасти
42.
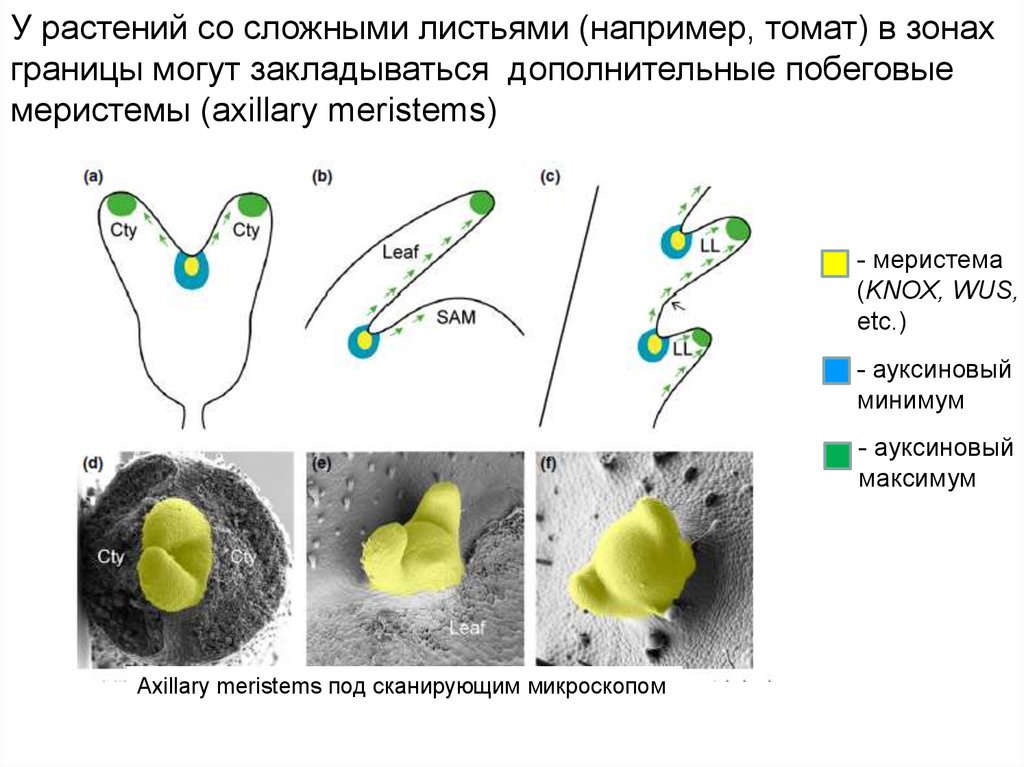
У растений со сложными листьями (например, томат) в зонахграницы могут закладываться дополнительные побеговые
меристемы (axillary meristems)
- меристема
(KNOX, WUS,
etc.)
- ауксиновый
минимум
- ауксиновый
максимум
Axillary meristems под сканирующим микроскопом
43.
Определение формы сложного листаmiR164
miR164 ov
CUC2
44.
Мутации, приводящиек упрощению листа у
томата
gob (goblet)
c (potato leaf)
Продукты генов GOB
и С – ортологи CUC