
































































































































 biology
biologySimilar presentations:









")
О структуре клетки и механизмах ее воспроизведения
1.
Исходя из сегодняшних знаний о структуре клетки имеханизмах ее воспроизведения, проблему возникновения
жизни надо рассматривать через проблему возникновения
клетки, а не ее фрагментов.
Клетка
это
единый,
жидкокристаллический,
апериодический,
гомеостатический комплекс,
состоящий из десятков и сотен тысяч разнообразных
молекул нуклеиновых кислот, белков, углеводов и липидов,
работающих как единый механизм, управляемый едиными
биохимическими
процессами,
самовоспроизведению,
имеющий
генетический
код,
информации,
независимо
прокариотического
способный
близкие
или
от
универсальный
механизмы
реализации
уровня
организации
эукариотического
Отдельные фрагменты данного
к
типа.
кмплекса, в том числе
вирусы или фаги, не являются жизнеспособными. Поэтому,
ответив на вопросы характеризующие клетку, как единую
структуру мы сможем понять, что такое жизнь и как она
возникла. Пока на поставленные вопросы ответа нет.
2.
3 часть –что такое Апатит,
свойства Апатита,
почему для изучения происхожденя жизни
был выбран Апатит.
Теоретически возникновение клетки может быть
следствием
различной
событий.
же
Мы
последовательности
остановимся
на
событиях,
которые реально, одномоментно могли привести
к образованию протоклетки со всеми ее органоминеральными структурами
специфической
в результате их
агрегации
минеральных кристаллов
в
структуре
в соответствии с
физико-химическими законами ,
путем диффузии
свободных
газовой
радикалов
и
ионов
фазы
в
кристаллическую решетку. Полученная протоклетка, как
и ее прародительница (матрица минералов), оказалась
способной
к
самовоспроизведению
транскрипционно-трансляционного аппарата.
за
счет
3. ПРОИСХОЖДЕНИЕ ПРОТОКЛЕТОК И ИХ СТРУКТУРНЫХ ЭЛЕМЕНТОВ НА ОСНОВЕ АПАТИТА И СОКРИСТАЛИЗУЮЩИХСЯ С НИМ МИНЕРАЛОВ
Посвящается памятиВ.В.Чернобровкина
ПРОИСХОЖДЕНИЕ ПРОТОКЛЕТОК И
ИХ СТРУКТУРНЫХ ЭЛЕМЕНТОВ НА
ОСНОВЕ АПАТИТА И
СОКРИСТАЛИЗУЮЩИХСЯ С НИМ
МИНЕРАЛОВ
• Костецкий Э.Я.
Отдел Биохимии и Биотехнологии, Дальневосточного
Государственного университета. Владивосток, 690950, Октябрьская
27. Факс: 7-4232-429510. Е-mail: kostetsky@nt.pin.dvgu.ru
• «Теория оказывается тем более впечатляющей,
чем проще ее предпосылки, чем значительнее
разнообразие охватываемых ею
явлений и чем
шире ее применимость»
• А. Эйнштейн
• «Мир слишком совершенен,
возникнуть из хаоса»
чтобы
случайно
4.
Апатит в диопсиде Ковдор5.
Апатит кристаллы с кальцитом. Слюдянка.Сопутствующие минералы. Сфен, циркон, пироксен,
амфибол, шпинель, везувиан, флогопит, нефелин, магнез
ит, кварц, полевой шпат, слюды, кальцит.
6.
7.
8.
Апатит впороде
9.
ВАЖНЫЕ КЛЮЧЕВЫЕ МОМЕНТЫ по апатитуЧто такое апатит?
Слово апатит греческого происхождения и обозначает «обманываю», так
как апатит очень часто путали со многими минералами. Относится к
группе полудрагоценных минералов.
1. Апатит единственный из 1500 минералов - источник фосфора на
поверхности Земли, без которого живые системы не бывают.
2. Апатит занимает особое место в огромном минеральном мире, ибо
находится на грани живой и неживой природы.
3. Проблема включения фосфора в органические соединения живых
систем (нуклеиновые кислоты, фосфорсодержащие белки и
другие соединения) связана с его пребиотической локализацией в
апатите.
4. Современные обитатели Земли наряду с апатитом включают в
состав
минерализованных
структур
кальцит,
арагонит
и
кристобалит, которые имеют с ним систему согласованных
пропорций.
5. Апатит является минеральной составляющей костей и зубов
позвоночных животных и человека, а также основой многих
патологических обызвествлений.
6. Апатит в составе позвоночника это место где формируются
стволовые клетки.
10.
7. Элементарная ячейка апатита (с6,879Å)
и
двойная
спираль
параметром по с =
ДНК
имеют
сходную
периодичность в 3,4 А°.
8. Апатит- это кристаллы способные к изоморфизму, без
изменения, основных параметров своей кристаллической
структуры, что обеспечивает единство метаболизма и будущее
разнообразие живых систем сформированных на их основе.
9.
Для живых систем апатит может быть источником РО4, Са, Si,
Sr, Mg, Ge, Fe, Mn, Zn, Cu, K, Na и др. , а спиральное расположение
фосфатных групп вдоль гексагональной оси в элементарной ячейке
апатита обеспечивает спиральность будущей ДНК.
10. Кристаллохимические особенности апатита по своим
параметрам допускают одновременный синтез на свей
матрице цепей ДНК в комплексе с основными белками
(колаген, протамин и др.) или их раздельный синтез.
11.
Для
обоснования
предложенной
теории
были
сопоставленны данные рентгеноструктурного анализа апатита
с одной стороны [54,88],цепей ДНК Z-формы [90–91], цепей
коллагена, полиглицина II и полипролина II [76–79] – с
другой;построена и проанализирована модель апатита с
фрагментом ДНК Z-формы, в том числе компьютерная.
11.
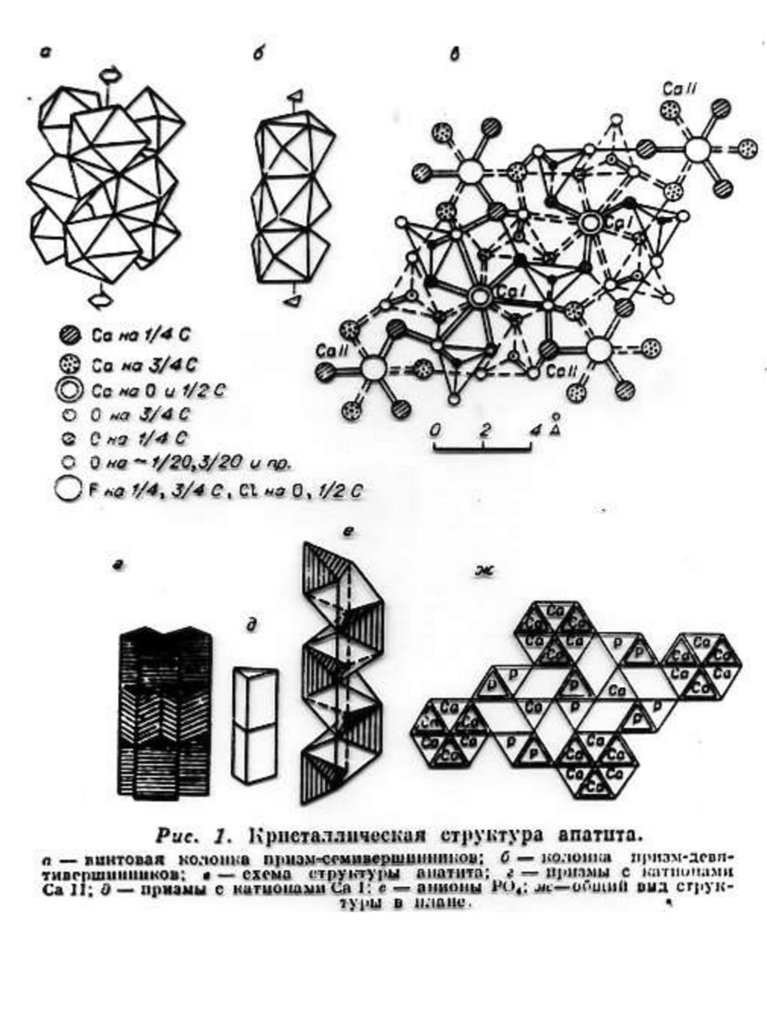
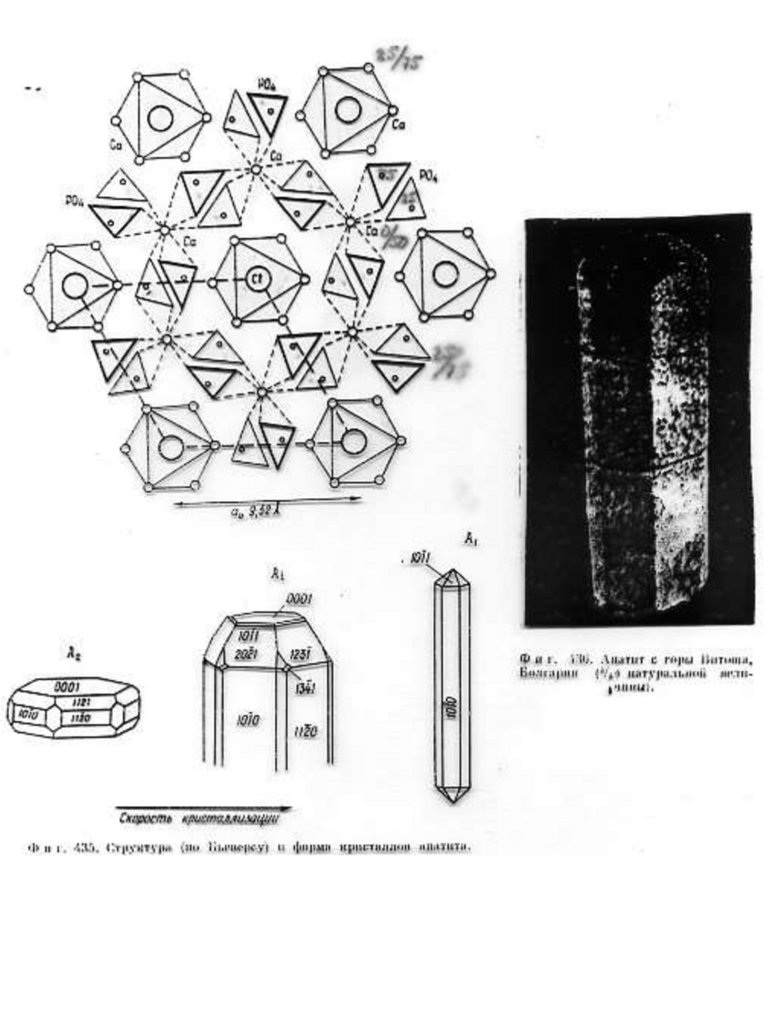
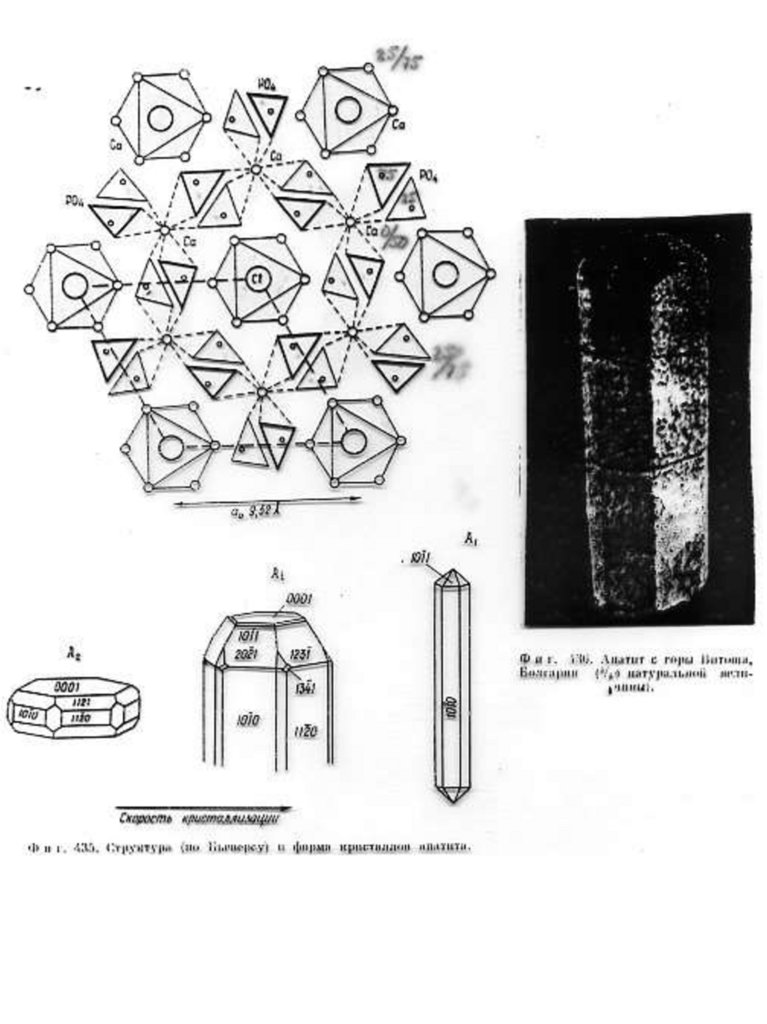
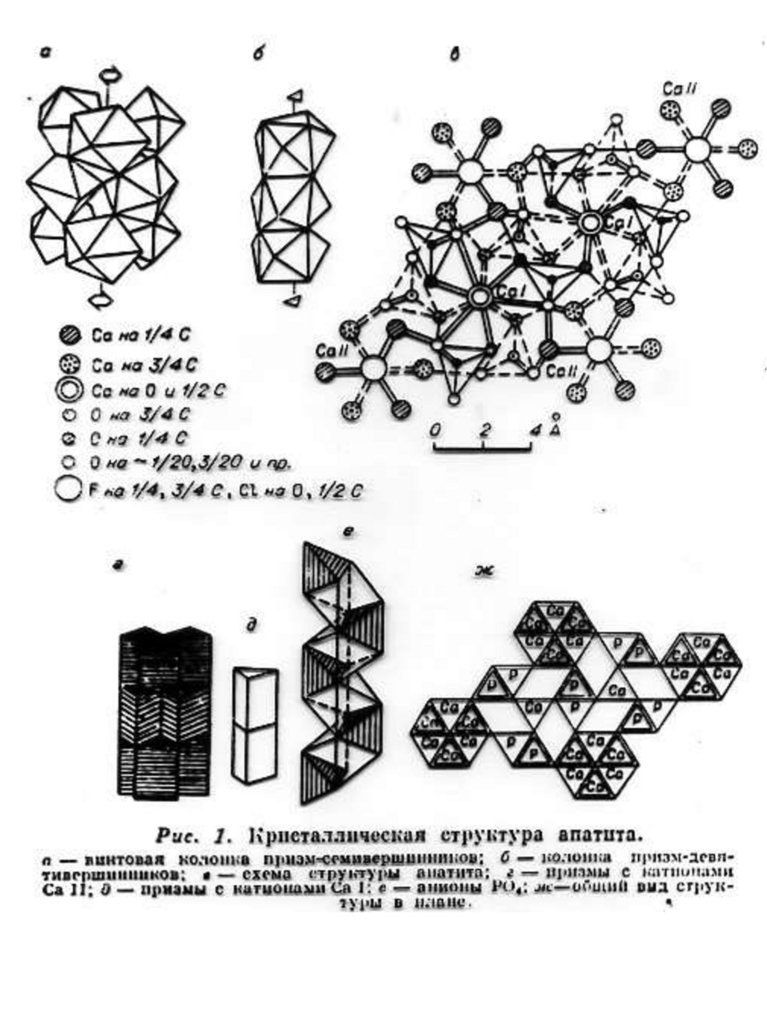
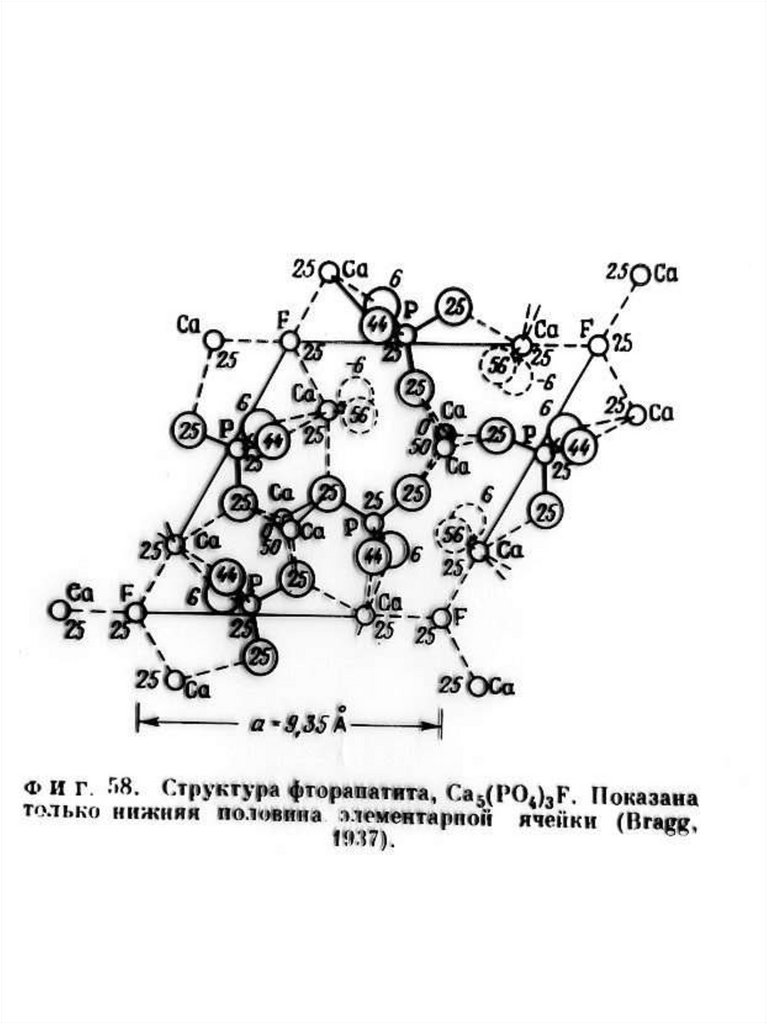
Краткое описание структурной модели апатита. Структура апатита была определена в 1930 г академиком Н.В.
Беловым. Идеализированная формула апатита Са10(PO4)6F2.
В элементарной ячейке содержится двойной набор формульных
единиц ( 42 атома), еѐ параметры равны: а = 9,364 Å, с = 6,879Å.
Структура апатита является гексагональной. На рис. 1. приведено
схематическое изображение структуры, предложенное Бъеверсом и
академиком Н.В. Беловым.
12.
Изредка апатит встречается в виде окаменевшего материалаископаемой древесины.
13.
14.
ДБ
В
Г
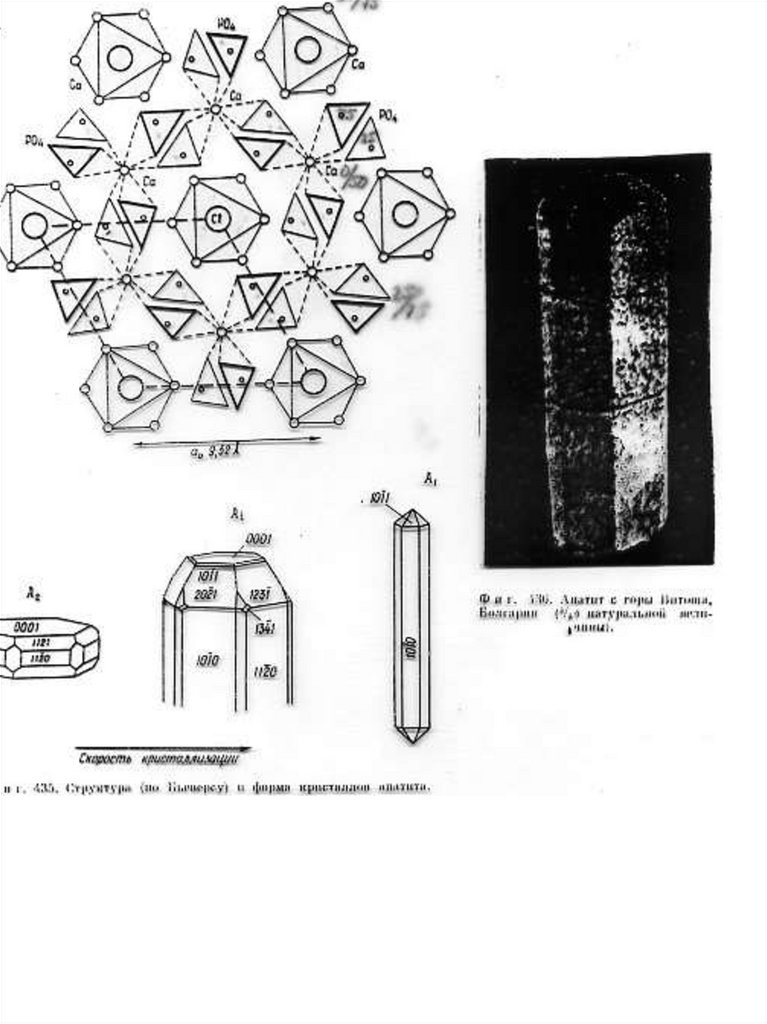
Рис.1. Б, В, Г - Cтруктура апатита по Н.В. Белову [Gilinskaya, Scherbacova 1975]:
Б - призмы с катионами Са II; В - призмы с катионами Са I; Г - анионы РО4. Д Апатит с горы Витоша, Болгария (4/5 натуральной величины). Числа указывают
доли от высоты элементарной ячейки.
15.
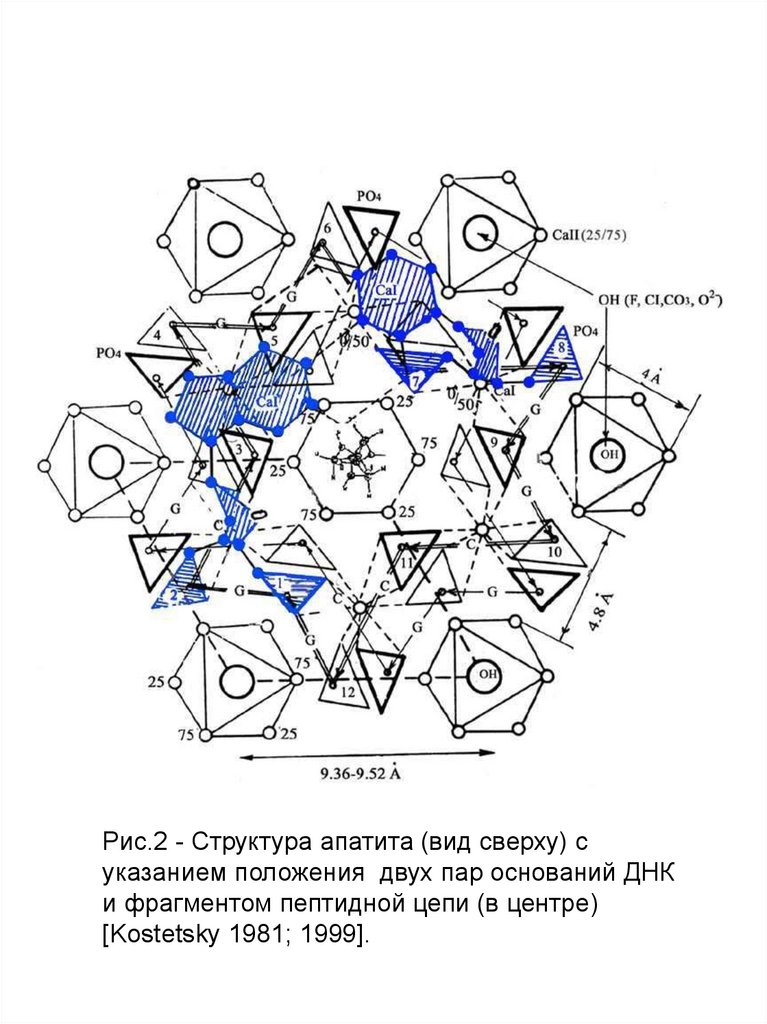
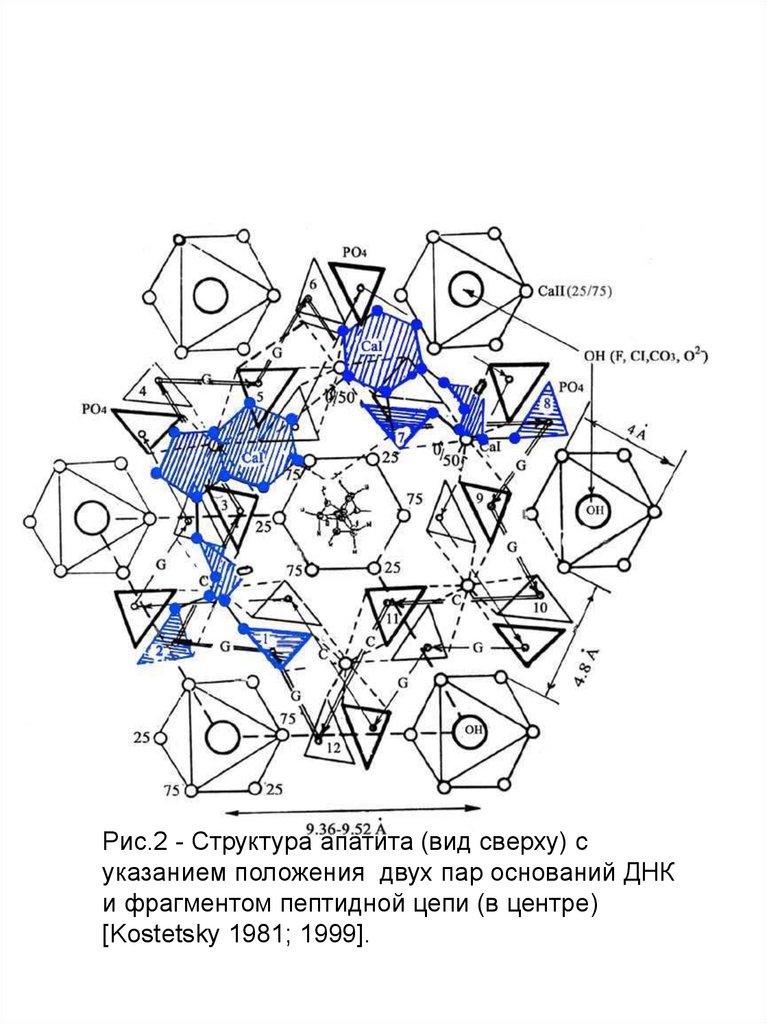
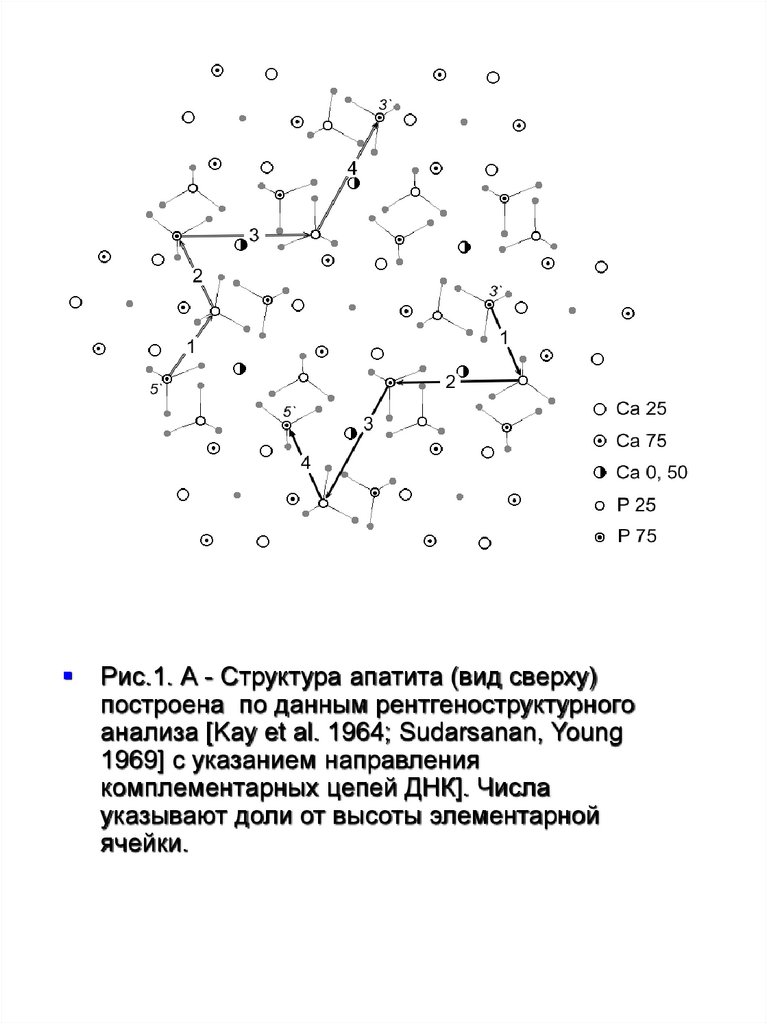
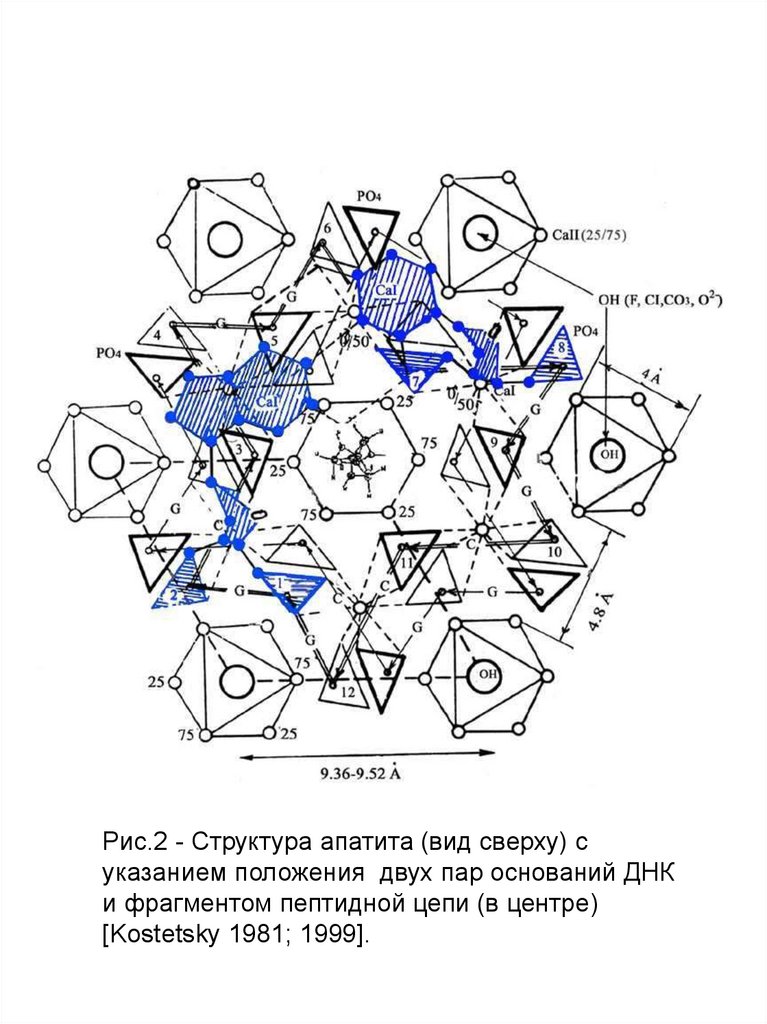
Рис.2 - Структура апатита (вид сверху) суказанием положения двух пар оснований ДНК
и фрагментом пептидной цепи (в центре)
[Kostetsky 1981; 1999].
16.
Изображение на рис.1 и 2 основано на такназываемых Паулинговских полиэдрах, в каждом углу
которых расположены атомы кислорода. В этой схеме
имеются три типа полиэдров: Са I, Са II (в
соответствии с двумя катионными позициями в
структуре) и (PO4)3- . Изолированные ортофосфатные
группы расположены вдоль оси второго порядка
между колонками Са I и Са II. Атомы фтора находятся в
центре Са II колонок вдоль гексагональной оси.
17.
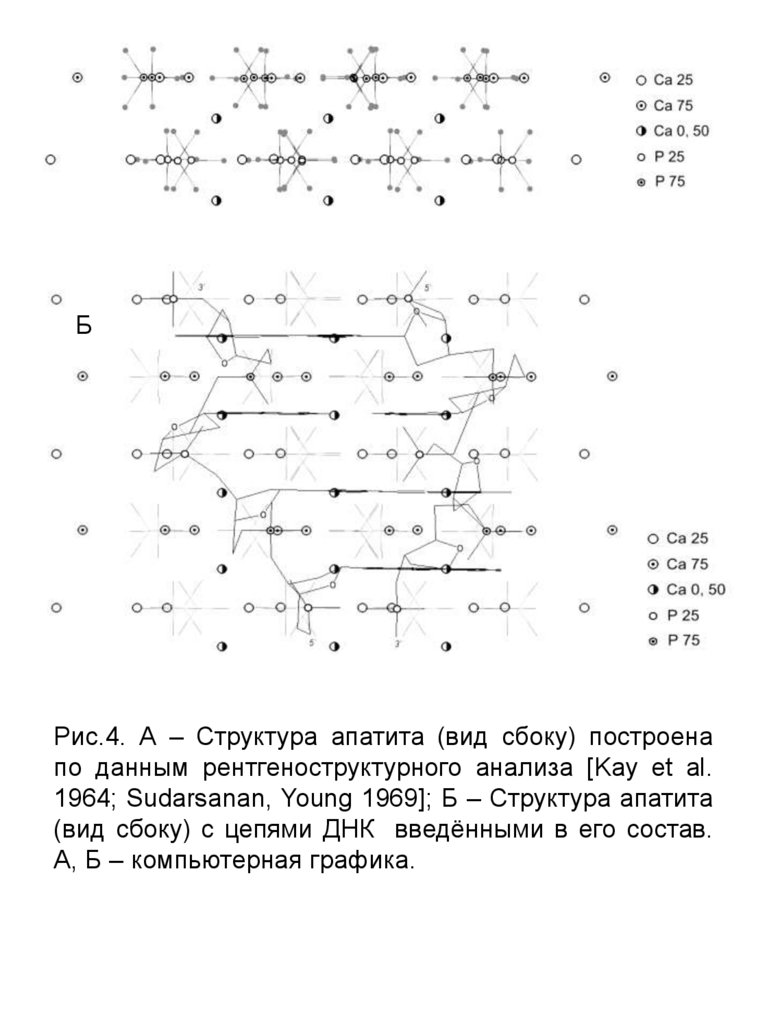
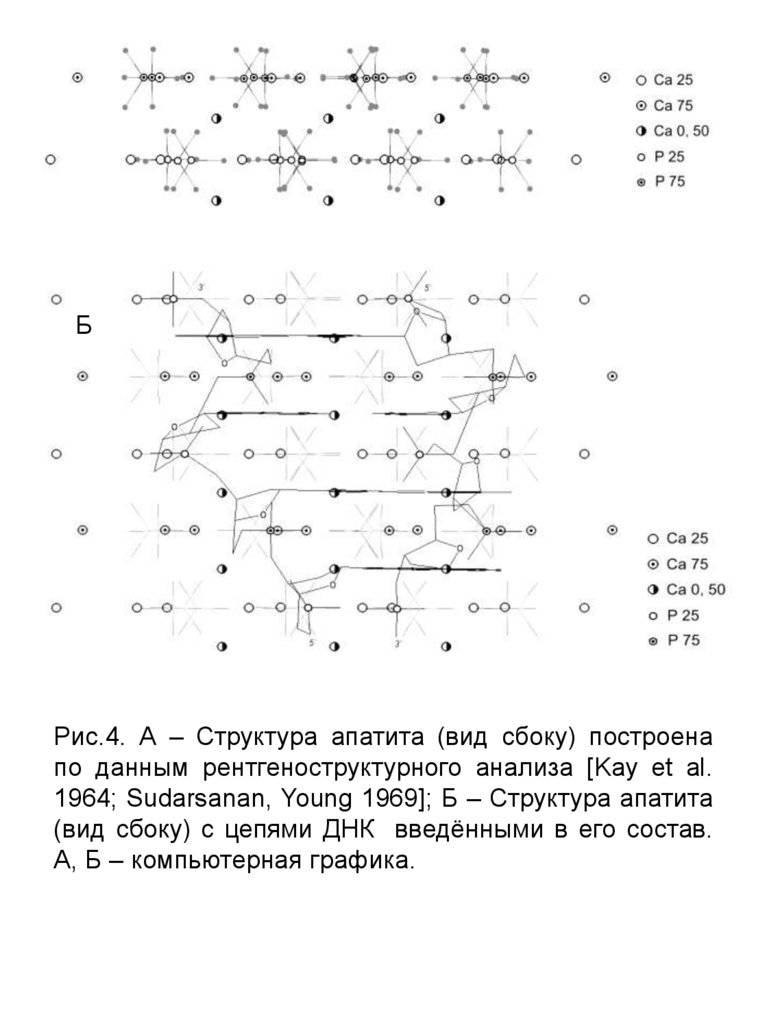
АБ
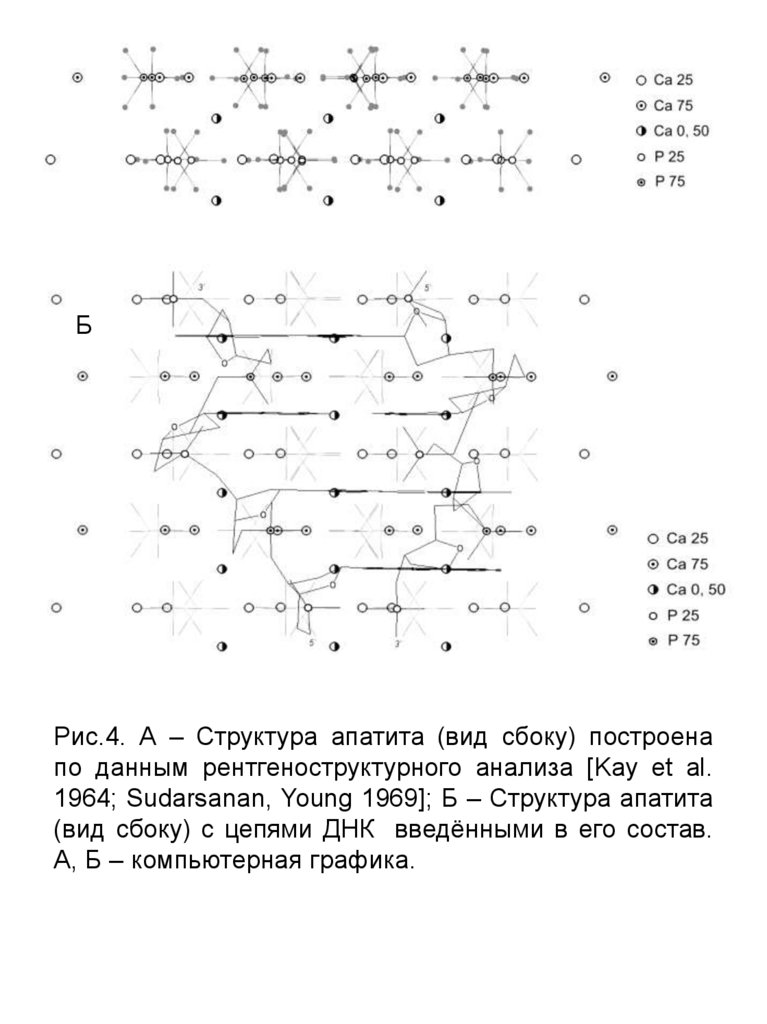
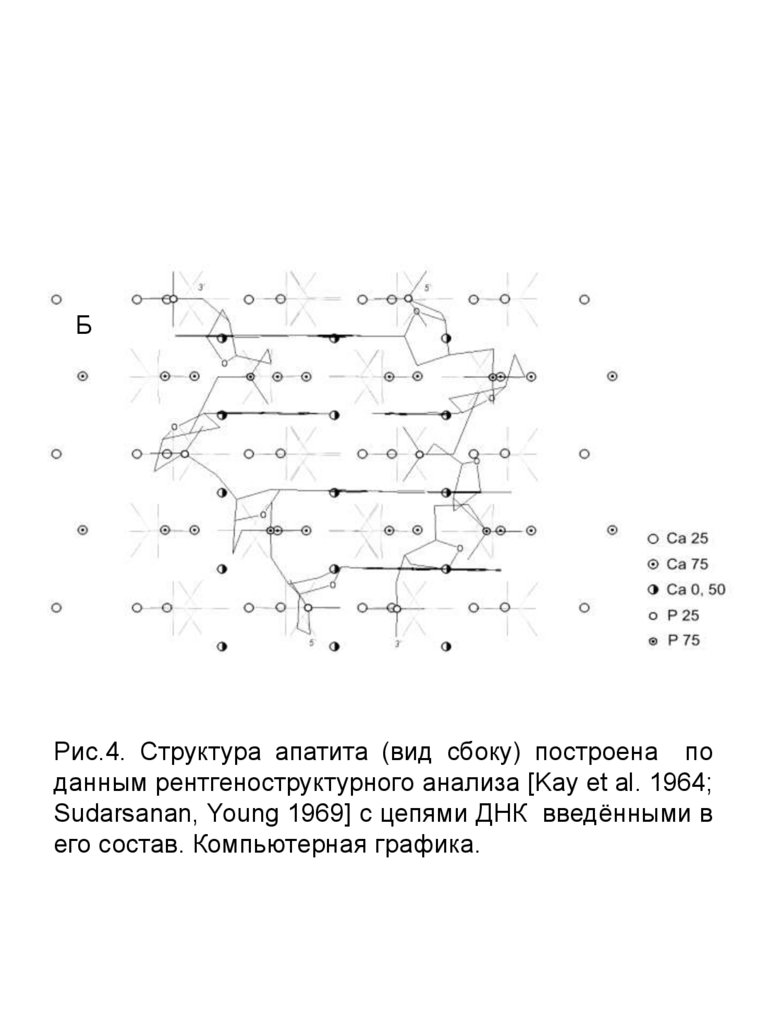
Рис.4. А – Структура апатита (вид сбоку) построена
по данным рентгеноструктурного анализа [Kay et al.
1964; Sudarsanan, Young 1969]; Б – Структура апатита
(вид сбоку) с цепями ДНК введёнными в его состав.
А, Б – компьютерная графика.
18.
Представление о структуре и связях в органоминеральном кристаллическом комплексе ДНК-РНКбелок дает кристаллическая структура взаимодействияРНКазы Н3 с РНК/ДНК субстратом вне минерального
комплекса (Рис. 6). Особо обращает на себя внимание
структура белка не имеющая жестких ограничений в
пространстве.
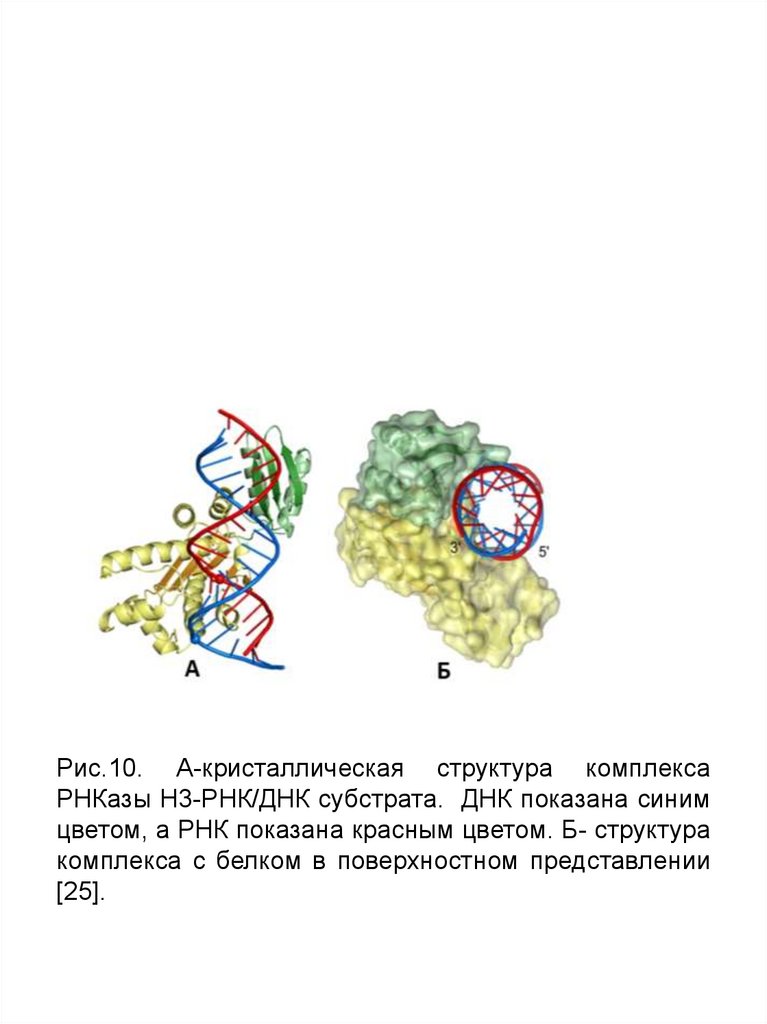
Рис.6. А-кристаллическая структура комплекса РНКазыH3-РНК/ДНК субстрата.
ДНК показана синим цветом, а РНК показана красным цветом. Б- структура
комплекса с белком в поверхностном представлении[26].
19.
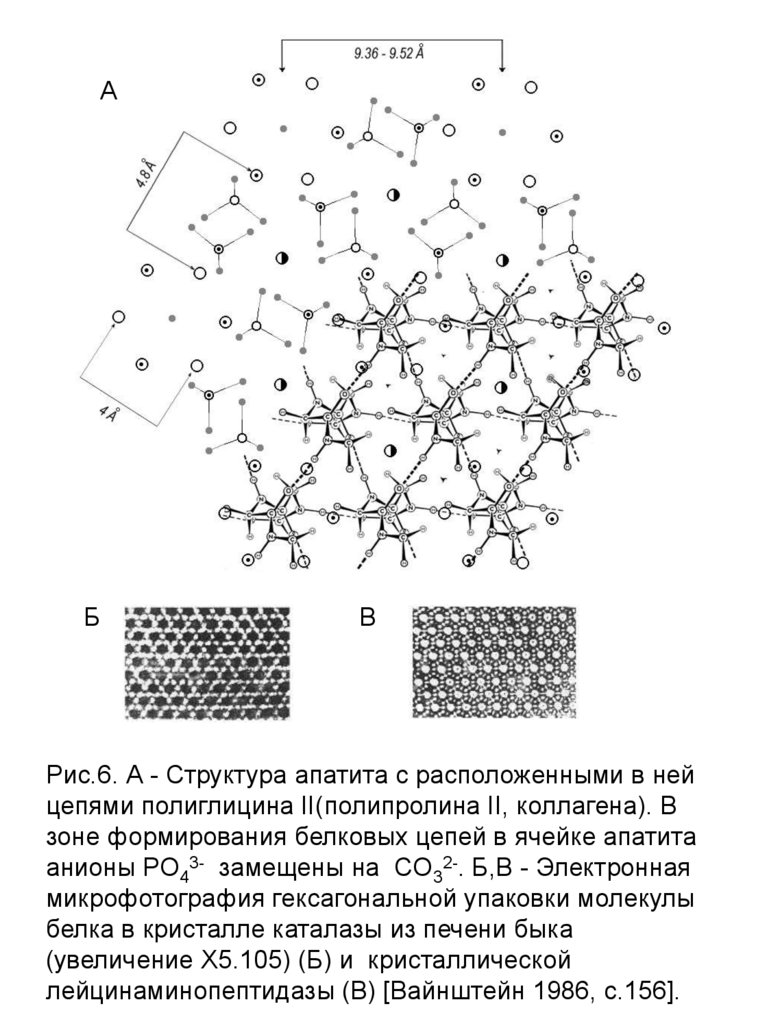
АБ
В
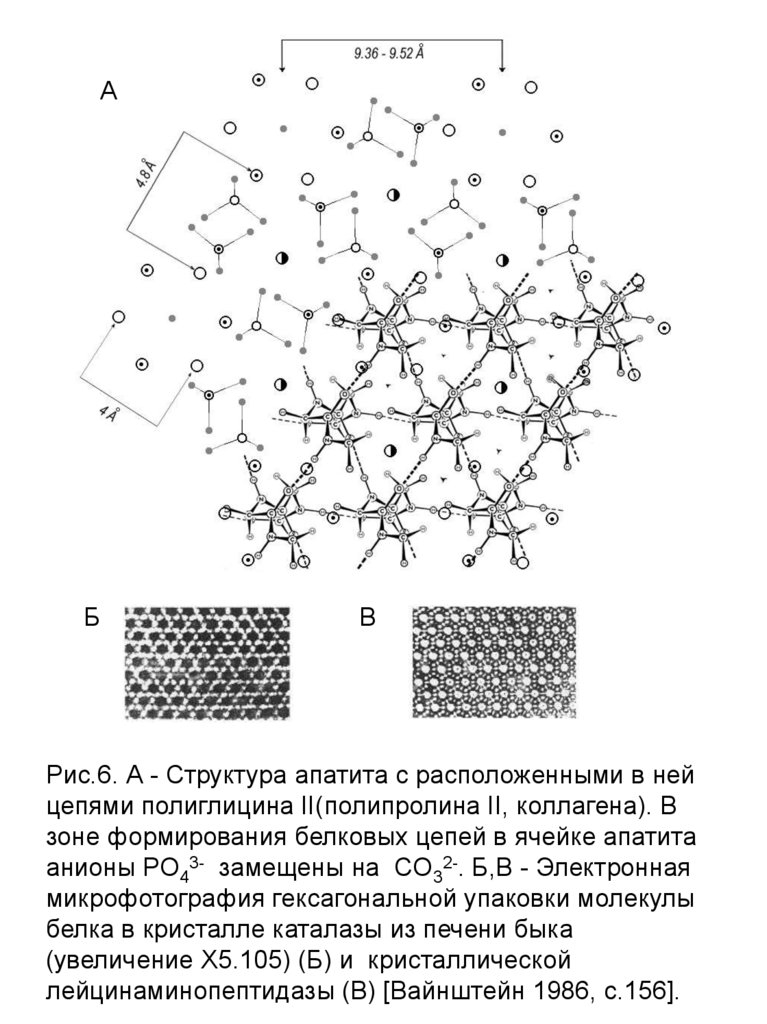
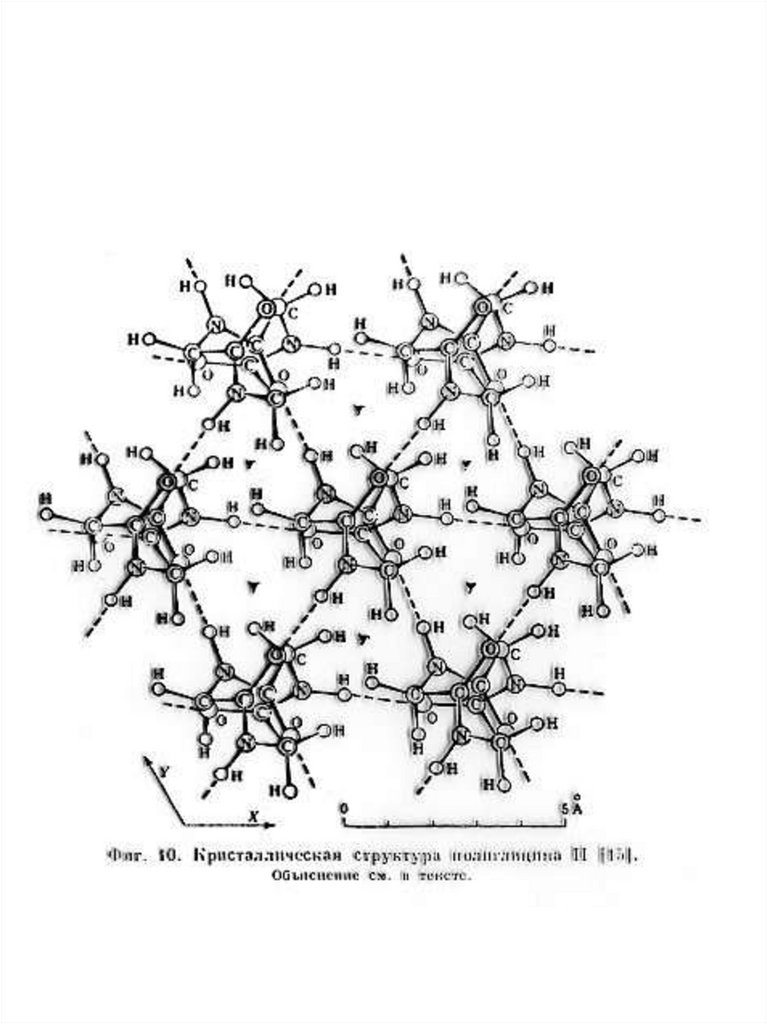
Рис.6. А - Структура апатита с расположенными в ней
цепями полиглицина II(полипролина II, коллагена). В
зоне формирования белковых цепей в ячейке апатита
анионы РО43- замещены на CО32-. Б,В - Электронная
микрофотография гексагональной упаковки молекулы
белка в кристалле каталазы из печени быка
(увеличение Х5.105) (Б) и кристаллической
лейцинаминопептидазы (В) [Вайнштейн 1986, с.156].
20.
21.
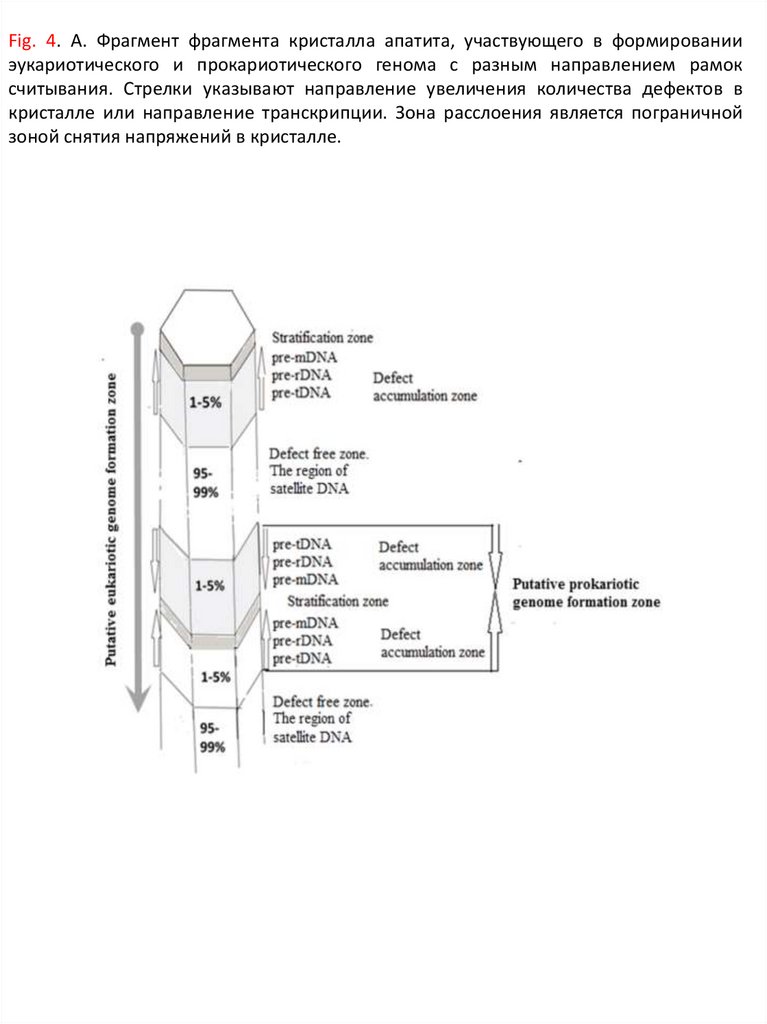
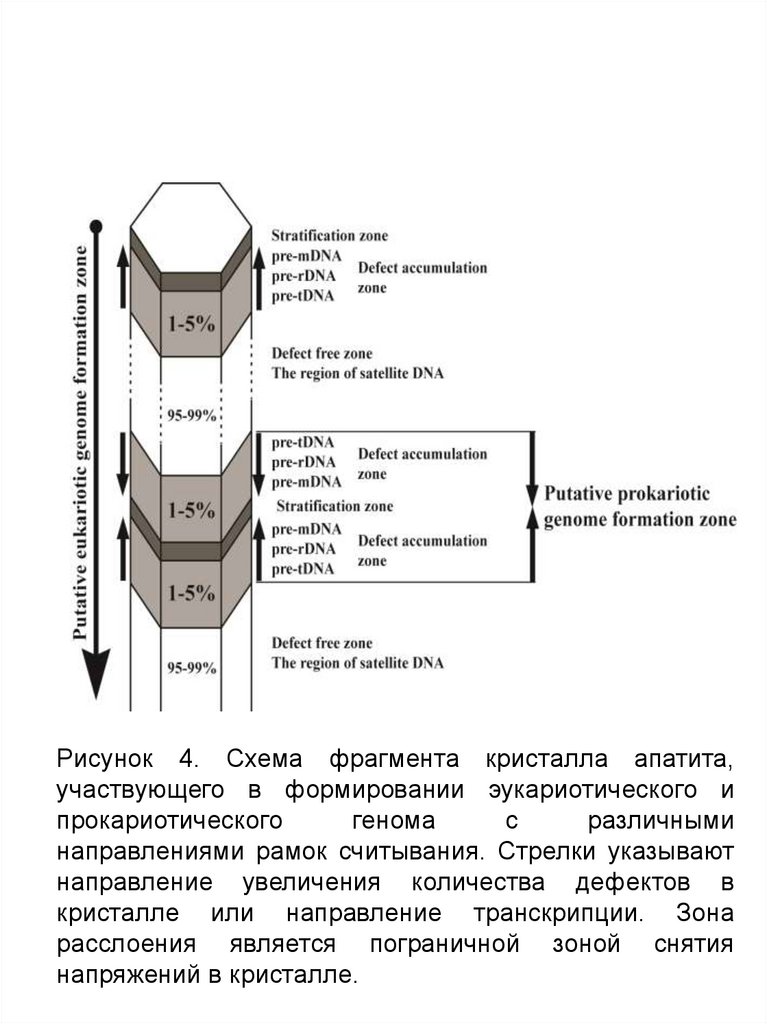
Fig. 4. А. Фрагмент фрагмента кристалла апатита, участвующего в формировании эукариотическогои прокариотического генома c разным направлением рамок считывания. Стрелки указывают
направление увеличения количества дефектов в кристалле или направление транскрипции. Зона
расслоения является пограничной зоной снятия напряжений в кристалле.
22.
Представление о структуре и связях в органоминеральном кристаллическом комплексе ДНК-РНКбелок дает кристаллическая структура взаимодействияРНКазы Н3 с РНК/ДНК субстратом вне минерального
комплекса (Рис. 6). Особо обращает на себя внимание
структура белка не имеющая жестких ограничений в
пространстве.
Рис.6. А-кристаллическая структура комплекса РНКазыH3-РНК/ДНК субстрата.
ДНК показана синим цветом, а РНК показана красным цветом. Б- структура
комплекса с белком в поверхностном представлении[26].
23.
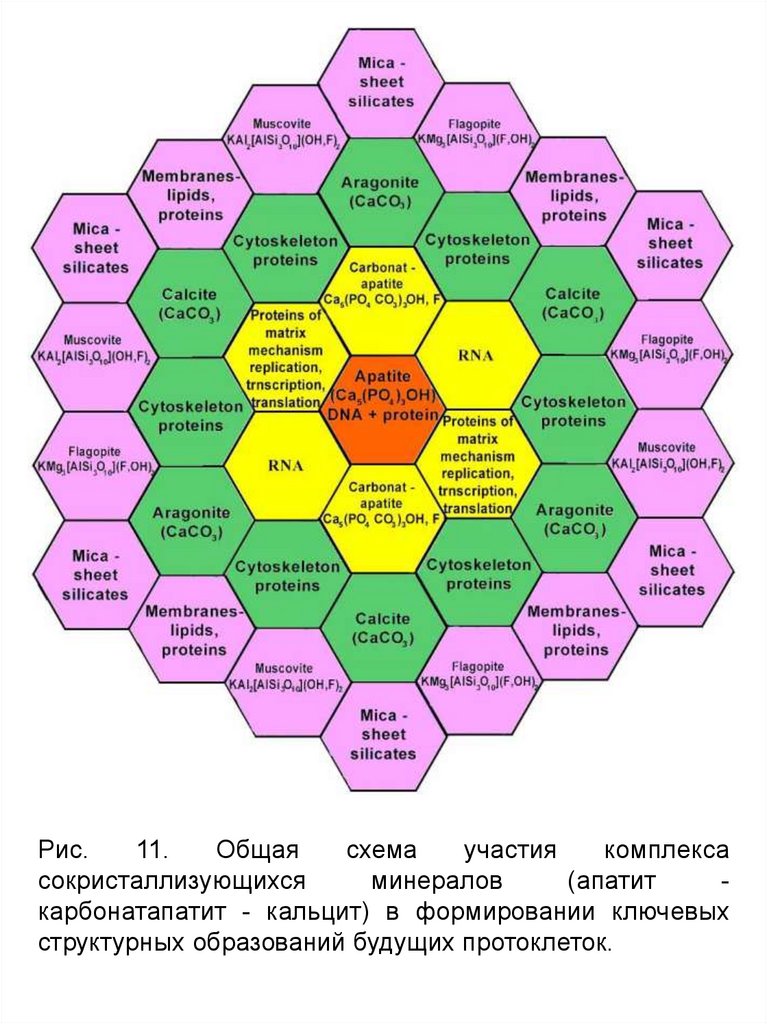
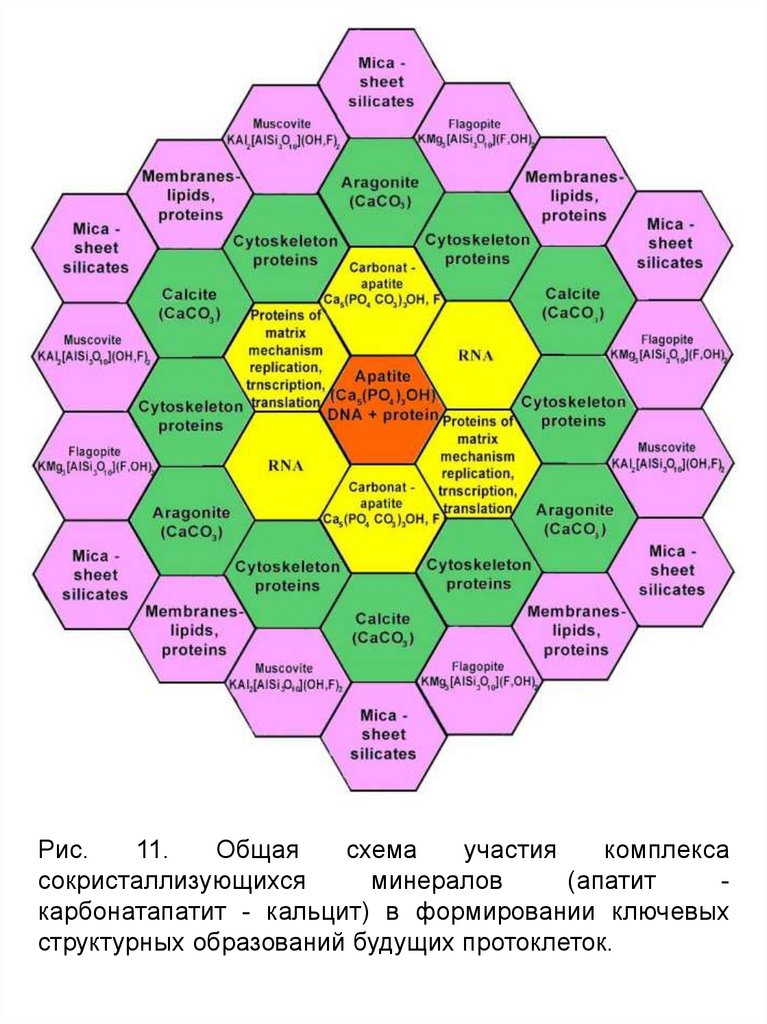
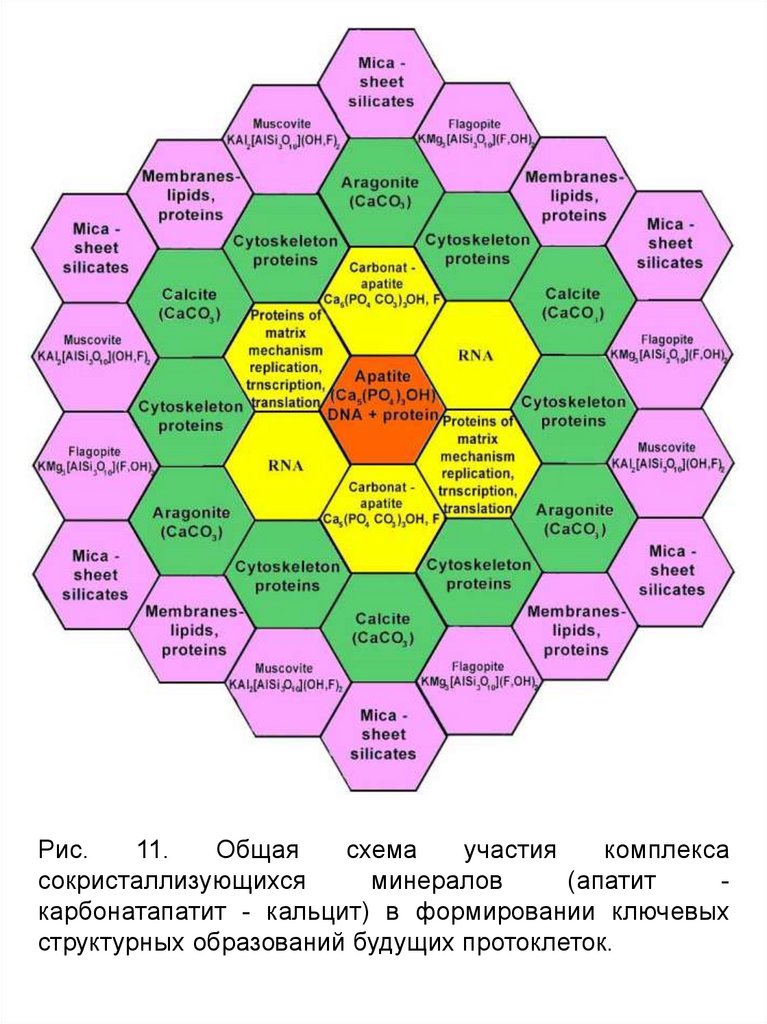
Рис.11.
Общая
схема
участия
комплекса
сокристаллизующихся
минералов
(апатит
карбонатапатит - кальцит) в формировании ключевых
структурных образований будущих протоклеток.
24.
25.
26.
3. Изоморфные замещения в апатите.Апатиты легко подвергаются различным изоморфным
замещениям. Более половины всех химических элементов
периодической
системы
Менделеева
обладают
способностью входить в структуру апатитов.
Изоморфизм анионов вдоль гексагональной оси. Атомы
фтора в апатите расположены вдоль гексагональной оси с
периодом 3,44Å. Вдоль этой оси имеются каналы, куда
могут входить многие ионы: гидроксильные группы, ионы
хлора, брома, кислород и сера в различных степенях
окисления, различные азотсодержащие ионы: CN22- , NCN-
, NCO и даже целая молекула аминокислоты глицина.
Всегда вдоль гексагональной оси присутствует более
чем один тип анионов. В минералах обычно много
фтора, некоторое количество гидроксильных групп, часто
кислород, иногда хлор.
27.
Изоморфныезамещения
фосфатных
групп.
Ортофосфатные группы (PO4)3- в апатите могут быть
замещены многими анионами: (SiO 4) 4- , (SO 4)2- ,
(AsO 4)3- , (VO 4)3- , (AlO 4)5- , (CrO 4)3- и др. Известны
парные замещения P на Si и S в фосфатных
минералах, (PO 4)3- на (SiO 4)4- и (CO 3)2- или (PO 4)3-
на (SiO 4)4- и CO 3OH 3- .
Изоморфные замещения катионов.
В апатитах Са
может быть замещѐн на многие катионы: Pb, Na, K, Sr,
Мn, Zn, Cd, Мg, Fe , Ba, редкоземельные элементы и
др. Согласно идеализированной структурной модели,
в апатите имеется два типа катионных позиций Са I и
Са II. Распределение примесных катионов между
этими позициями зависит от типа катиона, его
количества, а также от анионных замещений в
апатите.
28.
4. Особенности реальной структуры апатита.Водородные связи в апатите. В реальной структуре апатита
обнаружены Водородные связи в апатите OH…F , OH…Cl и
OH…O. Водородные связи присутствуют в апатитах
высокотемпературного происхождения, а в осадочных
образцах появляются после прокаливания.
Частичная
конденсация
пирофосфатных.
В
ортофосфорных
реальной
структуре
групп
до
природных
апатитов обнаружили наличие частичной конденсации
ортофосфатные группы (PO4) 3-- до пирофосфатных P-OP. Мы полагаем, что полимеризация фосфатных групп в
апатите происходит на дефектах структуры и процесс не
является ни регулярным, ни периодическим.
Карбонатионы в апатите. На сегодняшний день многими
авторами показано, что в апатите карбонатионы имеют две
неэквивалентные позиции: на гексагональной оси и вместо
(PO4) 3- .
29.
5.Динамика кристаллической решѐтки апатита.Мобильность анионов в каналах апатитовой
структуры была зафиксирована.
Динамика ортофосфатных групп.
Конденсация
ортофосфатных групп в соответствии с реакцией:
2HPO 4 → H 2O+P 2O 7 характерна для апатитов
высокотемпературного
происхождения,
в
осадочных образцах она происходит только после
нагревания. Динамика конденсации фосфатных
групп в апатите очень важна с биологической точки
зрения.
Физиологическая
низкомолекулярных
функция
полифосфатов,
особенно
пирофосфата, остаѐтся неясной.
Динамика карбонатных ионов.
Особенность
динамики карбонатионов в апатите – это их
способность к частичной перелокализации из
одной структурной позиции в другую.
перемещения-смерть
космонавтов.
при
Эффект
подготовке
30.
6.Зависимость свойств апатитов от реальной кристаллохимическойструктуры.
Известно, что гидроксилапатит поглощает фтор даже из
среды, бедной фтором. Вхождение фтора в структуру может
приводить к образованию водородных связей.
Водородные
связи
принадлежат
к
слабым
взаимодействиям (их энергия 5-10 ккал/моль), и поэтому они
очень важны для фазовых переходов, процессов адсорбции и
катализа.
Водородные связи влияют на растворимость апатитов.
Апатиты высокотемпературного происхождения всегда имеют
водородные связи и значительно меньшую растворимость,
чем минералы осадочного происхождения.
Водородные связи угнетают диффузию вдоль оси
структуры, активируют рост кристалла и его адсорбционную
способность.
.
31.
Пирофосфатныегруппы,
содержащие
лабильные связи Р-О-Р, активируют способность
апатита к адсорбции, изменяют патологические
процессы в зубной эмали.
Карбонатионы тоже оказывают сильное влияние
на растворимость, кристалличность апатита, но это
влияние
может
быть
противоположным
при
локализации карбонатионов в каждой из двух
возможных позиций в структуре.
Катионные замещения в апатите даже при
малых содержаниях заметно влияют на свойства, и
это влияние зависит не только от типа и количества
катионов, но и от их распределения по двум
структурным позициям в Са I и Са II
32.
7. О роли апатита при решении некоторыхмедицинских проблем
К
вопросу
о
невесомости.
Описаны
результаты
неоконченного исследования влияния невесомости на
состояние костной ткани человека.
В институте космических исследований было начато
изучение влияния невесомости на группе молодых
здоровых юношей. Мы изучали образцы костей этих
людей до и в течение эксперимента. Для всех образцов
костей
оказалась
частичная
перелокализация
карбонатионов с позиций ортофосфатных групп на
гексагональную
ось
структуры;
степень
этой
перелокализации увеличивалась по мере эксперимента
и была различной для разных людей. Через две недели
в образцах кости одного молодого человека произошла
полная перелокализация, и в структуре апатита не
осталось карбонатионов в позиции ортофосфатных
групп. Этот молодой человек умер, и его смерть
оказалась загадочной и совершенно неожиданной.
Никаких причин медикам найти не удалось
.
33.
К вопросу о мозговом песке. Долгое времясчиталось, что так называемый мозговой песок,
расположенный в центре мозга, в эпифизе,
является отбросом организма, а сам эпифиз –
единственная часть мозга, не имеющая левой и
правой части, – орган рудиментарный.
В настоящее время установлено, что эпифиз
вырабатывает
мелатонин,
серотонин,
норадреналин, гистамин.
Основная
суточных
функция
биоритмов,
метаболизма
и
эпифиза
–
регуляция
эндокринных
функций,
приспособление
организма
к
меняющимся условиям освещѐнности.
Ночной
пик
секреции
мелатонина
коррелирует
с
ночными
импульсами
магнитного поля Земли около 2 часов ночи.
Получается, что организм человека с помощью
эпифиза достаточно жестко сцеплѐн с гео- и
гелиокосмическими процессами.
34.
Эпифиз чувствителен не только к магнитному полю,но также к миллиметровому диапазону волн, к инфра- и
ультразвукам. Эпифиз моделирует активность гипофиза,
панкреатических островков, паращитовидных желѐз и
щитовидной железы. Влияние эпифиза на эндокринную
систему происходит через действие его гормонов на
систему гипоталамус-гипофиз-гонады. Эпифиз часто
называли третьим глазом. Его устройство напоминает
глаз.
В
эпифизе
стекловидное
тело,
удаѐтся
подобие
обнаружить
хрусталик,
сетчатки,
сосудистой
оболочки и глазной нерв. Символ всевидящего ока
всегда
сопровождает
мифологию,
телепатию
и
телекинез. Известна американская учительница Эми
Хансон,
которая
основатель
видит
затылком.
патанатомии,
Ещѐ
итальянский
в
18
веке
физиолог
Морганьи высказал мнение, что мозговой песок,
расположенный в эпифизе, связан с умом человека.
Песок отсутствует у детей до 7 лет, у престарелых и
у слабоумных. В последнее десятилетие установили, что
эти песчинки являются апатитом.
35.
Обнаружили,что
эпифиз
–
главное
место
аккумуляции фтора F в организме. Фтора в мягких
тканях в эпифизе оказалось 300 ppm, а в твѐрдых (в
гидрoксилапатите) 21000 ppm, что больше, чем в зубах и
костях . Установлено, что с возрастом изменения в
эпифизе
носят
более
функциональный,
чем
органический характер [14]. Функция мозгового песка
в эпифизе ещѐ ждѐт своего исследования. Детальное
кристаллохимическое исследование мозгового песка
сможет пролить свет на удивительную функцию этого
песка, хотя ещѐ в прошлом веке учѐные считали его
отбросом организма.
36.
Оквантовом
составной
компьютере.
частью
Эпифиз
квантового
является
компьютера
в
нашей голове, а мозговой песок в нѐм – это
физическая
основа
квантового
процессора.
Сознание человека способно под свой контроль
квантовый
режим
работы
своего
мозгового
компьютера и научиться им управлять. Профессор
МГУ, ученый - физикохимик Н.И. Кобозев (19031974) считал, что ”человек мыслит не по своей
воле, а потому что у него есть эпифиз с мозговым
песком, улавливающим космические излучения”.
Профессор, доктор физико-математических наук И.
Волович
считает,
индивидуальный
надеяться,
что
что
квантовый
“сознание
эффект”.
дальнейшие
–
это
Будем
физические
исследования помогут найти ответ на самую
интригующую тайну нашего бытия – что такое
сознание и как оно функционирует.
37.
Заключение Реальная структура апатита значительноотличается от идеализированной модели: энергетическим
разнообразием
гидроксильных
групп,
неэквивалентностью различных позиций атомов фтора,
частичной
конденсацией
ортофосфатных
групп
до
пирофосфатных, локализацией карбонатионов в двух
позициях
-
вместо
ортофосфатных
групп
и
на
гексагональной оси структуры. Распределение ионов
между различными позициями существенно влияет на
свойства минерала. Каждый анион в апатите может
переходить
из
одной
позиции
в
другую
при
энергетическом воздействии на минерал. Решающее
влияние
на
анионную
подвижность
в
кристалле
оказывает количество фтора на гексагональной оси. Для
предотвращения технологических сбоев в процессах
извлечения и переработки фосфатного сырья и даже при
поиске алмазов целесообразно кристаллохимическое
изучение апатита. Детальное исследование апатита
может
оказаться
полезным
при
решении
таких
медицинских проблем, как влияние невесомости на
костную ткань, роль мозгового песка.
38.
Словарь терминовМагматические
горные
породы
–
породы,
образовавшиеся
непосредственно
из
магмы
(расплавленной массы, образованной в глубинных
зонах Земли) в результате еѐ поступления в
верхние
горизонты
Земли,
охлаждения
и
застывания.
Метасоматические горные породы (от греческих
слов: metá- за, после и soma - родительный падеж
от somatos - тело). Это породы, образовавшиеся
при
замещении
одних
минералов
другими
с
существенным изменением химического состава
породы в процессе взаимодействия с жидкостями.
Осадочные горные породы – породы, возникшие в
результате осаждения различных веществ в водной
среде.
39.
Изоморфные замещения (от греческих слов iso одинаковый и morphosis - образовывать, приниматьформу). Это замещения элементов в минералах без
изменения формы кристалла. Водородная связь –
форма
ассоциации
между
электроотрицательным
атомом, например N, O, F, и атомом водорода H,
связанным
ковалентно
с
другим
электроотрицательным атомом.
Валентные колебания – колебания молекул, основной
вклад в которые вносят колебания ядер вдоль
направления валентных связей.
Либрационные колебания (от латинского слова libratio-
раскачивание)
–
колебания
одного
атома,
раскачивающегося относительно другого.
Типоморфизм
–
минерала
условий
от
явление
зависимости
образования.
свойств
Признаки
минерала, по которым можно судить о тех или иных
условиях его образования, называются типоморфными
признаками.
40.
Далее смотри слайды о третьемглазе эпифизе41.
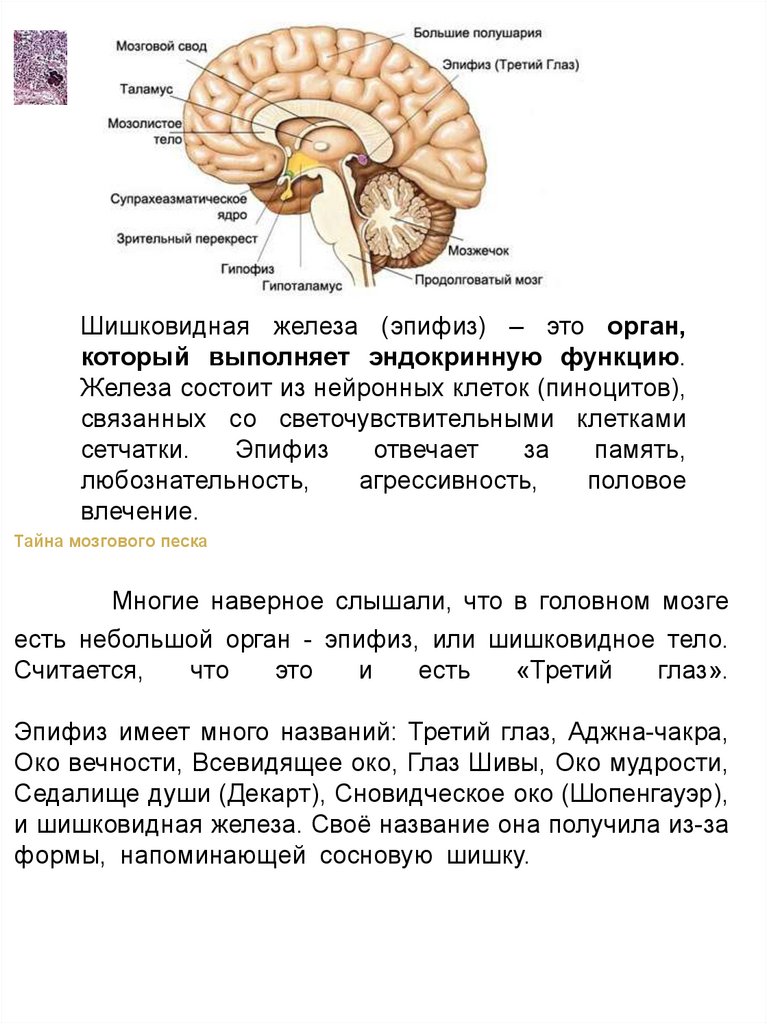
Шишковидная железа (эпифиз) – это орган,который выполняет эндокринную функцию.
Железа состоит из нейронных клеток (пиноцитов),
связанных со светочувствительными клетками
сетчатки.
Эпифиз
отвечает
за
память,
любознательность,
агрессивность,
половое
влечение.
Тайна мозгового песка
Многие наверное слышали, что в головном мозге
есть небольшой орган - эпифиз, или шишковидное тело.
Считается,
что
это
и
есть
«Третий
глаз».
Эпифиз имеет много названий: Третий глаз, Аджна-чакра,
Око вечности, Всевидящее око, Глаз Шивы, Око мудрости,
Седалище души (Декарт), Сновидческое око (Шопенгауэр),
и шишковидная железа. Своё название она получила из-за
формы, напоминающей сосновую шишку.Многие наверное
слышали, что в головном мозге есть небольшой орган - эпифиз, или шишковидное тело. Считается, что это
и
есть
Эпифиз имеет много названий: Третий глаз
«Третий
глаз».
42.
Восточныеоккультисты
утверждают,
что
шишковидная железа со своим особым устройством
нервных клеток и маленькими крупинками мозгового
песка тесно связана с волевой передачей и приёмом
ментальной
вибрации.
Шишковидная железа представляет собой массу
нервной ткани, находящуюся в мозге почти в центре
черепа и прямо над верхним концом позвоночника. Она
имеет форму небольшого конуса и красновато-серый
цвет. Находится перед мозжечком и прикреплена к
третьему желудочку мозга. В ней содержится большое
количество твёрдых частиц, похожих на песчинки,
известных
под
названием
мозговой
песок.
Исследования показали, что этого вещества нет у
детей примерно до 7 лет, у людей слабоумных и у тех
кто
страдает
теми
или
иными
расстройствами
умственной организации. Оккультисты знают, что этот
песок
является
человека.
Он
ключом
служит
сознанием и телом.
к
духовному
связующим
сознанию
звеном
между
43.
Учёныетоже
предположение,
что
неоднократно
кристаллы
высказывали
мозгового
песка
способны принимать излучения не электромагнитной
природы. Так, ещё в начале 70-х годов ХХ века
известный
советский
профессор
Московского
учёный-физикохимик,
университета
Николай
Иванович Кобозев, анализируя феномен сознания
пришел к выводу, что молекулярная материя мозга
сама по себе не способна обеспечить мышление, для
этого
необходим
внешний
источник
потоков
сверхлёгких частиц - психонов.
Согласно этой гипотезе, человек мыслит не по
своей воле, а потому, что у него есть эпифиз с
мозговым
песком
излучения.
А
носителями
и
эмоциональных
улавливающий
психоны
являются
переносчиками
космические
главными
мыслительных
и
импульсов.
44.
Выдвигается гипотеза, согласно которой мозговой песокв эпифизе является управляющим центром и носителем
информационной голограммы организме человека и
других высокоорганизованных животных. Это уже совсем
близко
к
понятию
квантового
компьютера.
В процессе жизнедеятельности живые кристаллы
постепенно обрастают органо-фосфорно-кальциевыми
оболочками,
то
есть
внутри
эпифиза,
в
среде
перенасыщенной солями кальция и фосфора, они
постепенно трансформируются в агрегаты мозгового
песка. Необычные информационные свойства мозгового
песка
замеченные
при
проведении
опытов,
свидетельствуют по мнению авторов о том, что в них
остаётся записанной вся информация о человеке.
В
настоящее
время
гистохимики выяснили
какова
структура мозгового песка. Песчинки по размеру бывают
от 5 мкм до 2 мм, по форме они часто напоминают
тутовую ягоду, то есть имеют фестончатые края.
45.
Песчинки Состоят из органической основы - коллоида,который считается секретом пинеалоцитов и пропитан
солями
кальция
и
магния,
преимущественно
фосфатами. Методом рентгенокристаллографического
анализа
было
показано,
что
соли
кальция
на
дифрактограммах эпифиза аналогичны кристаллам
гидроксиапатита.
Мозговые
поляризованном
свете
лучепреломление
с
песчинки
обнаруживают
образованием
в
двойное
мальтийского
креста.
Благодаря
песчинки
наличию
первично
фосфорнокислого
кальция,
флуоресцируют
в
ультрафиолетовых лучах как и капельки коллоида
голубовато-белым свечением. Подобную же голубую
флуоресценцию дают миелиновые оболочки нервных
стволов.
Самое интересное, что оказывается песок содержит
в своём составе гидроксиапатит кальция. Именно о
нём шла речь как об одном из самых подходящих
кандидатов на роль физической основы квантового
компьютера! Поразительное совпадение, и вероятно
не случайное
46.
пропуститьПосле
рождения
ребёнка,
его
квантовый
компьютер ещё чист, он не загружен никакими
программами позволяющими ориентироваться в
нашем плотном мире.
Да и сам компьютер, как устройство которым
можно пользоваться, ещё не готов к работе окончательная «сборка» ещё не завершена. Это
как в физике квантовой информации: какой толк в
том, что любые взаимодействующие системы
связаны нелокальными корреляциями - квантовыми
компьютерами
от
становятся.
Чтобы
получился
квантовый
компьютер,
нужно
организовать
кубиты,
которыми
можно
этого
было
они
бы
для
нас
не
избирательно
манипулировать, выполнять логические операции
и получать результат.
47.
Объединяя данные по элементной базе квантовогокомпьютера с данными биологии об эпифизе и структуре
мозгового песка, можно сделать очень интересное
предположение:
эпифиз
головного
мозга
является
составной частью квантового компьютера в нашей
голове,
а
мозговой
песок
-
физической
основой
квантового процессора.
Так же и ребёнок - поначалу он ближе к Тонкому
миру, в его эпифизе ещё нет кубитов, над которыми он
мог бы выполнять логические операции. Мозговой песок
и кристаллы гидроксиапатита как физические носители
кубитов формируются постепенно по мере взросления
ребёнка, когда он начинает овладевать ментальными
конструкциями и логическими операциями.
Человек
имеет
возможность
воспользоваться
«волшебными» нелокальными свойствами запутанных
состояний кубитов своего квантового компьютера.
Получается, что все эзотерические практики по своей
сути означают, что человек пытается переключить
головной
мозг
из
классического
функционирования в режим квантовый.
режима
48.
Занимаясьмистической
практикой,
он
пытается
задействовать нелокальный ресурс запутанных состояний и
управлять ими, делая практически то же самое, к чему
стремятся
сейчас
физики,
реализацией
работая
над
квантового
технической
компьютера!
Из теоретических основ квантовой механики следует, что для
появления дополнительных квантовых корреляций необходимо
наличие классических взаимодействий. То есть для того чтобы
наша душа имела возможность осознать себя и развиваться
далее, она должна иметь материальную основу, проводника в
предметном мире. Таким проводником вполне могут являться
кристаллы гидроскиапатита в мозговом песке, выступающие в
качестве физической основы квантового компьютера в нашем
головном мозге.
шишковидная
Кстати, при ультразвуковом исследовании
железа
становится
видна
в
человеческом
зародыше на 49-й день после его зачатия, примерно в то же
время когда становится различим пол ребёнка. Природа в
первую очередь начинает формировать процессор нашего
будущего квантового компьютера, на который «накручивается»
уже остальное «железо».
49.
Формирование начинается с тонких квантовыхуровней, и если имеет место реинкарнация, то есть
перевоплощение, то именно в это время
происходит улавливание квантового астросома
(Астральное тело или астросом служит в человеке
посредником между духовным принципом и
физическом телом) для очередного воплощения.
Согласно буддийским представлениям, жизненной
силе умершего необходимо как раз 49 дней для
вступления в следующее воплощение.
Расшифровка
Реинкарнация
Религиозная вера
Описание
Реинкарна́ция, то есть перевоплощение; также
переселе́ние душ, метемпсихо́з, — группа
религиозно-философских
представлений
и
верований,
согласно
которым
бессмертная
сущность живого существа перевоплощается снова
и снова из одного тела в другое.
Астральное тело или астросом служит в человеке
посредником между духовным принципом и
физическом телом, так как дух слишком отличен от
материи, ... Дух человека создаёт для себя
астральную оболочку, затем, с её помощью
облекается в более плотную материю. Астросом
управляет физической жизнью нашего тела.
50.
Онзаведует
всеми
дыханием,
жизненоважными
пищеварением,теплоотдачей
процессами.
Он
и
выделяет
отслужившие клетки и заменяет их новыми, восстанавливает
ткани при лёгких ранах и уничтожает микробов. Главный
орган
Астросома:
Большой
Симпатический
нерв.
Представитель Астросома есть инстинкт. Во время сна
Астросом бодрствует и продолжает наблюдать за функциями
физического
тела.
бодрствования,
Во
Астросом
сне,
а
иногда
выходит
из
и
во
время
тела
и
может
значительно удаляться от него, но в этом случае между ними
остаётся связь, которую может прервать только физическая
смерть. Так как Астросом состоит из эфирной материи и его
молекулы могут исходить из тела, мы можем видеть
эфирным зрением выход энергии из пальцев человека, ауру
предметов и т.д.Соотношение между Духом, Астросомом и
физической
материей,
определяется
их
вибрацией.
Вибрация Духа сходит в материю постепенно сгущаясь.
Вибрация Духа — есть мысль. Всякая мысль отпечатывается
в Астрале на ментальном плане, где сохраняется в
зависимости от ясности и силы.
51.
Получается, что в какой-то мере справедливомнение оккультистов, что мозговой песок является
отложением психической энергии, и точка зрения,
согласно которой эпифиз является связующим
звеном тела и сознания (седалищем души), тоже
кажется
вполне
разумной.
Этот вывод подтверждает цитата из книги М.П.
Холла:
«Маленький
преимущественно
в
ребёнок
незримых
живет
мирах.
Его
физический организм пока ещё управляется с
трудом, однако в тех мирах, с которыми он связан
посредством открытых врат шишковидной железы,
ребёнок
осознаёт
себя
и
активно
действует.
Постепенно определённые проявления его высшего
сознания поглощаются физическим организмом и
кристаллизуются
в
виде
мельчайшего
песка,
обнаруживаемого в этой железе. Но до тех пор, пока
сознание не войдёт в организм, никакого песка в
этой
железе
нет».
52.
А что происходит с человеком, у которого удаленэпифиз?
После удаления эпифиза люди испытывают так
называемое «би-размещение». Вот одно из таких
описаний:
наблюдал
...Я
пациентов,
у
вследствие
много
которых
нейрохирургических
был
опухоли.
удалён
Они
эпифиз
классически
демонстрируют виртуальное би-размещение, при
котором
они
существуют
одновременно
и
в
призрачной реальности и в настоящем. Они
существуют в ярком сновидческом состоянии, пока
находятся в сознании, и могут чередовать два эти
состояния своего сознания.
При тестировании этих пациентов выявляется,
что
их ориентация в
этой
действительности
отличается от нормы и может казаться странной
случайному наблюдателю.
Любопытно, что эти пациенты демонстрируют
полностью зафиксированный пристальный взгляд
со
слабо
различимым
движением
глаз.
53.
Иещё
более
любопытно
то,
что
когда
они
перемещаются в этой реальности, то и в другой
реальности перемещаются на такое же расстояние.
Один джентльмен, которому я помогал дойти до
ванной, остановился на полпути и некоторое время не
мог идти дальше вследствие того, что в его другой
реальности он был на скачках, и то место где мы
находились
в
воспринималось
коридоре
им
как
госпиталя,
граница
одновременно
трека.
Мы
не
двигались до тех пор, пока путь не стал свободен от
лошадей, которые могли его сбить...
Источник:
http://paranormal-
news.ru/news/tajna_mozgovogo_peska/2013-06-27-7175
54.
Основные моменты теории
Нобель Даниел Карлтон Гайдузек «Наиболее серьезные ученые ,
занимающиеся теорией происхождения жизни , уже давно
отказались от всей этой чепухи насчет того, что в океане мол, был
бульон из пуринов , пиримидинов и аминокислот и его
активировало ультрафиолетовое излучение.
2. Апатит является единственным существенным источником
фосфора на поверхности Земли.
3. Элементарная ячейка апатита и двойная спираль ДНК
имеют сходную периодичность в 3,4 Å и параметры РО43ˉ- групп.
4.Современные обитатели Земли, как уже упоминалось, наряду
с апатитом включают в состав минерализованных структур кальцит,
арагонит и кристобалит, которые имеют систему согласованных
пропорций
5. Для обоснования предложенной теории были сопоставлены
данные рентгеноструктурного анализа апатита, с одной стороны
[26, 43], цепей ДНК Z-формы [45, 46], цепей коллагена,
полиглицина II и полипролина II [37–39] – с другой; построена и
проанализирована модель апатита с фрагментом ДНК Z-формы, в
том числе компьютерная.
.
55.
6. В апатите можно отметить:гексагональную сингонию с пространственной группой С26n–
Р63/m,
определенный размер элементарной ячейки (Э. Я.): а = 9,36–
9,52 Å, с = 6,85–6,93 Å;
двойной набор формульных единиц в Э. Я. с периодом
трансляции вдоль оси С = 3,4 Å;
наличие фосфатных спиралей для РО43ˉ- групп
двух типов Са-полиэдров: девятивершинников – СаО9 (СаI) и
семивершинников – СаО6 (ОН, F, CI, CО3, О2-) (СаII),
высóты 0 и 50 – для СаI, 25 и 75 – для СаII,
по отношению к атомам фосфора – высóты 25 и 75;
изоморфизм,
т.
е.
способность
значительно
нарушать
стехиометрию состава (Са/Р = 1,67–1,50) при изменении рН
без изменения основных параметров структуры, и др
7. Кристаллохимические особенности апатита [6] допускают
в цепях «апатитовой» ДНК структуру, состоящую только из
комплементарных пуриновых и пиримидиновых оснований
типа Г Ц и А=Т с диаметром спирали 18 Å, а также
возможность выбора в апатите левой спирали из фосфатов
56.
8. Нами была построена пространственная модель кристаллаапатита с встроенным в него гипотетическим фрагментом Ап-ДНК
из 4 пар нуклеотидов в виде модели Дрейдинга и компьютерной
модели, построенной по данным рентгеноструктурного анализа.
9. В качестве оснований в модели использовали гуанин и
цитозин, как главные структурообразующие компоненты Z-формы
ДНК, а также в связи с тем, что они способны кодировать
основные аминокислоты коллагеновых белков (глицин – ГГГ, ГГЦ;
аланин – ГЦГ, ГЦЦ; пролин – ЦЦЦ, ЦЦГ), участвующих в синтезе
апатита в организме.
10.
Кристаллохимические особенности апатита наряду с
синтезом ДНК допускают одновременный синтез белковых цепей
гистонов, коллагена и других белков
Расположение и Параметры Ca II в элементарной ячейке
полностью совпадают с параметрами коллагеновых цепей
11. В случае гистоновых белков синтез, по-видимому, шел
одновременно с ДНК в тех же Э. Я. апатита (спираль в спираль)
по принципу стереохимической комплементарности.
12. Расположение и Параметры Ca II в элементарной ячейке
полностью совпадают с параметрами коллагеновых цепей.
13.
Подтверждение Формирования НП-комплекса на матрице апатита
аналогиями, при синтезе апатита в костях и зубах позвоночных животных
путем обращения процесса.
57.
14.
Механизм
сопряжения
решеток
основных
скелетных
минералов с органическим веществом базируется на физике
жидкокристаллических конфокальных текстур. Конфокальная микроскопия
— это оптический метод получения трёхмерных изображений текстур поверхностей и
объектов. С помощью метода отражённого света шероховатость поверхности измеряется
точно и бесконтактно.
15. Четыре основных скелетных минерала (апатит, кальцит,
арагонит
и
кристобалит),
взаимосогласованные
пропорции,
имеют
совпадающие
которые
сопрягаются
и
с
параметрами матричных конфокальных текстур жидких кристаллов.
16.При обращении модели матрица твердая, реактантами
являются радикалы и ионы (СН3˙, :СН2, СН3ˉ, СН22ˉ, СН3ˉ, С4ˉ(4+), Н+, N3ˉ,
NН2˙, NН2ˉ NН2ˉ, СО2ˉ и т. д.), меньшего радиуса, чем ионы в
минеральной матрице, а продуктами – молекулы с химическими
связями, существенно более короткими, чем атомные расстояния у
предшественников.
17. Участие минералов в процессе возникновения жизни делает
его закономерным, а время процесса весьма кратким.
58.
18.Время возникновения жизни совпадает с прогрессивным метаморфозом
базальтовой
коры,
формированием
гранитной
оболочки и уходом плотной первичной атмосферы. В этот момент
из остаточного магматического расплава происходило формирование пегматитов (апатита, кальцита, кристобалита, слюды,
кварца, полевого шпата и др.). Пегматитовые минералы обладают
крупнозернистой структурой. Размеры их могут сильно колебаться
– от субмикроскопических до метровых, вытянутых по оси С.
Апатит сокристаллизуется, как правило, именно с пегматитовыми
минералами. В тот период его решетка была достаточно подвижна
т.к. . поверхность Земли была нагрета до 600–700 °С, а из недр ее
шло активное выделение элементов восстановительной газовой
фазы в виде свободных радикалов.
19. Формирование органического кристалла в минералах идет без
воды под полным ориентационным контролем силовых полей
минеральной матрицы по правилу замещения вакансий «объем на
объем» (метасоматоз).
Особая роль в формировании органоминерального комплекса на
комплексе минералов отводится вакансиям.
59.
Вакансии в кристалле продукт флуктуации энергии вызванныйпрежде всего изоморфными замещениями, атомами в аномальном
валентном
состоянии,
атомами
смещенными
в
междоузлия,
дислокациями, внешнее энергетическое воздействие (космическое,
ядерное и УФ-излучение). Поглощенная энергия легко мигрирует на
большие расстояния в кристалле .
20. В кристалле происходит диффузное замещение элементов
кристаллической решетки свободными радикалами и ионами газовой
фазы (СН3˙, :СН2, СН3ˉ, СН22ˉ, СН3ˉ, С4ˉ(4+), Н+, N3ˉ, NН2˙, NН2ˉ NН2ˉ, СО2ˉ
и т. д.) по принципу «объем на объем».
В объеме вакансии от Ca2+ (1,05 Å) могли разместиться до 7 атомов
C4+ и N5+ (0,15 Å – радиус иона по Белову и Бокию); в объеме двух и
более
вакансий
от
Са2+
могли
разместиться
пуриновое
и
пиримидиновое основания, соединенные водородными связями.
Положение
таких
молекул
в
элементарной
ячейке
апатита
предопределено положением СаI на уровне 0, 50, 100 (см. ч. 1, рис. 1,
2). Расстояние между атомными плоскостями СаI в минеральных кри-
сталлах апатита и парами оснований в ДНК совпадает и равно 3,4 Å.
21. В результате ковалентного взаимодействия между атомами и
молекулами газовой фазы, ориентированными в силовом поле вакансий, синтезировались органические молекулы, которые способны образовывать друг с другом ковалентные, ионные или водородные
связи.
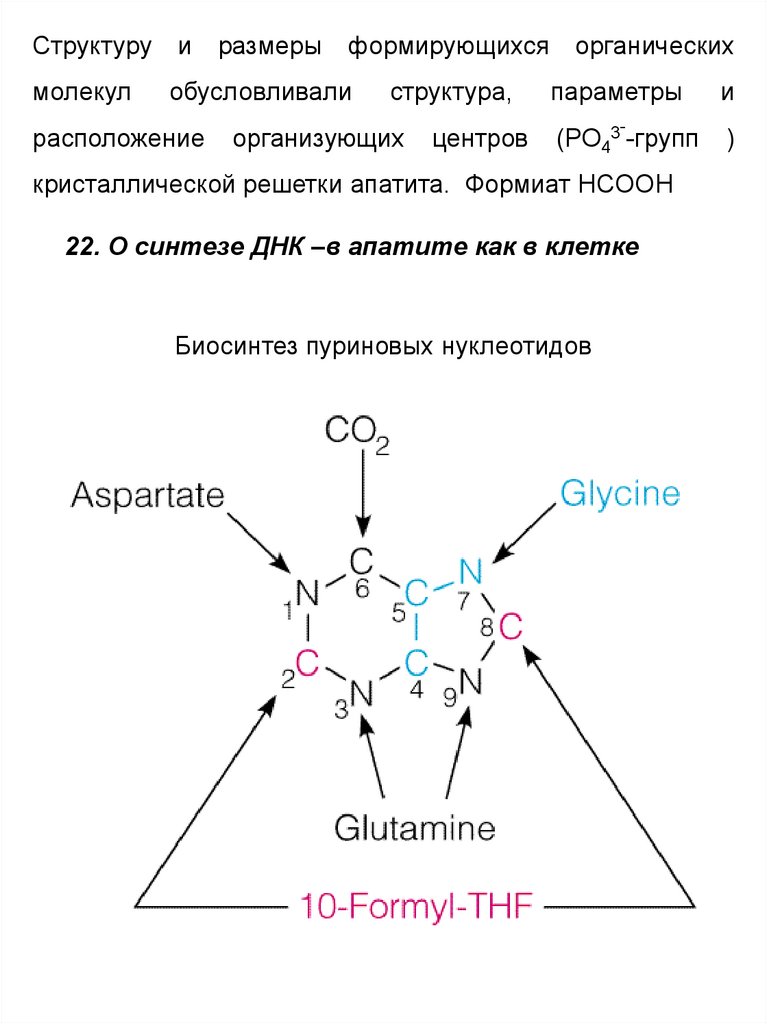
60.
Структуру и размеры формирующихся органическихмолекул
обусловливали
расположение
структура,
организующих
центров
параметры
и
(РО43ˉ-групп
)
кристаллической решетки апатита. Формиат HCOOH
22. О синтезе ДНК –в апатите как в клетке
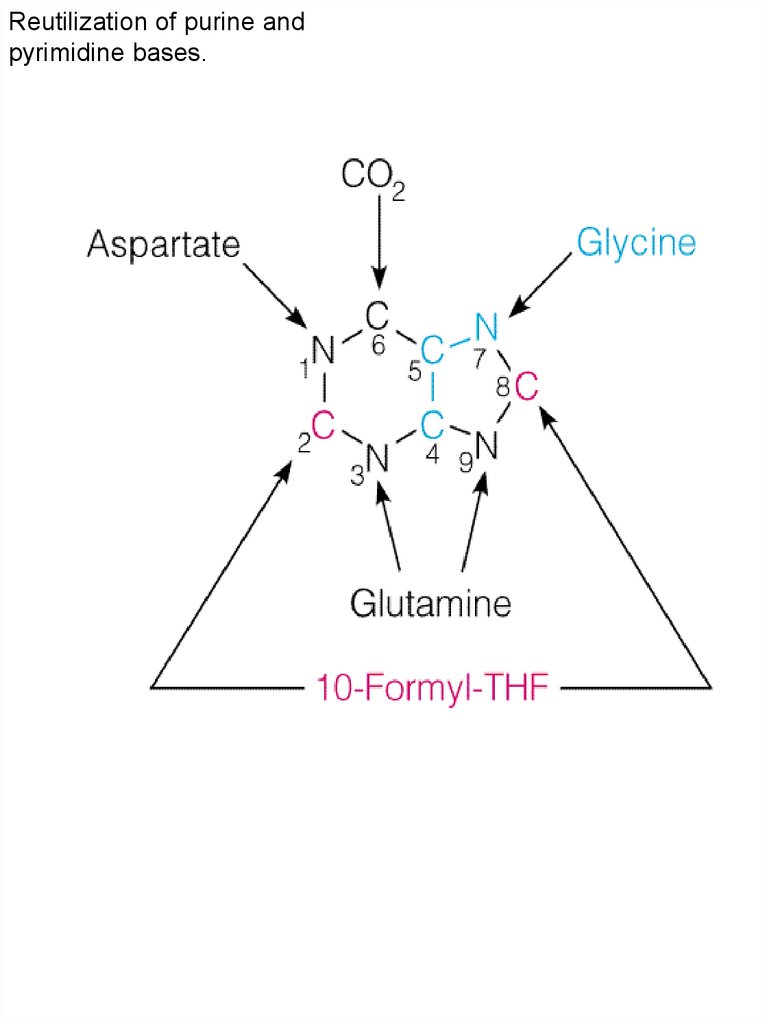
Биосинтез пуриновых нуклеотидов
61.
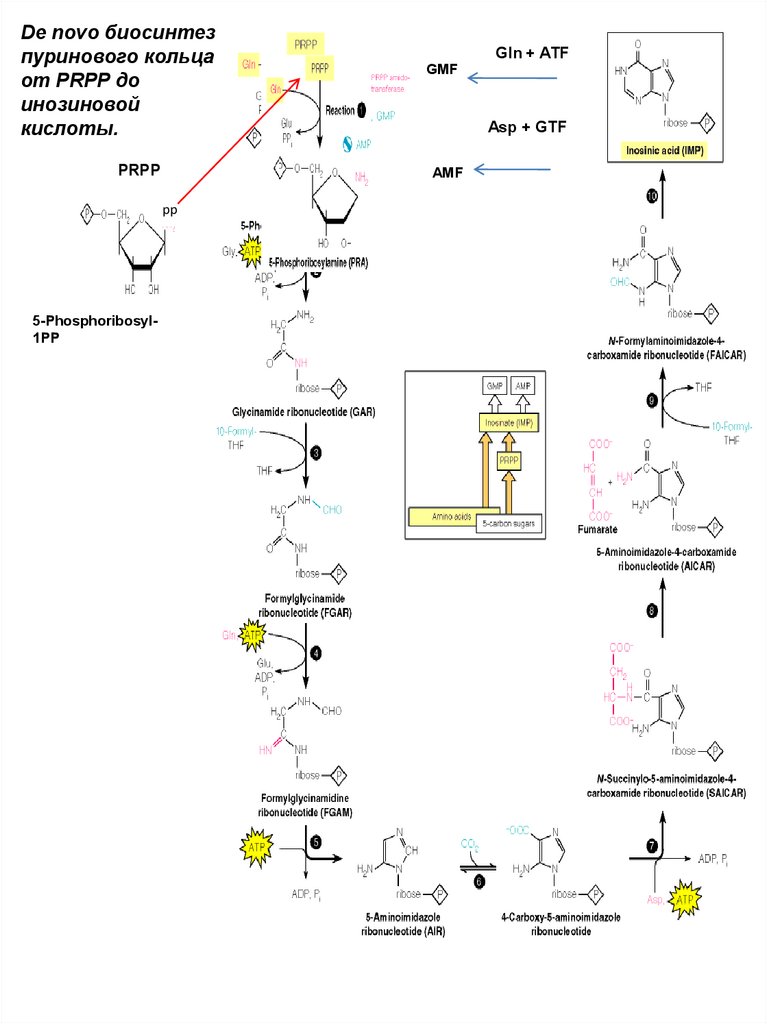
Формиат HCOOH62.
De novo биосинтезпуринового кольца
от PRPP до
инозиновой
кислоты.
PRPP
Asp + GTF
AMF
pp
5-Phosphoribosyl1PP
Gln + ATF
GMF
63.
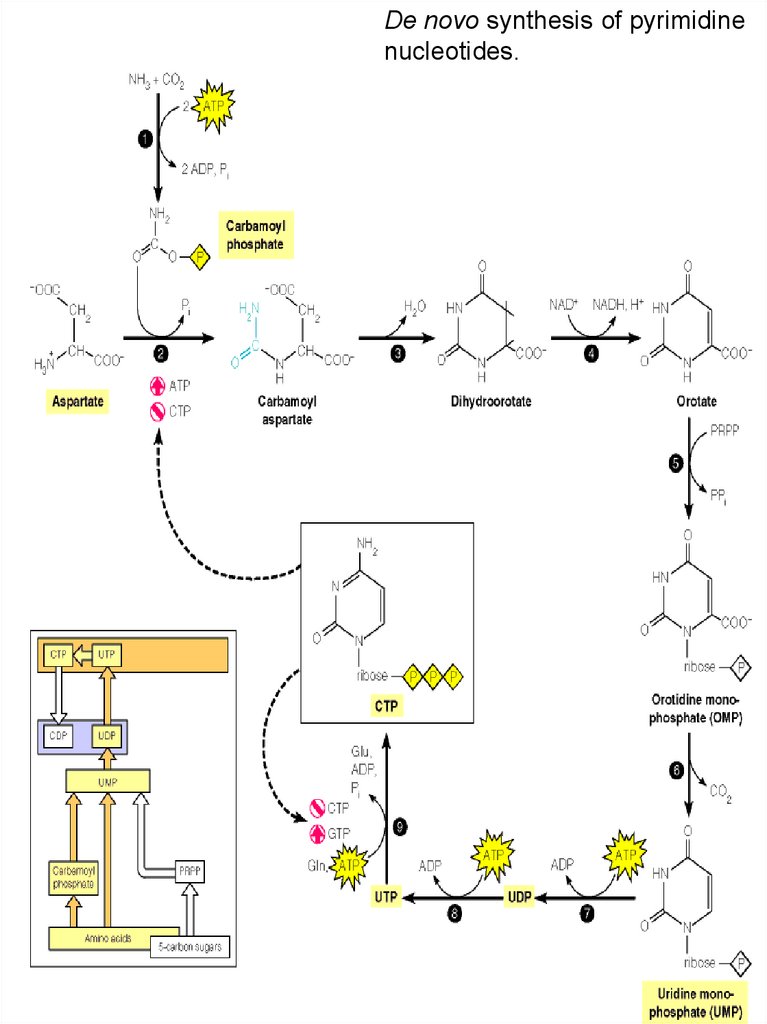
De novo synthesis of pyrimidinenucleotides.
64.
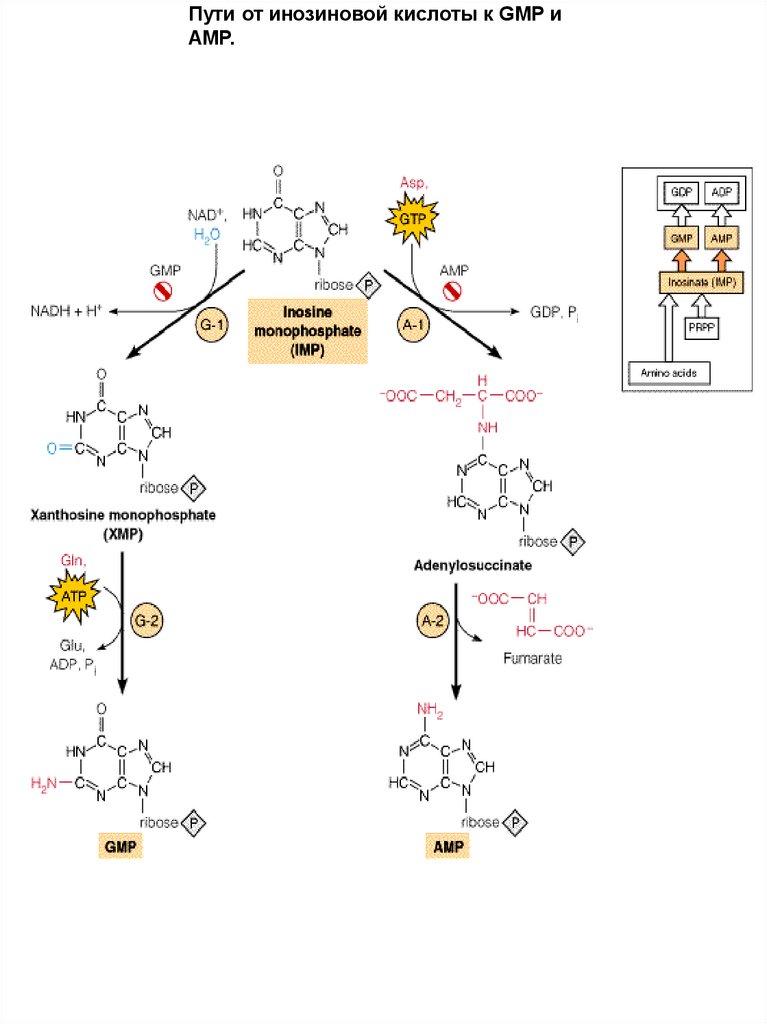
Пути от инозиновой кислоты к GMP иAMP.
65.
23. О синтезе РНК, белков матричного аппарата ицитоскелета.
Синтез происходил, видимо, по той же схеме, но на
смешанных
кристаллах,
имеющих
с
апатитом
совпадающие и взаимосогласованные пропорции. Это
карбонатапатит, кальцит, арагонит, кристобалит, кварц и
слюда.
Совпадающие
пропорции
что
разных
способствуют
тому,
структуры
органоминеральных
минералов
жидкокристаллические
комплексов,
возникающих на их основе, оказываются способными
соединиться в единую упорядоченную систему.
Отсутствие – РО43ˉ-групп полное или частичное,
делает невозможным формирование двойной спирали
ДНК, но не препятствует формированию цепей РНК,
нуклеотидных фрагментов типа АТФ, ГТФ, ЦТФ, УДФ,
НАД
или
ФАД,
белков
и
ферментов
аппарата
репликации, транскрипции, трансляции и процессинга,
связанных с РНК или ее фрагментами. Это будущие
ферменты, которые в качестве кофакторов могут иметь
НАД, ФАД, РО43ˉ, АТФ и т. д.
66.
Для
синтеза
белков
цитоскелета
совершенно
не
обязательно
регулярное расположение РО43ˉ- групп или даже их присутствие в
кристаллической решетке. Сокристаллизация апатита, карбонатапатита
и кальцита не оказывает влияния на появление белков с пропорциями
или параметрами, отличными от белков апатитовой природы, т. к.
решетки этих минералов имеют совпадающие и взаимосогласованные
пропорции.
Изоморфные и дефектные участки карбонатапатита и кальцита могли
стать
основой
для
формирования
белков,
обладающих
специфичностью в центрах связывания по отношению к ионам (Са, Mg,
Sr, Si и др.) и определяющих будущую структуру минеральной
составляющей живых систем (кальциевый, стронциевый, кремниевый,
апатитовый
скелет).
Пропорции
и
соответствия
таких
белков,
полученные на матрице кристалла, не могут быть изменены в силу
того, что подобные белки попадают в будущую протоклетку не из
окружающей среды, а находятся в едином органоминеральном
кристаллическом комплексе с остальными структурными элементами
будущих протоклеток. Весь набор структур, необходимый будущим
протоклеткам,
представлял
собой
существовало
комплементарное
единое
целое,
соответствие,
в
котором
обеспеченное
сокристаллизующимися минералами, имеющими совпадающие и
взаимосогласованные пропорции. В целом это была гомеостатическая
структура, стремящаяся поддерживать свою целостность.
67.
Органоминеральные
комплексы
будущих
протоклеток,
сформиованные при участии общих минеральных структур со
своим набором изоморфных замещений, дефектов и вакансий,
должны быть близки по принципу структурной организации.
24. Наследство протоклеток от минерального кристалла. В наследство
от
минерального
кристалла
органоминеральный
комплекс будущей пробиотической системы получил механизмы,
определяющие
и
составляющие
диффузионный
процесс
в
кристалле, а именно воздействие системы квантов света,
электронов, протонов, ядерных излучений, рН, тепла, способы
миграции по свободным полостям или каналам путем замещения
других атомов или вакансий либо по границам кристаллических
зерен, направление и последовательность процессов, т. е. основы
будущего метаболизма и гомеостаза в клетке.
Разнообразие
структурных
особенностей
минерала
обусловливает разнообразие будущих живых систем: изоморфизм
– специфичность белков к определенным ионам (Са, Fe, Ge, Mg, Si
и т. д.) и минеральную составляющую скелета (Sr, Si, Ca и т. д.);
степень дефектности – потенциальные возможности будущего
организма, поскольку, как уже отмечалось, чем выше степень
дефектности,
тем
труднее
состояния такой системы.
поддержание
гомеостатического
68.
Применительно к происхождению жизни к тем жекристаллам может восходить энергетика на фосфатах,
роль кальция в регуляции процессов в живых системах и
т. д.
Энергетическое перенесение электрона и
протона может использоваться митохондриями либо
для образования гидроксилапатита, либо для синтеза
АТФ (т. е. либо жизнь, либо возврат к минералу)
25.
От
органоминерального
комплекса
к
протоклетке. Переход произошел после появления
воды. Наиболее вероятно, что мембрана протоклеток
формировалась на месте, т. к. источник РО43ˉ- групп,
необходимых для фосфолипидов мембран, оставался
прежним
–
апатит.
Жидкокристаллический
органоминеральный комплекс содержал все компоненты,
необходимые
протоклеткам
для
обеспечения
промежуточного обмена. Органические вещества, не
попавшие в орбиту будущих протоклеток, фагов или
вирусов в силу своего родственного происхождения на
тех же минеральных матрицах, стали хорошей основой
для поддержания гомеостаза протоклеток. Движущей
силой дальнейшей эволюции протоклеток.
69.
О= писанный
выше
механизм
формирования
органоминерального комплекса будущей протоклетки
свидетельствует о тенденции природы к беспорядку, т. е.
возрастанию
энтропии
в
соответствии
со
вторым
законом термодинамики. В новом, органоминеральном,
комплексе
появляется
значительное
водородных
связей,
ковалентных,
изменяется
которые
количество
на
порядок
слабее
жесткая
система
связей
кристаллической решетки минералов. В целом энтропия
нового
комплекса
значительно
выше
энтропии
кристаллической решетки.
26. Проблемы биологической асимметрии. По нашему
мнению,
причина
несимметричным
киральности
связана
(однонаправленным
поверхности)
воздействием
формирование
комплекса
от
с
центра
температурного
киральных
внешним,
Земли
фактора
минералов
к
на
(апатит,
карбонатапатит, кальцит – оптически отрицательные). На
кристаллах
шел
однонаправленный
(снизу
вверх)
и
одновременный синтез нуклеиновых кислот и белков по
принципу «спираль в спираль», что практически исключало
возникновение рацемической смеси, но не препятствовало
возникновению правой или левой спирали.
70.
27. Предполагаемая схема возникновения матричногомеханизма в протоклетках.
Еще в 1936 г. Н.К. Кольцов [8] при формулировании принципа
матричного воспроизведения клеток использовал аналогию с
кристаллами.
Предлагаемый
нами
механизм
есть
нечто
большее, чем аналогия.
Как возник матричный механизм, как произошел его запуск,
откуда взялись первичные белки (рибосомальные) и белки-
ферменты (ААтРНК-синтетаза, ДНКзависимая РНК-полимераза и
др.) для его запуска?.
О дефектах в кристалле, как носителях информации
будущей протоклетки
Дефекты в кристалле – это прежде всего наличие изоморфных
замещений
и
вакансий
следствием
которых
является
возникновение флуктуации энергии и определенных структур в
зоне вакансий.
71.
Место возникновения матричного механизма – дефектная зона
апатита и зона сокристаллизации апатита, карбонатапатита,
кальцита. Система согласованных пропорций в решетках этих
минералов и наличие дефектных зон задают единый алгоритм
построения
будущих
протоклеток.
Благодаря
ритмичному
изменению химического состава смешанных кристаллов в них часто
проявляется
зональность.
Включения
в
кристалле
имеют
упорядоченное расположение. Реализация энергетических свойств
упорядоченно расположенных дефектов в структуре кристалла
приводит
в
конечном
итоге
к
регулярно
повторяющимся
стратификационным (слоистым) зонам.
Флуктуация энергии тепловых колебаний атомов
особенно
значительна в зоне дефектов кристаллической решетки, что ведет к
возникновению
очага
беспорядка
и
необходимости
сброса
энергетического потенциала. Для этих целей в любом кристалле,
имеющем дефекты (бездефектные кристаллы в природе не
встречаются), существуют страты.
72.
73.
.
Страты
нужны
кристаллу
для нивелирования
его
не-
совершенства. Переход от бездефектной зоны к дефектной идет от
нарастания беспорядка и далее к самоорганизации в виде
упорядоченно организованных в кристалле стратификационных
зон. Все это реализуется в структуре органоминерального
комплекса, в данном случае в структуре нуклеопротеидного
комплекса и транскрипционно-трансляционного аппарата клетки.
В зоне апатита и карбонатапатита образовывался тройной
комплекс – ДНК, РНК, белок. Самоорганизация такого комплекса
автоматически вытекает из того факта, что его синтез идет
одновременно на комплексе сокристаллизующихся минералов,
имеющих
взаимосогласованные
пропорции.
Взаимодействие
структур в тройном комплексе осуществлялось по принципу
стереоспецифической комплементарности, «спираль в спираль».
Его основа – двойная спираль ДНК.
74.
Рис.11.
Общая
схема
участия
комплекса
сокристаллизующихся
минералов
(апатит
карбонатапатит - кальцит) в формировании ключевых
структурных образований будущих протоклеток.
75.
76.
Fig. 4. А. Фрагмент кристалла апатита, участвующего в формировании эукариотического ипрокариотического генома c разным направлением рамок считывания. Стрелки указывают
направление увеличения количества дефектов в кристалле или направление транскрипции. Зона
расслоения является пограничной зоной снятия напряжений в кристалле.
77.
78.
79.
Рис.2 - Структура апатита (вид сверху) суказанием положения двух пар оснований ДНК
и фрагментом пептидной цепи (в центре)
[Kostetsky 1981; 1999].
80.
АБ
Рис.4. А – Структура апатита (вид сбоку) построена
по данным рентгеноструктурного анализа [Kay et al.
1964; Sudarsanan, Young 1969]; Б – Структура апатита
(вид сбоку) с цепями ДНК введёнными в его состав.
А, Б – компьютерная графика.
81.
АБ
В
Рис.6. А - Структура апатита с расположенными в ней
цепями полиглицина II(полипролина II, коллагена). В
зоне формирования белковых цепей в ячейке апатита
анионы РО43- замещены на CО32-. Б,В - Электронная
микрофотография гексагональной упаковки молекулы
белка в кристалле каталазы из печени быка
(увеличение Х5.105) (Б) и кристаллической
лейцинаминопептидазы (В) [Вайнштейн 1986, с.156].
82.
Переход в кристалле апатита от бездефектной области кпостепенному накоплению дефектов (усилению беспорядка)
реализовывался в особенностях чередующихся нуклеотидов в
формировавшихся
цепях
ДНК.
Наблюдался
постепенный
переход от сателлитной зоны (неинформативной части ДНК у
эукариот) к многократно повторенной (зоне тДНК), умеренно
повторенной (зоне рДНК) и уникальной (зоне генов мДНК) по
принципу стереоспецифической комплементарности, «спираль
в спираль».
Каждая
пре-tРНК
в
матрице
кристалла
взаимодействовала со своим белком (будущей ААтРНКсинтетазой), который уже имел в своем составе АТФ и
аминокислоту. Донором АТФ выступал карбонатапатит, а
аминокислот – карбонатапатит и кальцит. Возникает очень
интересное предположение, что структуру антикодона в
тРНК,
определяет
аминокислоту.
Она
ААтРНК-синтетаза,
расположена
в
узнающая
свою
органоминеральном
комплексе на пре-тРНК и для нее не очень важна исходная,
нуклеотидная последовательность пре-тРНК. Последняя может
выравниваться с помощью компенсаторной петли в тРНК.
83.
84.
Каждый пре-рРНК и пре-мРНК тоже был связан со
своими белками, которых было много в силу большей
протяженности этих фрагментов по сравнению с претРНК.
За
первыми
белками,
непосредственно
взаимодействующими с ДНК или РНК, следовало
большое
количество
других
комплементарно
связанных с ними белков. Не здесь ли лежит ответ на
вопрос,
откуда
брались
первые
белки
для
формирования рибосом и ацилирования tРНК и почему
их так много в клетке, притом что система синтеза
белков еще не готова?
85.
О возникновении матричного механизма в протоклеткахи ключевые проблемы молекулярной биологии.
Eduard Y.Kostetsky
Far Eastern Federal University, Russian Federation
kostetskiy.yeya@dvfu.ru
Аннотация. Рассмотрена теория возникновения матричного механизма в
составе протоклеток на комплексе минералов (апатит, карбонат-апатит, кальцит,
кварц), имеющих взаимосогласованные пропорции при участии радикалов газовой
фазы (NH3, CH4 и СO). Структура апатита и карбонат-апатита предопределила
формирование двойной спирали ДНК, имеющей комплементарные пары пуринпиримидиновых оснований; цепи РНК комплементарные ДНК; спиральные цепи
белков объединенные в супрамолекулярные структуры с РНК. Предполагается, что
в период архейской эры шло постепенное замещение минеральной матрицы
органическим веществом. Место возникновения матричного механизма бездефектная и нарастающая дефектная зона апатита и карбонат-апатита. Размер
и специфика генов в ДНК и комплементарно связанной РНК нарастали по мере
накопления дефектов в структуре минералов. Размер транскриптов РНК
сопровождался увеличением количества супрамолекулярно связанных молекул
белка. На первом, безводном, этапе на матрице апатита и карбонатапатита шло
формирование кристаллического органо-минерального, супрамолекулярного
комплекса - ДНК, РНК, белок по принципу «спираль в спираль» из элементов
газовой фазы. Фактически это был исходный транскрипционно-трансляционный
аппарат имеющий в готовом виде все типы РНК и белки-ферменты способные
обеспечить первый и последующий синтез белков. Появление воды определило
запуск транскрипционного аппарата и превращение органо-минерального
кристаллического комплекса в жидко-кристаллический или мезоморфное
состояние, которое сопровождалось запуском апарата трансляции. Дано
подробное описание подготовкии запуска матричного механизма. Обсуждаются
проблемы: происхождение рибосомальных белков и роли сверхспецифичной
аминоацил-тРНК-синтетазы, как истинного носителя генетической информации;
свойства генетического кода и синтез протоклеток без нарушения второго закона
термодинамики; возникновения биологической ассиметрии; нанобактерий и
темной материи.
86.
Развернутая аннотация. Рассмотрена теория возникновенияпротоклеток и матричного механизма в их составе на комплексе
минералов (апатит, карбонат-апатит, кальцит, кварц), имеющих
взаимосогласованные пропорции при участии радикалов газовой фазы
(NH3, CH4 и СO). позволяющая по новому осветить ключевые вопросы
молекулярной биологии. Структура апатита и карбонат-апатита
предопределила формирование двойной спирали ДНК, имеющей
комплементарные пары пурин-пиримидиновых оснований; цепи РНК
комплементарные ДНК; спиральные цепи белков объединенные в
супрамолекулярные структуры с РНК. Предполагается, что в период
архейской эры шло постепенное замещение минеральной матрицы
органическим веществом.
Место возникновения матричного механизма - бездефектная и
нарастающая дефектная зона апатита и карбонат-апатита. Первая зона
— это зона будущей сателлитной части ДНК у эукариот, вторая зона —
это будущая область транскрипционно-трансляционного аппарата.
Размер и специфика генов в ДНК и комплементарно связанной РНК
нарастали по мере накопления дефектов в структуре минералов.
Увеличение размера транскриптов РНК сопровождалось увеличением
количества белковых молекул в надмолекулярных белково-РНК
комплексах. От неинформативной или сателлитной части ДНК
(уэукариот) шло нарастание размеров геномов от пре-нкДНК к пре-тДНК,
пре-рДНК и пре-мДНК. Комплементарно и специфично с этими
областями, в зоне карбонатапатита, сокристаллизующейся с апатитом,
последовательно формировались транскрипты в виде белокнекодирующих пре-нкРНК, пре-тРНК, пре-рРНК и пре-мРНК.
Транскрипты РНК, помимо комплементарной связи с ДНК,
посредством межмолекулярных взаимодействий, формировали в зоне
карбонат-апатита супрамолекулярные структуры с белками, количество
которых росло в зависимости от размера транскрипта РНК. Так
формировался спиральный ДНК-РНК-белковый органо-минеральный
кристаллический
комплекс.
Синтез
шел
по
принципу
стереоспецифической комплементарности (спираль в спираль) из
элементов газовой фазы. Фактически это был готовый
транскрипционный аппарат, сформированный на матрице апатитом
и карбонат-апатита.
87.
Появление воды определило запуск транскрипционного итрансляционного аппарата и превращение органо-минерального
кристаллического комплекса в жидко-кристаллический. Вода нарушила
ионный баланс в сформированной системе и привела в движение ДНКРНК-белковый комплекс.
Произошло ослабление ДНК-РНК
взаимодействия и усиление РНК- белкового взаимодействия из-за
большого числа обращенных повторов в тРНК и рРНК. Изменение
ионного баланса системы позволило белкам превратить тРНК, рРНК и
мРНК в структурно-функциональные единицы аппарата и осуществить
его запуск.
Была сформулирована новая идея об истинном носителе
генетической информации - суперспецифической АА-тРНК-синтетазе,
которая стереоспецифически и комплементарно связывается со своей
кодонной тРНК, АТФ и аминокислотой. Сверхспецифичность будущей
АА-тРНК-синтетазы предопределила судьбу тРНК, обеспечив ее не
только собственной аминокислотой, но и превратив ее кодон в
антикодон, а саму тРНК в инструмент доставки аминокислот для
синтеза белка на кодонах мРНК рибосом, под контролем синтетазы.
Проблема с первыми рибосомальными белками была решена
еще на стадии формирования органо-минеральногоДНК-РНКбелкового комплекса.
Рибосомальная РНК, под влиянием обращенных повторов и
большого
количества
стереоспецифично
связанных
белков,
превратилась в рибосомальные единицы, которые начали
продвижение в сторону мРНК для подготовки встречи АА-тРНКсинтетазного комплекса и начала этапа трансляции. Этап трансляции
ознаменовался появлением новых рибосомальных и других белков,
произведенных прокариотами. Фактически это были первые стволовые
клетки, рожденные в недрах карбонат-апатитовой матрицы.
Процесс формирования матричного механизма у разных
таксономических групп мог продолжаться в течении всей архейской
эры формирования земной коры и гидросферы от 4 млрд.л.н. до 2.5
млрд.л.н.
88.
Рассмотрение процесса возникновения жизни на ранних этапахформирования земной коры позволило предположить, что механизм
возникновения структурной ассиметрии биомолекул обусловлен
однонаправленностью (от горячего к холодному) и одновременностью
синтеза НК и белков в объеме минералов по принципу «спираль в
спираль»; предположить ответ на вопрос о синтезе протоклеток без
нарушения второго закона термодинамики, путем перехода от
кристаллического
органоминерального
матрикса
к
жидкокристаллическому органоминеральному с потерей части энергии
кристалла и увеличения энтропии за счет смены связей кристаллической
решетки на супрамолекулярное взаимодействие, с помощью
стереоспецифической
комплиментарности.
Универсальный
генетический код рассматривается как следствие единой матрицы
апатита для всех живых и не живых систем (вирусы, фаги).
Предполагается, что вырожденность генетического кода по третьему
нуклеотиду, есть следствие расположения в элементарной ячейке
апатита только трех пар пурин-пиримидиновых оснований, позиция
третьей пары всегда на границе ячеек апатита, в зоне повышенной
электронно-плотностной напряженности. Следствием предложенной
теории является новый взгляд на понятие нанобактерий и темной
материи РНК.
Все выше рассмотренные, в виде предположений, единые
особенности происхождения и организации всех живых систем,
являются
следствием
единых
физико-химических
законов
структурной
организации
кристаллической
решетки
рассматриваемых нами минералов, ставших матрицей для
происхождения протоклеток.
89.
Мы предполагаем, что предложенный механизмвозникновения живых и неживых систем (вирусов, фагов),
гуминового скелета почв, углеводородов нефти и угля – все
это результат взаимодействия компонентов газовой
фазы(H2, NH3, CH4 и СO) с решетками разных минералов. Для
биомолекул это апатит, карбонатапатит, кальцит,
кристобалит, арагонит, слюда; для гумуса почв –
алюмосиликаты; для углеводородов нефти –минералы с
различной структурой кристаллической решетки; углей –
минерализованные органические скелеты.
90.
Предполагаемая схема возникновения матричного механизма впротоклетках.
Принято считать, что тот кто ответит на вопрос: «Как возник матричный
механизм», тот ответит на вопрос как возникла жизнь. При решении данной
проблемы Н. К. Кольцов [22]еще в 1936 г. использовал аналогию с кристаллами. В
данной работе эта аналогия приобрела реальные очертания. Перед нами стояли
следующие вопросы: когда, где, как и почему возник матричный механизм. Откуда
взялись первые белки для рибосом и будущие белки-ферменты транскрипционнотрансляционного аппарата (ААтРНК-синтетаза, ДНКзависимая РНК-полимераза и
др.) для его запуска и как произошел его запуск. По ходу обсуждения были
затронуты такие важные вопросы молекулярной биологии, как: почему истинным
носителем генетической информации являются ААтРНК-синтетазы, а не ДНК;
почему протоклетка формировалась без нарушения 2 закона термодинамики; как
возникла структурная ассиметрия, не кодирующая РНК(нкРНК) и нанобактерии.
Попытаемся ответить на поставленные выше вопросы, используя информацию,
полученую нами ране, о возможности происхождения первичных протоклеток на
комплексе минералов [1-3]. Для интерпретации наших результатов в качестве
базовой модели была взята прокариотическая система генома Escherichia coli. По
ходу обсуждения, иногда, будут делаться отступления для эукариот. Гены у E. Сoli
распологаются в следующем порядке.Это 76 объединенных в кластеры одинаковых
или разных генов тРНК; 7 оперонов с 3 разными рРНК, включающих 16S-, 23S- и
5S-рРНК фрагменты, отдельные тРНК и спейсеры. Далее идут мРНК. В
формировании рибосом участвуют 55 разных белков, гены которых объединены в 7
разных оперонах с индивидуальной регуляцией[23]. Исходные формы всех РНК в
виде пре-РНК подвергаются процессингу. От тРНК (73–93 н) к рРНК (1500–2900 н) и
мРНК (тысячи нуклеотидов) нарастает специфика и количество нуклеотидов в генах.
У эукариот вся эта система многократно повторена, и к ней добавляются тандемно
организованные повторы нуклеотидных последовательностей нетранслируемой
области сателлитной ДНК.
91.
Где и почему в кристаллах может появиться область снарастанием беспорядка?
У любого кристалла периодичность достигает 94-95% при
дефектной зоне в 5-6%. Бездефектные кристаллы в природе не
существуют. Дефектная зона – это потенциальная информация.
Бездефектная зона не несет никакой информации. Вопрос состоит в
причине и последовательности накопления дефектов. Дефекты в
кристалле – это, прежде всего, наличие изоморфных замещений,
наличие атомов в аномальном валентном состоянии, атомов,
смещенных в междоузлия,
наличие дислокаций. Подобные
дефекты приводят к появлению флуктуации энергии и
возникновению вакансий с делокализованными электронами.
Подобные электроны принадлежат кристаллу в целом, а не
отдельному атому. Дискретные энергетические уровни этих атомов
оказывают влияние на формирование вполне определенных
структур
в
зоне
вакансий.
Матричный
механизм
транскрипции и трансляции возникает в дефектной зоне,
где происходит сокристаллизация апатита, карбонатапатита
и кальцита, имеющих взаимосогласованные пропорции. В
этой зоне ритмично изменяется химический состав
смешанных кристаллов, образуются вакансии, происходят
изоморфные замещения, появляются атомы в аномально
валентном состоянии, возникают значительные изменения
энергии тепловых колебаний атомов в структуре минералов
и происходитнарастание очага беспорядка. В структуре
кристаллов появляются регулярно повторяющиеся слоистые
или стратификационные зоны. В этих зонах происходит
сброс несбалансированного энергетического потенциала.
Страты позволяют сглаживать несовершенство кристалла.
На
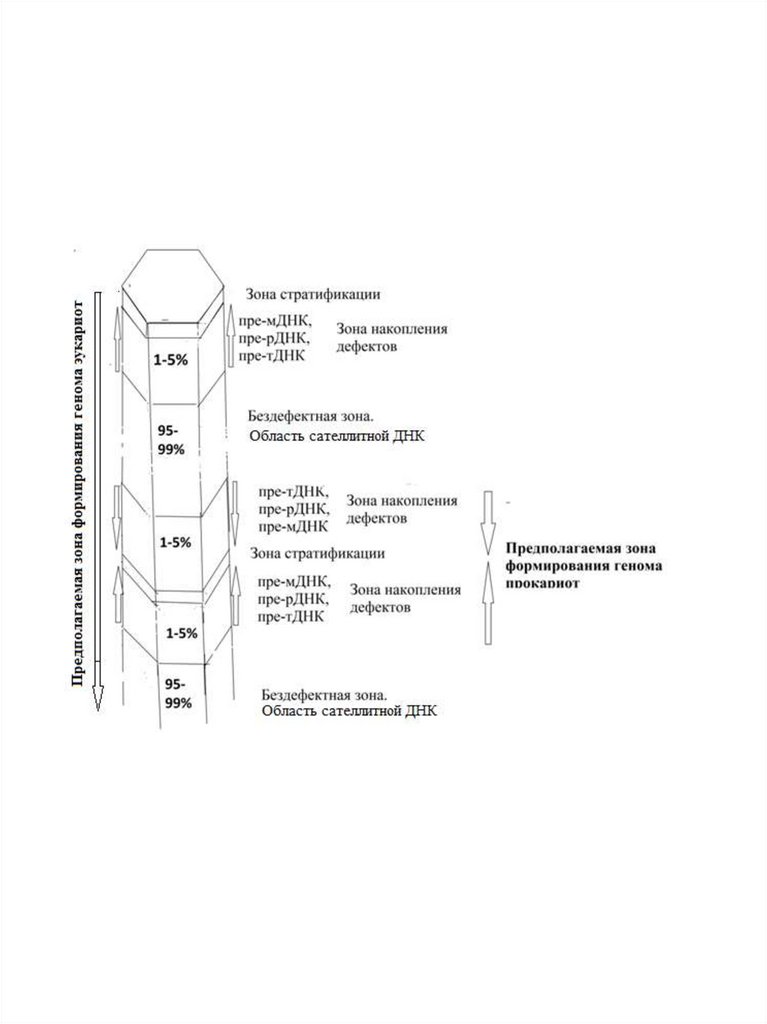
рис.
1D(F)
показаны
зоны
стратификации,
проявляющиеся на макроуровне. Все выше описанные
переходы с постепенным нарастанием
92.
беспорядка в дефектной зоне до зоны страт реализуются в структуреформирующегося
органоминерального
кристаллического
нуклеопротеидного комплекса и транскрипционно-трансляционного
аппарата клетки. Наличие дефектных зон и система согласованных
пропорций в решетках комплекса минералов задают единый алгоритм
построения структуры и процесса образования молекулярных и
надмолекулярных комплексов будущих протоклеток.
Общие принципы формирования ДНК-РНК-белкового
кристаллического органо-минерального комплекса и матричного
аппарата будущих протоклеток
Первый этап формирования органо-минерального кристаллического
комплекса будущих протоклеток происходил в отсутствии воды, под
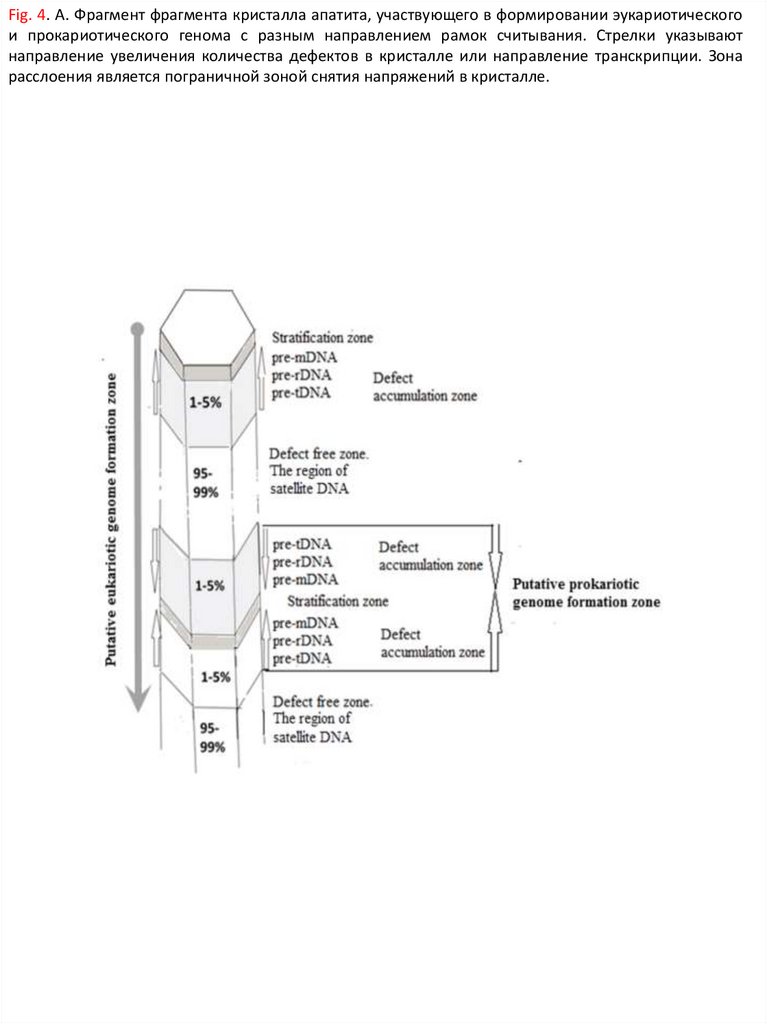
управлением комплекса минеральных ячеек. На Figure 4 приведена
схема фрагмента кристалла апатита, участвующего в формировании
эукариотического и прокариотического генома. Стрелками указано
направление увеличения количества дефектов в кристалле,
совпадающее с будущим направлением транскрипции, а также
предполагаемая зона формирования бактериального генома или его
фрагментов в виде плазмид (Fig.4). Два фрагмента дефектной части
кристалла, имеющие разное направление нарастания дефектов до зоны
страт, способны сформировать кольцевой прокариотный геном с разным
направлением рамок считывания (Fig.4). Стратификационная зона
является пограничной зоной снятия напряжений в кристалле и
одновременно зоной изменения направления снятия считывания
информации или транскрипции.
93.
Fig. 4. А. Фрагмент фрагмента кристалла апатита, участвующего в формированииэукариотического и прокариотического генома c разным направлением рамок
считывания. Стрелки указывают направление увеличения количества дефектов в
кристалле или направление транскрипции. Зона расслоения является пограничной
зоной снятия напряжений в кристалле.
94.
На рис.5 приведен один из фрагментов дефектной зоныкристаллической решетки апатита и карбонат-апатита с общей
схемой
формирования
матричного
аппарата
будущих
протоклеток. На рисунке не приводится бездефектный
фрагмент зоны апатита ответственной за сателлитную
(нетранслируемая) область ДНК. При участии элементов
газовой фазы (NH3, CH4, СО, H2S) в безводной среде, в зоне
апатита и карбонат-апатита по принципу «спираль в спираль»,
образовывался
тройной
–
ДНК,
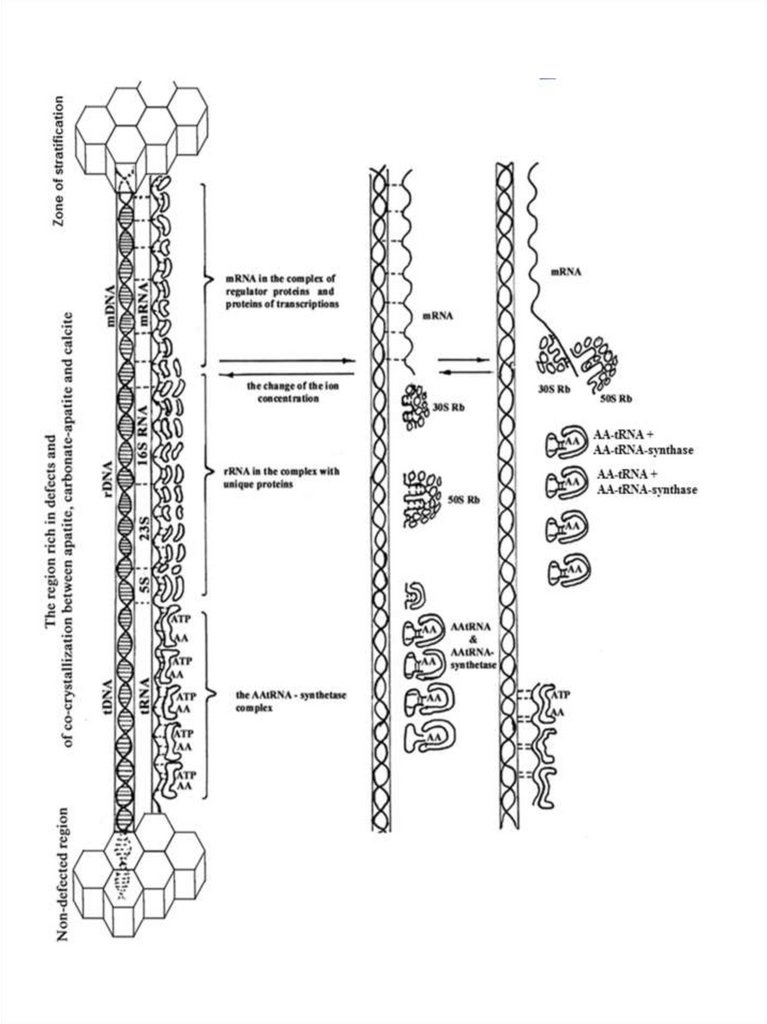
РНК,
белковый
кристаллический органо-минеральный комплекс. Спираль ДНК
в
структуре
комплекса
играла
ключевую
роль.
Супрамолекулярное взаимодействие в спиральном комплексе,
осуществлялось
по
принципу
комплементарности.
Гидрофобные
и
электростатические
взаимодействия
заряженных функциональных групп, водородные связи и силы
Ван-дер-Ваальса, обеспечивали большую суммарную энергию
и устойчивость молекулярных комплексов. У будущих эукариот
двойная спираль ДНК с помощью тех же связей дополнительно
взаимодействовала с гистоновыми и протаминовыми белками в
сателлитной части ДНК (Fig.1,А,В). В случае гистонов взаимодействие происходило с максимальным участием
водородных связей, в случае протаминов - в основном, за счет
вклада в стабильность комплекса электростатического
взаимодействия отрицательно заряженных фосфатных групп
полианиона
нуклеиновой
кислоты
с
положительно
заряженными аминокислотными остатками белка.
95.
96.
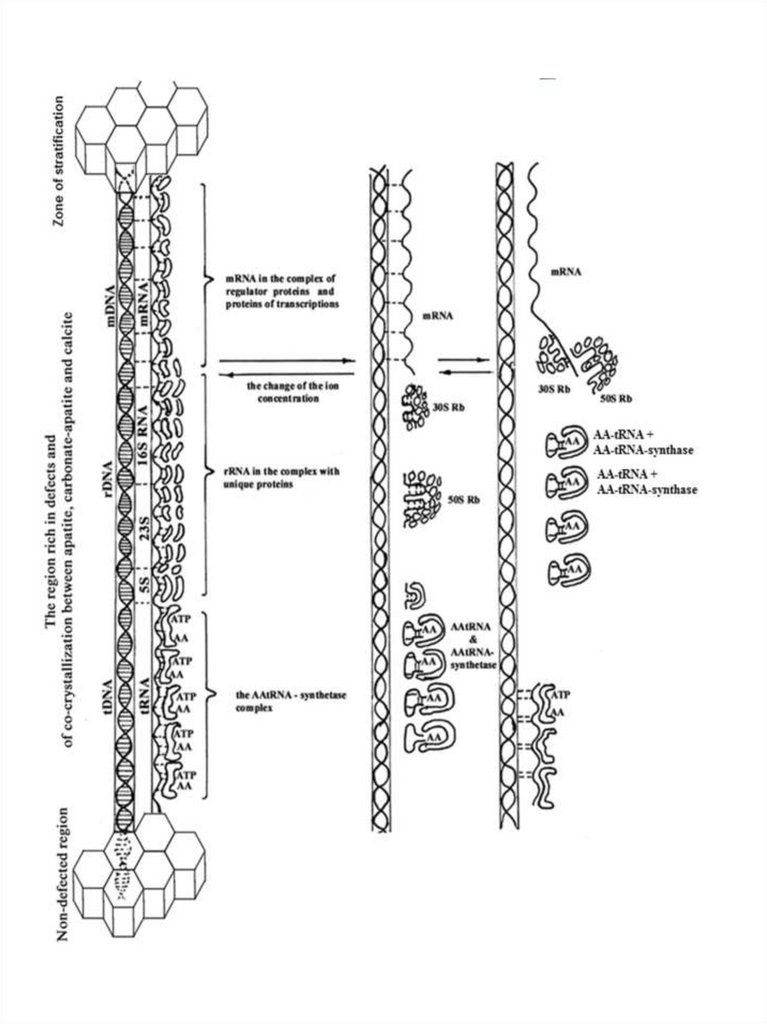
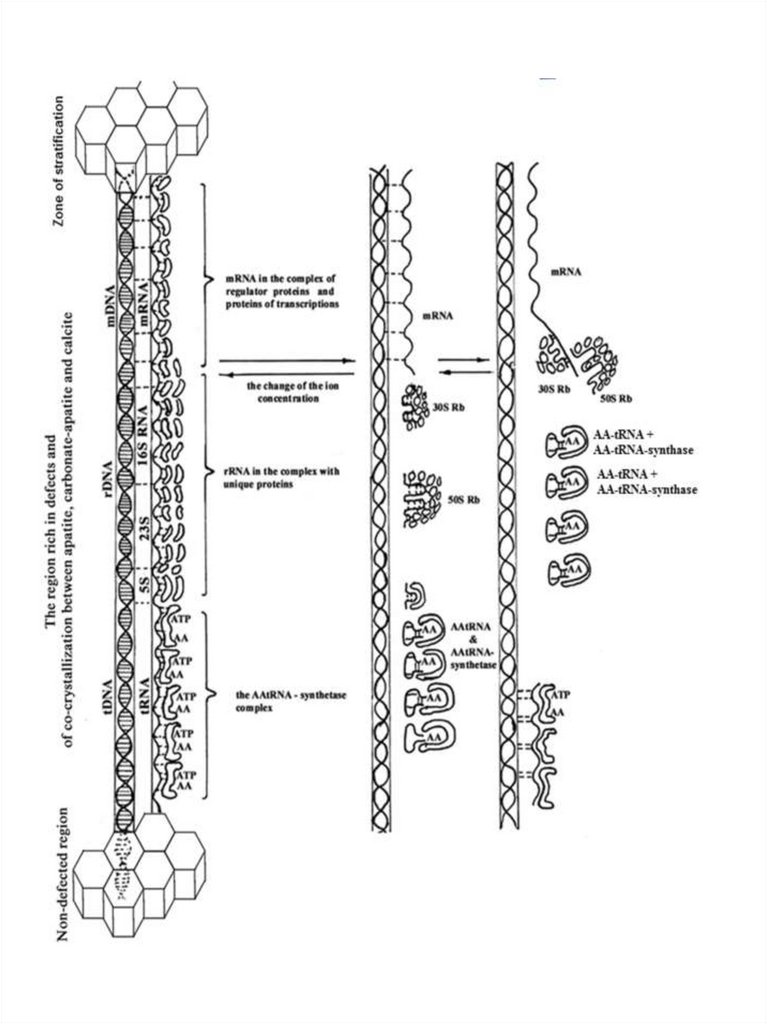
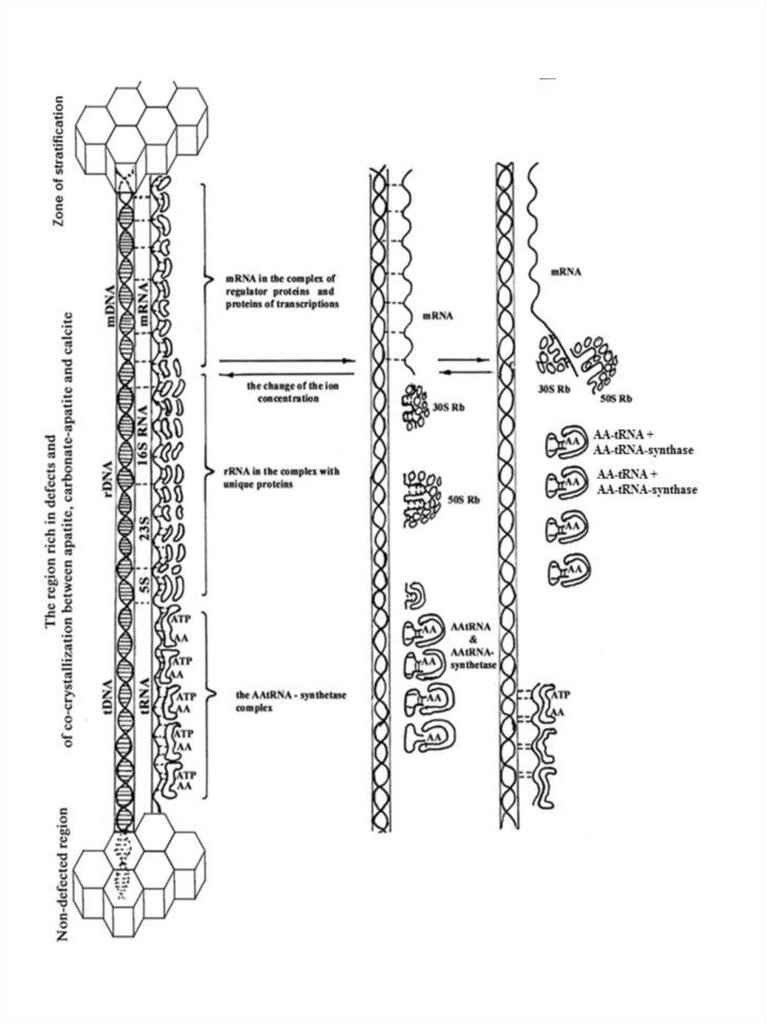
Рис. 5. Предполагаемая схема появления матричного (транскрипционнотрансляционного) механизма в протоклетках в области накопления дефектов исокристаллизации минералов. 1- образование тройного комплекса – ДНК, РНК,
белки; 2 - запуск матричного механизма при появлении воды и изменении
концентрации ионов, путем раскручивания спирализованного комплекса,
формирования рибосом и ААтРНК при участии ААтРНК-синтетазы; 3 –
последовательное продвижение рибосомальных частиц 30S Rb, 50S Rbв
комплексе со своими белками и комплекса ААтРНК с ААтРНК-синтетазой в сторону
мРНК; нарушение связывания мРНК с Ап-ДНК при подходе рибосом и
ААтРНКсинтетазного комплекса и начало синтеза белка.
Тройной комплекс ДНК, РНК, белок будущего
транскрипционно-трасляционного
аппарата
протоклеток
формировался при участии двойной спирали ДНК, спирали
РНК и белков на уровне вторичной структуры. Cотни, а
возможно и большее число различных белков, в том числе
будущие белки-ферменты, связанные с ДНК и РНК,
объединялись
с
помощью
стереоспецифической
комплементарности и максимального участия водородных и
ионных связей. Все белки, кроме гистонов и протаминов,
формировались на карбонат-апатите в комплексе с разными
РНК. Уже на первом, безводном этапе формирования
комплекса, имелись все белки, необходимые для репликации,
транскрипции и трансляции, связанные со своими РНК. В
случае
стереоспецифической
комплементарности
определённый участок белка связывался с нуклеотидной
последовательностью ДНК или РНК. Вклад водородных
связей,
образующихся
между
нуклеотидными
и
аминокислотными остатками белка, максимален, благодаря
пространственному соответствию фрагментов.
97.
Представление о структуре и связях в органоминеральном кристаллическом комплексе ДНК-РНКбелок дает кристаллическая структура взаимодействияРНКазы Н3 с РНК/ДНК субстратом вне минерального
комплекса (Рис. 6). Особо обращает на себя внимание
структура белка не имеющая жестких ограничений в
пространстве.
Рис.6. А-кристаллическая структура комплекса РНКазыH3-РНК/ДНК субстрата.
ДНК показана синим цветом, а РНК показана красным цветом. Б- структура
комплекса с белком в поверхностном представлении[26].
98.
Рис.11.
Общая
схема
участия
комплекса
сокристаллизующихся
минералов
(апатит
карбонатапатит - кальцит) в формировании ключевых
структурных образований будущих протоклеток.
99.
Синтез тройного комплекса идет однонаправленно и одновременнона
комплексе
сокристаллизующихся
минералов,
имеющих
взаимосогласованные пропорции. Продукт такого комплекса - самоорганизация
в единое целое. Такая самоорганизация находится под прямым воздействием
структурных изменений в комплексе минералов.
В структуре минералов идет постепенный переход от бездефектной
области к максимальному накоплению беспорядка в зоне страт (рис.4,5).
Изменение в структуре минералов сопровождается формированием
генов с разными размерами и спецификой. Происходил постепенный переход
от сателлитной зоны ДНК у эукариот к многократно повторенной зоне пре-тДНК,
умеренно повторенной зоне пре-рДНК и уникальной зоне пре-мДНК.
Одновременно
с
этими
областями
в
зоне
карбонат-апатита
сокристаллизующейся с апатитом, комплементарно и специфично,
формировались транскрипты некодирующих РНК (пре-нкРНК), пре-тРНК,
пре-рРНК и пре-мРНК. Транскрипты пре-нкРНК вполне могли локализоваться в
виде нерегулярных промежутков разной величины от сотен до тысяч
нуклеотидных пар в сателитной области ДНК, предшествующей пре-тДНК.
Нуклеотидный размер функциональных РНК нарастал от десятков в
тРНК до тысяч в рРНК и десятков и сотен тысяч в мРНК.
Все транскрипты РНК, помимо комплементарной связи с ДНК,
посредством межмолекулярных взаимодействий, формировали в зоне карбонатапатита супрамолекулярные РНК – белковые и белок-белковые комплексы,
многие из которых – будущие белки-ферменты. Формирование РНК-белковых
комплексов на карбонат-апатите, стало возможным благодаря замене частиРО43групп в структуре ячейки на СО32– группы[1], Fig.1,С.
Свободные
фосфатные группы в составе карбонат-апатита, не
задействованные в цепях РНК, могли быть использованы в образовании
АТФ, ГТФ, ЦТФ, УТФ, а также фосфарилированных белков. Карбонат-
апатит, как ключевая матрица для создания транскрипционно-трансляционного
аппарата будущих протоклеток крайне нуждался в том чтобы иметь в своем
составе макроэрги и фосфорилированные белки. Без этих структур превращения
органо-минерального кристаллического комплекса в жидкокристаллический
органоминеральный
невозможно.
комплекс
или
формирование
протоклетки
было бы
100.
Количество белков взаимодействующих со своими РНКопределялось количеством нуклеотидов в их составе. Для
комплексов пре-тРНК-белок речь могла идти о нескольких белках
при количестве нуклеотидов в РНК до 90-100 штук. Пре-рРНК и премРНК, тоже формировали супрамолекулярные комплексы с
белками, но значительно более крупные чем у тРНК в силу большей
протяженности их фрагментов. Для комплексов пре-рРНК-белок
речь уже идет о десятках белков при количестве нуклеотидов в РНК
до 3000-4000 штук. Для комплексов пре-мРНК-белок о количестве
белков говорить затруднительно, так как размерность мРНК сильно
варьирует, но речь может идти о сотнях белков. Будущаяструктура и
функция подобных комплексов в значительной степени зависит от
качества нуклеотидного состава, которое определяет в цепях РНК
обращенные и прямые повторы, интронные вставки. О влиянии
количества и качества нуклеотидных повторов на структуру и
функцию сформированных комплексов мы поговорим подробнее
во второй части где речь пойдет о превращении
органоминерального комплекса в жидкокристаллический в
присутствии воды. Стабильность структур в таких комплексах у
различных нуклеопротеидов осуществлялась по тому же принципу
стереоспецифичного и стереонеспецифичного нуклеино-белкового
взаимодействия, но более разнообразного: преимущественно за
счет водородных связей и электростатического взаимодействия. В
случае специфичного взаимодействия вклад водородных связей
был максимален. Мы предполагаем, что ячейка карбонат апатита, в
которой шел синтез одной спирали РНК, комплементарной ДНК, за
первым слоем белков, непосредственно взаимодействующих с РНК,
могли следовать слои других стереоспецифично связанных с ними
белков. Ячейка карбонат апатита это вполне допускала. Многие из
этих белков, в будущем, при появлении воды, могли принимать
участие в процессинге пре-рРНК и пре-мРНК, транскрипции и
трансляции, формировании рибосом, осуществлении первичного
метаболизма и гомеостаза клетки.
Именно так, по нашему мнению, появились те десятки, сотни, а
возможно и тысячи индивидуальных белков для создания рибосом
и будущего транскрипционно-трансляционного аппарата, хотя
система синтеза белков к тому времени еще не была готова.
101.
Переход к формированию транскрипционно-трансляционногоаппарата у будущихпротоклеток
Появление на поверхности земли воды со слабо кислой pHи ее
проникновение
в
состав
ДНК-РНК-белкового
органо-минерального
кристаллического комплекса, привело к разрушению минеральной основы
комплекса
и выходуДНК-РНК-белкового кристаллического комплекса.
Последний, в присутствии воды, осуществил переход в жидкокристаллическое
или мезоморфное состояние. Так появились протоклетки, находящиеся в
функционально активном жидкокристаллическом состоянии. Вода и изменение
концентрации ионов приводили к
частичному раскручиваниюДНК-РНКбелкового спирального комплекса и его переходу в метастабильное состояние
или в состояние неустойчивого равновесия. Любоеизменение концентрации
ионов могло способствовать изменению равновесия в системе и запуску
матричного механизма.Запуск сопровождался дальнейшим раскручиванием
компонентов
спирального
комплекса.
Происходило
ослабление
комплементарного и стереоспецифического взаимодействия компонентов
комплекса
и,
как
следствие,
изменение
ихконформации.
Часть
белковприобретала
свойства
ферментов
и
осуществляла
запуск
ферментативных реакций.Компактность комплекса предопределила единство
процессов в протоклеткеи ее будущий гомеостаз. Перестройка конформации в
цепях РНК и прилегающих к ним белков сопровождалась движением
фрагментов комплекса по цепи ДНК. Вода, ионы и подвижка спирального
комплекса наиболее эффективно воздействовали на фолдинг белков.
Первичная структура белков имела слабую стереоспецифическую связь с
фрагментами РНК, в отличии от связи ДНК-РНК и легко переходила в другие
типы структур, активно влияя на стереоспецифически связанные фрагменты
РНК.
Формирование АА-тРНКсинтетазного комплекса. .Рассмотрение запуска
матричного механизма мы начнем с процесса связанного с тРНК. Максимальное
количество нуклеотидов входящих в состав пре-тРНК около 100. Количесство
нуклеазных белков-ферментов, участвующих в подготовке тРНК к синтезу
на рибосоме, исходя из литературных данных около 5 штук. Они должны быть
способны распознавать и связываться с 2'-ОН-группами цепи РНК и фосфатными
группами ДНК после индуцирования ее конформации [5]. Это должен быть
плотный комплекс взаимозависимых друг от друга белков, подготавливающих
тРНК. Предполагается, что пре-тРНК в тройном минеральном комплексе
стереоспецифически взаимодействовала слева (в области, 5'-конца тРНК) с
будущуйРНКазойР, в состав которой войдет отщепленный фрагмент РНК из
пре-тРНК и сделает белковый фермент рибозимом. Его функция –
отщепление лидерной зоны от пре-тРНК.
102.
Справа (в области 3'-конца тРНК) пре-тРНК взаимодействовала сбудущей РНКазойD, функция которой заключается в формировании на
3'-конце последовательности ЦЦА. В подготовке тРНК могли
участвовать и другие
белки-ферменты, например, участники
модификации
оснований
тРНК.Между
5'
и
3'-концами
тРНК,располагалась будущая аминоацил(АА)-тРНК-синтетаза. Это
уникальная аминокислота, которая имела три активных центра: центр
связывания аминокислоты, АТФ и тРНК.Синтетазаизменяла свою
конформацию одновременно со своей тРНК, активировала
аминокислоту с помощью АТФ, ацилировалатРНК активированной
аминокислотой и формировала антикодон на тРНК под свою
аминокислоту с помощью компенсаторной петли.Заслуга в
формировании антикодона у тРНК, с большой вероятностью,
принадлежит АА-тРНК-синтетазе, так как исходно тРНК и мРНК
транскрибируются в виде кодонов с одной цепи ДНК. Изменению
конформациитРНК
и
ослаблению
ее
комплементарного
взаимодействия с ДНК, активно способствовало наличие в цепи
большого числа обращенных повторов [27] и усиление
стереоспецифичного взаимодействия с будущейААтРНК-синтетазой.
Первичная структура тРНК легко превращалась в третичную. Третичная
структура тРНК и специфичность ее антикодона по отношению к
аминокислоте взаимозависимы от ААтРНК-синтетазы [23]. Фермент,
как конформационно более активная часть своего комплекса, сам
подгонял под себя структуру тРНКс помощью компенсаторной петли и
определял ее антикодоновую специфичность, т. е. формировал
антикодоновую петлю, оставляя необходимый для антикодона
триплет. Видимо, в этом кроется ответ на вопрос о том, как столь
различные для одной аминокислоты изоакцепторные тРНК
узнавались АА-тРНК-синтетазой. В различиях между изоакцепторными
тРНК не видно никакой закономерности. Иногда они минимальны,
иногда столь велики, что сопоставление первичных структур тРНК не
дает ни малейших указаний на их одинаковую специфичность.
Сформированный АА-тРНК-синтетазный комплекс пришел в движение
в направлении рибосом. .
103.
Формирование рибосом.Неактивная на первом этапе пре-рРНК, послепоявления воды, для своего процессинга, как и тРНК, нуждалась в помощи
рибосомных или других белков [28], без наличия которых синтез белков
транскрипционно-трансляционного аппарата в клетках невозможен. Эти белки, как
было отмечено выше, в большом количестве уже могли сформироваться в ДНК-РНКбелковом комплексе и послужить основой для запуска транскрипционнотрансляционного аппарата.
Пре-рРНК, в отличие от пре-тРНК, взаимодействовали с большим количеством
белков, стереоспецифически связанных как с рРНК, так и междусобой (рис.10). Повидимому, в этом состоит ответ на вопрос, откуда брались белки для структур
рибосом еще до начала их функционирования. Рибосомальные РНК отличает очень
большое число (около 85%) обращенных повторов [27], поэтому при раскрутке
спирали ДНК-РНК-белкового комплекса, в присутствии воды, РНК-белковая часть
комплекса, взаимодействуя между собой, легко переходила из линейной структуры
в структуру «гусеницы», несущей на себе стереоспецифично и комплементарно
связанные белки (рис.10). Эти РНК-белковые комплексы фактически превращались в
рибосомальные субъединицы, которые, двигаясь по ДНК, взаимодействовали с
мРНК и уже были готовы к синтезу белков.
104.
Запуск транскрипционно-трасляционного аппаратаВ данном разделе для рибосом мы будем оперировать размерами взятыми
для E.coli, хотя это могли быть любые размеры других рибосомальных
частиц.Длинный фрагмент пре-рРНКподвергался процессингу с выделением малой
рРНК 16S в комплексе со своими уникальными белками (30SRb субъединица) и
большой рРНК 23S с малой частицей 5S тоже в комплексе со своими уникальными
белками (50SRb субъединица). Уникальность белков состоит в том, что они не
повторяются в составе рибосомальных субъединиц. Малая рибосомальная частица
(30SRb) первой подошла к мРНК и перевела ее спаривание с апатитовой ДНК на
себя. Этот процесс соответствует образованию первичного комплекса между 30SRb
субъединицей и мРНК при синтезе белка. Далее, наиболее вероятно, подошел ААтРНК-синтетазный комплекс, который передал30SRbаминокислоту и уступил свое
место подошедшей 50SRb субъединице. К образованному полному
рибосомальному комплексу с мРНК в большом количестве подходили АА-тРНК со
своими АА-тРНК-синтетазами (рис.5). Из литературы известно, что большая часть
АА-тРНК-синтетаз неспецифически ассоциированна с функционирующими белоксинтезирующими рибосомами или полирибосомами. Синтетазы при этом
проявляют сродство к их мРНК и рРНК и обеспечивают их частичную
компартментализацию [23]. Ассоциация синтетазного комплекса с рибосомами
довольно лабильна и обратима.
Слоистая упаковка белков, комплементарных РНК, была способна на первом
этапе обеспечить запуск транскрипционного аппарата. Последующий синтез белков,
входящих в состав рибосом, мог осуществлятся за счет их синтеза на рибосомах с
помощью АА-тРНК-синтетаз, гены которых, рассеянны у бактерий по хромосоме.
Известно, что бактериальные рРНК, как правило, представлены
несколькими оперонами РНК (у E. coliих 7), а рибосомальные белки собраны
в полицистронные опероны [23,27]. Предполагается, что первые, уже
готовые рибосомы (из рРНК-белкового комплекса), вступают во
взаимодействие с мРНК, образуя полирибосомные комплексы. Матричные
РНК могут быть полицистронными оперонамирибосомальных белков.
Происходит первый синтез рибосомальных белков для
семи новых рибосом у E. coli. Это и есть момент запуска
истинного матричного механизма, который будет
сохраняться живыми системами все последующие
миллиарды лет в виде готовых резервных рибосом.
105.
Предполагается, что на месте ушедших тРНК и рРНК вновь активноосуществлялась полимеразная сборка их цепей, РНК полимераз нового
рибосомального синтеза. Мы не исключаем, что в такой самосборке могли вновь
участвовать тРНК и рРНК или их фрагменты, уже выполнившие свою функцию в
предыдущем цикле. Количество новых рибосом всегда было не менее числа
рРНК. Часть из новых рибосом могла вступать во взаимодействие с оперонами
мРНК, а часть невостребованных рибосом могла накапливаться в клетке, как
резерв для запуска матричного механизма. Возможно, это те тысячи рибосом,
освободившиеся после синтеза 40000–60000 молекул АА-тРНК-синтетаз (по 2000–
3 000 на одну аминокислоту, или по 500–800 на 1 ген тРНК), которые присутствуют
в клетках E. coli[23, 29]. Наличие рибосом, АА-тРНК-синтетаз и мРНК в
протоклетках обеспечило их будущее существование и развитие.
Восстановление концентрации ионов, вызвавшей запуск матричного
механизма в ДНК-РНК-белковом комплексе, после проведения цикла реакций
должно приводить к восстановлению метастабильного состояния. В дальнейшем
подобные циклы в протоклетках будут соответствовать циклам экспрессии генов.
Описанный выше процесс формирования матричного механизма у разных
таксономических групп мог продолжаться в течении всей архейской эры
формирования земли от 4 млрд.л.н. до 2.5 млрд.л.н. Это период активного
формирования земной коры и гидросферы, появления первых прокариотных
анаэробных гетеротрофов – цианобактерий, одноклеточных нитчатых водорослей
– сфероформид, первых эукариот - микроорганизмов царства грибов, схожих с
дрожжевыми грибами. К концу архея появляются многоклеточные организмы и
условия для их существования.
АА-тРНК-синтетаза, как ключевой носитель генетической
информации
Сформулировано новое представление об истинном носителе
генетической информации-АА-тРНК-синтетазе, единственном белкеферменте,
обладающем
сверхспецифичностью.Синтетазастереоспецифически и комплементарно
связана со своей кодоновойтРНК (структурно сформированной при
непосредственном участии синтетазы), АТФ и аминокислотой. Она
способна превращать кодоновуютРНК в антикодоновуюво всех
изоакцепторныхтРНК и определять аминокислоту для своей тРНК, которая
будет представлена совместно с АА-тРНК-синтетазой виртуальному
носителю генетической информации - ДНК, в лице кодонов мРНК в составе
рибосом.
106.
Принято считать, что ключевым носителем генетическойинформации
является
ДНК.
Уэукариот
97–99%
ДНК
самовоспроизводится, но не несет генетической информации и
только 1–3% через мРНК имеют доступ к информации в виде
белка, но не несут ее. У прокариот информационная часть
достигает 98%. Информационная часть ДНК в виде кодонов
мРНК не способна определить какая аминокислота закодирована
ее кодонами. Она способна только подбирать комплементарные
антикодоны со своей аминокислотой. Эту функцию выполняет
АА-тРНК-синтетаза узнающая необходимую аминокислоту,
формирующая для нее структуру тРНК с необходимым
антикодоном и ацилирующая эту тРНК с помощью АТФ.
Возникает вопрос, если не ДНК, то какие структуры несут
генетическую информацию? Возможно это будут 20 АА-тРНКсинтетаз, узнающих только свою аминокислоту и свою тРНК для
нее, сформированную ферментом из различных изоакцепторных
форм тРНК с нужным антикодоном, вырожденным по третьему
нуклеотиду или это будет структура ДНК в виде мРНК,
являющаяся виртуальным носителем генетической информации.
Задача такого носителя сводится к наличию в своем составе
нужного кодонового триплета. Здесь следует напомнить, что все
наши рассуждения и выводы связаны с синтезом ДНК-РНКбелкового
жидкокристаллического
органо-минерального
комплекса
и
началом
функционирования
его,
как
транскрипционно-трансляционный аппарат. АА-тРНК-синтетазы,
единственные
из
ферментов,
которые
обладают
сверхспецифичностью и явяются истинными носителями
генетической информации. Их 20 по одной на каждую
аминокислоту. Они обладают тремя центрами связывания и
узнавания: АТФ, своей аминокислоты и тРНК. Задача
сверхспецифичности синтетазой решается на стадии активации
аминокислоты в процессе ее специфического отбора[30]и на
стадии взаимодействия с тРНК, так как тРНК больше 60 штук и во
многих чертах они имеют сходную структуру.
107.
Не вполне понятен источник нужной аминокислоты, появляющейся внужное время и в нужном месте. Invitro этот механизм изучен, но invivo– не
известен [30].Это одна из 20 кодируемых аминокислот, входящих в состав
белка, хотя в живых системах их около 500. Складывается впечатление, что
нужная аминокислота присутствует в составе АА-тРНК-синтетазы до начала ее
включения в состав тРНК и разгадка генетического кода аминокислот лежит
именно в особенностях структуры синтетазы.Взаимодействие АА-тРНКсинтетазы с тРНК — это ключевая проблема белково-нуклеинового узнавания
или переноса генетической информации в клетке. Синтетазный белок будущей
АА-тРНК-синтетазы, на этапе безводной фазы, плотно взаимодействует с ДНКРНК спиралью в области пре-тРНК и ему на этом этапе пока безразлична
величина и изоморфная форма будущей тРНК, но не аминокислота.
Мы полагаем, что при появлении воды, будущие белки-ферменты,
связанные с «незрелой» пре-тРНК, помогли удалению концевых 5' и 3'
фрагментов и способствовали взаимодействию АА-тРНК-синтетазы с
центральной частью пре-тРНК. Появление третичной структуры у синтетазы,
сопровождалось усилением стереоспецифического взаимодействия с тРНК и
ослаблением комплементарного ДНК-пре-тРНК взаимодействия. Этому активно
содействовало наличие большого числа обращенных повторов у тРНК. Для
синтетазы важно наличие в кодоновойобласти пре-тРНК последовательности,
которая станет антикодоновой для кодируемой ею аминокислоты. Третичная
структура тРНК и специфичность ее антикодона по отношению к аминокислоте
взаимозависимы
от
ААтРНК-синтетазы
[5].
Нужный
антикодон
изоакцепторныеАА-тРНК легко получали с помощью компенсаторной петли,
формируемой АА-тРНК-синтетазой.При этом синтетазе важны в антикодоне
только две первых позиции триплетного антикодона, но не вырожденная
третья. Соответствие антикодона и аминокислоты определялось с помощью
стереоспецифичного взаимодействия АА-тРНК-синтетазы, комплементарно
связанной с антикодоном своей тРНК, АТФ и кодируемой аминокислоты, которая
будет представлена кодону мРНК. Кстати, что и делает с успехом АА-тРНКсинтетаза в клетке, не допуская подмены своей АК и изменения антикодона.
Получается, что за формирование антикодона на тРНК в тройном комплексе
отвечает АА-тРНК-синтетаза, так-как именно она узнает своюАК, а не премДНК, с которой связана исходная пре-тРНК. Встреча антикодона АА-тРНК со
своим кодом на мРНК рибосомы, произойдет позднее, по мере продвижения по
спирали ДНК совместно со своей АА-тРНК-синтетазой. Из литературных данных
следует, что большая часть АА-тРНК-синтетазнеспецифическиассоциированна с
функционирующими рибосомами или полирибосомами. Ассоциация
синтетазного комплекса с рибосомами довольно лабильна и обратима. При
этом,синтетазы проявляют сродство к мРНК и рРНК рибосом обеспечивая их
частичную компартментализацию [23].
108.
Таким образом получается, что истиннымносителем генетической информации, является
аминоацил-тРНК-синтетаза, а кодон мРНК (область
антикодонов пре-мДНК) это виртуальный носитель,
который не знает, какая аминокислота ей будет
предложена тРНК, также, как и сама тРНК не знает, что
приготовила для нее АА-тРНК-синтетаза.
109.
К вопросу о темной генетической материи эукариотических систем.Участки ДНК, которые не отвечают за синтез белков, но
присутствуют в виде белок-некодирующих РНК (нкРНК, ncRNA),
долгое время считались чем-то вроде генетического мусора или
«генетической темной материи». Однако, оказалось, что они
способны выполнять инфраструктурную и регуляторную функцию в
различных процессах жизнедеятельности клеток, таких как
пролиферация, дифференцировка, апоптоз и др., а их количество в
геноме человека превышает белок-кодирующие гены в два раза [3133]. Исходя из предложенной нами теории происхождения жизни на
минеральных кристаллах [1-3], в бездефектной и дефектной зоне
органо-минерального жидко-кристаллического комплекса будущих
эукариот, генам пре-тРНК, пре-рРНК и пре-мРНК могут
предшествовать нерегулярные промежутки белок некодирующих
РНК (ncRNA), включающие от сотен до тысяч нуклеотидных пар,
которые
могут
рассматриваться
как
«темная
материя»
эукариотических систем.
Некодирующие РНК, как и остальные РНК могут формировать
ДНК-РНК-белковые комплексы, белки которых при появлении воды
помогут процессингу с освобождением нкРНК. Подобные нкРНК,
являясь
структурными
единицами
жидкокристаллического
комплекса, способны выполнять инфраструктурную и регуляторную
функцию и имеют прямое отношение к сохранению целости жидкокристаллического,
гомеостатического
состояния
будущих
протоклеток, без которых клетки существовать не смогут.
110.
К вопросу о нанобактериях?Термин нанобактерии впервые ввёл Ричард Морита (RichardMorita) в
1988 году, однако «отцом» нанобактерий считается Роберт Фолк
(RobertFolk), который начиная с 1992 опубликовал серию работ по
нанобактериям.Нанобактерии-круглые либо овальные минеральные
структуры размером от 30 до 200 нм, которые вызвали один из самых
значительных споров в современной микробиологии. В настоящее время
окончательно
установленно,
что
нанобактерииэто
неживые
кристаллизованные наночастицы минералов и органических молекул,
появление которых связано с процессами биоминерализации в живых
системах аморфных сферических частиц гидроксиапатита и карбоната
кальция. Молекулы апатита, при этом, являются центром кристаллизации, с
чем и связан наблюдаемый «рост» и «размножение»кристаллов. Процесс
образования подобных мелких частиц из минералов в комплексе с
органическими молекулами, является частным случаем описанной выше
теории зарождения жизни на минеральных кристаллах апатита и карбонатапатита так же, как образование неживых частиц фагов и вирусов[1-3].
Почему генетический код универсален и триплетен для всех
АК при вырожденности по третьему нуклеотиду?
АА-тРНК-синтетаза, стереоспецифически и комплементарно
связана со своей тРНК, АТФ, АК и определяет для своей тРНК
аминокислоту и структуру единого вырожденного антикодона в
изоакцепторныхтРНК. В антикодоне значащими остаются только
два первых нуклеотида для любых АК. Мы предполагаем, что
универсальность и триплетность кода определяется размерами
элементарной ячейки апатита и карбонатапатита, в структуре
которых формируются 3 пары пурин – пиримидиновых оснований
на высоте 0, 50,100 (область CaI), при этом третья пара всегда
оказывается на границе ячеек. Эта область характеризуется
повышенной флуктуацией энергии и, по нашему мнению,
является причиной вырожденности генетического кода по
третьему нуклеотиду [1-3]. Никакого пространственного
соответствия между тремя или двумя нуклеотидами и одной
аминокислотой нет. Это соответствие выполняется АА-тРНКсинтетазой.
111.
О синтезе протоклеток без нарушения второго закона термодинамикиСогласно предложенной нами теории происхождения жизни при участии
элементов газовой фазы (NH3, CH4 и СO), апатитовой матрицы и
сокристаллизующихся с ней минералов (карбонат-апатит, кальцит), шло
постепенное замещение минеральной матрицы на органо-минеральный
кристаллический комплекс с последующим его переходом, в присутствии воды,
в жидкокристаллическое или мезоморфное состояние (протоклетка).
Смена фаз проходила без нарушения 2го закона термодинамики и
сопровождалась потерей части энергии кристаллической решетки апатита,
карбонат-апатита, кальцита за счет смены ковалентных и координационных
связей кристаллической решетки на супрамолекулярное взаимодействие с
помощью стереоспецифической комплиментарности. Последняя обеспечивала
образование водородных и ван-дер-ваальсовых связей, гидрофобных и
электростатических взаимодействий отрицательно заряженных фосфатных
групп нуклеиновой кислоты с положительно заряженными аминокислотными
остатками белка. Перечисленные взаимодействия
характеризовались
достаточно большой суммарной энергией, что привело к образованию
устойчивых молекулярных комплексов без образования ковалентной
химической связи. В целом энтропия вновь образованного комплекса
оказалась значительно выше энтропии кристаллической решетки.
112.
ЗаключениеОпираясь на базовые элементы теории, описанные ранее Kostetsky(2005),
рассмотрена теория возникновения матричного механизма на комплексе
минералов, позволяющая по новому осветить ключевые вопросы молекулярной
биологии. В наследство от минералов мы получили ответ на вопрос когда, где,
как и почему возникли будущие протоклетки, а сами протоклетки получили
матричный механизм. Для возникновения протоклеток понадобился комплекс
минералов состоящий из апатита, карбонат-апатита, кальцита, слюды и кварца и
радикалы газовой фазы представленной NH3, CH4 и СO. Для возникновения
матричного механизма в составе протоклеток понадобилась бездефектная и
дефектная зона апатита и карбонат-апатита. Первая зона — это зона будущей
сателлитной части ДНК уэукариот, вторая зона — это будущая область
транскрипционно-трансляционного аппарата. Переход в кристалле апатита от
бездефектной области к постепенному накоплению беспорядка реализовывался
в размерах и специфике формирующихся генов в ДНК. От неинформативной или
сателлитной части ДНК (уэукариот) шло нарастание размеров геномов от пренкДНК к пре-тДНК, пре-рДНК и пре-мДНК. Комплементарно и специфично с
этими областями, в зоне карбонатапатита, сокристаллизующейся с апатитом,
последовательно формировались транскрипты в виде нерегулярных
промежутков белок-некодирующие пре-нкРНК, пре-тРНК, пре-рРНК и пре-мРНК.
Размер транскриптов нарастал по мере «нарастания беспорядка» в апатите и
карбонатапатите от десятков в тРНК до тысяч в рРНК и десятков и сотен тысяч
нуклеотидов в мРНК. Транскрипты РНК, помимо комплементарной связи с ДНК,
посредством межмолекулярных взаимодействий, формировали в зоне карбонатапатитасупрамолекулярные структуры с белками, количество которых росло в
зависимости от размера транскрипта РНК. Так формировался спиральный ДНКРНК-белковый органо-минеральный кристаллический комплекс. Синтезшел по
принципу стереоспецифической комплементарности (спираль в спираль).
Фактически это был готовый транскрипционный аппарат, сформированный
на матрице апатитом и карбонат-апатита из газовой фазы представленной
NH3, CH4 и СO.
Запуск транскрипционного аппарата и превращение органоминерального кристаллического комплекса в жидко-кристаллическкий
комплекс произошло после появления воды. Вода нарушила ионный
баланс в сформированной системе и привела в движение ДНК-РНКбелковый комплекс. Произошло ослабление ДНК-РНК взаимодействия и
усиление РНК- белкового взаимодействия из-за большого числа обращенных
повторов в тРНК и рРНК. Белки такого комплекса смогли превратить тРНК и рРНК
в структурно-функциональные единицы трансляционного аппарата и
осуществить его запуск.
113.
Сверхспецифичность будущей АА-тРНК-синтетазы, обращенные повторы ималые размеры тРНК, с их супрамолекулярным взаимодействием,
предопределили судьбу тРНК, дав ей не только свою аминокислоту, но и
антикодон и сделав ее инструментом доставки аминокислот для синтеза белка на
рибосомах под своим контролем. Рибосомальная РНК, под влиянием обращенных
повторов и большого количества стереоспецифично связанных белков,
превратилась в рибосомальные единицы, которые начали продвижение в сторону
мРНК для подготовки встречи АА-тРНК-синтетазного комплекса и начала этапа
трансляции. Проблема с первыми рибосомальными белками была решена еще на
стадии формирования органо-минеральногоДНК-РНК-белкового комплекса. Этап
трансляции ознаменовался появлением новыхрибосомальных и других белков,
произведенных прокариотами. Рибосомы, сформированные при участии новых
белков, никогда больше не покидали протоклетки и передавались по наследству
(естственно постоянно обновляясь).Востановление концентрационного градиента
ионов вызвавших запуск матричного механизма в ДНК-РНК-белковом комплексе,
после проведения цикла реакций, должно приводить к восстановлению
метастабильного состояния протоклетки. В дальнейшем подобные циклы в
протоклетках будут соответствовать циклам экспрессии генов.
Описанный процесс формирования матричного механизма у разных
таксономических групп мог продолжаться в течении всей архейской эры
формирования земной коры и гидросферы от 4 млрд.л.н. до 2.5 млрд.л.н. Это
период
появления
первых
прокариотных
анаэробных
гетеротрофов
(цианобактерий и одноклеточных нитчатых водорослей) и первых эукариот (
микроорганизмы царства грибов). К концу архея появляются многоклеточные
организмы и условия для их существования.
114.
В ходе проведенного исследования, сформулировано новое представлениеоб истинном носителе генетической информации -сверхспецифичнойАА-тРНКсинтетазе, стереоспецифически и комплементарно связанной со своей
кодоновойтРНК, АТФ и аминокислотой. Показаны ее способности превращать
кодоновуютРНК в антикодоновуютРНК во всех изоакцепторныхтРНК и
определять аминокислоту для своей тРНК, которая будет представлена совместно
с АА-тРНК-синтетазой виртуальному носителю генетической информации - ДНК, в
лице кодоновмРНК.
Рассмотрение процесса возникновения жизни на ранних этапах
формирования земной коры позволило предположить, что механизм
возникновения
структурной
ассиметриибиомолекул
обусловлен
однонаправленностью (от горячего к холодному) и одновременностью синтеза НК
и белков в объеме минералов по принципу «спираль в спираль»; предположить
ответ на вопрос о синтезе протоклеток без нарушения второго закона
термодинамики, путем перехода от кристаллического органоминерального
матрикса к жидкокристаллическому органоминеральному с потерей части
энергии кристалла и увеличения энтропии за счет смены связей кристаллической
решетки
на
супрамолекулярное
взаимодействие,
с
помощью
стереоспецифической комплиментарности. Универсальный генетический код
рассматривается как следствие единой матрицы апатита для всех живых и не
живых систем (вирусы, фаги). Предполагается, что вырожденность генетического
кода по третьему нуклеотиду, есть следствие расположения в элементарной
ячейке апатита только трех пар пурин-пиримидиновых оснований, с позицией
третьей пара всегда на границе ячеек апатита, в зоне повышенной электронноплотностной напряженности. Следствием предложенной теории является новый
взгляд на понятие нанобактерий и темной материи РНК.
115.
Все выше рассмотренные, в виде предположений, единые особенностипроисхождения и организации всех живых систем, являются следствием
единых физико-химических законов структурной организации
кристаллической решетки рассматриваемых нами минералов, ставших
матрицей для происхождения протоклеток.
Мы предполагаем, что предложенный механизм возникновения живых
и неживых систем (вирусов, фагов), гуминового скелета почв, углеводородов
нефти и угля – все это результат взаимодействия компонентов газовой
фазы(H2, NH3, CH4 и СO) с решетками разных минералов. Для биомолекул это
апатит, карбонатапатит, кальцит, кристобалит, арагонит, слюда; для
гумуса почв – алюмосиликаты; для углеводородов нефти –минералы с
различной структурой кристаллической решетки; углей –
минерализованные органические скелеты. Этот механизм является
универсальным для возникновения органических веществ как на Земле так и
других планетах типа Земля.
Acknowledgments
The authorwould like to thank Prof. Nina M. Sanina (Far
Eastern Federal University, Vladivostok), Dr.ArthurYu.Nikulin,
Dr.Vyacheslav Yu. Nikulin (Federal Scientific Center of the East
Asia Terrestrial Biodiversity FEB RAS, Vladivostok)for their
criticalreading and fruitful discussions of the manuscript. The
present work was supported in part by the U.S. Civilian Research &
Development Foundation for the Independent States of the Former
Soviet Union (CRDF) and Ministry of Education of the Russian
Federation (grant No.REC-003) and NATO (PDD (CP)–(NR.
CLG.980842).
116.
117.
118.
Апатит зернистые агрегаты в кварцево-диопсидовойпороде. Забайкалье Слюдянка.
119.
Апатит с нефелином и сфеном Кольский полуостров120.
Апатит в диопсиде Ковдор121.
Апатит кристаллы с кальцитом. Слюдянка.Сопутствующие минералы. Сфен, циркон, пироксен,
амфибол, шпинель, везувиан, флогопит, нефелин, магнез
ит, кварц, полевой шпат, слюды, кальцит.
122.
123.
124.
125.
Апатит впороде
126. ПРОИСХОЖДЕНИЕ ПРОТОКЛЕТОК И ИХ СТРУКТУРНЫХ ЭЛЕМЕНТОВ НА ОСНОВЕ АПАТИТА И СОКРИСТАЛИЗУЮЩИХСЯ С НИМ МИНЕРАЛОВ
Посвящается памятиВ.В.Чернобровкина
ПРОИСХОЖДЕНИЕ ПРОТОКЛЕТОК И
ИХ СТРУКТУРНЫХ ЭЛЕМЕНТОВ НА
ОСНОВЕ АПАТИТА И
СОКРИСТАЛИЗУЮЩИХСЯ С НИМ
МИНЕРАЛОВ
• Костецкий Э.Я.
Отдел Биохимии и Биотехнологии, Дальневосточного
Государственного университета. Владивосток, 690950, Октябрьская
27. Факс: 7-4232-429510. Е-mail: kostetsky@nt.pin.dvgu.ru
• «Теория оказывается тем более впечатляющей,
чем проще ее предпосылки, чем значительнее
разнообразие охватываемых ею
явлений и чем
шире ее применимость»
• А. Эйнштейн
• «Мир слишком совершенен,
возникнуть из хаоса»
чтобы
случайно
127.
128.
Изредка апатит встречается в виде окаменевшего материалаископаемой древесины.
129.
130.
ДБ
В
Г
Рис.1. Б, В, Г - Cтруктура апатита по Н.В. Белову
[Gilinskaya, Scherbacova 1975]: Б - призмы с
катионами Са II; В - призмы с катионами Са I; Г анионы РО4. Д - Апатит с горы Витоша, Болгария
(4/5 натуральной величины). Числа указывают доли
от высоты элементарной ячейки.
131.
АРис.4. А – Структура апатита (вид сбоку) построена
по данным рентгеноструктурного анализа [Kay et al.
1964; Sudarsanan, Young 1969]; А, Б – компьютерная
графика.
132. Пиримидины: структура
Все пиримидины являются производнымигетероциклического соединения - пиримидина. Основные
пиримидины, найденные в РНК,- урацил и цитозин; в ДНК тимин и цитозин.
Биологическая роль пиримидинов не ограничена
нуклеиновыми кислотами. Некоторые пиримидиновые
нуклеотиды играют важную роль в процессах обмена
углеводов и липидов. Витамин В1 (тиамин) - пиримидиновое
производное.
Все пиримидины способны к таутомерным превращениям.
132
133. Пурины: структура
• Пурин, давший название всей группе, содержитшестичленное пиримидиновое кольцо, сочлененное
со вторым кольцом, образованным еще двумя
атомами азота и одним атомом углерода. Аденин и
гуанин - основные пурины нуклеиновых кислот.
• Подобно пиримидинам, пурины склонны к
таутомерии.
133
134.
135.
136.
Рис.1. А - Структура апатита (вид сверху)построена по данным рентгеноструктурного
анализа [Kay et al. 1964; Sudarsanan, Young
1969] с указанием направления
комплементарных цепей ДНК]. Числа
указывают доли от высоты элементарной
ячейки.
137.
Рис.2 - Структура апатита (вид сверху) суказанием положения двух пар оснований ДНК
и фрагментом пептидной цепи (в центре)
[Kostetsky 1981; 1999].
138. Различные конформации сахара
Пятичленноефуранозное кольцо
может принимать
конформацию,
подобную конверту, в
которой четыре атома из
пяти находятся в одной
плоскости, а один атом
выходит из нее на
расстояние до 0.5 Å,
либо изогнутую твистконформацию, в которой
любые два соседних
атома смещены в разные
стороны от плоскости,
построенной по трем
оставшимся атомам
сахарного кольца.
В обоих случаях
атомы, выходящие
из плоскости в ту же
сторону, что и атом
С5', называют эндоатомами, а атомы,
смещенные в
противоположную
сторону, экзоатомами.
Фуранозное кольцо в составе нуклеозидов замещено
несимметрично, поэтому в нуклеозидах и нуклеотидах, фазовые
углы псевдовращения Р распределены по циклу неравномерно.
Они образуют две группы, концентрируясь в интервалах от -1 до
34 градусов (С3'-эндо) и от 137 до 194 градусов (С2'-эндо). В
соответствии с расположением в цикле псевдовращения эти
предпочтительные конформации называют северными (N) и
южными (S).
•Для ДНК характерны две основные конформации сахарного
кольца: С3'-эндо и С2'-эндо (или ее разновидность, С3'-экзо конформация),
переход
между
которыми
определяется
активностью молекул воды окружающей среды, а для РНК
138
всегда наблюдается С3'-эндо-конформация.
139.
Рис.3 - Структура апатита (вид сверху) суказанием положения четырёх пар оснований
ДНК и фрагментом пептидной цепи (в центре)
[Kostetsky 1981; 1999].
140.
141.
АБ
Рис.4. А – Структура апатита (вид сбоку) построена
по данным рентгеноструктурного анализа [Kay et al.
1964; Sudarsanan, Young 1969]; Б – Структура апатита
(вид сбоку) с цепями ДНК введёнными в его состав.
А, Б – компьютерная графика.
142.
БРис.4. Структура апатита (вид сбоку) построена по
данным рентгеноструктурного анализа [Kay et al. 1964;
Sudarsanan, Young 1969] с цепями ДНК введёнными в
его состав. Компьютерная графика.
143. Геометрия связанных пар
•Обычноцентр
пар не совпадает
с осью спирали,
а смещен от нее
на
некоторое
расстояние
D.
Кроме
того,
плоскость пар не
строго
перпендикулярна
оси.
Ее
отклонение
от
перпендикулярн
ого
положения
определяется
двумя углами углом
наклона
(tilt)
и
углом
крена (roll) пары.
Спаренные
основания
не
обязятельно
компланарны.
Они могут быть
повернуты друг
относительно
друга
(вокруг
длинной
оси
пары,
проходящей
через
С6-атом
пиримидина
и
С8-атом пурина)
на
угол
пропеллера
(propeller),
который
равен
двугранному
углу
между
нормалями
к
плоскостям
оснований.143
144.
Рис. 5. Структура связей в комплементарных цепях Ап-ДНК(Рис.2Б). Для наглядности основания даны под небольшим
углом. син-, анти- угол вращения вокруг гликозидной связи
С1`-N ; g - торсионный угол вращения вокруг С4`-С5` связи
имеет an,+ск, -ск конформацию; С2` - эндо- , или когда С2`
более чем С3` удалён от плоскости С1`- О1`- С4` и находится
по одну сторону с С5`; О1` у d-рибоз обеих цепей, имеющих
син- конформацию гликозидной связи, всегда направлен в дГ и
дЦ вниз или вверх относительно оси спирали, а у d-рибоз
имеющих анти- конформацию О1` направлен в сторону от оси
спирали
145.
146.
147.
148.
Сравнение спиралей B-, Z- ДНК и Aп- ДНКПараметры
B- ДНК*(14,15)
Z- ДНК*(14,15)
Aп-ДНК
Направление спирали
Правое
Левое
Левое
Число остатков на виток
10
12 (6 димеров)
12(3
тетраме
р)
Диаметр спирали,
20
18
18
Трансляция на нуклеотид,
3,4
3,7
3,44
Шаг спирали,
34
45
41,3
Наклон пары оснований
6
7
0
Поворот на нуклеотид, град
36
-60 (на димер)
-120 (per
тетраме
р)
d-гуанозин
Anti
Syn
Anti, Syn
d-цитидин
Anti
Syn
Anti, Syn
Sugar pucker
d-гуанозин
d-цитидин
C2 endo
C endo
2
C3 endo
C endo
2
C2 endo
C endo
2
an
+ck
an, +ck
an, +ck, -ck
Угол вращения вокруг гликозидной
связи
Торсионный угол вращения вокруг
C4 -C5 связи
d-гуанозин
d-цитидин
Расстояние фосфора от оси,
d Гф Ц
d ЦфГ
d ГфГ
d ЦфЦ
9
9
9
9
8
6,9
-
8
8,5
5,8
3,6
Расстояние между фосфорами
комплементарных пар,
d (ГфЦ)
d (ЦфГ)
d (ГфГ)
d (ЦфЦ)
17,5
17,5
17,5
17,5
15
12,5
15,3
13,1
8,9
7,9
149.
150.
АБ
В
Рис.6. А - Структура апатита с расположенными в ней
цепями полиглицина II(полипролина II, коллагена). В
зоне формирования белковых цепей в ячейке апатита
анионы РО43- замещены на CО32-. Б,В - Электронная
микрофотография гексагональной упаковки молекулы
белка в кристалле каталазы из печени быка
(увеличение Х5.105) (Б) и кристаллической
лейцинаминопептидазы (В) [Вайнштейн 1986, с.156].
151.
Рис.10. А-кристаллическая структура комплексаРНКазы H3-РНК/ДНК субстрата. ДНК показана синим
цветом, а РНК показана красным цветом. Б- структура
комплекса с белком в поверхностном представлении
[25].
152.
Рис. 7. Предполагаемая схема появления разногочисла хромосом в будущих протоклетках.
153.
Рис. 8. Предполагаемая схема появления разногочисла хромосом в будущих протоклетках.
154.
Роль минералов в возникновении протоклеток
Вести речь о реальных механизмах возникновения
жизни можно и нужно основываясь на твердофазных
эффектах в минеральных и жидких кристаллах.
Участие минералов в процессе возникновения
жизни делает его закономерным, а время процесса
весьма кратким. В клетках, какими мы их
представляем сегодня, все элементы системы так тесно
связаны друг с другом, что отсутствие даже одного из
них нарушает работу всей системы. Первичные
пробионты,
по-видимому,
должны
были
представлять
собой
хорошо
упорядоченные
образования. Возникает вопрос, а не были ли
предшественники
первичных
протоклеток,
органическими
кристаллами?
Подобное
предположение было высказано и А.Кэрнс-Смитом [17,
c.68]. Ведь кристаллы - это образования, способные
к самосборке. У кристаллов есть основной тип
строения,
для
которого
характерна
высокая
периодичность, но в каждом реальном кристалле эта
структура имеет дефекты. Присутствие дефектов,
малых или больших, в структуре кристаллов делает
их потенциально высокоинформативными. Дефекты
кристаллов могут приводить к образованию множества
стабильных альтернативных конфигураций, что
является необходимым условием для хранения
информации.
155.
Рис. 9. Схема образования биогенных кристаллов.Вначале образуется органическая матрица (I),
состоящая из стопки полисахаридных пластин
гексагональной
или
эллиптической
формы
и
«прошита» фибриллами белка. Затем внутри фибрилл
и
между
белковыми
фибриллами
в
самой
полисахаридной
матрице
возникают
зародыши
минеральных
кристаллов;
которые
растут
и
формируют элементарный минеральный кристаллит
(2). Конечная стадия минерализации - образование
минерализованного поликристалла (3). Совокупность
таких кристаллов и составляет скелетную ткань кость, раковину, панцирь и т.п. Схема по С.Н. Голубеву
[13].
156.
Reutilization of purine andpyrimidine bases.
157.
De novo биосинтезпуринового кольца
от PRPP до
инозиновой
кислоты.
PRPP
Asp + GTF
AMF
pp
5-Phosphoribosyl1PP
Gln + ATF
GMF
158.
De novo synthesis of pyrimidinenucleotides.
159.
Рис. 10.Схема действия факторов ослабляющих
кристаллическую
решетку
апатита,
облегчающих
и
ускоряющих
процесс
диффузии
элементов
восстановительной газовой фазы внутрь кристалла при
формировании органоминерального комплекса.
Процесс
идет однонаправленное (от центра Земли к поверхности) по
свободным полостям или каналам в структуре апатита либо
по границе кристаллических зерен, замещая атомы или
вакансии в кристаллической решетке в безводной фазе при
температуре » 200°С.
160.
Рис.11.
Общая
схема
участия
комплекса
сокристаллизующихся
минералов
(апатит
карбонатапатит - кальцит) в формировании ключевых
структурных образований будущих протоклеток.
161.
162.
Рисунок 4. Схема фрагмента кристалла апатита,участвующего в формировании эукариотического и
прокариотического
генома
с
различными
направлениями рамок считывания. Стрелки указывают
направление увеличения количества дефектов в
кристалле или направление транскрипции. Зона
расслоения является пограничной зоной снятия
напряжений в кристалле.
163.
164.
Рис.Предполагаемая
5.
матричного
модель
возникновения
(транскрипционно-трансляционного)
механизма в протоклетках в области накопления
дефектов
и
сокристаллизации
минералов.
(А)
образование тройного комплекса, содержащего ДНК,
РНК и белки; (B) запуск матричного механизма при
появлении воды и изменении концентраций ионов
путем
раскручивания
формирования
рибосом
спирального
и
AAtRNAs
комплекса,
с
участием
AAtRNAsynthetase; (C) последовательное продвижение
рибосомальных частиц 30S и 50S в комплексе с их
белками и комплексом AAtRNA с AAtRNAsynthetase к
мРНК; искажения в связывании мРНК с апатит-ДНК
при сближении рибосом и комплекса AAtRNA с
AAtRNAsynthetase и начале синтеза белка.
165.
Figure 5.The proposed model of the appearance of thematrix
(transcription-translational)
mechanism
in
protocells in the region of accumulation of defects and
co-crystallization of minerals. (A) formation of a triple
complex containing DNA, RNA, and proteins; (B)
triggering the matrix mechanism at the appearance of
water and changes of the ion concentrations, by
unwinding the helical complex, forming ribosomes and
AAtRNAs with the participation of AAtRNAsynthetase;
(C) sequential advancement of 30S and 50S ribosomal
particles in complex with their proteins and the complex
of AAtRNA with AAtRNAsynthetase towards mRNA;
distortions in the binding of mRNA to apatite-DNA at the
approach
of
ribosomes
and
of
AAtRNA
with
AAtRNAsynthetase complex and the beginning of
protein synthesis.
166.
Предложенный механизм формирования органоминерального
комплекса будущих протоклеток на комплексе минералов
позволяет ответить на
основные вопросы проблемы
происхождения жизни: как и почему клетки являются
апериодическими,
гомеостатическими,
самовоспроизводимыми жидкими кристаллами. Отпадают
вопросы относительно синтеза протобиополимеров в воде и
создания упорядоченной живой системы против законов
термодинамики. Однонаправленность и одновременность
синтеза НК и белков в объеме минералов по принципу
«спираль в спираль», по-видимому, сыграла ключевую роль в
возникновении структурной асимметрии мономерных звеньев
и апериодичности биомолекул. В наследство от минералов
будущие протоклетки получили: 1) матричный механизм
синтеза на участке накопления дефектов в решетке апатита и
сокристаллизующихся с ним минералов; 2) универсальный
генетический код (единая матрица апатита) и отсутствие
универсальности в строении структур, участвующих в
реализации этого кода; 3) особенности структуры ДНК
эукариот (сателлитные, умеренно повторенные и уникальные
гены); 4) невозможность удалить сателитные участки ДНК в
силу
необходимости
поддержания
возникшего
гомеостатического состояния живых систем: 5) ключевую роль
Са2+- и РО43--групп в жизнедеятельности живых систем; 5)
белки, специфически связывающие только Са2+ или другие
ионы; 6) живые системы, использующие для построения своего скелета только Sr, Si, Са, РО43--ионы или другие элементы.
Мы предполагаем, что предложенный механизм является
универсальным для возникновения органических веществ на
Земле или других планетах типа Земля. Биомолекулы живых
систем, гуминовый скелет почв, углеводороды нефти – все это
результат взаимодействия компонентов газовой фазы с
решетками апатита, карбонатапатита, кальцита, кристобалита,
для гумуса почв – алюмосиликаты; для углеводородов нефти –
минералы с различной структурой кристаллической решетки.
167.
168. Публикации Э.Я.Костецкого по проблеме возникновения жизни.
Э.Я.Костецкий, Алексаков С.А. О возможности синтеза
нуклеопротеинов на матрице апатита //Докл. Акад. Наук СССР.
1981. Т.260. С.1013-1018.
Э.Я.Костецкий. О происхождении жизни и возможности
формирования протоклеток и их структурных элементов на
кристаллах апатита //Журн. Эволюц. Биохимии и Физиологии.
1999. Т.35. С.249-256.
EDUARD Y. KOSTETSKY. The Possibility of the Formation of Protocells
and Their Structural Components on the Basis of the Apatite Matrix
and Cocrystallizing Minerals // Journal of Biological Physics 2005. 31:
607–638,
Э.Я. КОСТЕЦКИЙ . КАК ВОЗНИКЛА ЖИЗНЬ. ТЕОРИЯ
ВОЗНИКНОВЕНИЯ ПРОТОКЛЕТОК И ИХ СТРУКТУРНЫХ
КОМПОНЕНТОВ // ВЕСТНИК ТИХООКЕАНСКОГО
ГОСУДАРСТВЕННОГО ЭКОНОМИЧЕСКОГО УНИВЕРСИТЕТА.
ВЛАДИВОСТОК, 2008. N1, ЧАСТЬ 1, С.79-102; N2, ЧАСТЬ 2, С. 86104.
Э.Я. КОСТЕЦКИЙ . КАК ВОЗНИКЛА ЖИЗНЬ. ТЕОРИЯ
ВОЗНИКНОВЕНИЯ ПРОТОКЛЕТОК И ИХ СТРУКТУРНЫХ
КОМПОНЕНТОВ ПРИ УЧАСТИИ АПАТИТОВОЙ МАТРИЦЫ И
СОКРИСТАЛИЗУЮЩИХСЯ С НЕЙ МИНЕРАЛОВ // ТРУДЫ
ПРОФЕССОРСКОГО КЛУБА. ВЫПУСК 11. ВЛАДИВОСТОК, 2008. С.5377.
169.
170.
• Кристалл ДНК —апериодический, так как
последовательность пар
оснований нерегулярна, как
нерегулярна
последовательность букв в
осмысленном .. тексте книги. Но
подобно буквам в печатном
шрифте, пары оснований А-Т и
Г-Ц имеют одинаковые
размеры как в ширину, так и в
высоту.[c.36]
• Шредингер назвал организм
апериодическим кристаллом.