





















































































































 biology
biologySimilar presentations:










Биология: её задачи, объект и методы исследования
1.
1) Биология: её задачи, объект и методы исследованияБиология - совокупность наук о жизни, о живой природе (греч. bios - жизнь, logos - учение). Биология – наука, изучающая свойства живых систем.
Объектом исследования биологии являются все живые организмы, включая микроорганизмы, растения, животных и человека.
Метод – это путь исследования, который проходит ученый, решая какую-либо научную задачу, проблему.
К основным методам науки относятся следующие:
Моделирование – метод, при котором создается некий образ объекта, модель, с помощью которой ученые получают необходимые сведения об объекте. Так, например, при установлении структуры молекулы ДНК
Джеймс Уотсон и Френсис Крик создали из пластмассовых элементов модель – двойную спираль ДНК, отвечающую данным рентгенологических и биохимических исследований.
Наблюдение – метод, с помощью которого исследователь собирает информацию об объекте. Наблюдать можно визуально. Можно наблюдать с помощью приборов за изменениями, происходящими в живых объектах.
Эксперимент (опыт) – метод, с помощью которого проверяют результаты наблюдений, выдвинутые предположения – гипотезы. Эсперимент – это всегда получение новых знаний с помощью поставленного опыта.
Проблема – вопрос, задача, требующие решения. Решение проблемы ведет к получению нового знания. Научная проблема всегда скрывает какое-то противоречие между известным и неизвестным. Решение проблемы
требует от ученого сбора фактов, их анализа, систематизации.
Гипотеза – предположение, предварительное решение поставленной проблемы. Выдвигая гипотезы, исследователь ищет взаимосвязи между фактами, явлениями, процессами. Именно поэтому гипотеза чаще всего
имеет форму предположения: «если … тогда». Гипотеза проверяется экспериментально.
Теория – это обобщение основных идей в какой-либо научной области знания. Со временем теории дополняются новыми данными, развиваются. Некоторые теории могут опровергаться новыми фактами. Верные
научные теории подтверждаются практикой.
Частными научными методами в биологии являются:
Генеалогический метод – применяется при составлении родословных людей, выявлении характера наследования некоторых признаков.
Исторический метод – установление взаимосвязей между фактами, процессами, явлениями, происходившими на протяжении исторически длительного времени (несколько миллиардов лет). Эволюционное учение
развивалось в значительной мере благодаря этому методу.
Палеонтологический метод – метод, позволяющий выяснить родство между древними организмами, останки которых находятся в земной коре, в разных геологических слоях.
Центрифугирование – разделение смесей на составные части под действием центробежной силы. Применяется при разделении органоидов клетки, легких и тяжелых фракций (составляющих) органических веществ и т.
д.
Цитологический, или цитогенетический, – исследование строения клетки, ее структур с помощью различных микроскопов.
Биохимический – исследование химических процессов, происходящих в организме.
2.
2) Сущность жизни, уровни организации живого. Фундаментальные свойства живого, клетка - элементарная биологическая единицаВсеобщим методологическим подходом к пониманию сущности жизни в настоящее время является понимание жизни в качестве процесса, конечным результатом которого является
самообновление, проявляющееся в самовоспроизведении. Все живое происходит только из живого, а всякая организация, присущая живому, возникает только из другой подобной
организации. Следовательно, сущность жизни заключается в ее самовоспроизведении, в основе которого лежит координация физических и химических явлений и которое
обеспечивается передачей генетической информации от поколений к поколениям. Именно эта информация обеспечивает самовоспроизведение и саморегуляцию живых существ.
Поэтому жизнь -- это качественно особая форма существования материи, связанная с воспроизведением. Явления жизни представляют собой форму движения материи, высшей по
сравнению с физической и химической формами его существования.
Живое построено из тех же химических элементов, что и неживое (кислород, водород, углерод, азот, сера, фосфор, натрий, калий, кальций и другие элементы). В клетках они
находятся в виде органических соединений. Однако организация и форма существования живого имеет специфические особенности, отличающие живое от предметов неживой
природы.
Уровни организации живого На всех уровнях проявляются все свойства, характерные для живого. Каждый из этих уровней характеризуется особенностями, присущими другим
уровням, но каждому уровню присущи собственные специфические особенности.
1. Молекулярный уровень. Элементарными единицами этого уровня организации жизни являются химические вещества; нуклеиновые кислоты, белки, углеводы, липиды и др. На
этом уровне в основном проявляются такие важнейшие процессы жизнедеятельности, как передача наследственной информации, биосинтез, превращение энергии и др. Основная
стратегия жизни на молекулярном уровне - способность создавать живое вещество и кодировать информацию, приобретенную в меняющихся условиях среды.
2.Клеточный уровень. На клеточном уровне организации структурными элементами выступают различные органеллы. Способность к воспроизведению себе подобных, включение
различных химических элементов Земли в состав клетки, регуляция химических реакций, запасание и потребление энергии - основные процессы этого уровня. Стратегия жизни на
клеточном уровне - вовлечение химических элементов Земли и энергии Солнца в живые системы.
3. Тканевый уровень. Ткань совокупность клеточных элементов различных клеточных типов и межклеточного вещества, специализированная на выполнении специфических функций.
4. Органный уровень. Орган - совокупность тканей, которые связаны выполнением общих функций и занимают определенное место в многоклеточном организме.
5. Организменный уровень организации присущ одноклеточным и многоклеточным биосистемам (растениям, грибам, животным, в том числе человеку и разнообразным
микроорганизмам). У живых организмов проявляются такие свойства, как питание, дыхание, выделение, раздражимость, рост и развитие, размножение, поведение,
продолжительность жизни, взаимоотношения с окружающей средой. Все перечисленные процессы в совокупности характеризуют организм как целостную саморегулирующуюся
биосистему. Основная стратегия жизни на этом уровне - ориентация организма (особи) на выживание в постоянно меняющихся условиях среды.
6. Популяционно-видовой уровень организации характеризуется объединением родственных особей в популяции, а популяций - в виды, что приводит к возникновению новых
свойств системы. Основные свойства этого уровня: рождаемость, смертность, выживание, структура (половая, возрастная, экологическая), плотность, численность, функционирование
в природе. Основная стратегия популяционно-видового уровня проявляется в более полном использовании возможностей среды обитания, в стремлении к возможно более
длительному существованию, в сохранении свойств вида и самостоятельном развитии.
7. Биогеоценотический (экосистемный) уровень организации основными структурными элементами являются популяции разных видов. Данный уровень характеризуется множеством
свойств. К ним относятся: структура экосистемы, видовой и количественный состав ее населения, типы биотических связей, пищевые цепи и сети", трофические уровни,
продуктивность, энергетика, устойчивость и др. Организующие свойства проявляются в круговороте веществ и потоке энергии, саморегулировании и устойчивости, автономности,
открытости системы, сезонных изменениях. Основная стратегия этого уровня - активное использование всего многообразия окружающей среды и создание благоприятных условий
развития и процветания жизни во всем ее многообразии.
8. Биосферный уровень. Самый высокий уровень организации жизни. Основными структурными единицами этого уровня являются биогеоценозы (экосистемы) и окружающая их
среда, т. е. географическая оболочка Земли (атмосфера, гидросфера, почва, солнечная радиация и др.) и антропогенное воздействие. Для этого уровня организации характерны:
активное взаимодействие живого и неживого вещества планеты; биологический круговорот веществ и потоки энергии с входящими в него геохимическими циклами; хозяйственная и
этнокультурная деятельность человека. Основная стратегия, жизни на биосферном уровне - стремление обеспечить динамичную устойчивость биосферы как самой большой
экосистемы нашей планеты.
3.
2) Сущность жизни, уровни организации живого. Фундаментальные свойства живого, клетка - элементарная биологическая единицаСвойства живого Для живого характерен ряд свойств, которые в совокупности «делают» живое живым.
1.Химический состав. Живые существа состоят из тех же химических элементов, что и неживые, но в организмах есть молекулы веществ, характерных только для живого
(нуклеиновые кислоты, белки, липиды).
2. Дискретность и целостность. Любая биологическая система (клетка, организм, вид и т.д.) состоит из отдельных частей, т.е. дискретна. Взаимодействие этих частей
образует целостную систему (например, в состав организма входят отдельные органы, связанные структурно и функционально в единое целое).
3. Структурная организация. Живые системы способны создавать порядок из хаотичного движения молекул, образуя определенные структуры. Для живого характерна
упорядоченность в пространстве и в ремени. Это комплекс сложных саморегулирующихся процессов обмена веществ, протекающих в строго определенном порядке,
направленном на поддержание постоянства внутренней среды — гомеостаза.
4. Обмен веществ и энергии. Живые организмы—открытые системы, совершающие постоянный обмен веществом и энергией с окружающей средой. При изменении
условий среды происходит саморегуляция жизненных процессов по принципу обратной связи, направленная на восстановление постоянства внутренней среды —
гомеостаза. Например, продукты жизнедеятельности могут оказывать сильное и строго специфическое тормозящее воздействие на те ферменты, которые составили
начальное звено в длинной цепи реакций.
5. Самовоспроизведение. Самообновление. Время существования любой биологической системы ограничено. Для поддержания жизни происходит процесс
самовоспроизведения, связанный с образованием новых молекул и структур, несущих генетическую информацию, находящуюся в молекулах ДНК.
6.Наследственность. Молекула ДНК способна хранить, передавать наследственную информацию, благодаря матричному принципу репликации, обеспечивая
материальную преемственность между поколениями.
7. Изменчивость. При передаче наследственной информации иногда возникают различные отклонения, приводящие к изменению признаков и свойству потомков. Если
эти изменения благоприятствуют жизни, они могут закрепиться отбором.
8. Рост и развитие. Организмы наследуют определенную генетическую информацию о возможности развития тех или иных признаков. Реализация информации
происходит во время индивидуального развития—онтогенеза. На определенном этапе онтогенеза осуществляется рост организма, связанный с репродукцией молекул,
клеток и других биологических структур. Рост сопровождается развитием.
9. Раздражимость и движение. Все живое избирательно реагирует на внешние воздействия специфическими реакциями благодаря свойству раздражимости.
Организмы отвечают на воздействие движением. Проявление формы движения зависит от структуры организма.
Клетка – элементарная биологическая единица Клетка представляет собой обособленную, наименьшую по размерам структуру, которой присуща вся совокупность
свойств жизни и которая может в подходящих условиях окружающей среды поддерживать эти свойства в самой себе, а также передавать их в ряду поколений. Клетка,
таким образом, несет полную характеристику жизни. Вне клетки не существует настоящей жизнедеятельности. Поэтому в природе планеты ей принадлежит роль
элементарной структурной, функциональной и генетической единицы.Это означает, что клетка составляет основу строения, жизнедеятельности и развития всех живых
форм — одноклеточных, многоклеточных и даже неклеточных. Благодаря заложенным в ней механизмам клетка обеспечивает обмен веществ, использование
биологической информации, размножение, свойства наследственности и изменчивости, обусловливая тем самым присущие органическому миру качества единства и
разнообразия.
Занимая в мире живых существ положение элементарной единицы, клетка отличается сложным строением. При этом определенные черты обнаруживаются во всех без
исключения клетках, характеризуя наиболее важные стороны клеточной организации как таковой.
4.
3) Клеточная теория: основные этапы развитияКл теория сформулирована немецким исследователем, зоологом Т. Шванном (1839). Поскольку при создании этой теории Шванн широко пользовался работами ботаника М Шлейдена, последнего
по праву считают соавтором кл теории. Исходя из предположения о схожести (гомологичности) растительных и животных клеток, доказываемой одинаковым механизмом их возникновения, Шванн
обобщил многочисленные данные в виде теории, согласно которой клетки являются структурной и функциональной основой живых существ.
В конце 19 века немецкий патолог Р. Вирхов на основе новых фактов пересмотрел клеточную теорию. Ему принадлежит вывод о том, что клетка может возникнуть лишь из предшествующей клетки.
Им создана вызвавшая критику концепция «клеточного государства», согласно которой клеточный организм состоит из относительно самостоятельных единиц (клеток), поставленных в своей
жизнедеятельности в тесную взаимосвязь друг от друга.
Клеточная теория в современном виде включает четыре главных положения:
Клетка – основная структурно-функциональная и генетическая единица живых организмов, наименьшая единица живого
Клетки одноклеточных и многоклеточных организмов сходны по строению, химическому составу и важнейшим проявлениям процессов жизнедеятельности
Каждая новая клетка образуется в результате деления исходной (материнской) клетки
Клетки многоклеточных организмов специализированны: они выполняют разные функции и образуют ткани
Основные этапы развития клеточной теории
История изучения клетки.
История изучения клетки неразрывно связана с развитием микроскопической техники и методов исследования.
Галилео Галилей в 1609 – 1610 гг. сконструировал первый микроскоп.
Роберт Гук в 1665 г. Впервые описал строение коры пробкового дуба и стебля растений, ввел в науку термин «клетка» .
М. Мальпиги и Н. Грю описали микроструктуру некоторых органов растений.
Н. Грю ввел в науку термин «ткань» для обозначения совокупности однородных клеток.
Антоний Ван Левенгук (1632 – 1723) – голландский купец, подарил науке величайшие открытия.
В 1693 г. Петр I в Дельфе
В 1725 году организована Петербургская академия наук.
В 1831 г. Р. Броун открыл в клеточном соке ядро
Русский ученый П. Ф. Горянинов
Клеточная теория
В 1839 г. Теодор Шванн
М. Шлейдена в 1838 г.
Немецкий ученый Рудольф Вихров в 1858 г.
5.
3) Клеточная теория: основные этапы развития6.
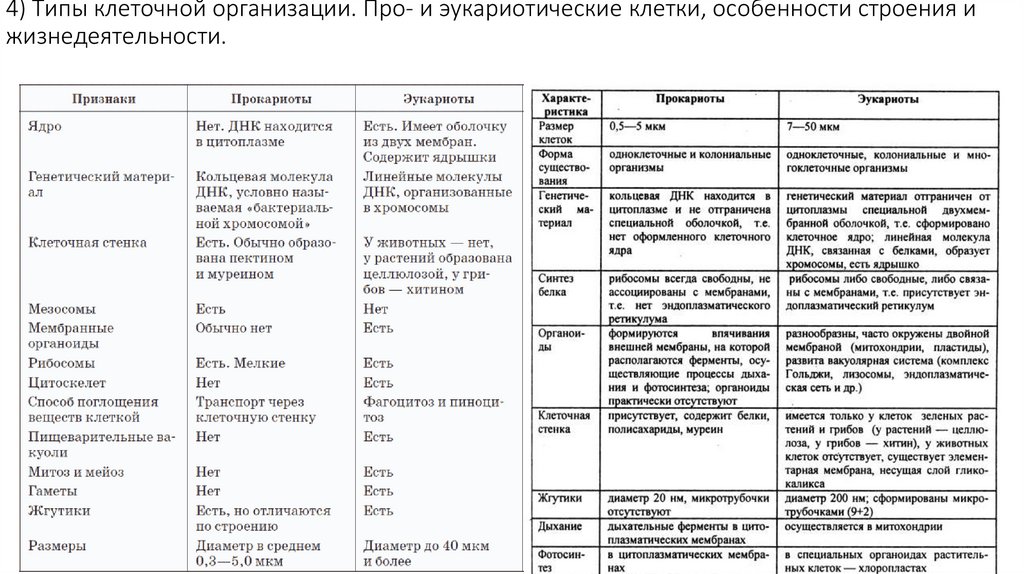
4) Типы клеточной организации. Про- и эукариотические клетки, особенностистроения и жизнедеятельности.
В настоящее время установлены 2 вида клеточной
организации: прокариоты и эукариоты. Они
существенно отличаются друг от друга. К
прокариотическим организмам относят бактерии, СЗО
и архебактерии (бактерии, выживающие в крайне
тяжелых условиях). 0,5-0,3 мкм – размер. Генетическая
информация в одной хромосоме – двуцепочечная
ДНК, кольцевой формы. Состав хромосом: нет
гистоновых белков. Хромосома «голая».
Распространены повсеместно. Короткая регенерация,
короткое время размножения, быстрый рост, большое
биохимическое разнообразие. Эукариотические
клетки имеют сильно разветвленные внутриклеточные
мембраны. Ядра содержат ядрышки и хромосомы
(количество хромосом больше 2). В состав хромосом
также входят белки-гистоны, РНК и др.
эукариотические клетки способны существовать
вместе с другими эукариотическими клетками и
являются субъединицами многоклеточного организма.
Прокариоты и эукариоты относятся к кислороду поразному. Большинство прокариот – облигатные
анаэробы, реже – факультативные анаэробы, есть и
облигатные аэробы. Среди эукариот – единообразие –
облигатные аэробы.
Прокариотические клетки:
- Прокариотические клетки являются самыми простыми и наиболее
древними формами жизни.
- У них отсутствует ядро, а генетический материал находится в
цитоплазме в виде кольцевой ДНК.
- Прокариоты обладают меньшим размером и менее сложными
мембранными органеллами, такими как митохондрии или
хлоропласты.
- Большинство прокариотических клеток имеют клеточную стенку
из пептидогликана.
- Прокариоты включают бактерии и археи.
Эукариотические клетки:
- Эукариотические клетки более сложные и содержат ядро,
отграниченное мембраной, где находится генетическая
информация в виде хромосом.
- Они также имеют множество мембранно-ограниченных
внутриклеточных органелл, таких как митохондрии, клеточная
мембрана, эндоплазматическая сеть, Гольджи аппарат и другие.
- Эукариотические клетки могут быть одноклеточными (например,
протисты, такие как амеба) или многоцеллюлярными (как растения,
животные и грибы).
- У эукариотических клеток могут быть специализированные
структуры, такие как центриоли, смычки, рибосомы и другие,
которые выполняют различные функции.
- Эукариотические клетки могут обмениваться информацией между
собой и с другими клетками, также они способны к более сложной
метаболической активности.
7.
4) Типы клеточной организации. Про- и эукариотические клетки, особенности строения ижизнедеятельности.
8.
5) Вирусы: строение, организация генетического материала,медицинское значение
Вирусы были обнаружены в 1892году русским учёным Д.И.Ивановским. В 1917году француз Ф.Д’Эрель открыл
бактериофаг-вирус,поражающий бактерии.
Особенности вирусов заключаются в их незначительных размерах,отсутствии клеточного строения,обмена веществ и
энергии.Но самым характерным критерием является наличие у вирусов только одной нуклеиновой кислоты – РНК или
ДНК(у остальных организмов всегда имеются и ДНК, и РНК).Вирусы самостоятельно не способны синтезировать
белки.Вирусы не растут. Простая вирусная частица состоит из образованной белками оболочки - капсида и нуклеиновой
кислоты. Некоторые более сложные вирусы(герпеса,гриппа) помимо белков капсида и нуклеиновой кислоты могут
содержать липопротеиновую мембрану,углеводы и ряд ферментов. Различают два вида вирусов РНК-содержащие и ДНКсодержащие, но независимо от того,какая из нуклеиновых кислот содержится в вирусе,она выполняет функции носителя
наследственной информации. Объём генетической информации вируса может быть очень мал,например у самых малых
вирусов он состоит из 3500нуклеотидов.
Бактериофаг состоит из головки(белковая оболочка и заключённая в ней ДНК или РНК)и отростка.
Биологическое значение вирусов в первую очередь связывается с их патогенным действием, т.е. способностью вызывать
различные заболевания у человека, животных и растений. Сегодня специалисты насчитывают не менее 500 различных
болезней человека, в которых в той или иной мере повинен вирус. Среди них такое заболевание как бешенство, оспа,
желтая лихорадка, энцефалиты, гепатиты, злокачественные опухоли, СПИД, корь и др.Помимо того, вирусы способны
оказывать влияние на генетический аппарат клетки, вызывая генные мутации.
Строение вирусов включает:
1. Генетический материал: Вирусы могут иметь ДНК или РНК в качестве своего генетического материала. У некоторых
вирусов геном состоит из двухцепочечной двухкаталной ДНК или мРНК, тогда как у других генетический материал может
быть одноцепочечным или разная структура РНК.
2. Капсид: Это белковая оболочка, которая окружает генетический материал вируса. Капсид обеспечивает защиту
генетического материала и его передачу внутрь клетки-хозяина.
3. Оболочка: Некоторые вирусы имеют внешнюю оболочку вокруг капсида, состоящую из липидного материала, взятого
из клетки-хозяина. Эта оболочка помогает вирусу обойти иммунную систему хозяина.
9.
6) Клетка как открытая система: Потоки вещества,энергии и информации в клетке
Клетка — открытая система, поскольку ее существование возможно только в условиях постоянного обмена веществом и энергией с окружающей
средой. Жизнедеятельность клетки обеспечивается процессами, образующими три потока: информации, энергии веществ.
Благодаря наличию потока информации клетка приобретает структуру, отвечающую критериям живого, поддерживает ее во времени, передает в
ряду поколений. В этом потоке участвуют ядро, макро молекулы, переносящие информацию в цитоплазму (мРНК), цитоплазматический аппарат
транскрипции (рибосомы и полисомы, тРНК, ферменты активации аминокислот). Позже полипептиды, синтезированные на полисомах,
приобретают третичную и четвертичную структуру, и используется в качестве катализаторов или структурных белков. Также функционируют
геномы митохондрий, а в зеленых растениях — и хлоропластов.
Поток энергии обеспечивается механизмами энергообеспечения — брожением, фото — или хемосинтезом, дыханием. Дыхательный обмен
включает реакции расщепления низкокалорийного органического «топлива» в виде глюкозы, жирных кислот, аминокислот, использование
выделяемой энергии для образования высококалорийного клеточного «топлива» в виде аденозинтрифосфата (АТФ). Энергия АТФ в
разнообразных процессах преобразуется в тот или иной вид работы — химическую (синтезы), осмотическую (поддержание перепадов
концентрации веществ), электрическую, механическую, регуляторную. Анаэробный гликолиз— процесс бескилородного расщепления глюкозы.
Фотосинтез— механизм преобразования энергии солнечного света в энергию химических связей органических веществ.
Дыхательный обмен одновременно составляет ведущее звено потока веществ, объединяющего метаболические пути расщепления и образования
углеводов, белков, жиров, нуклеиновых кислот.
Биологически активные вещества — гормоны, ферменты, адреналин, серотонин и т. д.
Потоки вещества:
В клетке происходит непрерывный поток веществ, необходимых для поддержания жизнедеятельности. Нутриенты, такие как глюкоза,
аминокислоты и липиды, входят в клетку через мембрану, а отходы и продукты метаболизма выводятся из клетки.
Потоки энергии:
Энергия в клетке обеспечивается через метаболические пути, такие как гликолиз, цикл Кребса и окислительное фосфорилирование. Вещества,
полученные из пищи, разлагаются и превращаются в энергию в форме АТФ (аденозинтрифосфата), которая затем используется для выполнения
различных клеточных функций, таких как синтез белка, перенос веществ и двигательная активность.
Потоки информации:
Клетка также обменивается информацией с окружающей средой. Гены, находящиеся в ДНК, содержащейся в ядре клетки, кодируют информацию,
необходимую для синтеза белка. Эта информация передается через процесс транскрипции (перенос генетической информации с ДНК на РНК) и
трансляции (синтез белка по информации, содержащейся в РНК).
10.
7) Элементарный химический состав живого. Вода и низкомолекулярные соединения клетки.Клеточное вещество является сложным полифазным коллоидом, т. е. представляет собой систему из 2х несмешивающихся фаз.
В элементарном составе клетки насчитывается более 80 элементов. Их делят на 3 группы:
•Макроэлементы – содержание которых в клетках составляет до 10-3 % (кислород, углерод, водород, азот, фосфор, сера, кальций, натрий и магний); на их долю приходится свыше 99% массы клеток.
•Микроэлементы – содержание которых колеблется от 10-3% до 10-6% (железо, марганец, медь, цинк, кобальт, никель, иод, фтор); на их долю приходится менее 1 % массы клеток.
•Ультрамикроэлементы – содержание которых составляет менее 10-6% (золото, серебро, уран, цезий, бром, ванадий, селен); на их долю приходится менее 0,01% массы клетки.
Все перечисленные элементы входят в состав неорганических и органических веществ или содержатся в виде ионов. Важными являются катионы калия, натрия, кальция, магния; анионы дигидрофосфата, хлора и гидрокарбоната.
Неорганическими соединениями клетки являются вода и минеральные соли.
Вода составляет около 70% массы клетки. У отдельных организмов (медуз) её содержание составляет 95%. В теле человека вода составляет 60%, из которой 40 % приходится на внутриклеточную воду.
Функции воды:
1.связанная вода (4-5%) образует водные (сольватные) оболочки вокруг молекул белков, препятствуя склеиванию их друг с другом;
2.свободная вода является универсальным растворителем и способствует трунспорту растворенных в ней веществ;
3.вода принимает непосредственное участие в реакциях гидролиза;
4.вода регулирует тепловой режим и осмотическое давление в клетках.
Минеральные соли и хим элементы в определенных концентрациях необходимы для нормальной жизнедеятельности клеток. Например, азот и сера входят в состав молекул белков, фосфор – в ДНК, РНК и АТФ, магний – во многие ферменты и
хлорофилл, железо – в гемоглобин, цинк – в гормон поджелудочной железы (инсулин), иод – в гормоны щитовидной железы и т.д. Нерастворимые соли кальция и фосфора обеспечивают прочность костной ткани, катионы натрия, калия и
кальция – раздражимость клеток. Ионы кальция принимают участие в свертывании крови.
Органические низкомолекулярные соединения – это аминокислоты, липиды, органические кислоты, витамины, коферменты (производные витаминов, обусловливающие активность ферментов) и другие.
Органические кислоты — органические вещества, проявляющие кислотные свойства. К ним относятся карбоновые кислоты, содержащие карбоксильную группу -COOH, сульфоновые кислоты, содержащие сульфогруппу -SO3H и некоторые
другие. Самыми известными органическими кислотами являются уксусная, муравьиная, лимонная и молочная.
Витами́ ны (от лат. vita -«жизнь») — группа низкомолекулярных органических соединений относительно простого строения и разнообразной химической природы. Это сборная по химической природе группа органических веществ,
объединённая по признаку абсолютной необходимости их для гетеротрофного организма в качестве составной части пищи. Витамины содержатся в пище в очень малых количествах, и поэтому относятся к микронутриентам. Витамины участвуют
во множестве биохимических реакций, выполняя каталитическую функцию в составе активных центров большого количества разнообразных ферментов либо выступая информационными регуляторными посредниками, выполняя сигнальные
функции экзогенных прогормонов и гормонов.
Витамины не являются для организма поставщиком энергии и не имеют существенного пластического значения. Однако витаминам отводится важнейшая роль в обмене веществ.
Концентрация витаминов в тканях и суточная потребность в них невелики, но при недостаточном поступлении витаминов в организм наступают характерные и опасные патологические изменения.
Большинство витаминов не синтезируются в организме человека. Поэтому они должны регулярно и в достаточном количестве поступать в организм с пищей или в виде витаминно-минеральных комплексов и пищевых добавок. Исключения
составляют витамин К, достаточное количество которого в норме синтезируется в толстом кишечнике человека за счёт деятельности бактерий, и витамин В3, синтезируемый бактериями кишечника из аминокислоты триптофана.
С нарушением поступления витаминов в организм связаны 3 принципиальных патологических состояния: недостаток витамина — гиповитаминоз, отсутствие витамина — авитаминоз, и избыток витамина — гипервитаминоз.
Известно около полутора десятков витаминов. Исходя из растворимости, витамины делят на жирорастворимые — A, D, E, F, K и водорастворимые — все остальные(B, C и др.). Жирорастворимые витамины накапливаются в организме, причём их
депо являются жировая ткань и печень. Водорастворимые витамины в существенных количествах не депонируются (не накапливаются), и при избытке выводятся с водой. Это объясняет то, что гиповитаминозы довольно часто встречаются
относительно водорастворимых витаминов, а гипервитаминозы — чаще наблюдаются относительно жирорастворимых витаминов.
Витамины отличаются от других органических пищевых веществ тем, что не включаются в структуру тканей и не используются организмом в качестве источника энергии (не обладают калорийностью).
11.
8) Строение и биологические функции белков клетки.Белки (протеины) составляют 50% от сухой массы живых организмов.
Белки состоят из аминокислот. У каждой аминокислоты есть аминогруппа и кислотная (карбоксильная) группа, при взаимодействии которых
получается пептидная связь, поэтому белки еще называют полипептидами.
Структуры белка
Первичная – цепочка из аминокислот, связанных пептидной связью (сильной, ковалентной). Чередуя 20 аминокислот в разном порядке, можно
получать миллионы разных белков. Если поменять в цепочке хотя бы одну аминокислоту, строение и функции белка изменятся, поэтому
первичная структура считается самой главной в белке.
Вторичная – спираль. Удерживается водородными связями (слабыми).
Третичная – глобула (шарик). Четыре типа связей: дисульфидная (серный мостик) сильная, остальные три (ионные, гидрофобные, водородные) –
слабые. Форма глобулы у каждого белка своя, от нее зависят функции. При денатурации форма глобулы меняется, и это сказывается на работе
белка.
Четвертичная – имеется не у всех белков. Состоит из нескольких глобул, соединенных между собой теми же связями, что и в третичной структуре.
(Например, гемоглобин.)
Денатурация
Это изменение формы глобулы белка, вызванное внешними воздействиями (температура, кислотность, соленость, присоединение других веществ
и т.п.)
•Если воздействия на белок слабые (изменение температуры на 1°), то происходит обратимая денатурация.
•Если воздействие сильное (100°), то денатурация необратимая. При этом разрушаются все структуры, кроме первичной.
Функции белков
Их очень много, например:
•Ферментативная (каталитическая) – белки-ферменты ускоряют химические реакции.
•Строительная (структурная) – клетка, если не считать воду, состоит в основном из белков.
•Защитная – антитела борются с возбудителями болезней (иммунитет).
12.
9) Строение и биологические функции липидов клеткиПод термином липиды (греч. lipos - жир) объединяют жиры и жироподобные вещества.
Липиды - органические соединения с различной структурой, но общими свойствами. Они нерастворимы в воде, но хорошо растворяются в органических растворителях: эфире, бензине, хлороформе и др. Липиды очень широко представлены в
живой природе и играют чрезвычайно важную роль в клетке и организме. Они содержатся в любых клетках. Содержание жира в них обычно невелико и составляет 5 - 15% от сухой массы. Существуют, однако, клетки, содержание жира в
которых достигает почти 90% от сухой массы. Эти буквально набитые жиром клетки имеются в жировой ткани.
В химическом отношении большинство липидов представляет собой сложные эфиры высших карбоновых кислот и ряда спиртов. Наиболее известны среди них жиры. Каждая молекула жира образована молекулой трехатомного спирта
глицерола и присоединенными к ней эфирными связями трех молекул высших карбоновых кислот. Согласно принятой номенклатуре, жиры называют триацилглщеролами. Все жирные кислоты делятся на две группы: насыщенные, т. е. не
содержащие двойных связей, и ненасыщенные, или непредельные, содержащие двойные связи. К насыщенным кислотам принадлежат пальмитиновая и стеариновая кислоты, а к ненасыщенным - олеиновая. Свойства жиров определяются
качественным составом жирных кислот и их количественным соотношением. Растительные жиры или масла богаты непредельными жирными кислотами, поэтому в подавляющем большинстве случаев они являются легкоплавкими - жидкими
при комнатной температуре. Например, в оливковом масле глицерин связан с остатками олеиновой кислоты. Животные жиры при комнатной температуре твердые, так как содержат главным образом насыщенные жирные кислоты. Например,
говяжье сало состоит из глицерина и пальмитиновой и стеариновой кислот (пальмитиновая кислота плавится при 43*С, а стеариновая при 60*С). Из формулы жира видно, что его молекула, с одной стороны, содержит остаток глицерина вещества, хорошо растворимого в воде, а с другой стороны - остатки жирных кислот, углеводородные цепочки которых практически нерастворимы в воде. При нанесении капли жира на поверхность воды в сторону воды обращается
глицериновая часть молекулы жира, а из воды "торчат" вверх цепочки жирных кислот.
Такая ориентация по отношению к воде играет очень важную роль. Тончайший слой этих веществ, входящих в состав клеточных мембран, препятствует смешиванию содержимого клетки или отдельных ее частей с окружающей средой.
Особенно биологически важными жироподобными веществами являются фосфолипиды. Это тоже сложные соединения глицерина и жирных кислот. От настоящих жиров они отличаются тем, что содержат остаток фосфорной кислоты.
Фосфолипиды имеют полярные головки и неполярные хвосты. Группы, образующие полярную головку, гидрофильны, а неполярные хвостовые группы гидрофобны. Они основные компоненты мембран клеток.
Большое значение для жизнедеятельности клетки и организма имеют и такие сложные соединения, как гликолипиды , состоящие из углеводов и липидов. Особенно их много в составе ткани мозга и нервных волокон. Здесь же надо назвать и
липопротеиды , представляющие собой комплексные соединения различных белков с жирами.
Функции липидов:
1) Энергетическая Основная функция триглицеридов. При расщеплении 1 г липидов выделяется 38,9 кДж. 2)Структурная
Фосфолипиды, гликолипиды и липопротеины принимают участие в образовании клеточных мембран.
3) Запасающая Жиры и масла являются резервным пищевым веществом у животных и растений. Важно для животных, впадающих в холодное время года в спячку или совершающих длительные переходы через местность, где нет источников
питания.
Масла семян растений необходимы для обеспечения энергией проростка.
4)Защитная Прослойки жира и жировые капсулы обеспечивают амортизацию внутренних органов.
Слои воска используются в качестве водоотталкивающего покрытия у растений и животных.
5) Теплоизоляционная
Подкожная жировая клетчатка препятствует оттоку тепла в окружающее пространство. Важно для водных млекопитающих или млекопитающих, обитающих в холодном климате.
6) Регуляторная Гиббереллины регулируют рост растений.
Половой гормон тестостерон отвечает за развитие мужских вторичных половых признаков.
Половой гормон эстроген отвечает за развитие женских вторичных половых признаков, регулирует менструальный цикл.
Минералокортикоиды (альдостерон и др.) контролируют водно-солевой обмен.
Глюкокортикоиды (кортизол и др.) принимают участие в регуляции углеводного и белкового обменов.
7)Источник метаболической воды При окислении 1 кг жира выделяется 1,1 кг воды. Важно для обитателей пустынь.
8) Каталитическая Жирорастворимые витамины A, D, E, K являются кофакторами ферментов, т.е. сами по себе эти витамины не обладают каталитической активностью, но без них ферменты не могут выполнять свои функции.
13.
10) Строение и биологические функции углеводов клетки.Углеводы, или сахара, являются важными молекулами в клетке. Они имеют разнообразные структуры, включая простые сахара (моносахариды,
такие как глюкоза и фруктоза), сложные сахара (олигосахариды, такие как мальтоза и целлюлоза) и полисахариды (такие как крахмал и гликоген).
Общая формула Сn (H2O)n. Следовательно, углеводы содержат в своем составе только три химических элемента.
Растворимые в воде углеводы.
Моносахариды: глюкоза – основной источник энергии для клеточного дыхания. Фруктоза – составная часть нектара цветов и фруктовых соков.
Рибоза и дезоксирибоза – структурные элементы нуклеотидов, являющихся мономерами РНК и ДНК.
Дисахариды: сахароза (глюкоза + фруктоза) – основной продукт фотосинтеза, транспортируемый в растениях. Лактоза (глюкоза + галактоза) –
входит в состав молока млекопитающих. Мальтоза (глюкоза + глюкоза) – источник энергии в прорастающих семенах.
Полисахариды: крахмал, гликоген, целлюлоза, хитин. Они не растворимы в воде..
Крахмал состоит из разветвленных спирализованных молекул, образующих запасные вещества в тканях растений.
Целлюлоза – полимер, образованный остатками глюкозы, состоящими из нескольких прямых параллельных цепей, соединенных водородными
связями. Такая структура препятствует проникновению воды и обеспечивает устойчивость целлюлозных оболочек растительных клеток.
Хитин состоит из аминопроизводных глюкозы. Основной структурный элемент покровов членистоногих и клеточных стенок грибов.
Гликоген – запасное вещество животной клетки. Гликоген еще более ветвистый, чем крахмал и хорошо растворимы в воде.
ФУНКЦИИ:
1)входит в состав жизненно-важных веществ
2)участвует в фиксации углерода
3)резервная
4)структурная
5)защитная
6)имеют энергетическую функцию 1г глюкозы=17кДж
14.
11) Строение и биологические функции нуклеиновых кислотНуклеиновые кислоты - природные высокомолекулярные органические соединения, обеспечивающие хранение и передачу наследственной(генетической) информации в живых организмах.
В природе существует два вида нуклеиновых кислот — дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Различие в названиях объясняется тем, что молекула ДНК содержит пяти-углеродный сахар дезоксирибозу, а молекула РНК—
рибозу. В настоящее время известно большое число разновидностей ДНК и РНК, отличающихся друг от друга по строению и значению в метаболизме.
ДНК находится преимущественно в хромосомах клеточного ядра (99% всей ДНК клетки), а также в митохондриях и хлоропластах. РНК входит в состав рибосом; молекулы РНК содержатся также в цитоплазме, матриксе пластид и митохондрий.
Нуклеотиды — структурные компоненты нуклеиновых кислот. Нуклеиновые кислоты представляют собой биополимеры, мономерами которых являются нуклеотиды. В состав каждого нуклеотида входит азотистое основание, пятиуглеродный
сахар (рибоза или дезоксирибоза) и остаток фосфорной кислоты. Существует пять основных азотистых оснований: аденин, гуанин, урацил, тимин и цитозин. Первые два являются пуриновыми; их молекулы состоят из двух колец, первое
содержит пять членов, второе — шесть. Следующие три являются пиримидинами и имеют одно пятичленное кольцо.
ДНК — представляет собой двухцепочечный биологический полимер, мономерами которого являются нуклеотиды, содержащие одно из азотистых оснований, дезоксирибозу и остаток фосфорной кислоты. Полинуклеотидные цепи молекулы
ДНК антипараллельны и соединены друг с другом водородными связями по принципу комплиментарности. Двойная спираль, открытая в 1953г. Уотсоном и Криком, содержит шаг размером 3,4 нм, включающем 10 пар комплементарно
связанных оснований.
ДНК состоит из Нуклеотидов: пуриновых оснований аденина(А) и гуанина (Г) и пиримидиновых оснований цитозина(Ц) и тимина(Т). РНК состоит из тех же оснований с различием лишь в то, что у РНК вместо тимина присутствует урацил(У).
(Тимин отличается от урацила наличием метильной группы (-СН3), которой нет в урациле).
Функцияу ДНК одна - хранение генетической информации
РНК - также полимер, мономерами которой являются нуклеотиды. РНК представляет собой однонитевую молекулу. Она построена таким же образом, как и одна из цепей ДНК. Нуклеотиды РНК очень близки, хотя и не тождественны,
нуклеотидам ДНК. Их тоже четыре и они состоят из азостистого основания, пентозы и фосфорной кислоты. Три азотистых основания совершенно такие же, как в ДНК: — Аденин, Гуанин и Цитозин. Однако вместо Тимина у ДНК, в РНК
присутствует близкий к нему по строению пиримидин - урацил. Различие между ДНК и РНК существует также в характере углевода: в нуклеотидах ДНК углевод — дезоксирибоза, у РНК — рибоза
В отличие от ДНК, содержание которой в клетках конкретных организмов относительно постоянно, содержание РНК сильно в них колеблется. Оно заметно повышено в клетках, в которых происходит синтез белка.
Функции РНК По выполнению функций выделя-ют несколько видов РНК.
Транспортная РНК(т-РНК). Молекулы т-РНК самые короткие: они состоят всего из 80—100 нуклео-тидов. Молекулярная масса таких частиц равна 25—30 тыс. Транспортная РНК в основном содержится в цитоплазме клетки. Функция состоит в
переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетки на долю т-РНК приходится около 10%.
Рибосомная РНК (р-РНК). Это самые крупные РНК в их молекулы входит 3—5 тыс. нуклеотидов, соответственно их молекулярная масса достигает 1,0—1, 5 млн. Рибосомная РНК составляет существенную часть структуры рибосомы. Из общего
содержания РНК в клетке на долю р-РНК приходится около 90%.
Информационная РНК (и-РНК), или матричная (м-РНК). Содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах. На долю и-РНК приходится примерно 0,5—1% от
общего содержания РНК клетки.
Все виды РНК синтезируются на ДНК, которая служит своего рода матрицей.
АТФ - аденозинтрифосфорная кислота. Аденозинмонофосфорная кислота (АМФ) входит в состав всех РНК; при присоединении еще двух молекул фосфорной кислоты (НзРО4) она превращается в АТФ и становится источником энергии, которая
запасается в двух последних остатках фосфатов: Каквовсякийнуклеотид, вАТФвходитостаток азотистого основания (аденин), пентоза (рибоза) и остатки фосфорной кислоты (у АТФ их три). Из состава АТФ под действием фермента АТФ-азы
отщепляются остатки фосфорной кислоты.При отщеплении одной молекулы фосфорной кислоты АТФ переходит в АДФ (аденозиндифосфорная кислота), а если отщепляются две молекулы фосфорной кислоты, АТФ переходит в АМФ
(аденозинмонофосфорная кислота). Реакции отщепления каждой молекулы фосфорной кислоты сопровождаются освобождением 419 кДж/моль. Для того чтобы подчеркнуть высокую энергетическую «стоимость» фосфорнокислородной связи
в АТФ, ее принято обозначать знаком ~ и называть макроэргической связью. В АТФ имеются две макроэргические связи.
Значение АТФ в жизни клетки велико, она играет центральную роль в клеточных превращениях энергии. В реакциях с участием АТФ она, как правило, теряет одну молекулу фосфорной кислоты и переходит в АДФ. А далее АДФ может
присоединить остаток фосфорной кислоты с поглощением 419 кДж/моль, восстановив запас энергии. Основной синтез АТФ происходит в митохондриях.
15.
12) Строение и биологические функции плазматической мембраны. Реснички и жгутики,микроворсинки
Биологическая мембрана имеет толщину 6-10 нм. Наружный и внутренний слои мембраны образованы молекулами белков, а средний – бимолекулярным слоем липидов. Липиды расположены
строго упорядоченно: гидрофильные концы молекул обращены к белковым слоям, а гидрофобные – друг к другу. Белковые молекулы могут располагаться периферически, полуинтегрально и
интегрально. Биол. мембрана обладает избирательной проницаемостью, которая обусловлена особенностями её строения. Большинство интегральных белков являются ферментами. Через
гидрофильные поры проходят водорастворимые вещества. Для избирательного поступления веществ необходим надмембранный комплекс гликокаликс (гликопротеины) - воспринимают
определенные химические вещества, окружающие клетку. Также обеспечивает иммунный ответ и другие реакции. Функции биологической мембраны – структурная, разделительная, транспортная,
рецепторная, ферментативная.
Органоиды движения клеток представлены жгутиками и ресничками. Они содержат по 20 микротрубочек, образующих 9 пар по периферии и 2 одиночные, расположенные в центре. Жгутики и
реснички покрыты элементарной мембраной. У основания органоидов движения расположены базальные тела, образующие микротрубочки. Реснички и жгутики служат для передвижения бактерий,
протистов, ресничных червей и сперматозоидов. Реснички мерцательного эпителия дыхательных путей освобождают их от попавших частиц.
Микроворсинка — вырост эукариотической (обычно животной) клетки, имеющий пальцевидную форму и содержащий внутри цитоскелет из актиновых микрофиламентов. Микроворсинки нередко
путают с ресничками, однако они резко отличаются по строению и функциям. Реснички имеют базальное тело и цитоскелет из микротрубочек, способны к быстрым движениям (кроме
видоизмененных неподвижных ресничек) и служат у крупных многоклеточных обычно для создания токов жидкости или восприятия раздражителей, а у одноклеточных и мелких многоклеточных
животных также для передвижения. Микроворсинки не содержат микротрубочек и способны лишь к медленным изгибаниям (в кишечнике) либо неподвижны.
За упорядочение актинового цитоскелета микроворсинок отвечают вспомогательные белки, взаимодействующие с актином — фимбрин, спектрин, виллин и др. Микроворсинки также содержат
цитоплазматический миозиннескольких разновидностей.
Микроворсинки кишечника (не путать с многоклеточными ворсинками) во много раз увеличивают площадь поверхности всасывания. Кроме того.у позвоночных на их плазмалемме закреплены
пищеварительные ферменты, обеспечивающие пристеночное пищеварение.
Микроворсинки внутреннего уха (стереоцилии) интересны тем, что образуют ряды с различной, но строго определенной в каждом ряду длиной. Вершины микроворсинок более короткого ряда
соединены с более длинными микроворсинками соседнего ряда с помощью белков - протокадгеринов. Их отсутствие или разрушение может приводить к глухоте, так как они необходимы для
открывания натриевых каналов на мембране волосковых клеток и, следовательно, для преобразования механической энергии звука в нервный импульс
Плазматическая мембрана - это тонкая двухслойная структура, образующая внешнюю границу живой клетки. Она состоит в основном из фосфолипидов, белков и углеводов.
Биологические функции плазматической мембраны включают:
1. Регуляция переноса веществ
2. Клеточное распознавание
3. Экскреция и эндоцитоз
4. Сигнальная передача
Ресничек и жгутиков это важные структуры, которые имеются у некоторых видов клеток. Они играют роль в передвижении и перемещении клеток. Реснички и жгутики содержат внутри себя
микротрубочки и микрофиламенты, которые обеспечивают их движение.
Микроворсинки, или микроворсинчатая (микроворсинковая) поверхность, представляет собой структуру, содержащую множество микроскопических выростов, называемых микроворсинками. Они
увеличивают поверхность клетки, что особенно важно для клеток, участвующих в поглощении питательных веществ из окружающей среды (например, некоторые клетки кишечника).
16.
13) Транспорт через плазматическую мембрану: активный и пассивный, их виды,экзоцитоз, эндоцитоз.
Транспорт через плазматическую мембрану представляет собой основные
способы передвижения веществ внутрь и из клетки. Он может быть активным
или пассивным, и включает в себя экзоцитоз и эндоцитоз.
1. Пассивный транспорт:
- Диффузия: Простая диффузия происходит без участия белков и основана на
случайном перемещении молекул через мембрану в обе стороны, пока не
достигнется равновесие. Облегченная диффузия, с другой стороны,
происходит через каналы или переносчики.
- Осмотический транспорт: Это процесс, при котором вода движется через
мембрану для выравнивания осмотического давления.
- Фильтрация: Транспорт через мембрану происходит под воздействием
давления.
2. Активный транспорт:
- Этот процесс требует затраты энергии и происходит против
концентрационного градиента. Примеры включают натриево-калиевую помпу,
которая поддерживает градиент ионообмена в клетке, а также перенос
веществ через мембрану.
3. Экзоцитоз:
- Это процесс, при котором клетка выделяет вещества, такие как белки, за
пределы мембраны путем слияния везикул, содержащих эти вещества, с
плазматической мембраной.
4. Эндоцитоз:
- Это процесс, при котором клетка захватывает вещества из внешней среды
путем образования везикул, которые затем перемещаются внутрь клетки. Три
основных типа эндоцитоза - фагоцитоз, пиноцитоз и рецепторномедиатизированный эндоцитоз - различаются по типу захватываемых частиц и
механизму, используемому клеткой.
17.
13) Транспорт через плазматическую мембрану: активный и пассивный, их виды, экзоцитоз,эндоцитоз.
-обеспечивают различные типы межклеточных контактов: простые щелевидные контакты, межклеточные «замки», десмосомы и плотные контакты в эпителиальных тканях, синапсы.
Наличие толстых клеточных стенок у грибов и растений затрудняет контакт между клетками. Поэтому клеточные стенки обычно пронизаны порами, через которые проходят тяжи цитоплазмы –
плазмодесмы. Через плазмодесмы эндоплазматическая сеть одной клетки переходит в эндоплазматическую сеть смежных клеток.
Транспорт через плазматическую мембрану: активный и пассивный, их виды, экзо- и эндоцитоз.
Важнейшее свойство мембран — избирательная проницаемость, т.е. мембраны хорошо проницаемы для одних веществ или молекул и плохо проницаемы (или совсем непроницаемы) для других.
Это свойство лежит в основе регуляторной функции мембран, обеспечивающей обмен веществ между клеткой и внешней средой. Процесс прохождения веществ через клеточную мембрану
называют транспортом веществ. Различают: 1) пассивный транспорт — процесс прохождения веществ, идущий без затрат энергии; 2) активный транспорт — процесс прохождения веществ, идущий с
затратами энергии.
При пассивном транспорте вещества перемещаются из области с более высокой концентрацией в область с более низкой, т.е. по градиенту концентрации. В любом растворе имеются молекулы
растворителя и растворенного вещества. Процесс перемещения молекул растворенного вещества называют диффузией, перемещения молекул растворителя — осмосом. Если молекула заряжена, то
на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Можно выделить следующие виды пассивного транспорта: 1) простая диффузия — транспорт веществ непосредственно через липидный бислой (кислород, углекислый газ); 2) диффузия через
мембранные каналы — транспорт через каналообразующие белки (Na+, K+, Ca2+, Cl-); 3) облегченная диффузия — транспорт веществ с помощью специальных транспортных белков, каждый из
которых отвечает за перемещение определенных молекул или групп родственных молекул (глюкоза, аминокислоты, нуклеотиды); 4) осмос — транспорт молекул воды (во всех биологических
системах растворителем является именно вода).
Необходимость активного транспорта возникает тогда, когда нужно обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется особыми
белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ. К активному транспорту относят: 1) Na+/К+-насос (натрий-калиевый насос), 2)
эндоцитоз, 3) экзоцитоз.
Активный транспорт веществ в клетке
Работа Na+/К+-насоса. Для нормального функционирования клетка должна поддерживать определенное соотношение ионов К+ и Na+ в цитоплазме и во внешней среде. Концентрация К+ внутри
клетки должна быть значительно выше, чем за ее пределами, а Na+ — наоборот. Следует отметить, что Na+ и К+ могут свободно диффундировать через мембранные поры. Na+/К+-насос
противодействует выравниванию концентраций этих ионов и активно перекачивает Na+ из клетки, а K+ в клетку. Na+/К+-насос представляет собой трансмембранный белок, способный к
конформационным изменениям, вследствие чего он может присоединять как K+, так и Na+. Цикл работы Na+/К+-насоса можно разделить на следующие фазы: 1) присоединение Na+ с внутренней
стороны мембраны, 2) фосфорилирование белка-насоса, 3) высвобождение Na+ во внеклеточном пространстве, 4) присоединение K+ с внешней стороны мембраны, 5) дефосфорилирование белканасоса, 6) высвобождение K+ во внутриклеточном пространстве. На работу натрий-калиевого насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки. За один цикл
работы насос выкачивает из клетки 3Na+ и закачивает 2К+.
Эндоцитоз — процесс поглощения клеткой крупных частиц и макромолекул. Различают два типа эндоцитоза: 1) фагоцитоз — захват и поглощение крупных частиц (клеток, частей клеток,
макромолекул) и 2) пиноцитоз — захват и поглощение жидкого материала (раствор, коллоидный раствор, суспензия). Явление фагоцитоза открыто И.И. Мечниковым в 1882 г. При эндоцитозе
плазматическая мембрана образует впячивание, края ее сливаются, и происходит отшнуровывание в цитоплазму структур, отграниченных от цитоплазмы одиночной мембраной. К фагоцитозу
способны многие простейшие, некоторые лейкоциты. Пиноцитоз наблюдается в эпителиальных клетках кишечника, в эндотелии кровеносных капилляров.
Экзоцитоз — процесс, обратный эндоцитозу: выведение различных веществ из клетки. При экзоцитозе мембрана пузырька сливается с наружной цитоплазматической мембраной, содержимое
везикулы выводится за пределы клетки, а ее мембрана включается в состав наружной цитоплазматической мембраны. Таким способом из клеток желез внутренней секреции выводятся гормоны, у
простейших — непереваренные остатки пищи.
18.
14) Контакты и межклеточные коммуникацииэукариотической клетки
Межклеточные контакты — соединения между клетками,
образованные при помощи белков. Межклеточные контакты
обеспечивают непосредственную связь между клетками или между
клеткой и матриксом. Кроме того, контакты служат для передачи
сигналов между клетками. В статье рассмотрена общая структура
межклеточного контакта, типы контактов и их функции, а также
основные белки, входящие в их состав.
Типы межклеточных контактов
Все межклеточные контакты можно разделить на четыре группы в
зависимости от выполняемой функции:
Прикрепляющие (заякоривающие) контакты. Данные соединения
обычно нужны для связывания клеток друг с другом (или с
внеклеточным матриксом);
Запирающие контакты необходимы для того, чтобы сделать
пространство между соседними клетками как можно менее
проницаемым;
Каналообразующие контакты (или соединения-каналы) позволяют
производить транспорт различных веществ между соседними
клетками;
Коммуникационные контакты представляют собой сложные
молекулярные комплексы, которые важны для передачи сигнала от
клетки к клетке.
19.
15) Клетка как целостная структура. Коллоидная система цитоплазмы (гиалоплазма).Клетка как целостная структура представляет собой сложную и организованную систему, где внутренние компоненты взаимодействуют, сотрудничают и осуществляют свои функции для обеспечения жизнедеятельности клетки.
Цитоплазма - это внутренняя жидкость клетки, которая окружает ядро и различные клеточные органеллы. Она имеет коллоидную структуру, что означает, что вещества в цитоплазме образуют дисперсную фазу мельчайших частиц (коллоиды) в жидкой среде. Гиалоплазма состоит из воды, белков, липидов,
углеводов, органических кислот, солей, и других веществ, которые поддерживают жизнедеятельность клетки.
Гиалоплазма также обеспечивает механическую поддержку для клеточных органелл и структур, что позволяет им выполнять свои функции. Она также служит для транспорта молекул внутри клетки, поддержания внутриклеточной среды и осуществления многих других процессов, необходимых для
жизнедеятельности клетки.
Взаимодействуя с окружающей средой и отвечая на регуляторные стимулы, клетка ведет себя как целостная структура. Об этом свидетельствует однотипность реакции разных видов клеток на действие раздражителей, вызывающих переход клетки в возбужденное состояние. Важная роль в функциональном
объединении структурных компонентов и компартментов клетки принадлежит свойствам живой протоплазмы (содержимое живой клетки, включающее ее ядро и цитоплазму).
Клетка любого организма представляет собой целостную живую систему. Она состоит из трех неразрывно связанных между собой частей: оболочки, цитоплазмы и ядра. Оболочка клетки осуществляет непосредственное взаимодействие с внешней средой и взаимодействие с соседними клетками (в
многоклеточных организмах).
Цитоплазма эукариотических клеток состоит из полужидкого содержимого и органелл. Основное полужидкое вещество цитоплазмы называют гиалоплазмой (от греч.хиалос — стекло) или матриксом. Гиалоплазма является важной частью клетки, ее внутренней средой. Она представляет собой сложную
коллоидную систему, которая образована белками, нуклеиновыми кислотами, углеводами, водой и другими веществами.
В гиалоплазме в растворенном состоянии содержится большое количество аминокислот, нуклеотидов и других строительных биополимеров, а также множество промежуточных продуктов, возникающих при синтезе и распаде макромолекул. Гиалоплазма содержит большое количество ионов неорганических
соединений, таких как Na+,K+, Ca2+, Mg2+, CI-, HCO3- ,HPO42- и др.
Несмотря на то, что в электронном микроскопе гиалоплазма выглядит гомогенным веществом, она не является однородной. Гиалоплазма состоит из двух фаз — жидкой и твердой. Жидкая фаза представляет собой коллоидный раствор различных белков и других веществ. В жидкой фазе содержится ситема
тонких белковых нитей (~2 нм толщиной), пересекающих цитоплазму в различных направлениях - микротрабекулярная система. Микротрабекулярная система связывает все внутриклеточные структуры клетки: мембранные органеллы, различные фибриллярные и трубчатые структуры. В местах пересечения или
соединения концов трабекул располагаются группы рибосом.
Вместе с трубчатыми (микротрубочки) и фибриллярными (микрофиламенты) органеллами микротрабекулярная система образует внутриклеточный цитоплазматический скелет (цитоскелет). Цитоскелет способствует упорядоченному размещению всех структурных компонентов клетки. Микротрубочки
обеспечивают определенную форму клетки, отвечают за направленное движение клеточных структур.
Микротрабекулярная система очень динамична. В определенных участках клетки ее нити могут легко распадаться на отдельные молекулы белка, которые переходят в раствор и изменяют физические свойства гиалоплазмы (изменяется агрегатное состояние отдельных участков цитоплазмы с жидкого на
гелеобразное, и, наоборот, изменяется ее вязкость и текучесть). Это происходит при изменении внешних и внутренних условий.
С распадом и сборкой микротрабекул связывают также движение цитоплазмы, которое имеет очень важное значение при перемещении веществ и структурных элементов клетки.
Микротрубочки представляют собой полые неразветвленные цилиндры. Внешний диаметр их не превышает 30 нм; толщина стенки микротрубочки составляет около 5 нм. В длину они могут достигать нескольких микрометров. Микротрубочки вместе с микротрабекулярной системой выполняет опорную
функцию в клетке, придавая ей определенную форму (при обработке клеток колхицином происходит разрушение микротрубочек; животные клетки, например, лишенные такой внутренней опоры, приобретают шаровидную форму). Они также образуют веретено деления и обеспечивают расхождение хромосом
к полюсам клетки. Перемещение хромосом (хроматид) осуществляется благодаря способно микротрубочек скользить одна по одной. Это скольжение обеспечивается благодаря энергии АТФ. Одни микротрубочки (хромосомные) прикрепляются к хромосомам и скользят по другим микротрубочкам (полюсным),
в результате чего хромосомы во время деления клетки растаскиваются к ее полюсам. Микроотрубочки отвечают также за перемещение клеточных органелл, которые с помощью микротрубочек направляются в нужные места подобно тому, как поезд следует в определенном направлении по рельсам.
Микрофиламенты представляют собой тонкие нити, встречающиеся во всей цитоплазме клеток. Особенно много их в поверхностном слое цитоплазмы, в ложноножках подвижных клеток, где они образуют густую сеть тонких нитей, которые пересекаются в разных направлениях. Пучки микрофиламентов
обнаруживаются в микроворсинках эпителия кишечника. Микрофиламенты образуются из белка актина, глобулярные молекулы которого полимеризуются в длинную тонкую фибриллу (толщиной 6 нм), состоящую из двух спирально закрученных вокруг друг друга нитей. В клетках содержание актина составляет
10—15 % от общего количества всех белков. В гиалоплазме обнаруживаются также нити другого важного белка — миозина, которые образуют вместе с актиновыми микрофиламентами комплекс, способный к сокращению при расщеплении АТФ. Взаимодействие актина и миозина лежит в основе сокращения
мышц. Микрофиламенты актина взаимодействуют с микротрубочками поверхностного слоя цитоплазмы и с плазмалеммой, обеспечивая двигательную активность гиалоплазмы. Считается также, что они участвуют в эндоцитозе, в образовании перетяжки при делении клеток животных и обеспечении
амебоидного движения.
Функции гиалоплазмы:
Является внутренней средой клетки, в которой происходит многие химические процессы.
Объединяет все клеточные структуры и обеспечивает химическое взаимодействие между ними.
Определяет местоположение органелл в клетке.
Обеспечивает внутриклеточный транспорт веществ и перемещение органелл (например, движение хлоропластов в растительных клетках.
Основное вместилище и зона перемещения молекул АТФ.
Определяет форму клетки.
20.
16) Ультраструктурная организация клеток человека.Ультраструктурная организация клеток человека относится к микроскопическому уровню организации живых систем. В каждой клетке человека есть различные органеллы, которые выполняют
специфические функции и имеют свою ультраструктуру.
Ядро обычно находится внутри мембраны, называемой ядерной оболочкой. Ядро имеет характерную ультраструктуру, включая ядрышко, хроматин и ядерную матрицу.
Митохондрии - это энергетические органеллы клеток, которые производят АТФ при окислении пищевых веществ. Они имеют двойную мембрану и содержат свою собственную ДНК.
Эндоплазматическая сеть (ЭПС) - это система мембранных структур, расположенных в цитоплазме клетки. Она включает гладкую ЭПС и шероховатую ЭПС. Гладкая ЭПС отвечает за синтез липидов и
метаболические функции, в то время как шероховатая ЭПС связана с синтезом белков и модификацией.
Гольджи - это стопка мембранных отделений, связанных с обработкой, сортировкой и упаковкой молекул для транспортировки внутри и вне клетки.
Лизосомы - это специализированные вакуоли, содержащие различные гидролазы, которые разрушают и перерабатывают внутриклеточные отходы и вещества.
Цитоплазма - это жидкая среда, заполняющая клетку. В ней находятся различные органеллы и молекулы, включая цитоскелет, который обеспечивает механическую поддержку и форму клетки.
Клетка- элементарная структурная и функциональная единица растительных и животных организмов, способная к самовоспроизведению и развитию. В традиционном изложении клетку
растительного или животного организма описывают как объект, отграниченный оболочкой, в котором выделяют ядро и цитоплазму.
Основные компоненты эукариотической клетки:
Наружная мембрана. Клетки многоклеточных организмов, как животных, так и рас¬тительных, обособлены от своего окружения оболочкой. Клеточная оболочка, или плазмалемма, животных клеток
образована мембра¬ной, покрытой снаружи слоем гликокаликса толщиной 10—20 нм. Плазмалемма выполняет отграничивающую, барьерную и транс¬портную функции. Благодаря свойству
избирательной проницаемо¬сти она регулирует химический состав внутренней среды клетки. В плазмалемме размещены молекулы рецепторов, которые избира¬тельно распознают определенные
биологически активные вещества (гормоны). Наличие в обо¬лочке рецепторов дает клеткам возможность воспринимать сигналы извне, чтобы целесообразно реагировать на изменения в
окружаю¬щей их среде или состоянии организма.
Цитоплазма. В цитоплазме различают основное вещество (матрикс, гиалоплазма), включения и органеллы. Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной
оболочкой и другими внутриклеточными структурами. Оно образует истинную внутрен¬нюю среду клетки, которая объединяет все внутриклеточные струк¬туры и обеспечивает взаимодействие их
друг с другом.
Органеллы— это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции. Выделяют органеллы общего значения и специальные. Последние в значительном
количестве присутствуют в клетках, специализированных к выполнению определенной функции, но в незначительном количестве могут встречаться и в других типах клеток (микроворсинки,
реснички, синаптические пузырьки).
К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и
полисомы, лизосомы, пероксисомы, микрофибриллы и микротру¬бочки, центриоли клеточного центра. В растительных клетках вы¬деляют также хлоропласты, в которых происходит фотосинтез.
Включениями называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из
клетки (гранулы секрета), балластными веществами (некоторые пигменты).
Ядро. Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Функциональ¬ная роль ядерной оболочки заключается в обособлении генетического материала (хромосом)
эукариотической клетки от цитоплазмы с присущими ей многочисленными метаболическими реакциями, а также регуляции двусторонних взаимодействий ядра и цитоплазмы.
Основу ядерного сока, или матрикса, составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования
генетического материала. В составе ядерного сока присутствуют нитчатые, или фибршиярные, белки, что указывает на выполнение ими опорной функции.
Ядрышко представляет собой структуру, в которой происходит образование и созревание рибосомалъных РНК (рРНК).
Хроматин является интерфазной формой существования хромо¬сом клетки.
21.
17) Структурная организация эукариотической клетки: 1-мембранные, 2-мембранные и немембранные органеллы клетки.Включения
Эукариотическая клетка имеет сложную внутреннюю структуру, которая включает в себя множество мембранных и немембранных органелл.
Мембранные органеллы клетки:
Ядро: содержит генетическую информацию клетки в виде ДНК и участвует в регуляции всех клеточных функций.
Митохондрии: осуществляют процесс аэробного дыхания, в результате которого происходит синтез АТФ - основного энергетического источника клетки.
Хлоропласты: присутствуют только в растительных клетках и занимаются процессом фотосинтеза, при котором световая энергия превращается в химическую, а углекислый газ превращается в органические вещества.
Эндоплазматическая сеть: состоит из гладкого и шероховатого эндоплазматического ретикулума. Гладкий эндоплазматический ретикулум синтезирует липиды, участвует в метаболизме и детоксикации, а шероховатый эндоплазматический
ретикулум содержит рибосомы и занимается синтезом белков.
Аппарат Гольджи: участвует в синтезе, модификации, сортировке и транспорте белков и липидов.
Лизосомы: содержат гидролитические ферменты и участвуют в переработке и утилизации органических молекул, а также в защите клетки от патогенов.
Немембранные органеллы клетки:
Цитоскелет: сеть полимерных белков, обеспечивающая поддержку и форму клетки, участвует в движении органелл, клеток и транспорте веществ.
Рибосомы: клеточные органеллы, где происходит синтез белков.
Включения: Гликоген: сложный углевод, запасающийся в клетках для обеспечения энергией. Крахмал: сложный углевод, запасающийся в клетках растений в виде гранул и служащий источником энергии. Жиры: энергетический запас, который
хранится в клетках в виде триглицеридов. Пигменты: химические соединения, которые дают цвет различным органеллам и клеткам. Кристаллы: минеральные соединения, которые накапливаются в клетках и выполняют различные функции,
например, хранение кальция или фосфатов.
Органоиды - это постоянные структуры клетки. По строению можно выделить немембранные органоиды (рибосомы, центриоли, микротрубочки, филаменты...) и мембранные, среди которых различают одномембранные (эндоплазматическая
сеть, комплекс Гольджи, лизосомы, пероксисомы...) и двухмембранные (митохондрии, пластиды).
Рибосомы. Диаметр рибосом около 20 нм. Состоят из двух субъединиц: малой и большой. В клетке эукариот два вида рибосом - 80 и 70 S (S - единицы седиментации) (табл. 1). В состав рибосом входят рРНК, рибосомальные белки, Синтез рРНК
и сборка субъединиц рибосом осуществляется в ядрышке. Функции рибосом - синтез белка.
Эндоплазматическая сеть (ЭПС) - система соединенных сплющенных цистерн. Выделяют две структурно взаимосвязанные разновидности ЭПС: гладкую и гранулярную (шероховатую). Гладкая ЭПС имеет трубчатое строение, ее мембраны более
контрастны (при электронной микроскопии), т. к. содержат рабочие ферменты. Функции гладкой ЭПС: компартментализация, первичный синтез липидов, синтез олигосахаридов, синтез предшественников стероидов, транспорт синтезированных
веществ, детоксикация. Гранулярная ЭПС представлена уплощенными цистернами с рибосомами. Мембрана менее контрастна в сравнении с гладкой ЭПС. Функции гранулярной ЭПС: компартментализация, синтез экспортного белка,
созревание белка, транспорт синтезированного белка и др.
КомплексГолъджи (КГ) состоит из дискоидных цистерн, собранных в стопки, и пузырьков по периферии. Пузырьки представляют собой формирующиеся первичные лизосомы или секреторные гранулы. При митозе КГ делится пополам, т. е.
имеет преемственное строение. Функции КГ: созревание, сортировка и упаковка экспортного белка; формирование первичных лизосом и секреторных гранул; синтез полисахаридов и липидов; детоксикация; компартментализация.
Лизосомы - округлые тельца с гомогенным содержимым, окруженные мембраной. Размер лизосом 0,2-1 мкм. Содержат около 60 гидролитических ферментов (20% в мембране, 80% внутри). Функции лизосом: ауто- и гетерофагия. Пероксисомы
- округлые тельца с кристаллоподобной сердцевиной. Содержат разнообразные ферменты, большинство из которых относятся к группе каталаз. Выделяют два вида пероксисом: 0,15 - 0,25 мкм - универсальные мелкие, локализуются во всех
клетках; 0,3 -1,5 мкм - крупные (в клетках печени, почек). Пероксисомы участвуют в метаболизме Н202, которая используется для последующего окисления разнообразных веществ.
Цитоскелет включает опорные органоиды - микротрубочки, микрофиламенты, промежуточные филаменты. Микротрубочки (рис. 5) - полый цилиндр диаметром 24 нм, стенка которого построена из спирально упакованных субъединиц белка
тубулина. Растут микротрубочки путем добавления с одного конца тубулиновых субъединиц. Кроме того, микротрубочки являются структурными компо-нентами центриолей, ресничек, жгутиков, базальных телец, митотического веретена.
Микрофиламенты - белковые нити диаметром 5 - 7 нм состоят из актина и миозина. Микрофиламенты обеспечивают двигательные функции
Включения -временные элемеаты, возникающие в клетке на определенной стадии ее жизнедеятельности в результате синтетической функции. Они либо используются, либо выводятся из клетки. Включениями являются также запасные
питательные вещества: в растительных клетках-крахмал, капельки жира, блки, эфирные масла, многие органические кислоты, соли органических и неорганических кислот; в животных клетках - гликоген (в клетках печени и мышцах), капли жира
(в подкожной клетчатке); Некоторые включения накапливаются в клетках как отбросы - в виде кристаллов, пигментов и др.
Промежуточные филаменты, их диаметр 8-10 нм, состоят из нитей собранных в пучки. Данные структуры тканеспецифичны, т. е. в каждой ткани свои промежуточные филаменты.
Митохондрии - двухмембранный органоид, присутствующий только у эукариот. Размер и форма митохондрий варьирует в широких пределах, их типичное строение см. рис. 6. Функции митохондрий: синтез АТФ (энергетическая),
22.
18) Одномембранные органеллы клетки: канальцевая и вакуолярная система клетки — ЭПС, Комплекс Гольджи,диктиосомы, лизосомы, микротельца, пероксисомы. Их строение и функции.
Одномембранные органеллы клетки представляют собой структуры, окруженные мембраной, состоящей из одного слоя липидов. В данной категории находятся:
Канальцевая и вакуолярная система клетки - эндоплазматическое ретикулум (ЭПС) и комплекс Гольджи. ЭПС представляет собой сеть мембран, расположенных в цитоплазме клетки. Он выполняет функции синтеза, обработки и транспорта
белков и липидов.
Комплекс Гольджи - это стопка плоских мембранных пузырьков, которые обрабатывают и модифицируют белки, а также участвуют в их сортировке и упаковке для дальнейшего транспорта.
Диктиосомы - это специализированные участки ЭПС и комплекса Гольджи, отвечающие за образование везикул, содержащих молекулы для транспорта.
Лизосомы - это пузырьки, содержащие гидролитические ферменты, которые разрушают и перерабатывают различные вещества, такие как бактерии, вирусы, старые клеточные органеллы и остатки пищи.
Микротельца - это пузырьки, образующиеся на поверхности плазматической мембраны и содержащие различные молекулы, такие как гормоны или нейромедиаторы. Они выполняют функцию транспорта и обмена веществ между клетками.
Пероксисомы - это органеллы, содержащие перекись водорода и ферменты, участвующие в окислительных реакциях. Они играют важную роль в обработке и расщеплении различных органических веществ, в том числе токсичных продуктов
обмена веществ.
Функции одномембранных органелл включают синтез, модификацию и транспорт белков и липидов, утилизацию различных молекул, регуляцию внутриклеточного обмена веществ, образование и транспорт молекул между клетками, а также
участие в образовании химических реакций с участием специфических ферментов.
Эндоплазматическая сеть (ЭПС), илиэндоплазматический ретикулум (ЭПР) – это система цистерн и трубочек, связанных между собой в единое внутриклеточное пространство, отграниченное от остальной части цитоплазмы замкнутой
внутриклеточной мембраной. ЭПС открыл К. Портер (1945 г.) с помощью электронного микроскопа.
Эндоплазматическая сеть (эндоплазматический ретикулум) имеется у всех эукариот и существует в виде двух типов: гранулярного (шероховатого) эндоплазматического ретикулума и агранулярного (гладкого) эндоплазматического ретикулума.
Мембраны эндоплазматического ретикулума тесно связаны с ядерной оболочкой, внутренние полости цистерн и трубочек эндоплазматического ретикулума связаны с перинуклеарным пространством.
Основной функцией ЭПС является биосинтез и транспортировка различных веществ. От цистерн и трубочек ЭПС отшнуровываются одномембранные мелкие пузырьки, дальнейшая судьба и функции которых зависят от их содержимого (см.
ниже).
Шероховатая ЭПС (гранулярный ЭПР) представлен системой плоских цистерн, на поверхности которых расположены рибосомы. Главной функцией гранулярного ЭПР является биосинтез, транспортировка и начальная модификация белков.
Дополнительной функцией является сборка компонентов биологических мембран.
Если на рибосомах гранулярного ЭПР идет синтез клеточных белков, то синтезированные полипептиды поступают в цитоплазматический матрикс или внедряются в мембраны. Если на рибосомах гранулярного ЭПР идет синтез экспортных
белков, то синтезированные полипептиды поступают в полость ретикулума через специальные поры – каналы, контролируемые специфическими белками–рецепторами. В полости гранулярного ЭПР полипептиды модифицируются:
отщепляется начало полипептидной цепи, образуются белковые гранулы, полипептиды образуют комплексы с другими веществами и т.д.
Гладкая ЭПС (агранулярный ЭПР) образована системой разветвленных трубочек. В полости агранулярного ЭПР происходит биосинтез липидов и полисахаридов. В агранулярном ретикулуме сократимых клеток происходит накопление ионов
кальция, а в агранулярном ретикулуме печени происходит детоксикация ядовитых веществ.
Аппарат Гольджи (комплекс Гольджи, пластинчатый комплекс). Назван в честь К. Гольджи, который в 1898 г. обнаружил его в нервных клетках.
Основой аппарата Гольджи является диктиосома – стопка уплощенных одномембранных цистерн. Количество диктиосом в клетке может достигать 20. Если диктиосомы расположены независимо друг от друга, то такая структура аппарата
Гольджи называется диффузной. Если диктиосомы связаны между собой каналами в единую трехмерную систему, то такая структура называется сетчатой.
В зоне аппарата Гольджи наблюдается множество мелких вакуолей. Часть вакуолей имеет ретикулярное происхождение, то есть они образуются путем отшнуровывания от эндоплазматического ретикулума. Путем слияния этих вакуолей и
образуются цистерны аппарата Гольджи. Другая часть вакуолей (обычно более крупных) образуется путем отшнуровывания от цистерн аппарата Гольджи.
В цистернах аппарата Гольджи завершается формирование компонентов плазмалеммы. Здесь же завершается модификация экспортных белков. От аппарата Гольджи отшнуровываются секреторные вакуоли и первичные лизосомы (см. ниже).
Таким образом, функции аппарата Гольджи сводятся к накоплению разнообразных веществ, их модификации и сортировке, упаковке конечных продуктов в одномембранные пузырьки, выведению секреторных вакуолей за пределы клетки и
формированию первичных лизосом.
У одноклеточных организмов расширенные цистерны аппарата Гольджи образуют сократительные вакуоли. В передней части сперматозоидов расширенная цистерна аппарата Гольджи образует акросому, которая содержит литические
ферменты, растворяющие оболочки яйцеклетки.
23.
18) Одномембранные органеллы клетки: канальцевая и вакуолярная система клетки — ЭПС, Комплекс Гольджи,диктиосомы, лизосомы, микротельца, пероксисомы. Их строение и функции.
Лизосомы – это одномембранные пузырьки диаметром 0,1...0,5 мкм, содержащие гидролитические ферменты (протеазы, нуклеазы, липазы и кислые фосфатазы).
Лизосомы открыл биохимик Де Дюв (1955). Дальнейшее их изучение велось с помощью биохимических и электронно-микроскопических методов. Лизосомы хорошо
изучены у животных. Существование настоящих лизосом у растений не доказано.
Первичные лизосомы образуются при отшнуровывании от периферической части аппарата Гольджи. Их размеры очень малы (около 0,1 мкм). Затем эти первичные
лизосомы сливаются с фагосомами (фагоцитарными вакуолями), образуя вторичные лизосомы (пищеварительные вакуоли). Вторичные лизосомы могут сливаться
между собой. Вещества, поглощенные клеткой, подвергаются гидролизу, продукты которого через мембрану вторичной лизосомы поступают в цитоплазматический
матрикс. Лизосома, содержащая непереваренные вещества, превращается в остаточное тельце. Остаточные тельца выводятся из клетки путем экзоцитоза или остаются
в ее составе вплоть до гибели клетки.
Первичные лизосомы могут изливать свое содержимое за пределы клетки (при внеклеточном пищеварении) или превращаться в автолизосомы. Автолизосомы
(фагосомы) образуются при слиянии первичных лизосом и отработанных внутриклеточных структур: фрагментов эндоплазматической сети, митохондрий, пластид,
рибосом, включений и т.д. Автолизосомы выполняют роль внутриклеточных чистильщиков, их количество возрастает при повреждении клеток, при стрессах, при
различных генетических и инфекционных заболеваниях.
Секреторные вакуоли (или секреторные гранулы) – это короткоживущие одномембранные пузырьки, которые образуются путем отшнуровывания от периферической
части аппарата Гольджи.
Секреторные вакуоли содержат разнообразные вещества (неактивные ферменты, или проферменты, полисахариды, липиды), выводимые за пределы клетки путем
экзоцитоза. Секреторные вакуоли хорошо видны в специализированных клетках экзокринных желез.
Пероксисомы (микротельца) – это одномембранные пузырьки диаметром 0,3...1,5 мкм, которые образуются путем отшнуровывания от цистерн гранулярной
эндоплазматической сети. Пероксисомы заполнены гранулярным матриксом и содержат разнообразные ферменты, например, каталазу, разлагающую пероксид
водорода. В ряде случаев пероксисомы содержат и другие системы ферментов.
У проростков некоторых растений встречаются и другие органоиды, содержащие каталазу – глиоксисомы. Глиоксисомы участвуют в липидно-углеводном обмене
веществ.
Сферосомы – это одномембранные пузырьки диаметром около 1 мкм, которые образуются путем отшнуровывания от эндоплазматической сети. Сферосомы
характерны для клеток растений. Первичная сферосома (просферосома) накапливает липиды, увеличивается в размерах, затем утрачивает мембрану и превращается в
масляную каплю.
Кроме липидов в составе сферосом имеются ферменты липазы, контролирующие превращения липидов.
Вакуоли – это заполненные жидкостью крупные одномембранные полости. Настоящие вакуоли имеются только у растений.
Первично вакуоли образуются при слиянии мелких пузырьков, отшнуровывающихся от эндоплазматической сети. В ходе функционирования вакуолей в их состав могут
включаться пузырьки, отшнуровывающиеся от аппарата Гольджи. Мембрана крупных вакуолей имеет собственное название – тонопласт. Содержимое вакуолей
называется клеточным соком. В состав клеточного сока входят неорганические соли, пигменты, растворимые углеводы, органические кислоты, некоторые белки.
Функции вакуолей разнообразны: регуляция водно-солевого режима, накопление пигментов (например, антоциана), накопление алкалоидов, таннидов, латекса,
минеральных солей, некоторых отходов жизнедеятельности.
24.
19) Трубчатые структуры клетки: центриоли, базальные тельца, жгутики, реснички, элементы цитоскелета.Центрио́ ль — внутриклеточный органоид эукариотической клетки, представляющий тельца в структуре клетки, размер которых находится на границе разрешающей способности светового
микроскопа.
Эти органеллы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках центриоли часто определяют полярность клеток
эпителия и располагаются вблизи комплекса Гольджи.
Термин был предложен Теодором Бовери в 1895 году. Тонкое строение центриолей удалось изучить с помощью электронного микроскопа. В некоторых объектах удавалось наблюдать центриоли,
обычно расположенные в паре (диплосома), и окруженные зоной более светлой цитоплазмы, от которой радиально отходят тонкие фибриллы (центросфера). Совокупность центриолей и
центросферы называют клеточным центром.
Чаще всего пара центриолей лежит вблизи ядра. Каждая центриоль построена из цилиндрических элементов (микротрубочек), образованных в результате полимеризации белка тубулина. Девять
триплетов микротрубочек расположены по окружности.
Центриоли принимают участие в формировании цитоплазматических микротрубочек во время деления клетки и в регуляции образования митотического веретена. В клетках высших растений и
большинства грибов центриолей нет, и митотическое веретено образуется там иным способом. Кроме того, ученые полагают, что ферменты клеточного центра принимают участие в процессе
перемещения дочерних хромосом к разным полюсам в анафазе митоза.
Базальные тельца. Зернышки в клетках, имеющих жгутики или реснички, находящиеся в основании данных жгутиков и ресничек. Для растений является нормальным нахождение базальных телец в
зооспорах водорослей, сперматозоидах папоротников, мхов, цикадовых и харовых. Согласно мнениям Эннеги-Леноссека и Шаудина, базальные тельца происходят благодаря центральному аппарату
клеток и соответственно их центросомам. Данная связь, прежде всего, существует во время образования сперматозоидов растительного и животного мира, во время образования жгутиков,
воротничковых клеток губок и так далее. Базальные тельца вполне могут возникнуть в глубине клеток или наружном слое клеток – в эпителии дыхательных путей или так называемом альвеолярном
слое. Могут они возникать и в области клеточного ядра, как например у инфузорий. При делении сперматозоидов, эпителиальных и прочих клеток, в некоторых случаях происходит так же деление
базальных телец. Они - динамические центры, отвечающие за движение жгутиков и ресничек. Согласно теории Гартмана, базальные тельца служат именно в качестве механической опоры ресничек
и жгутиков. Это может доказывать то, что в клетках, относящихся к ресничному эпителию, от базальных телец могу отходить не только реснички, но и корневые фибрилли, которые образуют
внутриклеточный пучок, выполняющий опорную функцию. Ботаническая цитология для базальных телец использует название – блефаропласты.
Цитоскеле́т — это клеточный каркас или скелет, находящийся в цитоплазме живой клетки. Он присутствует во всех клетках как у эукариот, так и у прокариот. Это динамичная, изменяющаяся
структура, в функции которой входит поддержание и адаптация формы клетки ко внешним воздействиям, экзо- и эндоцитоз, обеспечение движения клетки как целого, активный внутриклеточный
транспорт и клеточное деление.
Цитоскелет образован белками. В цитоскелете выделяют несколько основных систем, называемых либо по основным структурным элементам, заметным при электронно-микроскопических
исследованиях (микрофиламенты, промежуточные филаменты, микротрубочки), либо по основным белкам, входящим в их состав (актин-миозиноваясистема, кератины, тубулин-динеиновая
система).
Порядка 7 нм в диаметре, микрофиламенты представляют собой две цепочки из мономеров актина, закрученные спиралью. В основном они сконцентрированы у внешней мембраны клетки, так как
отвечают за форму клетки и способны образовывать выступы на поверхности клетки (псевдоподии и микроворсинки). Также они участвуют в межклеточном взаимодействии (образовании
адгезивных контактов), передаче сигналов и, вместе с миозином — в мышечном сокращении. С помощью цитоплазматических миозинов по микрофиламентам может осуществляться везикулярный
транспорт. Диаметр промежуточных филаментов составляет от 8 до 11 нанометров. Они состоят из разного рода субъединиц и являются наименее динамичной частью цитоскелета. Микротрубочки
представляют собой полые цилиндры порядка 25 нм диаметром, стенки которых составлены из 13 протофиламентов, каждый из которых представляет линейный полимер из димера белка тубулина.
Димер состоит из двух субъединиц — альфа- и бета- формы тубулина. Микротрубочки — крайне динамичные структуры, потребляющие ГТФ в процессе полимеризации. Они играют ключевую роль
во внутриклеточном транспорте (служат «рельсами», по которым перемещаются молекулярные моторы — кинезин и динеин), образуют основу аксонемы ундилиподий и веретено деления при
митозе и мейозе.
Трубчатые структуры клетки включают центриоли, базальные тельца, жгутики, реснички и элементы цитоскелета. Центриоли - это парные цилиндрические органеллы, состоящие из микротрубочек,
которые участвуют в делении клетки и образовании ворсинок и ресничек. Базальные тельца - это структуры, располагающиеся у основания жгутиков и ресничек. Они обеспечивают поддержку и
ориентацию этих структур. Жгутики - это длинные и тонкие образования, состоящие из микротрубочек. Их функции варьируются в зависимости от типа клетки, но, в основном, они играют роль в
движении органелл и целого организма. Реснички - это короткие и волнообразные образования, состоящие из микротрубочек. Они выполняют разнообразные функции, включая обеспечение
подвижности клетки и создание потока жидкости вокруг клетки.
Элементы цитоскелета - это сеть микротрубочек, интерфиламентных филаментов и микрофиламентов, которые обеспечивают структурную поддержку клетки, участвуют в ее движении и транспорте
внутриклеточных структур и молекул. Элементы цитоскелета также играют роль в клеточной делении и передвижении органелл и везикул внутри клетки.
25.
20) Строение и функции митохондрий.Митохондрии являются двухмембранными органеллами, присутствующими в клетках эукариотических организмов. Они обладают своей собственной ДНК и способностью к делению, что
свидетельствует об их эволюционной происхождении от прокариотических организмов.
Строение:
Внешняя мембрана. Состоит из фосфолипидного бислоя, содержит ряд белков, включая каналы и переносчики веществ.
Пространство интермембранного пространства. Объемное пространство между внешней и внутренней мембранами митохондрии.
Внутренняя мембрана. Обладает сложным строением, включает в себя многочисленные взаимосвязанные складки, называемые христа. Здесь расположены ферменты, необходимые для процессов
дыхания и фосфорилирования.
Матрикс. Жидкая гель-подобная субстанция, заполняющая внутреннюю полость митохондрии. Здесь располагается митохондриальная ДНК, ферменты клеточного дыхания и другие необходимые
молекулы.
Функции:
Энергетическое обеспечение
Регуляция клеточного метаболизма.
Участие в программированной клеточной гибели
Роль в регуляции кальция
В целом, митохондрии считаются "энергетическими централизаторами" клетки, обеспечивая энергию для ее выживания и функционирования.
Митохондрия образована двумя мембранами — внешней и внутренней, между которыми расположено межмембранное пространство. Внутренняя мембрана образует множество впячиваний —
крист, представляющих собой либо пластины, либо трубочки. Такая ее организация обеспечивает огромную площадь внутренней мембраны. На ней располагаются ферменты, обеспечивающие
преобразование энергии, заключенной в органических веществах (углеводах, липидах), в энергию АТФ, необходимую для жизнедеятельности клетки. Следовательно, функция митохондрий —
участие в энергетических клеточных процессах. Именно поэтому большое количество митохондрий присуще, например, мышечным клеткам, выполняющим большую работу.
Между наружной и внутренней мембранами возникает неширокое (около 15 нм) пространство, которое называется наружной камерой митохондрии; внутренняя мембрана ограничивает
внутреннюю камеру. Содержимое наружной и внутренней камер митохондрии различно, и так же, как и сами мембраны, существенно отличается не только по рельефу поверхности, но и по ряду
биохимических и функциональных признаков. Наружная мембрана по химическому составу и свойствам близка к другим внутриклеточным мембранам и плазмолемме.
Ее характеризует высокая проницаемость, благодаря наличию гидрофильных белковых каналов. Эта мембрана имеет в своем составе рецепторные комплексы, распознающие и связывающие
вещества, поступающие в митохондрию.
Внутренняя мембрана митохондрий в большинстве клеток тканей различных органов формирует кристы в виде пластин (ламеллярные кристы), что значительно увеличивает площадь поверхности
внутренней мембраны. В эндокринных клетках надпочечников и половых желез митохондрии участвуют в синтезе стероидных гормонов. В этих клетках митохондрии имеют кристы в виде трубочек
(тубул), упорядоченно расположенных в определенном направлении. Поэтому кристы митохондрий в стероидпродуцирующих клетках названных органов именуются тубулярными.
Матрикс митохондрии, или содержимое внутренней камеры, представляет собой гелеобразную структуру, содержащую около 50 % белков. Матрикс содержит ферменты цикла лимонной кислоты,
катализирующие окисление жирных кислот, синтез рибосом, ферменты, участвующие в синтезе РНК и ДНК
В клетке митохондрии выполняют функцию дыхания. Клеточное дыхание — это последовательность реакций, с помощью которых клетка использует энергию связей органических молекул для
синтеза макроэргических соединений типа АТФ.
26.
21) Включения клеток.Включения клеток - это различные вещества, структуры или органеллы, которые находятся внутри клетки и выполняют определенные функции. Включения могут быть различных типов и выполнять
различные функции, в зависимости от типа клетки и ее потребностей.
Примеры включений клеток включают:
Хромосомы - это структуры, содержащие генетическую информацию клетки. Хромосомы находятся в ядре клетки и отвечают за передачу наследственной информации от поколения к поколению.
Митохондрии - это органеллы, которые выполняют функцию производства энергии в клетке. Митохондрии производят АТФ, основной энергетический носитель клетки.
Лизосомы - это органеллы, которые выполняют функцию переработки и утилизации отходов в клетке. Лизосомы содержат ферменты, которые разлагают молекулы и органические соединения.
Рибосомы - это структуры, где происходит синтез белка. Рибосомы считывают информацию из РНК и строят белковые цепи.
Вакуоли - это органеллы, которые выполняют функцию хранения веществ в клетке. Вакуоли могут содержать в себе воду, питательные вещества, а также различные отходы и токсины.
Эндоплазматическое ретикулум - это система мембранных каналов в клетке, которые выполняют функцию синтеза и транспорта биохимических веществ внутри клетки.
Гольджи аппарат - это органелла, ответственная за сортировку, изменение и упаковку молекул, которые будут отправлены в другие части клетки или внеклеточное пространство.
В состав цитоплазмы входят включения – структуры, которые не являются ее обязательными компонентами. Включения разнообразны по химическому составу, происхождению и функциям.
Эргастические включения содержат энергию для жизнедеятельности клетки. К ним относятся: жировые капли, гранулы гликогена и крахмальные зерна, гранулы белка и алейроновые зерна.
Неэргастические включения не служат источниками энергии. К ним относятся: некоторые пигменты (меланин, антоциан), эфирные масла, кристаллы оксалата кальция.
Особый вид включений клетки — остаточные тельца — продукты деятельности лизосом.
По характеру все включения - это продукты клеточного метаболизма. Они накапливаются главным образом в форме гранул, капель и кристаллов. Химический состав включений очень разнообразен.
Липоиды обычно откладываются в клетке в виде мелких капель. Большое количество жировых капель встречается в цитоплазме ряда простейших, например инфузорий. У млекопитающих жировые
капли находятся в специализированных жировых клетках, в соединительной ткани. Часто значительное количество жировых включений откладывается в результате патологических процессов,
например при жировом перерождении печени. Капли жира встречаются в клетках практически всех растительных тканей, очень много жира содержится в семенах некоторых растений.
Включения полисахаридов имеют чаще всего формулу гранул разнообразных размеров. У многоклеточных животных и простейших в цитоплазме клеток встречаются отложения гликогена . Гранулы
гликогена хорошо видны в световом микроскопе. Особенно велики скопления гликогена в цитоплазме поперечнополосатых мышечных волокон и в клетках печени, в нейронах. В клетках растений из
полисахаридов наиболее часто откладывается крахмал. Он имеет вид гранул различной формы и размеров, причем форма крахмальных гранул специфична для каждого вида растений и для
определенных тканей. Отложениями крахмала богата цитоплазма клубней картофеля, зерен злаков; каждая крахмальная гранула состоит их отдельных слоев, а каждый слой, в свою очередь,
включает радиально расположенные кристаллы, почти невидимые в световой микроскоп.
Белковые включения встречаются реже, чем жировые и углеводные. Белковыми гранулами богата цитоплазма яйцеклеток, где они имеют форму пластинок, шариков, дисков, палочек. Белковые
включения встречаются в цитоплазме клеток печени, клеток простейших и многих других животных.
К клеточным включениям относятся некоторые пигменты, например распространенный в тканях желтый и коричневый пигмент липофусцин , круглые гранулы которого накапливаются в процессе
жизнедеятельности клеток, особенно по мере их старения. Сюда же относятся пигменты желтого и красного цвета - липохромы . Они накапливаются в виде мелких капель в клетках коркового
вещества надпочечников и в некоторых клетках яичников. Пигмент ретинин входит в состав зрительного пурпура сетчатки глаза. Присутствие некоторых пигментов связано с выполнением этими
клетками особых функций. Примерами могут служить красный дыхательный пигмент гемоглобин в эритроцитах крови или пигмент меланин в клетках меланофорах покровных тканей животных.
В качестве включений во многих животных клетках присутствуют гранулы секрета, вырабатываемого в клетках разных типов, в первую очередь в железистых. Секреторные включения могут быть
белками, сахаридами, липопротеидами и т. д.
27.
22) Строение и функции клеточного ядра. Структура и организация хроматина: эухроматин и гетерохроматин, его роль в регуляцииактивности генов.
Ядро — это один из структурных компонентов эукариотической клетки, содержащий генетическую информацию (молекулы ДНК). В ядре происходитрепликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на
молекуле ДНК. В ядре же синтезированные молекулы РНК подвергаются ряду модификаций, после чего выходят в цитоплазму. Образование субъединиц рибосом также происходит в ядре в специальных образованиях - ядрышках.
Считается, что в ядре существуют так называемые функциональные домены хроматина(ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». К
сожалению, вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом
закреплены на белках ядерной ламины.
От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких
компартментов. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белкамиламинами, к которой прикреплены нитихромосомной ДНК. Ламины прикрепляются к внутренней мембране ядерной оболочки при помощи заякоренных в ней трансмембранных белков — рецепторов ламинов. В некоторых местах внутренняя и
внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Пора не является дыркой в ядре, а имеет сложную структуру,
организованную несколькими десятками специализированных белков —нуклеопоринов. Под электронным микроскопом она видна как восемь связанных между собой белковых гранул с внешней и столько же с внутренней стороны ядерной
оболочки.
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются
специальные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных
субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. Nucleolus Localization Signal). Следует
отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для
осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного
фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
Ядро выполняет 2 главные функции:
1)
хранение, воспроизведение и передачу наследственной информации;
2)
регуляцию процессов обмена веществ, протекающих в клетке.
Хроматин (от греч. chroma - краска) мелкие зернышки и глыбки материала, который обнаруживается в ядре клеток и окрашивается ос¬новными красителями. Хроматин состоит изКомплекса ДНК и белка И соответствует хромосомам, которые в
интерфазном ядре представлены длинными, тонкими перекрученными нитями и неразличимы как индивидуальные структуры. Выраженность спирализапии каждой из хромо¬сом неодинакова по их длине. Различают два вида хроматина Эухроматин и гетерохроматин.
Эухроматин. Соответствует сегментам хромосом, которыеДеспира-лизованы и открыты для транскрипции. Эти сегментыНе окрашива¬ются И не видны в световой микроскоп.
Гетерохроматин. Соответствует Конденсированным, Плотно скру¬ченным сегментам хромосом (что делает их Недоступными для транс¬крипции). Он Интенсивно окрашиваетсяОсновными красителями, и в световом микроскопе имеет вид
гранул.
Таким образом, По морфологическим признакам ядра (соотноше¬нию содержания эу - и гетерохроматина) можно оценить активность процессов транскрипции, а, следовательно, синтетической функции клетки. При ее повышении это
соотношение изменяется в пользу эухроматина, при снижении - нарастает содержание гетерохроматина. При полном подавлении функции ядра (например, в поврежденных и гибну¬щих клетках, при ороговении эпителиальных клеток
эпидермиса - кера-тиноцитов, при образовании ретикулоцитов крови) оно уменьшается в размерах, содержит только гетерохроматин и окрашивается основны¬ми красителями интенсивно и равномерно. Такое явление называется
Кариопикнозом (от греч. karyon - ядро и pyknosis - уплотнение).
Распределение гетерохроматина (топография его частиц в яд¬ре) и соотношение содержания эу - и гетерохроматина Характерны для клеток каждого типа, что позволяет осуществлять их идентификацию как визуально, так и с помощью
автоматических анализаторов изобра¬жения. Вместе с тем, имеются определенные общие закономерности распределения гетерохроматина В ядре: его скопления располагают¬сяПод кариолеммой, прерываясь в области пор (что обусловлено
его связью с ламиной) и вокруг ядрышка (Перинуклеолярный гетерохроматин), более мелкие глыбки разбросаны по всему ядру .
Тельце Барра - Скопление гетерохроматина, соответствующее од¬ной Х-хромосоме у особей женского пола, которая в интерфазе плотно скручена и неактивна. В большинстве клеток оно лежит у кариолеммы, а в гранулоцитах крови имеет вид
маленькой добавочной дольки ядра ("барабанной палочки"). Выявление тельца Барра (обычно в эпителиальных клетках слизистой оболочки полости рта) используется как ди¬агностический тест для определения генетического пола
(обязателен, в частности, для женщин, участвующих в Олимпийских Играх).
28.
22) Строение и функции клеточного ядра. Структура и организация хроматина: эухроматин игетерохроматин, его роль в регуляции активности генов.
Упаковка хроматина в ядре. В дсконденсированном состоянии длина одной молекулы (двойной спирали) ДНК, образующей каждую хромосому, равна в среднем, около
5 см, а общая длина молекул ДНК всех хромосом в ядре (диаметром около 10 мкм) составляет более 2 м (что сравнимо с укладкой нити длиной 20 км в теннисный
мячик диа¬метром около 10 см), а в S-период интерфазы - более 4 м. Конкретные механизмы, препятствующие спутыванию этих нитей во время транс¬крипции и
репликации, остаются нераскрытыми, однако очевидна необ¬ходимость Компактной упаковки молекул ДНК, В клеточном ядре это осуществляется благодаря их связи
со специальными основными (гистоновыми) белками. Компактная упаковка ДНК в ядре обеспечивает:
(1) Упорядоченное расположение Очень длинных молекул ДНК в небольшом объеме ядра;
(2) функциональный Контроль активности генов (вследствие вли¬яния характера упаковки на активность отдельных участков генома.
Уровни упаковки хроматина . Начальный уровень упа¬ковки хроматина, обеспечивающий образованиеНуклеосомной нити Ди-аметром 11 нм, обусловлен намоткой
двойной нити ДНК (диаметром 2 нм) на блоки дисковидной формы из 8 гистоновых молекул (нуклеосомы). Нуклеосомы разделены короткими участками свободной
ДНК. Второй уровень упаковки также обусловлен гистонами и приводит к скручиванию нуклеосомной нити с формированием Хроматиновой фибриллы Диаметром 30
нм. В интерфазе хромосомы образованы хроматиновыми фибриллами, причем каждая хроматида состоит из одной фибриллы. При дальнейшей упаковке
хроматиновые фибриллы образу¬ют Петли (петельные домены) Диаметром 300 нм, каждый из которых соответствует одному или нескольким генам, а те, в свою
очередь, в результате еще более компактной укладки, формируют участки конденси¬рованных хромосом, которые выявляются лишь при делении клеток.
В хроматине ДНК связана помимо гастонов также и с Негистоновыми белками, Которые Регулируют активность генов. Вместе с тем, и гистоны, ограничивая доступность
ДНК для других ДНК-связьвзающих белков, могут участвовать в регулядии активности генов.
Функция хранения генетической информации В ядре в неизме¬ненном виде имеет исключительно важное значение для нормальной жизнедеятельности клетки и всего
организма. Подсчитано, что при ре¬пликации ДНК и в результате ее повреждений внешними факторами в каждой клетке человека ежегодно происходят изменения 6
нуклеотидов. Возникшие повреждения молекул ДНК могут исправляться в ре¬зультате процессаРепарации Или путем Замещения После Распознава¬ния и маркировки
соответствующего участка.
В случае невозможности репарации ДНК при слишком значитель¬ных повреждениях включается механизм запрограммированной гибели клетки . В этой ситуации
"поведение" клетки можно оценить как своего рода "альтруистическое самоубийство": ценой своей гибели она спасает организм от возможных негативных
последствий реплика¬ции и амплификации поврежденного генетического материала.
Способность к репарации ДНК у Взрослого человека снижается примерно на 1% с каждым годом. Это снижение может отчасти объяс¬нить, почему старение является
фактором риска развития злокачест¬венных заболеваний. Нарушения процессов репарации ДНК Характерно для ряда наследственных болезней, при которых резко
Повышены Как Чувствительность к повреждающим факторам, Так и Частота разви¬тия злокачественных новообразований.
Функция Реализации генетической информации В интерфазном ядре осуществляется непрерывно благодаря процессам Транскрипции. Геном млекопитающих содержит
около ЗхЮ9 нуклеотидов, однако не более 1% его объема кодирует важные белки и принимает участие в ре¬гуляции их синтеза. Функции основной некодирующей
части генома не¬известны.
При транскрипции ДНК образуется очень крупная молекула РНК (первичный транскрипт), которая связывается с ядерными белками с образованием
Рибонуклеопротеинов (РНП). В первичном РНК-транс¬крипте (как и в матричной ДНК) имеются дискретные значащие после¬довательности нуклеотидов (экзоны),
Разделенные длинными некодирующими вставками (нитронами). Процессинг РНК-транскрипта включает отщепление нитронов и стыковку экзонов - сплайсинг (от англ,
splicing - сращивание). При этом очень крупная молекула РНК превращается в достаточно мелкие молекулы иРНК, отделяющиеся от связанных с ни¬ми белков при
переносе в цитоплазму.
29.
23) Уровни организации хроматина: нуклеосомная нить, элементарная хроматиновая фибрилла, интерфазная хромонема, метафазнаяхроматида, их значение в митотическом цикле.
Хроматин (греч. chroma — цвет, краска и греч. nitos — нить) — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у
прокариот. Именно в составе хроматина происходитреализация генетической информации, а также репликация и репарация ДНК.
Нуклеосомиая нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных гистонов: Н2А, Н2В, НЗ, Н4. Они образуют напоминающие по форме шайбу белковые тела
— коры,состоящие из восьми молекул (по две молекулы каждого вида гистонов) (рис. 3.46).
Молекула ДНК комплектируется с белковыми корами, спирально накручиваясь на них. При этом в контакте с каждым кором оказывается участок ДНК, состоящий из 146 пар нуклеотидов (п.н.).
Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают от 15 до 100 п.н. (в среднем 60 п.н.) в зависимости от типа клетки.
Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому. Благодаря такой организации в основе структуры хроматина лежит нить, представляющая собой
цепочку повторяющихся единиц — нуклеосом (рис. 3.46, Б). В связи с этим геном человека, состоящий из 3 · 109 п. н., представлен двойной спиралью ДНК, упакованной в 1,5 · 107 нуклеосом.
Вдоль нуклеосомной нити, напоминающей цепочку бус, имеются области ДНК, свободные от белковых тел. Эти области, расположенные с интервалами в несколько тысяч пар нуклеотидов, играют
важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками.
В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм.
Хроматиновая фибрилла. Дальнейшая компактизация нуклеосомной нити обеспечивается пистоном HI, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их
друг с другом. В результате образуется более компактная структура, построенная, возможно, по типу соленоида. Такая Хроматиновая фибрилла, называемая также элементарной, имеет диаметр 20—
30 нм
Интерфазная хромонема. Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли. В их образовании, по-видимому, принимают
участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар
нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы (рис. 3.48). Участок ДНК, соответствующий одной
петле, содержит от 20 000 до 80 000 п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки Хроматиновая фибрилла диаметром 20—30 нм
преобразуется в структуру диаметром 100—200 нм, называемую интерфазной хромонемой.
Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяющие соседние петли с одинаковой организацией (рис. 3.49). Они
выявляются в интерфазном ядре в виде глыбок хроматина. Возможно, существование таких структурных блоков обусловливает картину неравномерного распределения некоторых красителей в
метафазных хромосомах, что используют в цитогенетических исследованиях
Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина. Отдельные хромосомы становятся хорошо различимы. Этот процесс начинается в
профазе, достигая своего максимального выражения в метафазе митоза и анафазе (см. разд. 2.4.2). В телофазе митоза происходит декомпак-тизация вещества хромосом, которое приобретает
структуру интерфазного хроматина. Описанная митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза. Степень компактизации
хроматина в разные периоды митотического цикла клетки можно оценить по данным.
Нуклеосомная нить: это основная структурная единица хроматина, состоящая из ДНК, намотанной на белки гистоны. Нуклеосомы сгруппированы в более крупные структуры.
Элементарная хроматиновая фибрилла: это длинная нить, образованная нуклеосомами, которые связаны друг с другом. Она образует петли, которые могут пролезать через поры ядра.
Интерфазная хромонема: это суперспирализованная структура хроматина, которая образуется в период интерфазы клеточного цикла. Хромонема состоит из нескольких элементарных хроматиновых
фибрилл, связанных друг с другом.
Метафазная хроматида: в митотическом цикле хроматин конденсируется и формирует хромосомы, состоящие из двух одинаковых нитей - метафазных хроматид. В процессе деления клетки на две
новые клетки, метафазные хроматиды разделяются между дочерними клетками.
Уровни организации хроматина играют важную роль в митотическом цикле, который включает в себя процессы деления клетки. Конденсация хроматина в метафазные хроматиды позволяет
правильно распределить генетическую информацию между дочерними клетками. Интерфазная хромонема обеспечивает компактность и упорядоченность хроматина в период между делениями,
чтобы генетическая информация могла быть скопирована и подготовлена к делению клетки.
30.
24) Политенные хромосомы, хромосомы типа ламповых щеток, их строение ифункциональное значение.
Политенные хромосомы (гигантские хромосомы) содержат во много раз больше ДНК, чем обычные. Они не изменяют своей формы на протяжении цикла деления и достигают длины до 0,5 мм и толщины 25 мкм. Они встречаются, например, в
слюнных железах двукрылых (мух и комаров), в макронуклеусе инфузорий и в тканях завязи бобов. Чаще всего они видны в гаплоидном числе, так как гомологичные хромосомы тесно спарены.
Политения возникает в результате эндорепликации. По сравнению с эндомитозом это еще более редуцированный процесс деления - после репликации хроматиды не разделяются. Этот процесс повторяется многократно.
При этом разные отрезки ДНК умножаются в различной степени:
•участки центромер - незначительно;
•большинство информативных областей - приблизительно в 1000 раз;
•некоторые - более чем в 30000 раз.
Поэтому политенные хромосомы представляют собой пучки из бесчисленных хроматид, разделенных не полностью. Эти хроматиды растянуты, гомологичные хромомеры образуют темные диски, тесно расположенные вдоль хромосомы. Эти
диски разделены более светлыми полосами. Вероятно, на хроматиде один диск и одна промежуточная полоса образуют, помимо спейсера, один ген (реже несколько генов), который, по-видимому, находится в диске. Политенные хромосомы
чрезвычайно бедны гетерохроматином.
На политенных хромосомах отдельные диски временами раздуваются в пуфы (кольца Бальбиани). Там гомологичные хроматиды отделяются друг от друга, гомологичные хромомеры раздвигаются и возникает разрыхленная структура
транскрипционно-активного хроматина. В пуфах содержится меньше гистона Н1, чем в дисках, вместо него здесь находится фермент РНК-полимераза (что указывает на синтез РНК).
В промежуточных полосах тоже мало гистона Н1, но есть РНК-полимераза и, возможно, происходит хотя бы незначительный синтез РНК.
Хромосомы типа ламповых щеток, впервые обнаруженные В. Флеммингом в 1882 году, — это специальная форма хромосом, которую они приобретают в растущих ооцитах (женских половых клетках) большинства животных, за исключением
млекопитающих
В растущих ооцитах всех животных, за исключением млекопитающих, во время протяженной стадии диплотены профазы мейоза I активная транскрипция многих последовательностей ДНК приводит к преобразованию хромосом в хромосомы,
по форме напоминающие щетки для чистки стёкол керосиновых ламп (хромосомы типа ламповых щёток). Они представляют собой сильно деконденсированные полубиваленты, состоящие из двух сестринских хроматид. Хромосомы типа
ламповых щеток можно наблюдать с помощью световой микроскопии, при этом видно, что они организованы в виде серии хромомеров (содержат конденсированный хроматин) и исходящих из них парных латеральных петель (содержат
транскрипционно активныйхроматин).
Наиболее подробно описана организация хромосом типа ламповых щеток хвостатых и бесхвостых амфибий, доместицированных видов птиц и некоторых видов насекомых. Хромосомы типа ламповых щёток амфибий и птиц могут быть
изолированы из ядра ооцита с помощью микрохирургических манипуляций.
Хромосомы типа ламповых щёток производят огромное количество РНК, синтезируемой на латеральных петлях. Каждая латеральная петля всегда содержит одну и ту же последовательность ДНК и остаётся в вытянутом состоянии на
протяжении всего роста ооцита, вплоть до начала конденсации хромосом. Латеральная петля может содержать одну или несколько транскрипционных единиц с поляризованным РНП-матриксом, покрывающим ДНП-ось петли. Вместе с тем,
большая часть ДНК остается в конденсированном состоянии и организована в хромомеры в осях хромосом типа ламповых щёток.
Благодаря гигантским размерам и выраженной хромомерно-петлевой организации, хромосомы типа ламповых щёток на протяжении многих десятилетий служат удобной моделью для изучения организации хромосом, работы генетического
аппарата и регуляции экспрессии генов во время профазы мейоза I. Кроме того, хромосомы этого типа широко используются для картирования последовательностей ДНК с высокой степенью разрешения, изучения феномена транскрипции
некодирующих белки тандемных повторов ДНК, анализа распределения хиазм и др.
Политенные хромосомы - это особый тип хромосом, образующихся в ходе процесса политении. При политении хромосомы удлиняются и происходит их парный склеивание, что приводит к образованию хромосом типа ламповых щеток.
Строение политенных хромосом отличается от обычных хромосом. Они имеют многоочковое утолщение (блошинки), в результате склеивания дублированных хроматид. Эти дублированные хроматиды содержат повторяющиеся
последовательности ДНК.
Функциональное значение политенных хромосом заключается в увеличении активности генов. При политении дублированные хроматиды вытягиваются и становятся параллельными друг другу. Это позволяет более эффективно организовать
транскрипцию ДНК и, следовательно, увеличить синтез РНК и белков.
Политенные хромосомы обнаруживаются в клетках некоторых организмов, таких как некоторые насекомые и растения. Этот процесс является временным и протекает на определенных стадиях развития организма или в ответ на определенные
условия. Например, политенные хромосомы могут образовываться в саливарных железах мух при росте их личинок.
31.
25) Обмен веществ и энергии: роль АТФ вжизнедеятельности клеток.
Обязательным условием существования любого организма является постоянный приток питательных веществ и постоянное выделение конечных продуктов химических реакций, происходящих в
клетках. Питательные вещества используются организмами в качестве источника атомов химических элементов (прежде всего атомов углерода), из которых строятся либо обновляются все
структуры. В организм, кроме питательных веществ, поступают также вода, кислород, минеральные соли.Поступившие в клетки органические вещества (или синтезированные в ходе фотосинтеза)
расщепляются на строительные блоки — мономеры и направляются во все клетки организма. Часть молекул этих веществ расходуется на синтез специфических органических веществ, присущих
данному организму. В клетках синтезируются белки, личиды, углеводы, нуклеиновые кислоты и другие вещества, которые выполняют различные функции (строительную, каталитическую,
регуляторную, защитную и т. д.).Другая часть низкомолекулярных органических соединений, поступивших в клетки, идет на образование АТФ, в молекулах которой заключена энергия,
предназначенная непосредственно для выполнения работы. Энергия необходима для синтеза всех специфических веществ организма, поддержания его высокоуно-рядоченной организации,
активного транспорта веществ внутри клеток, из одних клеток в другие, из одной части организма в другую, для передачи нервных импульсов, передвижения организмов, поддержания постоянной
температуры тела (у птиц и млекопитающих) и для других целей.В ходе превращения веществ в клетках образуются конечные продукты обмена, которые могут быть токсичными для организма и
выводятся из него (например, аммиак). Таким образом, все живые организмы постоянно потребляют из окружающей среды определенные вещества, преобразуют их и выделяют в среду конечные
продукты.Совокупность химических реакций, происходящих в организме, называется обменом веществ нли метаболизмом.В зависимости от общей направленности процессов выделяют
катаболизм и анаболизм.
Катаболизм (диссимиляция) —совокупность реакций, приводящих к образованию простых соединений из более сложных. К катаболическим относят, например, реакции гидролиза полимеров до
мономеров и расщепление последних до углекислого газа, воды, аммиака, т. е. реакции энергетического обмена, в ходе которого происходит окисление органических веществ и синтез АТФ.
Анаболизм (ассимиляция) — совокупность реакций синтеза сложных органических веществ из более простых. Сюда можно отнести, например, фиксацию азота и биосинтез белка, синтез углеводов
из углекислого газа и воды в ходе фотосинтеза, синтез полисахаридов, липидов, нуклеотидов, ДНК, РНК и других веществ.
Синтез веществ в клетках живых организмов часто обозначают понятием пластический обмеи, а расщепление веществ и их окисление, сопровождающееся синтезом АТФ, —энергетическим
обменом. Оба вида обмена составляют основу жизнедеятельности любой клетки, а следовательно, и любого организма и тесно связаны между собой. С одной стороны, все реакции пластического
обмена нуждаются в затрате энергии. С другой стороны, для осуществления реакций энергетического обмена необходим постоянный синтез ферментов, так как продолжительность их жизни
невелика. Кроме того, вещества, используемые для дыхания, образуются в ходе пластического обмена (например, в процессе фотосинтеза).
Молекула АТФ представляет собой нуклеотид, образованный азотистым основанием аденином, пятиуглеродным сахаром рибозой и тремя остатками фосфорной кислоты. Фосфатные группы в
молекуле АТФ соединены между собой высокоэнергетическими (макроэргическими) связями :
Связи между фосфатными группами не очень прочные, и при их разрыве выделяется большое количество энергии. В результате гидролитического отщепления от АТФ фосфатной группы образуется
аденозиндифосфорная кислота (АДФ) н высвобождается порция энергии:
АДФ также может подвергаться дальнейшему гидролизу с отщеплением еще одной фосфатной группы и выделением второй порции энергии; при этом АДФ преобразуется в аденозин-монофосфат
(АМФ), который далее не гидролизуется
АТФ образуется из АДФ и неорганического фосфата за счет энергии, освобождающейся при окислении органических веществ и в процессе фотосинтеза. Этот процесс называется
фосфорилированием. При этом должно быть затрачено не менее 40 кДж/моль энергии, которая аккумулируется в макроэргических связях:
Следовательно, основное значение процессов дыхания и фотосинтеза определяется тем, что они поставляют энергию для синтеза АТФ, с участием которой в клетке выполняется большая часть
работы.
Таким образом, АТФ — это главный универсальный поставщик энергии в клетках всех живых организмов.
АТФ чрезвычайно быстро обновляется. У человека, например, каждая молекула АТФ расщепляется и вновь восстанавливается 2 400 раз в сутки, так что ее средняя продолжительность жизни менее 1
мин. Синтез АТФ осуществляется главным образом в митохондриях и хлоропластах (частично в цитоплазме). Образовавшаяся здесь АТФ направляется в те участки клетки, где возникает потребность
в энергии.
32.
26) Передача наследственной информации при делении соматических клеток. Жизненный цикл клетки. Интерфаза. Митоз.Митотический индекс. Нарушение митоза.
В опухолях атипичные клетки делятся митотическим способом. В результате деления образуются идентичные измененной клетки. Деление происходит многократно. В итоге опухоль быстро растет.
Жизненный цикл клетки. Интерфаза. Митоз. Жизненный цикл клетки – это период ее жизни от деления до деления. Клетки размножаются путем удвоения своего содержимого с последующим делением пополам. Клеточное деление лежит в
основе роста, развития и регенерации тканей многоклеточного организма. Клеточный цикл подразделяют на интерфазу, сопровождающуюся точным копированием и распределением генетического материала и митоз – собственно деление
клетки после удвоения других клеточных компонентов. Длительность клеточных циклов у разных видов, в разных тканях и на разных стадиях широко варьирует от одного часа (у эмбриона) до года (в клетках печени взрослого человека).
Интерфаза – период между двумя делениями. В этот период клетка готовится к делению. Удваивается количество ДНК в хромосомах. Удваивается количество других органоидов, синтезируются белки, причем наиболее активно те из них,
которые образуют веретено деления, происходит рост клетки.
К концу интерфазы каждая хромосома состоит из двух хроматид, которые в процессе митоза станут самостоятельными хромосомами.
Митоз – это форма деления клеточного ядра. Следовательно, происходит он только в эукариотических клетках. В результате митоза каждое из образующихся дочерних ядер получает тот же набор генов, который имелародительская клетка. В
митоз могут вступать как диплоидные, так и гаплоидные ядра. При митозе получаются ядра той же плоидности, что и исходное. Митоз состоит из нескольких последовательных фаз.
Профаза. К разным полюсам клетки расходятся удвоенные центриоли. От них к центромерам хромосом протягиваются микротрубочки, образующие веретено деления. Хромосомы утолщены и каждая хромосома состоит из двух хроматид.
Метафаза. В этой фазе хорошо видны хромосомы, состоящие из двух хроматид. Они выстраиваются по экватору клетки, образуя метафазную пластинку.
Анафаза. Хроматиды расходятся к полюсам клетки с одинаковой скоростью. Микротрубочки укорачиваются. Телофаза. Дочерние хроматиды подходят к полюсам клетки. Микротрубочки исчезают. Хромосомы деспирализуются и снова
приобретают нитевидную форму. Формируются ядерная оболочка, ядрышко, рибосомы.
Цитокинез – процесс разделения цитоплазмы. Клеточная мембрана в центральной части клетки втягивается внутрь. Образуется борозда деления, по мере углубления которой клетка раздваивается.
В результате митоза образуются два новых ядра с идентичными наборами хромосом, точно копирующими генетическую информацию материнского ядра.
В опухолевых клетках ход митоза нарушается.
Митотический индекс (Mitotic index,MI, %) — процент делящихся клеток от общего числа проанализированных клеток. Данный индекс можно вычислить используя световой микроскоп, просчитав в поле зрения клетки с видимыми
хромосомами и разделив его на общее число клеток в поле зрения.
Правильное течение митоза может быть нарушено различными внешними воздействиями: высокими дозами радиации, некоторыми химическими веществами. Например, под действием рентгеновых лучей ДНК хромосом может разорваться.
Хромосомы в таком случае тоже разрываются. При этом могут возникнуть хромосомы без центромерного района. Такие хромосомы лишены способности двигаться в прометафазе и анафазе. В зависимости от того, в каком месте ядра
бесцентромерная хромосома находилась накануне деления,будет складываться ее дальнейшая судьба. Если хромосома была смещена к одному из полюсов клетки, то при формировании дочерних клеток она может целиком включиться в
одну из них, т. е. обе сестринские хроматиды окажутся в одном ядре. Если хромосома, лишенная центромерного района, окажется вблизи центральной части клетки, то велика вероятность того, что она не попадет ни в одно из формирующихся
ядер, так как в анафазе не сможет последовать к полюсу. В обоих случаях вновь возникшие клетки будут иметь хромосомный набор, отличающийся от набора хромосом в исходной клетке. Некоторые химические соединения, не свойственные
живым организмам (спирты, эфиры), нарушают согласованность митотических процессов. Одни хромосомы начинают двигаться быстрее, другие отстают. Отставшие хромосомы могут не включиться в формирующиеся дочерние ядра. Иногда в
делящейся клетке образуется не два, а три или четыре полюса, что ведет к возникновению соответственно трех или четырех дочерних клеток. При таком делении нарушается весь слаженный механизм распределения хромосом. Метафазная
хромосома, состоящая из двух сестринских хроматид, может взаимодействовать одновременно только с двумя полюсами. Если полюсов больше, то каждая хромосома вынуждена "выбирать", с какими двумя полюсами из трех или четырех ей
взаимодействовать. Этот выбор совершается случайно. В результате каждая дочерняя клетка получает не весь набор хромосом, а только его часть. Клетки, получившие неполный набор хромосом, как правило, оказываются
нежизнеспособными и погибают. Изучение нарушений митоза, вызванных различными факторами, с одной стороны, помогает лучше понять митотические процессы, с другой - позволяет устанавливать механизмы повреждающего действия
этих факторов и, следовательно, создает условия для целенаправленного поиска методов устранения таких нарушений
Предача наследственной информации при делении соматических клеток:
При делении соматических клеток (клеток, составляющих тело организма) наследственная информация передается через процесс деления, известный как митоз. Это обеспечивает, что каждая новая клетка содержит идентичную генетическую
информацию.
Жизненный цикл клетки включает три основные фазы: интерфаза (период роста и подготовки к делению), митоз (деление клетки) и цитокинез (деление цитоплазмы).
Интерфаза - это фаза, в течение которой клетка растет и подготавливается к делению. Включает в себя три стадии: Г1 (период роста), S (синтез ДНК) и G2 (подготовка к делению).
Митоз - это процесс деления клетки, состоящий из четырех последовательных фаз: профаза, метафаза, анафаза и телофаза. В результате митоза образуются две генетически идентичные дочерние клетки.
Митотический индекс - это отношение числа клеток, проходящих митоз, к общему числу клеток в образце. Он используется для оценки активности деления клеток в ткани или клеточной культуре.
Нарушение митоза может привести к различным проблемам, включая неправильное распределение хромосом, что может привести к мутациям, аномалиям и неспособности клетки разделиться корректно. Такие нарушения могут стать
причиной раковых заболеваний и других болезней.
33.
27) Прямое деление клеток: амитоз. К-митоз, эндомитоз, политения.АМИТОЗ (amitosis, is, f)- прямое деление ядра клетки, которое чаще всего происходит в соматических клетках, без формирования веретена деления. Является наиболее характерным для лейкоцитов
и чаще всего символизирует злокачественные клетки.
Впервые процесс деления клеток был охарактеризован Р.Ремаком – знаменитым немецким биологом в 1841 году. В 1882 году в медицине появляется научный термин – амитоз.
Чаще всего такой процесс деления клеток наблюдается в стареющих и патологически измененных клетках, которые обречены на гибель. К примеру: амитоз зародышевых клеток млекопитающих,
амитоз опухолевых клеток. В процессе амитоза ядро и ядерная оболочка остаются неизменными. Более того ядро не разделяется, а перешнуровывается или в нем образуется перегородка, вместо
веретена деления. Отсутствует удвоение количества ДНК, вследствие чего клетки оказываются наследственно неполноценными, не наблюдается спирализация хроматина, не обнаруживаются
хромосомы. При амитозе клетка остается функционально активной, однако деление клетки распределяет наследственный материал в хаотичном порядке.
При амитозе отсутствует цитокинез, что ведет к образованию двуядерных клеток. Образовавшиеся клетки лишены, способности вступать в нормальный митотический цикл. Если амитоз происходит
постоянно, то вполне допустимо появление многоядерных клеток.
Изучение амитоза осложняется еще и неточностью его определения, так как не каждая перетяжка, образовавшаяся в ядре, символизирует начало данного процесса. Наблюдаются случаи, когда
характерные перетяжки ядра являются результатом неправильно протекающего митоза.
Амитоз также наблюдается у одноклеточных организмов, а также в клетках растений и животных, у которых появляется ослабленная физиологическая активность, с признаками отклонений.
На самом деле, амитоз не был исследован на современном уровне. При этом многие ученые и биологи оспаривают взгляд на то, что амитоз является особой формой деления клеток. Правильнее
было бы судить об амитозе как о внутриклеточной регулятивной реакции.
К-МИТОЗ Mитоз, заторможенный (или остановленный) на стадии метафазы в результате инактивации веретена (вероятно, за счет ацетилирования входящего в него тубулина <acetylatedtubulin>) под
действием колхицина или аналогичного по действию вещества; при неполной блокировке деления (частичный С-м.) возможность правильного анафазного расхождения хромосом все равно
оказывается нарушенной, в результате чего образуются многополюсные фигуры.
ЭНДОМИТОЗ(отэндо... и митоз), удвоение числа хромосом внутри ядерной оболочки без разрушения ядрышка и без образования веретена деления клетки. Ранее эндомитоз считали способом
образования полиплоидных и политенных ядер, однако позднее было установлено, что полиплоидия — результат неоконченных обычных митозов, а политения — повторяющихся циклов синтеза
ДНК без митоза. Распространённость и механизм эндомитоза пока не ясны.
ПОЛИТЕНИЯ (от поли. . . и лат. taenia — повязка, лента), образование в ядре соматич. клеток нек-рых двукрылых, простейших и растений гигантских многонитчатых (политенных) хромосом,
превышающих по размерам в сотни раз обычные. За счёт многократной репликации исходной хромосомы без последующего её расхождения число хромонем (иногда св. 1000) и кол-во ДНК
увеличиваются, что и приводит к увеличению диаметра и длины хромосом. П. впервые описана Э. Бальбиани в 1881. Благодаря неравномерной спирализации в политенных хромосомах образуются
диски (тёмные поперечные полосы), выявляемые при окраске хромосом. Число, размер и характер расположения дисков специфичны для вида. П. используют для построения карт хромосом,
обнаружения хромосомных перестроек; сравнение цитологич. карт политенных хромосом позволяет определять видовую принадлежность особей разных популяций и способствует пониманию
процессов микроэволюции и видообразования.
Прямое деление клеток, или амитоз, представляет собой процесс деления клеток, которое происходит без образования хромосом и спиндельного аппарата. В результате процесса амитоза
образуется две дочерние клетки с одинаковым генетическим материалом.
К-митоз - процесс деления клеток, который происходит с образованием спиндельного аппарата и хромосом. Этот процесс протекает в несколько этапов, включая профазу, метафазу, анафазу и
телофазу, и в результате образуются две дочерние клетки с одинаковым генетическим материалом.
Эндомитоз - тип деления клеток, который включает в себя процесс образования новых ядер внутри клетки без деления цитоплазмы. Этот процесс может привести к образованию многоядерных
клеток.
Политения - это процесс деления клеток, который включает в себя многократное деление ядра без последующего деления цитоплазмы. В результате этого процесса образуются клетки с большим
числом ядер.
34.
28) Мейоз, его биологическое значение и цитологическая и цитогенетическаяхарактеристики: редукция числа хромосом, конъюгация, кроссинговер, случайное
расхождение хромосом в дочерние клетки
.
Мейоз – разновидность полового размножения, в результате которого из диплоидных соматических клеток половых желез образуются
гаплоидные клетки. При оплодотворении ядва гамет сливаются и восстанавливается диплоидный набор хромосом.
Мейоз обеспечивает сохранение постоянного для каждого вида набора хромосом и количества ДНК.
Мейоз состоит из 2х последовательных делений – мейоза 1 и мейоза 2. В каждом делении выделают профазу, метафазу, анафазу и телофазу. В
результате число хромосом уменьшается вдвое (редукционное деление); при мейозе 2 гаплоидность клеток сохраняется (эквационное деление).
Клетки, вступающие в мейоз, содержат генетическую информацию 2n2xp.
В профазе мейоза 1 происходит постепенная спирализация хроматина с образованием хромосом. Гомологичные хромосомы сближаются,
образуя общую структуру, состоящую из 2х хромосом (бивалент) и 4х хроматид (тетрада). Соприкосновение 2х гомологичных хромосом по всей
длине называется конъюгацией. Затем между гомологичными хромосомами появляются силы отталкивания и хромосомы сначала разделяются в
области центромер, оставаясь соединенными в области плеч, и образуют перекресты (хиазмы). Расхождение хроматид постепенно
увеличивается и перекресты смещаются к их концам. В процессе конъюгации между некоторыми хроматидами гомологичных хромосом может
происходить обмен участками – кроссинговер, приводящий к перекомбинации генетического материала. К концу профазы растворяются ядерная
оболочка и ядрышки, формируется ахроматиновое веретено деления. Содержание генетического материала остается прежним (2n2xp).
В метафазе мейоза 1 биваленты хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация их достигает
максимума. Содержание генетического материала не изменяется.
В анафазе мейоза 1 гомологичные хромосомы, состоящие из 2х хроматид, окончательно отходят друг от друга и расходятся к полюсам клетки.
Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна – число хромосом уменьшается вдвое
(происходит редукция). Содержание генетического материала становится 1n2xp у каждого полюса.
В телофазе происходит формирование ядер и разделение цитоплазмы – образуются 2 дочерние клетки. Дочерние клетки содержат гаплоидный
набор хромосом, каждая хромосома – 2 хроматиды (1n2xp).
Интеркинез – короткий промежуток между первым и вторым мейотическими делениями. В это время не происходит репликация ДНК, и 2
дочерние клетки быстро вступает в мейоз 2, протекающий по типу митоза.
В профазе меоза 2 происходит незначительная спирализация хромосом (в интеркинезе они не успевают деспирализоваться). В метафазе
хромосомы располагаются в экваториальной плоскости. Изменений содержания генетического материала не происходит. В анафазе мейоза 2
хроматиды каждой хромосомы отходят к противоположным полюсам клетки и содержание генетического материала у каждого полюса
становится 1n1xp. В телофазе образуются 4 гаплоидные клетки (1n1xp).
35.
29) Бесполое размножение, его виды и биологическое значениеРазмножение – универсальное свойство всех живых организмов, способность воспроизводить себе подобных. С его помощью происходит сохранение во времени
видов и жизни в целом. Оно обеспечивает смену поколений. Жизнь клеток, составляющих организм, намного короче жизни самого организма, поэтому его
существование поддерживается только за счет размножения клеток. Различают два способа размножения – бесполое и половое. При бесполом размножении главным
клеточным механизмом, обеспечивающим увеличение числа клеток, является митоз. Родителем является одна особь. Потомство представляет собой точную
генетическую копию родительского материала.
1. Биологическая роль бесполого размножения
Поддержание наибольшей приспособленности в малоизменяю-щихся условиях окружающей среды. Оно усиливает значение стабилизирующего естественного отбора;
обеспечивает быстрые темпы размножения; используется в практической селекции. Бесполое размножение встречается как у одно-, так и у многоклеточных
организмов. У одноклеточных эукариот бесполое размножение представляет собой митотическое деление, у прокариот – деление ну-клеоида, у многоклеточных форм
– вегетативное размножение.
2. Формы бесполого размножения
У одноклеточных организмов выделяют следующие формы бесполого размножения: деление, эндогонию, шизогонию (множественное деление) и почкование,
спорообразование.
Деление характерно для таких одноклеточных, как амебы, инфузории, жгутиковые. Сначала происходит митотическое деление ядра, затем цитоплазма делится
пополам все более углубляющейся перетяжкой. При этом дочерние клетки получают примерно одинаковое количество цитоплазмы и органоидов.
Эндогония (внутреннее почкование) характерно для токсо-плазмы. При образовании двух дочерних особей материнская дает лишь двух потомков. Но может быть
внутреннее множественное почкование, что приведет к шизогонии.
Шизогония развивается на основе предыдущей формы. Встречается у споровиков (малярийного плазмодия) и др. Происходит многократное деление ядра без
цитокинеза. Затем вся цитоплазма разделяется на части, которые обособляются вокруг новых ядер. Из одной клетки образуется очень много дочерних.
Почкование (у бактерий, дрожжевых грибов и др.). При этом на материнской клетке первоначально образуется небольшой бугорок, содержащий дочернее ядро
(нуклеоид). Почка растет, достигает размеров материнской особи, а затем отделяется от нее.
Спорообразование (у высших споровых растений: мхов, папоротников, плаунов, хвощей, водорослей). Дочерний организм развивается из специализированных клеток –
спор, содержащих гаплоидный набор хромосом. В царстве бактерий тоже встречается спорообразование. Cпоры, покрытые плотной оболочкой, защищающей ее от
неблагоприятных воздействий окружающей среды, не способ размножения, а способ переживания неблагоприятных условий.
3. Вегетативная форма размножения
Характерна для многоклеточных организмов. При этом новый организм образуется из группы клеток, отделяющихся от материнского организма. Растения
размножаются клубнями, корневищами, луковицами, корнеклубнями, корнеплодами, корневой порослью, отводками, черенками, выводковыми почками, листьями. У
животных вегетативное размножение встречается у самых низкоорганизованных форм. У губок и гидр оно идет путем почкования. За счет размножения группы клеток
на материнском теле образуется выпячивание (почка), состоящее из клеток экто– и эндодермы. Почка постепенно увеличивается, на ней возникают щупальца, и
отделяется от материнского организма. Ресничные черви делятся на две части, и в каждой из них восстанавливаются недостающие органы за счет неупорядоченного
деления клеток. Кольчатые черви могут восстанавливать целый организм из одного членика. Этот вид деления лежит в основе регенерации – восстановления
утраченных тканей и частей тела (у кольчатых червей, ящериц, саламандр). Особая форма бесполого размножения – стробиляция (у полипов). Полипоид-ный организм
довольно интенсивно растет, при достижении определенных размеров начинает делиться на дочерние особи. В это время он напоминает стопку тарелок.
Образовавшиеся медузы отрываются и начинают самостоятельную жизнь.
36.
30) Биологическое значение и сущность полового размножения, его виды.Половое размножение – процесс, в котором объединяется генетическая информация от двух особей.
Объединение генетической информации может происходить при конъюгации (временном соединении
особей для обмена информацией, как это происходит у инфузорий) и копуляции (слиянии особей для
оплодотворения) у одноклеточных животных, а также при оплодотворении у представителей разных царств.
Особым случаем полового размножения является партеногенез у некоторых животных (тли, трутни пчел). В
этом случае новый организм развивается из неоплодотворенного яйца, но до этого всегда происходит
образование гамет.
Половое размножение у покрытосеменных растений происходит путем двойного оплодотворения. Дело в
том, что в пыльнике цветка образуются гаплоидные пыльцевые зерна. Ядра этих зерен делятся на два –
генеративное и вегетативное. Попав на рыльце пестика, пыльцевое зерно прорастает, образуя пыльцевую
трубку. Генеративное ядро делится еще раз, образуя два спермия. Один из них, проникая в завязь,
оплодотворяет яйцеклетку, а другой сливается с двумя полярными ядрами двух центральных клеток
зародыша, образуя триплоидный эндосперм.
При половом размножении особи разного пола образуют гаметы. Женские особи производят яйцеклетки,
мужские – сперматозоиды, обоеполые особи (гермафродиты) производят и яйцеклетки, и сперматозоиды. У
большинства водорослей сливаются две одинаковых половых клетки. При слиянии гаплоидных гамет
происходит оплодотворение и образование диплоидной зиготы. Зигота развивается в новую особь.
Все вышеперечисленное справедливо только для эукариот. У прокариот тоже есть половое размножение, но
происходит оно по-другому.
Таким образом, при половом размножении происходит смешивание геномов двух разных особей одного
вида. Потомство несет новые генетические комбинации, что отличает их от родителей и друг от друга.
Различные комбинации генов, проявляющиеся в потомстве в виде новых, интересующих человека признаках,
отбираются селекционерами для выведения новых пород животных или сортов растений. В некоторых
случаях применяют искусственное оплодотворение. Это делается и для того, чтобы получить потомство с
заданными свойствами, и для того, чтобы преодолеть бездетность некоторых женщин.
37.
31) Нерегулярные типы полового размножения.У животных и растений встречаются так называемые нерегу¬лярные типы полового размножения. Это прежде всего апомиксис (от греч. «апо» — без, «миксис» —
смешение), т.е. половое раз¬множение без оплодотворения. Апомиксис противоположен амфи¬миксису («амфи» — разделенный), т. е. половому размножению,
происходящему путем слияния разнокачественных гамет. Синоним апомиксиса — партеногенез, т. е. девственное размножение от греч. «партенос» — девственница).
Термин апомиксис чаще упот¬ребляют в отношении растений, а партеногенез — в отношении животных.
Партеногенез может быть гаплоидным и диплоидным. При гаплоидном, или генеративном, партеногенезе новый организм раз¬вивается без оплодотворения, из
гаплоидной яйцеклетки. Разви¬вающиеся при этом особи могут быть только мужскими, только женскими или теми и другими. Это зависит от хромосомного механизма
определения пола. Например, у пчел, паразитических ос, червецов, клещей самцы появляются в результате партеноге¬неза. Партеногенез может быть постоянным
(облигатным), как в упомянутых случаях, или циклическим (факультативным). У дафний, тлей, коловраток партеногенетические поколения чере¬дуются с половыми. У
дафний, в частности, самки диплоидны, а самцы гаплоидны. В благоприятных условиях у дафний не про¬исходит мейоза. Яйцеклетки диплоидны. Они развиваются без
оплодотворения и дают начало только самкам. Это пример дипло¬идного, или соматического, партеногенеза. В неблагоприятных условиях (понижение температуры,
нехватка корма) самки начи¬нают откладывать гаплоидные яйца, из которых выводятся самцы. В результате полового процесса образуются диплоидные зиготы, вновь
дающие начало самкам.
Наряду с партеногенезом наблюдается и развитие яйцеклетки, активируемое сперматозоидом, не участвующим в оплодотворении. Мужской пронуклеус погибает, а
организм развивается за счет женского пронуклеуса. Это явление называется гиногенезом, который встречается у гермафродитных круглых червей и у неко¬торых рыб.
Противоположность гиногенеза — андрогенез — развитие толь¬ко за счет мужского пронуклеуса в случае гибели женского пронук¬леуса. Гаплоидный андрогенез
встречается очень редко. Развитие андрогенных особей до взрослого состояния наблюдали только у наездника Habrobracon и у тутового шелкопряда.
У тутового шелкопряда при оплодотворении в яйцеклетку про¬никает несколько сперматозоидов, но ядро лишь одного из них сливается с ядром яйцеклетки, остальные
погибают. Если неопло¬дотворенные яйцеклетки активировать температурным шоком, как это описано выше, и облучить рентгеновскими лучами, то ядро яйцеклетки
погибнет. Если далее такие энуклеированные яйца осеменить, то два мужских пронуклеуса, проникшие в яйцеклетку, сливаются между собой. За счет образовавшегося
диплоидного ядра развивается зигота. Как показал Б. JI. Астауров, такие андро- генетические зиготы всегда превращаются в самцов, поскольку они несут две
одинаковые половые хромосомы — ZZ. Получение чисто мужского потомства у шелкопряда экономически выгодно, так как самцы продуктивнее самок.
Гиногенез — частный случай партеногенеза, особая форма полового размножения, при которой после проникновения спермия в яйцеклетку их ядра не сливаются, и в
последующем развитии участвует только ядро яйцеклетки, либо не происходит оплодотворения. При этом нет объединения наследственного материала родителей
посредством слияния ядер их половых клеток. Роль сперматозоида ограничивается активацией осеменённого яйца к развитию. В природе гиногенез встречается крайне
редко. Известен у нескольких видов рыб (голомянка, серебряный карась и др.), земноводных, круглых червей и растения семейства амариллисовых (Atamosco
mexicana).
Андрогене́з — развитие яйцеклетки с мужским ядром, привнесённым в неё спермием в процессе оплодотворения. Андрогенез наблюдается у отдельных видов
животных (шелкопряд) и растений (табак, кукуруза) в тех случаях, когда материнское ядро погибает до оплодотворения, которое при этом является ложным, то есть
женское и мужское ядра не сливаются (Псевдогамия) и в дроблении участвует только мужское ядро. Андрогенез — особый случай девственного развития, или
партеногенеза; иногда его называют «мужской партеногенез».
Апомиксис – партеногенез у растений (одуванчик).
38.
32) Биологические аспекты репродукции человека.· развитие человека: прямое, внутриутробное
· в женском организме формируется плацента
· половая зрелость наступает к 15-18 годам
· плодовитость невысока, редкая смена поколений
· наблюдается многоплодие, созревает несколько яйцеклеток
· полиэмбриония, однояйцовые близнецы формируются в результате
расхождения бластомеров размеры сперматозоидов: 20 мкм в длину и 2 мкм
в толщину, в сутки формируется более 120 млн., для нормальной
репродукции более 50% должно быть без аномалий. Аномалии вызываются
повреждающими факторами от высокой температуры (плотная одежда).
· яйцеклетка, диаметр 100-150 мкм, окружена 3 оболочками
· овогенез начинается на 3 мес. эмбрионального развития.
39.
33) Половой диморфизм: генетический, морфофизиологический, эндокринный и поведенческий аспекты.Под половым диморфизмом понимают подразделение людей на лиц женского и мужского пола (мужчин и женщин). Наличие в природе полового диморфизма вообще отражает различия в задачах, решаемых в процессе полового
размножения мужской и женской особью. У человека с появлением культуры половой диморфизм стал проявляться и в разделении труда, или вернее экологических функций в популяции (добывание пищи, рождение и воспитание потомства,
приготовление пищи, постройка жилья и так далее). В силу биологических особенностей мужчина был более приобщен к поддержанию эколого-экономического благополучия семьи и общины. Женщине достался примат воспроизводства
популяции, отсюда её ведущая роль в биологическом существовании человека. Лишь в последнее время возникли тенденции стирания социальных (но не биологических) различий между мужчиной и женщиной.
На организменном уровне половой диморфизм проявляется в половых признаках. Выделяют первичные половые признаки и вторичные. К первичным половым признакам относят внутренние половые органы (половые железы (семенники и
яичники) вместе с проводящими путями (семяпроводы и яйцепроводы), маткой) и внешние половые органы. Формирование половых морфологических и функциональных половых признаков определяется наличием в кариотипе данной особи
в 23 паре хромосом X- или У-хромосомы. Особи, имеющие кариотип ХУ, развиваются по мужскому типу и у них формируются мужские половые признаки. Особи, имеющие кариотип XX, развиваются по женскому типу.
Считается, что половые признаки начинают формироваться на 7 неделе с момента оплодотворения, когда под воздействием генов У-хромосомы ранее не дифференцированная гонада начинает превращаться в яичко. Роль гормонов в этом
процессе пока не известна. На 9 неделе в яичке появляются клетки Лейдига, которые с 10 недели начинают продуцировать мужской половой гормон тестостерон. Под действием этого гормона ранее недифференцированные наружные половые
органы превращаются в пенис и мошонку.
У женщин дифференцировка яичника и наружных половых органов происходит, по-видимому, не столь бурно. В отсутствие У-хромосомы на 7 неделе ничего не происходит, а на 8 неделе гонада превращается в яичник. Формирование наружных
половых органов по женскому типу происходит примерно на 12 неделе, очевидно, без участия гормонов.
Известен ещё один аспект половой дифференцировки, изученный пока только на животных, но существование которого нельзя исключить и у человека. Установлено, что секреция меток Лейдига воздействует не только на половые органы, но и
на мозг. Происходит дифференциация структуры гипоталамуса, от которой у всех изученных млекопитающих зависят гормональные и в значительной степени поведенческие половые признаки. Недостаток соответствующего полового гормона
или отсутствие в гипоталамусе рецепторов к нему могут привести к формированию морфологических или поведенческих признаков, более характерных для противоположного пола.
Окончательное формирование признаков, характерных для данного вида происходит к концу пубертатного периода. В это время устанавливаются ритмы и уровни секреции половых гормонов – андрогенов (тестостерона) у мужчин и эстрогена и
прогестерона у женщин. Под влиянием этих гормонов формируются все вторичные половые признаки.
Из них наиболее важными являются следующие.
Особенности строения скелета – высокий рост, массивные кости и узкий таз у мужчин, низкий рост, тонкие кости, широкий таз у женщин.
Мышцы у мужчин развиты сильнее, что обеспечивает большую физическую силу. У женщин мышцы развиты в меньшей степени.
Половой диморфизм проявляется также в разном характере распределения жировой ткани у мужчин и женщин. У женщин жировая ткань наиболее выражена в области ягодиц, бедер, нижней части живота. В результате этого типичная женская
фигура имеет форму груши. Жировая ткань у женщин в норме составляет около 25% от массы тела. У женщин, имеющих слабовыраженную жировую прослойку, значительно чаще наблюдаются бесплодие и невынашивание беременности. Это
объясняется некоторыми авторами выработкой в жировой ткани веществ, похожих по действию на гормон второй половины менструального цикла и беременности – прогестерон.
У мужчин жировая ткань обычно откладывается в области верхней части туловища и на животе (по типу «яблока»). Такое отложение связывают с действием мужских половых гормонов.
5. Под действием мужских половых гормонов у мальчиков в пубертатном периоде увеличивается щитовидный хрящ гортани. В результате происходит перестройка голосового аппарата, и голос приобретает низкий тембр (мутация голоса). Так
как у женщин нет мужских половых гормонов, у них голос остается высоким.
6. У женщин во время полового созревания под влиянием женских половых гормонов развиваются молочные железы.
7. Половые гормоны определяют структуру волос и характер оволосения у людей. У мужчин волосы более толстые и жесткие, характерен их рост на лице. Кроме того, у мужчин сильнее, чем у женщин выражено оволосение туловища (хотя
имеются большие вариации в разных этнических группах). С действием мужских половых гормонов связывают и облысение.
У женщин волосы тоньше, обычно более мягкие, чем у мужчин данной этнической группы. Меньше выражено и оволосение туловища и конечностей.
8. К проявлениям полового диморфизма относятся и особенности роста и развития у мальчиков и девочек, разная продолжительность жизни и другие особенности, о которых уже говорилось в соответствующих разделах (см. Разд.
«Закономерности протекания онтогенеза у человека», «Видовая продолжительность жизни человека»)
Генетический аспект: У человека половой диморфизм проявляется в наличии хромосом. Женщины обычно имеют два одинаковых Х-хромосомы (46,XX), в то время как мужчины имеют один Х-хромосому и одну Y-хромосому (46,XY).
Морфофизиологический аспект: : Морфологические различия включают в себя анатомические особенности, такие как различия в теле, лице, голо.се, размере и форме скелета, росте волос и другие физические характеристики.
Эндокринный аспект: Половые гормоны (эстрогены у женщин и тестостерон у мужчин) играют важную роль в формировании и поддержании половых характеристик и функций организма.
Поведенческий аспект: В разных культурах могут существовать социокультурные различия в поведении мужчин и женщин, включая предпочтения, интересы, стереотипы и роли, которые также могут быть связаны с половыми характеристиками.
40.
34) Морфологическое строение хромосом. Кариотип.Морфология хромосом лучше всего видна в клетке на стадии метафазы. Хромосома состоит из двух палочкообразных телец - хроматид. Обе хроматиды каждой хромосомы идентичны друг другу по
генному составу.
Хромосомы дифференцированы по длине. Хромосомы имеют центромеру или первичную перетяжку, две теломеры и два плеча. На некоторых хромосомах выделяют вторичные перетяжки и
спутники. Движение хромосомы определяет Центромера, которая имеет сложное строение.
ДНК центромеры отличается характерной последовательностью нуклеотидов и специфическими белками. В зависимости от расположения центромеры различают акроцентрические,
субметацентрические и метацентрические хромосомы. Как говорилось выше, некоторые хромосомы имеют вторичные перетяжки. Они, в отличие от первичной перетяжки (центромеры), не служат
местом прикрепления нитей веретена и не играют никакой роли в движении хромосом. Некоторые вторичные перетяжки связаны с образованием ядрышек, в этом случае их называют ядрышковыми
организаторами. В ядрышковых организаторах расположены гены, ответственные за синтез РНК. Функция других вторичных перетяжек еще не ясна.
У некоторых акроцентрических хромосом есть спутники — участки, соединенные с остальной частью хромосомы тонкой нитью хроматина. Форма и размеры спутника постоянны для данной
хромосомы. У человека спутники имеются у пяти пар хромосом. Концевые участки хромосом, богатые структурным гетерохроматином, называются теломерами. Теломеры препятствуют слипанию
концов хромосом после редупликации и тем самым способствуют сохранению их целостности. Следовательно, теломеры ответственны за существование хромосом как индивидуальных
образований. Хромосомы, имеющие одинаковый порядок генов, называют гомологичными. Они имеют одинаковое строение (длина, расположение центромеры и т. д.). Негомологичные хромосомы
имеют разный генный набор и разное строение.
Исследование тонкой структуры хромосом показало, что они состоят из ДНК, белка и небольшого количества РНК. Молекула ДНК несет отрицательные заряды, распределенные по всей длине, а
присоединенные к ней белки — гистоны заряжены положительно. Этот комплекс ДНК с белком называют хроматином. Хроматин может иметь разную степень конденсации. Конденсированный
хроматин называют гетерохроматином, деконденсированный хроматин — эухроматином. Степень деконденсации хроматина отражает его функциональное состояние. Гетерохроматиновые участки
функционально менее активны, чем эухроматиновые, в которых локализована большая часть генов. Различают структурный гетерохроматин, количество, которого различается в разных хромосомах,
но располагается он постоянно в околоцентромерных районах. Кроме структурного гетерохроматина существует факультативный гетерохроматин, который появляется в хромосоме при
сверхспирализации эухроматических районов. Подтверждением существования этого явления в хромосомах человека служит факт генетической инактивации одной Х-хромосомы в соматических
клетках женщины. Его суть заключается в том, что существует эволюционно сформировавшийся механизм инактивации второй дозы генов, локализованных в Х-хромосоме, вследствие чего, несмотря
на разное число Х-хромосом в мужском и женском организмах, число функционирующих в них генов уравнено. Максимально конденсирован хроматин во время митотического деления клеток, тогда
его можно обнаружить в виде плотных хромосом
Размеры молекул ДНК хромосом огромны. Каждая хромосома представлена одной молекулой ДНК. Они могут достигать сотен микрометров и даже сантиметров. Из хромосом человека самая
большая — первая; ее ДНК имеет общую длину до 7 см. Суммарная длина молекул ДНК всех хромосом одной клетки человека составляет 170 см.
Несмотря на гигантские размеры молекул ДНК, она достаточно плотно упакована в хромосомах. Такую специфическую укладку хромосомной ДНК обеспечивают белки гистоны. Гистоны
располагаются по длине молекулы ДНК в виде блоков. В один блок входит 8 молекул гистонов, образуя нуклеосому (образование, состоящее из нити ДНК, намотанной вокруг октамера гистонов).
Размер нуклеосомы около 10 нм. Нуклеосомы имеют вид нанизанных на нитку бусинок. Нуклеосомы и соединяющие их участки ДНК плотно упакованы в виде спирали, на каждый виток такой
спирали приходится шесть нуклеосом. Так формируется структура хромосомы.
Наследственная информация организма строго упорядочена по отдельным хромосомам. Каждый организм характеризуется определенным набором хромосом (число, размеры и структура), который
называется кариотипом. Кариотип человека представлен двадцатью четырьмя разными хромосомами (22 пары аутосом, Х- и Y-хромосомы). Кариотип — это паспорт вида. Анализ кариотипа
позволяет выявлять нарушения, которые могут приводить к аномалиям развития, наследственным болезням или гибели плодов и эмбрионов на ранних стадиях развития. Длительное время
полагали, что кариотип человека состоит из 48 хромосом. Однако в начале 1956 г. было опубликовано сообщение, согласно которому число хромосом в кариотипе человека равно 46.
Хромосомы человека различаются по размеру, расположению центромеры и вторичных перетяжек. Впервые подразделение кариотипа на группы было проведено в 1960 г. на конференции в г.
Денвере (США). В описание кариотипа человека первоначально были заложены два следующих принципа: расположение хромосом по их длине; группировка хромосом по расположению
центромеры (метацентрические, субметацентрические, акроцентрические).
Точное постоянство числа хромосом, их индивидуальность и сложность строения свидетельствуют о важности выполняемой ими функции. Хромосомы выполняют функцию основного генетического
аппарата клетки. В них в линейном порядке расположены гены, каждый из которых занимает строго определенное место (локус) в хромосоме. В каждой хромосоме много генов, но для нормального
развития организма необходим набор генов полного хромосомного набора.
Кариотип — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данного биологического вида (видовой кариотип), данного организма
(индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы).
41.
35) Генетическая сущность полового размножения. Образование половых клеток (гаметогенез).Оплодотворение.
При половом размножении материнским и отцовским организмами вырабатываются специализированные половые клетки — гаметы. Женские неподвижные гаметы называют яйцеклетками,
мужские неподвижные — спермиями, а подвижные — сперматозоидами. Эти половые клетки сливаются с образованием зиготы, то есть происходит оплодотворение. Половые клетки, как правило,
имеют половинный набор хромосом (гаплоидный), так что при их слиянии восстанавливается двойной (диплоидный) набор, из зиготы развивается новая особь. Если суть и биологическое значение
полового процесса едины для всех организмов, то его формы очень разнообразны и зависят от уровня эволюционного развития, среды обитания, образа жизни и некоторых других особенностей.
Возникновение примитивного полового процесса произошло еще у простейших животных, и многоклеточных нитчатых водорослей. Например, у хламидомонады при неблагоприятных условиях в
материнской клетке образуются двужгутиковые гаметы. Они выходят из оболочки материнской клетки и сливаются попарно с гаметами других особей, в результате чего образуется зигота, покрытая
плотной оболочкой. При улучшении условий из зиготы образуется несколько молодых хламидомонад.
Гаметогенез (образование половых клеток). Зрелые сперматозоид и яйцеклетка различаются по своему строению, похожи у них только ядра; тем не менее, и мужские, и женские гаметы (развитые
половые клетки) образуются из одинаковых на вид первичных половых клеток. У всех организмов, размножающихся половым путем, эти первичные половые клетки на ранних стадиях развития
отделяются от других клеток и развиваются особым образом, готовясь к выполнению своей функции – продуцированию предназначенных для образования «детей» половых, или зародышевых,
клеток. Поэтому такие первичные половые клетки называют зародышевой плазмой – в отличие от всех других клеток, составляющих соматоплазму, то есть «материал» для построения тела самого
организма, а не его будущих потомков. Однако понятно, что и зародышевая плазма, и соматоплазма происходят из оплодотворенного яйца – зиготы, из которой начал развиваться новый организм.
То есть изначально все клетки одинаковы. Что, в конечном итоге, определяет, какие клетки станут половыми, а какие – соматическими, до сих пор не известно. Как бы то ни было, в конечном итоге
половые клетки приобретают достаточно четкие отличия, возникающие в процессе гаметогенеза.
Оогенез и оплодотворение (на примере человека)
Оогенез включает 3 периода (фазы): размножения, роста и созревания. Начинается оогенез на 2…3 месяце развития зародыша женского пола, а завершается для каждой данной яйцеклетки только
после ее оплодотворения.
В фазе размножения диплоидные оогонии делятся несколько раз путем митоза. Образовавшиеся клетки вступают в фазу роста, увеличиваются в размерах и превращаются в диплоидные
ооцитыпервогопорядка (ооциты I). Ооциты I находятся на поверхности яичников в пузырьках–фолликулах. (Всего образуется около 2 миллионов таких фолликулов, но лишь около 450 из них
завершает свое развитие.)
В фазе созревания в ооцитах I начинается мейоз. Однако первое деление мейоза блокируется во время рождения девочки на стадии метафазы I вплоть до полового созревания.
После завершения полового созревания примерно каждый месяц один ооцит I увеличивается в размерах (такая крупная клетка (независимо от уровня плоидности) обычно называется яйцом на всех
дальнейших стадиях оогенеза) и окружается слоем фолликулярных клеток. Фолликул увеличивается в размерах до 1 см и превращается в Граафов пузырек, который выступает на поверхности
яичника в виде бугорка. Внутри Граафова пузырька ооцит I окружен рядом оболочек: плазматической мембраной, затем желточной оболочкой (или прозрачной оболочкой) и слоем фолликулярных
клеток, образующих лучистый венец (яйценосный бугорок).
Перед овуляцией ооцит I завершает первое деление мейоза. В результате образуется две гаплоидные клетки (1n): ооцитвторогопорядка (ооцит II) и первое полярное тельце. При овуляции Граафов
пузырек лопается, и ооцит II, окруженный фолликулярными клетками вместе с первым полярным тельцем, выходит в брюшную полость и попадает в фаллопиеву трубу. (На месте лопнувшего
Граафова пузырька образуется желтое тело.)
Затем начинается второе деление мейоза, доходит до стадии метафазы II, но не продолжается до тех пор, пока ооцит второго порядка не сольется со сперматозоидом. Проникновение сперматозоида
в ооцит II служит стимулом для завершения второго деления мейоза. После окончания мейоза ооцит II образует крупную оплодотворенную яйцеклетку и второе полярное тельце. Параллельно
происходит деление первого полярного тельца. Полярные тельца дегенерируют, а все структуры сперматозоида, кроме ядра, рассасываются.
Ядра обеих гамет превращаются в пронуклеусы и сближаются. Слияние пронуклеусов – кариогамия – происходит позднее. На этой стадии оплодотворенное яйцо получает название зиготы. Зигота
вступает в первый митоз, в ходе которого происходит объединение женского и мужского хромосомных наборов.
42.
36) Менделирующие признаки человека.Признаки, наследование к-рых подчиняется перечисленным закономерностям, принято
называть менделирующими(по имени Г. Менделя).
У человека менделирующими признаками являются, напр. альбинизм (отсутствие
пигментации, вызываемое рецессивным геном; встречается у всех человеческих рас с
частотой 1 на 20— 30 тыс. новорожденных), цвет глаз, характер волос (курчавые или
гладкие), групповые отличия по различным факторам в крови (см. Группы крови) и др.
Законам Менделя подчиняются и гены, обусловливающие наследственные болезни
человека.
Мультифакторные признаки формируются при сложном взаимодействии большого числа
различных генов и факторов среды, особенностей жизнендеятельности.
Мультифакторные признаки очень сложные, к ним можно отнести различные
интеллектуальные особенности: внимание, память, речь. В 1865 г. английский ученый Ф.
Гальтон разработал подход для оценки мультифакторных признаков, который был назван
биометрическим. В соответствии с эти подходом заключение о наследование такого
признака основано на его сравнительной количественной оценке у разных членов семьи
(у родителей, детей, внуков) с использованием статистических методов. Количественный
принцип оценки мультифакторных признаков, введенный Ф. Гальтоном, не потерял
актуальности и по сей день.
43.
37) Наследование признаков при полном и неполном доминировании и кодоминировании.Наследование признаков при полном доминировании:
В случае полного доминирования один из родителей полностью определяет признак потомка.
Потомок наследует только признаки, которые присутствуют в доминирующем родителе.
При этом, признаки, которые отсутствуют у доминирующего родителя, не будут наследоваться.
Наследование признаков при неполном доминировании:
В случае неполном доминировании ни один из родителей не полностью определяет признак потомка.
Потомок наследует признаки от обоих родителей.
Если признак представлен в обоих родителях, может возникнуть конфликт и два разных значения признака могут быть унаследованы потомком.
Наследование признаков при кодоминировании:
В случае кодоминирования два родителя полностью влияют на признак потомка.
Потомок наследует признаки от обоих родителей.
Если признак представлен в обоих родителях, значения двух родителей могут быть комбинированы или конкатенированы для создания нового значения признака у потомка.
Полное доминирование — это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не отличается от фенотипа гомозигот по доминанте, то есть в фенотипе гетерозигот
присутствует продукт доминантного гена. Полное доминирование широко распространено в природе, имеет место при наследовании, например, окраски и формы семян гороха, цвета глаз и цвета
волос у человека, резус-антигена и мн. др.
Наличие резус-антигена (резус-фактора) эритроцитов обусловливается доминантным геном Rh. То есть генотип резус-положительного человека может быть двух видов: или RhRh, или Rhrh; генотип
резус-отрицательного человека — rhrh. Если, например, мать — резус-отрицательная, а отец резус-положительный и гетерозиготен по этому признаку, то при данном типе брака с одинаковой
вероятностью может родиться как резус-положительный, так и резус-отрицательный ребенок.
Между резус-положительным плодом и резус-отрицательной матерью может возникнуть резус-конфликт.
Неполное доминирование
Так называется вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву и имеет
среднее (промежуточное) значение между ними. Имеет место при наследовании окраски околоцветника ночной красавицы, львиного зева, окраски шерсти морских свинок и пр.
Сам Мендель столкнулся с неполным доминированием, когда скрещивал крупнолистный сорт гороха с мелколистным. Гибриды первого поколения не повторяли признак ни одного из
родительских растений, они имели листья средней величины.
При скрещивании гомозиготных красноплодных и белоплодных сортов земляники все первое поколение гибридов имеет розовые плоды. При скрещивании этих гибридов друг с другом получаем:
по фенотипу — 1/4 красноплодных, 2/4 розовоплодных и 1/4 белоплодных растений, по генотипу — 1/4 АА, 1/2 Аа, 1/4 аа (и по фенотипу, и по генотипу соотношение 1:2:1). Соответствие
расщепления по генотипу расщеплению по фенотипу является характерным при неполном доминировании, так как гетерозиготы фенотипически отличаются от гомозигот.
Кодоминирование — вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву, и в
фенотипе гетерозигот присутствуют продукты обоих генов. Имеет место при формировании, например, IV группы крови системы (АВ0) у человека.
Для того чтобы представить, как происходит наследование групп крови у человека, можно посмотреть, рождение детей с какой группой крови возможно у родителей, имеющих один — вторую,
другой — третью группы крови и являющихся гетерозиготными по этому признаку.
44.
38) Законы Г. Менделя. Типы и варианты наследования признаков, контролируемых ядерными генами.Законы Г. Менделя описывают основные принципы наследования и были открыты через его исследования растений, в частности гороха, в 19 веке. Эти законы включают
в себя:
1. Закон гомозиготности
2. Закон разделения
3. Закон независимого расщепления
Варианты наследования признаков могут быть контролируемыми ядерными генами и могут описываться разными типами наследования, вот некоторые из них:
А) Кодоминирование: оба аллеля одного гена являются доминантными и оба проявляются в фенотипе гетерозиготного организма.
B) Полное доминирование: один аллель полностью доминирует над другим, и фенотип гетерозигота идентичен фенотипу гомозиготной доминантной особи.
C) Неполное доминирование: гетерозигот имеет промежуточный фенотип между двумя гомозиготными родителями.
Эти принципы и варианты наследования признаков, контролируемых ядерными генами, лежат в основе понимания наследственности и эволюции живых организмов.
Первый закон Менделя (правило единообразия).
Скрещивание двух организмов называют гибридизацией. Моногибридным ( моногенным) называют скрещивание 2 организмов, при котором прослеживают
наследование одной пары альтернативных проявлений какого-либо признака( развитие этого признака обусловлено парой аллелей одной гена).
Рассмотрим наследование цвета семян ( его альтернативное варианты желтый или зеленый- кодируются следующими парами аллелей одного гена: АА – ж, аа – з, Аа–
гибрид .
Гибриды первого поколения оказались единообразными по исследуемому признаку. В F1 проявился лишь один( желтый) из пары альтернативных вариантов признака
цвета семян, названный доминантным. Эти результаты иллюстрируют первый закон Менделя – закон единообразия первого поколения , а также правило
доминирования. Первый закон Менделя: при скрещивании гомозиготных особей, отличающихся одной или несколькими парами альтернативных признаков, все
гибриды первого поколения окажутся по этим признакам единообразными. У гибридов проявятся доминантные признаки родителей.
Во втором поколении ( F2) обнаружилось расщепление по исследуемому признаку.
Решетка Пеннета.
Р: АаXАа
Гаметы: А а и Аа
F2: АА, Аа, Аа, аа.
Появились семена как с желтой, так и с зеленой окраской семядолей. У части гибридов F2 вновь возник признак, не обнаруженный у гибридов в F1. Этот признак (
зеленый) назван рецессивным. Соотношение потомков с доминантным и рецессивным проялением признака оказалось близко к ¾ : ¼ .
45.
38) Законы Г. Менделя. Типы и варианты наследования признаков, контролируемыхядерными генами.
Второй закон Менделя: при моногибридном скрещивании гетерозиготных особей ( гибридов F1) во втором поколении наблюдается расщепление по вариантам анализируемого признакак в
отношении №:1 по фенотипу и 1:2:1 по генотипу.
Гипотеза «Чистота гамет».
Мендель предположил, что каждый наследственный признак зависит от наличия в соматических клетках 2 наследственных факторов, полученных от отца и матери. К настоящему времени
установлено, что наследственные факторы Менделя соответствуют генам – локусам хромосом, поэтому сходство между поведением факторов и поведением хромосом стало вполне понятным.
Гомозиготные растения АА образуют гаметы одного сорта с аллелем А ; растения с зелеными семенами аа образуют гаметы а. В момент оплодотворения происходит слияние половых клеток и
возникают гетерозиготные диплоидные особи Аа, образующие семена с доминантной желтой окраскойю
В F1 во время анафазы 1-ого деления мейоза гомологичные хромосомы с аллелями А и а оходят к разным полюсам клетки и затем попадают в разные гаметы , причем яйцеклеток с аллелем А и с
аллелем а образуется примерно в равном количестве, так же как и спермиев с Аи а. Вероятность оплодотворения яйцеклеток с А и а спермиями с А равна вероятности оплодотворения их спермиями
с а. Таким образом, самоопылении растений F1 количество гибридов F2 с определенным соотношение аллелей А и а зависит от вероятности сочетания гамет с А и а.
Соотношение генотипов особей гибридных поколений можно показать графически при помощи решетки Пеннета. При её составлении гаметы одного из родителей выпитсывают снаружи решетки по
вертикали, гаметы другого – по горизонтали. Возникающие в результате скрещивания гамет зиготы вписываются внутри в клетках на пересечении линий, идущих от соответствующих гамет.
Таким образом, пользуясь современной терминологией , гипотезу « чистоты гамет» можно сформулировать : « В процессе образования половых клеток в каждую гамету попадает только один ген из
аллельной пары» , потому что, в процессе мейоза в гамету попадает одна хромосома и пары гомологичных хромосом.
Третий закон Менделя. Правило независимого наследования.
Скрещивание, при котором прослеживается наследование по двум парам альтернативных признаком, называют дигибридным, по нескольким парам признакам- полигибридным. В опытах Менделя
при скрещивании сорта гороха, имевшего желтые А и гладкие В семена, с сортом гороха с зелеными а и морщинистыми b семенами , гибриды F1 имели желтые и гладкие семена, т.е. проявлялись
доминантные признаки( гибриды единообразны).
Гибридные семена второго поколения F2 распределились на 4 фенотипические группы в соотношении : 315- с гладкими желтыми семенами, 101 – с морщинистыми желтыми , 108- с гладкими
зелеными , 32- с зелеными морщинистыми семенами. Если число потомков в каждой группе разделить на чисто потомков в самой малочисленной группе , то в F2 соотношение фенотипических
классов составит приблизительно 9:3:3:1
При образовании гамет двойная гетерозиготная особь(дигетерозигота) АаВbдаст 4 типа разных половых клеток. Вcпомните, что в анафазе 1-го деления мейоза гомологичные хромосомы каждой пары
расходятся к разным полюсам клетки независимо от других пар гомологичных хромосом. Поэтому члены одной пары аллельных генов распределяется по гаметам независимо от членов других пар. В
данном конкретном случае у аллеля А равные шанcы попасть в гамету с аллелем В или с аллелемb. То же справедливо и для аллеля а. Поэтому каждая дигетерозигота в F1 образует 4 сорта гамет
примерно в одинаковом количественном соотношению
Подчеркнем, что каждая гамета может получить только один из каждой пары. Соответственно независимому распределению аллельных генов каждой пары гомологичных хромосом осуществляется
и независимо наследование обусловленных ими признаков. Любая женская гамета имеет равные шансы быть оплодотворенной любой мужской, и в F2 общее число возможных генотипов равно 9, а
число возможных фенотипов - четырем.
Итак, третий закон Менделя: гены разных аллельных пар и соответствующие им признаки передаются потомству независимо друг от друга, комбинируясь во всевозможных сочетаниях.
46.
39) Возвратное скрещивание, анализирующее скрещивание, их использование в генетике.Возвратное скрещивание и анализирующее скрещивание - это два важных
метода, используемых в генетике для изучения наследственности и
определения генетических свойств организмов.
1. Возвратное скрещивание:
- Этот метод заключается в скрещивании фенотипически проявляющего
признака с беглецом (особью с гомозиготным рецессивным генотипом).
2. Анализирующее скрещивание:
- Это метод, используемый для определения генотипов организмов на
основе фенотипических данных.
Использование возвратного скрещивания и анализирующего скрещивания в
генетике позволяет установить типы наследования, выявить рецессивные
гены, определить генотипы, провести анализ генетических связей и выявить
другие аспекты наследственности.
47.
42) Статистический характер расщепления. Критерий χ2.Статистический характер расщепления определяется вероятностным характером комбинирования в мейозе хромосом, а значит, и заключеннных в них генов. Поэтому законы расщепления для любого числа генов при полигибридном
скрещивании могут быть выведены математически. Действительно, число типов гамет (и число фенотипов) при моногибридном скрещивании равно 2, при дигибридном -4 (2^), а при полигибридном — 2“; число генотипов равно соответственно
3, ЗЧ9) и 3“; расщепление по фенотипу — для моногибридного 3: t, дигибридного — 9: 3: 3: 1 (3: 1)^ и для полигибридного — (3: 1)». Поскольку характер расщепления статистический, полученные экспериментальные данные редко
соответствуют абсолютно точно теоретически ожидаемым, например 3:1 или 9: 3 ; 3 ; 1 и т.д. (см. результаты Менделя). Специальные методы статистической обработки позволяют установить достоверность соответствия практически полученных
результатов теоретически ожидаемым.
Критерий Хи-квадрат позволяет сравнивать распределения частот вне зависимости от того, распределены они нормально или нет. Под частотой понимается количество появлений какого-либо события. Обычно, с частотой появления события
имеют дело, когда переменные измерены в шкале наименований и другой их характеристики, кроме частоты подобрать невозможно или проблематично. Другими словами, когда переменная имеет качественные характеристики. Так же
многие исследователи склонны переводить баллы теста в уровни (высокий, средний, низкий) и строить таблицы распределений баллов, чтобы узнать количество человек по этим уровням. Чтобы доказать, что в одном из уровней (в одной из
категорий) количество человек действительно больше (меньше) так же используется коэффициент Хи-квадрат.
Разберем самый простой пример.
Среди младших подростков был проведён тест для выявления самооценки. Баллы теста были переведены в три уровня: высокий, средний, низкий. Частоты распределились следующим образом:
Высокий (В) 27 чел. Средний (С) 12 чел. Низкий (Н)
11 чел.
Очевидно, что детей с высокой самооценкой большинство, однако это нужно доказать статистически. Для этого используем критерий Хи-квадрат.
Наша задача проверить, отличаются ли полученные эмпирические данные от теоретически равновероятных. Для этого необходимо найти теоретические частоты. В нашем случае, теоретические частоты – это равновероятноые частоты, которые
находятся путём сложения всех частот и деления на количество категорий.
В нашем случае: (В + С + Н)/3 = (27+12+11)/3 = 16,6
Формула для расчета критерия хи-квадрат: Хи-квадрат = ∑(Э - Т)² / Т
Строим таблицу:
Эмпирич. (Э)
Теоретич. (Т)
(Э - Т)² / Т
Высокий
27 чел.
16,6
6.41
Средний
12 чел.
16,6
1,31
Низкий
11 чел.
16,6
1,93
Находим сумму последнего столбца:
Хи-квадрат = 9,64
Теперь нужно найти критическое значение критерия по таблице критических значений. Для этого нам понадобится число степеней свободы (df)
df = (R - 1) * (C - 1), где R – количество строк в таблице, C – количество столбцов.
В нашем случае только один столбец (имеются в виду исходные эмпирические частоты) и три строки (категории), поэтому формула изменяется – исключаем столбцы.
df = (R - 1) = 3-1 = 2
Для вероятности ошибки p≤0,05 и df = 2 критическое значение хи-квадрат = 5,99. Полученное эмпирическое значение больше критического – различия частот достоверны (хи-квадрат = 9,64; p≤0,05). Как видим, расчет критерия очень прост и не
занимает много времени. Практическая ценность критерия хи-квадрат огромна. Этот метод оказывается наиболее ценным при анализе ответов на вопросы анкет.
Статистический характер расщепления, или расщепление в соответствии с ожидаемой частотой, является методом, используемым для сравнения наблюдаемых и ожидаемых значений в пределах определенного распределения.
Критерий χ2 используется для определения того, насколько хорошо наблюдаемые значения соответствуют ожидаемым значениям, и измеряет разницу между двумя наборами данных.
Чтобы применить критерий χ2, необходимо сравнить наблюдаемые частоты с ожидаемыми частотами. Разница между наблюдаемыми и ожидаемыми частотами выражается в виде суммы квадратов отклонений, деленных на ожидаемые
частоты.
48.
43) Наследование признаков при взаимодействии неаллельных генов: комплементарность, эпистаз, полимерия.Плейотропия и модифицирующее действие генов.
Вот некоторые виды взаимодействия
генов:
1. Комплементарность: В этом типе
взаимодействия гены влияют на один
признак, и полный набор аллелей обоих
генов необходим, чтобы данный признак
проявился. Если у организма есть хотя бы
одна рецессивная аллель на одном из
генов, признак не проявляется.
2. Эпистаз: В этом случае один ген
блокирует или подавляет проявление
признака, определяемого другим геном.
Ген, который подавляет выражение
другого гена, называется эпистатическим
геном, а ген, выражение которого
подавляется, - гипостатическим.
3. Полимерия: В этом типе
взаимодействия несколько независимых
генов влияют на один и тот же признак.
Каждый ген вносит свой собственный
вклад в формирование признака.
- Плейотропия: Это свойство гена влиять
на несколько различных признаков в
организме. Один ген может оказывать
влияние на различные аспекты фенотипа.
- Модифицирующее действие генов: Это
взаимодействие неаллельных генов, при
котором один ген изменяет проявление
другого гена или усиливает его эффект.
Это вид взаимодействия двух и более пар неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака. Полимерное действие
генов может быть кумулятивным и некумулятивным. При кумулятивной полимерии интенсивность значения признака зависит от суммирующего действия генов: чем больше
доминантных аллелей, тем больше степень выраженности признака. При некумулятивной полимерии количество доминантных аллелей на степень выраженности признака не
влияет, и признак проявляется при наличии хотя бы одного из доминантных аллелей. Полимерные гены обозначаются одной буквой, аллели одного локуса имеют одинаковый
цифровой индекс, например А1а1А2а2А3а3.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака
зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 по фенотипу происходит в
соотношении 1: 4: 6: 4: 1.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на
степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15: 1.
Принято различать следующие основные типы взаимодействия неаллельных генов:
Комплементарность, Эпистаз, Полимерия.
Комплементарность это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена, и они вместе
определяют новый признак, который отсутствует у родителей. Причем соответственный признак развивается только в присутствии обоих неаллельних генов. Например, сера
окраска шерсти у мышей контролируется двумя генами (А и В). Ген А детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа) - альбиносы. Другой ген
В обеспечивает скопления пигмента преимущественно у основания и на кончиках волос. Скрещивания дигетерозигот (АаВЬ х АаВЬ) приводит к расщеплению гибридов в
соотношении 9:3:4. Числовые соотношения при комплементарном взаимодействии могут быть как 9:7; 9:6:1 (видоизменение менделивского расщепления).
Примером комплементарного взаимодействия генов у человека может быть синтез защитного белка - интерферона. Его образование в организме связано с комплементарным
взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
Эпистаз -это такое взаимодействие неаллельных генов, при котором один ген подавляет действие другого неаллельного гена. Угнетение могут вызывать как доминантные, так и
рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого розличают эпистаз доминантный и рецессивный. Подавляющий ген получил названиеингибитора или
супрессора. Гены-ингибиторы в основном не детерминируют развитие определенного признака, а лишь подавляют действие другого гена.
Плейотропное действие генов - это зависимость нескольких признаков от одного гена, то есть множественное действие одного гена. В дрозофилы ген белого цвета глаз
одновременно влияет на цвет тела, длины, крыльев, строение полового аппарата, снижает плодовитость, уменьшает продолжительность жизни. У человека известна
наследственная болезнь - арахнодактилия ("паучьи пальцы"-очень тонкие и длинные пальцы), или болезнь Марфана. Ген, отвечающий за эту болезнь, вызывает нарушение
развития соединительной ткани и одновременно влияет на развитие нескольких признаков: нарушение строения хрусталика глаза, аномалии в сердечно-сосудистой системе.
Плейотропное действие гена может быть первичным и вторичным. При первичной плейотропии ген проявляет свой множественный эффект. Например, при болезни Хартнупа
мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишечнике и его реабсорбции в почечных канальцах. При этом поражаются одновременно
мембраны эпителиальных клеток кишечника и почечных канальцев с расстройствами пищеварительной и выделительной систем.
При вторичной плейотропии есть один первичный фенотипний проявление гена, вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к
множественным эффектам. Так, при серповидно клеточной анемии у гомозигот наблюдается несколько патологических признаков: анемия, увеличенная селезенка, поражение
кожи, сердца, почек и мозга. Поэтому гомозиготы с геном серповидно клеточной анемии гибнут, как правило, в детском возрасте. Все эти фенотипные проявления гена составляют
иерархию вторичных проявлений. Первопричиной, непосредственным фенотипним проявлением дефектного гена является аномальный гемоглобин и эритроциты серповидной
формы. Вследствие этого происходят последовательно другие патологические процессы: слипание и разрушение эритроцитов, анемия, дефекты в почках, сердце, мозге - эти
патологические признаки вторичны. При плейотропии, ген, воздействуя на какой то один основнй признак, может также менять, модифицировать проявление других генов, в связи
с чем введено понятие о генах-модификаторах. Последние усиливают или ослабляют развитие признаков, кодируемых "основным" геном.
49.
44) Сцепленное наследование. Закон Т. Моргана. Группы сцепления. Методы генетического картирования. Соматическая гибридизация, еёзначение в установлении групп сцепления человека.
Сцепленное наследование является
типом наследования, при котором гены,
расположенные на одной хромосоме,
наследуются вместе. Этот закон был
открыт ученым Т. Морганом в начале 20
века и получил название "Закон Т.
Моргана о сцепленном наследовании".
Группы сцепления представляют собой
гены, которые находятся на одной и той
же хромосоме и, следовательно,
обычно наследуются вместе
Методы генетического картирования
позволяют установить расположение
генов на хромосомах и определить их
порядок.
Соматическая гибридизация имеет
большое значение в установлении групп
сцепления человека. Путем создания
гибридных клеток и анализа их
генетического материала,
исследователи могут определить, какие
гены находятся вместе на одной
хромосоме и на сколько далеко они
находятся друг от друга. Это позволяет
построить карту групп сцепления и
отслеживать передачу наследуемых
черт от поколения к поколению.
Мендель изучил наследование только семи пар признаков у душистого горошка. Его законы подтвердились на самых разных видах организмов, т. е. было признано, что
эти законы носят всеобщий характер. Однако позже было замечено, что у душистого горошка два признака — форма пыльцы и окраска цветков — не дают независимого распределения в потомстве. Потомки оставались похожими на родителей. Постепенно таких исключений из третьего закона Менделя накапливалось
все больше. Стало ясно, что принцип независимого распределения в потомстве и свободного комбинирования распространяется не на все гены. Действительно, у любого организма признаков очень много, а число хромосом невелико.
В каждой хромосоме должно локализоваться много генов. Каковы же закономерности наследования генов, локализованных & одной хромосоме? Вопрос этот был
изучен выдающимся американским генетиком Т. Морганом.
Предположим, что два гена — А и В находятся в одной хромосоме и организм, взятый для скрещивания, гетерозиготен по этим генам:
В анафазе I мейотического деления гомологичные хромосомы расходятся к разным полюсам и образуются два типа гамет вместо четырех, как должно быть при
дигибридном скрещивании в соответствии с третьим законом Менделя.
При скрещивании с организмом, рецессивным по обоим генам aabb, получается расщепление 1:1 вместо ожидаемого при дигибридном анализирующем скрещивании
1:1:1:1. Такое отклонение от независимого распределения означает, что гены, локализованные в одной хромосоме, наследуются совместно.
Явление совместного наследования генов, локализованных в одной хромосоме, называется сцепленным наследованием, а локализация генов в одной хромосоме —
сцеплением генов. Сцепленное наследование генов, локализованных в одной хромосоме, установил Морган.
Таким образом, третий закон Менделя применим лишь к наследованию аллельных пар, находящихся в негомологичных хромосомах.
Все гены, входящие в состав одной хромосомы, передаются по наследству совместно и составляют группу сцепления. Поскольку в гомологичных хромосомах находятся одинаковые гены, группу сцепления составляют две гомологичные хромосомы. Число групп сцепления у данного вида организмов соответствует числу
хромосом в гаплоидном наборе. Так, у человека 46 хромосом в диплоидном наборе — 23 группы сцепления, у дрозофилы 8 хромосом — 4 группы сцепления, у гороха
14 хромосом — 7 групп сцепления. Однако при анализе наследования сцепленных генов было обнаружено, что в определенном проценте случаев сцепление может нарушаться.
Вспомним, что в профазе I мейотического деления гомологичные хромосомы конъюгируют. В этот момент может произойти обмен участками гомологичных хромосом.
Предположим, что в одной из гомологичных хромосом локализуются пять известных нам доминантных генов, а в другой — пять их рецессивных аллелей. Если
проследить распределение в потомстве двух генов — А и В, то в результате расхождения гомологичных хромосом в анафазе I мейотического деления
дигетерозиготный организм в случае сцепления генов А и В должен давать два типа гамет: АВ и ab. Но если в результате кроссин-говера в некоторых клетках
происходит обмен участками хромосом между генами А и В, то появляются гаметы АЬ и аВ, и в потомстве образуются четыре группы фенотипов, как при свободном
комбинировании генов. Отличие заключается в том, что числовое отношение фенотипов не соответствует отношению 1:1:1:1, установленному для дигибридного
анализирующего скрещивания.
Таким образом, сцепление генов может быть полным и неполным. Причина нарушения сцепления — кроссин-говер, т. е. перекрест хромосом в профазе I мейотического деления. Чем дальше друг от друга расположены гены в хромосоме, тем выше вероятность перекреста между ними и тем больше процент гамет с перекомбинированными генами. В генетике принято определять расстояние между генами в процентах гамет, при образовании которых в результате кроссинговера
произошла перекомбинация генов в гомологичных хромосомах. Кроссинговер — важный источник комбинативной генетической изменчивости.
Закон Моргана
Сцепленные гены, локализованные в одной хромосоме, наследуются совместно и не обнаруживают независимого распределения
Группы сцепления.
Гены, находящиеся в одной хромосоме и наследующиеся сцепленно, составляют группу сцепления. Количество групп сцепления каждого вида должно соответствовать
числу пар хромосом. Кроссинговер возникает со вполне определенной частотой для каждой пары генов, расположенных в одной группе сцепления. Причем, чем ближе
в хромосоме расположены гены друг удругу, тем она выше. На основании анализа частоты кроссинговера между генами можно вычислить расстояние между генами и,
таким образом, определить их локализацию в хромосоме План расположения генов в хромосоме называется картой хромосомы.
Хромосомные карты построены для животных ряда видов, однако для собаки их нет, так как для их составления необходимы фундаментальные исследования генетики
вида и большой статистический материал, получение которого у собак затруднительно. Для кошки установлено положение на хромосомах приблизительно для 50 генов
(Бородин, 1995).
50.
На основании анализа результатов многочисленных экспериментов с мухой дрозофилой Т. Морган сформулировал свою хромосомную теорию наследственности,сущность которой заключается в следующем:
1. Материальные носители наследственности — гены — находятся в хромосомах, располагаются в них линейно на определенном расстоянии друг от друга.
2. Гены, расположенные в одной хромосоме, наследуются сцепленно. Число групп сцепления соответствует гаплоидному числу хромосом.
3. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно.
4. В потомстве гетерозиготных родителей новое сочетание генов, расположенных в одной паре хромосом, может возникнуть в результате кроссинговера в процессе
мейоза.
5. Частота кроссинговера, определяемая по проценту кроссоверных особей, зависит от расстояния между генами.
6. На основании линейного расположения генов в хромосоме и частоты кроссинговера как показателя расстояния между генами можно построить карты хромосом.
Сцепление генов приводит к одновременному наследованию блока признаков. Эти блоки могут сохраняться на протяжении нескольких поколений. Именно они
способствуют передаче сложных признаков, например определяют внешнее сходство дедов и внуков, или обусловливают то, что все метисы колли или кокеров имеют
совершенно определенную внешность независимо от внешнего вида второго родителя.
Сцепление признаков может происходить в силу целого ряда причин, среди них и сцепление генов и плейотропия и действие определенных каналов морфогенеза.
Сцепление признаков в значительной степени обусловливают тип собаки, препотентность производителя и делают возможным существование заводских линий.
Методы генетического картирования
Современные методы картирования сложнонаследуемых признаков и мультифункциональных заболеваний включают 5 основных способов:
анализ сцепления , основанный на проверке конкретной модели наследования болезни в родословных;
Метод идентичных по происхождению аллелей (параметрический метод) , который заключается в оценке того, насколько часто больные родственники наследуют
идентичный участок генома;
анализ ассоциаций в популяциях и семьях ,
анализ неравновесия по сцеплению и
анализ экспериментов по скрещиванию модельных организмов.
Соматическая гибридизация
Гибридизация соматических клеток основана на слиянии совместно культивируемых клеток разных типов, образующих гибридные клетки со свойствами обоих
родительских видов. Используются клетки от разных людей, а также от человека и других животных (мыши, крысы, морской свинки, обезьяны и т.д.). Гибридные
клетки, содержащие два полных генома, при делении обычно «теряют» хромосомы одного из видов. Например, в гибридных клетках «человек – мышь» постепенно
утрачиваются все хромосомы человека. Можно получать клетки с желаемым набором хромосом, что дает возможность изучать сцепление генов и их локализацию в
определенных хромосомах. Можно изучать механизмы первичного действия и взаимодействия генов, регуляцию генной активности. Они позволяют судить о генной
активности. Это позволяет судить о генетической гетерогенности наследственных болезней, изучать их патогенез.
51.
45) Типы определения пола. Типы хромосомного определения пола. Наследование признаков, сцепленных с полом.Существует несколько типов
определения пола, включая:
1. Гонадальное определение пола:
Определяется развитием гонад органов, которые производят половые
клетки. У мужчин гонады называются
яички, а у женщин - яичники.
2. Хромосомное определение пола:
Основано на комбинации половых
хромосом XX у женщин и XY у мужчин.
У женщин оба набора хромосом
являются одинаковыми (XX), а у
мужчин одна из хромосом является
отличной от другой (XY).
3. Гормональное определение пола:
Зависит от уровня и типа половых
гормонов, таких как эстрогены и
тестостерон. Эти гормоны определяют
развитие и функцию половых органов
и вторичных половых признаков.
В отношении наследования признаков,
сцепленных с полом, многие такие
признаки находятся на половых
хромосомах. Например, ген,
ответственный за цвет волос, может
быть сцеплен с полом, то есть
находиться на половых хромосомах.
Наследование, сцепленное с полом — наследование какого-либо гена,
находящегося в половых хромосомах. Наследование признаков,
проявляющихся только у особей одного пола, но не определяемых генами
находящимися в половых хромосомах,- называется наследованием,
ограниченным полом.
Наследованием, сцепленным с X-хромосомой, называют наследование
генов в случае, когда мужской пол гетерогаметен и характеризуется
наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют
две X-хромосомы (XX). Таким типом наследования обладают все
млекопитающие (в том числе человек), большинство насекомых и
пресмыкающихся.
Наследованием, сцепленным с Z-хромосомой, называют наследование
генов в случае, когда женский пол гетерогаметен и характеризуется
наличием W-хромосомы (ZW), а особи мужского пола гомогаметны и
имеют две Z-хромосомы (ZZ). Таким типом наследования обладают все
представители класса птиц.
Если аллель сцепленного с полом гена, находящегося в X-хромосоме или
Z-хромосоме, является рецессивным, то признак, определяемый этим
геном, проявляется у всех особей гетерогаметного пола, которые
получили этот аллель вместе с половой хромосомой, и у гомозиготных по
этому аллелю особей гомогаметного пола. Это объясняется тем, что
вторая половая хромосома (Y или W) у гетерогаметного пола не несет
аллелей большинства или всех генов, находящихся в парной хромосоме.
Таким признаком гораздо чаще будут обладать особи гетерогаметного
пола. Поэтому заболеваниями, которые вызываются рецессивными
аллелями сцепленных с полом генов, гораздо чаще болеют мужчины, а
женщины часто являются носителями таких аллелей.
Примеры заболеваний человека, сцепленного с полом
Гемофилия A Гемофилия В Дальтонизм Лекарственная гемолитическая
анемия, связанная с дефицитом глюкозо-6-фосфатдегидрогеназы (Г6ФД)
Синдром Леша-Найхана X-связанный ихтиоз
Большинство живых организмов нашей планеты размножаются половым путем. Отсюда интерес,
который проявили ученые к объяснению генетических основ определения пола и соотношения
полов (мужского и женского) в природе.
Было показано, что у насекомых, животных, человека и некоторых растений существуют
специфические хромосомы, определяющие пол особи. Эти хромосомы получили название
половых, а все остальные хромосомы - аутосом. При этом, у подавляющего числа организмов
клетки женских особей имеют две одинаковые (морфологически) половые хромосомы, а клетки
мужских особей разные, одна из которых полностью соответствует половым хромосомам женской
особи. Естественно, что в результате в половые гаметы (яйцеклетки) женской особи попадают
одинаковые половые хромосомы, а в мужские половые гаметы (сперматозоиды, спермин) - разные.
Отсюда и определение у подобных организмов женского пола, как гомогаметный (гаметы по
половым хромосомам похожи), а мужского, как гетерогаметный (гаметы несут разные половые
хромосомы).
Женские половые хромосомы были обозначены как X -хромосомы, а мужские как Y.
Следовательно, женские гомогаметные особи несут две X хромосомы (XX), а мужские, являясь
гетерогаметными, X и Y хромосомы (XY). Подобное хромосомное определение пола было
установлено у человека, всех видов млекопитающих, дрозофилы и некоторых видов растений
(конопля, дрема, щавель и др.).
Наряду с этим оказалось, что у птиц, бабочек и некоторых растений (земляника) гомогаметным
является мужской пол (ZZ), а гетерогаметным - женский (ZW). Третий тип определения пола был
описан у кузнечика и многих видов полужесткокрылых (Hemiptera), прямокрылых (Orthotera). У
них самки гомогаметны и несут две Х- хромосомы (XX), а вот самцы, несут только одну Ххромосому (Х0). И наконец, у мхов в клетках женских особей присутствует одна Х-хромосома, а
мужских - одна Y-хромосома. Во всех, вышеописанных случаях, женские хромосомы по размерам
больше мужских.
Большинство видов растений являются однодомными. У них в одном цветке образуются мужские и
женские половые органы и одновременно формируются мужские и женские гаметы. И, независимо
от вида опыления (самоопыление или перекрестное опыление), такие формы получили название
гермафродитных форм.
И наконец, интересно определение пола у пчел и муравьев. У этих насекомых нет половых
хромосом. Самки - диплоидные особи и развиваются из оплодотворенных яиц, а самцы (трутни у
пчел) из неоплодотворенных.
52.
46) Доказательство ведущей роли ДНК в наследственности. Трансформация и трансдукция.Доказательства ведущей роли ДНК в наследственности были получены благодаря экспериментам, проведенным различными учеными:
1. Эксперимент Гриффитса (1928 год): В ходе этого эксперимента использовались два штамма бактерий Streptococcus pneumoniae, один патогенный (капсулированный)
и другой непатогенный (некапсулированный). Гриффитс провел эксперимент, в котором воздействовал на непатогенные бактерии теплом, которое обычно убивает
патогенные бактерии. После этого он вводил мертвые патогенные бактерии вместе с живыми непатогенными бактериями в организм мышей. Результаты показали, что
непатогенные бактерии стали патогенными, что свидетельствовало о передаче генетической информации между штаммами. Это доказало, что передача генетической
информации осуществляется через какую-то вещество, которое переносится от одной бактерии к другой.
2. Эксперименты Герши и Чейза (1952 год): В ходе этих экспериментов использовались бактериофаги, вирусы, инфицирующие бактерии. Герши и Чейз проводили
эксперименты, в которых они меченными изотопами обозначали либо ДНК, либо белки вирусов. В результате они обнаружили, что только меченная ДНК переносится
из вируса в зараженные бактерии, в то время как меченные белки остаются снаружи. Это доказало, что вирусная ДНК является главным носителем генетической
информации, отвечающей за наследственность.
- Трансформация: Это процесс, при котором бактерии могут поглощать и интегрировать в свою ДНК иностранную ДНК из окружающей среды. Этот процесс был впервые
описан Фредериком Гриффитсом и является одним из доказательств ведущей роли ДНК в наследственности.
- Трансдукция: Это процесс, в ходе которого генетический материал может быть передан от одной бактерии к другой с помощью бактериофагов (вирусов,
инфицирующих бактерии). В процессе инфекции вирусом, его ДНК может интегрироваться в ДНК бактерии-хозяина. При последующем взаимодействии с другими
бактериями, возможна передача генетической информации, что подтверждает роль ДНК в наследственности.
Хромосомная теория наследственности закрепила за генами роль элементарных наследственных единиц, локализованных в хромосомах. Однако химическая природа
гена долго еще оставалась неясной. В настоящее время известно, что носителем наследственной информации является ДНК. Убедительным доказательством того, что
именно с ДНК связана передача наследственной информации, получена при изучении вирусов. Проникая в клетку, они вводят в нее лишь нуклеиновую кислоту с очень
небольшой примесью белка, а вся белковая оболочка остается вне зараженной клетки. Следовательно, введенная в клетку ДНК передает генетическую информацию,
необходимую для образования нового поколения вируса такого же типа.
Далее было обнаружено, что чистая нуклеиновая кислота вируса табачной мозаики может заразить растение, вызывая типичную картину заболевания. Более того,
удалось искусственно создать вегетативные "гибриды" из вирусов, в которых белковый футляр принадлежал одному виду, а нуклеиновая кислота - другому. В таких
случаях генетическая информация "гибридов" всегда в точности соответствовала тому виду вирусов, чья нуклеиновая кислота входила в состав "гибрида". Важным
доказательством роли ДНК в передаче наследственной информации были получены на микробных клетках в явлениях трансформации и трансдукции.
Трансформация - включение чужеродной ДНК в бактериальную клетку. Это перенос наследственной информации от одной клетки прокариот посредством ДНК
бактерии-донора или клетки-донора. Трансформация происходит при обработке бактерии мертвыми клетками или экстрактами других штаммов. При этом бактерии
приобретают определенные свойства и сохраняют их. Например, при обработке невирулентного (т. е. не вызывающего заболевания) штамма пневмококков экстрактом
ДНК из вирулентного штамма он приобретал способность вызывать воспаление легких.
Трансдукция заключается в переносе вирусами части ДНК предыдущих бактериальных клеток-хозяев новым клеткам-хозяевам. Так, при инфицировании культуры
неподвижных бактерий вирусами, размножающимися в подвижной культуре, некоторые из бактерий становятся подвижными. Следовательно, изменение
наследственности бактерий при трансдукции достигается благодаря переносу ДНК от одних бактериальных клеток к другим с помощью ДНК или РНК вируса.
Описан переносДНК посредством вирусов, то есть трансдукции, у насекомых. Непигментированных личинок тутового шелкопряда заражали вирусами, которые до этого
паразитировали в окружающих насекомых. В результате у части потомства, полученного от непигментированных, но зараженных особей, появилась окраска. И в этом
случае изменение наследственных свойств было связано с переносом ДНК.
53.
47) Строение, локализация и функции нуклеиновых кислот.Нуклеиновые кислоты - это полимеры, состоящие из нуклеотидных мономеров. Они играют ключевую роль в хранении и передаче генетической информации в клетках.
Строение нуклеиновых кислот: Нуклеотиды состоят из трех компонентов: нитрогеновой базы (аденин, гуанин, цитозин, тимин или урацил), пятиугольного сахара (деоксирибоза в ДНК или рибоза в РНК) и фосфатной группы.
Локализация нуклеиновых кислот: ДНК (дезоксирибонуклеиновая кислота) находится в ядре клеток и несет генетическую информацию, которая определяет структуру и функцию организма. Она также может быть присутствующей в митохондриях и хлоропластах. РНК (рибонуклеиновая кислота) находится в
цитоплазме клеток и выполняет разнообразные функции в синтезе белка, регуляции генов и передаче генетической информации.
Функции нуклеиновых кислот^ Основная функция ДНК - хранить генетическую информацию, которая передается из поколения в поколение.
Значение нуклеиновых кислот очень велико. Особенности их химического строения обеспечивают возможность хранения, переноса в цитоплазму и передачи по наследству дочерним клеткам информации о структуре белковых молекул, которые синтезируются в каждой клетке. Белки обуславливают
большинство свойств и признаков клеток. Существуют два типа кислот: ДНК и РНК.
ДНК (дезоксирибонуклеиновая кислота) — биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие одно из четырех азотистых оснований: аденин
(А) или тимин (Т), цитозин (Ц) или гуанин (Г), пятиатомный сахар пентозу — дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов. РНК (рибонуклеиновая кислота), так же как и ДНК представляет собой полимер,
мономерами которого служат нуклеотиды. Азотистые основания те же самые, что входят в состав ДНК (аденин, гуанин, цитозин), четвертое — урацил — присутствует в молекуле РНК вместо тимина. Нуклеотиды РНК содержат вместо дезоксирибозы другую пентозу — рибозу. В цепочке РНК нуклеотиды
соединяются путем образования ковалентных связей между рибозой одного нуклеотида и остатком фосфорной кислоты другого.
Известны двух — и одно цепочечные молекулы РНК. Двухцепочечные РНК служат для хранения и воспроизведения наследственной информации у некоторых вирусов, т.е. у них выполняется функции хромосом. Одноцепочечные РНК осуществляют перенос информации о последовательности аминокислот в
белках от хромосомы к месту их синтеза и участвуют в процессах синтеза.
Экспрессия генов — реализация информации, записанной в генах, осуществляемой в два этапа: транскрипция, трансляция. Транскрипция — синтез РНК с использованием ДНК в качестве матрицы. В результате возникает 3 типа РНК: матричная (мРНК), рибосомная (рРНК), транспортная (тРНК).
Стадии транскрипции: 1). Инициация — образование нескольких начальных звеньев РНК. 2). Элонгация — продолжается дальнейшее расплетение ДНК и синтез РНК по кодирующей цепи. 3). Терминация — когда полимераза достигает терминатора (точки отсчета транскрипции), она немедленно отщепляется от
ДНК, локальный гибрид ДНК-РНК разрушается и новосинтезированная РНК транспортируется из ядра в цитоплазму. Транскрипция заканчивается.
Трансляция — синтез полипептидной цепи с использованием мРНК в роли матрицы. В трансляции участвуют все три основных типа РНК: м-, р -, тРНК. мРНК является информационной матрицей; тРНК «подносят» аминокислоты и узнают кодоны мРНК; рРНК вместе с белками образуют рибосомы, которые
удерживают мРНК, тРНК и белок и осуществляют синтез полипептидной цепи. Процессинг - совокупность биохимических реакций, при которых пре-РНК укорачиваются, подвергаются химическим модификациям, в результате которых образуются зрелые РНК. В этом процессе участвует четвертый тип РНК —
малая ядерная РНК (мяРНК). Подобно белкам, нуклеиновые кислоты — биополимеры, а их функция заключается в хранении, реализации и передаче генетической (наследственной) информации в живых организмах.
Существует два типа нуклеиновых кислот — дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Мономерами в нуклеиновых кислотах служат нуклеотиды. Каждый из них содержит азотистое основание, пятиуглеродный сахар (дезоксирибоза — в ДНК, рибоза — в РНК) и остаток фосфорной кислоты. В
ДНК входят четыре вида нуклеотидов, отличающихся по азотистому основанию в их составе, — аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т). В молекуле РНК также имеется 4 вида нуклеотидов с одним из азотистых оснований — аденином, гуанином, цитозином и урацилом (У). Таким образом, ДНК и РНК
различаются как по содержанию сахара в нуклеотидах, так и по одному из азотистых оснований (табл. 1).
Таблица 1
Компоненты нуклеотидов ДНК и РНК
Нуклеиновая кислота
Пятиуглеродный сахар Азотистые основания
Остаток фосфорной кислоты
ДНК
Дезоксирибоза
Аденин, гуанин, цитозин, тимин
Остаток фосфорной кислоты
РНК
Рибоза
Аденин, гуанин, цитозин, урацил
Остаток фосфорной кислоты
Молекулы ДНК и РНК существенно различаются по своему строению и выполняемым функциям. Молекула ДНК может включать огромное количество нуклеотидов — от нескольких тысяч до сотен миллионов (поистине гигантские молекулы ДНК удается «увидеть» с помощью электронного микроскопа). В
структурном отношении она представляет собой двойную спираль из полинуклеотидных цепей (рис. 1), соединенных с помощью водородных связей между азотистыми основаниями нуклеотидов. Благодаря этому полинуклеотидные цепи прочно удерживаются одна возле другой.
При исследовании различных ДНК (у разных видов организмов) было установлено, что аденин одной цепи может связываться лишь с тимином, а гуанин — только с цитозином другой. Следовательно, порядок расположения нуклеотидов в одной цепи строго соответствует порядку их расположения в другой.
Этот феномен получил название комплементарности (т. е. дополнения), а противоположные полинуклеотидные цепи называются комплементарными. Именно этим обусловлено уникальное среди всех неорганических и органических веществ свойство ДНК — способность к самовоспроизведению или удвоению
(рис. 2). При этом сначала комплементарные цепи молекул ДНК расходятся (под воздействием специального фермента происходит разрушение связей между комплементарными нуклеотидами двух цепей). Затем на каждой цепи начинается синтез новой («недостающей») комплементарной ей цепи за счет
свободных нуклеотидов, всегда имеющихся в большом количестве в клетке. В результате вместо одной («материнской») молекулы ДНК образуются две («дочерние») новые, идентичные по структуре и составу друг другу, а также исходной молекуле ДНК. Этот процесс всегда предшествует клеточному делению и
обеспечивает передачу наследственной информации от материнской клетки дочерним и всем последующим поколениям. Молекулы РНК, как правило, одноцепочечные (в отличие от ДНК) и содержат значительно меньшее число нуклеотидов.
54.
48) Типы РНК и их роль в синтезе белка клетки.В зависимости от локализации в клетке, функции различают 3 вида РНК: м-РНК (матричная, или информационная), транспортная – т-РНК, рибосомальная –рРНК.
м-РНК. Матричные РНК синтезируются на ДНК в ядре. Процесс называется транскрипция (списывание).Роль м-РНК – она несет информацию об аминокислотной последовательности (т.е. о первичной
структуре) синтезируемого белка. Место каждой аминокислоты в молекуле белка закодировано определенной последовательностью нуклеотидов в цепи м-РНК, т.е. в м-РНК имеются «кодовые
слова» для каждой аминокислоты – триплеты, или кодоны, или генетические коды.
р-РНК. Она входит в состав рибосом. Рибосомы – это РНП, состоящие на 65% из р-РНК и на 35% из белка. Полинуклеотидная цепь р-РНК легко изгибается и укладывается вместе с белком в
компактные тельца. Рибосома состоит из 2-х субъдиниц – большой и малой (соотношение их 2,5:1). В рибосоме различают 2 участка – А (аминокислотный, или участок узнавания) и Р – пептидный,
здесь присоединяется п/п цепь. Эти центры расположены на контактирующих поверхностях обеих субъдиниц. Рибосомы могут свободно перемещаться в клетке, что дает возможность синтезировать
белки в клетке там, где это необходимо. Рибосомы мало специфичны и могут считывать информацию с чужеродных м-РНК, вместе с м-РНК рибосомы образуют матрицу. Роль р-РНК – обуславливает
количество синтезируемого белка.
т-РНК. Этот вид т-РНК изучен лучше всего, составляет 10% всей клеточной РНК. Содержится в цитоплазме, мол.масса небольшая (20тыс.Da) состоит из 70-80 нуклеотидов. Основная роль – транспорт и
установка аминокислот на комплиментарном кодоне м-РНК. т-РНК специфичны к аминокислотам, что обеспечивается ферментом аминоацилсинтетазой. В неактивном состоянии она свернута в
клубочек, а в активном имеет вид трилистника (клеверного листа). В молекуле т-РНК различают несколько участков: а) акцепторный стебель с последовательностью нуклеотидов АЦЦ, к нему
присоединяется аминокислота. Б) участок для присоединения к рибосоме; в) антикодон – участок, комплиментарный кодону м-РНК, который кодирует аминокислоту, присоединенную к данной тРНК . Особенностью первичной структуры т-РНК является то, что они содержат минорные, или модифицированные основания (7-метилгуанин, гипоксантин, дигидроурацил, псевдоурацил, 4тиоурацил), которые способны к неклассическому спариванию. Это ускоряет белковый синтез. Т.о., т-РНК «метит» аминокислоту, придавая ей специфичность и способствует установлению
аминокислоты на определенный участок м-РНК.
Процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта РНК в зрелую РНК.
Наиболее известен процессинг матричных РНК, которые во время своего синтеза подвергаются модификациям: кэпированию, сплайсингу и полиаденилированию. Также модифицируются (другими
механизмами) рибосомные РНК, транспортные РНК и малые ядерные РНК.
Кэпирование
При кэпировании происходит присоединение к 5'-концу транскрипта 7-метилгуанозина посредством трифосфатного моста, соединяющего их в необычной позиции 5'-5', а также метилирование рибоз
двух первых нуклеотидов. Процесс кэпирования происходит во время транскрипции молекулы пре-мРНК. Кэпирование защищает 5'-конец первичного транскрипта мРНК от действия рибонуклеаз,
специфически разрезающихфосфодиэфирные связи в направлении 5’→3'.
Функции кэпа и связанных с ним белков:
экспорт мРНК из ядра;
защита 5'-конца транскрипта от экзонуклеаз;
участие в инициации трансляции;
участие в полиаденилировании.
Полиаденилирование
Фермент поли(А)-полимераза присоединяет 3'-концу транскрипта от 100 до 200 остатков адениловой кислоты. Полиаденилирование осуществляется при наличии сигнальной последовательности 5'AAUAAA-3' на 3'-конце транскрипта, за которой следует 5'-CA-3'. Вторая последовательность является сайтом разрезания.
Сплайсинг
После полиаденилирования мРНК подвергается сплайсингу, в ходе процессе которого удаляются интроны (участки, которые не кодируют белки), а экзоны (участки, кодирующие белки) сшиваются и
образуют единую молекулу. Сплайсинг катализируется крупным нуклеопротеидным комплексом — сплайсосомой, состоящей из белков и малых ядерных РНК. Многие пре-мРНК могут быть
подвергнуты сплайсингу разными путями, при этом образуются разные зрелые мРНК, кодирующие разные последовательности аминокислот (альтернативный сплайсинг).
Редактирование РНК — процесс, в ходе которого информация, содержащаяся в молекуле РНК изменяется путем химической модификации оснований.
55.
49) Генетический код. Основные свойства генетического кода. Расшифровка генетического кода в процессесинтеза белка в клетке.
Генетическая информация о синтезе белка содержится в молекулах ДНК и закодирована с помощью генетического кода. Код, его структура и свойства были открыты в
1960 году.
Структура генетического кода характеризуется тем, что он является триплетным, т. Е. состоит из триплетов азотистых оснований ДНК, получивших название кодонов. 61
сочетаний нуклеотидов кодируют место аминокислоты в полипептидах. Три кодона не кодируют места аминокислот в полипептиде, детерминируют лишь останову
синтеза полипептида. Это стоп-кодоны или терминирующие кодоны.
Свойства генетического кода – неперекрывающийся; линейный; не имеет пунктуации, обеспечивающей свободные пространства между кодонами; вырожденный.
Неперекрываемость означает, что любое азотистое основание является членом только одного кодона. Ни одно азотистое основание не входит сразу в 2 кодона.
Код является линейным по той причине, что молекулы ДНК являются линейными плимерами. Кодоны в виде триплетов азотистых оснований следуют вдоль молекулы
ДНК без перерывов по направлению от 5-конца к 3-концу, причем между кодонами нет свободных пространств, нет пунктуации.
Вырожденность кода определяется тем, что место в полипептиде одной и той же аминокислоты может кодироваться одновременно несколькими кодонами, но не
совместно, а раздельно. Это распространяется на все аминокислоты, кроме метионина и триптофана, которым соответствуют одиночные кодоны.
Расшифровка генетического кода в процессе синтеза белка в клетке.
Транскрипция – первый этап в передаче генетической информации, сущность которого заключается в синтезе мРНК, т.е. в переписывании генетической информации в
молекулы мРНК. Основными структурами которые участвуют в транскрипции, являются ДНК-матрица, РНК-полимераза и хромосомные белки (гистоновые и
негистоновые). Синтез молекул мРНК происходит в ядре и сходит с репликацией ДНК. Для переписывания используется только одна цепь ДНК. Копирование
информации может начинаться с любого участка, к которому прикрепляется РНК-полимераза, и который называют промотором. Фермент (РНК-полимераза) расщепляет
двойную цепочку ДНК, и на одной из её цепей (кодирующей) по принципу комплементарности выстраиваются нуклеотиды РНК. Синтезированная таким образом
(матричный синтез) молекула иРНК выходит в цитоплазму и на один её конец нанизываются малые субъединицы рибосом и происходит сборка рибосом (соединение
малой и большой субъединиц).
Трансляция – это перевод последовательности нуклеотидов в молекуле иРНК в последовательность аминокислот в полипептиде. Транспортные РНК (тРНК) «приносят»
аминокислоты в большую субъединицу рибосомы. Молекула тРНК имеет сложную конфигурацию. На некоторых участках её между комплементарными нуклеотидами
образуются водородные связи, и молекула по форме напоминает лист клевера. На её верхушке расположен триплет свободных нуклеотидов (антикодон), который
соответствует определенной аминокислоте, а основание служит местом прикрепления этой аминокислоты. Каждая тРНК может переносить только свою
аминокислоту.тРНК активируется специльными ферментами, присоединяет свою аминокислоту и транспортирует её в амминоацильный (аминокислотный) центр
рибосомы. Если антикодон тРНК является комплементарным кодону иРНК, находящемуся в аминоацильном центре рибосомы, то происходит временное
присоединение тРНК с аминокислотной к иРНК. После этого рибосома продвигается на один кодон вперед. Первая тРНК с аминокислотой оказывается в пептидильном
центре рибосомы. В освободившийся аминоацильный центр поступает вторая тРНК с аминокислотой. Внутри рибосомы в каждый данный момент находится всего два
кодона иРНК. Аминокислоты располагаются рядом в большой субъединице рибосомы, и с помощью ферментов между ними устанавливается пептидная связь.
Одновременно разрушается связь между первой аминокислотой и её тРНК, и тРНК уходит из рибосомы за следующей аминокислотой. Рибосома перемещается на один
триплет и процесс повторяется. Так постепенно наращивается молекула полипептида, в которой аминокислоты располагаются в строгом соответствии порядком
кодирующих их триплетов (матричный синтез). Регуляция синтеза белка осуществляется специальными кодонами. Начало синтеза определяется кодоном-инициатором
(АУГ), а окончание сборки молекулы белка – кодонами-терминаторами (УАА, УВГ, УГА). После завершения синтеза синтеза белковая молекула отделяется от рибосомы и
приобретает свойственную ей (вторичную, третичную или четвертичную) структуру.
56.
50) Генная инженерия. Синтез и выделение генов. Плазмиды. Достижения генной инженерии в медицине.— совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие организмы.
Генетическая инженерия не является наукой в широком смысле, но является инструментом биотехнологии, используя методы таких биологических наук, как молекулярная и клеточная биология, цитология, генетика, микробиология,
вирусология. Впервые химический синтез гена осуществил в 1969 г. работающий в США индийский ученый Х.Г.Корана с сотрудниками. Он синтезировал ген (участок молекулы ДНК), кодирующий синтез аланиновой т-РНК пекарских дрожжей.
Этот ген состоял из 77 пар нуклеотидов, последовательность которых была известна.
Вначале синтезировали мелкие фрагменты ДНК, содержащие от 4 до 13 нуклеотидных пар, затем с помощью фермента лигазы их соединили в соответствующем порядке, но данный ген был не в состоянии синтезировать аланиновую т-РНК, так
как не содержал акцепторной системы.
В 1976 г. в лаборатории Х.Г.Кораны был синтезирован фрагмент су-прессорной тирозиновой т-РНК длиной 126 пар нуклеотидов, промотор,состоящий из 52 пар нуклеотидов, и терминатор, имеющий 21 пару нуклеотидов.
К концам молекулы ДНК были прикреплены так называемые "липкие концы", состоящие из чередования нуклеотидов ААТТ (один конец) и ТТАА (другой конец). Благодаря этому данный ген был встроен в геном фага и нормально в нем фун
кционировал .Таки м образом, была показана возможность искусственного синтеза генов. Метод позволяет синтезировать относительно мелкие гены, содержащие небольшое число пар нуклеотидов.
Впервые выделить ген методом трансдукции удалось ДжБексвиту с сотрудниками. Они показали, что бактериофаг X при размножении в клетке Е.СоЫ может захватить и встроить в свой геном полный лактозный оперон бактерии вместе с
примыкающим к нему геном-регулятором.
Фрагмент ДНК E.Coli встраивается в геном фага X в строгом порядке: структурные гены, оператор, промотор и регулятор. Применением денатурации и центрифугирования из ДНК-фага был выделен лактозный оперон и ген-регулятор E.Coli.
К сожалению, метод оказался сугубо специфичным для данного гена и не может широко использоваться в генетической инженерии.
Ферментативный синтез. Гены, кодирующие ферменты или структурные белки, состоящие из тысячи и более нуклеотидных пар, рациональнее создавать методом ферментативного синтеза с помощью фермента обратной транскриптазы
(ревертазы). С помощью данного фермента с м-РНК могут быть получены точные копии ДНК (кДНК).
ДНК может быть выделена из любого биологического материала: из крови и других жидкостей организма, различных типов тканей и клеток, содержащих ядра. У человека ДНК чаще всего выделяют из лейкоцитов крови. Процесс выделения
ДНК состоит из нескольких этапов: быстрый лизис клеток, удаление фрагментов клеточных органелл и мембран, ферментативное разрушение и экстрагирование белков, осаждение молекул ДНК в этаноле с последующим их растворением в
буферном растворе. Оценку качества выделенной ДНК проводят на основании измерения оптической плотности раствора ДНК в области белкового и нуклеинового спектров поглощения. В чистых образцах ДНК соотношение А(260)/А(280) > 1,8;
где А(260) и А(280) — оптическая плотность раствора при длине волны 260 и 280 нм, соответственно. Для научных исследований ДНК не только вьщеляют из биологического материала. но и получают синтетически при использовании ДНКсинтезаторов - приборов для автоматического синтеза ДНК. Принципиальное отличие процесса химического синтеза от биологического состоит в присоединении каждого нового нуклеотида к 5'-гидроксильному концу цепи, а не к З'-концу.
Наиболее распространенным методом химического синтеза ДНК является фосфорамидитный (фосфиттриэфирный) метод. Для синтеза используют модифицированные дезоксирибонуклеозиды. Для предотвращения неспецифического
взаимодействия 5'-гидроксильной группы первого нуклеотида до добавления в реакционную смесь второго нуклеотида ее защищают с помощью диметокситритильной (ДМТ) группы. Каждый последующий присоединяемый к растущей цепи
нуклеотид добавляется в колонку в виде фосфорамидита, т.е. содержит ДМТ-группу и диизопропиламинную группу на З'-фосфитной группе, защищенную метильным остатком. Химический синтез необходим для получения одноцепочечных
олигонуклеотидов длиной < 50 п.н. Такие олигоиуклеотиды используют для конструирования целых геномов и их фрагментов, в качестве праймеров для амплификации специфических последовательностей и секвенировании ДНК, для сайтспецифического мутагенеза in vitro, в качестве зондов при гибридизации и в качестве линкеров, облегчающих клонирование. Химический синтез применяют при затруднении клонирования гена (если известна аминокислотная
последовательность белка). В тех случаях, когда ко-доны гена плохо считываются организмом хозяина, можно синтезировать ген с таким набором кодонов (оптимизация кодонов), который сохраняет аминокислотную последовательность
прежней, но транскрибируется более эффективно. Для того чтобы синтезировать ген, необходимо каждую из цепей составляющей его последовательности синтезировать отдельно. При большой протяженности гена его синтезируют
отдельными фрагментами, которые впоследствии «собирают» с помощью ДНК-полимеразы иДНК-лигазы. Все молекулярно-генетические методы основаны на использовании различных классов ферментов, два из которых имеют наибольшее
значение: ДНК-полимеразы и рестриктазы. ДНК-полимеразы осуществляют синтез ДНК, и для их работы необходима одноцепочечная матричная ДНК с двухцепочечным участком на 3'-конце молекулы и четыре типа
дезоксинуклеотидтрифосфатов (dNTP: dATP, dCTP, dGTP и dTTP). ДНК-полимеразы обладают различными видами активности, в том числе и экзонуклеазной в направлении 3' -» 5', что позволяет этим ферментам репарировать дефекты,
образовавшиеся при синтезе ДНК. Способность ДНК-полимеразы I E. colt инициировать репликацию в месте разрыва ДНК и замещать гомологичный участок в двойной цепи ДНК используется для введения в ДНК меченых нуклеотидов методом
никтрансляции, а также для заполнения брешей при сборке протяженного гена из химически синтезированных олигонуклеотидных последовательностей. Основное свойство ДНК-полимераз- осуществлять синтез ДНК — используют при
амплификации изучаемого фрагмента ДНК. Одним из видов амплификации является полимеразная цепная реакция.
Плазми́ ды — небольшие молекулы ДНК, физически отдельные от геномных хромосом и способные реплицироваться автономно. Как правило, плазмиды встречаются у бактерий и представляют собой двухцепочечные кольцевые молекулы, но
изредка плазмиды встречаются также у архей и эукариот. В природе плазмиды обычно содержат гены, повышающие устойчивость бактерии к неблагоприятным внешним факторам (в т. ч. устойчивость к антибиотикам), нередко они могут
передаваться от одной бактерии к другой (иногда даже к бактерии другого вида) и, таким образом, служат средством горизонтального переноса генов.
Генная инженерия оказалась очень перспективной для медицины, прежде всего, в создании новых технологий получения физиологически активных белков, используемых в качестве лекарств (инсулин, соматостатин, интерфероны,
соматотропин и другие). Инсулин используют для лечения больных диабетом, который стоит на третьем месте (после болезней сердца и рака) по частоте вызываемых смертельных случаев. Мировая потребность инсулина составляет несколько
десятков килограммов. Традиционно его получают из панкреатических желез свиней и коров, но гормоны этих животных слегка отличаются от инсулина человека
Генотерапия Неблагоприятная экологическая обстановка и целый ряд других подобных причин приводят к тому, что все больше детей рождается с серьезными наследственными дефектами. В настоящее время известно 4000 наследственных
заболеваний, для большинства из которых не найдено эффективных способов лечения. Генные инженеры уже внесли свой вклад в решение этой проблемы, разработав диагностические препараты, позволяющие обнаруживать генетические
аномалии в период беременности, что дает возможность предотвратить рождение больного ребенка. Однако более одного процента всех новорожденных имеют генетические заболевания, которые приводят к физическим и умственным
нарушениям, а также к ранней смерти..
Трансгенные растения С этой целью была разработана система переноса в растения различных чужих генов, которые могут сообщать растениям полезные свойства. Наиболее распространен перенос генов с помощью вируса, поражающего
фитопатогенную бактерию Agrobacterium tumefaciens. Плазмида найденного в клетках A. tumefaciens способна переносить часть своей ДНК в растительные клетки. Именно в эту ДНК встраивается необходимый "полезный" ген. Растения, в
хромосому которых встраивается чужой ген, называются трансгенными.
57.
51) Современное представление о гене как функциональной единице наследственности и изменчивости. Ген-регулятор, оперон, ген-оператор, структурные гены.Ген-регулятор отвечает за управление активностью гена. Он может продуцировать
молекулы, которые связываются с определенными участками ДНК и влияют на
степень транскрипции гена. Таким образом, ген-регулятор контролирует, когда и в
каком объеме ген будет экспрессирован.
Оперон представляет собой генетическую единицу, состоящую из гена-оператора и
группы структурных генов. Ген-оператор является участком ДНК, к которому
связывается специфический регуляторный белок. Этот белок может включать или
выключать оперон, контролируя транскрипцию структурных генов.
Структурные гены содержат информацию о последовательности аминокислот,
которые используются для синтеза определенных белков.
Ген - функциональная единица наследственного материала. Ген (от греч. genos -- род,
происхождение) - участок молекулы геномной нуклеиновой кислоты,
характеризуемый специфической для него последовательностью нуклеотидов,
представляющий единицу функции, отличной от функций других генов, и способный
изменяться путем мутирования.
От гипотетических дискретных наследственных факторов до локализованных в
хромосомах и молекулах ДНК генов. Долгое время ген рассматривали как
минимальную часть наследственного материала (генома), обеспечивающую развитие
определенного признака у организмов данного вида. Однако каким образом
функционирует ген, оставалось неясным. Термин ген предложен В. Иогансеном в 1909
году, однако проникновение в его сущность связано с именем Г. Менделя, который
еще в 1860-х гг. ввел термин «наследственный фактор» и на основе точных
экспериментов сделал гениальные обобщения относительно свойств и поведения
наследственных факторов при передаче информации от родителей потомкам,
которые в последующем легли в основу теории гена. Это следующие
фундаментальные свойства наследственных факторов - генов:
1) Наличие альтернативных наследственных факторов для развития каждого
конкретного признака организма (в современном представлении доминантный и
рецессивный аллели гена).
2) Парность наследственных факторов, определяющих развитие признака (у
диплоидного организма). Существенный вывод: наследуются не признаки, а от
родителей к потомкам передаются вместе с гаметами гены. Из этих двух положений
был развит принцип аллелизма.
3) Относительное постоянство гена.
58.
52) Реализация генетической информации: транскрипция, посттранскрипционные процессы.ДНК - носитель всей генетической информации в клетке - непосредственного участия в синтезе белков не принимает. В клетках животных и растений молекулы ДНК содержатся в хромосомах ядра и отделены ядерной мембраной от
цитоплазмы, где происходит синтез белков. К рибосомам - местам сборки белков - высылается из ядра несущий информацию посредник, способный пройтичерез поры ядерной мембраны. Таким посредником является информационная РНК (
и-РНК ). По принципу комплементарности она считывается с ДНК при участии фермента, называемого РНК-полимеразой . Процесс считывания (вернее, списывания), или синтеза РНК, осуществляемый РНК-полимеразой, называется
транскрипцией (лат. transcriptio - переписывание). Информационная РНК - это однонитевая молекула, й транскрипция идет с одной нити двунитевой молекулы ДНК. Если в транскрибируемой нити ДНК стоит нуклеотид Г, то РНК-полимераза
включает в РНК Ц, если стоит Т, включает А, если стоит А, включает у (в состав РНК не входит Т) ( рис. 46 ). По длине каждая из молекул и-РНК в сотни раз короче ДНК. Информационная РНК является копией не всей молекулы ДНК, а только части
ее - одного гена или группы рядом лежащих генов, несущих информацию о структуре белков, необходимых для выполнения одной функции. У прокариот такая, группа генов называется опероном . О том, как гены объединены в оперон и как
организовано управление транскрипцией, вы прочтете в в разделе о биосинтезе белков . В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором . Это специфическая
последовательность нуклеотидов ДНК, которую фермент узнает благодаря химическому сродству. Только присоединившись промотору, РНК-полимераза способна начать синтез и-РНК. Дойдя до конца оперона, фермент встречает сигнал (в виде
определенной последовательности нуклеотидов), означающий конец считывания. Готовая и- РНК отходит от ДНК и направляется к месту синтеза белков. В описанном процессе транскрипции можно выделить четыре стадии:
1) Связывание РНК-полимеразы с промотором;
2) Инициация - начало синтеза. Она заключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы и-РНК;
3) элонгация - рост цепи РНК, т. е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации достигает 50 нуклеотидов в
секунду;
4) терминация - завершение синтеза и-РНК.
Постранскрипционные модификации
Процессинг - это превращение первичного транскриптата в зре¬лую и-РНК, выполняющей роль матрицы при трансляции.
Стадии процессинга:
1. Отщепление концевых участков первичного транскриптата.
2. Формирование на 5' конце колпачка.
3. Формирование на 3' конце полиоадениловой последователь¬ности нуклеотидов.
4. Метилирование некоторых азотистых оснований в транскрип-тате, стабилизирующее молекулу РНК.
5. Вырезание неинформативных участков, соответствующих ин-тронам ДНК и сшивание (сплайсинг) участков, соответствую¬щих экзонам.
В результате процессинга у эукариот образуется зрелая и-РНК, имеющая следующее строение:
Колпачок - последовательность нуклеотидов с метилированны¬ми основаниями, которая обеспечивает узнавание малых субъединиц рибосом. Лидер - вводная последовательность нуклеотидов, компли-ментарная последовательности в
молекуле р-РНК малой субъединицы рибосомы, которая обеспечивает прикрепление и-РНК к малой субъ¬единице.
Стартовый кодон - триплет нуклеотидов, кодирующий в боль¬шинстве случаев аминокислоту формилметионин (АЦГ).
Кодирующая часть - последовательность кодонов, шифрующих определенную последовательность аминокислот в полипептидной цепи.
Трейлер - концевая часть молекулы и-РНК, включающая нон-сенс-кодон или поли-А-последовательность (А-А-А-А).
Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс вырезания определенных нуклеотидных последовательностей из молекул РНК и соединения последовательностей, сохраняющихся в «зрелой» молекуле, в ходе
процессинга РНК. Наиболее часто этот процесс встречается при созревании матричной, или информационной, РНК (мРНК) у эукариот, при этом путём биохимических реакций с участием РНК и белков из мРНК удаляются участки, не кодирующие
белок (интроны) и соединяются друг с другом кодирующие аминокислотную последовательность участки — экзоны. Таким образом незрелая пре-мРНК превращается в зрелую мРНК, с которой считываются (транслируются) белки клетки.
Большинство генов прокариот, кодирующих белки, не имеют интронов, поэтому у них сплайсинг пре-мРНК встречается редко. У представителей эукариот, бактерий и архей встречается также сплайсинг транспортных РНК (тРНК)[1] и других
некодирующих РНК.
59.
53) Уникальные свойства ДНК: репликация и репарация.ДНК - это линейный полимер, содержащий от 70-80 до 109 мононуклеотидов, которые соединяються ковалентными фосфодиэфирными связями, возникающими между
гидроксильной группой пентозы одного нуклеотида и фосфатной группойследующего нуклеотида.
Данные рентгеноструктурного анализа показали, что молекула ДНК большинства живых организмов, за исключением некоторых фагов, состоят из двух
полинуклеотидных цепей, антипараллельно напрвленных и ориентированных таким образом, что их сахарофосфатные остовы оказываютя снаружи, а азотистые
основания - внутри. Основания располагаются парами друг против друга и соединяются водородными связями. Спаривание происходит только между
комплементарными (подходящими друг другу) основаниями: одним пуриновым и одним перимединовым. Пара А-Т соединяется двумя, а Г-Ц тремя водородными
связями. Молекула ДНК имеет форму двойной спирали, в котором полинуклеотидные цепи закручены вокруг воображаемой центральной оси.
Спираль ДНК характеризуется рядом параметров:
- ширина спирали около 2 нм;
- шаг или полный оборот спирали составляет 3,4 нм и содержит 10 пар комплементарных нуклеотидов.
ДНК обладает уникальными свойствами: способностью к самоудвоению (репликации) и способностью ксамовосстановлении (репарации).
20 белков: узнающих измененные участки ДНК и удаляющие их из цепи, восстанавливающих правильную последовательность нуклеотидов и сшивающих
восстановленный фрагмент с остальной молекулой ДНК.5% всей клеточной РНК.
Репликация осуществляется под контролем ряда ферментов и протекает в несколько этапов. Она начинается в определенных точках молекулы ДНК. Специальные
ферменты разрывают водородные связи между комплементарными азотистыми, и спираль раскручивается. Поинуклеотидные цепи материнской молекулы
удерживаются в раскрученном состоянии и служат матрицей для синтеза новых цепей.
С помощью фермента ДНК-полимеразы из имеющихся в среде трифосфатов дезоксирибонуклеотидов (дАТФ, дГТФ, дЦТФ, дТТФ) комплементарно материнским цепям
собираются дочерние цепи. Репликация осуществляется одновременно на обеих материнских цепях, но с разной скоростью и снекоторыми отличиями. На одном из
цепей (лидирующей) сборка дочерней цепи идет непрерывно, на другой (отставщей) - фрагментароно. В последующем синтезированные фрагменты смешиваются с
помощью фермента ДНКлигазы. В результате из одной молекулы ДНК образуется две, каждая и которых имеет материнскую и дочернюю цепи. Синтезированные
молекулы являются точными копиями друг друга и исходной молекулы ДНК. Такой способ репликации ДНК называется полуконсервативным и обеспечивает точное
воспроизведение в дочерних молекулах той информации, которая была записана в материнской молекуле.
Репарацией называется способность молекулы ДНК "исправлять" возникающие в ее цепях изменения. В восстановлении исходной молекулы ДНК участвует не менее
Перечисленные особенности химической структуры и свойств ДНК обусловливают выполняемые ею функции. ДНК записывает, хранит, воспроизводит генетическую
информацию, участвует в процессах ее реализации между новыми поколениями клеток и организмов.
1. Репликация ДНК: Репликация является процессом, при котором ДНК молекула копируется для передачи генетической информации при делении клеток. Это
происходит в ходе синтеза новой цепи ДНК на основе существующей цепи.
2. Репарация ДНК: Репарация ДНК - это механизм восстановления поврежденной ДНК-структуры, который возникает из-за мутаций, воздействия вредных факторов
окружающей среды или ошибок в процессе репликации. Организм обладает специализированными ферментами, которые могут опознавать и исправлять повреждения
ДНК.
60.
54) Цитоплазматические гены и их роль в цитоплазматической наследственности.В случае партеногенеза или наследованиее при полиплоидии наследуемые признаки передавались через ядерные структуры - хромосомы . Различия состояли только в том, сколько таких хромосом
имеется в клетке и получает ли организм их от обоих родителей или только от одного. Но, кроме того, существует еще особый тип передачи наследуемых признаков - не через ядро, а через
цитоплазму клетки. В этом случае говорят о нехромосомной, или цитоплазматической наследственности. Наиболее важные случаи нехромосомной наследственности - это наследование пластид и
митохондрий .
Растительные клетки содержат особые органеллы, так называемые пластиды , которые имеют собственную кольцевую хромосому и размножаются делением. Если клетка утратила пластиды, то она
не способна образовать их заново. Например, обычно эвглена зеленая содержит около 100 хлоропластов . При выращивании эвглены в темноте ее хлоропласты не делятся, в то время как сами
одноклеточные продолжают делиться. В результате этого процесса появляются эвглены, не имеющие хлоропластов. У таких эвглен новые хлоропласты не образуются.
Пластиды обычно передаются с яйцеклеткой , но не передаются со спермиями, практически лишенными цитоплазмы. (Однако имеются и исключения, например, спермин герани содержат
цитоплазму и пластиды).
Ясно, что наследование пластид подчиняется особым правилам. Этот тип наследования был описан немецкими учеными, (переоткрывшими законы Менделя) еще в 1908 г. при изучении передачи по
наследству пестролистности у растений. Рассмотрим этот пример более подробно.
Пестролистные растения состоят из клеток с нормальными пластидами, содержащими хлорофилл и имеющими зеленый цвет, и из клеток с мутантными пластидами, которые не содержат
хлорофилла и имеют белый цвет. Листья таких растений "пестрые", т.е. состоят из участков с разной окраской, от чисто зеленой до чисто белой. Нередко одна ветка такого растения несет зеленые
листья, а другая - белые. Сами по себе белые листья не могли бы выжить, так как в них не идет процесс фотосинтеза. Но на пестролистном растении они выживают, и на ветках с такими листьями
даже могут развиваться цветки, так как они получают питательные вещества от нормальных частей растения.
Пластиды наследуются только по материнской линии ( рис. 116 ). Поскольку пыльцевые клетки не содержат пластид, то, например, при опылении цветка нормального зеленого растения пыльцой
цветков, развившихся на ветках с зелеными или с бесцветными листьями, все равно получаются гибриды с нормальными пластидами, т.е. с фенотипом материнского растения.
Теми же особенностями, что и пластиды, обладают митохондрии , имеющие собственную ДНК. Митохондрии сперматозоида при оплодотворении не проникают внутрь клетки или разрушаются в
ней. Так что все митохондрии организм получает от матери. Поскольку подавляющее большинство клеток эукариот содержат митохондрии, нехромосомная наследственность - обычное явление. Этот
тип наследственности зависит от двух факторов: во-первых, от характера распределения данных митохондрий по дочерним клеткам при делении материнской клетки; во-вторых, от свойств генов,
которые локализованы в ДНК пластид или митохондрий. Например, в одну из дочерних клеток может попасть больше мутантных пластид, а в другую меньше или не попасть совсем. В результате
потомки этих дочерних клеток будут обладать разными признаками.
В ряде случаев показано, что хромосомная наследственность и нехромосомная могут комбинироваться, давая сложные случаи наследования признаков. Дело в том, что не все белки, необходимые
для функционирования митохондрий, закодированы в их ДНК. Большая часть таких белков (до 90%) закодирована в ядерной ДНК клетки. Те признаки митохондрий, которые закодированы в
хромосомах ядра клетки, наследуются по законам Менделя, а те признаки, которые закодированы в ДНК самих митохондрий, наследуются (вместе с самими митохондриями) с цитоплазмой
яйцеклетки, т.е. по материнской линии.
У бактерий тоже есть генетический материал ( плазмиды ), который не связан с их единственной хромосомой.
В конце XIX века биологи потратили много труда, чтобы сначала доказать, что носителем наследственности является ядро клетки, а затем конкретизировать это утверждение и доказать хромосомную
теорию наследственности. Противники этой точки зрения пытались доказать, что наследственные признаки передаются через цитоплазму клетки. В этой дискуссии было придумано и проведено
много экспериментов и теорий. Проводилась пересадка ядер из одних клеток в другие, удаление отдельных хромосом из яйцеклеток и т.д. В результате хромосомная теория восторжествовала, а
идея цитоплазматическои наследственности, хотя и не была отвергнута полностью, но влачила жалкое существование. Однако в последние десятилетия ХХ столетия было показано, что такие важные
органеллы, как пластиды и митохондрии, имеют собственный генетический материал и передаются в дочерние клетки с цитоплазмой.
Цитоплазматические гены – гены, локализованные в ДНК органелл, например, в хлоропластной ДНК (в ДНК пластид) и ДНК митохондрий.
61.
55) Генетически-модифицированные объекты. Их медико-биологическое значение.ГМО — генетически модифицированные организмы, еще один термин — трансгенные культуры, или трансгеники. Это
организмы, в которые встраивают чужеродные гены с целью получения хозяйственно-полезных свойств.
-Генетически модифицированные организмы используются в медицине. Зарегистрирован в качестве лекарства генноинженерный человеческий инсулин, получаемый с помощью генетически модифицированных бактерий. Ведутся работы
по созданию генетически модифицированных растений, продуцирующих компоненты вакцин и лекарств против опасных
инфекций (чумы, ВИЧ). Успешно прошло испытания и одобрено к использованию лекарство против тромбозов. Бурно
развивается новая отрасль медицины — генотерапия. В её основе лежат принципы создания ГМО, но в качестве объекта
модификации выступает геном соматических клеток человека.
-Генная инженерия используется для создания новых сортов растений, устойчивых к неблагоприятным условиям среды и
вредителям, обладающих лучшими ростовыми и вкусовыми качествами. Создаваемые новые породы животных
отличаются ускоренным ростом и продуктивностью. Созданы сорта и породы, продукты из которых обладают высокой
питательной ценностью и содержат повышенные количества незаменимых аминокислот и витаминов.
-В настоящее время генетически модифицированные организмы широко используются в фундаментальных и прикладных
научных исследованиях. С помощью ГМО исследуются закономерности развития некоторых заболеваний (болезнь
Альцгеймера, рак), процессы старения и регенерации, изучается функционирование нервной системы, решается ряд
других актуальных проблем биологии и современной медицины.
Генетически-модифицированные объекты (ГМО) представляют собой организмы или продукты, в которых генетический
материал был изменен с помощью технологий генной инженерии.
Медико-биологическое значение ГМО заключается в их потенциальной способности улучшать сельскохозяйственные
культуры, создавать новые лекарства и пищевые продукты или предоставлять другие полезные свойства.
В целом, медико-биологическое значение ГМО заключается в их потенциале улучшить сельское хозяйство, науку и
медицину, но требуется осторожность и грамотный подход к их разработке и применению.
62.
56) Использование генетической информации в процессе жизнедеятельности: трансляция, этапы биосинтеза белка.Генетическая информация. Важнейшим процессом ассимиляции в клетке является синтез белков. Так как белки выполняют в организме целый ряд функций, то необходимо синтезировать тысячи различных белков, тем более что большинство
белков имеют ограниченный срок функционирования и синтез таких белков (компонентов мембран, гормонов, ферментов) не прекращается ни на минуту. Так, например, за сутки в организме человека распадается около 400 г различных
белков, следовательно, такую же массу нужно синтезировать снова.
Каждый вид живых существ имеет свой собственный, строго определенный набор белков. Белки являются основой уникальности каждого вида, хотя некоторые белки, выполняющие одну и ту же функцию в разных организмах, могут быть
похожими и даже одинаковыми.
С другой стороны, все особи одного вида хоть немного, но отличаются друг от друга. На Земле нет, например, двух абсолютно одинаковых людей или амеб. Индивидуальную неповторимость каждой особи определяют различия в структуре
белков.
Свойства белков определяются прежде всего их первичной структурой, т. е. последовательностью аминокислот в молекуле белка. Наследственная информация о первичной структуре белка заключена в последовательности нуклеотидов в
молекулах двуцепочечной ДНК. Следовательно, информация о строении и жизнедеятельности как каждой клетки, так и всего многоклеточного организма в целом заключена в нуклеотидной последовательности ДНК. Эта информация получила
название генетической информации, а участок ДНК, в котором содержится информация о первичной структуре одного белка, называется геном.
Генетический код. Каждой аминокислоте белка соответствует последовательность из трех расположенных друг за другом нуклеотидов ДНК — триплет, или кодон. К настоящему времени составлена карта генетического кода, т. е. известно, какие
триплеты в ДНК соответствуют той или иной из 20 аминокислот, входящих в состав белков (табл. 6), Как известно, в состав ДНК могут входить четыре азотистых основания: аденин (А), гуанин (Г), тимин (Т) и цитозин (Ц). Число сочетаний из 4 по 3
составляет 43 = 64, т. е. ДНК может кодировать 64 аминокислоты. Однако всего кодируется только 20 аминокислот, Оказалось, что многим аминокислотам соответствует не один, а несколько кодонов. Предполагается, что такое свойство
генетического кода — вырожденность — повышает надежность хранения и передачи генетической информации при делении клеток. Например, аминокислоте аланину соответствуют 4 кодона — ЦГА, ЦГГ, ЦГТ и ЦГЦ. Получается, что случайная
ошибка в третьем нуклеотиде кодона не сможет привести к изменениям в структуре белка — все равно это будет кодон аланина.
Примечание: первый нуклеотид триплета берут из левого вертикального ряда, второй — из горизонтального ряда, третий — из правого вертикального. Так как в молекуле ДНК содержатся сотни генов, то в ее состав обязательно входят
триплеты, являющиеся «знаками препинания» и обозначающие начало или конец того или иного гена.
Очень важное свойство генетического кода — специфичность, т. е. один триплет всегда кодирует только одну аминокислоту. Генетический код универсален для всех живых организмов от бактерии до человека.
Транскрипция. Носителем генетической информации является ДНК, расположенная в клеточном ядре. Сам же синтез белка происходит в цитоплазме на рибосомах. Из ядра в цитоплазму информация о структуре белка поступает в виде
информационной РНК (иРНК). Для того чтобы синтезировать иРНК, участок двуцепочечной ДНК раскручивается, а затем на одной из цепочек ДНК по принципу комплементарности синтезируется молекула иРНК. Это происходит следующим
образом: против, например, Г молекулы ДНК становится Ц молекулы РНК, против А молекулы ДНК — У молекулы РНК (вспомните, что вместо тимина РНК несет урапил, или У), против Т молекулы ДНК — А молекулы РНК и против Ц молекулы
ДНК — Г молекулы РНК. Таким образом, формируется цепочка иРНК, представляющая собой точную копию второй (нематричной) цепочки ДНК (только вместо тимина включен урацил). Так информация о последовательности аминокислот в
белке переводится с «языка ДНК» на «язык РНК». Этот процесс получил название транскрипции.
Для транскрипции, т. е. для синтеза иРНК, необходим особый фермент — РНК-полимераза. Так как в одной молекуле ДНК может находиться множество генов, очень важно, чтобы РНК-полимера- за начала синтез иРНК со строго определенного
места ДНК, иначе в структуре иРНК будет записана информация о белке, которого нет в природе и который конечно же клетке не нужен. Поэтому в начале каждого гена находится особая специфическая последовательность нуклеотидов,
называемая про мотором. РНК-полимераза «узнает» промотор, взаимодействует с ним и, таким образом, начинает синтез цепочки иРНК с нужного места. Фермент продолжает синтезировать иРНК, присоединяя к ней новые нуклеотиды, до тех
пор, пока не дойдет до очередного «знака препинания» в молекуле ДНК — терминатора. Это последовательность нуклеотидов, указывающая на то, что синтез иРНК нужно прекратить.
У прокариот синтезированные молекулы иРНК сразу же могут взаимодействовать с рибосомами и участвовать в синтезе белков, У эукариот иРНК синтезируется в ядре, поэтому сначала она взаимодействует со специальными ядерными белками
и переносится через ядерную мембрану в цитоплазму.
В цитоплазме обязательно должен иметься полный набор аминокислот, необходимых для синтеза белков. Эти аминокислоты образуются в результате расщепления белков, получаемых организмом с пищей, а некоторые могут синтезироваться
в самом организме,
Необходимо помнить, что любая аминокислота может попасть в рибосому, только прикрепившись к специальной транспортной
Трансляция. В цитоплазме происходит процесс синтеза белка, который по-другому называют трансляцией. Трансляция — это перевод последовательности нуклеотидов молекулы иРНК в последовательность аминокислот молекулы белка .С тем
концом иРНК, с которого должен начаться синтез белка, взаимодействует рибосома. При этом начало будущего белка обозначается триплетом АУГ, который является знаком начала трансляции. Так как этот кодон кодирует аминокислоту
метионин, то все белки (за исключением специальных случаев) начинаются с метионина. После связывания рибосома начинает двигаться по иРНК, задерживаясь на каждом ее участке, который включает в себя два кодона (т. е. 3 + 3 = 6
нуклеотидов). Время задержки составляет всего 0,2 с. За это время молекула тРНК, антикодон которой комплементарен кодону, находящемуся в рибосоме, успевает распознать его. Та аминокислота, которая была связана с этой тРНК,
отделяется от «черешка» и присоединяется с образованием пептидной связи к растущей цепочке белка. В тот же самый момент к рибосоме подходит следующая тРНК, антикодон которой комплементарен следующему триплету в иРНК, и
следующая аминокислота, принесенная этой тРНК, включается в растущую цепочку. После этого рибосома сдвигается по иРНК, задерживается на следующих нуклеотидах, и все повторяется сначала. Наконец, рибосома доходит до одного из так
называемых стоп-кодонов (УАА, УАГ или У Г А), Эти кодоны не кодируют аминокислот, они только лишь показывают, что синтез белка должен быть завершен. Белковая цепочка отсоединяется от рибосомы, выходит в цитоплазму и формирует
присущую этому белку вторичную, третичную и четвертичную структуры.
Все описываемые реакции происходят за очень маленькие промежутки времени. Подсчитано, что на синтез крупной молекулы белка уходит всего около двух минут.
Клетке необходима не одна, а много молекул каждого белка. Поэтому как только рибосома, первой начавшая синтез белка на молекуле иРНК, продвигается вперед, тут же на эту иРНК нанизывается вторая рибосома, которая начинает
синтезировать такой же белок. На ту же иРНК может быть нанизана и третья, и четвертая рибосома, и т. д. Все рибосомы, синтезирующие белок на одной молекуле иРНК, называются полисомой. Когда синтез белка окончен, рибосома может
связаться с другой молекулой иРНК и начать синтезировать новый белок, закодированный в этой молекуле иРНК. Таким образом, последовательность аминокислот в первичной структуре белка не зависит от рибосом, а определяется только
последовательностью нуклеотидов иРНК.
Таким образом, трансляция — это перевод последовательности нуклеотидов молекулы иРНК в последовательность аминокислот молекулы белка.
63.
57) Особенности организации генома прокариот.У прокариот, к которым относятся бактерии и археи, есть несколько особенностей в организации генома:
1. Круглая хромосома 2. Отсутствие ядра 3. Организация оперонов 4. Плиссировка хромосомы 5. Горизонтальный перенос генов
В целом, организация генома прокариот характеризуется простотой и компактностью, что позволяет им эффективно функционировать и адаптироваться к своей среде.
Прокариоты – это организмы, в клетках которых отсутствует оформленное ядро. Его функции выполняет нуклеоид (то есть «подобный ядру»); в отличие от ядра, нуклеоид не имеет собственной оболочки.
Тело прокариот, как правило, состоит из одной клетки. Однако при неполном расхождении делящихся клеток возникают нитчатые, колониальные и полинуклеоидные формы (бактероиды). В прокариотических клетках отсутствуют постоянные
двумембранные и одномембранные органоиды: пластиды и митохондрии, эндоплазматическая сеть, аппарат Гольджи и их производные. Их функции выполняют мезосомы – складки плазматической мембраны. В цитоплазме фотоавтотрофных
прокариот имеются разнообразные мембранные структуры, на которых протекают реакции фотосинтеза. Иногда их называют бактериальными хроматофорами.
Специфическим веществом клеточной стенки прокариот является муреин, однако у некоторых прокариот муреин отсутствует. Поверх клеточной стенки часто имеется слизистая капсула. Пространство между мембраной и клеточной стенкой
служит резервуаром протонов при фотосинтезе и аэробном дыхании.
Размеры прокариотических клеток изменяются от 0,1-0,15 мкм (микоплазмы) до 30 мкм и более. Большинство бактерий имеет размеры 0,2-10 мкм. У подвижных бактерий имеются жгутики, основой которых служит белки флагеллины.
Организация генома прокариот(на примере кишечной палочки)
Основу генетического аппарата кишечной палочки составляет бактериальная хромосома, входящая в состав нуклеоида – ядерноподобной структуры. Нуклеоид по морфологии напоминает соцветие цветной капусты и занимает примерно 30%
объема цитоплазмы. Бактериальная хромосома представляет собой кольцевую двуспиральную правозакрученную молекулу ДНК, которая свернута во вторичную спираль. Длина бактериальной хромосомы составляет примерно 4,7 млн.
нуклеотидных пар (п.н.), или ~ 1,6 мм. Вторичная структура хромосомы поддерживается с помощью гистоноподобных (основных) белков и РНК. Точка прикрепления бактериальной хромосомы к мезосоме (складке плазмалеммы) является
точкой начала репликации ДНК (эта точка носит название OriC). Бактериальная хромосома удваивается перед делением клетки, и сестринские копии распределяются по дочерним клеткам с помощью мезосомы. Репликация ДНК идет в две
стороны от точки OriC и завершается в точке TerC. Молекулы ДНК, способные себя воспроизводить путем репликации, называются репликоны.
Одна бактериальная хромосома содержит до 1000 известных генов. Обычно это гены «домашнего хозяйства», то есть необходимые для поддержания жизнедеятельности клетки.
Все множество известных генов делится на 10 групп, контролирующих следующие процессы (в скобках указано количество изученных генов):
1. Транспорт различных соединений и ионов в клетку 2. Реакции, поставляющие энергию, включая катаболизм различных природных соединений 3. Реакции синтеза аминокислот, нуклеотидов, витаминов, компонентов цепей переноса
электронов, жирных кислот, фосфолипидов и некоторых других соединений 4. Генерация АТФ при переносе электронов. 5. Катаболизм макромолекул 6.Аппарат белкового синтеза 7. Синтез нуклеиновых кислот, включая гены, контролирующие
реком¬бинацию и репарацию 8. Синтез клеточной оболочки 9. Хемотаксис и подвижность 10. Прочие гены, в том числе с неизвестной функцией
В лаг–фазе в клетке имеется одна бактериальная хромосома, но в фазе экспоненциального роста ДНК реплицируется быстрее, чем происходит деление клетки; тогда число бактериальных хромосом на клетку увеличивается до 2...4...8. Такое
состояние генетического аппарата называется полигаплоидностью.
При делении клетки сестринские копии бактериальной хромосомы распределяются по дочерним клеткам с помощью мезосомы.
Кроме бактериальной хромосомы в состав генетического аппарата прокариот входит множество мелких репликонов – плазмид – кольцевых молекул ДНК длиной в тысячи п.н. Плазмиды такого размера содержат несколько десятков генов.
Обычно это «гены роскоши», обеспечивающие устойчивость к антибиотикам, тяжелым металлам, кодирующие специфические токсины, а также гены конъюгации и обмена генетическим материалом с другими особями. Известны также мелкие
плазмиды длиной 2...3 тпн, кодирующие не более 2 белков. У многих бактерий открыты мегаплазмиды длиной порядка миллиона пн, то есть немногим меньше бактериальной хромосомы. Плазмиды могут быть прикреплены к мезосомам,
могут находиться в автономном состоянии и в интегрированном состоянии. В последнем случае плазмида включается в состав бактериальной хромосомы в определенных точках attB. Таким образом, одна и та же плазмида может включаться в
состав хромосомы и может вырезаться из нее. Существуют плазмиды, представленные одной копией – они реплицируются синхронно с ДНК бактериальной хромосомы. Другие плазмиды могут быть представлены многими копиями, и их
репликация происходит независимо от репликации бактериальной хромосомы. Репликация свободных плазмид часто протекает по принципу «катящегося кольца» – с одной кольцевой матрицы ДНК считывается «бесконечная» копия.
Репликация плазмид может быть синхронизирована с репликацией бактериальной хромосомы, но может быть и независимой. Соответственно, распределение плазмид по дочерним клеткам может быть точным или статистическим.
64.
58) Особенности экспрессии у прокариот.Особенности экспрессии генов у прокариот отличаются от особенностей у эукариот. Вот некоторые из них:
1. Отсутствие окружения ядра 2. Отсутствие сплайсинга РНК 3. Опероны 4. Присутствие репрессоров и активаторов 5. Транскрипционная и трансляционная связь
В целом, экспрессия генов у прокариот регулируется более простыми механизмами по сравнению с эукариотами, что делает их более удобными объектами изучения для исследований в области генетики и биологии.
Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидовДНК) преобразуется в функциональный продукт — РНК или белок. Экспрессия генов может регулироваться на всех стадиях
процесса: и во время транскрипции, и во время трансляции, и на стадии посттрансляционных модификаций белков.
Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений,
так как контроль завременем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме.
Одно из самых важных свойств гена - способность к экспрессии. За это свойство отвечают различные генетические элементы, которые мы должны встроить в векторную молекулу, несущую ген.
Многие бактериальные гены устроены таким образом, что они способны функционировать с существенно разной эффективностью. У E. coli, например, относительное содержание различных белков варьирует в очень широких пределах (от
менее чем 0.1% до 2%) в зависимости от их функций; при этом каждый белок в хромосоме E. coli кодируется единственным геном. Такие вариации обусловлены действием системы контроля генной экспрессии, которая осуществляется главным
образом на уровне транскрипции ДНК. Таким образом, чаще всего уровень активности гена связан с количеством синтезируемой на нем мРНК, то есть с активностью фермента РНК-полимеразы.
Последовательности ДНК, расположенные перед началом структурного гена и определяющие степень активности РНК-полимеразы, называются регуляторными последовательностями. Одна из таких последовательностей представляет собой
участок ДНК, с которым связывается РНК-полимераза. Этот участок называется промотором.
Последовательность оснований промотора определяет частоту инициации синтеза иРНК, причем замена одного основания в этой последовательности может привести к уменьшению частоты инициации в 1000 раз.
Промотор может быть сильным и слабым. Сильный промотор инициирует синтез иРНК часто, слабый - гораздо реже. С другой стороны, промотор может быть регулируемым и нерегулируемым. Например, промотор β-лактамазы
нерегулируемый, но сильный. Использование таких промоторов не всегда удобно. Дело в том, что большое количество белка может блокировать рост бактерий. Кроме того, интенсивная транскрипция рекомбинантной ДНК может помешать
репликации плазмиды, и она будет утрачена. Поэтому удобнее использовать регулируемые сильные промоторы (индуцибельные), включение которых, а значит и синтез чужеродного белка можно осуществить, когда получена большая
бактериальная масса.
Некоторые плазмидные векторы содержат промотор, работа которого регулируется температурочувствительным белковым продуктом гена-репрессора. Белок-репрессор активен при определенных температурах и блокирует действие
промотора. Повысив температуру до 42 оС, можно "включить" промотор и быстро получить большое количество требуемого белка.
В качестве индуцибельных промоторов используют также Trp-промотор триптофанового оперона, который регулируется триптофановым голоданием, lac-промотор лактазного оперона, который индуцируется субстратом (лактозой) и другие.
Интенсивность транскрипции определенных структурных генов может зависеть от эффективности ее терминации, в частности, от того, как часто РНК-полимераза прекращает синтез РНК, не дойдя до этих генов. Сравнительно недавно
обнаружено, что во многих оперонах Е.coli, контролирующих биосинтез аминокислот, между промотором и первым структурным геном имеется терминирующая последовательность. В определенных условиях происходит образование
терминирующего сигнала, ослабляющего интенсивность транскрипции.
Это явление получило название аттенуации, а участок ДНК - аттенуатор (ослабитель). Как и репрессия, аттенуация зависит от присутствия в среде соответствующих аминокислот. Например, избыток триптофана в клетках триптофанзависимых
мутантов, дефектных по репрессору, только 1 из 10 молекул РНК-полимеразы, начавших транскрипцию, преодолевает аттенуатор и считывает структуру генов. Удаление триптофана втрое повышает эффективность транскрипции генов. В отличие
от репрессии, антенуация зависит не от самой аминокислоты, а от триптофанил - тРНК (аминокилоты, присоединенной к соответствующей тРНК).
На эффективность продуктивности рекомбинантной ДНК в существенной степени влияет количество копий этой ДНК в расчете на клетку. Суммарная активность экспрессируемого гена растет с ростом копийности плазмиды. Таким образом,
используя многокопийные плазмиды, можно достичь сверхсинтеза нужных белковых продуктов. Обычно используемые плазмидные векторы (pBR 322 и др.) поддерживаются в клетке в количестве 20-50 копий. Сейчас исследователи имеют в
своем распоряжении температурно-чувствительные мутантные плазмиды, способные накопить до одной-двух тысяч копий на клетку, не нарушая ее жизненно-важных функций. Таким образом можно достичь сверхпродукции плазмидных генов
бактериальными штаммами-сверхпродуцентами.
Регуляция экспрессии у E. coli происходит также и на уровне трансляции. Последовательность оснований длиной 6-8 нуклеотидов, расположенная непосредственно перед инициирующим кодоном АУГ, определяет эффективность трансляции.
Эта последовательность представляет собой участок связывания мРНК с рибосомой. Как правило, он отстоит на 8 нуклеотидов от инициирующего кодона, и его сдвиг в ту или иную сторону может резко снижать эффективность трансляции
соответствующей мРНК. Описанный участок называется последовательностью Шайна-Дальгарно, по имени исследователей, впервые его идентифицировавших.
В состав вектора кроме всего прочего должен входить маркерный ген, позволяющий селектировать измененные клетки. Часто в качестве селективных используют широко распространенные в природе гены ферментов, модифицирующих
антибиотики и инактивирующие их действие.
65.
59) Методы изучения ДНК. Секвенирование генома. Современная геномика.МЕТОДЫ ИЗУЧЕНИЯ ДНК
1. Для выделения ДНК из гомогената тканей уда¬ляют фрагменты клеточных органелл и мембран с помощью центрифугирования. Белки, разрушенные
протеазами (чаще всего применяют протеиназу К), экстрагируют из раствора. Затем ДНК осаждают, на¬пример, этанолом и после удаления надосадочной
жидкости ДНК растворяют в буферном растворе.
2. Молекула ДНК среднего размера содержит 150 000 000 нуклеотидных пар и имеет длину 4 см.
Поэтому молекулы ДНК чувствительны к сдвиго¬вым усилиям, возникающим в растворе, и в процессе выделения ДНК из тканей она фрагменти-руется. Получаются молекулы ДНК значительно
меньше исходных, но все равно очень большие — тысячи или десятки тысяч пар нуклеотидов. Такие молекулы неудобны для исследований, и их при¬ходится дополнительно фрагментировать.
Для фрагментирования используют рестриктазы — ферменты, выделяемые из бактерий. У бак¬терий эти ферменты участвуют в уничтожении чужеродных для них ДНК. Рестриктазы «узнают»
специфические последовательности из 4—6 нук¬леотидов (сайты рестрикции), которые встреча¬ются в ДНК человека. Известно множество раз¬личных рестриктаз, причем каждая из них «узнает»
свой сайт рестрикции (рис. 3.3).
С помощью набора рестриктаз можно разрезать молекулу ДНК на фрагменты желаемой длины. На¬пример, для изучения первичной структуры удобны фрагменты размером около 300 нуклеотидных
пар н.п . Следовательно, цельную молекулу ДНК в 150 000 000 н.п. нужно разрезать на 500 000 фраг¬ментов и каждый из фрагментов изучать отдельно.
Полимеразная цепная реакция (ПЦР). Для прове¬дения некоторых исследований необходимо боль¬шое количество хорошо очищенной высокомоле¬кулярной ДНК. Метод ПЦР дает возможность
избирательно синтезировать in vitro небольшие уча¬стки ДНК и получить за 3—4 ч несколько миллио¬нов копий исследуемого фрагмента. Объектами для выделения ДНК могут быть кровь, биоптат
ткани, слюна, моча, околоплодные воды и т.д. Подробно этот метод и его применение в ДНК-диагностике будут рассмотрены в теме 3.10.
Гибридизация. Для изучения видовой специфично¬сти нуклеиновых кислот применяют метод гибриди¬зации. Он основан на способности ДНК к денатура¬ции при нагревании (80—90 °С) и
ренативации при последующем охлаждении. Возможно использова¬ние метода для проведения гибридизации ДНК-ДНК и ДНК-РНК. Методом гибридизации можно установить сход¬ство и различия
первичной структуры разных об¬разцов нуклеиновых кислот.
Секвенирование генома.
Секвенирование генома здорового человека в большинстве случаев не способно предсказать развитие у него в будущем тех или иных заболеваний. Результаты этой работы были представлены на
Ежегодной встрече Американской Ассоциации по исследованию рака (Association for Cancer Research), а также опубликованы в журнале Science Translational Medicine.
Полное секвенирование генома представляет собой «каталогизирование» всех генов, полученных человеком от обоих родителей, и их проверку на наличие повреждений ДНК, которые могли бы
повысить восприимчивость человека к раку и иным заболеваниям. Поскольку стоимость секвенирования генома постоянно снижается (в настоящее время цена процедуры составляет 1-3 тысячи
долларов США), многие здоровые люди начали задумываться над тем, чтобы пройти такое обследование и определить для себя риск развития того или иного заболевания. Однако исследователи из
Центра по изучению рака Johns Hopkins Kimmel призывают их не торопиться.
Выводы ученых вовсе не означают, что секвенирование генома не имеет никакой медицинской ценности. «Во-первых, секвенирование генома остается лучшим средством для предсказания
„семейных“ заболеваний, таких как рак и некоторые другие, — говорит профессор онкологии Берт Фогельштайн (Bert Vogelstein). — Во-вторых, детальное изучение генома индивидуума помогает
лучше понять механизм уже протекающего у него заболевания и точнее подобрать для него индивидуальную терапию. Однако в случае со здоровым человеком геном перестает быть надежным
предсказателем».
Исследователи проследили развитие 24 заболеваний у более чем 50 тысяч близнецов из 5 стран, прошедших процедуру секвенирования генома. Результаты показали, что в случае 23 заболеваний
анализ генома дал отрицательный результат и отнес риск их развития к разряду «низкий». Однако, по словам профессора Фогельштайна, это вовсе не означает, что данное заболевание у данного
человека не разовьется. «Это означает всего лишь, что его персональный риск немного ниже, чем средний риск по популяции, который может быть очень существенным, — говорит исследователь. —
Таким образом, даже негативный результат не гарантирует отсутствия заболевания в будущем». При этом риск 4 из 24 заболеваний— заболевания сердца у мужчин, аутоиммунный тиреоидит,
диабет I типа и болезнь Альцгеймера — устойчиво определялся в ¾ случаев, что позволяет назвать секвенирование генома достаточно надежным средством определения предрасположенности к
этим болезням.
66.
59) Методы изучения ДНК. Секвенирование генома. Современная геномика.Современная геномика.
Длительное время геномом называли гаплоидный набор хромосом. Накопление сведений об информационной роли внехромосомной ДНК изменило определение термина «геном». В настоящее время он означает полный состав ДНК клетки,
т.е. совокупность всех генов и межгенных участков. Можно считать, что геном - полный набор инструкций для формирования и функционирования индивида.
Общие принципы построения геномов и их структурно-функциональную организацию изучает геномика, которая проводит секвенирование, картирование и идентификацию функций генов и внегенных элементов. Методы геномики направлены
на расшифровку новых закономерностей биологических систем и процессов. Геномика человека является основой молекулярной медицины и имеет важнейшее значение для разработки методов диагностики, лечения и профилактики
наследственных и ненаследственных болезней. Для медицины первостепенное значение имеют исследования в области геномики патогенных микроорганизмов, поскольку они проливают свет на природу инфекционного процесса и создание
лекарств, направленных на специфические мишени бактерий.
Геномика, несмотря на её «молодой возраст», подразделяется на несколько почти самостоятельных направлений: структурную, функциональную, сравнительную, эволюционную, медицинскую геномику. Структурная геномика изучает
последовательность нуклеотидов в геномах, определяет границы и строение генов, межгенных участков и других структурных генетических элементов (промоторов, энхансеров и т.д.), т.е. составляет генетические, физические и транскриптные
карты организма.
Функциональная геномика. Исследования в области функциональной гено-мики направлены на идентификацию функций каждого гена и участка генома, их взаимодействие в клеточной системе. Очевидно, это будет осуществляться путём
изучения белковых ансамблей в разных клетках. Эту область исследований называют протеомикой. Сравнительная геномика изучает сходства и различия в организации геномов разных организмов с целью выяснения общих закономерностей
их строения и функционирования.
Эволюционная геномика объясняет пути эволюции геномов, происхождение генетического полиморфизма и биоразнообразия, роль горизонтального переноса генов. Эволюционный подход к изучению генома человека позволяет проследить
за длительностью формирования комплексов генов, отдельных хромосом, стабильностью его частей, недавно обнаруженными элементами «непостоянства» генома, процессом расообразования, эволюцией наследственной патологии.
Медицинская геномика решает прикладные вопросы клинической и профилактической медицины на основе знания геномов человека и патогенных организмов (например, диагностика наследственных болезней, генотерапия, причины
вирулентности болезнетворных микроорганизмов и т.д.).
Все шаги эволюции живой природы, несомненно, должны были закрепляться в информационной системе ДНК (а для некоторых существ - в РНК), а также в организации её в клетке для выполнения консервативной функции сохранения
наследственности и противоположной функции - поддержания изменчивости. Такое представление о формировании генома каждого вида наиболее обоснованно. Применительно к геному человека можно сказать, что эволюция человека - это
эволюция генома. Такое представление подтверждается теперь многочисленными молекулярно-генетическими исследованиями, поскольку стало возможным сопоставление геномов разных видов млекопитающих, в том числе
человекообразных обезьян, а также в пределах вида Homo sapiens геномов разных рас, этносов, популяций человека и отдельных индивидов. Организация генома каждого эукариотического вида представляет собой последовательную
иерархию элементов: нуклеотидов, кодонов, доменов, генов с межгенными участками, сложных генов, плеч хромосом, хромосом, гаплоидного набора вместе с внехромосомной и внеядерной ДНК. В эволюционном преобразовании генома
каждый из этих иерархических уровней мог вести себя совершенно дискретно (изменяясь, комбинируясь с другими и т.д.).
Наши представления о геноме человека - обширная область генетики человека, включающая по меньшей мере понятия «инвентаризации» генов, групп сцепления, картирования генов (локализация), секвенирования всей ДНК (генов, их мутаций
и хромосом в целом), мейотических преобразований, функционирования отдельных генов и их взаимодействий, интеграции структуры и функции генома в целом. На решении всех этих вопросов была сосредоточена обширная многолетняя
международная программа «Геном человека» (с 1990 по 2000 г.). Главным направлением работ были последовательное секвенирование участков генома и их «состыковка». Успешные разработки в этой области придали программе клиникогенетический аспект.
Клинические приложения сведений о геноме человека
Этапы изучения наследственной болезни
Клинические приложения
Регистрация болезни как наследственной формы
Медико-генетическое консультирование
Локализация гена в хромосоме
Выделение гена
Дифференциальная диагностика на основе анализа сцепления генов
Генотерапия
Определение дефекта гена
Диагностика (ДНК-специфическая)
Обнаружение первичного продукта гена
Диагностика (биохимическая). Улучшение лечения на основе понимания патогенеза
Систематическое изучение генома человека фактически началось с применения менделевского анализа наследственных признаков человека (начало XX века). Генеалогический метод вошел тогда в широкую практику, и шаг за шагом стал
накапливаться материал по «инвентаризации» дискретных наследственных признаков человека, но этот процесс постепенно замедлялся (за 50 лет было открыто не более 400 менделирующих признаков и 4 группы сцепления), возможности
клинико-генеалогического метода в чистом виде были исчерпаны. Бурный прогресс цитогенетики человека, биохимической генетики и особенно генетики соматических клеток в 60-х годах в комплексе с генеалогическим подходом поставил
изучение генома человека на новые теоретические основы и высокий методический уровень. Обнаружение новых менделирующих признаков человека стало быстро продвигаться, особенно на биохимическом и иммунологическом уровне,
появились возможности изучения сцепления и локализации генов. Особый импульс изучению генома человека придали молекулярно-генетические методы, или технология генной инженерии (70-е годы). Процесс познания генома углубился до
выделения гена в чистом виде и его секвенирования. В отличие от классической, в новой генетике изменился подход к анализу генов. В классической генетике последовательность была следующей: идентификация менделирующего признака —
> локализация гена в хромосоме (или группе сцепления) —> первичный продукт гена —> ген. В современной генетике стал возможным и обратный подход: выделение гена —> секвенирование —> первичный продукт, в связи с чем был введён
новый термин для определения такого направления исследований: «обратная генетика» или «генетика наоборот». Продолжаются совершенствование молекулярно-генетических методов и, что не менее важно, их автоматизация. В США и
Великобритании были разработаны и внедрены автоматические приборы по секвенированию геномов. Их назвали геномотронами. В них осуществляется до 100 000 полимеразных реакций в час. Это означает, что в течение недели может быть
просеквенирован участок (или участки) длиной в несколько миллионов пар нуклеотидов. Большую роль в расшифровке генома человека играют вычислительная техника и информационные системы. Благодаря им решаются вопросы
накопления информации (базы данных) из разных источников, хранения её и оперативного использования исследователями из разных стран.
67.
60) Регуляция синтеза белка в клетке прокариотов по Жакобу и МоноРегуляция у прокариот
Регуляция биосинтеза белка упрокариот осуществляется на уровне изменения скорости синтеза мРНК. В настоящее время принята теория оперона,
сформулированная Франсуа Жакобом и Жаком Моно. В основе теории лежат следующие понятия:
оперон – группа тесно связанных между собой генов, которые программируют образование структурных белков и ферментов в клетке,
конституитивные ферменты – те, которые присутствуют в клетках всегда, независимо от ее активности и условий,
индуцибельные ферменты – те, которые программируются опероном и синтезируются при необходимости,
ген-регулятор – ген, регулирующий работу оперона, но не входящий в его состав. Он синтезирует белок-регулятор (чаще называемый белокрепрессор), который может быть в активной или неактивной форме,
ген-оператор – участок ДНК, способный связываться с белком-регулятором, и "решающий" нужно работать РНК-полимеразе или нет.
Схема строения и работы оперона
Предложены две схемы регуляции скорости транскрипции: по механизму индукции(лактозный оперон) и по механизму репресии (триптофановый оперон).
Лактозный оперон
Лактозный оперон в целом отвечает за катаболизм лактозы.
При изучении E.coli было замечено, что в клетке может быть две взаимоисключающие ситуации:
активность одного из ферментов катаболизма лактозы низка, если в среде имеетсяглюкоза.
активность этого фермента резко повышается в обратной ситуации, т.е. при отсутствии глюкозы и при наличии лактозы.
На основании наблюдений была предложена схема регуляции оперона по механизму индукции:
1. В отсутствие лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма лактозы. В результате
эти ферменты не образуются.
2. Если глюкозы нет, а лактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая связаться с геном-оператором. Это позволяет РНКполимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать мРНК.
Таким образом, лактоза является индуктором транскрипции.
Схема работы лактозного оперона при наличии и отсутствии лактозы
Триптофановый оперон
Триптофановый оперон в целом отвечает за синтез триптофана.
Функционирование триптофанового оперона в некотором смысле противоположно лактозному. Регуляция осуществляется по механизму репрессии.
1. В отличие от лактозного оперона, белок-репрессор синтезируется в неактивномсостоянии и не может заблокировать транскрипцию генов, кодирующих
ферменты синтеза триптофана. Синтез этой аминокислоты будет в клетке продолжаться до тех пор, пока в питательной среде не появится триптофан.
2. Триптофан соединяется с белком-репрессором и активирует его. Далее такой активный комплекс присоединяется к гену-оператору и блокирует транскрипцию.
Таким образом, при наличии триптофана в среде прекращается его внутриклеточный синтез, экономятся ресурсы и энергия бактериальной клетки.
В этом случае триптофан является репрессором транскрипции.
Схема работы триптофанового оперона при наличии и отсутствии триптофана
68.
61) Мутационная изменчивость. Мутационная теория Г. Де Фриза. Закон гомологических рядов в наследственной изменчивости Н.Н.Вавилова. Спонтанные и индуцированные мутации. Классификация мутаций.
Мутационная изменчивость относится к изменениям в генетической информации организма, которые могут происходить в процессе репликации ДНК.
Мутационная теория Г. Де Фриза представляет собой концепцию, согласно которой мутации являются основной причиной наследственного изменчивости. Г. Де Фриз предложил классификацию мутаций на основе их влияния на фенотип и наследование.
Закон гомологических рядов, предложенный Н.Н. Вавиловым, говорит о том, что виды, находящиеся ближе к средней точке гомологического ряда, имеют большую генетическую изменчивость и больше подвержены мутациям. Это может объясняться наличием общих предков в гомологических рядах и более
высокой изменчивостью в более прогрессивных группах.
Мутации могут происходить спонтанно (естественно, без воздействия внешних факторов) или индуцироваться (искусственно вызываться факторами, такими как радиация или химические вещества).
Классификация мутаций включает генные мутации (изменения в структуре конкретного гена), хромосомные мутации (изменения в структуре и количестве хромосом) и геномные мутации (изменения, затрагивающие всю генетическую информацию организма).
Мутационной называется изменчивость, вызванная возникновением мутации. Мутации — это наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
Основные положения мутационной теории разработаны Гуго Де Фризом в 1901—1903 гг. и сводятся к следующему:
•Мутации возникают внезапно как дискретные изменения признаков;
•Новые формы устойчивы;
•В отличие от ненаследственных изменений мутации не образуют непрерывных рядов. Они представляют собой качественные изменения;
•Мутации проявляются по-разному и могут быть как полезными, так и вредными;
•Вероятность обнаружения мутаций зависит от числа исследованных особей;
•Сходные мутации могут возникать повторно;
•Мутации ненаправленны (спонтанны), т. е. мутировать может любой участок хромосомы, вызывая изменения как незначительных, так и жизненно важных признаков.
По характеру изменения генома различают несколько типов мутаций — геномные, хромосомные и генные.
Геномные мутации (анеуплоидия и полиплоидия) — это изменение числа хромосом в геноме клетки.
Хромосомные мутации, или хромосомные перестройки, выражаются в изменении структуры хромосом, которые можно выявить и изучить под световым микроскопом. Известны перестройки разных типов (нормальная хромосома — ABCDEFG):
•нехватки, или дефишенси, — это потеря концевых участков хромосомы;
•делеции — выпадение участка хромосомы в средней ее части (ABEFG);
•дупликации — двух- или многократное повторение набора генов, локализованных в определенном участке хромосомы (ABCDECDEFG);
•инверсии — поворот участка хромосомы на 180° (ABEDCFG);
•транслокации — перенос участка к другому концу той же хромосомы либо к другой, негомологичной хромосоме (ABFGCDE).
Гомологические ряды в наследственной изменчивости — понятие, введенное Н. И. Вавиловым[1] при исследовании параллелизмов в явлениях наследственной изменчивости по аналогии сгомологическими рядами органических соединений.
Генетически близкие виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов.
Закономерности в полиморфизме у растений, установленные путем детального изучения изменчивости различных родов и семейств, можно условно до некоторой степени сравнить с гомологическими рядами органической химии, например с углеводородами (CH4, C2H6, C3H8…).[2]
Суть явления состоит в том, что при изучении наследственной изменчивости у близких групп растений были обнаружены сходные аллельные формы, которые повторялись у разных видов (например, узлы соломины злаков с антоциановой окраской или без, колосья с остью или без и т. п.). Наличие такой
повторяемости давало возможность предсказывать наличие ещё не обнаруженных аллелей, важных с точки зрения селекционной работы. Поиск растений с такими аллелями проводился в экспедициях в предполагаемые центры происхождения культурных растений. Следует помнить, что в те годы
искусственная индукция мутагенеза химическими веществами или воздействием ионизирующих излучений ещё не была известна, и поиск необходимых аллелей приходилось производить в природных популяциях.
Мутации делятся на спонтанные и индуцированные. Спонтанные мутации возникают самопроизвольно на протяжении всей жизни организма в нормальных для него условиях окружающей среды. Индуцированными мутациями называют наследуемые изменения генома, возникающие в результате тех или иных
мутагенных воздействий в искусственных (экспериментальных) условиях или при неблагоприятных воздействиях окружающей среды.
Мутации появляются постоянно в ходе процессов, происходящих в живой клетке. Основные процессы, приводящие к возникновению мутаций – удвоение ДНК, нарушения деления ДНК и генетическая перестройка. Для устранения последствий подобных повреждений имеется специальные репарационные
механизмы (например, ошибочный участок ДНК вырезается и на этом месте восстанавливается исходный). Мутации возникают лишь тогда, когда репарационный механизм по каким-то причинам не работает или не справляется с устранением повреждений.
69.
62) Хромосомные аберрации, их типы. Значение хромосомных аберраций в изменчивости.Хромосомные аберрации – изменение структуры хромосом, связанное с разрывом хромосом (при
воздействии на ядро радиации или химических веществ). Можно выделить основные типы хромосомных
мутаций:
1.Делеция- утрата одного, нескольких нуклеотидов или целого гена.
потеря 5 хромосомы - синдром "кошачьего крика" (плачь ребенка напоминает мяуканье кошки)
2. транслокация — обмен участками между негомологичными хромосомами, как результат — изменение
группы сцепления генов
3.Дупликация- повторное дублирование участка ДНК размером от одного до нескольких нуклеотидов или
целого гена
Семейная Х-сцепленная кардиомиопатия
4.Инверсия- встраивание от одного до двух нуклеотидов на прежнее место ДНК после поворота на 180
градусов. В результате нарушается порядок нуклеотидов в гене.
Гемофилия А
5. фрагментация – разрыв хромосомы в нескольких местах.
Хромосомные мутации часто приводят к патологическим нарушениям в организме, но в то же время
хромосомные перестройки сыграли одну из ведущих ролей в эволюции. Так, у человека 23 пары хромосом, а
у обезьяны - 24. Таким образом, различие составляет всего одна хромосома. Ученые предполагают, что в
процессе эволюции произошла хотя бы одна перестройка. Подтверждением этого может служить и тот факт,
что 17 хромосома человека отличается от такой же хромосомы шимпанзе лишь одной перецентрической
инверсией. Такие рассуждения во многом подтверждают теорию Дарвина. Таким образом, хромосомные
мутации приводят к изменению функционирования генов и имеют значение в эволюции вида.
Хромосомные аберрации играют важную роль в изменчивости организмов. Они могут вызывать
генетические нарушения, такие как синдромы Дауна или наследственные раковые заболевания.
70.
63) Точковые мутации. Репарирующие системы клетки.Генные мутации могут быть подразделены на точковые и структурные. Точковые мутации (point mutation) затрагивают один нуклеотид либо 1-2 соседних нуклеотида, тогда как к структурным
мутациям относятся более протяженные дефекты гена. Среди точковых мутаций чаще всего встречаются нуклеотидные замены в кодирующей области гена, т.е. в экзонах. В том случае, если
нуклеотидная замена сопровождается изменением аминокислотного шифра соответствующего кодона, такая мутация ведет к изменению состава полипептидной цепи вследствие встраивания в нее
«ошибочной» аминокислоты. Мутации данного типа называются миссенс-мутация (missense - т.е. с изменением «смысла» повреждаемого кодона). Тяжесть фенотипических проявлений болезни в
этом случае будет определяться функциональной значимостью мутантного участка белка; наиболее тяжелые последствия имеют обычно миссенс-мутации, затрагивающие активные центры белковой
молекулы. Иногда нуклеотидная замена приводит к замещению информационно значимого кодона на стоп-кодон; в результате этого на мутантном участке соответствующей молекулы мРНК
происходит преждевременный обрыв трансляции и образование «усеченной» (truncated), функционально дефектной молекулы белка. Такие нуклеотидные замены обозначаются термином нонсенсмутации (nonsense - «бессмысленные», нарушающие информационную роль кодона). Достаточно распространенным типом мутантных аллелей являются аллели, образующиеся в результате
выпадения (делеции) или вставки (инсерции) нуклеотидов в кодирующей области гена. Если число нуклеотидов при делециях и вставках некратно трем (например, в случае точковой мутации с
выпадением одного нуклеотида), такая мутация приводит к нарушению нормального отсчета кодирующих триплетов, в результате чего нее последующие кодоны в полинуклеотидной цепи
приобретают другой аминокислотный смысл.
Другими словами, данный тип мутации сопровождается сдвигом рамки считывания генетического кода (frameshift); мутации со сдвигом рамки ведут к синтезу «бессмысленного» чужеродного белка,
который быстро деградирует под воздействием внутриклеточных протеаз. Если мутация со сдвигом рамки либо нопсенс-мутация расположены в 5'-области гена (т.е. ближе к его началу), они
сопровождаются отсутствием синтеза с данной конии гена сколько-нибудь значимой в функциональном отношении полипептидной молекулы и полной потерей активности белка - так называемые
нулевые мутации (null mutation). Разумеется, такие мутации являются весьма тяжелыми и нередко либо несовместимы с жизнью, либо характеризуются развитием выраженных мультисистемных
проявлений болезни. В неврологии ярким примером такого рода является атаксия-телеангиэктазия, обусловленная повреждением обеих копий гена ATM, кодирующего белок семейства
фосфатидилинозитолкиназ. Около 90% известных на сегодня мутаций в гене ATM представляют собой нулевые мутации, приводящие к полной инактивации белка и развитию множественных
симптомов поражения головного мозга, кожи, сосудистой, иммунной, эндокринной, костной систем, онкологических осложнений [Gilad S. el al., 1996]. Точковые и структурные мутации могут
происходить не в собственно кодирующих участках гена, а на самом стыке экзонов и интронов - в сайтах сплайсинга (сплайсинговые мутации). Такие мутации нарушают нормальные механизмы
созревания первичного РНК-транскрипта и приводят к неправильному вырезанию нитрона либо к удалению из молекулы РНК информационно значимой экзонной последовательности. В результате
этого происходит значительное нарушение структуры белка, ведущее обычно к развитию тяжелых клинических проявлений болезни. В редких случаях мутации могут затрагивать различные
регуляторные последовательности (например, в области промотора или интронов), влияющие на характер экспрессии гена; такие регуляторные мутации обычно сопровождаются не нарушением
структуры или функции белковой молекулы, а количественными изменениями содержания белка в клетке.\
Однако организмы обладают репарирующими системами, которые помогают исправлять такие точковые мутации.
Существуют несколько типов таких систем:
1. Mismatch repair (MMR) система
2. Base excision repair (BER) система
3. Nucleotide excision repair (NER) система
Все эти репарирующие системы являются важными механизмами защиты клетки от повреждения ДНК и помогают поддерживать его целостность и стабильность.
Репарация (от лат.репарацио – восстановление) – 1) ликвидация повреждения генетических структур (ДНК, хромосом), осуществляется специальными ферментами и находится под контролем генов;
процесс, направленный против возникновения мутаций; репарация свойственна всем живым организмам; 2) восстановление тканей тела и состава популяций организмов, поврежденных или
изреженных ионизирующим излучением или ультрафиолетовыми лучами. Происходит посредством размножения клеток и организмов, уцелевших после облучения.
Повреждения в ДНК сводятся к минимуму благодаря существованию систем, которые узнают эти повреждения и исправляют их. Поэтому определяемая частота мутаций – спонтанных и
индуцированных – отражает равновесие между числом повреждающих событий, происходящих в ДНК, и количеством повреждений, которые были исправлены (или неправильно исправлены).
Значение репарирующих систем в жизни клетки так велико, что, вероятно, по сложности они не отличаются от репликационного аппарата.
Под "повреждением" понимают любое изменение ДНК, которое вызывает отклонение от обычной двухцепочечной структуры. Неправильные нуклеотиды, как правило, удаляются из ДНК системой
эксцизионной репарации (от англ. excision – вырезание). На первом этапе поврежденная структура узнается либо ферментом специфической эндонуклеазой, либо ферментом специфической ДНКгликозилазой. К специфическим эндонуклеазам относится, например, фермент УФ-эндонуклеаза, узнающий тиминовый димер. В каждой клетке существует также не менее 20 различных
специфических ДНК-гликозилаз, узнающих какой-либо один тип измененных оснований в ДНК.
В том случае, когда нарушение структуры ДНК заключается в неправильном спаривании обычных оснований, репарирующая система не может определить, какое из оснований правильное. В этом
случае возможен случайный выбор для удаления одного из неспаренных оснований. Но во многих случаях может быть применена рекомбинационная репарация, которая использует материал одной
молекулы ДНК (из гомологичной хромосомы) для восстановления другой.
71.
64) Индуцированный мутагенез и понятие о мутагенах.Мутагенез — это внесение изменений в нуклеотидную последовательность ДНК (мутаций). Различают естественный (спонтанный) и искусственный (индуцированный) мутагенез.
Искусственный мутагенез широко используют для изучения белков и улучшения их свойств.
Ненаправленный мутагенез Методом ненаправленного мутагенеза в последовательность ДНК вносятся изменения с определенной вероятностью. Мутагенными факторами (мутагенами) могут быть различные химические и физические
воздействия — мутагенные вещества, ультрафиолет, радиация. После получения мутантных организмов производят выявление (скрининг) и отбор тех, которые удовлетворяют цели мутагенеза. Ненаправленный мутагенез более трудоемок и его
проведение оправдано, если разработана эффективная система скрининга мутантов.
Направленный мутагенез В направленном (сайт-специфическом) мутагенезе изменения в ДНК вносятся в заранее известный сайт. Для этого синтезируют короткие одноцепочечные молекулы ДНК (праймеры), комплементарные целевой ДНК за
исключением места мутации.
Мутагенез по Кункелю Для бактериальной плазмиды (внехромосомной кольцевой ДНК) получают уридиновую матрицу, то есть такую же молекулу, в которой остатки тимина заменены на урацил. Праймер отжигают на матрице, проводят его
достройкуin vitro с помощью полимеразы до кольцевой ДНК, комплементарной уридиновой матрице. Двухцепочечной гибридной ДНК трансформируют бактериальные клетки, внутри клетки уридиновая матрица разрушается как чужеродная, и
на мутантной одноцепочеченой кольцевой ДНК достраивается вторая цепь. Эффективность такого способа мутагенеза менее 100 %.
Мутагенез с помощью ПЦР Полимеразная цепная реакция позволяет проводить сайт-направленный мутагенез с использованием пары праймеров, несущих мутацию (рис. 1), а также случайный мутагенез. В последнем случае ошибки в
последовательность ДНК вносятся полимеразой в условиях, понижающих ее специфичность.
Мутагены Индуцированными являются те мутации, которые возникают после обработки клеток мутагенными факторами.
Мутагены бывают:
•физические
•химические
•биологические.
Среди физических мутагенов различают ионизирующую радиацию и ультрафиолетовое излучение. Эти факторы ответственны примерно за 10% всех индуцирующих мутаций.
Ионизирующее излучение – рентгеновское излучение (х-лучи), протоны и нейтроны космических лучей, а также альфа, бета и гамма лучи, освобождаемые радиоактивными элементами изотопов (плутония, фосфора 32, углерода 14, водорода 3
и т.д.). Источником ионизирующего излучения также являются радиоактивные отходы ядерных реакторов. Это излучение обладает высокой энергией и может проникать в ткани, в которых сталкивается с атомами и вызывает освобождение
электронов, оставляя положительно заряженные свободные радикалы или ионы. Они вызывают дальнейшее освобождение электронов.
Мутагенный эффект этого излучения впервые был показан в нашей стране на микроорганизмах Г. А. Надсоном и Г. С. Филипповым в 1925 году. Позднее он был показан в США на дрозофиле Г. Д. Мёллером.
Уф-излучение характеризуется меньшей энергией, проникает только через поверхностные слои клеток животных и растений не вызывает ионизации тканей.
Обычно различают вида излучения:
с длиной волны 180-290 нм. Это излучение не найдено в солнечном свете, так как адсорбируется озоновым слоем. Оно продуцируется искусственно бактерицидными лампами, используемыми для стерилизации
с длиной волны 290-320 нм. Это основная фракция солнечного света. Она мутагена
с длиной волны 320 нм. Оно также обладает повреждающим эффектом
Это излучение не опасно для половых клеток, но опасно для кожи, т.к. вызывает образование в ней тиминовых диметров, которые вызывают мутации не прямо, а нарушением точности репликации.
Химическими мутагенами являются органические и неорганические кислоты, щелочи, перекиси, соли металлов, этиленамины, формальдегид, фенолы, акридиновые красители, алкилирующие соединения, аналоги пуриновых и пиримидиновых
оснований. Аналоги оснований включаются в ДНК вместо нормальных оснований в течение репликации ДНК, изменяя её структуру. На реплицирующую ДНК действуют аналоги азотистых оснований и акридиновые красители. Многие
химические мутагены нарушают процесс мейоза, что приводит к нерасхождению хромосом; а также вызывают разрывы хромосом и генные мутации. Например, алкилирующие соединения могут входить в состав лекарственных препаратов,
поэтому при их приеме и в течение 3 месяцев после окончания лечения необходимо избегать зачатия.
Биологическими мутагенами являются вирусы, которые вызывают хромосомные аберрации в культивируемых клетках, например, вирус гриппа.
72.
65) Множественный аллелизм, наследование признаков и взаимодействие аллелей при множественном аллелизме.Взаимодействие между аллельными генами осуществляется в виде 3 форм: полное доминирование, неполное доминирование и независимое
проявление(кодоминирование).
Опыты Менделя, выявившие полное доминирование одного аллеля и рецессивность другого.
Рецессивный аллель гены может проявиться в результате мутации. Измененный участок ДНК либо не кодирует белок, либо кодирует белок, лишенный активности , что
приводит к нарушению экспрессии рецессивного аллеля. У особи, гомозиготной по рецессивному аллелю, соответствующий белок не образуется , а поэтому обычная
экспрессия данного признака невозможна. В ряде случаев рецессивный фенотип не влияет на жизнеспособность и плодовитость особей, но если белок необходим для
жизни данного организма , то мутантный ген является в этом случае летальным.
Доминантные летальные аллели существуют в генофондах популяций многих видов, но в большинстве случаев они элиминируются, так как вызывают гибель имеющего
их организма. Рецессивные летальные гены не проявляются у гетерозиготного организма, вследствие чего они могут передаваться следующим поколениям и
распространяться в популяции достаточно широко. Подсчитано, что в среднем человек гетерозиготе по 30 летальным рецессивным генам. Это объясняет тот факт, что
среди потомков от браков между близкими родственниками наблюдают высокую частоту летальных наследственных признаков. Одним из леталей человека ялвяется
ген брахидактилииb, обуславливающий в гетерозиготном состоянии Bbукорочение средней фаланги пальцев. У гомозиготbb по этому гену проявляются и другие
нарушения в формировании скелета, приводящие к гибели в раннем возрасте.
В первом случае, когда брахидактилией обладают оба родителя, можно ожидать , что из 4 детей 1 будет нормальным 2 будут гетерозиготны и 1 умрет в раннем
возварсте ю Полученное соотношение фенотипов 1:2:1 совпадает с соотношением генотипов и типично для тех случаев, когда ген обладает неполным
доминированием.
Неполное доминирование наблюдается в том случае, когда одни ген из пары аллелей не обеспечивает образование в достаточном для нормального проявления
признака его белкового продукта. При этой форме взаимодействия генов все гетерозиготы и гомозиготы значительно отличаются по фенотипу друг от друга. Еще одним
примером расщепления при неполном доминировании может служить наследование окраски цветов Ночной красавицы.
При скрещивании растений с красным цветами АА и растений с белыми аа гибриды F1 имеют розовые цветки Аа. Таким образом, имеет место неполное
доминирование ; в F2 наблюдается расщепление 1:2:1 как по фенотипу, так и по генотипу.
Кроме полного и неполного доминирования известны случаи отсутствия доминантно- рецессивных отношений или кодоминирования. При кодоминировании у
гетерозиготных организмов каждый из аллельных генов вызывает формирование в фенотипе контролируемого им признака. Примером этой формы взаимодействия
аллелей служит наследование групп крови человека по системе АВ0, детерминируемых геном I.сущестуют три аллеля этого гена Io, Ia,Ib, определяющие антигены групп
крови.
Если человек гетерозиготенIaIb, его эритроциты имеют оба поверхностных антигены: А и В ( IVгруппа). Аллель Ioрецессивен как по отношению к аллелюIa, так и по
отношению к аллелюIb.
Наследование групп крови иллюстрирует также явление множественного аллелизма: в генофондах популяций человека ген Iсуществует в виде 3 разных аллелей,
которые комбинируются у отдельных индивидуумов только попарною ДО этого примера мы говорили о генах, существующих только в двух разных аллельных формах.
Однако многие гены состоят из сотен пар нуклеотидов , так что мутации могут проходить во многих участках гена и порождать множество различных его аллельных
форм. Так как в каждой из гомологичной зромосом имеется по одному аллельному гену, то, разумеется, диплоидный организм имеет не более двух из серии аллелей
генофонда популяции.
73.
66) Модификационная изменчивость. Норма реакции. Методы изучения модификационной изменчивости.— изменения в организме, связанные с изменением фенотипа вследствие влияния окружающей среды и носящие, в большинстве случаев, адаптивный характер. Генотип при этом не изменяется. В целом современное понятие «адаптивные
модификации» соответствует понятию «определенной изменчивости», которое ввел в науку Чарльз Дарвин.
Модификационная изменчивость — это результат не изменений генотипа, а его реакции на условия окружающей среды. При модификационной изменчивости наследственный материал не изменяется, — изменяется проявление генов.
Основные черты модификационной изменчивости:
1.
модификационные изменения не передаются из поколения в поколение (А.Вейсман);
2.
модификационные изменения проявляются у многих особей вида и зависят от действия на них условий внешней среды;
3.
модификационные изменения возможны только в пределах нормы реакции, т.е. в конечном счете они определяются генотипом.
Изменчивость многих признаков поддается количественному изучению. При исследовании количественных признаков из наблюдаемых объектов составляют вариационный ряд. При составлении вариационного ряда изучаемые особи
располагают друг за другом в порядке возрастания или убывания значения исследуемого признака. Величину изучаемого признака называют вариантой (v). Число особей, соответствующей каждой варианте, называется частотой встречаемости
варианты (p). Важной величиной, характеризующей изменчивость изучаемого признака, является средняя величина, вычисляемая по формуле M = Σ(v•p)/n, где М – средняя величина; Σ - знак суммы, n - общее число вариант вариационного
ряда. Когда изучается изменчивость признака, выражаемая целым числом, варианты называются определенными. Если варианты выражены дробным числом или вариационный ряд очень большой, пользуются классными вариантами (в один
класс объединяют несколько вариант).
Норма реакции
Предел проявления модификационной изменчивости организма при неизменном генотипе — норма реакции. Норма реакции обусловлена генотипом и различается у разных особей данного вида. Фактически норма реакции — спектр
возможных уровней экспрессии генов, из которого выбирается уровень экспрессии, наиболее подходящий для данных условий окружающей среды. Норма реакции имеет пределы или границы для каждого биологического вида (нижний и
верхний) — например, усиленное кормление приведет к увеличению массы животного, однако она будет находиться в пределах нормы реакции, характерной для данного вида или породы. Норма реакции генетически детерминирована и
наследуется. Для разных признаков пределы нормы реакции сильно различаются. Например, широкие пределы нормы реакции имеют величина удоя, продуктивность злаков и многие другие количественные признаки), узкие пределы —
интенсивность окраски большинства животных и многие другие качественные признаки.
Тем не менее, для некоторых количественных признаков характерна узкая норма реакции (жирность молока, число пальцев на ногах у морских свинок), а для некоторых качественных признаков — широкая (например, сезонные изменения
окраски у многих видов животных северных широт). Кроме того, граница между количественными и качественными признаками иногда весьма условна.
Модификационная изменчивость тесно связана с естественным отбором. Естественный отбор имеет четыре направления, три из которых непосредственно нацелены на выживание организмов с разными формами ненаследственной
изменчивости. Это стабилизирующий, движущий и дизруптивный отбор.
Стабилизирующий отбор характеризуется обезвреживанием мутаций и формирования резерва этих мутаций, что обуславливает развитие генотипа при постоянном фенотипе. Вследствие этого организмы со средней нормой реакции
доминируют в неизменных условиях существования. Например, у генеративных растений сохраняется форма и размер цветка, которые отвечают форме и размеру насекомого, которое опыливает растение.
Дизруптивный отбор характеризуется раскрытием резервов с обезвреженными мутациями и последующим отбором этих мутаций для формирования новых генотипа и фенотипа, которые подходят под окружающую среду. Вследствие этого
выживают организмы с крайней нормой реакции. Например, насекомые с большими крыльями имеют большую устойчивость к порывам ветра, тогда как насекомых того же вида со слабыми крыльями сдувает.
Движущий отбор характеризуется тем же механизмом, что и дизруптивный, однако он нацелен на формирование новой средней нормой реакции. Например, у насекомых появляется стойкость к химикатам.
Для изучения модификационной изменчивости существует несколько методов:
1. Экспериментальный подход
2. Наблюдательный подход
3. Исследования на уровне генома:
В целом, изучение модификационной изменчивости требует комбинации различных методов, которые позволяют нам понять, как организмы могут изменять свои характеристики в ответ на окружающую среду.
74.
67) Особенности человека как объекта генетических исследований, его биосоциальная природа человека.Человек как объект генетического исследования
Раздел генетики, изучающий наследственность и изменчивость у человека, называется антропогенетикой или генетикой человека. Генетика
человека - это наука о наследственно обусловленных различиях между людьми. Из генетики человека выделяется медицинская генетика,
исследующая механизмы развития наследственных болезней, возможности их лечения и профилактики. В настоящее время человек хорошо
изучен морфологически, физиологически, биохимически, что способствует рассмотрению его генетических особенностей.
Изучение генетики человека связано с биологическими и социально-этическими особенностями.
Биологические особенности: позднее половое созревание, малочисленное потомство у одной пары родителей, в основном моноплодная
беременность (исключение - близнецы); большой срок беременности, медленная смена поколений (20 - 25 лет), особенности кариотипа (большое
число хромосом и др.), фенотипический полиморфизм.
Социально-этические особенности: невозможность направленных скрещиваний в интересах исследователя, отсутствие точной регистрации
наследственных признаков (проводится не всегда и не везде), невозможность создания одинаковых условий жизни для всех людей.
У человека есть и преимущества перед другими генетическими объектами: способность воспринимать информацию и абстрактно мыслить;
значительное число и разнообразие мутаций; высокая численность популяций, доступных для изучения; возможность регистрации
наследственных признаков в течение длительного времени; использование гибридизации соматических клеток для генетического анализа.
Биосоциальная природа человека отражается в том, что его жизнь определяется единой системой условий, в которую входят как биологические,
так и социальные элементы. Это вызывает необходимость не только его биологической, но и социальной адаптации, т. е. приведения
межиндивидуального и группового поведения в соответствие с господствующими в данном обществе, классе, социальной группе нормами и
ценностями в процессе социализации (путем усвоения знаний об этом обществе, классе и т. д.) . Эту область человеческой природы изучает
большая группа социальных дисциплин, с которыми экология весьма тесно связана (социально-экономические науки и др.) . Биологическая
адаптация человека весьма отличается от таковой в животном мире, так как стремится сохранить не только его биологические, но и социальные
функции при возрастающем значении социального фактора. Последнее обстоятельство имеет важное экологическое значение и нашло свое
отражение в экологическом подходе к определению понятия «человек» .
Человек — один из видов животного царства со сложной социальной организацией и трудовой деятельностью, в значительной мере
«снимающими» (делающими малозаметными) биологические, в том числе этологические (первично — поведенческие) свойства организма
(Реймерс, 1990).
Общие законы взаимоотношения человека (или группы людей) и биосферы, влияние на человека (или группы людей) природной и социальной
сред изучает наука экология человека.
75.
68) Генетический полиморфизм человека. Мутации и их роль в развитии заболеваний.Генетический полиморфизм, сосуществование в пределах популяциидвух или нескольких различных наследственных форм, находящихся в динамическом равновесии в
течение нескольких и даже многих поколений. Чаще всего Генетический полиморфизм обусловливается либо варьирующими давлениями и векторами
(направленностью) отбора в различных условиях (например, в разные сезоны), либо повышенной относительной жизнеспособностью гетерозигот. Один из видов
Генетический полиморфизм — сбалансированный Генетический полиморфизм — характеризуется постоянным оптимальным соотношением полиморфных форм,
отклонение от которого оказывается неблагоприятным для вида, и автоматически регулируется (устанавливается оптимальное соотношение форм).В состоянии
сбалансированного Генетический полиморфизм у человека и животных находится большинство генов. Различают несколько форм Генетический полиморфизм, анализ
которых позволяет определять действие отбора в природных популяциях.
Одной из главных причин пороков развития являются мутации. В организме они происходят постоянно (спонтанные мутации) под воздействием естественного фона
радиации и процессов тканевого метаболизма. При дополнительном воздействии на организм ионизирующего излучения или химических мутагенов происходят
индуцированные мутации. Мутации могут быть генными, хромосомными и геномными. Первые представляют собой новые молекулярные состояния гена. С мутацией
единичных генов связано около 13% пороков. Хромосомные мутации — это изменения хромосом в виде транслокации, делеции, дупликации и инверсии.
Геномные мутации — изменение числа хромосом или хромосомных наборов. Хромосомные и геномные мутации индуцируют развитие хромосомных болезней. Под
«перезреванием» половых клеток понимают комплекс изменений в яйцеклетках и сперматозоидах, произошедших от момента их полного созревания до образования
зиготы. Они наблюдаются в основном при увеличении времени от эякуляции до слияния спермия с яйцеклеткой и связаны преимущественно с изменением рН среды в
половых путях, снижением подвижности сперматозоидов, с нарушением проходимости труб. Следствием «перезревания», по-видимому, является нерасхождение
хромосом, что в дальнейшем проявляется геномными мутациями. Среди эндокринных заболеваний, вызывающих пороки развития, главную роль играет сахарный
диабет. Пороки развития у детей возникают как при клинически проявляющихся, так и при латентных формах заболевания у матери, но особенно часто — у женщин,
которые заболели в препубертатном периоде. Зависимость состояния ребенка от возраста родителей, в котором произошло его зачатие, хорошо известна. Так, у
женщин старше 35 лет и мужчин старше 40 лет значительно увеличивается риск рождения ребенка с хромосомными болезнями, обусловленными числовыми
изменениями хромосом. У отцов с возрастом повышается риск рождения ребенка с пороками, обусловленными вновь возникшими доминантными мутациями.
Генетический полиморфизм человека означает наличие множества различий в генетической информации между индивидами одного вида.
Мутации, в свою очередь, являются изменениями в генетической последовательности, которые могут возникнуть случайно или под воздействием внешних факторов.
Мутации могут быть вредными, благоприятными или нейтральными, в зависимости от их влияния на функцию гена.
Роль мутаций в развитии заболеваний проявляется в следующих аспектах:
1. Генетически обусловленные заболевания
2. Восприимчивость к болезням
3. Фармакогенетика
Объединение генетического полиморфизма и мутаций дает значительное разнообразие генетической информации в популяции и имеет важное значение для
понимания генетической основы заболеваний и разработки персонализированной медицины.
76.
69) Роль наследственности и среды в формировании нормального и патологически измененного фенотипачеловека. Наследственные болезни человека: хромосомные, генные, болезни с наследственной
предрасположенностью. Мультифакториальные заболевания.
Роль наследственности и среды в формировании фенотипа человека включает взаимодействие между
наследственными факторами (гены) и окружающей средой.
Наследственные болезни человека могут быть классифицированы как хромосомные, генные и болезни с
наследственной предрасположенностью.
Хромосомные болезни связаны с изменениями в структуре или количестве хромосом и включают такие
состояния, как синдром Дауна, синдром Тёрнера и синдром Клайнфельтера.
Генные болезни связаны с наличием изменений (мутаций) в генах и могут включать состояния, такие как
муковисцидоз, гемофилия, наследственный рак и синдромы, связанные с нарушениями работы
определенной группы генов.
Болезни с наследственной предрасположенностью возникают при взаимодействии наследственных
факторов с окружающей средой. Например, особенные гены могут увеличить риск развития
определенных заболеваний, при этом окружающая среда может сыграть роль в том, разовьется ли
болезнь или нет.
Мультифакториальные заболевания возникают при взаимодействии наследственных и окружающих
факторов. Это такие состояния, как сердечно-сосудистые заболевания, диабет, некоторые виды рака и
психические расстройства.
В целом, формирование нормального и патологически измененного фенотипа человека является
результатом сложного взаимодействия генетических и окружающих факторов.
77.
70) Биосоциальная природа человека. Методы генетики человека и их характеристика. Цитогенетический метод, его сущность ивозможности.
Биосоциальная природа человека означает, что его здоровье и характеристики определяются взаимодействием генетических, биологических, социальных и окружающих факторов. Методы генетики
человека позволяют изучить генетическую природу человека и понять, как гены влияют на его фенотип и здоровье.
Один из методов генетики человека - цитогенетический метод, основанный на изучении хромосом. Он позволяет исследовать хромосомные аномалии, мутации и наследственные болезни.
Сущность цитогенетического метода заключается в анализе числа, структуры и расположения хромосом в клетках организма. Для этого берутся образцы тканей, такие как кровь, кожа или
амниотическая жидкость, и проводится цитогенетическое исследование.
Цитогенетический метод имеет свои возможности и ограничения. Он может помочь в диагностике наследственных заболеваний, определении риска возникновения определенных генетических
нарушений и изучении хромосомных изменений, но не всегда позволяет точно определить гены, ответственные за конкретные болезни.
Генетика человека – это особый раздел генетики, который изучает особенности наследования признаков у человека, наследственные заболевания (медицинская генетика), генетическую структуру
популяций человека. Генетика человека является теоретической основой современной медицины и современного здравоохранения.
В настоящее время твердо установлено, что в живом мире законы генетики носят всеобщий характер, действительны они и для человека.
Однако, поскольку человек – это не только биологическое, но и социальное существо, генетика человека отличается от генетики большинства организмов рядом особенностей:
– для изучения наследования человека неприменим гибридологический анализ (метод скрещиваний); поэтому для генетического анализа используются специфические методы: генеалогический
(метод анализа родословных), близнецовый, а также цитогенетические, биохимические, популяционные и некоторые другие методы;
– для человека характерны социальные признаки, которые не встречаются у других организмов, например, темперамент, сложные коммуникационные системы, основанные на речи, а также
математические, изобразительные, музыкальные и иные способности;
– благодаря общественной поддержке возможно выживание и существование людей с явными отклонениями от нормы (в дикой природе такие организмы оказываются нежизнеспособными).
Генетика человека изучает особенности наследования признаков у человека, наследственные заболевания (медицинская генетика), генетическую структуру популяций человека. Генетика человека
является теоретической основой современной медицины и современного здравоохранения. Известно несколько тысяч собственно генетических заболеваний, которые почти на 100% зависят от
генотипа особи. К наиболее страшным из них относятся: кислотный фиброз поджелудочной железы, фенилкетонурия, галактоземия, различные формы кретинизма, гемоглобинопатии, а также
синдромы Дауна, Тернера, Кляйнфельтера. Кроме того, существуют заболевания, которые зависят и от генотипа, и от среды: ишемическая болезнь, сахарный диабет, ревматоидные заболевания,
язвенные болезни желудка и двенадцатиперстной кишки, многие онкологические заболевания, шизофрения и другие заболевания психики.
Для изучения генетики человека разработаны и успешно используются следующие методы: клинико-генеалогический (метод родословных), популяционно-статистический(методы, используемые для
установления частот генов и генотипов в популяции, демонстрирующие характер их изменения под влиянием окружающей среды и различных факторов популяционной динамики, называются
популяционно-статистические), близнецовый(с помощью близнецового метода можно выявить наследственный характер признака, определить пенетрантность аллеля, оценить эффективность
действия на организм некоторых внешних факторов (лекарственных препаратов, обучения, воспитания), дерматоглифики и пальмоскопии(в настоящее время установлена наследственная
обусловленность кожных узоров. Вероятно, признак наследуется полигенно, на характер пальцевого и ладонного узоров оказывает влияние материнский организм через механизм
цитоплазматической наследственности. Дерматоглифические исследования важны при идентификации зиготности близнецов), цитогенетический(методы генетики соматических клеток в
значительной мере компенсируют невозможность применения к человеку метода гибридологического анализа. Благодаря разработке методов генетики соматических клеток человек оказался
включенным в группу объектов экспериментальной генетики), молекулярно-цитогенетические: биохимические, молекулярно-генетические методы и метод генетики соматических клеток. На
основании совокупности методов основана пренатальная диагностика наследственных заболеваний. На лабораторных занятиях подробно изучаются клинико-гениалогический, цитогенетический и
популяционно-статистический методы, а также статистическое изучение модификационной изменчивости. Для формулировки научно обоснованных выводов необходимо использовать методы
биометрии.
Задачи медицинской генетики заключаются в своевременном выявлении носителей этих заболеваний среди родителей, выявлении больных детей и выработке рекомендаций по их лечению.
Большую роль в профилактике генетически обусловленных заболеваний играют генетико-медицинские консультации и пренатальная диагностика (то есть выявление заболеваний на ранних стадиях
развития организма).
Существуют специальные разделы прикладной генетики человека (экологическая генетика, фармакогенетика, генетическая токсикология), изучающие генетические основы здравоохранения. При
разработке лекарственных препаратов, при изучении реакции организма на воздействие неблагоприятных факторов необходимо учитывать как индивидуальные особенности людей, так и
особенности человеческих популяций.
78.
71. Генеалогический метод изучения наследования признаков у человека. Составление и анализ родословных.Клинико-генеологический метод был предложен в 1865 году Ф. Гальтоном. Метод основан на прослеживании интересующего нас признака (нормального или патологического) в семье, с указанием
родственных связей между отдельными членами этой семьи (составлением родословной).
Клинико-генеалогический метод дает возможность:
- выявлять наследственный характер признака;
- определять тип наследования;
- определять зиготность членов родословной;
- определять особенности взаимодействия генов;
- устанавливать сцепленное наследование и проводить картирование хромосом;
- определять пенетрантность гена;
- изучать закономерности мутирования отдельных генов;
- устанавливать носительство мутантного гена тем или иным членом семьи;
- определять вероятность генетически обусловленных событий и рассчитывать риск наследования патологического гена (признака) при медико-генетическом консультировании.
Клинико-генеалогический метод часто осложняется невозможностью сбора достаточного количества информации из-за малодетности семей, либо из-за прерывания связей между
поколениями, отсутствия связей между родственниками, либо по морально-этическим причинам.
Клинико-генеалогический метод лежит в основе медико-генетического консультирования и включает 3 этапа:
1 этап – клиническое обследование;
2 этап – составление родословной;
3 этап – генетический анализ родословной.
Первый этап – клиническое обследование. При составлении родословной сбор сведений о семье начинается с человека, которого называют пробанд (обычно это больной с изучаемым
заболеванием или признаком). В сведениях о пробанде указывается анамнез заболевания, включающий начальные признаки и возраст их манифестации, последующее течение болезни; если
пробанд – ребенок – сведения о раннем психомоторном и последующим умственном и физическом развитии. Чем больше поколений удается проследить и чем более полно охватить членов
родословной при сборе сведений, тем больше вероятность получения достоверных сведений о характере наследования изучаемого признака.
Сбор генетической информации проводится путем опроса, анкетирования, личного собеседования. Опрос начинается обычно с родственников по материнской линии. В родословную вносят
сведения о выкидышах, абортах, мертворожденных, бесплодных браках, внебрачных детях и др. При сборе генетической информации о проявлении изучаемого признака ведется краткая запись
данных о каждом члене рода с указанием его родства по отношению к пробанду. Обычно указывается фамилия (для женщин девичья фамилия), имя, отчество, дата рождения и смерти. Полученные
данные записываются в медико-генетическую карту. При сборе информации необходимо внимательно анализировать сообщения об инфекциях и травмах, следует учитывать гетерогенность и
варьирующую экспрессивность наследственных заболеваний. Необходимо выяснять акушерский анамнез, учитывать наличие и характер профессиональных вредностей, возраст, национальность,
место жительства семьи, профессию, наличие хронических заболеваний в семье, причину смерти умерших и др. На основании изученных данных составляется анамнез (греч. – anamnesis –
воспоминание).
79.
71. Генеалогический метод изучения наследования признаков у человека. Составление и анализ родословных.Второй этап – составление родословной. После сбора сведений составляется графическое изображение родословной,
для этого используется система символов, предложенная в 1931 году Г. Юстом. (Рис. 1).
При составлении графического изображения родословной важно соблюдать следующие правила:
1.Составление родословной начинают с пробанда. Братья и сестры (сибсы) располагаются в порядке рождения слева
направо, начиная со старшего.
2.Все члены родословной располагаются строго по поколениям, в один ряд.
3.Поколения обозначаются римскими цифрами слева от родословной сверху вниз.
4.Арабскими цифрами нумеруется потомство одного поколения (одного ряда) слева направо. Благодаря такой
нумерации каждый член семьи имеет свой шифр (например: I-1, I-2, II-2, II-4 и др.)
5.Указывается возраст членов семьи (родословной), в связи с тем, что некоторые болезни проявляются в разные
периоды жизни.
6.Отмечаются лично обследованные члены родословной.
Графическое изображение родословной может быть вертикально-горизонтальным или расположенным по кругу (в
случае многочисленных данных). Схема родословной сопровождается описанием обозначений под рисунком
(легендой).
Третий этап – генетический анализ родословной.
Этот этап требует хороших знаний критериев
установленных типов наследования. Задача генетического анализа – установление наследственного характера
заболевания и типа наследования, выявление гетерозиготных носителей мутационного гена, установление генотипа
пробанда и , как заключение, прогнозирование потомства.
Анализ родословной рекомендуется проводить в следующей последовательности:
1.
Установление, является ли данный признак (заболевание) наследственным. Если признак встречается
несколько раз в разных поколениях (имеет семейный характер), то можно предполагать, что признак имеет
наследственную природу.
2.
Определение типа наследования признака. Для этого учитывают:
- во всех ли поколениях и как часто среди членов родословной встречается признак;
- одинакова ли частота признака у обоих полов и если нет, то у какого пола встречается чаще;
- детям какого пола передается признак от больного отца и от больной матери;
- есть ли семьи, в которых от больных родителей рождаются здоровые дети, или наоборот, от здоровых родителей
рождаются больные дети;
- какая часть потомства имеет наследуемый признак в семьях, где болен один из родителей.
80.
72. Генетика человека. Популяционно-статистический метод.Теоретическая часть. Методы, используемые для установления частот генов и генотипов в популяции, демонстрирующие характер их изменения под влиянием
окружающей среды и различных факторов популяционной динамики, называются популяционно-статистические.
С помощью этих методов можно:
- определить частоты генов, степень гомозиготности и полиморфизма;
- установить, как меняются частоты генов под действием отбора;
- выявить влияние факторов популяционной динамики на частоты тех или иных фенотипов и генотипов;
- проанализировать влияние факторов окружающей среды на экспрессию генов;
- определить степень межпопуляционного генетического разнообразия и вычислить расстояние между популяциями.
Генетическое изучение популяций человека предполагает знание их демографических характеристик (размер популяций, рождаемость, смертность,
возрастная структура, национальный состав), географических и климатических условий жизни, религиозных убеждений и др.
Это связано с некоторыми особенностями популяций человека, которые могут быть панмиксными (случайные браки) и инбредными (высокая частота
кровнородственных браков).
В популяции человека формирование субпопуляций связано с такими формами изоляции, которые свойственны только человеку: расовая, социальная
(специальное положение, экономические, этнические, языковые, административные особенности), религиозно-конфессиональная и идеологическая.
Для изучения достоверных результатов популяция должна быть достаточно большой, для генетических исследований оптимальным является размер
популяции с численностью от 0,5 до 5,0 млн. человек.
Для сбора материала используется обзорный метод и его различные модификации, т.е. можно исследовать всю наследственную патологию, или
отдельную группу заболеваний, или только одно заболевание, но изучая при этом все население выбранного региона.
Наследственные заболевания распределены по различным регионам земного шара, среди разных рас и народностей неравномерно, а знания о
распространении частот заболеваний и количестве гетерозиготных носителей в регионе имеет большое значение, т.к. способствует организации профилактических
мероприятий.
Популяционно-статистический метод позволяет определить, какие гены и аллели встречаются в популяции чаще или реже, а также оценить уровень генетического
разнообразия.
В целом, популяционно-статистический метод в генетике человека является важным инструментом для изучения генетической изменчивости, наследственности и
эволюции.
81.
73. Генетика человека. Близнецовый метод, сущность и значение.Генетика человека – это особый раздел генетики, который изучает особенности наследования признаков у человека, наследственные заболевания (медицинская
генетика), генетическую структуру популяций человека. Генетика человека является теоретической основой современной медицины и современного
здравоохранения.
Близнецовый метод
Метод предложен в 1875г. Гальтоном первоначально для оценки роли наследственности и среды в развитии психических свойств человека. В настоящее
время метод широко применяется в изучении наследственности и изменчивости человека для определения соотносительной роли наследственности и среды в
формировании различных признаков.
С помощью близнецового метода можно выявить наследственный характер признака, определить пенетрантность аллеля, оценить эффективность
действия на организм некоторых внешних факторов (лекарственных препаратов, обучения, воспитания).
Сущность метода заключается в сравнении проявления признака в разных группах близнецов при учете сходства или различия их генотипов.
Монозиготные близнецы (МБ) генетически идентичны, т.к. развиваются из одной зиготы, поэтому среди монозиготных близнецов наблюдается высокий процент
конкордантных пар, в которых признак развивается у обоих близнецов, что свидетельствует о генетической обусловленности признака. По признакам, в
формировании которых ведущая роль принадлежит факторам среды между близнецами наблюдается несовпадение или дискордантность. Количественную оценку
относительной роли наследственности и среды можно сделать на основе расчета коэффициента наследственности Н и коэффициента влияния среды Е. с помощью
формулы К. Хольцингера: Н=(КМБ-КДБ)/(100-КДБ), где КМБ – конкорданность признака (в %) для МБ; КДБ – то же для ДБ. Если Н=1, т.е. 100%, то можно считать, что
экспрессия признака определяется только генотипом индивидуума. Если Н=70%, то 70% - в формировании признака отводится наследственности, а 30% - факторам
среды.
Существуют таблицы конкордантности близнецов по различным признакам и заболеваниям.
Трудности близнецового метода связаны с низкой рождаемостью близнецов в популяциях (1:86 – 1:88), с идентификацией монозиготности близнецов, что важно для
достоверности выводов.
Для идентификации монозиготности применяют полисимптомный метод сравнения близнецов по многим признакам, методы основанные на
иммунологической идентичности близнецов по эритроцитарным антигенам (АВ0, МN, резусу), по сывороточным белкам, -глобулину), наиболее достоверный
критерий – трансплпнтационный тест с применением перекрестной пересадки кожи.
Близнецовый метод является одним из методов изучения наследственности в генетике человека.
Существуют два типа близнецов: однояйцевые (монозиготные) и двояйцевые (дизиготные). Однояйцевые близнецы образуются, когда одно оплодотворенное яйцо
делится на две части, в то время как двояйцевые близнецы образуются от разных яйц и оплодотворяются разными сперматозоидами.
Близнецовый метод используется для изучения генетического влияния на различные фенотипические характеристики и заболевания.
Использование близнецового метода в генетике человека помогает в понимании наследственности различных фенотипических свойств и может быть полезным
инструментом при изучении генетических механизмов, лежащих в основе различных болезней.
82.
74. Генетическая структура менделевской популяции. Закон Харди-Вайнберга.Генетическая структура популяций
Каждая популяция обладает собственной генетической структурой. Генетическая структура популяций определяется исходным соотношением аллелей, естественным отбором и элементарными эволюционными факторами (мутационный
процесс и давление мутаций, изоляция, популяционные волны, генетико-автоматические процессы, эффект основателя, миграции и др.). Для описания генетической структуры популяций используются понятия «аллелофонд» и «генофонд».
Аллелофонд. Аллелофонд популяции – это совокупность аллелей в популяции. Если рассматриваются два аллеля одного гена: А и а, то структура аллелофондаописывается уравнением: pA + qa = 1. В этом уравнении символом pA
обозначается относительная частота аллеля А, символом qa – относительная частота аллеляа.
Популяции, в которых структура аллелофонда остается относительно постоянной в течение длительного времени, называются стационарными.
Генофонд. Термин генофонд употребляется в разных значениях. Основоположник учения о генофонде и геногеографии Александр Сергеевич Серебровский называл генофондом «совокупность всех генов данного вида..., чтобы подчеркнуть
мысль о том, что в лице генофонда мы имеем такие же национальные богатства, как и в лице наших запасов угля, скрытых в наших недрах» (1928). Однако это выражение в настоящее время используется для определения генетического
потенциала, а генофондом называют совокупность всех генотипов в популяции.
При изучении природных популяций часто приходится сталкиваться с полным доминированием: фенотипы гомозигот АА и гетерозигот Аа неразличимы. Кроме того, в природе широко распространено полигенное определение признаков,
причем типы взаимодействия неаллельных генов (комплементарность, эпистаз, полимерия) не всегда известны. Поэтому на практике часто изучают не генофонд, а фенофонд популяций, то есть соотношение фенотипов. В настоящее время
развивается раздел генетики популяций, который называется фенетика популяций.
Закон Харди–Вайнберга – основной закон популяционной генетики
При статистической обработке материала основой для определения генетической структуры популяции является закон генетического равновесия Харди-Вайнберга, 1908г. Закон отражает закономерность, в соответствии с которой «В
популяции из бесконечно большого числа свободно скрещивающихся особей, в отсутствии мутаций, избирательной миграции организмов с различными генотипами и давления естественного отбора первоначальные частоты аллелей
сохраняются из поколения в поколение».
В основе сохранения частот аллелей лежат статистические закономерности случайных событий в больших выборках.
Для расчета частот генотипов, фенотипов и аллелей в диаллельной системе применяются формулы pA+qa=1 и (pA+qa)2=1 или p2AA+2pqAa+q2aa=1, где pA – частота доминантного аллеля, qa – частота рецессивного аллеля,
p2 – частота гомозиготной АА, 2pq – частота гетерозигот Аа. Закон Харди-Вайнберга описывает условия генетической стабильности популяции. Популяцию, генофонд которой не меняется в ряду поколений, называют Менделевской. В
природе эти популяции не встречаются, но их выделение имеет чисто теоретическое значение.
Применение закона Харди-Вайнберга
Одно из важнейших применений закона Харди-Вайнберга состоит в том, что он дает возможность рассчитать некоторые из частот генов и генотипов в том случае, когда не все генотипы могут быть идентифицированы вследствии
доминантности некоторых аллелей.
Пример 1: альбинизм у человека обусловлен редким рецессивным геном. Если аллель нормальной пигментации обозначить А, а аллель альбинизма а, то генотип альбиносов будет аа, а генотипы нормально пигментированных людей – АА и
Аа. Предположим, что в популяции людей (Европейской части) частота альбиносов составляет 1 на 10000. Согласно закону Харди-Вайнберга, в данной популяции частота гомозигот q2аа=1:10000=0,0001 (0,1%), а частота рецессивных гомозигот
qa=√(q^2 )=0,01. Частота доминантного аллеля рА=1-qa=1-0,01=0,99. Частота нормально пигментированных людей составляет р2АА=0,992=0,98(98%), а частота гетерозигот 2pqАа=2 0,99 0,1=0,198(1,98%).
Важное следствие из закона Харди-Вайнберга состоит в том, что редкие аллели присутствуют в популяции главным образом в гетерозиготном состоянии. Рассмотрим приведенный пример с альбинизмом (генотип аа). Частота альбиносов
равна 0,0001, а гетерозигот Аа 0,00198. Частота рецессивного аллеля у гетерозигот составляет половину частоты гетерозигот, т.е. 0,0099. Следовательно, в гетерозиготном состоянии содержится примерно в 100 раз больше рецессивных
аллелей, чем в гомозиготном состоянии. Таким образом, чем ниже частота рецессивного аллеля, тем большая доля этого аллеля присутствует в популяции в гетерозиготном состоянии.
Практическое значение закона Харди–Вайнберга
1. В здравоохранении – позволяет оценить популяционный риск генетически обусловленных заболеваний, поскольку каждая популяция обладает собственнымаллелофондом и, соответственно, разными частотами неблагоприятных аллелей.
Зная частоты рождения детей с наследственными заболеваниями, можно рассчитать структуру аллелофонда. В то же время, зная частоты неблагоприятных аллелей, можно предсказать риск рождения больного ребенка.
2. В селекции – позволяет выявить генетический потенциал исходного материала (природных популяций, а также сортов и пород народной селекции), поскольку разные сорта и породы характеризуются собственными аллелофондами,
которые могут быть рассчитаны с помощью закона Харди-Вайнберга. Если в исходном материале выявлена высокая частота требуемого аллеля, то можно ожидать быстрого получения желаемого результата при отборе. Если же частота
требуемого аллеля низка, то нужно или искать другой исходный материал, или вводить требуемый аллель из других популяций (сортов и пород).
3. В экологии – позволяет выявить влияние самых разнообразных факторов на популяции. Дело в том, что, оставаясь фенотипически однородной, популяция может существенно изменять свою генетическую структуру под воздействием
ионизирующего излучения, электромагнитных полей и других неблагоприятных факторов. По отклонениям фактических частот генотипов от расчетных величин можно установить эффект действия экологических факторов. (При этом нужно
строго соблюдать принцип единственного различия. Пусть изучается влияние содержания тяжелых металлов в почве на генетическую структуру популяций определенного вида растений. Тогда должны сравниваться две популяции,
обитающие в крайне сходных условиях. Единственное различие в условиях обитания должно заключаться в различном содержании определенного металла в почве).
83.
75. Морфофункциональная характеристика и классификация хромосом. Кариотип человека. Цитогенетический метод.Денверская и Парижская номенклатура кариотипа человека.
Различают четыре типа строения хромосом:
телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце);
акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
субметацентрические (с плечами неравной длины, напоминающие по форме букву L);
метацентрические (V-образные хромосомы, обладающие плечами равной длины).
Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода.
Кариоти́ п — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление
полного хромосомного набора (кариограммы).
У человека нормальный кариотип состоит из 46 хромосом.
Определение кариотипа
Внешний вид хромосом существенно меняется в течение клеточного цикла: в течение интерфазы хромосомы локализованы в ядре, как правило, деспирализованы и труднодоступны для наблюдения, поэтому для определения кариотипа используются клетки в одной из стадий их деления — метафазе митоза
Метафазная хромосома (спирализованный хроматин) состоит из двух хроматид. Форма определяется наличием первичной перетяжки - центромеры. Она разделяет хромосому на 2 плеча.
Расположение центромеры определяет основные формы хромосом:
- метацентрические, - субметацентрические, - акроцентрические, - телоцентрические.
Степень спирализации хромосом не одинакова. Участки хромосом со слабой спирализацией называют эухроматиновыми. Это зона высокой метаболической активности, где ДНК состоит из уникальных последовательностей. Зона с сильной спирализацией - гетерохроматиновыйучасток, способный к
транскрипции. Различают конститутивныйгетерохроматин-генетический инертный, не содержит генов, не переходит в эухроматин, а так же факультативный, который может переходить в активный эухроматин. Концевые отделы дистальных участков хромосом называют теломеры.
Хромосомы подразделяются на аутосомы (соматических клеток) и гетерохромосомы (половых клеток).
По предложению Левитского (1924) диплоидный набор соматических хромосом клетки был назван кариотипом. Он характеризуется числом, формой, размерами хромосом. Для описания хромосом кариотипа по предложению С.Г. Навашина их располагают в виде идиограммы - систематизированного
кариотипа. В 1960 году была предложена Денверская международная классификация хромосом, где хромосомы классифицированы по величине и расположению центромеры.
Согласно Парижской классификации хромосомы разделены на группы по их размерам и форме, а также линейной дифференцировке. (На Парижской конференции по стандартизации кариотипов вместо морфологических терминов «метацентрики» или «акро-центрики» в связи с разработкой новых методов
получения «по¬лосатых» хромосом предложена символика, в которой всем хро¬мосомам набора присваивается ранг (порядковый номер) по по¬рядку убывания величины и в обоих плечах каждой хромосомы (р — короткое плечо, q— длинное плечо) нумеруются участки плеч и полосы в каждом участке по
направлению от центромеры. Такая система обозначений позволяет детально описывать ано¬малии хромосом.)
Хромосомы обладают следующими свойствами (правила хромосом):
1. Индивидуальности - отличия негомологичных хромосом. 2. Парности. 3. Постоянством числа - характерным для каждого вида. 4. Непрерывности - способности к репродукции.
Цитологический метод основан на микроскопическом изучении хромосом в клетках человека. Цитогенетический метод широко применяется с 1956 года, когда Дж. Тио и Л. Леван установили, что в кариотипе человека 46 хромосом.
Цитогенетический метод основывается на данных о хромосомах. В 1960 году на научной конференции в Денвере была принята классификация идентифицируемых хромосом, в соответствии с которой им были даны номера, увеличивающиеся по мере уменьшения размеров хромосом. Эта
классификация была уточнена на конференции в Лондоне (1963) и Чикаго (1966). Применение цитогенетического метода позволяет изучать нормальную морфологию хромосом и кариотипа в целом, определять генетический пол организма, и, главное, диагностировать различные хромосомные болезни,
связанные с изменением числа хромосом или с нарушением структуры хромосом. Цитогенетический метод позволяет изучать процессы мутагенеза на уровне хромосом и кариотипа. Метод широко применяется в медико-генетическом консультировании для целей пренатальной диагностики хромосомных
болезней. В соматических клетках человека диплоидный набор хромосом, 2n=46, а в половых – гаплоидный n=23. При оплодотворении диплоидный набор хромосом восстанавливается. В хромосоме выделяют короткое (р) и длинное (q) плечи. Концы обоих плеч хромосомы называют теломерами. В метафазе
митоза хромосомы представлены двумя сестринскими хроматидами, соединенными центромерой. В центромере содержится вещество – кинетохор, участвующее в формировании нитей веретена при клеточном делении.
При изучении кариотипа определяют следующие морфометрические характеристики хромосом: Lа – абсолютная длина хромосомы в мкм; Lр – длина короткого плеча; Lg – длина длинного плеча. Iв – плечевой индекс, Iс – центромерный индекс, Lr – относительная длина хромосомы, Ih процент гетерохроматиновой зоны, Is – индекс спирализации.
По значению плечевого индекса определяется форма хромосом. При Iв 1-1,9 хромосома называется равноплечей (метацентрической), 2-4,9 – слабонеравноплечей (субметацентрической), 5 и более – акроцентрической или резко неравноплечей.
Для кариотипирования подбирают метафазные пластинки в количестве не менее 30 с одинаковым индексом спирализации.
На основании различий в длине выделены 23 пары хромосом. По форме в кариотипе человека имеются метацентрические, субметацентрические и акроцентрические хромосомы. Отнесение хромосом к той или иной группе производится на основе расчета центромерного индекса. На
основании размеров и комбинации плечевого и центромерного индексов хромосомы человека в соответствии с Международной Денверской классификацией (1960) сгруппированы в 7 групп, обохзначаемых буквами английского алфавита: A, B, C, D, E, F, G.
84.
76. Предмет и история эмбриологии. Преформизм и эпигенез.Как и многие другие естественные науки, эмбриология зародилась в античности. В трудах Аристотеля имеются довольно подробные описания развития куриного зародыша. В это же время возникли и две основные точки зрения на процессы
развития - преформизм и эпигенез . Эти два взгляда на развитие полностью сформировались к XVII в., и между ними началась борьба. Тогда в связи с появлением микроскопа стали накапливаться фактические данные о строении зародышей и
процессах развития разных организмов.
Голландец Ян Сваммердам изучил развитие и метаморфоз насекомых. Его соотечественник Антони ван Левенгук открыл партеногенез у тлей и наблюдал сперматозоиды человека. Итальянец Марчелло Мальпиги переисследовал развитие
куриного эмбриона и изучил микроскопическую анатомию многих органов и тканей животных и растений. Большинство сделанных наблюдений истолковывались их авторами с позиций преформизма (предсуществования). Согласно этой точке
зрения, в процессе развития не возникает ничего действительно нового, все части зародыша в уже сформированном виде присутствуют в яйце, и если их нельзя увидеть, то только из-за малых размеров; в дальнейшем происходит лишь их рост.
Так, Сваммердам считал, что внутри яйца бабочки спрятана гусеница, внутри гусеницы - куколка, а внутри куколки - взрослая бабочка. Отметим, что с точки зрения преформистов казалось маловероятным самозарождение организмов. Ясно, что
если зародыш предсуществует, он может находится только либо в яйцеклетке, либо в сперматозоиде. Поэтому преформисты разделились на два лагеря - овистов (от лат. ovum - яйцо) и анималькулистов (от лат. animal-culum - "зверек", одно из
названий сперматозоида). При этом оказывалось совершенно непонятным, как же передаются потомкам признаки второго родителя.
Доведенные до логического завершения взгляды преформистов привели к появлению "теории вложения", одним из авторов которой был математик Г.Лейбниц . Если в яйцеклетке есть полностью сформированный зародыш, то в его яичниках
должны быть сформированные яйцеклетки, в них - зародыши следующего поколения, и так до бесконечности. С точки зрения современной науки эти взгляды, конечно, совершенно нелепы.
Согласно точки зрения, получившей название эпигенеза, в ходе развития происходит постепенное усложнение формы; из бесструктурного, однородного материала яйца возникают различающиеся части зародыша. Автором термина "эпигенез"
был У.Гарвей , открывший круги кровообращения и выдвинувший в своей книге "О произрождении животных" знаменитый принцип "Все (живое) из яйца". Гарвей писал: "Ни одна часть плода не существует в яйце актуально, но все части
находятся в нем потенциально". Яйцо не содержит органов зародыша, а лишь "потенции" - задатки их развития.
В XVIII в. продолжал господствовать преформизм. Однако в 1759 г. в работе малоизвестного тогда ученого Каспара Фридриха Вольфа появились факты, прямо противоречащие преформистским воззрениям. Вольф доказывал, что такие органы
куриного зародыша, как нервная трубка и кишечник, первоначально закладываются в виде плоских пластов, а лишь затем сворачиваются в трубки. Таким образом, в ходе развития происходит формообразование. Однако даже эти четкие
наблюдения отвергались преформистами.
В первой половине XIX в. факты, противоречащие преформизму, уже стали общепризнанными. Так, было твердо установлено, что все органы позвоночных развиваются из трех первичных зародышевых пластов, или листков (см. " Органогенез ").
Согласно современной точке зрения, истина лежит где-то посередине между преформизмом и эпигенезом. Высказывание Гарвея оказалось гениальным предвидением. Генетическая программа развития, определяющая его ход, полностью
"преформирована" в оплодотворенной яйцеклетке. Кроме того, и цитоплазма яйца не бесструктурна - в разных ее участках содержатся вещества, предопределяющие ход развития частей зародыша (см. " Оплодотворение "), но, конечно,
никаких структур будущего зародыша в яйце нет.
В первой половине XIX в. большие успехи делали систематика и сравнительная анатомия. Одним из главных методов в биологии стал сравнительный метод. Под влиянием этих успехов зародилась сравнительная эмбриология. Наиболее важные
результаты этого направления эмбриологии были получены российским ученым Карлом Бэром . Изучив эмбриональное развитие всех классов позвоночных, Бэр установил, что на некоторой довольно ранней стадии все эти зародыши наиболее
сходны между собой, а различия приобретаются в ходе дальнейшего развития. Это позволило Бэру сформулировать закон зародышевого сходства: зародыши животных, относящихся к одному типу, более сходны, чем взрослые формы.
Признаки типа появляются в ходе развития раньше, чем признаки класса, отряда и т.д.; последними появляются видовые признаки.
Новый импульс это направление исследований получило после выхода в свет книги Ч.Дарвина "О происхождении видов" и быстрого распространения идей эволюционизма. Зародышевое сходство стали рассматривать как показатель общности
происхождения, а изучение эмбрионального развития - как один из главных способов изучения филогении (путей эволюционного развития) организмов. Э.Геккель сформулировал биогенетический закон : "Онтогенез есть быстрое и краткое
повторение филогенеза (исторического развития вида)". Геккель утверждал, что филогенез есть причина онтогенеза: идивидуальное развитие полностью обусловлено историей развития вида. В дальнейшем эти взгляды были частично
отвергнуты наукой, а частично видоизменены и дополнены.
Особенно большой вклад в эволюционную эмбриологию внесли отечественные ученые А.О.Ковалевский и И.И.Мечников . Ковалевский изучил раннее развитие ланцетника - представителя хордовых - и доказал его сходство с развитием
асцидий и некоторых других беспозвоночных. Мечников изучил раннее развитие примитивных многоклеточных ( губок и кишечнополостных ) и доказал сходство ранних стадий развитая всех многоклеточных животных.
Увлечение восстановлением хода эволюции на основе эмбриологических данных привело к тому, что эмбриология во многом утратила самостоятельность, став частью сравнительной анатомии и теории эволюции. Однако в конце XIX в. вновь
усилился интерес к непосредственным причинам развития, его механизмам. Новому направлению одним из его основателей, Вильгельмом Ру , было дано название механики развития. В основу этого подхода Ру положил причинноаналитический метод, согласно которому, чтобы понять механизмы развития, надо разбить его на отдельные звенья, выяснить механизмы каждого звена и причинно-следственные связи между ними. Ру подчеркивал, что сделать это можно
только путем экспериментов. Например, следовало выяснить, может ли каждая часть зародыша развиваться самостоятельно или требует воздействий извне. Для этого можно изолировать часть зародыша, пересадить ее в другой участок и т.д.
Он утверждал также, что процессы развития нельзя объяснить без знания законов химии и физики. Эти законы лежат в основе функционирования живых клеток, а развитие, в свою очередь, складывается из небольшого числа разных форм
клеточной активности - роста, деления, миграции и т. д.
Предметом эмбриологии является изучение развития организмов, начиная с момента зачатия до полного формирования их структуры и функций. Эмбриология исследует процессы, которые происходят на различных стадиях эмбрионального
развития, а также механизмы, по которым разные клетки и ткани формируются и организуются в органы и системы организма.
Преформизм и эпигенез являются двумя основными теориями, объясняющими развитие организма.
Преформизм - это теория, согласно которой все органы и структуры будущего организма уже содержатся в зародыше в миниатюрной, заранее заданной форме, которая просто разворачивается в процессе развития. Приверженцы преформизма
считали, что эмбрионы представляют собой предопределенные "микроскопические человеческие существа", содержащиеся в сперматозоиде или яйцеклетке.
Эпигенез - это теория, согласно которой организм формируется постепенно, поэтапно, при активном взаимодействии генов с окружающей средой. Приверженцы эпигенеза считают, что организм строится посредством последовательных
изменений и дифференцировки клеток, а окружающая среда и взаимодействие с ней играют существенную роль в определении формы и функции органов.
85.
77. Онтогенез. Периодизация онтогенеза. Видоизменения онтогенеза: эмбрионизация, деэмбрионизация, ноотения.Онтогенез - это процесс развития
организма от оплодотворения до
полноценной зрелости. Онтогенез
состоит из нескольких стадий и
подразделяется на периоды.
Периодизация онтогенеза:
1. Эмбриональный период
2. Перинатальный период
3. Детский период
4. Пубертатный период
5. Взрослый период
Видоизменения онтогенеза:
1. Эмбрионизация: Это процесс, когда
некоторые аспекты развития организма
сохраняются или повторяются на более
поздних стадиях эволюции.
2. Деэмбрионизация: Это процесс, когда
некоторые аспекты развития зародыша
или эмбриона ослабляются или исчезают,
не оставляя следов во взрослой особи.
3. Ноотения: Это процесс развития
нервной системы и мозга. Ноотения
включает в себя формирование, рост и
созревание нервных клеток и связей
между ними на протяжении всего
периода онтогенеза.
Диапауза. В эмбриональном периоде ряда позвоночных существует диапауза, т.е. остановка развития на более или менее
продолжительный период. Она имеет приспособительное значение. Так, у сумчатых и грызунов развитие эмбрионов останавливается в
случае совпадения по времени развития нового потомства со вскармливанием самкой предыдущего помета. Это лактационная пауза,
продолжающаяся обычно несколько суток. В других отрядах млекопитающих наблюдаются длительные, до 7 мес. обязательные диапаузы.
К примеру, у соболя оплодотворение происходит в июле. Оплодотворенное яйцо дробится, процесс развития доходит до стадии
бластоцисты и тормозится. Имплантация и дальнейшее развитие начинаются только в марте. Описанная диапауза возникла в эволюции
названного вида животных путем отбора на рождение потомства в сезон, наиболее благоприятный для вскармливания.
Деэмбрионизация. Под этим термином понимают сильное укорочение собственно эмбрионального периода, протекающего под
оболочками яйца. Деэмбрионизация, наблюдающаяся у плацентарных млекопитающих, сочетается с резким уменьшением желтка в их
яйцеклетках и установлением связи с материнским организмом через плаценту.
Эмбрионизация. Процесс эмбрионизации заключается в удлинении времени защищенности зародыша от внешней среды благодаря
зародышевым оболочкам и материнскому организму. Зародыш в это время проходит периоды, соответствующие собственно
эмбриональному и личиночному. Развивающийся организм защищен до момента формирования ювенильной стадии. После рождения не
происходит существенной перестройки организма и развитие идет прямое, т.е. без метаморфоза.
Эмбрионизация развития особенно проявилась в процессе эволюции наземных позвоночных. Уже у многих земноводных, например у
рогатой лягушки Соломоновых островов, все развитие происходит под яйцевыми оболочками, из которых вылупляется уже претерпевшее
метаморфоз животное. Полная Эмбрионизация имеет место у пресмыкающихся и птиц в связи с переходом их к наземному
существованию. Особое изменение эмбрионального и личиночного периодов произошло у плацентарных млекопитающих. Наряду с
деэмбрионизацией, о чем говорилось выше, у них произошла максимальная Эмбрионизация, но не под оболочками яйца, а
внутриутробно. Период свободного личиночного развития полностью исчез.
Проходящие под яйцевыми оболочками или внутриутробно предплодный и плодный периоды развития вместе равнозначны личиночному
периоду. Они следуют непосредственно за эмбриональным периодом, включают средние и более поздние фазы морфогенеза,
характеризуются наличием провизорных органов, переходом к активному питанию, началом функционирования органов чувств (слуха,
вкуса и др.).
К провизорным органам у млекопитающих относят зародышевые оболочки, некоторые отделы кровеносной системы, временные
структуры кожного покрова. У яйцекладущих и сумчатых млекопитающих зародыш на предплодных этапах уже переходит к питанию
молоком матери, а у птиц и плацентарных млекопитающих — заглатывает амниотическую жидкость. Начало функционирования органа
слуха описано у зародышей страуса эму, которые энергично реагируют на крик взрослых птиц уже на последней четверти инкубационного
периода.
Период метаморфоза у наземных позвоночных соответственно упрощается, так как плод в значительной степени похож на взрослую особь.
Однако отбрасывание зародышевых оболочек, открытие эмбриональных заращений, изменения в органах дыхания, кровообращения и в
коже в момент родов и в первую неделю после родов представляют собой перестройки, соответствующие метаморфозу.
Неотения. Это выпадение ювенильного и взрослого периодов развития. В эволюции многих групп позвоночных наблюдается тенденция к
более раннему достижению половой зрелости. Наиболее ярко эта тенденция выражена у хвостатых земноводных. Так, в семействе
Ambistomatidae личинки (аксолотли) приобрели возможность размножаться до наступления периода метаморфоза и превращения во
взрослую стадию, однако способности превращаться во взрослую стадию они полностью не потеряли.
Известны популяции таких животных, в которых сосуществуют неотенические и взрослые формы. Это частичная неотения. При полной
неотении ювенильный и взрослый периоды выпадают полностью и о его существовании у предков судят только по близкородственным
видам. Некоторые ученые полагают, что на основе преобразования личиночных стадий мог возникнуть ряд крупных таксонов. Считают, что
неотения могла явиться основным направлением эволюции бескилевых бегающих птиц и человека.
86.
78. Гаметогенез. Сперматогенез. Оогенез, особенности строения половых клеток.Гаметогенез — процесс образования яйцеклеток (овогенез) и сперматозоидов (сперматогенез) — подразделяется на ряд
стадий (рис. 5.4).
Гаметогенез имеет несколько стадий. Сходство сперматогенеза и овогенеза как раз и заключается в том, что три стадии у
них одинаковы.
1. Стадия размножения. Первичные клетки на этой стадии называются сперматогониями и овогониями, из них в
последующем образуются мужские и женские половые клетки. Половые клетки несколько раз делятся путем митоза, и
количество их значительно возрастает. Сперматогонии размножаются у мужчины в течение всего репродуктивного
периода, а размножение овогоний происходит в эмбриональном периоде и наиболее интенсивно происходит во 2 - 5
месяц внутриутробного развития.
2. Стадия роста. В этот период клетки значительно увеличиваются в размерах. Сперматогонии и овогонии превращаются в
сперматоциты и овоциты I порядка. Овоциты I порядка достигают больших размеров, поскольку накапливают питательные
вещества.
3. Стадия созревания. На этой стадии происходят два следующих друг за другом деления - мейоз I и мейоз II. После первого
деления образуются сперматоциты и овоциты II порядка, а после второго деления - сперматиды и зрелые яйцеклетки с
тремя полярными тельцами, которые в процессе размножения не участвуют и погибают. При созревании один
сперматоцит I порядка дает четыре сперматиды, а один овоцит I порядка образует одну яйцеклетку и три полярных тельца.
Эти особенности сперматогенеза и овогенеза имеют биологический смысл, который связан с разным назначением мужских
и женских гамет. Неравномерное деление клеток при овогенезе (меньше) обеспечивает формирование крупной
яйцеклетки, в ней накапливается большее количество питательных веществ, так как из оплодотворенного яйца будет
развиваться новый организм.
При сравнительной характеристике овогенеза и сперматогенеза можно заметить, что сперматозоидов образуется
значительно больше, и это также имеет биологический смысл.
Яйцеклетку достигает только один сперматозоид, проникает в нее и доставляет свой набор хромосом. Остальные же в
процессе поиска яйцеклетки массово погибают.
При сравнении овогенеза и сперматогенеза становится понятным, почему сперматозоидам нет необходимости в запасании
питательных веществ - их существование кратковременно, а подвижность должна быть высокой.
4. Стадия формирования. Она характерна только для сперматогенеза. Незрелая сперматида превращается в сперматозоид,
приобретая свойственный ему вид. Образование сперматозоидов у мужчин начинается только в период полового
созревания и происходит в течение всего года. Период развития сперматогоний в зрелые сперматозоиды составляет 74
дня.
признаки
сперматогенез
овогенез
Половые
железы,
половые
клетки
Яички,
сперматозоиды
Яичники,
55 мк
130-169 мк
подвижные
неподвижные
округлые со
жгутиком
округлые
Характерные
особенности
половых
клеток:
относительные
размеры
- подвижность
- форма
яйцеклетки
имеется
отсутствует
наличие
питательных
веществ
Особенности
развития
половых
на
клеток
разных
стадиях:
путем митоза
образуются
сперматоциты;
путем митоза
образуются
овоциты;
стадия
размножения
увеличиваются
в размерах;
увеличиваются
в размерах;
- стадия роста
путем мейоза
образуются
гаплоидные
сперматиды,
которых
их
формируются
сперматозоиды
путем мейоза
образуются
гаплоидные
овоциты, их
которых
формируется
яйцеклетка
стадия
созревания
87.
78. Гаметогенез. Сперматогенез. Оогенез, особенности строения половых клеток.СТРОЕНИЕ ПОЛОВЫХ КЛЕТОК (ГАМЕТ)
Гаметы представляют собой высокодифференцированные клетки. В про¬цессе эволюции они приобрели приспособления для выполнения специ-фических функций Ядра как мужских, так и женских гамет в равной мере содержат
наследственную информацию, необходимую для развития организма Но дру¬гие функции яйцеклетки и сперматозоида раз¬личны, поэтому по строению они резко отли¬чаются,
Яйцеклетки неподвижны, имеют шарообраз¬ную или слегка вытянутую форму (рис. 34). В яйце содержатся все типичные клеточные «органоиды, но строение его отличается от дру¬гих клеток, так как приспособлено для реали¬зации
возможности развития целого организ¬ма. Размеры яйцеклеток значительно крупнее, чем соматических. Внутриклеточная структура цитоплазмы в яйца спе¬цифична для каждого вида животных, чем обеспечиваются видовые (а нередко и
индивидуальные) особенности развития. В яйцах содер¬жится ряд веществ, необходимых для развития зародыша. К их числу относится питательный материал (желток). У некоторых видов живот¬ных накапливается столько желтка в яйцах, что
они могут быть видимы невооруженным глазом. Таковы икринки рыб и земноводных, яйца реп¬тилий и птиц. Из современных животных наиболее крупные яйца у сель¬девой акулы - 29 см в диаметре; диаметр яйца страуса 10,5 см, куриЦЫ 3,5 см. Небольшие размеры имеют яйца животных, у которых раз¬вивающийся зародыш получает питание из окружающей среды, на¬пример у высших млекопитающих. Диаметр яйца мыши 60 ц., коровы 100 ц. Яйцо человека в поперечнике
имеет 130-200 ц.
Сперматозоиды обладают способностью к движению, чем в известной мере обеспечивается возможность встречи гамет. По внешней морфоло¬гии и малому количеству цитоплазмы сперматозоиды резко отличаются от всех других клеток, но
все основные органоиды в них имеются.
Типичный сперматозоид (рис. 35) имеет головку, шейку и хвостовую нить. На переднем конце головки расположена акросомя, состоящая из видоизмененного комплекса Гольджи. Основную массу головки зани¬мает ядро. В шейке находится
центриоль и спиральная нить, образован¬ная митохондриями.
При исследовании сперматозоидов под электронным микроскопом об¬наружено, что протоплазма головки сперматозоида имеет не коллоидное, а жидко-кристаллическое состояние. Этим достигается' устойчивость сперматозоидов к
неблагоприятным влияниям внешней среды. Напри¬мер, они в меньшей степени повреждаются ионизирующей радиацией по сравнению с незрелыми половыми клетками.
Размеры сперматозоидов всегда микроскопические. Наиболее крупные сперматозоиды у тритона - около 500 ц,, у домашних животных: собаки,. быка, лошади, барана - от 40 до 75 ц. Размеры сперматозоидов чело¬века колеблются в пределах
52-70 ц. Все сперматозоиды несут одно¬именный (отрицательный) электрический заряд, что препятствует их склеиванию. Число сперматозоидов, образуемых животными, колоссаль¬но. Например, при половом акте собака выделяет около 60
млн. сперма¬тозоидов, баран - до 2 млрд., жеребец - около 10 Млрд. человек - около 200 млн.
Процесс формирования половых клеток (гамет) известен под общим» названием гаметогенеза. Он характеризуется рядом весьма важных био-логических процессов и протекает несколько по-разному при созрева¬нии сперматозоидов
(сперматогенез) и яиц (овогенез).
Гаметогенез - это процесс образования половых клеток (гамет) - сперматозоидов у мужчин и яйцеклеток у женщин. Сперматогенез происходит в мужских половых железах (яичках), а оогенез - в женских половых железах (яичниках).
Сперматогенез:
- Сперматогенез начинается при половом созревании мальчиков в подростковом возрасте и продолжается вплоть до старости.
- В процессе сперматогенеза происходит многократное подразделение половых клеток, называемых сперматогониями, последующее превращение их в сперматоциты, и затем в сперматиды.
- Сперматиды затем проходят процесс дифференцировки и превращения в сперматозоиды.
- Сперматогенез происходит в специальных структурах яичек, называемых половыми тубулями.
Оогенез:
- Оогенез начинается еще до рождения девочек и прекращается во время месячных циклов после достижения половой зрелости.
- В процессе оогенеза формируются ооциты (первичные яйцеклетки), которые затем превращаются в вторичные ооциты и, при оплодотворении, в оосферы (зрелые яйцеклетки).
- Оогенез протекает в специальных микроскопических мешочках в яичниках, называемых фолликулами.
Особенности строения половых клеток:
- Сперматозоиды являются мужскими половыми клетками и имеют форму длинного хвостика, который позволяет им плавать и перемещаться к яйцеклетке. Они содержат половые хромосомы X или Y.
- Яйцеклетки (ооциты) являются женскими половыми клетками и имеют крупный размер и форму сферы. Они содержат половые хромосомы X.
88.
79. Генетическая сущность оплодотворения. Нарушения оплодотворения, нерегулярные типы оплодотворения.Генетическая сущность
оплодотворения заключается в
объединении генетического
материала мужской и женской
клеток для создания зародыша.
Оплодотворение происходит, когда
сперматозоиды, содержащие
половые хромосомы X или Y,
соединяются с яйцеклеткой,
содержащей только половую
хромосому X.
Однако некоторые нарушения
оплодотворения могут привести к
нерегулярным типам
оплодотворения:
1. Нежелательная беременность
2. Бесплодие
3. Эктопическая беременность
4. Генетические аномалии
Все эти нарушения оплодотворения
требуют диагностики и лечения со
стороны медицинских
специалистов, таких как генетики
или репродуктологи.
Оплодотворение представляет собой процесс слияния сперматозоида с яйцеклеткой, в результате чего возникает диплоидная зигота;
каждая пара хромосом в ней представлена одной отцовской и другой материнской. Сущность оплодотворения заключается в
восстановлении диплоидного набора хромосом и в объединении наследственного материала обоих родителей, в результате чего потомство,
соединяющее в себе полезные признаки отца и матери, более жизнеспособно.
Нарушение оплодотворения, его последствия.
Оплодотворение - одно из звеньев биологического существования вида. Этому предшествует длительная и сложная подготовка двух
особей, во время которой они подвергаются разнообразной действия окружающей среды, что отрицательно повлиять на процесс
оплодотворения.
Яйцеклетка и сперматозоид имеют ограниченную продолжительность жизни и еще меньшую продолжительность способности к
оплодотворению. Так, у млекопитающих, и у человека частности, освобождена из яичника яйцеклетка сохраняет способность к
оплодотворению в течение 24 ч. Нарушение этого временного срока неизбежно приведет к потере способности к оплодотворению.
Сперматозоиды мужчины в половых путях женщины остаются подвижными более 4 суток, но оплодотворяющей способность они теряют
уже через 1 - 2 суток. С увеличением длительности во времени незащищенные клетки испытывают негативного влияния различных
факторов.
Последние могут вызывать нарушения восходящего состояния генофонда гамет, что неизбежно приведет к незапрограммированным
отклонениям развития зиготы с соответствующими последствиями для вида в целом.
Скорость движения сперматозоидов, в обычных условиях составляет 1,5-3 мм / мин. Разное отклонение от такого поступательного
перемещение вызывает потерю способности к оплодотворению. К этому приводит также изменение рН среды влагалища, воспалительные
явления и др. В эякуляте мужчины в среднем содержится 350 млн сперматозоидов, способных к оплодотворению. Если количество
сперматозоидов меньше 150 млн (или меньше 60 млн в 1 мл), то вероятность оплодотворения резко снижается. Итак, чрезмерная
концентрация сперматозоидов в эякуляте имеет исключительное значение в механизме оплодотворения.
Нарушение оплодотворения возникает при патологических изменениях морфологии сперматозоидов. На биологическую полноценность
гамет существенно влияет срок пребывания их в половых путях женщины. Так, перезревания сперматозоидов и яйцеклеток в женском
половом тракте при различных причинах обусловливает рост частоты хромосомных аберраций в абортированных плодах.
Нерегулярные типы полового размножения.
Классификация нерегулярных типов полового размножения.
К нерегулярным типам полового размножения можно отнести партеногенетическое, гиногенетическое и андрогенетическое размножение
животных и растений (рис. 27).
Партеногенез — это развитие зародыша из неоплодотворенной яйцеклетки. Явление естественного партеногенеза свойственно низшим
ракообразным, коловраткам, перепончатокрылым (пчелам, осам) и др. Известен он также у птиц (индейки). Партеногенез можно
стимулировать искусственно, вызывая активацию неоплодотворенных яиц путем воздействия различными агентами.
Различают партеногенез соматический, или диплоидный, и генеративный, или гаплоидный. При соматическом партеногенезе яйцеклетка не
претерпевает редукционного деления или если и претерпевает, то два гаплоидных ядра, сливаясь вместе, восстанавливают диплоидный
набор хромосом (автокариогамия) ; таким образом в клетках тканей зародыша сохраняется диплоидный набор хромосом.
При генеративном партеногенезе зародыш развивается из гаплоидной яйцеклетки. Например, у медоносной пчелы (Apis mellifera) трутни
развиваются из неоплодотворенных гаплоидных яиц путем партеногенеза.
89.
80. Оплодотворение и ооплазматическая сегрегация.Оплодотворение - это процесс, при Половой процесс, или оплодотворение, или амфимиксис (др.-греч. ἀμφι- — приставка со значением
двойственности и μῖξις — смешение) — процесс слияния гаплоидных половых клеток, или гамет,
котором сперматозоиды (мужские обоюдности,
приводящий к образованию диплоидной клетки зиготы, которая способна развиваться в новый организм.
репродуктивные клетки)
При образовании половых клеток в мейозе возникают гаметы с разным сочетанием хромосом, поэтому после
оплодотворения новые организмы могут сочетать в себе признаки обоих родителей в самых различных
соединяются с яйцеклеткой
В результате этого происходит колоссальное увеличение наследственного разнообразия
(женской репродуктивной клеткой), комбинациях.
организмов.
чтобы образовать зиготу, первую
Половой процесс закономерно встречается в жизненном цикле всех организмов, у которых отмечен мейоз. Мейоз
стадию развития нового организма. приводит к уменьшению числа хромосом в два раза (переход от диплоидного состояния к гаплоидному), половой
процесс — к восстановлению числа хромосом (переход от гаплоидного состояния к диплоидному).
Симметричная ооплазматическая
Различают несколько форм полового процесса:
изогамия — гаметы не отличаются друг от друга по размерам, подвижны, жгутиковые или амебоидные;
сегрегация происходит в случае,
анизогамия(Гетерогамия) — гаметы отличаются друг от друга по размерам, но оба типа гамет
когда ооплазма равномерно
и микрогаметы) подвижны и имеют жгутики;
распределяется по всей зиготе, без (макрогаметы
оогамия — одна из гамет (яйцеклетка) значительно крупнее другой, неподвижна, деления мейоза,
выраженной асимметрии. Этот
приводящие к её образованию, резко асимметричны (вместо четырёх клеток формируется одна яйцеклетка и
два абортивных «полярных тельца»); другая (спермий, или сперматозоид) подвижна, обычно жгутиковая
процесс может быть связан с
или амебоидная.
равномерным распределением
молекул и структур внутри
СЕГРЕГАЦИЯ ооплазматическая возникновение локальных различий в свойствах ооплазмы,
осуществляющееся в периоды роста и созревания ооцита, а также в оплодотворённом яйце. С.- основа для
зародыша.
последующей дифференцировки зародыша: в процессе дробления яйца участки ооплазмы, различающиеся по
своим свойствам, попадают в разные бластомеры; взаимодействие с ними одинаковых по своим потенциям ядер
Ооплазматическая сегрегация
дробления приводит к дифференциальной активации генома. У разных животных С. наступает неодновременно и
является важным механизмом
бывает выражена в разной степени. Наиболее ярко она проявляется у животных с мозаичным типом развития
раннего развития организма и
(Мозаичные яйца), но наблюдается и у животных с регуляционным типом развития (Регуляционные яйца).
способствует формированию
Примеры С.: образование полярных плазм у моллюсков, концентрация РНК в будущем спинном полушарии яйца
млекопитающих.
различных клеточных типов и
структур в разных областях зиготы,
что далее определяет дальнейшее
развитие эмбриона.
90.
81. Дробление. Нарушения дробления.Дробление. Образование морулы. Дробление представляет собой начальный период развития зиготы (оплодотворенного яйца). Поскольку
яйцеклетки обладают центриолями, то оно заключается в делении зиготы путем митоза, которое начинается, например у человека, через 30
часов после осеменения. У человека деление начинается с движения оплодотворенной яйцеклетки по фаллопиевой трубе и заключается в
появлении на поверхности яйцеклетки борозды. Первая борозда приводит к образованию двух клеток -- двух бластомеров, вторая -- четырех
бластомеров, третья -- восьми бластомеров и т. д. Группа клеток, образовавшаяся в результате последовательных дроблений зиготы, получила
название морулы (от греч. morum -- тутовая ягода).
Стадию морулы проходят все многоклеточные животные, размножающиеся половым путем. В зависимости от видовой принадлежности
деление идет по-разному. Различают радиальное (позвоночные, иглокожие), билатеральное (грибневики, некоторые хордовые) и спиральное
дробление (немертины, кольчатые черви, многие моллюски), причем эти формы дробления зависят от плоскостей дробления. Поэтому их
морулы состоят из разного количества клеток. Кроме того, из части клеток образуется структура, называемая трофобластом, клетки которого
питают зародыш, а благодаря ферментам обеспечивают также внедрение последнего в стенку матки. У человека прикрепление морулы к
стенке матки происходит на 7-й день после оплодотворения. Позднее клетки трофобласта отслаиваются от зародыша и образуют пузырек,
который заполняется жидкостью тканей матки.
Характерная особенность дробления заключается в том, что при нем значительного роста клеток не происходит. Поэтому биологическое
значение этой стадии заключается в том, что из крупной клетки, которой является яйцеклетка, образуются более мелкие клетки, в которых
уменьшено отношение цитоплазмы к ядру. В результате этого происходит изменение топологии цитоплазматических комплексов в
бластомерах, что создает новое цитоплазматическое окружение для ядер.
Дробление зиготы завершается образованием многоклеточной структуры, получившей название бластулы (от греч. blastos -- росток). Эта
структура имеет форму пузырька, состоящего из одного слоя клеток, называемого бластодермой. Теперь эти клетки называют
эмбриональными. По размерам бластула сходна с яйцеклеткой. В период дробления увеличивается количество ядер, возрастает общее
количество ДНК. В конце стадии бластулы синтезируется также небольшое количество мРНК и тРНК, но новые рибосомы и рибосомная РНК до
начала гаструляции еще не обнаруживаются, либо если обнаруживаются, то в ничтожных количествах.
Нарушения дробления могут возникать в различных ситуациях, их причины могут быть разными. Вот несколько примеров нарушений
дробления в разных контекстах:
1. Математика
2. Химия
3. Физика
4. Технические системы
5. Логистика и управление процессами
91.
82. Гаструляция и органогенез. Возможные нарушения.Гаструляция и органогенез являются двумя ключевыми процессами в развитии эмбриона. Гаструляция происходит после зиготы, и в результате образуется гаструла - многоклеточный эмбрион,
состоящий из трех зародышевых листков: эктодермы, мезодермы и эндодермы.
Органогенез - это процесс формирования и дифференциации органов и тканей из зародышевых листков. В этот период развиваются все основные органы и системы организма.
Возможные нарушения в процессе гаструляции и органогенеза могут привести к различным врожденным аномалиям и дефектам развития. Некоторые из них могут быть следующими:
1. Спина бифида
2. Гидроцефалия
3. Гастросхизис
4. Аненцефалия
5. Кластерная шишка
6. Расщепление губы и/или неба
Гаструляция — это процесс образования зародышевых листков. Гаструляция у человека происходит в два этапа. Первый этап идет путем деляминации (расщепления), а второй — путем
миграции.
В процессе первого этапа образуются два зародышевых листка (экто- и энтодерма), два провизорных органа (амнион и желточный мешок). Кроме того, непосредственно перед началом первого
этапа происходит образование такого провизорного органа, как хорион. Формирование хориона — это второй этап в образовании плаценты.
Во время второго этапа образуется еще один зародышевый листок — мезодерма и его производное — мезенхима, провизорный орган — аллантоис и идет дальнейшее образование еще одного
провизорного органа — плаценты: формируются третичные ворсинки хориона, которые в последующем соединяются с decidua basalis и формируют плаценту. Образуются осевые органы — хорда,
нервная трубка, кишечная трубка, мезодерма.
Первый этап гаструляции
Клетки эмбриобласта подразделяются на две части (Рис. Д. Т3). Одна часть представляет собой пласт клеток, расположенных в один ряд. Эта группа клеток получает название зародышевого диска
(щитка). Все остальные клетки эмбриобласта превращаются во внезародышевую мезенхиму (внезародышевую мезодерму).
Образование хориона Внезародышевая мезенхима подрастает к трофобласту, и получаются вторичные ворсинки хориона. Хорион — это трофобласт + внезародышевая мезенхима. Вторичные
ворсинки хориона состоят из трофобласта и внезародышевой мезенхимы (Рис. Д).
бразование эктодермы и энтодермы Клетки зародышевого диска делятся в тангенциальной плоскости, то есть происходит его расщепление (деляминация) на два слоя (Рис. Е, Т4). В результате
этого зародышевый диск уже состоит из двух слоев клеток. Верхний слой клеток — это эктодерма (эпибласт), нижний слой — энтодерма (гипобласт).
Образование желточного мешка Под энтодермой скапливается жидкость, которая отодвигает внезародышевую мезенхиму. В результате этого образуется пузырек, заполненный жидкостью,
называемый желточным пузырьком (Рис. Ж, Т4). Энтодерма разрастается за пределы зародышевого диска, обрастает поверхность желточного пузырька и получается желточный мешок (Рис. З, И,
Т5). Его стенка состоит из внезародышевой энтодермы и внезародышевой мезенхимы. Внезародышевая энтодерма называется внезародышевой, так как она вышла за пределы зародышевого
диска.
Образование амниона Клетки эктодермы выселяются из зародышевого диска, отодвигая внезародышевую мезенхиму (Рис. З, Т4). Между клетками эктодермы скапливается жидкость. Она
раздвигает выселившиеся из зародышевого диска клетки эктодермы. В результате образуется одна заполненная жидкостью полость, по периферии которой располагаются клетки
внезародышевой эктодермы и внезародышевой мезенхимы (Рис. З, И, Т5). Это — амнион. Стенка амниона состоит из внезародышевой эктодермы и внезародышевой мезенхимы.
Внезародышевая эктодерма называется внезародышевой потому, что она находится за пределами зародышевого диска.
92.
82. Гаструляция и органогенез. Возможные нарушения.Второй этап гаструляции
В эктодерме клетки начинают двигаться (мигрировать) с двух сторон от головного конца зародыша к каудальному (хвостовому). В области каудального конца клеточные потоки встречаются и начинают двигаться кпереди. При движении
клеток эктодермы в срединной части образуется нагромождение клеток, которое получает название первичной полоски. В средней части эктодермы движение клеток останавливается, и в передней части этой полоски имеется еще большее
нагромождение клеток, которое получает название первичного узелка.
Образование мезодермы происходит из клеток первичной полоски. Клетки первичной полоски, образовавшейся в эктодерме, прорастают в пространство между экто- и энтодермой и там разрастаются, образуя мезодерму (Рис. К, Т6).
Образование хорды происходит из клеток первичного узелка. Клетки первичного узелка, образующегося в эктодерме, прорастают в пространство между экто- и энтодермой, там разрастаются вперед и назад и образуют хорду (Рис. К, Т6).
Образование нервной трубки Уже после образования мезодермы и хорды в срединной части эктодермы образуется углубление — нервная пластинка (Рис. Л). Нервная пластинка погружается под эктодерму, а края эктодермы смыкаются, и
образуется нервная трубка (Рис. М).
Образование кишечной трубки происходит в процессе формирования туловищной складки. Туловищная складка формируется путем подгибания всех имеющихся зародышевых листков под тело зародыша (Рис. Л, М, Н, Т7). В результате
этого кишечная (зародышевая) энтодерма отделяется от желточного мешка (желточная энтодерма) (Рис. О). Кишечная трубка остается связанной с желточным мешком только в одном небольшом участке — желточном протоке (стебельке).
Часть желточного протока может остаться после рождения в виде дивертикула подвздошной кишки.
Дифференцировка мезодермы и образование мезенхимы Сразу после своего образования мезодерма подразделяется на два главных отдела (Рис. Л-С, Т6): сомиты — спинной отдел и спланхнотом — брюшной отдел. Между сомитами и
спланхнотомом имеется еще один отдел — сегментная ножка, с помощью которой они соединяются (Рис. С). Сомиты разделяются на три части: дерматом, склеротом, миотом. Дерматом дает начало мезенхиме дерматома, из нее
образуется дерма — собственно кожа. Миотом является источником поперечнополосатых мышц. Из склеротома образуется мезенхима склеротома, которая дает начало костям и хрящам. Спланхнотом делится на висцеральный и
париетальный листки, между которыми находится вторичная полость тела — целом (Рис. С). Висцеральный и париетальный листки дают начало висцеральным и париетальным серозным оболочкам. Кроме того, из висцерального листка
спланхнотома выселяются клетки, которые дают начало мезенхиме спланхнотома. Из нее развивается соединительная ткань. Таким образом, мезенхима образуется как минимум из трех источников — висцерального листка спланхнотома,
дерматома, склеротома. Из мезенхимы развивается вся соединительная ткань. Из сегментных ножек, находящихся в грудном отделе зародыша (первые 8–10 сегментов), закладывается предпочка и мезонефральный (вольфов) проток, из
которого образуются каналец придатка семенника и семявыносящий проток. Из сегментных ножек, находящихся в туловищных отделах зародыша, развивается первичная почка, которая сначала функционирует у зародыша, а потом из
канальцев первичной почки образуются прямые канальцы, канальцы сети семенника, выносящие канальцы придатка семенника. Сегментные ножки каудального отдела зародыша не отделяются одна от другой (не сегментируются), а
формируют нефрогенный тяж, из которого развивается окончательная почка.
Образование аллантоиса Аллантоис образуется как вентральный вырост передней части задней кишки. Он растет кпереди, дорастает до желточного стебелька, и в этой области выходит из тела зародыша (Рис. П, Т8). Стенка аллантоиса
состоит из энтодермы и внезародышевой мезенхимы. Урахус и верхушка мочевого пузыря — это остатки аллантоиса.
Образование плаценты (III этап) По аллантоису к вторичным ворсинкам хориона подрастают кровеносные сосуды, и такие ворсинки, состоящие уже из трофобласта, внезародышевой мезенхимы и кровеносных сосудов, называются
третичными ворсинками хориона (Рис. Р, Т8). Хорион прикрепляется к подлежащему участку слизистой оболочки матки — decidua basalis и вместе с ним формирует плаценту. Таким образом, плацента в буквальном смысле представляет
собой соединение третичных ворсинок хориона (плодная часть) и decidua basalis (материнская часть).
Органогенез — это формирование органов в процессе эмбрионального развития организма. Процесс формирования органов в течение онтогенеза (см.), т. е. онтогенетический органогенез, изучает эмбриология (см.), а на протяжении
исторического развития вида (филогенетический органогенез) — сравнительная анатомия.
Органогенез (от греч. organon — орган, genesis — развитие, образование)— процесс развития, или формирования, органов у зародыша человека и животных.
Органогенез следует за более ранними периодами зародышевого развития (см. Зародыш) — дроблением яйца, гаструляцией и наступает после того, как обособятся основные зачатки (закладки) органов и тканей. Органогенез протекает
параллельно с гистогенезом или развитием тканей. В отличие от тканей, из которых каждая имеет своим источником какой-либо один из эмбриональных зачатков, органы, как правило, возникают при участии нескольких (от двух до
четырех) различных зачатков (см. Зародышевые листки), дающих начало разным тканевым компонентам органа. Например, в составе стенки кишки эпителий, выстилающий полость органа, и железы развиваются из внутреннего
зародышевого листка — энтодермы (см.), соединительная ткань с сосудами и гладкая мышечная ткань — из мезенхимы (см.), мезотелий, покрывающий серозную оболочку кишки,— из висцерального листка спланхнотома, т. е. среднего
зародышевого листка — мезодермы, а нервы и ганглии органа — из неврального зачатка. Кожа образуется при участии наружного зародышевого листка — эктодермы из которой развиваются эпидермис и его производные (волосы, сальные
и потовые железы, ногти и др.), и дерматомов, из которых возникает мезенхима, дифференцирующаяся в соединительнотканную основу кожи (дерму). Нервы и нервные окончания в коже, как и всюду,— производные неврального зачатка.
Некоторые органы формируются из одного зачатка, например кость, кровеносные сосуды, лимфатические узлы — из мезенхимы; однако и здесь в закладку врастают производные зачатка нервной системы — нервные волокна,
формируются нервные окончания.
Если гистогенез заключается главным образом в размножении и специализации клеток, а также в образовании ими межклеточных веществ и других неклеточных структур, то основными процессами, лежащими в основе органогенеза,
являются образование зародышевыми листками складок, впячиваний, выпячиваний, утолщений, неравномерный рост, срастание или разделение (обособление), а также взаимное прорастание различных закладок.
У человека органогенез начинается с конца 3-й недели и завершается в основных чертах к 4-му месяцу внутриутробного развития. Однако развитие ряда провизорных (временных) органов зародыша — хориона, амниона, желточного мешка
— начинается уже с конца 1-й недели, а некоторые дефинитивные (окончательные) органы формируются позже других (например, лимфатические узлы— начиная с последних месяцев внутриутробного развития и до наступления полового
созревания).
Нарушение гаструляции - нарушение клеточных перемещений, приводящее к массовым уродствам развивающихся зародышей
93.
83. Дифференциация и интеграция в развитии. Аномалии и пороки развития.Основным принципом эволюции органических структур является принцип дифференциации. Дифференциация представляет собой разделение однородной структуры
на обособленные части, которые в силу различного положения, связей с другими органами и различных функций приобретают специфическое строение. Таким
образом, усложнение структуры всегда связано с усложнением функций и специализацией отдельных частей. Дифференцированная структура выполняет несколько
функций, и строение ее сложно.
Примером филогенетической дифференциации может являться эволюция кровеносной системы в типе хордовых. Так, у представителей подтипа бесчерепных она
построена очень просто: один круг кровообращения, отсутствие сердца и капилляров в системе жаберных артерий.
В надклассе рыб имеются двухкамерное сердце и жаберные капилляры. У земноводных впервые появляется разделение кровеносной системы на два круга
кровообращения, а сердце становится трехкамерным. Максимальная дифференциация характерна для кровеносной системы млекопитающих, сердце которых
четырехкамерное, а в сосудах достигается полное разобщение венозного и артериального кровотоков.
Отдельные части дифференцирующейся, ранее однородной структуры, специализируясь на выполнении одной функции, становятся функционально все более
зависимыми от других частей данной структуры и от организма в целом. Такое функциональное соподчинение отдельных компонентов системы в целостном организме
называют интеграцией.
Четырехкамерное сердце млекопитающих представляет собой пример высокоинтегрированной структуры: каждый отдел выполняет лишь свою специальную функцию,
не имеющую никакого смысла в отрыве от функций других отделов. Поэтому сердце снабжено автономной системой функциональной регуляции в виде
парасимпатического атриовентрикулярного нервного узла и при этом строго подчинено нейрогуморальной системе регуляции организма в целом.
Таким образом, одновременно с дифференциацией наблюдается и подчинение частей целостной системе организма, т.е. процесс интергации.
Пороки развития представляют собой стойкие морфологические изменения органа или организма в целом, выходящие за пределы вариаций нормы и возникающие
внутриутробно в результате нарушения развития зародыша либо плода, иногда — после рождения ребенка вследствие нарушения дальнейшего формирования
органов. Эти изменения вызывают нарушения соответствующих функций. Синонимами термина «пороки развития» являются «врожденные пороки», «аномалии
развития», «дисплазии».
Однако под аномалиями развития и дисплазиями понимают только такие пороки, при которых анатомические изменения не приводят к существенному нарушению
функций, например деформации ушных раковин, не обезображивающие лица больного и существенно не отражающиеся на восприятии звуков. Грубые пороки
развития, при которых обезображивается внешний облик ребенка, нередко называют уродствами. Однако термин «уродство» — понятие скорее социальное, чем
медицинское.
Дифференциация - это процесс, в результате которого клетки организма становятся специализированными и приобретают определенные функции. То есть, из
множества клеток появляются различные типы клеток, которые выполняют разные задачи в организме.
Интеграция - это процесс, в результате которого специализированные клетки и ткани организма взаимодействуют и работают вместе для выполнения различных
функций организма. Например, в организме различные системы (нервная, сердечно-сосудистая, дыхательная и т. д.) должны интегрироваться и сотрудничать друг с
другом для обеспечения правильной работы организма.
Аномалии и пороки развития возникают, когда процессы дифференциации и интеграции не протекают нормально. Это может быть вызвано генетическими мутациями,
воздействием внешних факторов или другими причинами. Аномалии развития могут проявляться как структурные или функциональные изменения в организме,
которые могут привести к различным заболеваниям, порокам или нарушениям функций органов.
94.
84. Роль наследственности и среды в онтогенезе.Онтогенез протекает в конкретных условиях окружающей среды, и на любом его этапе организм наитеснейшим образом взаимосвязан со средой. Под средой
понимают совокупность конкретных абиотических и биотических факторов (условий), в которых обитает данная особь (популяция, вид). Эти взаимосвязи организма
и среды складываются и изменяются в процессе эволюции. Развитие каждого конкретного организма - это, по сути, формирование фенотипа (совокупности внешних
и внутренних признаков), или реализация генотипа в конкретных условиях среды. Фенотип организма не только обусловлен генотипом, обеспечивающим
материальную преемственность между поколениями, но и зависит от факторов внешней среды, в которой формируется и существует данный организм.
В течение всего онтогенеза происходит взаимодействие между генотипом и факторами среды, которые в конечном счёте и детерминируют все биологические
признаки данного организма. При этом обе эти группы факторов имеют одинаково важное значение, хотя для отдельных признаков доминирующей может
выступать одна из двух групп факторов. Так, группы крови (фенотипический признак) имеют у человека исключительно генетическую природу: при любых условиях
среды данный генотип проявляется одинаково и обусловливает строго определённую группу крови. С другой стороны, существуют признаки, обусловленные
исключительно факторами среды. Например, количество эритроцитов в циркулирующей крови у людей с разнообразными генотипами прямо зависит от высоты
местности проживания над уровнем моря: с увеличением высоты их число у всех возрастает. Тем не менее сама способность к изменению числа эритроцитов в
зависимости от парциального давления кислорода в атмосферном воздухе обусловлена генетически. Однако подобные крайние случаи очень редки. В большинстве
случаев различия особей определяются факторами обеих групп - наследственными и средовыми. Так, различия в росте обусловлены как генетически, так и
конкретными средовыми факторами (климат, характер питания и т.п.).
Значительными могут быть влияния абиотических факторов, или условий среды (атмосферное давление, излучение, температура, влажность, газовый состав,
степень освещённости и др.). При снижении температуры с +20° до +15°С зародыши лягушки не могут развиваться дальше стадии нейрулы. Прекращение доступа
кислорода к эмбриону аскариды приостанавливает его развитие. Такие реакции позволяют характеризовать подобные изменения внешней среды как
неблагоприятные. К последним можно отнести также действие сильных доз облучения. Если неблагоприятные изменения будут сопутствовать многим поколениям,
то может произойти отбор на повышение сопротивляемости этим факторам, при условии, что такие организмы из поколения в поколение не будут погибать.
В процессе эволюции выработались приспособления, уменьшающие зависимость развивающегося организма от прямого воздействия факторов среды. Эмбрион
характеризуется определённой степенью автономности, которая увеличивается у более высокоорганизованных животных и достигает максимума у млекопитающих.
Эмбрион млекопитающих, развиваясь в утробе материнского организма и осуществляя опосредованную взаимосвязь с внешней средой через плаценту,
максимально защищен от прямого действия факторов среды. Его развитие характеризуется максимальной автономизацией.
Часто характер изменений развивающегося организма, вызываемых либо наследственными, либо средовыми факторами, бывает сходным. Например, у женщин,
перенесших краснуху на ранних сроках беременности, часто рождаются глухонемые дети или дети с врождённой катарактой, причём эти аномалии не отличимы от
соответствующих аномалий, обусловленных генетически. Изменения фенотипа, сходные с изменениями генетической природы, но обусловленные только
факторами внешней среды, получили название фенокопий.
Роль наследственности заключается в передаче генетической информации от родителей к потомству. Гены, содержащиеся в ДНК, кодируют информацию о
различных аспектах структуры и функции организма. Эти гены могут определять такие характеристики, как форма и цвет глаз, цвет волос, рост и склонность к
определенным заболеваниям. Влияние наследственности проявляется в том, что некоторые особенности могут быть унаследованы от родителей.
Среда влияет на онтогенез путем создания определенных условий для развития организма. Эта среда может включать физические, химические и биологические
факторы, такие как питание, температура, воздух, освещение, образование и социальное взаимодействие.
95.
85. Механизмы онтогенеза на клеточном и организменном уровнях: размножение, рост, дифференцировка,морфогенез.
Рост, увеличение массы индивидуума (особи), происходящее за счёт увеличения числа клеток, массы клеток и неклеточных образований. Рост живой системы происходит в результате преобладания анаболизма над катаболизмом.
У животных в процессе индивидуального развития, или онтогенеза, Рост тесно связан с качественными изменениями - дифференцировкой. Рост и дифференцировка часто осуществляются асинхронно, но не исключают друг друга. Показателями Рост обычно служат изменение массы (весовой Рост) или
линейных размеров (линейный Рост) особи или органа. Для описания Рост используют: кривые Рост (характеризующие изменение массы или длины тела в течение онтогенеза); величину абсолютного и относительного приростов за определённый отрезок времени; удельную скорость
Нарушение любого этапа может привести к возникновению пороков развития и уродства
Дифференцировка – это процесс формирования структурно-функциональной организации клеток многоклеточных животных и растений, в результате которого клетки приобретают способность к выполнению отдельных функций в сложном организме. Дифференцированное состояние в норме
стабильно: клетки нервной ткани не превращаются в печеночные или эпителиальные клетки кишечника, и наоборот.
Конечная дифференцировка часто связана с утратой способности клеток к размножению. Активная пролиферация и функционирование – процессы в норме, как правило, исключают друг друга. Это, вероятно, и служит причиной того, что, например, нервные клетки млекопитающих делятся только в
эмбриональном и раннем постэмбриональном периодах. При развитии взрослого организма формируются комплексы, состоящие из многих нервных клеток, выполняющих разные функции, начиная от чувствительного нейрона и кончая двигательным. Если бы нервные клетки постоянно делились во
взрослом организме, то поддержание целостности этих комплексов было бы невозможно, что, несомненно, имело бы катастрофические последствия для нервной регуляции.
Итак, дифференцированное состояние проявляется в специфическом “портрете” соматических клеток и их функциональной характеристике. Детерминация клеток начинается в раннем эмбриогенезе и постепенно сужает число возможных превращений клеток до одного какого-либо
дифференцированного состояния или очень немногих.
Детерминированные клетки постепенно вступают на путь развития (неспециализированные эмбриональные клетки превращаются в дифференцированные клетки организма). Дифференцированные клетки в отличие от детерминированных обладают специальными морфологическими и
функциональными организациями. В них происходят строго определенные биохимические реакции и синтез специальных белков.
Как правило, дифференциация происходит в эмбриональном периоде и приводит к необратимым изменениям полипотентных клеток эмбриона.
Синтез специальных белков начинается на очень ранних стадиях развития. Касательно стадии дробления: бластомеры отличаются друг от друга цитоплазмой. В цитоплазме различных бластомеров имеются разные вещества. Ядра всех бластомеров несут одну и ту же генетическую информацию, т.к.
имеют одинаковое количество ДНК и идентичный порядок расположения пар нуклеотидов. Вопрос о специализации до сих пор не нашел ответа.
1939 год Томас Морган выдвинул гипотезу: « дифференцировка клеток связана с активностью разных генов одного и того же генома». В настоящее время известно, что в дифференцированных клетках работает около 10% генов, а остальные неактивны. В силу этого в разных типах специализированных
клеток функционируют свои определенные гены. Специальными опытами по пересадки ядер из клеток кишечника головастика в безъядерную яйцеклетку было доказано, что в дифференцированных клетках сохраняется генетический материал и прекращение функционирования определенных генов
обратимо. Из яйца лягушки удаляли ядро, брали ядро из клетки кишечника головастика. Развитие не происходило, иногда эмбриогенез происходил нормально. Строение взрослой лягушки полностью определялось ядром.
На функционирование генов в процессе развития многоклеточного организма оказывают влияние сложные и непрерывные взаимодействия ядра и цитоплазмы и межклеточные взаимодействия.
Регуляция дифференцировки происходит на уровне транскрипции и на уроне трансляции
Морфогенез – возникновение и развитие органов, частей тела организмов как в онтогенезе, так и в филогенезе. Изучение особенностей морфологии на разных этапах онтогенеза в целях управления развитием организмов составляет основную задачу биологии развития, а также генетики, молекулярной
биологии, эволюционной физиологии и др. и связано с изучением закономерностей наследственности.
Морфогенез — это процесс возникновения новых структур и изменения их формы в ходе индивидуального развития организмов. Морфогенез, как рост и клеточная дифференцировка, относится к ациклическим процессам, т. е. не возвращающимся в прежнее состояние и по большей части
необратимым. Морфогенез на надклеточном уровне начинается с гаструляции. У хордовых животных после гаструляции происходит закладка осевых органов. В этот период, как и во время гаструляции, морфологические перестройки охватывают весь зародыш. Следую¬щие затем органогенезы
представляют собой местные процессы. Внутри каждого их них происходит расчленение на новые дискрет¬ные (отдельные) зачатки. Так последовательно во времени и в пространстве протекает индивидуальное развитие, приводящее к формированию особи со сложным строением и значительно
более богатой информацией, нежели генетическая информация зиготы. Таким образом, морфогенез представляет собой многоуровне¬вый динамический процесс. В настоящее время уже многое известно о тех структурных превращениях, которые происходят на внутри¬клеточном и межклеточном
уровнях и которые преобразуют хими¬ческую энергию клеток в механическую, т. е. об элементарных движущих силах морфогенеза.
периоды развития.
Критический период – период, который связан с изменением обмена веществ (переключение генома).
В онтогенезе человека выделяют:
1. развитие половых клеток 2. оплодотворение 3. мплантация (7-8 неделя) 4. развитие осевых органов и формирование плаценты(3-8 недели) 5. стадия роста головного мозга (15-20 недели). 6. формирование основных функциональных систем организма и дифференцировка полового аппарата(10-14
недели). 7. рождение(0-10 дней) 8. период грудного возраста – максимальная интенсивность роста, функционирование системы энергопродукции и др. 9. дошкольный (6-9 лет) 10. пубертатный - для девочек 12. для мальчиков 13 лет. 11. окончание репродуктивного периода, у женщин – 55, у мужчин –
60 лет.
В критические периоды развития проявляются мутации, поэтому надо быть внимательным к этим периодам. Все генетические программы связаны с детскими учреждениями. Наследственные пороки (уродства) вызваны изменениями у родителей в ходе гаметогенеза в генотипе. Наследственные
уродства – выражены из-за повреждающих факторов среды. Описано около 50 форм наследственной глухоты. Около 250 аномалий глаза, около 100 аномалий скелета. В развитии организма большое значение имеют иммунная, эндокринная и нервная системы. Иммунная система способствует
сохранению и зарождению жизни, контролирует генотипическое постоянство, выполняет контрольные функции. На ранних стадиях эмбриогенеза формируется из стволовых клеток. До 2 месячного возраста развивается тимус, угасает к периоду полового созревания. Иммунная система очищает
организм от мутирующих генотипов.
96.
86. Постнатальный онтогенез.Постнаталъный период
Период новорожденности — от рождения до 28 дней: адаптация к внешним условиям жизни, самые быстрые для всего периода постнатального роста скорости роста и созревания.
Младенчество — 2-й месяц жизни до конца грудного вскармливания (в среднем до 3 лет): постепенное уменьшение скоростей роста, вскармливание материнским молоком, прорезывание молочных зубов, освоение многих ключевых
стадий развития в психологическом и поведенческом контекстах.
Детство — с 3 до 7 лет: умеренные скорости роста, полуростовой скачок, прорезывание первых постоянных моляров и резцов, окончание роста мозга в конце этой стадии; хотя и отнятые от груди, дети нуждаются в защите и опеке
взрослых.
Ювенильная стадия — девочки — 7-10; мальчики — 712; замедление скоростей роста, способность к самостоятельному пропитанию, переход к овладению социальными навыками.
Пубертат — начинается сразу же после окончания ювенальной стадии и длится всего несколько дней или недель. Реактивация механизмов центральной нервной системы, запускающая процесс полового созревания, резкое увеличение
секреции половых гормонов.
Подростковый период — от 5 до 8 лет после начала пубертата: подростковый скачок роста длины и веса тела, фактическое завершение прорезывания постоянных зубов, развитие вторичных половых признаков, социосексуаль-ная зрелость,
рост интереса и переход к социальной, экономической и сексуальной жизни взрослых.
Зрелость и переходный период — от 20 лет до окончания периода деторождения: гомеостаз физиологических, поведенческих, интеллектуальных функций, менопауза у женщин в возрасте около 50 лет.
Пожилой возраст и старость — с окончания периода деторождения до смерти: затухание функций многих тканей, органов и систем органов
Постнатальный онтогенез у человека подразделяют на следующие периоды:
1) новорождения – 1-й-10-й дни; сложный период адаптации к совершенно новым условиям существования;
2) грудной – 11-й день – 1-й год; ребёнок вскармливается молоком матери, в котором содержатся помимо питательных веществ, солей и витаминов готовые антитела;
3) раннего детства — 2-й – 3-й годы; ребёнок учится нормально ходить, говорить, начинает познавать окружающий мир;
4) первый период детства — 4-й – 6-й годы; ребёнка интересует всё
окружающее и он стремится его понять;
5) второй период детства — девочки 7-й – 11-й годы, мальчики 7-й –12-й годы; школьный период до полового созревания;
6) подростковый — девочки 12-й – 15-й годы, мальчики 13-й – 16-й годы; период наступления полового созревания;
7) юношеский — девушки 16-й – 20-й годы, юноши 17-й – 21-й годы; период окончания роста, полового и физического созревания
8) средний возраст (I период) — мужчины 22-й – 35-й годы; женщины21-й – 35-й годы — наилучший период для деторождения;
9) средний возраст (II период) — женщины 36-й – 55-й годы, мужчины 36-й – 60-й годы; это период максимального профессионализма; после 35 лет обнаруживаются изменения некоторых физиологических и биохимических реакций
обмена, которые предшествуют инволюции; к концу этого периода происходят изменения, определяющие начало процессов старения и включаются механизмы, обеспечивающие перестройку организма и его адаптацию;
10) пожилой возраст — женщины 56-й – 75-й годы, мужчины 61-й -75-й
годы; в этот период многие люди ещё сохраняют достаточную профессиональную трудоспособность, хотя процессы старения продолжают развиваться;
11) старческий возраст – 76-й – 90-й годы; заметно выражены старческие изменения, однако и в этом возрасте многие люди сохраняют ясность ума и способность к творческому труду;
12) долгожители — свыше 90 лет; до этого последнего периода онтогенеза доживают преимущественно женщины.
97.
87. Биологическое старение на различных уровнях организации организма. Проблемы долголетия.Биологическое старение является нормальным процессом, который происходит на различных уровнях организации организма. Вот некоторые из них:
1. Клеточный уровень: С возрастом клетки в организме подвергаются повреждениям и накоплению мутаций в ДНК. Также уменьшается способность клеток воспроизводиться и восстанавливаться.
2. Тканевый уровень: С возрастом ткани теряют свою структуру и функцию. Например, мышцы и кости становятся слабее и более хрупкими, кожа теряет свою эластичность
3. Органный уровень: Биологическое старение оказывает влияние на работу и функцию различных органов, таких как сердце, легкие, печень и почки. Они начинают функционировать менее эффективно.
4. Уровень системы: Возрастные изменения в разных системах организма, таких как иммунная система или эндокринная система, особенно влияют на общее состояние организма.
Проблемы долголетия связаны с возрастным процессом и включают различные заболевания и ограничения, которые могут снижать качество жизни стареющего человека. Некоторые из них включают сердечно-сосудистые заболевания, диабет, онкологические заболевания, нейродегенеративные
заболевания и проблемы с опорно-двигательным аппаратом.
Старение человека как и старение других организмов, это биологический процесс постепенной деградации частей и систем тела человека и последствия этого процесса. Тогда как физиология процесса старения аналогична физиологии старения других млекопитающих, некоторые аспекты этого процесса,
например, потеря умственных способностей, имеют большее значение для человека. Кроме того, большое значение приобретают психологические, социальные и экономические эффекты.
Старение представляет собой всеобъемлющий процесс, охваты¬вающий все уровни структурной организации особи —от макромолекулярного до организменного.
В дифференцированных клетках млекопитающих животных старение сопровождается в целом снижением транскрипционной активности.
Изменение трансляции в процессе старения изучают по содержанию рРНК (показатель общей белокобразующей способности клетки), мРНК (набор образуемых белков), активности аминоацил-тРНК-синтетаз (ферменты активации аминокислот). Оказалось, что в возрасте от 12 до 70 лет у людей утрачивается
до половины генов рРНК, относящихся, как известно, к умеренно повторяющимся нуклеотидным последовательностям, которые продублированы в геноме более 300 раз. Сохраняющееся число генов, по-видимому, способно обеспечить образование требуемого количества рРНК. Интенсивность белкового
синтеза в целом снижается в зрелом возрасте.
Субклеточный уровень
Митохондрии
Рейнер изучал влияние возраста на углеводный обмен гомогенатов тканей крысы и обнаружил снижение потребления кислорода мозгом лишь в возрасте старше 2 лет. Он наблюдал также постепенное снижение интенсивности анаэробного гликолиза. Рейнер ставил свои опыты с избытком субстратов.
Рафский и др. изучали возрастные изменения дыхания гомогенатов на тканях морской свинки. Эти авторы измеряли потребление кислорода в присутствии глюкозы и раствора Рингера. В опытах с гомогенатами печени, взятой у животных различного возраста (от 8 до 100 недель), существенных возрастных
изменений не наблюдалось; на гомогенатах почки удалось выявить довольно существенные возрастные различия.
Ядро
Фалыконе и др. недавно провели работу с целью изучить влияние возраста на плоидность ядер крысиной печени путем определения их объема и среднего содержания ДНК. Существенных возрастных изменений не обнаружено, хотя Эндрью отмечал отдельные аномальные кариотипы. Мы уже указывали
на более высокую частоту амитоза. В тканях, подвергающихся атрофии, пикноз и кариолиз нередко сопровождают процесс отмирания клеток. Однако, по мнению автора, многие изменения формы ядра являются побочным результатом нарушений на тканевом уровне, а не отражением процесса старения,
свойственного самим клеткам.
Микросомы, эндоплазматический ретикулум и аппарат Гольджи
О возрастных изменениях микросом и эндоплазматического ретикулума свидетельствуют данные об уменьшении с возрастом содержания вещества Ниссля в нервной ткани животных и человека. По данным Пеляи и Палада тельца Ниссля представляют собой не что иное, как содержащие РНК гранулы
эндоплазматического ретикулума. В той же работе авторы указывают, что им не удалось идентифицировать аппарат Гольджи, но они склонны принять за него лежащую вблизи ядра часть гладкого эндоплазматического ретикулума, компоненты которого отличаются более плотной «упаковкой». Это
предположение согласуется с полученными при помощи фазово-контрастной микроскопии данными Гетенби о расположении этой структуры в симпатических ганглиях мыши. Выше мы привели данные Эндрью о наблюдавшихся им изменениях аппарата Гольджи в нервных клетках старых мышей.
Характерная для этой структуры непрерывная сеть превращается в отдельные разбросанные в беспорядке гранулы. Есть основания ожидать, что исследование возрастных изменений аппарата Гольджи при помощи электронного микроскопа значительно расширит наши представления.
Ядерная и клеточная мембраны
Данные о возрастных изменениях ядерной и клеточной мембран весьма скудны, если не считать работ Лансинга о накоплении кальция в кортикальном слое клеток. Вполне вероятно, что ядерные и клеточные мембраны непрерывно обновляются. Об этом свидетельствуют результаты киносъемки клеток в
культуре, которые дают представление о подвижности мембран в процессе фагоцитоза и пиноцитоза. Мы не располагаем достоверными данными о скорости диффузии через мембраны клеток, если не считать данных Кирка и Ларсена о проницаемости клеток мозжечка и аорты у людей различного
возраста; этим авторам не удалось выявить существенные возрастные различия. Необходимо провести более точные исследования скорости диффузии в различных тканях интактных животных различного возраста.
На клеточном уровне старение проявляется в замедлении деления клеток. Частично этот эффект является результатом так называемого предела Хейфлика деления соматических клеток. Эта граница связана с отсутствием активной теломеразы, в результате чего конечные участки хромосом, теломеры,
сокращаются при каждом делении. У человека соматическая клетка может делиться около 52 раз, после чего теломеры исчезают, и в клетках запускается программа апоптоза, «альтруистического самоубийства» клеток. Хотя граница Хейфлика и считается основной причиной уменьшения уровня клеточного
деления, даже стволовые клетки, в которых эта граница отсутствует, становятся менее активными, замедляют свое деление и не так часто дифференцируются в соматические клетки. Серьёзной причиной, тормозящей прогресс в области изучения старения клеток, является то, что исследования в этой сфере
в основном выполняются на большой совокупности клеток в один из моментов роста клеточной популяции. Однако такой подход не позволяет проследить всю последовательность изменений, начиная с их возникновения в отдельных клетках и заканчивая экспансией этих изменений в клеточной
популяции.
Геронтология — это наука, изучающая биологические механизмы и процессы, обуславливающие и сопровождающие старение живых существ, а также способы замедления старения и увеличения продолжительности жизни.
Гериатрия — медицинская дисциплина, занимающаяся изучением особенностей заболеваний у лиц пожилого и старческого возраста и их лечением.
98.
88. Регенерация органов и тканей, физиологическая и репаративная регенерация.Регенерация — способность живых организмов со временем восстанавливать повреждённые ткани, а иногда и целые потерянные органы.
Регенерацией также называется восстановление целого организма из его искусственно отделённого фрагмента (например, восстановление
гидры из небольшого фрагмента тела или диссоциированных клеток). У протистов регенерация может проявляться в восстановлении
утраченных органоидов или частей клетки.
Регенерацией называется восстановление организмом утраченных частей на той или иной стадии жизненного цикла. Регенерация,
происходящая в случае повреждения или утраты какого-нибудь органа или части организма, называется репаративной. Регенерацию в
процессе нормальной жизнедеятельности организма, обычно не связанную с повреждениями или утратой, называют физиологической.
Физиологическая регенерация
В каждом организме на протяжении всей его жизни постоянно идут процессы восстановления и обновления. У человека, например, постоянно
обновляется наружный слой кожи. Птицы периодически сбрасывают перья и отращивают новые, а млекопитающие сменяют шерстный покров.
У листопадных деревьев листья ежегодно опадают и заменяются свежими. Такие процессы носят название физиологической регенерации.
Репаративная регенерация
Репаративной называют регенерацию, происходящую после повреждения или утраты какой-либо части тела. Выделяют типичную и атипичную
репаративную регенерацию.
При типичной регенерации утраченная часть замещается путём развития точно такой же части. Причиной утраты может быть внешнее
воздействие (например, ампутация), или же животное намеренно отрывает часть своего тела (автотомия), как ящерица, обламывающая часть
своего хвоста, спасаясь от врага.
При атипичной регенерации утраченная часть замещается структурой, отличающейся от первоначальной количественно или качественно. У
регенерировавшей конечности головастика число пальцев может оказаться меньше исходного, а у креветки вместо ампутированного глаза
может вырасти антенна.
Регенерация органов и тканей - это процесс восстановления или замены поврежденных или утраченных клеток, тканей или органов организма.
Есть два типа регенерации: физиологическая и репаративная.
Физиологическая регенерация происходит в некоторых органах организма, которые постоянно обновляют свои клетки. Это происходит путем
деления специализированных клеток, таких как эпителиальные клетки кожи или кишечника.
Репаративная регенерация происходит в органах или тканях, которые не могут нормально восстановить свои клетки. В случае повреждения или
потери клеток происходит образование рубца, который замещает потерянные клетки и восстанавливает структуру, но не полностью
восстанавливает функцию органа или ткани. Примером репаративной регенерации является заживление раны, которое может привести к
образованию шрама.
99.
89. Филогенез систем органов хордовых.ФИЛОГЕНЕЗ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Для хордовых характерна филогенетическая, эмбриональная и функциональная связь пищеварительной и дыхательной систем. Дыхательная система развивается на базе пищеварительной, у низших хордовых функционирует совместно с
ней, у взрослых наземных позвоночных системы перекрещиваются в области глотки. Обе системы закладываются у эмбрионов под хордой в виде прямой трубки, которая в дальнейшем делится на три части, передняя из которых называется
stomodeum.
У рыб и земноводных крышей ротовой полости служит основание мозгового черепа. У рептилий и млекопитающих объем ротовой полости увеличивается за счет складок верхнечелюстных и небных костей. У млекопитающих обе складки
срастаются, образуя вторичное твердое небо, которое разделяет ротовую и носовую полости. Если это срастание нарушается у человека, формируется порок развития – «волчья пасть». Этот порок имеет генетические механизмы
возникновения и может наследоваться.
Зубы по происхождению связаны с плакоидной чешуей рыб, у которых они располагаются в несколько рядов. У рептилий уже имеется один ряд зубов, но они все одинаковые - гомодонтная зубная система. Такие зубы могут многократно
сменяться в течение жизни. У млекопитающих зубы дифференцированы и выполняют различные функции - гетеродонтная зубная система. У человека имеются резцы, клыки, премоляры и моляры. Зубы расположены в ячейках альвеолярных
дуг, общее их число прогрессивно уменьшилось до 32, имеется лишь однократная смена зубов. Гомодонтные зубы являются редкой аномалией у человека, но часто происходит закладка сверхкомплектных зубов, которые обычно
уменьшены в размерах. На дне рта имеется язык, у рыб он лишен мышц. У наземных позвоночных язык подвижен и закладывается из трех зачатков. Если парные зачатки не срастаются, возникает редкая аномалия у человека – раздвоенный
язык.
Глотка выполняет дыхательную и пищеварительную функции. У рыб 5-7 жаберных щелей. Навстречу выростам глотки жаберным мешком выпячиваются кожные жаберные карманы. Между ними расположены щели. Они закладываются и у
наземных хордовых, но щели прорываются только у личинок амфибий, у пресмыкающихся и млекопитающих щели прорываются только при нарушении развития. У человека при этом формируются аномалии - латеральные свищи и кисты
шеи, которые часто малигнизируются. Первая жаберная щель у наземных хордовых превращается в евстахиеву трубу, барабанную полость и наружный слуховой проход.
Кишечная трубка в филогенезе удлиняется, дифференцируется на отделы, увеличивается количество одноклеточных желез, образуются крупные многоклеточные железы. У ланцетника длина кишечной трубки составляет 1/3 длины тела, у
человека она в 10 раз длиннее тела. У рептилий впервые появляется слепая кишка, в которой поселяются симбиотические бактерии и простейшие. Это позволяет значительно расширить рацион питания. У человека встречаются такие пороки,
как укорочение и недоразвитие любых отделов кишечника и пищеварительных желез. Тяжелые аплазии несовместимы с жизнью. Нередко встречаются свищи между трахеей и пищеводом, а при персистировании клоаки прямая кишка и
мочеполовые пути объединены. При замедлении миграции зачатков поджелудочной железы из пищеварительной трубки наблюдается гетеротопия поджелудочной железы в тонкий кишечник и желудок.
ФИЛОГЕНЕЗ ОРГАНОВ ДЫХАНИЯ
Жабры – наиболее ранние органы дыхания. Среди наземных позвоночных они функционируют лишь у личинок земноводных.Эволюция шла по пути увеличения дыхательной поверхности жабр. У ланцетников имеются только жаберные
щели, у круглоротых формируются жаберные мешки, у рыб на стенках щелей появляются пронизанные капиллярами жаберные лепестки. Позади жаберных дуг у кистеперых рыб формируется парный плавательный пузырь. Он выполняет
гидростатическую функцию, но в нем также происходит газообмен между кровью и воздухом, который может беспрепятственно проходить в пузырь из глотки. Кистеперые рыбы вышли на сушу, от них произошли первые амфибии –
стегоцефалы, а из плавательного пузыря – первые крупноячеистые легкие, поэтому газообмен у амфибий в немалой степени осуществляется также и через кожу. В дальнейшем из материала жаберных дуг формируются дыхательные пути –
гортань, трахея, бронхи, которые постепенно удлиняются, в них происходит очистка, согревание и увлажнение воздуха. Легкие у рептилий становятся мелкоячеистыми, появляются межреберные мышцы. У человека альвеолярная
поверхность легких достигает 90 м2, а диафрагма приобретает мышечные волокна и становится основной дыхательной мышцей.
Т.о., эволюция вновь шла по пути увеличения дыхательной поверхности: крупноячеистые дыхательные мешки у земноводных , легкие с большим количеством внутренних перегородок у рептилий, губчатые легкие у птиц,
мелкоальвеолярные легкие у млекопитающих с преддвериями, альвеолярными ходами, альвеолами. Также удлинились и дифференцировались дыхательные пути, совершенствовалась дыхательная мускулатура. Среди аномалий
встречается эзофаготрахеальные и бронхолегочные свищи, а также кистозная гипоплазия легких, при которой бронх связан с кистой, имеющей очень малую дыхательную поверхность. Могут быть недоразвитие или полная аплазия
диафрагмы, которая несовместима с жизнью.
ФИЛОГЕНЕЗ СИСТЕМЫ КРОВООБРАЩЕНИЯ
Для крупных высокоорганизованных организмов необходимо эффективное и быстрое перемещение жидкой внутренней среды. Это обеспечивается системой кровообращения. У хордовых она замкнутая. У ланцетника основные сосуды –
брюшная и спинная аорты. По брюшной венозная кровь движется к органам дыхания, по спинной – артериальная кровь к органам. Часть брюшной аорты периодически сокращается, проталкивая кровь по сосудам. В брюшную аорту с левой и
с правой стороны впадают парные кювьеровы протоки. Венозная кровь от желудочно-кишечного тракта оттекает по воротной вене к печени, из которой выходит печеночная вена, впадающая в брюшную аорту. В дальнейшем правый
кювьеров проток преобразуется в верхнюю полую вену, а левый – в коронарный синус сердца.
У рыб рядом с жаберным аппаратом формируется двухкамерное сердце, но сохраняется один круг кровообращения, как у ланцетника. Сердце амфибий расположено каудальнее, рядом с легкими, оно трехкамерное – два предсердия и
желудочек, из которого выходит артериальный конус. Он делится на три пары сосудов: кожно-легочные артерии, дуги аорты, сонные артерии, они несут соответственно венозную, смешанную и почти артериальную кровь. Такое разделение
объясняется особым строением желудочка и спирального клапана в конусе. У рептилий в сердце возникает неполная межжелудочковая перегородка, а от сердца отходит не один, а три непарных сосуда: из левой части желудочка – правая
дуга аорты с артериальной кровью, из правой – легочная артерия с венозной кровью, из середины – левая дуга аорты со смешанной кровью. У млекопитающих происходит полное разделение венозного и артериального кровотоков, сердце
становится четырехкамерным, правая дуга аорты редуцируется, остается левая, несущая артериальную кровь.
Сердце у человека закладывается в области шеи, затем перемещается с переднее средостение. Редкие пороки, несовместимые с жизнью - шейное положение сердца и двухкамерное сердце. Гораздо чаще встречаются дефекты
межпредсердной и межжелудочковой перегородок, вплоть до трехкамерного сердца. У человека, как у большинства амниот, закладывается шесть пар жаберных артериальных дуг, они никогда не функционируют одновременно.
Сохраняются левые части IV и IV дуги – аорта и легочная артерия. Очень редко сохраняется правая половина IV дуги, это клинически не проявляется. Если сохраняются обе части IV дуги, они срастаются позади пищевода, сдавливая трахею, что
проявляется нарушением глотания и удушьем. Самым частым пороком является сохранение боталлова протока между спинной аортой и легочной артерией. Через проток артериальная кровь сбрасывается в венозный малый круг. Такой
проток имеется у взрослых хвостатых амфибий, у более развитых позвоночных он облитерируется. Редким пороком является отхождение от сердца единственного артериального ствола (первичного эмбрионального), порок летален.
Существует множество других аномалий артериальных и венозных сосудов.
100.
89. Филогенез систем органов хордовых.ФИЛОГЕНЕЗ МОЧЕПОЛОВОЙ СИСТЕМЫ
Выделительная и половая системы выполняют функции выделения продуктов обмена и размножения соответственно, и имеют единую закладку – нефротом, который формируется в области ножки
сомита и тесно связан с вторичной полостью тела – целомом.Органом выделения являются почки, которые в филогенезе проходят три этапа: пронефрос (головная, предпочка), мезонефрос
(туловищная), метанефрос (тазовая). Предпочка существует только у личинок рыб и амфибий, туловищная почка – у взрослых рыб и амфибий, у рептилий и млекопитающих – тазовая почка.
При развитии предпочки от головного конца тела к клоаке тянется пронефритический канал, который впоследствии расщепляется на два – вольфов, который вступает с соединение с нефронами, и
мюллеров, который образует яйцевод, а передним концом открывается в целом. У рептилий и млекопитающих из каудальной части вольфова канала образуются почка и мочеточник, а у самцов
также семенные протоки. Мюллеров канал у самцов редуцируется, а у самок из него формируется матка с придатками и влагалище. У яйцекладущих и сумчатых имеется по два влагалища, матки и
яйцевода, у остальных эти образования срастаются в одно (кроме яйцеводов).
Почки состоят из нефронов, которые фильтруют жидкость внутренней среды, поэтому связаны с кровеносной системой. Пронефрос и мезонефрос, который имеет нефроны с капсулой и сосудистым
клубочком, сохраняют связь с целомом, а метанефрос – полностью эту связь утрачивает. Для экономии жидкости выделительные канальцы в процессе эволюции удлиняются, формируется также
петля Генле, что обеспечивает эффективное обратное всасывание не только воды, но и солей, глюкозы, гормонов. Метанефрос рептилий сохраняет сегментарное строение, у млекопитающих оно
утрачено, почки перемещаются вниз – в забрюшинное пространство поясничной области. У анамний первоначально закладываются только две почки – первичная и туловищная, последняя остается
функционировать, у амниот – три почки, при этом у взрослых особей функционируют тазовые.
Нередкими аномалиями являются сегментирование или удвоение почки и мочевыводящих путей, а также опущение почки – нефроптоз или ее тазовое положение.
Половые органы закладываются в виде парных складок. Первоначально яичники имеют фолликулярное строение, семенники – трубчатое. Иногда развивается гермафродитизм, он нередко
встречается у круглоротых и рыб, у человека – крайне редко (ovotestis). У большинства млекопитающих мужские гонады из брюшной полости через паховый канал перемещаются в мошонку. При
нарушении такого перемещения возникает крипторхизм – неопущение яичек.
У человека нередко встречается аномалия - двурогая матка, а также удвоенная матка. Закладка полового члена также парная, в дальнейшем зачатки срастаются. Редкая аномалия - полное удвоение
полового члена.
ФИЛОГЕНЕЗ НЕРВНОЙ СИСТЕМЫ
У большинства многоклеточных животных существует две интегрирующие системы – нервная и эндокринная. Последняя филогенетически более древняя, она осуществляет медленные
приспособительные реакции. У позвоночных большое значение приобрела нервная системы, которая обеспечивает быстрое реагирование внутри организма и при взаимодействии с внешней
средой.
Нервная система хордовых формируется из эктодермы, закладывается сначала в виде пластинки. Затем преобразуется в трубку над хордой с полостью внутри – невроцелем. передний конец трубки
расширен. Здесь формируется головной мозг, который у взрослых позвоночных состоит из 5 отделов – переднего, промежуточного, среднего, заднего и продолговатого. У рыб и амфибий самым
большим и интегрирующим отделом является средний мозг, связанный с центрами зрения - ихтиопсидный тип мозга. У рептилий увеличивается передний отдел, в котором особенно развиты
полосатые тела, которые являются высшим интегрирующим центром. На поверхности крыши формируются зачатки древней коры - arhicortex, или arhipallium. Это зауропсидный тип мозга. У
млекопитающих появляется новая кора – neocortex, или neopallium, со сложным строением, у высших приматов она формирует множество извилин. Это маммалийный тип мозга. Между
полушариями имеется комиссура. Промежуточный мозг включает гипоталамус, гипофиз, эпифиз и управляет вегетативными функциями. В среднем мозге располагаются подкорковые центры
зрения и слуха. Хорошо развит мозжечок, который имеет свою кору, и осуществляет распределение тонуса мышц и координацию движений.
В процессе развития из переднего утолщения формируется вначале три мозговых пузыря – передний, средний и задний. Передний подразделяется на два – telencephalon (1) – передний и
diencephalon (2) – промежуточный. Из среднего формируется средний мозг - mesencephalon (3). Из заднего – metencephalon (4) – задний мозг и medulla oblongata (5) – продолговатый. Это стадия
пяти мозговых пузырей. У человека уже на ранних стадиях опережающими темпами начинает развиваться передний мозг.
Нервная система столь важна, что многие пороки ее развития несовместимы с жизнью. Среди них рахисхиз – незамыкание нервной трубки и прозенцефалия – недоразвитие полушарий и коры. При
агирии (отсутствие извилин), а также олигогирии и пахигирии (уменьшение числа и утолщение извилин) развивается грубая олигофрения с нарушением многих рефлексов. Такие дети обычно
умирают в течение первого года жизни.
101.
90. Трансплантация эмбрионов. Аллофенные животные.Трансплантация эмбрионов в настоящее время является одной из наиболее актуальных проблем в области животноводства. С помощью пересадки эмбрионов можно
резко увеличить выход числа потомков от высокопродуктивных коров. Трансплантация эмбрионов, или эмбриотехнология, заключается в получении одного или
нескольких эмбрионов из матки племенных животных (доноров) и пересадке в матку коров (рецепиентов), где эмбрионы развиваются до отела. Этот метод в сочетании
с суперовуляцией у доноров позволяет получить большое потомство от высокопродуктивных животных. Этим способом эмбрионы можно внедрить в ту или иную
породу в другие регионы, используя в качестве рецепиентов коров мясных пород. Применение этого метода также упрощает обмен генофондом сельскохозяйственных
животных между странами и континентами. Пересадка эмбрионов может быть использована для получения потомства от ценных, но бесплодных коров, утративших
способность к размножению в результате несчастного случая, болезни или по возрасту.
Когда было установлено, что кролик обладает иммунитетом по отношению к ящуру, была выдвинута идея использования метода трансплантации для оздоровления
потомства зараженных ящуром животных. Половые пути кролика, куда трансплантируются эмбрионы, способны разрушать вирус ящура в эмбрионах. Трансплантация
может быть использована и для временного хранения эмбрионов. В яйцеводах крольчих удается осуществлять трансконтинентальную перевозку эмбрионов овец.
Извлечение эмбрионов до 70-х годов производили в основном хирургическим путем, впоследствии он был заменен менее травматичным и трудоемким
нехирургическим, основанным на введении в матку особого зонда по естественному каналу. Зонд имеет три канала. Один из каналов предназначен для надувания
баллончика, который закупоривает рог матки, препятствуя вытеканию жидкости. По другому каналу вводится физиологический раствор с температурой 25-30оС,
который вымывает эмбрионы и возвращается вместе с ними через третий канал зонда в пробирку, помещенную в водяную баню с температурой 35оС. Из этой
жидкости извлекаются эмбрионы. В среднем при суперовуляции от донора можно получить от 5 до 7 эмбрионов.
Трансплантацию производят с помощью специального зонда или пистолета для осеменения. Эмбрионы помещаются в рога матки. Стельность у самок - рецепиентов
проверяется по уровню прогестерона в плазме крови на 21-й день.
Регулирование пола. В практике разведения животных очень важно научиться управлять образованием в потомстве мужских и женских особей. Метод разделения
эмбрионов по полу основан на определении белков, специфичных для самцов. Этом метод широко применяется в животноводческой практике многих стран. В Канаде
уже с 1975 года рождаются телята, разделенные по полу на стадии эмбрионов. В перспективе для целенаправленного получения особей мужского или женского пола
может быть применен метод микрохирургической замены Х и У хромосом. Такие манипуляции уже проводились на растительных клетках и яйцеклетках земноводных.
Аллофенными называют химерные организмы, содержащие разные ткани, произошедшие из клеток, полученных от разных родителей
Б.Минтц получил аллофенных мышей путем образования смешанной бластулы из клеток черных и белых мышей. В последующих опытах соединяли бластомеры
животных, различающихся по другим признакам -окраске радужной оболочки, длине ушей, хвоста и др.
Для получения аллофернных потомков у беременных мышей, имеющих четко выраженные альтернативные признаки, извлекали эмбрионы на стадии восьми
бластомеров и с помощью фермента проназы отделяли бластомеры. Комбинируя бластомеры от двух (и более) эмбрионов, создали в специальной питательной среде
единый комплексный эмбрион, который ввели в матку мыши, гормонально подготовленной к имплантации зародыша. Рождавшиеся мышата представляли собой
мозаиков, у них проявлялись признаки всех родительских форм.
Методика, разработанная на мышах, в последние годы используется для получения аллофенных овец.
102.
91. Трансплантация органов и тканей, тканевая несовместимость.Пересадка органов и тканей (синоним трансплантация органов и тканей). Пересадка органов и тканей в пределах одного организма называется
аутотрансплантацией, от одного организма другому в пределах одного вида — гомотрансплантацией, от организма одного вида организму другого вида —
гетеротрансплантацией.
Пересадка органов и тканей с последующим приживлением трансплантата возможна только при биологической совместимости — сходстве антигенов (см.),
входящих в состав тканевых белков донора и реципиента. При ее отсутствии тканевые антигены донора вызывают выработку в организме реципиента антител (см.).
Возникает особый защитный процесс — реакция отторжения, с последующей гибелью пересаженного органа. Биологическая совместимость может быть только при
аутотрансплантации. Ее нет при гомо- и гетеротрансплантации. Поэтому основной задачей при осуществлении пересадки органов и тканей является преодоление
барьера тканевой несовместимости. Если в эмбриональном периоде на организм воздействовать каким-нибудь антигеном, то после рождения этот организм уже не
вырабатывает антитела в ответ на повторное введение того же антигена. Возникает активная толерантность (переносимость) к чужеродному тканевому белку.
Уменьшить реакцию отторжения можно различными воздействиями, подавляющими функции систем, вырабатывающих иммунитет против чужеродного органа. С
этой целью применяют так называемые иммунодепрессантные вещества — имуран, кортизон, антилимфоцитарную сыворотку, а также общее рентгеновское
облучение. Однако при этом происходит угнетение защитных сил организма и функции кроветворной системы, что может привести к тяжелым осложнениям.
В настоящее время широко применяют аутотрансплантацию кожи для закрытия послеожоговых дефектов, успешно пересаживают кости, хрящи, сухожилия и т. д.
Гомотрансплантация находит применение при пересадке роговицы, хряща. Все большее распространение приобретает пересадка почек от одного человека другому.
Наибольшие шансы на приживление возникают в случаях, когда ткани донора и реципиента близки по своему антигенному составу. Самые идеальные условия
существуют у однояйцевых близнецов. Однако почки пересаживают не только от живых лиц, но и от трупов. Важное значение имеет подбор донора, осуществляемый
путем определения совместимости по эритроцитарным и лейкоцитарным антигенам крови. Существует целый ряд других проб, позволяющих установить степень
сходства между органами и тканями донора и реципиента.
Показания к пересадке почек возникают при резком нарушении их функции в связи с тяжелым заболеванием (хронический нефрит, поликистоз и т. д.). Произведено
уже много операций пересадки почек, причем некоторые больные живут после операции более трех лет и вполне трудоспособны.
В 1967 г. Барнард (С. N. Barnard) с сотрудниками произвел первую в мире успешную гомотрансплантацию сердца человеку. Дальнейшие успехи по пересадке органов
связаны с изысканием путей преодоления барьера тканевой несовместимости.
Тканевая несовместимость — комплекс иммунных реакций организма к трансплантируемым чужеродным клеткам, тканям или органам. Тканевая несовместимость
возникла в процессе эволюции как развитие защитной реакции организма от бактерий и других чужеродных биологических агентов. В основе тканевой
несовместимости лежат явления иммунитета (см.). Тканевая несовместимость наблюдается при различных видах пересадки органов и тканей (см.). В случае гетероили ксенотрансплантации (пересадки органа от другого вида животного) основной причиной несовместимости являются видовые различия организмов донора и
реципиента. При гомо- или аллотрансплантации (пересадке органов от животного того же вида или от человека человеку) тканевая несовместимость обусловлена
индивидуальными особенностями специфичности белков, полисахаридов и липидов клетки. Эти особенности обусловлены присутствием во всех тканях организма
различных антигено
Гистосовместимость, тканевая совместимость (histocompatibility) [греч. histos — ткань] — иммунологическое сходство тканей донора и реципиента, благодаря
которому после пересадки клеток, тканей или органов отсутствует реакция отторжения трансплантата; Г. обусловлена генетически детерминированной
адекватностью антигенных свойств клеток донора и реципиента. Г. необходима для пересадки тканей или трансплантации органов без их последующей гибели и
отторжения.
103.
92. Понятие о гомеостазе. Генетические, клеточные, и системные основыгомеостатических реакций многоклеточного организма.
ГОМЕОСТАЗ - свойство живого организма сохранять относительное динамичное постоянство внутренней среды. Гомеостаз выражается в
относительном постоянстве химического состава, осмотическом давлении, устойчивости основных физиологических функций. Гомеостаз
специфичен и обусловлен генотипом.
Установление гомеостаза клеточной среды обеспечивается мембранными системами, с которыми связаны биоэнергетические процессы и
регулирование транспорта веществ в клетку и из нее. В клетке непрерывно идут процессы изменения и восстановления органоидов. Это
происходит и в обычных условиях среды, но особенно интенсивно при действии различных повреждающих факторов (изменение температуры,
гипоксия, недостаток питательных веществ). В основе реакций, осуществляемых в клетке на ультраструктурном уровне, лежат генетические
механизмы гомеостаза. Важнейшее свойство живого — са¬мовоспроизведение — основано на про¬цессе редупликации ДНК. Репарация играет
важнейшую роль в восстановлении структуры генетического материала и сохранении нормальной жизнеспособности клетки. При повреждении
механизмов репарации происходит нарушение гомеостаза как на клеточном, так и на организменном уровнях. Важным механизмом
сохранения гомеостаза является диплоидное состояние соматических клеток у эукариот. Стабилизация сложной системы генотипа
обеспечивается и явлениями полимерии, а также другими видами взаимодействия генов. У прокариот, имеющих более примитивную
организацию генотипа, наблюдается меньшая автономность организмов от колебания внешней среды и более низкая стабильность самого
генетического аппарата.
Системные мех-мы гом-за.
Этот уровень обеспечивается взаимодействием важнейших регуляторных систем: нервной, эндокринной и иммунной.
В организме имеются различные ме¬ханизмы, обеспечивающие поддержание кислотно-щелочного равновесия. Большую роль в раз¬работке
вопросов регуляции функций сыграла выдвинутая П. К. Анохиным теория функциональных систем, явив¬шаяся предпосылкой к построению
фи¬зиологической кибернетики
Гомеостазис - это способность организма поддерживать стабильность своей внутренней среды, несмотря на изменения во внешней среде.
Гомеостатические реакции происходят на генетическом, клеточном и системном уровнях в организме.
Генетические основы гомеостазиса связаны с наследственностью и передачей генетической информации от одного поколения к другому.
Некоторые гены могут кодировать белки, которые играют важную роль в регуляции гомеостаза.
Клеточные основы гомеостаза заключаются во взаимодействии и регуляции различных клеток в организме. Клетки имеют механизмы обратной
связи, которые позволяют им реагировать на изменения внешней и внутренней среды и поддерживать стабильность внутренней среды.
104.
93. Иммунологические механизмы гомеостаза. Проблемы трансплантации.Иммунологические механизмы гомеостаза включают различные процессы, которые поддерживают устойчивость иммунной системы и ее способность реагировать
на патогены, не принося вред организму. Примеры таких механизмов включают иммунотолерантность, иммунорегуляцию и иммунокомпетентность.
Проблемы, связанные с трансплантацией, возникают из-за иммунологической реакции организма на противоестественное присутствие чужеродного трансплантата.
Когда ткань или орган пересаживаются из одного организма в другой, иммунная система получателя может реагировать на трансплантат как на инородное вещество
и начать атаковать его.
Основные проблемы, связанные с трансплантацией, включают:
1. Отторжение трансплантата
2. Инфекции
3. Силовые проблемы
4. Длительность применения препаратов
Каждая из этих проблем требует дополнительных исследований и технологических разработок, чтобы обеспечить более безопасные и эффективные методы
трансплантации.
В клинике известны заболевания, связанные с врожденными дефектами иммунной системы. Например, при недоразвитии тимуса отсутствуют Т-лимфоциты,
нарушен трансплантационный и противоопухолевый иммунитет. При недоразвитии В-системы тимоциты сохранены, но в крови отсутствуют гамма-глобулины,
организм не вырабатывает антител. В норме иммунная система не продуцирует антител против собственных тканевых белков. Существует ряд заболеваний, которые
рассматривают как следствие извращения иммунной реакции — так называемые аутоиммунные болезни. Например, множественный склероз — очень опасное
заболевание, при котором развиваются аутоиммунные реакции, направленные против тканей центральной нервной системы. 5. Иммуногенетика. Иммуногенетика
— это направление, изучающее закономерности наследования антигенной специфичности и генетическую обусловленность иммунных реакций. 6. Общие
закономерности гомеостаза. Способность сохранять гомеостаз — одно из важнейших свойств живой си¬стемы, находящейся в состоянии динамического
равновесия с условиями внешней среды. Способность к поддержанию гомеостаза неодинакова у раз¬личных видов. Контроль за генетическим постоян¬ством
осуществляется иммунной систе¬мой. Эта система состоит из анатомиче¬ски разобщенных органов, представля¬ющих функциональное единство. Свой¬ство
иммунной защиты достигло высше¬го развития у птиц и млекопитающих. В процессе индивидуального разви¬тия характер внутренней среды клеток, органов,
организма меняется. Каждый возрастной период характеризуется спе¬цифическими особенностями обмена ве¬ществ и энергии и механизмами гомео-стаза.
Иммунный гомеостаз - это часть общего гомеостаза. Поддерживается иммунный гомеостаз при помощи иммунной системы, главная задача которой не допустить в
организм любых «чужаков». К «чужакам» могут относиться любые возбудители инфекции, инородные тела, чужеродные белки и даже собственные измененные
клетки (например, клетки злокачественной опухоли). Иммунная система отслеживает появление таких элементов и беспощадно уничтожает, поддерживая тем
самым постоянство внутренней среды. Иногда иммунная система дает сбои и нападает на «своих» (например, на клетки какого-то органа), в результате возникают
аутоиммунные заболевания.
Успех трансплантации зависит от многих факторов и, в первую очередь, от уровня идентичности по молекулам (антигенам) MHC между донором трансплантата и
больным реципиентом. Подбор пар для пересадки труден, т.к. слишком высока антигенная индивидуальность среди людей. Даже максимально возможное сходство
по МНС между донором и реципиентом не исключает значительных различий по минорным антигенам гистосовместимости .
Вторым осложняющим моментом при трансплантации является возможное присутствие у пациента антител к антигенам донора трансплантата. Это обстоятельство
определяет необходимость предварительного тестирования реципиента на наличие у него антител к антигенам трансплантируемого органа.
105.
94. Иммунологическая несовместимость. Резус конфликт.Причина иммунологического бесплодия
Причина иммунологического бесплодия — образование антиспермальных антител, как у мужчин, так и у женщин. У мужчин аутоантитела к сперматозоидам образуются в семенной плазме, что приводит к их аутоагглютинации. У женщин
антитела синтезируются в слизистой оболочке цервикального канала (реже в эндометрии и трубах) и вызывают полную иммобилизацию сперматозоидов, их агглютинацию; антителообразование происходит в результате попадания в секрет
женского репродуктивного тракта спермоспецифических антигенов (поверхностные и внутриклеточные антигены сперматозоидов, ферменты сперматозоидов, а также антигены групп крови системы ABO, MNSS, Rh-Hr, антигены системы
гистосовместимости HLA, содержащиеся в сперме). Также возможно возникновение в организме женщины аутоиммунной или аллергической реакции на антигены соответственной фолликулярной жидкости и блестящей оболочки фолликула
Такую реакцию иммунной системы действительно можно сравнить с аллергией . Она возникает, когда, попадая в организм женщины, сперматозоид воспринимается ее иммунитетом как чужеродный агент. И организм начинает с ним активно
бороться, вырабатывая специальные вещества – антиспермальные антитела. В результате этой борьбы сперматозоиды становятся неподвижными и не могут оплодотворить яйцеклетку.
Часто супружеская пара, имеющая такую проблему, не может зачать, а если беременность наступает, то прерывается на раннем сроке. Обследование лучше начинать до того, как вы планируете стать мамой. И первое, с чего надо начать, это
тест на совместимость. Проводится он очень легко. Через 2–3 часа после полового акта после 3 дней воздержания у женщины берется специальный мазок, в котором определяют количество живых сперматозоидов. Если их более 7, то тест
считается положительным и, значит, "аллергии" нет. Если 2–6, то слабоположительным. А если живых сперматозоидов вообще нет, то тест отрицательный.
При беременности может возникать иммунологическая несовместимости крови матери и плода по резус-фактору, реже по системе АВ0 и еще реже по некоторым другим (система Келл, Даффи, Лютеран, Льюис, система Рр) факторам крови. В
результате такой несовместимости возникает гемолитическая болезнь плода и новорожденного.
Большинство людей плохо представляют себе, что такое резус-фактор, поскольку в обычной жизни его наличие или отсутствие не влечет за собой никаких болезненных последствий. Однако в случае беременности неподходящее сочетание
резусов родителей может привести к так называемому резус-конфликту.
Резус-фактор — это антиген (белок), который находится на поверхности красных кровяных телец — эритроцитов. Он может присутствовать (положительный резус), или отсутствовать (резус отрицательный). Согласно медицинской статистике,
около 85% людей являются резус-положительными, остальные 15% — резус-отрицательными.
Резус-конфликт возникает либо при переливании резус-несовместимой крови, либо при беременности женщины с отрицательным резусом, если кровь у плода — резус-положительная.
Симптомы
У плода могут быть выявлены увеличение печени, селезенки и сердца с помощью УЗИ, наблюдается анемия, в лёгких случаях — ретикулоцитоз, в более тяжёлых — эритробластоз, желтуха. В наиболее тяжёлых случаях развиваются водянка
плода и отёчный синдром новорождённых, что может привести к мертворождению или смерти новорождённого.
Что происходит?
Когда в кровь матери с отрицательным резусом попадают эритроциты плода, несущие на себе белки системы резус, они воспринимаются ее иммунной системой как чужеродные. Организм начинает вырабатывать антитела для уничтожения
эритроцитов ребенка. При этом в его крови появляется большое количество вещества, называемого билирубином, которое может повредить его мозг. Поскольку эритроциты плода непрерывно уничтожаются, его печень и селезенка стараются
ускорить выработку новых эритроцитов, увеличиваясь при этом в размерах. В конце концов, и они не справляются с восполнением убыли эритроцитов. Наступает сильное кислородное голодание, раскручивается новый виток серьезных
нарушений. В самых тяжелых случаях это может привести к гибели плода.
Кому угрожает резус-конфликт?
Поскольку наличие резус-фактора передается по наследству, угроза резус-конфликта существует только в том случае, если будущая мать резус-отрицательна (Rh-), а отец резус-положителен (Rh+). При таком раскладе в 75% случаев мать и
ребенок будут резус-несовместимыми.
Но если у женщины отрицательный резус-фактор, а у мужчины положительный — это еще не повод отказаться от создания семьи.
Первая беременность такой пары скорее всего пройдет нормально. Если ранее женщина не встречалась с резус-положительной кровью, то у нее нет антител, а следовательно, и риска резус-конфликта с плодом. При первой беременности
антител вырабатывается не так много (это ведь «первая встреча»). Если же количество проникших в кровь матери эритроцитов плода было значительным, в организме женщины остаются «клетки памяти», которые при последующих
беременностях организуют быструю выработку антител против резус-фактора.
При резус-несовместимой беременности многое зависит и от того, чем она закончилась. После выкидыша резус-сенсибилизация (выработка антител) возникает в 3-4% случаев, после медицинского аборта — в 5-6, после внематочной
беременности — приблизительно в 1% случаев, а после нормальных родов — в 10-15. Риск сенсибилизации увеличивается после кесарева сечения или, если были отслойки плаценты. То есть все зависит от того, сколько эритроцитов плода
проникнет в кровоток матери.
106.
95. Паразитизм как биологический феномен. Адаптации к паразитизму. Взаимодействие в системе паразит-хозяин.Это форма межвидовых взаимоотношений двух организмов, при которых организм использует другой в качестве среды обитания, либо в качестве источника пищи. Паразитические организмы поражают все органы и ткани человека. Они
обитают на наружных покровах (блохи, вши), в тканях (гельминты), в крови (малярийный плазмодий).
Организм хозяина для паразита является источником питания, местом обитания, защитой от врагов. Тело хозяина создает для паразитов благоприятный микроклимат, не подверженный колебаниям, которые имеются в природе.
Паразиты могут быть временными, когда организм хозяина подвергается нападению на короткий срок или на время питания (клопы). Всесветное распространение имеет постельный клоп. Существует поцелуйный клоп.
-обитает в тропиках. -ведет ночной образ жизни. -заселяет тростниковые или камышовые хижины. -нападая на человека, клопы прокалывают кожу около глаз или губы; напившись крови, клоп выпускает на место укуса каплю
экскрементов, в которых содержатся трипаносомы, возбудители тяжелой болезни.
К временным паразитам относятся слепни, комары, блохи. Они наносят организму хозяина ущерб, что может привести к гибели.
Есть и постоянные паразиты; к ним относятся простейшие – малярийный плазмодий, дизентерийная амеба, плоские черви (аскариды), членистоногие (вши), чесоточный зудень.
При постоянном паразитизме организм хозяина является единственным местом обитания для паразита. Например: в крови африканских антилоп находятся жгутиковые простейшие, переносчиком которых является кровососущая муха
ЦЦ. Антилопам жгутиковые простейшие вреда не приносят. Но если эти простейшие попадают в кровь человека, то развивается сонная болезнь (она приводит к смерти). Паразиты могут поселяться не только в крови, но и в тканях и
полостях тела. Например, личинки бычьего цепня с недостаточно проваренном или прожаренном мясом попадают в кишки человека, где они превращаются во взрослую особь, длина которой 9–10 метров. Цепни питаются содержанием
тонкого кишечника: они всасывает питательные вещества всей поверхностью тела, в результате лишая хозяина части пищи. Ленточные черви ведут паразитический образ жизни. Цикл их развития сопровождается сменой хозяев.
Например, черви длиной до 1 метра в половозрелом состоянии паразитируют в кишках рыбоядных птиц. С пометом птиц яйца попадают в воду, они проглатываются циклопом, в его теле образуется личинка, она вступает в следующую,
личиночную, стадию уже в полости тела пресноводных рыб. На этой стадии личинки достигают крупных размеров и сильно снижает жизнеспособность организма хозяина, вызывая даже массовую гибель рыбы. Паразитические
отношения встречаются и у растений. Широко распространены паразитические бактерии и грибы. Они поселяются на вегетативных органах древесных и травянистых растений, вызывая заболевания. Изучение паразитов необходимо для
предупреждения и лечения заболеваний человека. Паразитология изучает биологию, экологию паразитов, методы диагностики, лечения и профилактики паразитов. Медицинская паразитология подразделяется на: протозоологию
(простейшие), гельминтологию (паразитические черви) и арахноэнтомологию (паукообразные и насекомые).
Поражения тканей.
Кроме эрозии, вызванной инвазией и механическим разрушением клеток паразитом, можно различить три главных типа повреждений клеток и тканей.
1) Паренхиматозная и альбуминовая дегенерация характеризуется увеличением клеток из-за наполнения их гранулами альбумина или жира, нечетко видимым ядром и бледной цитоплазмой. Часто наблюдается при поражении
паразитами клеток печени, почек, сердца.
2) Жировая дегенерация - накопление в клетках аномальных количеств жира, придающее им желтоватую окраску, характерно для ивазированных паразитом клеток печени.
3) Некроз. Постоянная дегенерация любого типа вызывает смерть клеток и тканей. Так, некроз окружающих клеток вызывает инцистирование и кальцификацию личинок в мышечной ткани.
Изменения тканей.
Паразиты часто вызывают серьёзные изменения процессов роста и обновления тканей..
1) Гиперплазия. Повышение уровня метаболизма в клетках ведёт к усиленной клеточной пролиферации путем ускорения клеточных делений. Это связано с повышением репаративной активности тканей которая обычно сопровождает
вызванное паразитом воспаление.
2) Гипертрофия. Увеличение размеров клеток и тканей характерно при инвазиях внутриклеточных паразитов.
3) Метаплазия – преобразование одного типа тканей в другой.
4) Неоплазия. Аномальный рост может приводить к появлению опухолей, в том числе и злокачественных, вовлекающих как окружающие ткани, так и метастазирующие в другие органы.
Взаимодействия паразит-хозяин влекут физиологические, биологические, морфологические и иммунологические адаптации обоих. Они состоят в изменения энергетических процессов, энзиматической активности, развитии защитных
механизмов, экскреции и секреции ряда веществ и др. Способность организмов хозяев сопротивляться инвазиям паразитов может развиваться несколькими путями. Ряд физических и химических барьеров предотвращает внедрение
паразитов и их миграцию в теле хозяев. Например, птичьи сосальщики не могут внедриться в кровеносную систему человека. Если паразит преодолевает все барьеры, он подвергается действию реакций неспецифического и
специфического иммунитета. Врождённый иммунитет обусловлен химическими, физиологическими и другими особенностями хозяина. Физиологические состояния хозяина: рН и температура и другие также влияют на взаимодействия с
паразитом. Важнейшими факторами являются антитела сыворотки крови и фагоциты. Наличие антител является результатом предыдущих взаимодействий организма с бактериальными или другими агентами химически сходными с
имеющимися на поверхности паразита
107.
96. Тип Простейшие. Класс Саркодовые. Значение для медицины.Простейшие или протисты - это обширная группа одноклеточных организмов, которую можно
найти в различных средах, включая воду, почву и внутренние органы животных.
Класс Саркодовые включает в себя простейших, которые двигаются и питаются за счет
вытягивания псевдоподий - подвижных ветвей цитоплазмы.
Значение класса Саркодовых в медицине заключается в том, что некоторые из них являются
патогенными организмами и могут вызывать разные инфекционные заболевания у человека.
Для медицины важно изучать и понимать структуру и поведение саркодовых простейших,
чтобы разработать эффективные методы лечения и профилактики инфекций, вызываемых
этими организмами.
97. Тип Простейшие. Класс Жгутиковые. Значение для медицины
Простейшие (Protozoa) - это разнообразная группа одноклеточных микроорганизмов, которые
не образуют тканей или органов. Они могут быть свободно живущими или паразитирующими.
Класс Жгутиковые (Flagellata) относится к типу Простейшие и включает организмы, которые
двигаются с помощью жгутиков (жгутиками).
В медицине, некоторые виды жгутиковых простейших могут быть патогенными и вызывать
различные заболевания. Например, одноклеточный паразит, вызывающий венерическое
заболевание, называемое трихомониазом. Также существуют другие патогенные виды
жгутиковых простейших, которые могут вызвать инфекции у человека.
108.
98. Тип Простейшие. Класс Споровики. Значение для медициныSporozoa
Известно около 1400 видов споровиков. Все представители класса являются паразитами человека и животных. Многие споровики — внутриклеточные паразиты.
Питание:
Они не имеют никаких органов выделения и пищеварения. Питание происходит за счет поглощения пищи всей поверхностью тела. Продукты жизнедеятельности
также выделяются через всю поверхность мембраны.
Органелл дыхания нет. Общими чертами всех представителей класса являются отсутствие у зрелых форм каких-либо органелл движения, а также сложный
жизненный цикл.
Для споровиков характерны два варианта жизненного цикла — с наличием полового процесса и без него. Первый вариант цикла включает в себя стадии бесполого
размножения и полового процесса.
Размножение:
Бесполое размножение осуществляется простым делением с помощью митоза или множественным делением (шизогонией). При шизогонии происходит
многократное деление ядра без цитокинеза. Затем вся цитоплазма разделяется на части, которые обособляются вокруг новых ядер. Из одной клетки образуется
очень много дочерних. Перед половым процессом происходит образование мужских и женских половых клеток — гамет. Они называются гамонтами. Затем
разнополые гаметы сливаются с образованием зиготы. Она одевается плотной оболочкой и превращается в цисту, в которой происходит спорогония —
множественное деление с образованием клеток (спорозоитов). Именно на стадии спорозоита паразит и проникает в организм хозяина. Споровики, для которых
характерен именно такой цикл развития, обитают в тканях внутренней среды организма человека (например, малярийные плазмодии).
Второй вариант жизненного цикла намного проще и состоит из стадии цисты и трофозоита (активно питающейся и размножающейся формы паразита). Такой цикл
развития встречается у споровиков, которые обитают в полостных органах, сообщающихся с внешней средой.
В основном споровики, паразитирующие в организме человека и других позвоночных, обитают в тканях тела. Они могут поражать как человека, так и многих
животных (в том числе и диких). Таким образом, это зоо- и антропозоонозные заболевания, профилактика которых представляет собой сложную задачу. Эти
заболевания могут передаваться нетрансмиссивно (как токсоплаз-мы), т. е. не иметь специфического переносчика, или трансмиссивно (как малярийные плазмодии),
т. е. через переносчиков.
Диагностика заболеваний, вызываемых простейшими класса Споровики, довольно сложна, так как паразиты могут обитать в различных органах и тканях (в том
числе глубоких), что снижает вероятность их обнаружения. Выраженность симптомов заболевания невелика, поскольку они не являются строго специфичными.
Токсоплазмы (Toxoplasmagondii) — возбудители токсоплазмоза. Человек для этого паразита является промежуточным хозяином, а основные хозяева — это кошки и
другие представители семейства Кошачьи.
Малярийные плазмодии (Plasmodium) — возбудители малярии. Человек — промежуточный хозяин, окончательный — комары рода Anopheles.
Споровики имеют очень большое практическое значение для медицины, так как к этому отряду относится возбудитель тяжелого заболевания человека — малярии.
Существует четыре вида рода Plasmodium, вызывающие малярию (P. vivax, P. malaria, P. falciparum, P. ovale). Клиническая картина болезни, вызываемой разными
видами паразита, несколько различна.
109.
99. Тип Простейшие. Класс Инфузории. Значение для медицины.Вегетативная форма – имеет длину 30-200 мкм; округлую или овальную форму, но чаще в передним зауженным и задним расширенным концом. Питается углеводами, др. микроскопическими
организмами, а может фагоцитировать эритроциты и лейкоциты.
Может не наносить существенного вреда хозяину, а может внедряться в слизистые кишечника, образуя язвы размером до нескольких сантиметров, усиленно питаться и размножаться, тем самым
вызывая некроз тканей. Балантидиоз возникает чаще всего из-за обедненности микрофлоры кишечника.
Летальность этого заболевания высокая, диагностируют по обнаружению вегетативных форм в нативных мазках из фекалий путем прямого микроскопирования.
Распространено повсеместно, основной источник заражения – домашняя свинья. Заражение происходит при проглатывании цист, а иногда и вегетативных форм
Сохраняются вегетативных формы в свежих фекалиях в течение 30 часов, в водопроводной или застойной воде в течение 7 суток; в трупах в течение 3 суток со дня смерти.
Пути передачи совпадают с таковыми амебиаза. За переболевшими людьми ведется наблюдение в течение года, делаются 2-3 контрольные проверки.
Классическая систематика
Всего данный тип, по различным данным, содержит 6-7 тысяч видов. Тип содержит в себе два класса — класс Ресничных инфузорий (Ciliata) с тремя надотрядами и класс Сосущих инфузорий
(Suctoria)
Класс Ciliata — ресничные инфузории.
Надотряд Kinetofragminofora — самая примитивная и разнообразная группа инфузорий. Реснички располагаются по телу равномерно, обычно продольными рядами, но некоторые части тела могут
быть лишены ресничек.
Отряд Entodiniomorpha Отряд Gymnostomata Отряд Hypostomata Отряд Entodiniomorpha
Надотряд Oligohymenophora — для инфузорий относящихся к данному надотряду характерно образования в области ротового отверстия специфического аппарата тетрахимениума — слагаемого из
трех мембранелл (состоящих из мелких жгутиков) левее рта и трех мембранелл правее.
Отряд Hymenostomata Отряд Peritrichia (подотряд Sessilia (сидячие), подотряд Mobilia (свободноживущие и паразиты)
Надотряд Polyhemenophora — характеризуются усложнением тетрахимениума — спирально закрученная зона околоротовых мембранелл сопровождающаяся увеличением числа мембранелл.
Реснички или равномерно покрывают тело или образуют цирры.
Отряд Heterotrichia Отряд Hypotrichia Отряд Olygotrichi
Класс Suctoria — сосущие инфузории. Характеризуются отсутствием во взрослом состоянии ресничек, рта и глотки. Наличествуют щупальца, одно или несколько, ветвящиеся и не ветвящиеся, на
конце которых существует канал.
-Тело инфузории покрыто оболочкой – пелликулой. Последняя способствует сохранению определенной формы тела инфузории.
-Органоидами движения являются реснички, которые могут быть одинаковыми или различными по длине. Имеется сложно устроенный перистом (предротовой аппарат), связанный с клеточным
ртом – цитостомом.
-Органоидами пищеварения являются пищеварительные вакуоли; органоидами выделения – сократительные вакуоли. Помимо ресничек, характерным признаком этого класса простейших
является строение ядерного аппарата. В эндоплазме имеется два ядра: макронуклеус (большое ядро) и микронуклеус (малое ядро).
-Инфузории размножаются как бесполым, так и половым путем. Бесполое размножение – поперечное деление надвое, которому предшествует половое размножение – конъюгация. Процесс
конъюгации заключается в том, что временно сходятся две особи, между которыми происходит обмен микронуклеусами.
-Инфузории могут инцистироваться. Представителями этого класса является большинство свободно живущих организмов, но встречаются также и паразитические формы (например, паразит
кишечника человека – балантидий).
110.
100. Тип Плоские черви. Класс Сосальщики. Значение для медицины.Тип плоских червей относится к группе животных, известных как плоские черви или плоскожаберные. Один из классов плоских червей - это
класс сосальщиков, из которого наиболее известные представители - свиные и говяжьи цепни, а также цепень эхинококк.
Значение плоских червей класса сосальщики для медицины связано с тем, что они являются паразитами человека и животных, и могут
вызывать серьезные заболевания.
Плоские черви класса сосальщики могут вызывать различные заболевания, такие как цепная глистная инфекция и эхинококкоз. Эти
заболевания могут привести к различным симптомам, включая желудочно-кишечные расстройства, анемию, утомляемость, потерю веса и
даже внутренние органы.
Из-за важности понимания и контроля этих паразитических инфекций, медицинская наука и практика работают над методами диагностики,
лечения и предотвращения данных заболеваний. Диагноз основывается на клинических симптомах, анализе фекалий, использовании
образовательного материала и проведении лечебных процедур.
101. Тип Плоские черви. Класс Ленточные черви. Значение для медицины.
В классе Ленточные черви (Cestoda) находятся плоские черви, которые являются паразитами кишечника человека и животных. Эти черви имеют
длинное и полосатое тело, состоящее из сегментов или проглотов.
Перекосли и ленточные черви могут вызывать различные заболевания у человека. Одним из наиболее известных заболеваний, вызываемых
ленточными червями, является тенидиоз (теньез). Он вызывается заложником (булавоголовкой) Taenia solium. Инфекция происходит через
употребление сырого или недостаточно приготовленного мяса свиньи, содержащего кисты заложников.
Медицинское значение ленточных червей состоит в предупреждении и лечении инфекций. При подозрении на инфекцию червеобразными
паразитами важно обратиться к врачу для диагностики и эффективного лечения.
102. Тип Круглые черви. Значение для медицины.
Тип "Круглые черви" (Nematoda) относится к группе животных, известных как нематоды или круглые черви. Это разнообразная группа червей с
более чем 25 000 описанными видами. Они распространены повсеместно и обитают в различных средах, включая почву, пресноводные и
морские экосистемы, а также паразитируют в тканях животных и растений.
Некоторые виды круглых червей являются паразитами человека и имеют важное значение для медицины. Например, одним из наиболее
известных нематод, паразитирующих в организме человека, является гельминт Ascaris lumbricoides, вызывающий болезнь аскаридоз.
111.
103. Тип Членистоногие. Класс Паукообразные. Значение для медицины.Членистоногие - это огромная группа животных, включающая в себя такие классы, как насекомые, ракообразные и
паукообразные. Класс паукообразные включает аранеид, скорпионов, клещей и других подобных видов.
Паукообразные имеют значительное значение для медицины по нескольким причинам:
1. Аллергические реакции
2. Токсины
3. Медицинские исследования
4. Продукты
Таким образом, паукообразные имеют значительное значение для медицины и могут вносить важный вклад в
различные области медицинской науки и практики.
104. Тип Членистоногие. Класс Насекомые. Значение для медицины.
Членистоногие – это широкий тип животных, включающий в себя такие группы, как насекомые, паукообразные,
многоножки и ракообразные. Одной из крупнейших групп Членистоногих являются насекомые, составляющие
большую долю биоразнообразия на Земле.
Насекомые имеют огромное значение для медицины. Некоторые из этих значений включают:
1. Лекарственные свойства
2. Изучение заболеваний
3. Биомедицинские исследования
4. Использование в хирургии
В целом, насекомые играют важную роль в медицине, как объекты исследования, источники лекарственных веществ и
инструменты для развития новых медицинских технологий и лечений.
112.
105. Сущность эволюции. Микро - и макроэволюция. Характеристика механизмов и основных результатов.Термин «эволюция» (от лат.evolutio— развертывание) ввел в XVIII в. швейцарский натуралист Шарль Бонна.
Под эволюцией в биологии понимают необратимое историческое развитие живой природы. Эволюция организмов затрагивает все процессы жизни, происходящие на молекулярном уровне, на уровне отдельных биохимических или физиологических процессов, всего организма, вплоть
до процессов, развивающихся в сообществах организмов, экосистемах и биосфере в целом. В биологии эволюция рассматривается как сила, ведущая к образованию новых форм организмов, как процесс, благодаря которому доклеточные формы жизни, возникшие более 3 млрд лет
назад, дали начало исключительно сложным многоклеточным организмам нашего времени. Понятие «эволюция» пронизывает все области биологии как основная объединяющая идея наших представлений о жизни и о функциях организмов.
Микроэволюция
Эволюция на уровне вида (на внутривидовом уровне) получила название микроэволюции. В рамках учения о микроэволюции различают элементарную эволюционную единицу, элементарное эволюционное явление, элементарный эволюционный материал и элементарные факторы
эволюции. Элементарной единицей эволюции является популяция. Популяция (от фр. population) — это совокупность особей одного вида, длительно (в течение многих поколений) населяющих определенную территорию, свободно скрещивающихся между собой и отделенных от других
популяций этого же вида давлением изоляции. В составе вида может быть одна или несколько (много) популяций.
Элементарным эволюционным явлением служит мутация. В результате мутаций популяции становятся генетически гетерогенными, т. е. представляют собой смесь разных генотипов. Если условия существования популяции постоянны, ее генетический состав в среднем будет
неизменным. Однако, если имеется давление со стороны каких-либо внешних факторов, то тогда происходит изменение генотипического состава популяций.
Мутации случайны во времени и пространстве применительно к отдельным организмам. Однако мутационные изменения организмов в популяции не являются направляющими для эволюционного процесса. Направления изменений организмов определяются естественным отбором.
Элементарным эволюционным материалом служат любые генотипически различные особи и группы особей, т. е. мутантные организмы или особи-носители мутаций, различающиеся между собой генетически. С эволюционной точки зрения важнейшей характеристикой мутантных
организмов как элементарного эволюционного материала является частота появления их в природных популяциях. Наличие организмов-носителей мутаций создает различия между близкими природными группами особей (популяциями, подвидами, видами).
Элементарными факторами эволюции служат естественный отбор, мутационный процесс, популяционные волны, изоляция, дрейф генов, миграция, с действия которых начинается эволюция в популяциях. Естественный отбор является важнейшим направляющим фактором эволюции,
поскольку его основная функция заключается в устранении из популяций организмов с неудачными комбинациями генов и сохранение генотипов, которые не нарушают процесса приспособительного формообразования. Действие естественного отбора проявляется в пределах
популяции, но объектами приложения естественного отбора являются отдельные мутантные особи, которые являются элементарным материалом, на котором работает естественный отбор.
Естественный отбор является результатом борьбы за существование, представляющейся обычно в двух формах — внутривидовой и межвидовой. Внутривидовая борьба за существование является наиболее упорной, т. к. организмы одного и того же вида конкурируют за сходные условия
существования (за свет, воду, места охоты, самку и т. д.). Межвидовая борьба приобретает остроту у видов, обитающих в сходных экологических условиях.
Мутационный процесс в качестве элементарного фактора эволюции важен тем, что он является поставщиком элементарного эволюционного материала (мутантных организмов), поддерживая генетическую гетерогенность природных популяций. Однако важно заметить, что, выполняя эту
роль, мутационный процесс в качестве фактора эволюции также не направляет ход эволюционных изменений. Эту функцию осуществляет естественный отбор.
Популяционные волны или волны жизни (колебания численности особей в популяциях в сторону от средней численности) являются элементарными факторами эволюции по той причине, что они тоже служат поставщиками элементарного эволюционного материала для естественного
отбора.
Примерами их служат колебания численности вредителей полей и огородов в разные годы. Эволюционное значение волн жизни состоит в том, что они резко изменяют (повышают) концентрацию в популяциях редко встречающихся аллелей (генотипов), в результате чего под отбор
попадают организмы, обладающие редкими генами, которые обычно ускользают от действия отбора.
Дрейф генов, или генетико-автоматический процесс, является следствием популяционных волн и, как отмечено выше, заключается в том, что в малых популяциях имеет место либо потеря какого-то аллеля, либо резкое повышение его концентрации. Примером дрейфа генов является
утрата многих аллелей у баптистов, переселившихся около 250 лет назад из Германии в США. Но дрейф генов также не определяет направления эволюции.
Изоляция в качестве элементарного фактора обеспечивает барьеры, исключающие репродукцию. Различают географическую, экологическую и генетическую изоляцию, каждая из которых ведет в конечном итоге к репродуктивной изоляции на межвидовом уровне. При этом установлены
презиготические и постзиготические механизмы, предотвращающие скрещивания.
К презиготическим изолирующим механизмам относят географическую и экологическую изоляции, которые исключает встречи потенциальных брачных партнеров. Известна также поведенческая изоляция, когда потенциальные брачные партнеры встречаются, но из-за недостаточного
полового влечения эти встречи заканчиваются отрицательно, и сезонная изоляция, когда партнеры встречаются, но не спариваются, ибо спаривание у них в нормальных условиях обычно происходит в разное время. Наконец, известна гаметическая изоляция, при которой после
совокупления животных не происходит осеменения, т. к. сперматозоиды утрачивают жизнеспособность в женских половых путях.
Постзиготические изолирующие механизмы также различны. Например, после встречи партнеров яйцеклетки могут оплодотвориться, но зиготы погибают. Если зиготы развиваются, то гибриды все же не достигают половой зрелости.
Далее могут формироваться гибриды, но их жизнеспособность оказывается невысокой, либо они могут быть стерильными (не способными продуцировать гаметы, обладающие нормальными свойствами).
Важнейшей особенностью изоляции как элементарного фактора эволюции является ее длительность. Нарушая скрещивания и закрепляя различия в генотипах, изоляция как бы расчленяет популяцию на генотипически различные группы организмов, полностью исключая среди них обмен
генами, усиливая дивергенцию признаков. Изоляция является, по существу, усилителем начальных стадий генотипической дифференциации, развертывающейся в популяциях. Она подставляет разделенные части популяций под действие естественного отбора. Однако изоляция тоже не
направляет эволюцию. Это делает только естественный отбор.
Миграция аллелей (мутантных организмов) ведет к изменению в популяциях как частот аллелей, так и генотипов. Например, негритянское население США унаследовало около 30% генов от белых предков. Миграция способствует объединению генных пулов популяций.
Сущность микроэволюционных преобразований составляет изменение генетической структуры популяций. Видообразование, т. е. появление нового вида — это центральный и важнейший завершающий этап эволюции.
Макроэволюция
Процесс образования из видов новых родов, из родов — новых семейств и так далее называют макроэволюцией. Результатом макроэволюционных процессов становятся существенные изменения внешнего строения и физиологии организмов — такие, например, как формирование
замкнутой системы кровообращения у животных или появление устьиц и эпителиальных клеток у растений.
Макроэволюция — надвидовая эволюция, в отличие от микроэволюции, происходящей внутри вида, внутри его популяций. Однако принципиальных различий между этими процессами нет, так как в основе макроэволюционных процессов лежат микроэволюционные, В макроэволюции
действуют те же факторы — борьба за существование, естественный отбор и связанное с ним вымирание. Макроэволюция, так же как микроэволюция, носит дивергентный характер.
Макроэволюция происходит в исторически грандиозные промежутки времени, поэтому она недоступна непосредственному изучению. Несмотря на это, наука располагает множеством доказательств, свидетельствующий о реальности макроэволюционных процессов.
113.
106. Биологический вид и его определение. Критерии вида.Вид. С развитием биологии пришло понимание того, что по сравнению с бесконечным разнообразием условий, в которых протекает жизнь, разнообразие форм организмов конечно; оно как бы
собрано в «узлы» — биологические виды.
Биологический вид — это совокупность особей, обладающих способностью к скрещиванию с образованием плодовитого потомства; населяющих определенный ареал; обладающих рядом общих
морфологических и физиологических признаков и сходством во взаимоотношениях с биотической и абиотической средой.
Биологический вид — не только систематическая категория. Это целостный и обособленный от других видов элемент живой природы. Целостность вида проявляется в том, что его особи могут
жить и размножаться, лишь взаимодействуя друг с другом благодаря выработанным в процессе эволюции взаимным приспособлениям организмов: особенностям скоординированности строения
материнского организма и эмбриона, системам сигнализации и восприятия у животных, общности территории, сходству жизненных привычек и реакций на сезонные изменения климата и др.
Видовые приспособления обеспечивают сохранение вида, хотя иногда они могут повредить отдельным особям. Речной окунь, например, питается собственной молодью, за счет чего вид
выживает при недостатке корма, даже несмотря на утрату части приплода. Каждый вид существует в природе как исторически возникшее целостное образование.
Обособленность вида поддерживается репродуктивной изоляцией (см. § 59), которая препятствует его смешению с другими видами при размножении. Изоляция обеспечивается различиями в
строении половых органов, разобщенностью ареалов, расхождением в сроках или местах размножения, различиями поведения, экологической разобщенностью и другими механизмами, о
которых вы узнаете из последующих разделов. Обособленность видов препятствует возникновению промежуточных форм. Береза бородавчатая, например, не растет на моховых болотах, где
обычно растет береза карликовая. Благодаря изоляции виды не смешиваются друг с другом.
Критерии вида. Характерные признаки и свойства, по которым одни виды отличаются от других, называют критериями вида.
Морфологический критерий — это сходство внешнего и внутреннего строения организмов. Карл Линней, например, определял виды как целостные группы организмов, отличные от других
жизненных форм по признакам строения. Иными словами, наличие черт строения, которые делают некоторую группу организмов похожими друг на друга и одновременно отличными от всех
других групп, и есть критерий для причисления их к данному виду.
Особи в пределах вида иногда настолько изменчивы, что только по морфологическому критерию не всегда удается определить вид. Существуют виды морфологически сходные. Это — видыдвойники, которые открыты во всех систематических группах. Например, у черных крыс известно два вида-двойника — с 38 и 49 хромосомами; у малярийного комара — б видов-двойников, у
небольшой рыбки щиповки, широко распространенной в пресных водоемах, — 3 таких вида. Виды-двойники встречаются среди самых различных организмов: рыб, насекомых, млекопитающих,
растений, однако особи таких видов-двойников не скрещиваются между собой
Генетический критерий — это характерный для каждого вида набор хромосом; строго определенное их число, размеры и формы, состав ДНК. Хромосомный набор — главный видовой признак.
Особи разных видов имеют разные наборы хромосом, поэтому они не могут скрещиваться и репродуктивно ограничены друг от друга в естественных условиях.
Физиологический критерий — сходство реакций организма на внешние воздействия, ритмов развития и размножения. В основе этого критерия лежит сходство всех процессов жизнедеятельности,
и прежде всего размножения. Представители разных видов, как правило, не скрещиваются или их потомство бесплодно. Однако встречаются исключения. Например, собаки могут давать
потомство, спариваясь с волками. Плодовитыми могут быть гибриды некоторых видов птиц (канарейки, зяблики), а также растений (тополя, ивы). Следовательно, физиологический критерий также
недостаточен для определения видовой принадлежности особей.
Экологический критерий — это характерное для вида положение в природных сообществах, его связи с другими видами, наборы факторов внешней среды, необходимые для существования.
Географический критерий — область распространения, определенный ареал, занимаемый видом в природе.
Исторический критерий — общность предков, единая история возникновения и развития вида.
Критерии вида связаны между собой и определяют качественную особенность вида. Но ни один из них не является абсолютным. Например, два разных вида могут не различаться по
анатомическому строению и обладать одинаковыми хромосомными наборами. Но если они различаются по поведению, то не скрещиваются между собой и, следовательно, обособлены один от
другого. Лишь в совокупности перечисленные критерии позволяют с достаточной надежностью установить принадлежность организма к тому или иному виду.
Виды представляют собой определенный уровень организации живой материи — видовой.
114.
107. Популяция - элементарная единица эволюции.(От лат. populus - народ, населения) - совокупность всех представителей данного вида, занимающих определенную область в одно и то же
время. Важная особенность популяции - большое генотипическое сходство составляющих ее особей и как следствие сходство во всех свойствах
и признаках по сравнению с особями даже соседних популяций того же вида. Это объясняется тем, что внутри популяции случайное свободное
скрещивание и "перемешивание" особей осуществляются легче и чаще, чем между различными популяциями, из-за их территориальной
обособленности друг от друга. Например, одна дубрава от другой находится за несколько километров, а пыльца дуба разносится на несколько
сот метров. Однако в сильные бури пыльца, подхватываемая ветром, может переноситься на значительно большие расстояния и достигать
соседней популяции. Другими словами, обособленность популяций относительна. Понятно, почему важно знать расстояние, на которое
перемещаются особи в природе, т.е. как далеко они могут перенести свои гены и передать их следующему поколению.
Популяция - самая мелкая из групп особей, способная к эволюционному развитию, поэтому ее и называют элементарной эволюционной
единицей. Отдельно взятый организм не является элементарной эволюционной единицей, так как его генотип остается неизменным на
протяжении всей жизни. Популяция, благодаря большой численности особей, представляет собой непрерывный поток поколений.
Вид как целостная замкнутая система может существовать на протяжении длительного времени, и его можно было бы принять за
эволюционную единицу. Однако вид распадается на составляющие его части - популяции. Вот почему роль элементарной эволюционной
единицы приходится все-таки признать не за видом, а за популяцией.
Итак, для популяции характерна сильная, хотя и не абсолютная обособленность (изоляция) от других популяций этого вида, обеспечивающая
большое генотипическое сходство составляющих ее особей. Вместе с тем у отдельных особей популяции возникают наследственные
изменения, которые в результате свободного скрещивания распространяются в популяции и приводят к генетической разнородности
составляющих ее особей.
Неоднородность особей в популяции создает условия для действия естественного отбора. Следовательно, эволюционный процесс начинается в
недрах вида - в популяциях. Вот почему без понимания сущности вида, его популяционной структуры нельзя понять закономерности эволюции.
Вид - генетически целостная и замкнутая система. Наиболее существенной характеристикой вида является то, что вид представляет собой
генетически целостную и замкнутую систему. Благодаря целостности вида особи разных популяций могут скрещиваться между собой. Поэтому
в случае появления благоприятной мутации последняя, переходя от популяции к популяции, в сравнительно ограниченное число поколений
может быстро распространиться по всему ареалу вида, но она не может перейти из одного вида в другой из-за обособленности видов друг от
друга специальными барьерами (различия в сроках размножения, в брачном поведении, в хромосомных наборах и т.п.).
115.
108. Элементарные эволюционные факторы.Элементарные эволюционные факторы
1. Изменчивость
Мутационная изменчивость – основная форма; в виде генных, хромосомных и геномных мутаций. Хотя мутации происходят редко (вероятность генной мутации10-6 ), но в целом накапливается значительное количество. Большинство
накапливающихся мутаций – рецессивные (доминантные тут же проявляются и подвергаются жесткому отбору), при этом популяция остается генетически однородной – скрытый резерв наследственной изменчивости. Доля полезных
мутаций мала (1 на 109 вредных), но за период существования вида накапливаются значительные количество, что может привести к образованию нового вида, более приспособленного к новым условиям существования.
Комбинативная изменчивость – следствие механизмов мейоза (кроссинговер, случайное расхождение гомологичных хромосом, случайное сочетание гамет), большое разнообразие генотипов и фенотипов – исходный материал для
естественного отбора.
2.
Естественный отбор – избирательное (дифференциальное) воспроизведение определенных генотипов (выживание наиболее приспособленных), функционирует при наличии 2 факторов: интенсивного размножения особей и
борьбы за существование.
Интенсивность размножения – рождается намного больше особей, чем может выжить Гибель потомства – результат борьбы за существование.
Различают 3 вида борьбы за существование:
-
межвидовая (между хищниками-плотоядными и травоядными, травоядными и растениями, птицами и насекомыми, между культурными растениями и сорняками, между микроорганизмами разных видов и т.д.);
-
внутривидовая (между особями одного вида – рыжие тараканы вытесняют черных, серая крыса – черную, конкуренция за места гнездования, самцов – за самку, каннибализм у рыб);
с неблагоприятными условиями внешней среды (с засухой, заморозками, переувлажнением - растения, недостаток кислорода в воде – рыбы, резкие колебания температуры – возникновение теплокровности и шерстного
покрова).
Кто выживает в этой борьбе? Те виды, которые наиболее приспособлены к конкретным определенным условиям окружающей среды. Следовательно, эволюционный процесс носит приспособительный характер, а целесообразность
обусловлена взаимосвязью и соответствием всех органов в организме условиям существования в определенных условиях.
Примеры приспособлений: внешний вид – покровительственная окраска, маскировка, предупреждающая окраска, мимикрия.
В зависимости от направленности и результата различают следующие формы естественного отбора:
Направленный (движущий) – вызывает прогрессивное или направленное в одну строну изменение генетического состава популяции, проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления (ослабления),
характерен для популяций в процессе приспособления к новой среде (при медленно изменяющихся условиях внешней среды). Этот принцип используют при искусственном отборе. Пример: индустриальный меланизм бабочек.
Стабилизирующий отбор – сохраняет в популяции средний вариант фенотипа (признака), устраняет из репродуктивного процесса фенотипы, отклоняющиеся от нормы. Наиболее распространенная форма, характерна для постоянных условий
окружающей среды.
Дизруптивный (разрывающий) отбор – сохраняет крайние формы, направлен против «средних» особей.
Дарвин: «Естественный отбор ежедневно, ежечасно расследует по всему свету мельчайшие изменения, отбрасывая дурные, сохраняя и слагая хорошие, работая неслышно, невидимо, где бы и когда бы только ни представился к тому случай,
над усовершенствованием каждого органического существа.
2.
Изоляция – возникновение любых барьеров, препятствующих скрещиванию организмов, связана с другими факторами. Различают:
o Географическую (пространственную) – разрыв единого ареала водными или наземными преградами, обособление отдельных популяций, в каждой свои мутации, изменчивость, отбор, изменение генотипа, изменение фенотипа;
o Биологическую (репродуктивную) из-за несовпадения сезонов размножения, особенностей ритуала ухаживания, различия в строении органов размножения, несовместимость гамет.
3.
Популяционные волны – периодические колебания численности организмов в природных популяциях. Причины различны: соотношение численности хищников и травоядных, деятельность человека (размножение кроликов в
Австралии, колорадского жука и т.д.). Рост численности сопровождается расширением территории, возможно слияние с другими популяциями, выселение за пределы ареала, где могут быть другие условия существования. При спаде
численности распад крупных популяций, изоляция отдельных групп, в которых возможно сохранение редких мутационных аллелей, приводящих к возникновению новых видов.
4.
Дрейф генов (генетико-автоматические процессы). Когда особей в популяции мало, возможны случайные отклонения от общих закономерностей наследственности и изменчивости, которые могут привести к утрате или
закреплению признака вне его связи с приспособительной ценностью
116.
109. Микроэволюционные процессы в популяциях людей.Микроэволюционные процессы в популяциях людей - это изменения в генетическом составе популяций, которые происходят на уровне отдельных генов или аллелей со временем. Вот несколько примеров микроэволюционных процессов,
которые могут происходить в популяции людей:
1. Естественный отбор
2. Мутация
3. Генетический поток
4. Генетический дрейф
В целом, эти процессы могут приводить к изменению генетического разнообразия, адаптации к окружающей среде и эволюции популяций.
Микроэволюция – изучает эволюционные события, протекающие внутри вида.
Особенности мутационного процесса
В последнее время давление мутационного процесса на генофонд человеческих популяций усиливается, что связано с загрязнением окружающей среды мутагенными факторами (химические, ионизирующая радиация, аварии на АЭС).
Увеличение частоты мутаций ведет к увеличению частоты наследственных заболеваний.
Особенности действия изоляции
На ранних этапах становления человека в человеческих популяциях изолирующими барьерами являлись географические преграды (горы, моря и океаны). В настоящее время в качестве изолирующих барьеров выступают социальные факторы:
культура, религия, язык, экономический уклад, национальные обычаи и т.п. Они оказались очень стойкими.
Действие изоляции на протяжении длительного времени привело к морфологическим различиям отдельных популяций людей.
В человеческих популяциях действует стабилизирующая форма естественного отбора, направленная на сохранение генофонда популяций. Стабилизирующий отбор может быть как положительным, так и отрицательным.
Положительный естественный отбор направлен на сохранение особей, отрицательный – на устранение из популяции особей с нежелательным генотипом.
Отрицательный отбор в свою очередь может быть направлен против гетерозигот и против гомозигот.
Положительный отбор, направленный против отрицательного отбора, называется контротбор.
Генофонд любой популяции – результат длительных и сложных взаимодействий элементарных эволюционных факторов (мутационного процесса, дрейфа генов, популяционных волн, изоляции, естественного отбора). В связи с этим все
природные популяции генетически полиморфны.
Генетический полиморфизм – наличие в популяции нескольких генетических форм в состоянии длительного равновесия. Генетический полиморфизм бывает адаптационный и балансированный. Адаптационный (приспособительный).
Пример: у виноградной улитки часть особей популяции, обитающая на песчаных почвах, имеет белый вход в раковину, а другая часть – красный цвет, они обитают на глинистой почве. У двухточечной божьей коровки есть красные и черные
особи. Красные хорошо переносят зиму, а черные хорошо размножаются летом. Балансированный – гетерозиготный – отбор благоприятствует сохранению гетерозиготных особей. Имеет большой биологический смысл – обеспечивает
выживаемость особей в изменяющихся условиях окружающей среды, создает резерв наследственной изменчивости. Приспособленность любой реальной популяции всегда хуже приспособленности идеальной популяции, которая состояла
бы только из нужных в данный момент генотипов. Разность между приспособленностью идеальной и реальной популяций называется генетическим грузом. Различают генетический груз мутационный (обусловлен мутациями в данном
поколении) и сегрегационный (обусловлен мутациями, которые передались от предков).
В медицине наблюдаются следующие проявления генетического полиморфизма:
неравномерное распределение заболеваний на планете;
различная тяжесть течения заболеваний;
неодинаковая предрасположенность к наследственным заболеваниям.
117.
110. Происхождение жизни и эволюция органического мираХод развития органического мира на земле восстанавливается исследователями по данным палеонтологии, а также по данным накопленных морфологических и эмбриологических материалов. По установленным данным наша планета образовалась менее 7 млрд. лет назад.
Промежуток времени существования нашей планеты подразделяют на эры. Эры дополнительно подразделяют на периоды. На каждом этапе своего развития в органической жизни на Земле происходили определенные изменения.
1. Догеологическая эра
В этот период происходило формирование нашей планеты. Началось формирование около 7 млрд. лет назад, продолжалось менее 3 млрд. лет. В период зарождения и формирования планеты жизнь на Земле отсутствовала.
2. Архейская эра
В этот период на нашей планете в водной толще первых морей зародилась жизнь. К концу этой эры жизнь на Земле существовала в виде достаточно примитивных форм: одноклеточных бактерий и водорослей и лишь небольшим числом многоклеточных. Эволюция органического мира на этом этапе
претерпела сравнительно незначительные изменения. В эту эру произошло разделение на ветви развития животного и растительного мира, которые имели до этого общего прародителя – одноклеточных жгутиковых организмов. Разделение произошло на почве питания. Первичные животные остались
гетеротрофными организмами, а водоросли в своем развитии приобрели способности к фотосинтезу и стали автотрофными организмами.
3. Протерозойская эра
По своей продолжительности считается одной из самых длительных. В эту эру появились новые виды водорослей, которые постепенно стали исходными для всех групп растений. Массовое размножение этих видов водорослей в эту эру способствовало накоплению кислорода на планете, что сыграло
решающую роль в ходе эволюции животного мира. Эволюция органического мира на планете получила мощный толчок к своему развитию. Животный мир в ту эру прошел в своем развитии большой путь. На этом пути возникли новые типы червей и моллюсков. В конце протерозойской эры появились
простейшие членистоногие и бесчерепные хордовые. Основные формы жизни в этот период существовали только в воде.
4. Палеозойская эра
В эту эру происходили крупные события в развитии органического мира. Главным из них является выход растений и животных на сушу. Первыми на суше оказались бактерии, водоросли и низшие формы грибов. С их появлением на суше начались почвообразовательные процессы на планете. Достигнув
своего зенита в каменноугольный период, земноводные вынуждены были уступить место на суше пресмыкающимся. Наиболее интенсивное развитие пресмыкающихся наблюдалось в пермском периоде палеозойской эры. Эволюция органического мира в период этой эры заключалась в том, что растения
прошли путь от водорослей до голосеменных, а позвоночные животные от простейших хордовых до пресмыкающихся, находящихся как на суше. Получила свое развитие и одна из ветвей беспозвоночных животных. В своем развитии она прошла путь от простейших морских членистоногих до летающих
насекомых.
5. Мезозойская эра
По своему временному периоду была наполовину короче палеозойской. Развитие органического мира в эту эру происходило более быстрыми темпами. В триасовый период среди голосеменных растений появились хвойные растения. В юрский период появились покрытосеменные растения, быстро
развившиеся и которые приобрели разные формы. Эволюция органического мира не остановилась только на развитии растений. В триасовый период среди позвоночных животных появились первые млекопитающие животные, а в юрский период первые птицы.
6. Кайнозойская эра
Эта эра в историческом развитии планеты является более кроткой. Именно в эту эпоху появился на Земле человек. С появлением человека произошло изменение характера и направление эволюции органического мира на планете. В кайнозойскую эру произошла окончательная победа среди позвоночных
млекопитающих, птиц и костистых рыб. В эту эру происходило развитие высших представителей растительного и животного мира в тесном взаимодействии За время палеогена и неогена сформировалось современное очертание материков океанов и морей на земле. Последний период кайнозойской эры –
антропоген назван в честь человека, являющегося высшей формой развития живой материи и оказывающего на эволюцию и развитие органического мира наибольшее влияние.
Теория самопроизвольного (спонтанного) зарождения - таким образом, благодаря работам Луи Пастера теория самозарождения была признана несостоятельной и в научном мире утвердилась теория биогенеза, краткая формулировка которой — «все живое — от живого».
Теория креационизма предполагает, что все живые организмы (либо только простейшие их формы) были в определенный период времени сотворены («сконструированы») неким сверхъестественным существом (божеством, абсолютной идеей, сверхразумом, сверхцивилизацией и т.п.). Очевидно, что
именно этой точки зрения с глубокой древности придерживались последователи большинства ведущих религий мира, в частности христианской религии.
Теории стационарного состояния и панспермии
Обе эти теории представляют собой взаимодополняющие элементы единой картины мира, сущность которой заключается в следующем: вселенная существует вечно и в ней вечно существует жизнь (стационарное состояние). Жизнь переносится с планеты на планету путешествующими в космическом
пространстве «семенами жизни», которые могут входить в состав комет и метеоритов (панспермия). Подобных взглядов на происхождение жизни придерживался, в частности, основоположник учения о биосфере академик В.И. Вернадский.
Однако теория стационарного состояния, предполагающая бесконечно долгое существование вселенной, не согласуется с данными современной астрофизики, согласно которым вселенная возникла сравнительно недавно (около 16 млрд лет т.н.) путем первичного взрыва.
Очевидно, что обе теории (панспермии и стационарного состояния) вообще не предлагают объяснения механизма первичного возникновения жизни, перенося его на другие планеты (панспермия) либо отодвигая по времени в бесконечность (теория стационарного состояния).
Теория биохимической эволюции (теория А.И. Опарина)
Из всех теорий происхождения жизни наиболее распространенной и признанной в научном мире является теория биохимической эволюции, предложенная в 1924 г. советским биохимиком академиком А.И. Опариным (в 1936 г. он подробно изложил ее в своей книге «Возникновение жизни»).
Сущность этой теории состоит в том, что биологической эволюции — т.е. появлению, развитию и усложнению различных форм живых организмов, предшествовала химическая эволюция — длительный период в истории Земли, связанный с появлением, усложнением и совершенствованием взаимодействия
между элементарными единицами, «кирпичиками», из которых состоит все живое — органическими молекулами.
118.
112. Происхождение человека.Долгое время отсутствовали эмпирические данные о предках человека. Дарвин знал только дриопитеков (найденных в 1856 г. во Франции) и писал о них как о далеких предках человека. В XX веке раскопки позволили обнаружить остатки
ископаемых обезьян, живших примерно от 20 до 12 млн. лет назад. К ним относятся проконсулы (обнаруженные в Восточной Африке), ориопитек (находка скелета в 1958 г. в Италии), рамапитек (30-е годы XX века в Индии), сивапитеки и др.,
которые уже по многим признакам обнаруживают сходство с человеком.
В настоящее время большинство специалистов считает, что ближайшим предшественником человека являются австралопитеки - прямоходящие млекопитающие. Их костные остатки, возраст которых составляет от 5 до 2,5 млн. лет, впервые
были обнаружены в 1924 г. в Южной Африке. К настоящему времени обнаружены костные остатки около 400 особей австралопитековых. Австралопитеки были связующим звеном между животным миром и первыми людьми.
В современной антропологии наиболее распространенной является точка зрения, по которой "эволюция человеческой линии заняла не свыше 10 млн. лет, а обезьяний предок гоминид имел черты сходства с шимпанзе, был по существу
"шимпанзеподобен"... В качестве "модельного предка" человеческой и шимпанзоидной линии некоторые антропологи рассматривают карликового шимпанзе - бонобо - ... из джунглей Экваториальной Африки" (Хрисанова Е.Н., Перевозчиков
И.В. Антропология, М.: 1991, стр.37-38). В 1891 г. голландский исследователь Эжен Дюбуа на о.Ява впервые нашел окаменелости древнейшего человека - первого питекантропа, или человека прямоходящего. Уже в нашем веке на Яве
найдены еще несколько питекантропов, в Китае - близкие к ним синантропы и т.д. Все они представляют собой различные географические варианты человека прямоходящего, существовавшего приблизительно 0,5-2 млн.лет назад. Наряду с
добыванием растительной пищи у питекантропов большую роль играла охота. Они умели пользоваться огнем, сохраняли его от поколения к поколению. В 60-70-е годы нашего века в Африке были обнаружены остатки древнейших людей и
самые примитивные орудия труда из гальки. Этот древнейший предок человека получил название человека умелого. Человек умелый, судя по найденным останкам, датирующимся 2,6-3,5 млн. лет назад, существовал более полумиллиона
лет, медленно эволюционировал, пока не приобрел значительное сходство с человеком прямоходящим.
Древнейших людей - питекантропов - сменили древние люди, которых называют неандертальцами (по месту первой находки в долине реки Неандр, Германия). Их скелетные остатки открыты в Европе, Азии и Африке. Время существования
- 200-35 тысяч лет назад. Они могли не только поддерживать, но и добывать огонь. Шло развитие речи. С помощью изготовленных орудий древние люди охотились на животных, сдирали с них шкуры, разделывали туши, строили жилища. У
неандертальцев впервые встречаются захоронения. В гроте Кроманьон во Франции было обнаружено сразу несколько ископаемых людей современного типа. По месту находки их называют кроманьонцами. Самые ранние их костные остатки
датируются в 40 тысяч лет. Разнообразие типов орудий из камня и кости говорит о сложной трудовой деятельности. Человек уже умел сшивать шкуры животных и изготавливать из них одежду, жилье. На стенах пещер обнаружены
мастерские рисунки.
Происхождение человека
Этапы становления человека. В эволюции человека (Homo) различают 4 этапа:
1)Предлюди (протоантропы)
Вид человек умелый - Homo habilis
2)Древнейшие люди (архантропы), к которым относятся питекантроп, синантроп и гейдельбергский человек
Вид человек прямоходящий - Homo erectus
3)Древние люди (палеоантропы)
4)Современные (новые) люди, включающие ископаемых кроманьонцев и современных людей
Вид человек разумный - Homo sapiens Челове́к уме́лый (лат. Homo habilis) — высокоразвитый австралопитек или первый представитель рода Homo. Обнаружен археологами Лики (Мэри и Джонатаном) в 1960 году Человек умелый, судя по
найденным останкам, датирующимся 2,6—2,5 млн. лет назад, существовал более полумиллиона лет.
Масса головного мозга этого гоминида была 650 граммов, что было намного больше, чем у типичных австралопитеков. Он также отличался от австралопитеков строением таза, обеспечивавшим более совершеннуюбипедальность и рождение
более «головастых» детёнышей. Эндокранокруглый и точек роста не имеет. Однако у Человека умелого происходит перераспределение долей мозга — более примитивная затылочная доля мозга уменьшается в пользу увеличения более
прогрессивных долей — лобной, теменной, височной с ассоциативными долями. Отличался он от австралопитековых и строением черепа — череп расширен в подглазничной и теменно-затылочной областях. Размер зубов уменьшается,
зубная эмаль становится менее толстой. Структура кисти Homo habilis мозаична и сочетает в себе как прогрессивные черты, так и следы адаптации к лазанью по деревьям. Расширение ногтевых фаланг — прогрессивная черта,
свидетельствующая о формировании пальцевых подушечек как осязательного кинестетического аппарата. Формировался силовой захват, с помощью которого можно было изготовлять орудия труда. От ноги хабилиса остались пять фаланг
пальцев, пять костей ступни, пяточная кость и лодыжка. Это была примитивная нога по соединению костей, но всё же — нога человека. У Человека умелого первый палец стопы не был отведён в сторону, а так же, как у нас, располагался
вместе с другими пальцами. Это означало, что нога его была полностью приспособлена только к двуногому передвижению.
Объём мозга Человека умелого — 650-1100 см³. Рост составлял 1,0—1,5 м, вес — около 30—50 кг. Его лицо имело архаичную форму с надглазничными валиками, плоским носом и выступающими вперёд челюстями. Голова Человека умелого
стала более округлой формы, чем у австралопитеков; мозг также стал крупнее, хотя составлял все ещё лишь половину мозга современного человека. Выпуклость внутри тонкостенного черепа говорит о наличии у них центра Брока — центра
речи, но гортань, возможно, не была способна производить столько же звуков, сколько наша гортань. Челюсти были менее массивные, чем у австралопитека; кости рук и бёдер кажутся более современными, а ноги имели вполне
«современную» форму. У особей Человека умелого был заметный половой диморфизм — самки имели более широкие бёдра по сравнению с самцами. Рацион питания был в основном вегетарианский, но так же входило в него и мясо, в
частности некоторых видов австралопитеков.
Человек умелый, по-видимому, первое существо, сознательно изготовившее орудия труда и охоты: первые ещё грубо обработанные каменные гальки (орудия олдувайской культуры) неоднократно находили вместе с останками этого
существа. Именно Человек умелый перешагнул невидимую границу, отделяющую род Homo от всех других биологических существ — он сделал первый шаг по пути подчинения себе окружающей природы. Орудия, которые делал Человек
умелый, почти все были кварцевые, а кварц в местах стоянок этих людей не водился. Они приносили его с расстояния от 3 до 15 км. Это доказывало, что Человек умелый действительно был человеком. Он заранее подбирал камень для своих
орудий. Ни одно из животных не только не подбирает сырьё для своих орудий, но и вообще не додумывается раскалывать камень, чтобы сделать его острым, превратить в орудие. Однако в отличие от более поздних видов Homo, небережно
относились к изготовленным собой орудиям труда, и после использования попросту выбрасывали их. Учёные провели серию исследований и пришли к выводу, что кисть Человека умелого была способна к труду. Она обладала силовым
захватом большей мощности. Ни у одной обезьяны таких способностей нет.
119.
114) Понятие о расах и видовое единствоНесмотря на неоднократные попытки доказать, что человеческие расы представляют собой разные виды и даже разные роды, все же подавляющее большинство исследователей определяет
человечество как единый вид.
Видовое единство человечества может быть обосновано множеством убедительных доказательств.
Сравнительно-анатомическое изучение человеческих рас позволяет сделать следующие выводы:
1. В отношении признаков строения тела, связанных с общественной, трудовой деятельностью, все человеческие расы чрезвычайно сходны друг с другом.
2. Это сходство во многих случаях охватывает мельчайшие детали строения органов.
3. По строению органов тела ни одна раса ни в какой степени не приближается в целом к какой-либо антропоморфной обезьяне; все расы в равной степени обладают специфическими для
человека чертами строения.
4. Человеческие расы весьма сходны также по многим признакам, не имеющим сколько-нибудь важного жизненного значения, причем наблюдающиеся различия в частоте разных вариантов
между расами не нарушают их сходства в общем плане строения.
5. Факты сравнительной анатомии свидетельствуют в пользу теории монофилетического происхождения человека и его рас.
Все расы при смешении дают вполне плодовитое потомство. Предположения о том, что существуют расы, которые при скрещивании с другими расами якобы дают бесплодных или мало
плодовитых метисов, оказались ошибочными.
Все области соприкосновения разных расовых типов всегда являлись и являются одновременно и поясами смешения; даже искусственно воздвигавшиеся социальные преграды и
террористические меры, направленные к сохранению «чистоты» расы, никогда не могли полностью остановить стихийного процесса расового смешения и возникновения промежуточных типов.
Все ныне живущее человечество с точки зрения биологической науки представляет один вид Homo sapiens ("человек разумный"), распадающийся на более мелкие подразделения, именуемые
расами. Термин "раса" созвучен арабскому "ras" ("начало", "происхождение"), итальянскому "razza" ("раса", "племя").
Расы человека отличаются друг от друга рядом признаков - цветом кожи, волос, глаз, типом волос, чертами лица, формой черепа, пропорциями тела и другими не существенными для
жизнедеятельности признаками. Расовые признаки связаны с наследственностью и мало меняются под воздействием среды.
Расы связаны с определенной территорией (ареалом).В настоящее время данная связь может не обнаруживаться (черная раса представлена в населении Африки и Америки), однако она
существовала в прошлом. Не все биологические признаки человека являются расовыми признаками. Например, развитие жироотложения, мускулатуры, различия в осанке - не расовые отличия,
так как связаны с влиянием среды сейчас. На Земле уже насчитывается около 6 млрд. людей. Среди них нет, и не может быть двух совершенно одинаковых людей; даже близнецы, развившиеся
из одного яйца, несмотря на огромное сходство их внешнего облика, и внутреннего строения, всегда какими-то мелкими чертами отличаются друг от друга. Наука, изучающая изменения
физического типа человека, известна подименем "антропологии" (греч, "антропос" – человек). Особенно сильно заметны телесные различия между территориальными группами людей,
удаленными друг от друга и живущими в различной естественно-географической обстановке.Деление вида Homo Sapiens на расы произошло два с половиной столетия назад. Происхождение
термина "раса" точно не установлено; возможно, что он представляет собой видоизменение арабского слова "рас" (голова, начало, корень). Есть также мнение, что термин этот связан с
итальянским razza, что значит "племя". Слово "раса" примерно в том смысле, как оно употребляется теперь, встречается уже у французского ученого Франсуа Бернье, который опубликовал в 1684
г. одну из первых классификаций человеческих рас.Расы - это исторически сложившиеся группировки (группы популяции) людей разной численности характеризующиеся сходством
морфологических и физиологических свойств, а также общностью занимаемых ими территорий.Развиваясь под влиянием исторических факторов и относясь к одному виду (H.sapiens), раса
отличается от народа, или этноса, который, обладая определенной территорией расселения, может содержать несколько расовых комплексов. К одной и той же расе может принадлежать ряд
народов и носителей многих языков. Большинство ученых сходятся во мнении, что существует 3 крупных расы, которые в свою очередь распадаются на более мелкие. В настоящее время по
мнению разных ученых, насчитывается 34 – 40 рас. Расы отличаются друг от друга по 30-40 элементам. Расовые особенности наследственны и являются приспособительными к условиям
существования. Целью моей работы является систематизировать и углубить знания о человеческих расах.
120.
115)Этич-е пробл. мед. био и генетики.Больные с наследственными заболеваниями и их семьи составляют большую группу населения, по отношению к которой существует много этических вопросов, возникающих при оказании им врачебной помощи. Все элементы врачебной деонтологии и общечеловеческой морали, сформулированные
обществом со времен Гиппократа, и сейчас остаются в силе для этой группы больных. Однако своеобразный характер течения большинства наследственных болезней, а именно: их пожизненность, прогредиентность, тяжесть и особенно их свойство передаваться от поколения к поколению - ставят перед
врачами и обществом специфические этические вопросы на фоне новейших успехов в генетике человека. Суть успехов в том, что они обеспечили такие генетические технологии, которые позволяют вмешиваться в геном человека. Предпосылками для выработки правовых и законодательных регуляций
любого характера являются моральные нормы общества. Следовательно, биоэтическое рассмотрение новых научных достижений - первый шаг к предупреждению отрицательных последствий научных достижений. Данное положение особенно явно просматривается на примере бурно развивающихся
дисциплин, к которым, несомненно, относится генетика во всем ее многообразии. Формирование правовых положений для медико-биологической науки и практической медицины на основе моральных принципов общества не может быть отделено от формирования правосознания в разных социальных
группах (ученые, врачи, пациенты, политики и т.д.). Главным результатом биоэтических разработок становится своевременное обсуждение морально-правовых проблем, возникающих в новых областях науки и практики. На основе обсуждений и научных исследований разрабатываются национальные и
международные законы, рекомендации, правила - как для проведения исследований, так и для практической реализации их результатов. В генетике человека четко прослеживаются непосредственная связь научных исследований с этическими вопросами, а также зависимость научных поисков от этического
смысла их конечных результатов. Генетика шагнула вперед настолько, что человек рано или поздно сможет определять свою биологическую судьбу. В связи с этим использование всех потенциальных возможностей медицинской генетики реально только при строгом соблюдении этических норм.
Прогресс медицинской генетики поставил этические вопросы в связи с:
• генной инженерией (генодиагностика и генотерапия);
• разработкой методов ранней диагностики наследственных болезней (см. статью В.Л. Ижевской «Этико-правовые аспекты генетического тестирования и скрининга» на компакт-диске);
• новыми возможностями медико-генетического консультирования;
• пренатальной и предымплантационной диагностикой наследственных болезней (см. статью «Этические аспекты пренатальной диагностики» на компакт-диске);
• охраной наследственности человека от повреждающего действия новых факторов окружающей среды.
Поскольку медицинская генетика имеет дело с больным человеком или его семьей, она должна опираться на выработанные и проверенные веками принципы медицинской деонтологии. Однако в современных условиях этого недостаточно, потому что в биоэтике возникают новые вопросы:
• внедрение принципиально новых медицинских и генетических технологий (искусственное оплодотворение, суррогатное материнство, пренатальная диагностика, генетическое тестирование донора, генотерапия) стало массовым в медицинской практике;
• медико-генетическая помощь и генетические технологии все больше коммерциализируются как на Западе, так и у нас в стране;
• появились новые формы взаимоотношений врача и пациента, формируются общества пациентов и их родителей (с болезнью Дауна, муковисцидозом, фенилкетонурией и др.);
• потребовалось этическое и правовое регулирование научных исследований, их направлений и итогов, поскольку они затрагивают интересы общества (дополнительное финансирование, угроза войны и т.д.).
Большинство этических вопросов современной генетики человека можно решить в рамках 4 принципов (делай благо, не навреди, автономия личности, справедливость) и 3 правил (правдивость, конфиденциальность, информированное согласие).
Принцип «делай благо». Применение этого принципа на практике сталкивается с противоречием между благом конкретного человека и благом группы людей или общества в целом. На этой основе возникли евгенические программы насильственной стерилизации пациентов с отклонениями в умственном и
физическом развитии в США, Дании, Швеции, Германии и других странах. Главным обоснованием таких мероприятий был приоритет общего блага нации над индивидуальным. Современные моральные принципы обязывают искать компромисс между интересами общества и отдельного человека. В ряде
международных документов утверждается норма, согласно которой интересы пациента ставятся выше интересов общества. Соблюдая принцип «делай благо», не во всех случаях можно определить, что является благом для пациента, а что - благом для его семьи. Если раньше право решать принадлежало
врачу-генетику (например, директивное консультирование считалось нормой), то современная мораль общества принципиально изменила ситуацию. Принимает решение пациент вместе со своей семьей, а недирективное консультирование стало нормой работы врача-генетика. Принцип «не навреди»
запрещает исследовательские и терапевтические действия, связанные с неоправданным риском неблагоприятных последствий для пациента. Однако на стадии клинических испыта-ний моральная ответственность врача занимает большее место, чем правовая. Принцип автономии личности - это признание
свободы и достоинства пациентов или участников эксперимента. Их следует уважать как собственников своей жизни и здоровья. Никакие вмешательства нельзя проводить без их согласия. Применительно к медицинской генетике этот принцип может легко нарушаться врачом или исследователем при
передаче образцов ДНК по запросу, сохранении и размножении клеток и т.п. В современной генетике принцип автономии личности должен распространяться на потомков обследуемого в той же мере, в какой сохраняется право наследования имущества. Принцип справедливости учитывает равную
доступность ресурсов медико-генетической помощи через систему государственного здравоохранения, с одной стороны, и моральную оправданность неравенства уровня медико-генетической помощи в частном секторе здравоохранения, обусловленного рыночными отношениями, - с другой.
Наряду с 4 принципами современной биоэтики выделяют еще 3 правила.
Первое правило - правило правдивости. Моральный долг врача и ученого обязывает говорить правду пациентам или участникам эксперимента. Без этого они не могут сами принять правильное решение. В генетическое обследование вовлекается не только один человек, но и члены его семьи, что и создает
этически трудные ситуации для врача-генетика.. Доверие между врачом и пациентом может поддерживаться только при соблюдении обоюдно правдивых отношений между ними. Если пациент скрывает сведения своей родословной, это обязательно отразится на заключении врача.
Второе правило - правило конфиденциальности. На первый взгляд его легко соблюдать, но это не всегда так. Чем глубже обследуется пациент (например, на генном уровне), тем больше затруднений в соблюдении этого правила. Например, разглашение информации о генетической характеристике пациента
может нанести ему вред (отказ в приеме на работу, отказ от предстоящего брака). Правило конфиденциальности требует полного согласия пациентов на передачу полученной при генетическом исследовании информации. Наиболее трудные случаи соблюдения правила конфиденциальности создает изучение
родословной. Например, может ли пациент получить информацию от врача о генетическом здоровье своих родственников, если они на это не согласны, могут ли родственники узнать о генетическом диагнозе пациента. И в том и в другом случае это может затрагивать моральные интересы каждой стороны.
Третье правило - правило информированного согласия. Оно во многом уже вошло в правовые и юридические нормы, регламентирующие проведение медицинских испытаний и вмешательств. Любое генетическое обследование должно проводиться с согласия пациента или его законных представителей на
основе достаточной информации, выраженной в понятной для пациента форме.